CHƯƠNG III. ENZYME - tailieukhoathuy.files.wordpress.com · –2.3. Trung tâm hoạt động...
71
CHƢƠNG III. ENZYME
Transcript of CHƯƠNG III. ENZYME - tailieukhoathuy.files.wordpress.com · –2.3. Trung tâm hoạt động...

CHƢƠNG III. ENZYME

NỘI DUNGI. ĐẠI CƢƠNG VỀ ENZYME
– 1.1. Khái niệm
– 1.2. Tên gọi và phân loại enzymeII. CẤU TRÚC VÀ CÁC DẠNG ENZYME
– 2.1. Cấu trúc phân tử
– 2.2. Các cofactor
– 2.3. Trung tâm hoạt động – active siteIII. TÍNH ĐẶC HiỆU CỦA ENZYME
IV. CƠ CHẾ XÚC TÁC CỦA ENZYME
V. CÁC Y/TỐ Ả/HƢỞNG TỚI H/TÍNH X/T CỦA ENZYME
5.1. Động học các p.ứ. E (a/h của [E] và [S])
5.2. Các yếu tố lý hoá hoá của môi trƣờng
5.3. Các chất ả/hƣởng đến h/động của enzyme

I. ĐẠI CƢƠNG VỀ ENZYME
• 1.1. Khái niệm
• Enzyme: là chất xúc tác sinh học
(biocatalyst), làm tăng tốc độ các phản
ứng hoá sinh.
• Bản chất: protein (trừ ribozyme - ARN có
khả năng xúc tác)

ENZYME
Ƣu điểm
• Làm tăng tốc độ phản ứng, không tham gia vào sản phẩm cuối cùng
• Hiệu quả xúc tác lớn: Ví dụ, 2H2O2 2H2O + O2
• Không xúc tác, hằng số t.độ ph.ứng là 0,23/s, NLHH: 18kcal/mol
• Pt xúc tác: 1,3 x 103/s; NLHH: 11,7kcal/mol
• catalase xúc tác: 3,7.107/s; NLHH: 2kcal/mol

ENZYME
Ƣu điểm
• Có tính đặc hiệu theo kiểu phản ứng và cơ chất
• Xúc tác trong những điều kiện môi trƣờng tƣơng đối ổn định (to khoảng 20- 40o C, áp suất khoảng 1 at, pH 7).
• Tác dụng của enzyme có thể đƣợc điều khiển

ENZYME
Nhƣợc điểm
• Rất mẫn cảm với hàng loạt yếu tố
• Thƣờng xuyên đƣợc sử dụng rất nhiều, nhƣng luôn bị phân giải và tổng hợp trở lại theo nhu cầu.

1.2. Đơn vị hoạt lực của enzyme
• Năm 1961, IUB (International Union of Biochemistry) đưa ra đơn vị hoạt lực enzyme chuẩn: U (1U) là lượng enzyme cần thiết để biến đổi 1 mol cơ chất trong thời gian 1 phút ở điều kiện chuẩn (30o C, pH tối ưu, bão hoà cơ chất).
• Năm 1972, IUB đưa ra đơn vị mới là katal (kat): 1kat là lượng chất xúc tác làm biến đổi 1 mol cơ chất trong thời gian 1 giây ở điều kiện chuẩn.
• kat = 10-6 kat; ηkat = 10-9 kat
• Liên hệ giữa U và kat: 1U = 16,67 ηkat

1.3. Tên gọi và phân loại enzyme1.3.1. Tên gọi
• Pepsin
• Trypsin…Tên enzyme + in
• Tên gọi theo cơ chất:
• VD: amylase, protease, lipase
• Theo kiểu ph.ứng:
• VD: oxidase, hydrolase
Enzyme+ase
• E xúc tác cho cơ chất A nhờ dạng ph.ứng R có tên là ARase: VD: Glyceraldehyd-3-phosphate-hydrolase
• E x.tác ph.ứng của chất A với chất B (hay cofactor B) nhờ ph.ứng dạng R, có tên A:B-Rase
Tên hệ thống

1.3.2. Phân loại enzyme theo kiểu
phản ứng
ENZYME
OXIDOREDUCTASE
TRANSFERASE
HYDROLASE
LIASAE (SYNTHASE)
ISOMERASE
LIGASE (SYNTHETASE)

Lớp 1: Oxidoreductase
Xúc tác cho các phản ứng oxy
hoá khử
Bản chất: protein ph.tạp
Vận chuyển: hydro, e-, gắn
oxy vào cơ chấtPhân thành các phân lớp theo
nhóm chức năng nhƣờng hydro hay e-
Lớp lớn nhất
CH3 – CO – COO -
Pyruvate
NAD.H+H+ NAD+
CH3 – CH.OH – COO -
Lactat
VD:
Lactate dehydrogenase

Lớp 2: Transferase
Vận chuyển nhóm (CH3, NH2, vv…)
Bản chất: protein ph.tạp
Phân thành các phân lớp theo nhóm đƣợc vận chuyển
R1-CH.NH2-COOH
Amino acid
R2- CO-COOH
Keto acid
+ Aminotransferase
R1- CO - COOH R2-CH.NH2-COOH
Ketoacid mới Amino acid mới
+
VD:

Lớp 3: Hydrolase
Xúc tác cho các phản ứng
thuỷ phân
Bản chất: protein đơn
giản
Thuỷ phân các liên kết vốn hình thành nhờ sự ngƣng tụ
nhƣ peptide, glycoside, ester …
VD:
3 H.OH
lipase+
R1COOH
R2COOH
R3COOH
acid béoTriacylglycerol
CH2 -O-CO-R3
CH2-O-CO-R1
CH -O-CO-R2
Glycerol
CH2 – OH
CH2 - OH
CH - OH

Lớp 4: Liase (synthase)
Xúc tác cho các phản ứng:
phân giải (không thuỷ phân)
hình thành (không đòi hỏi NL) VDcác liên kết C- C, C- O, C- N, vv…
Bản chất: protein phức
tạp
Phân thành các phân lớp theo kiểu l/kết h/học đƣợc ph/giải
hay tạo thành.
VD: Pyruvate decarboxylase tách CO2 từ pyruvate tạo ra acetaldehyd.
CH3 – CO – COO -
Pyruvate decarboxylase
CO2
pyruvate acetaldehyde

Lớp 5: Isomerase
Xúc tác cho các phản ứng đồng phân hoá
Bản chất: protein đơn
giản
Vận chuyển: các ng/tử hay nhóm ng/tử trong nội bộ một ph/tử
Lớp nhỏ nhất
Dihydroxyacetonphosphate Glyceraldehyd-3-phosphate
Izomerase
CH2-O-PO32-
CH2OH
C = O
CH2-O-PO32-
CHO
CH.OH

Lớp 6: Ligase (Synthetase)
Xúc tác cho các q.trình sinhtổng hợp
Bản chất: protein phức
tạp
hình thành nên các l.kết nhờ tiêu tốn
n.lƣợng (VD: ATP)

II. CẤU TRÚC VÀ CÁC DẠNG ENZYME
• 2.1. Cấu trúc phân tử
• Là các protein hình cầu, phần lớn (60-
70%) có bản chất là protein phức tạp.
• Xét về c.trúc, có hai loại enzyme:• Đơn giản (một thành phần)
• Phức tạp (hai thành phần):
– Protein (apoenzyme)
– Cofactor

2.2. Các cofactor
• 2.2.1. Khái niệm
• Cấu trúc nhỏ, không đƣợc cấu tạo từ các aa.
• Thành phần của các enzyme phức tạp, làm
nhiệm vụ vận chuyển các nguyên tử hay e- trong
các phản ứng hóa học mà enzyme của nó xúc
tác.
• Cofactor gồm 2 loại:
– Nhóm ghép
– Coenzyme

• Nhóm ghép (prosthetic group)
– Loại liên kết chặt với apoenzyme, là thành
phần cố định của phân tử enzyme.
• VD: FMN; FAD của dehydrogenase
– PLP của aminotransferase
– Hem của cytochrome
• Coenzyme
– Loại gắn lỏng lẻo với apoenzyme, dễ tách ra
và nhập lại, chạy từ apoenzyme này tới
apoenzyme kia.
• VD: NAD+; NADP+ của nhiều dehydrogenase

2.2.2. Cấu trúc của cofactor
• Bản chất hóa học khác nhau; phân tử
thƣờng chứa dị vòng.
• Phần trực tiếp tham gia phản ứng hoặc có
chức năng nhận biết các đại phân tử.
• Nhiều cofactor là dẫn xuất của các
vitamin tan trong nƣớc và phần lớn
thƣờng chứa phosphate gắn trong
nucleotid.

2.2.2.1. Cofactor của các
oxidoreductaseNAD+ (Nicotinamid–Adenine-Dinucleotid) – Vit PP
NADP+ (Nicotinamid-Adenine-Dinucleotid-Phosphate) – Vit PP
FMN (Flavin - Mononucleotid) – Vit B2
FAD (Flavin – Adenine – Dinucleotid) – Vit B2
Lipoate (6,8 dithioctanate)
Coenzyme Q
Hem

Cofactor của các oxidoreductase
• NAD+ (Nicotinamid–Adenine-Dinucleotid)
• NADP+ (Nicotinamid-Adenine-Dinucleotid-Phosphate)
– Dẫn xuất của vit. PP (nicotinamid, niacin)
– NAD+ và NADP+ là coenzyme của khoảng
250 dehydrogenase.
• Cơ chế hoạt động


Cofactor của các oxidoreductase
• FMN: Flavin - Mononucleotid
• FAD: Flavin – Adenine - Dinucleotid
• Dẫn xuất của vit. B2 (Riboflavin)
• FMN và FAD liên kết chặt với apoenzyme,
tạo thành flavoprotein
• Dạng OXH (FAD, FMN) có màu vàng. Lõi
hoạt động là vòng isoalloxasine
(isoalloxasine ring)
•

FMN: Flavin mononucleotid
FAD: Flavin – Adenine - Dinucleotid

Cơ chế hoạt động


Lipoate (6,8 dithioctanate)
• Coenzyme Q
– Vận chuyển hydro
– thành viên của chuỗi hô hấp
+ 2H
- 2 H
COOH
S S
Dạng OXH
COOH
S H S H
Dạng khử


Hem
• Nhóm ghép của catalase, peroxidase, các cytochrome
• Vận chuyển e-

2.2.2.2. Cofactor của các
transferase
ATP (Adenosine Triphosphate)
TPP (thiamine pyrophosphate) – Vit B1
PLP (pyridoxal phosphate) – Vit B6
Coenzyme A, CoASH (coenzyme acyl hoá)

Cofactor của các transferase• ATP (adenosine triphosphate): cofactor của các
transferase có tên là kinase

TPP (thiaminepyrophosphate)
• Là dẫn xuất của vitamin B1
• Vòng pyrimidine gắn với thiazol nhờ cầu -CH2-.
• Phần th.gia p.ứ.là vòng thiazol, vòng pyrimidine và nhóm
diphosphate làm n/v gắn với apoenzyme.
Vòng thiazol
Vitamin B1 (thiamine)

PLP (pyridoxalphosphate)
• Dẫn xuất của vitamin B6
• Nhóm ghép của:
– transaminase - chuyển amin
– decarboxylase - khử carboxyl cho aa
• Ngoài NAD(P+), PLP là cofactor thứ 2 có nhân pyridine
Pyridoxal

Coenzyme A, CoASH (coenzyme acyl hoá)
• Vận chuyển acyl acyl đƣợc gắn vào nhóm thiol (-SH)
nhờ liên kết thioester
acyl.CoA (AB hoạt hoá)R - CO – S - CoA
CH3CO – S - CoA Acetyl – CoA (a. acetic hoạt động)

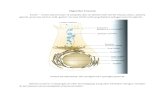
2.3. Trung tâm hoạt động – active site
• 2.3.1. Khái niệm
– Vùng không gian giới hạn nhỏ, chứa các nhóm
chức năng đƣợc phân bố, định hƣớng một cách
chính xác. Các nhóm chức năng này là thành phần
của các gốc aa đôi khi xa nhau trên chuỗi
polypeptide, song lại gần nhau trong không gian
nhờ sự cuộn lại, gấp nếp lại của chuỗi (nhờ ctb 3).
– Ở các enzyme phức tạp có coenzyme, TTHĐ có
vùng liên kết với coenzyme.

• Là nơi gắn cơ chất, chứa các nhóm chức
góp phần trực tiếp vào việc cắt đứt hay
hình thành các liên kết để tạo ra sản
phẩm.
• Các nhóm chức năng:
– :OH của Ser
– :SH của Cys
– COOH của Glu và Asp
– Vòng imidazol của His
– NH2 của Lys

Nhiệm vụ của các nhóm chức năng
• Một số nhóm xúc tác tích cực (tạo vùng
xúc tác)
• Một số nhóm gắn cơ chất (tạo vùng liên
kết
• Một số nhóm tạo môi trƣờng và cấu trúc
không gian thích hợp cho TTHĐ.

2.3.2. Đặc điểm của TTHĐ
• E có thể có 1; 2 hay nhiều TTHĐ. Các
TTHĐ của 1 p.tử E có thể giống nhau,
song có thể khác nhau về c/tạo và c/n, do
đó 1 p.tử E có thể xt nhiều p.ứ. h.học
khác nhau.
• Chỉ S đ/hiệu, có c/trúc p.tử thích hợp với
TTHĐ của E mới tạo ra đƣợc ph/hợp E-S
và q/trình xt chỉ xảy ra khi S đƣợc gắn vào
TTHĐ của E.

2.4. Các tổ hợp đa enzyme và các
enzyme dị lập thể
• 2.4.1. Các tổ hợp đa enzyme
(multienzyme complex)• Nhiều p.tử E chỉ là một chuỗi polypeptide (đơn nguyên).
• Nhiều E có ctb4, do nhiều chuỗi tổ hợp lại. Đây là những
E đa nguyên (oligomer). Các E này thay đổi h.tính xt khi
các chuỗi tách rời nhau.
• Phần lớn các E hđ theo 1 dây chuyền p.ứ trong tổ hợp
đa E. (SP của 1 p.ƣ. E là S cho p.ứ của E tiếp theo)
– VD: Tổ hợp pyruvate dehydrogenase
– Tổ hợp đa enzyme t/hợp acid béo

VD: Acyl synthase (phức hợp
enzyme tổng hợp acid béo)
• 6 enzyme và một protein mang gốc acyl
ACP (Acyl carrier protein)

Ý nghĩa của các tổ hợp đa enzyme
• C/trúc bậc cao của E tạo ra các t/chất mới mà
các c/trúc bậc thấp không có.
• P.tử có nhiều tiểu phần tạo ra k/n đ.khiển hoạt
tính x/t một cách l/hoạt và h/quả.
• Tạo đ.k. để S đƣợc b/đổi ở nhiều mức (trình tự
nhƣ 1 dây chuyền sx)
– SPTG chuyển từ 1 TTHĐ sang TTHĐ tiếp
theo, không bị g/ph ra khỏi kh/vực p.ứ.
– SPTG không dồn đống lại, q.t. xt xảy ra rất
nhanh.

2.4.2. Enzyme dị lập thể (allosteric enzyme)
• Ngoài TTHĐ, có TTĐK (TT dị lập thể) để đ/chỉnh
h/tính của E. Tuy có t.dụng t/hỗ, song 2 TT này
có c.trúc khác biệt nhau và định vị ở những
kh/vực khác nhau trên ptử E.
• TTĐK = nơi gắn chất có k/n làm b.đổi c/n xt của
E.
• H/tính đƣợc đ.kh bởi các chất gây h/ứng dị lập
thể (allosteric effectors) là chất có k/n b/đổi cấu
hình E và ả/h tới h/tính xt của E.

Chất ức chế làm mất khả năng xúc tác của E

2.5. Sự phân bố và các dạng enzyme
• 2.5.1. Sự phân bố của các enzyme
– Enzyme nội bào: chiếm đa số, có mặt và
th.hiện c/n bên trong TB nơi đã sinh ra nó
– Enzyme ngoại bào: Đƣợc TB tiết ra bên
ngoài, có mặt trong các dịch bào (t/hoá, não
tuỷ, máu).
• Một số VSV tiết E (th/phân các chất dd) vào mt
nuôi cấy.

2.5.1. Các dạng enzyme
Enzyme
Zimogen
(Proenzyme)
Dạng tiền chất, chƣa hoạt động
Isozyme
(Isoenzyme)
Nhiều dạng khác nhau của
1 enzyme

Các zimogen hay proenzyme
• Dạng tiền chất, dạng chƣa h/đ của E
• Có tiếp đầu ngữ "pro" hay đuôi “ogen" gắn
với tên E
– VD: Procarboxypeptidase, pepsinogen,
trypsinogen, …
• Các zymogen đƣợc v/c nhờ dịch bào từ
nơi hình thành đến nơi th/gia các hđ h/học

Cơ chế hoạt hóa các zimogen
• Cắt đi 1 đoạn p/tử chƣa hđ → bộc lộ TTHĐ
• Phá vỡ 1 hay một vài lk mấu chốt để s/xếp lại
c/trúc kh/gian, tạo TTHĐ
– VD: chymotrypsinogen
• Quá trình hoạt hoá thực hiện nhờ:
– Protease đặc hiệu
– Dạng hđ của chính E ấy (tự h/hoá).
• Đây là cơ chế điều hòa hàm lƣợng E, đồng thời
bảo vệ cho TB đã sản xuất ra E này.

Các isozyme hay isoenzyme
• Nhiều dạng khác nhau của 1 E cùng xt
cho 1 pứ nào đó (cùng c/năng x/tác).
– Có thể là các dạng của 1 E đƣợc thay thế
dần trong qt phát triển của cơ thể
– Hay các dạng của một E đảm nhiệm những
v/trò giống nhau ở những m/bào khác nhau.

Các isozyme hay isoenzyme
• VD: lactate dehydrogenasa (LDH), ptử có
4 tiểu phần thuộc 2 dạng: H và M.
– Các t/phần này có thể k/hợp tạo ra 5 isozyme:
H4, H3M1, H2M2, H1M3 và M4; chiếm t/lệ khác
nhau ở những mô khác nhau.
• Các isozyme của 1E khác nhau về ctb1 do
chúng là SP của các gen độc lập.

Ý nghĩa của các isozyme
• Để cùng 1 p.ứ ph/vụ những m/đích khác nhau hay đƣợc
đ/khiển theo 1 cách khác, đòi hỏi E khác đi một chút.
• Sự t/tại các dạng E đ/hiệu trong m/bào với k/năng x/t
đƣợc đ/hoà, biến đổi, làm SV có thể th/nghi với những
ĐK thay đổi của MT
– VD: LDH dạng M4 có k/n biến pyruvate thành lactate rất mạnh,
k/n này ở H4 lại thấp. M/bào nào đƣợc c/cấp đ/đặn và đủ O2 (cơ
tim), b/thƣờng không tạo lactate, chứa nhiều LDH dạng H4.
Ngƣợc lại, cơ vân có thể bị thiếu O2, lại cần dạng M4 (M =
muscle, cơ; H = heart, tim).

III. TÍNH ĐẶC HIỆU CỦA ENZYME
• 3.1. Đặc hiệu kiểu phản ứng
• Một S đƣợc chuyển hoá nhờ các E có tác
dụng đ/hiệu nhau thành những SP
nhau.
– VD: 1 aa có thể đƣợc ch/hoá theo kiểu p/ứ:
• khử nhóm amin oxidase
• chuyển amin transaminase
• khử carboxyl decarboxylase


• 3.2.1. Đặc hiệu cơ chất tuyệt đối
• Xúc tác cho 1 S duy nhất
– VD: urease chỉ xt sự th/phân urê NH3 và
CO2
• 3.2.2. Đặc hiệu cơ chất tƣơng đối (theo nhóm)
• Xúc tác 1 p.ứ nh/định cho 1 nhóm S cùng loại
– VD: alcol deh. x.t. sự oxy hóa của nhiều rƣợu
hexokinase chuyển phosphoryl từ ATP tới
các hexose khác nhau.
3.2. Đặc hiệu cơ chất

3.3. Các yếu tố quyết định tính đặc
hiệu của enzyme
• Tính đặc hiệu S do phần protein E quyết định
• Tính đặc hiệu phản ứng ở E ph.tạp do cả
apoenzyme và cofactor q/định. Một cofactor có thể
th.gia các p.ứ nhau, tuỳ đƣợc gắn vào protein
nào.
– VD: PLP trong transaminase th.gia p.ƣ. chuyển
amin, trong decarboxylase th.gia khử carboxyl.
• Ý nghĩa: các pứ h.sinh xảy ra theo một trật tự nhất
định; tuỳ từng lúc, từng đk mà diễn ra qt ph.giải,
hoặc là t/hợp.

IV. CƠ CHẾ XÚC TÁC CỦA ENZYME
• E + S ES
– Tạo NLHH
– Một phần E tham gia xuc tác, phần còn lại tạo
cấu trúc 3 chiều ổn định lk giữa E và S

∆G1: NL hoạt hóa của p.ứ không đƣợc xúc tác
∆G2: NL hoạt hoá của p.ứ. có E xúc tác
Hình 3.1: Tiến trình phản ứng và sự biến đổi năng lƣợng tự do

• Pứ tạo phức hợp ES diễn ra rất nhanh, ES
không bền vững.
• Chỉ có S có cấu trúc đặc hiệu mới gây
cảm ứng và kết hợp với TTHĐ của E.
• Khi S tƣơng tác với E:
– S bị biến đổi theo hƣớng hoạt động hơn
– Cấu tạo điện tử bị biến đổi:
• Lk căng ra
• Lk bị xoắn vặn
• Lk giảm độ bền vững
• S tăng tính hoạt động hóa học pứ xảy ra
nhanh chóng

E S+ ES1
ES*2 ES
EP3 ES*
P4 EP +E
Sự biến đổi các mức NL trong pứ xt của E trải qua
nhiều bƣớc (hình 3.2.):

Hình 3.2. Sucrase thủy phân sucrose thành
glucose và fructose

Yếu tố ảnh hƣởng đến hiệu quả
xúc tác và tính đặc hiệu của E
• TTHĐ có thể làm biến dạng hay phân cực các lk
của S S hoạt động hơn.
• TTHĐ [S] và cố định S trong 1 không gian
hình học chính xác với các nhóm xt.
• TTHĐ định hƣớng chính xác các S từng bƣớc
của pƣ diễn ra với sự dịch chuyển nhỏ nhất của
S.
• TTHĐ cố định S theo một phƣơng thức nhất
định NL cho pứ.

Bản chất hóa học của hiện tƣợng
xúc tác bằng E
• Các nhóm xt trong TTHĐ là nhóm ái nhân
- có các cặp e- tự do, có k/n tạo lk với các
nhóm ái e- của S:
– VD: OH, SH, N trong vòng imidazol
– Trong 1 số tr/hợp, ngƣợc lại, các nhóm xt lại
nhận e- từ nhóm ái nhân của S:
• VD: các ion kim loại và NH3+ trong TTHĐ

Bản chất hóa học của hiện tƣợng
xúc tác bằng E
• E làm việc theo ng/tắc xt acid-base h.tính của 1 trong
các S khi nhận H+ hoặc bị tách H+
- VD: nhóm carboxyl, amin, phenol, thiol và đ/biệt là vòng
imidazol
- Pứ E diễn ra nhờ các cofactor:
- Khi gắn với E, S tiếp xúc tr/tiếp với cofactor th/lợi
cho pứ xảy ra.
- các cofactor cũng c/cấp NL cho các pứ E
- VD: cofactor có mạch phosphate cao năng hay các
nucleotide.
- Nhiều E Vpứ nhờ tạo SPTG rất h/động với cơ chất.
Sau đó pứ phân thành 2 hay nhiều phần nhỏ, và NLHH
cũng chia thành những phần nhỏ hơn.

V. CÁC YẾU TỐ ẢNH HƢỞNG TỚI HOẠT
TÍNH XÚC TÁC CỦA ENZYM
• 5.1. Động học các p.ứ. E (ảnh hƣởng của [E] và [S])
[S] = constant
[E][E] = constant
[S]
Vmax [S]v =
Km + [S]
Phƣơng trình
Michaelis-Menten
Km = hằng số
Michaelis-Menten

5.2. Các yếu tố lý hoá hoá của môi trƣờng
• 5.2.1. Nhiệt độ– Tốc độ của pứ , khi t° . Nhƣng, khi t° tăng đ/thời có thể
làm E mất h/tính (apoenzyme b/tính, cofactor có thể bị tách
ra). Vì 2 h/tƣợng trái ngƣợc trên → E h/đ tốt nhất ở t°op

5.2.2. pH
• E là chất polyampholite, t/chất của E (cả h/tính x/tác) phụ thuộc vào pH; pứ của E và S ph/thuộc k/n tách H+
của các nhóm chức này. các E chỉ có kn x/tác ở một khoảng pH nh/định.
• E đạt t/độ x/tác cực đại ở pHop.
• Đa số E có pHop khoảng 5 - 7. Một số E tiêu hoá h/đ tốt nhất ngoài khoảng này:
– VD: pepsin pH = 1,5-2, trypsin và chymotrypsin pH = 8 -11, arginase pH = 9,5.
• Ở một số E, khoảng pHop khá hẹp, ở một số khác khá rộng. Đôi khi pHop có thể phụ thuộc vào loại S:
– VD: pHop ở các protease có thể th/đổi ph/thuộc loại protein đƣợc ph/giải.

5.3. Các chất ả/hƣởng đến h/động của enzyme
• 5.3.1. Chất hoạt hoá
– Làm tăng h/tính xúc tác enzyme, biến enzyme từ dạng khộng
hoạt động hoạt động
– Các chất h/hoá thƣờng không gắn cố định với E (không là
th/phần của E hay các nhóm ghép cố định).
– Chất hoạt hoá có thể là ion k/loại, các chất h/cơ; các anion
Cl-, -PO32-). VD
Pepsinogen PepsinHCL
Trypsinogen
Trypsin
Enterokinase
(cắt đoạn hexapeptide)
Chymotrypsinogen Chymotrypsin
Trypsin
(sắp xếp lại cấu trúc phân tử)

Sự h/thành các enzyme từ các zymogen t/ứng là c/chế đ/h h/lƣợng
E, đ/thời là c/chế b/vệ cho TB đã sinh ra loại E này.

5.3.2. Chất ức chế
• k/năng x/tác của enzyme
• Các chất ức chế (inhibitor) có b/chất khác nhau:
– VD: ion, các chất vô cơ hay hữu cơ
• Cơ chế:
– Làm thay đổi cấu trúc ph/tử enzyme làm enzyme mất
kh/năng x/tác (ức chế không cạnh tranh).
– Cạnh tranh với cơ chất về TTHĐ tốc độ ph/ứng,
hoặc không làm enzyme bị biến tính nhƣng làm cho
phức hợp ES không thể tạo ra s/phẩm và gi/phóng
enzyme đƣợc (ức chế cạnh tranh).

Ức chế cạnh tranh

Ức chế không cạnh tranh

• Ý nghĩa:
– Là công cụ điều hoà của TB và có rất nhiều ý nghĩa thực tiễn:
• Y học, vệ sinh (chống nhiễm trùng),
• Nông nghiệp (s/dụng thuốc trừ sâu, diệt côn trùng)...
– Ức chế E là hiện tƣợng rất phổ biến trong đời sống sv. Trên 90% các h/tƣợng trúng độc là do sự ức chế hoạt động của các E.
• Ví dụ : Các cyt. trong chuỗi h/hấp có nhóm ghép (hem) với Fe có thể b/đổi hoá trị linh động, cho và nhận điện tử trong q/trình v/c điện tử. Khi ngộ độc HCN, CN- kết hợp với sắt tạo phức hợp (Fe+3-CN) bền vững K/năng tr/đổi điện tử của các cyt. bị trở ngại, gây ngạt từ mô bào.