
Parallel and “Antagonistic” Complementation Structures in the ...

Complementation of the Fo c Subunit of Escherichia coli with That ofStreptococcus mutans and Properties of the Hybrid FoF1 ATP Synthase
Makoto Araki,a Kazuya Hoshi,a Masasuke Fujiwara,b Yuka Sasaki,c Hideo Yonezawa,d Hidenobu Senpuku,d Atsuko Iwamoto-Kihara,c
Masatomo Maedaa
Department of Molecular Biology, School of Pharmacy, Iwate Medical University, Nishitokuta, Yahaba, Shiwa-gun, Iwate, Japana; Laboratory of Biochemistry and MolecularBiology, Graduate School of Pharmaceutical Sciences, Osaka University, Suita, Osaka, Japanb; Department of Bioscience, Nagahama Institute of Bio-Science andTechnology, Nagahama, Shiga, Japanc; Department of Bacteriology I, National Institute of Infectious Diseases, Shinjyuku, Tokyo, Japand
The c subunit of Streptococcus mutans ATP synthase (FoF1) is functionally exchangeable with that of Escherichia coli, sinceE. coli with a hybrid FoF1 is able to grow on minimum succinate medium through oxidative phosphorylation. E. coli F1
bound to the hybrid Fo with the S. mutans c subunit showed N,N=-dicyclohexylcarbodiimide-sensitive ATPase activity sim-ilar to that of E. coli FoF1. Thus, the S. mutans c subunit assembled into a functional Fo together with the E. coli a and bsubunits, forming a normal F1 binding site. Although the H� pathway should be functional, as was suggested by thegrowth on minimum succinate medium, ATP-driven H� transport could not be detected with inverted membrane vesiclesin vitro. This observation is partly explained by the presence of an acidic residue (Glu-20) in the first transmembrane helixof the S. mutans c subunit, since the site-directed mutant carrying Gln-20 partly recovered the ATP-driven H� transport.Since S. mutans is recognized to be a primary etiological agent of human dental caries and is one cause of bacterial endo-carditis, our system that expresses hybrid Fo with the S. mutans c subunit would be helpful to find antibiotics and chemi-cals specifically directed to S. mutans.
FoF1-ATP synthase catalyzes both the synthesis and hydroly-sis of ATP coupled to H� transport across membranes. This
enzyme is widely distributed from prokaryotes to eukaryotes. Itis localized in bacterial plasma membranes and mitochondrialinner and chloroplast thylakoid membranes. F1 has three cata-lytic sites, and Fo is an H�-transporting pathway. The synthesisand hydrolysis of ATP at catalytic sites are coupled to H� trans-port through Fo. Bacterial FoF1 is composed of membrane-peripheral F1 (�3�3��ε) and membrane-integral Fo (ab2c10)(1). The c subunit of Fo forms a hairpin structure in the mem-brane (2). It is further suggested that the linker region of the csubunit joining two transmembrane helices constitutes a partof the F1-binding site together with other Fo subunits (3). Theessential residue for H� transport is believed to be a conservedacidic residue (Glu or Asp) in the second transmembrane helixof the c subunit (4).
Recent mechanistic studies demonstrated that the bacterialFoF1 is a molecular motor, which can be dissected into the rotor(�, ε, and c) and the stator (�, �, �, a, and b) (5). According tothe rotation mechanism of ATP synthases, an H� gradientdrives rotation of the c subunit ring accompanying H� trans-port from the periplasm to the cytoplasm through the H� path-way composed of the a and c subunits. The � subunit on the csubunit ring concomitantly rotates together with the ring, in-ducing changes in the three catalytic sites and ATP synthesis.On the contrary, ATP hydrolysis at the catalytic sites inducesrotation of the � subunit together with the c subunit ring, re-sulting in the transport of H� from the cytoplasm to theperiplasm through the H� pathway.
S. mutans is implicated as the principal causative agent ofhuman dental caries, which is one of the most common infec-tious diseases (6). This bacterium is able to survive in the lowpH environment of dental caries, which is ascribed to the pres-ence of FoF1 (7). FoF1 of S. mutans is suggested to pump H�
back out of the cell and thereby protect acid-sensitive glycolyticenzymes (8). Furthermore, S. mutans is recognized to be asso-ciated with infective endocarditis (9). It is reported that thebinding site for diarylquinoline is the c subunit of Mycobacte-rium tuberculosis (10, 11). Although the conserved acidic resi-due in the transmembrane helix of the c subunit functions inH� transport, the surrounding residues differ among species(4). Thus, the c subunit of M. tuberculosis is suggested to be aspecific target of antituberculosis agents. These observationsprompted us to express S. mutans FoF1 and find specific inhib-itors of this bacterial FoF1.
In this study, we expressed the S. mutans c subunit in E. colitogether with other E. coli FoF1 subunits and characterized theresulting hybrid FoF1.
MATERIALS AND METHODSConstruction of expression plasmids for the S. mutans c subunit. S.mutans DNA (7) was extracted, and its entire ATP operon was cloned asdescribed for Fig. S1 and S2 in the supplemental material. The resultingplasmid containing the cloned ATP operon, pP5-2-SM-BWU13 (see Fig.S2A), did not produce active ATP synthase in E. coli DK-8, which is adeletion strain of the ATP operon (12). Thus, we attempted to substitutethe E. coli c subunit for the corresponding one of S. mutans, since this is thesmallest FoF1 subunit and can be easily manipulated.
The c subunit gene of E. coli was deleted from pBWU13 (13), as indi-
Received 17 May 2013 Accepted 17 August 2013
Published ahead of print 23 August 2013
Address correspondence to Masatomo Maeda, [email protected].
Supplemental material for this article may be found at http://dx.doi.org/10.1128/JB.00542-13.
Copyright © 2013, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JB.00542-13
November 2013 Volume 195 Number 21 Journal of Bacteriology p. 4873–4878 jb.asm.org 4873
on March 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

cated in Fig. S3A in the supplemental material. The small HindIII-SphIfragment was cloned into pUC18, and then SpeI and NgoMIV sites wereintroduced between the a/c and c/b intersubunit genes, respectively, usinga PrimeSTAR mutagenesis basal kit (TaKaRa) with primers mSpeI-F/mSpeI-R and mNgoMIV-F/mNgoMIV-R, respectively (see Table S1). Af-ter SpeI and NgoMIV digestion, the large SpeI-NgoMIV fragment wastreated with Klenow enzyme and self-ligated. The HindIII-SphI fragment,from which the SpeI-NgoMIV fragment carrying the c subunit gene hadbeen deleted, then was reintroduced into pBWU13. The resulting plasmidwas named pBWU13�c (see Fig. S4C).
The expression plasmids for c subunits were constructed as follows(see Fig. S3B in the supplemental material). The S. mutans c subunit genewith a promoter was amplified from pP5-2-SM-BWU13 by means of PCRusing primer pair P5-(c)SD-F and SMC-R (see Table S1). The PCR con-ditions comprised preheating (96°C, 1 min), followed by 35 cycles ofdenaturation (96°C, 10 s), annealing (55°C, 5 s), and extension (72°C, 30s). The amplified fragment was cloned into the HincII site of pMW218(GenBank accession number AB005477) (Nippon Gene), and the result-ing plasmid was named pSmcEE (expression plasmid for S. mutans c sub-unit with glutamic acids at positions 20 and 53) (see Fig. S4B). The E. colic subunit gene with the promoter was amplified from pBWU13 usingprimer pair mSpeI-R and ECC-R. The resulting expression plasmid wasnamed pEcc (see Fig. S4A).
Mutations (E20Q, E20A, E53Q, and E53A) of the S. mutans c subunit genewere introduced by using a PrimeSTAR mutagenesis basal kit together withpSmcEE and one of the primer pairs (SMC-mE20Q-F/R, SMC-mE20A-F/R,SMC-mE53Q-F/R, or SMC-mE53A-F/R) listed in Table S1 in the supple-mental material, and then the mutated genes were amplified with primer pairP5-(c)SD-F and SMC-R as described above. The expression plasmids werenamed pSmcQE, pSmcAE, pSmcEQ, and pSmcEA, respectively. Second mu-tations (E20Q�E53A and E20Q�E53Q) were introduced with pSmcQE andthe primer pairs SMC-mE53A-F/R and SMC-mE53Q-F/R, respectively. TheDNA sequence was confirmed by the dideoxy chain termination method(14). The molecular biological techniques used for DNA manipulations werebased on standard procedures (15).
Growth conditions for E. coli. A rich medium (L broth) and mini-mum medium containing thymine (50 �g/ml), thiamine (2 �g/ml), iso-leucine (50 �g/ml), valine (50 �g/ml), and a carbon source (0.5% glucoseor 0.5% succinate) were used (16). For preparation of membrane vesicles,0.5% glycerol was used as the sole carbon source. Ampicillin (100 �g/ml)(Wako) and kanamycin (10 �g/ml) (Wako) were added to L broth toselect cells harboring expression plasmid pBWU13 (and its derivatives)and pMW218 (and its derivatives), respectively. Both antibiotics wereadded together to select cells with both plasmids.
Assay procedures. Membrane vesicles were prepared after passingcells through a French press (FA-078; AMINCO) using an FA-031 cell at1,250 kg/cm2 pressure, and the ATPase activity was determined by mea-suring the release of inorganic phosphate colorimetrically (17). N,N=-dicyclohexylcarbodiimide (DCCD; Wako) was added as an ethanol solu-tion prepared as a 20 mM stock solution. Formation of an H� gradientwas assayed by measuring the fluorescence quenching of acridine orange(Sigma) (18). Protein concentrations were determined by means of theBio-Rad protein assay (19) using bovine serum albumin (fraction V;Sigma) as a standard.
Detection of the �-subunit. Protein samples were mixed with 5�sample buffer and then heat treated at 98°C for 5 min. They were sepa-rated by SDS-polyacrylamide gel electrophoresis (small gel [10 by 10 cm],1 mm thick, consisting of a 3% [wt/vol] stacking gel and a 9% [wt/vol]separation gel [20]) and subjected to Western blotting (21). Proteins wereelectroblotted (25 V, 3 h; ATTO model AE-8130) onto a Hybond-P poly-vinylidene difluoride (PVDF) membrane (GE Healthcare). Mouse mono-clonal antibodies recognizing the E. coli F1 � subunit (Mito Science) werereacted for 1 h as the first antibodies at �1,000 dilution in TBS-T (25 mMTris-HCl [pH 7.6], 150 mM NaCl containing 0.1% [wt/wt] Tween-20).The membrane was washed in TBS-T for 5 min (3 times). Horseradish
peroxidase-linked sheep anti-mouse Ig (�5,000 dilution) was used as thesecond antibody with ECL prime western blotting detection reagent (GEHealthcare). The � subunit was visualized with a LAS-3000 (Fuji Film).
Chemicals. Restriction enzymes and calf intestine phosphatase werepurchased from NEB. The DNA ligation kit (version 1) and PrimeSTARHS DNA polymerase were supplied by TaKaRa. The Klenow enzyme andagarose-LE classic type were provided by Toyobo and Nacalai, respec-tively. Oligonucleotides were purchased from Hokkaido System Science.All other chemicals used were of the highest grade commercially available.
RESULTSComplementation of the c subunit of E. coli with that of S. mu-tans. We initially attempted to express S. mutans ATP synthase inDK-8, an E. coli strain lacking ATP operon production (12). How-ever, the cloned ATP operon of S. mutans could not function inDK-8 (see Fig. S1 and S2 in the supplemental material). Thus, weintroduced pSmcEE, carrying the S. mutans c subunit gene, intoDK-8 harboring only pBWU13�c, which could not grow on suc-cinate plates but could grow on glucose plates. Interestingly, thetransformant could grow on succinate plates as well as on glucoseplates, similar to DK-8 harboring both pBWU13�c and pEcc (Fig.1A). These results suggest that the c subunit of S. mutans assemblestogether with the E. coli a and b subunits into Fo and complementsthe function of the E. coli c subunit of ATP synthase.
Growth behavior of E. coli with the hybrid FoF1. We nextexamined the growth behavior of E. coli with the hybrid FoF1 inliquid medium of neutral and acidic pH. Since the S. mutans csubunit has an additional acidic residue, Glu-20, together withH�-transporting Glu-53 in transmembrane helices (Fig. 1C), itseems likely that the growth of DK-8 with the hybrid FoF1 wouldrespond to the medium pH. DK-8 expressing E. coli FoF1 grew wellon glucose medium at pH 7.5 compared to growth at pH 5.5 (Fig.2A and C). Without the c subunit (�c) or FoF1 (DK-8), E. colicould not grow on succinate medium (Fig. 2C), and the growthbehavior on glucose medium was similar to that with E. coli FoF1
(Fig. 2A).The growth of E. coli with the hybrid FoF1 on glucose medium
was similar to that with E. coli FoF1 (Fig. 2A, closed and opencircles, respectively). On succinate medium, cells grew better atpH 5.5 (Fig. 2C, closed and open circles, respectively). These re-sults clearly indicate that E. coli with the hybrid FoF1 could growwell at acidic pH on succinate medium, although the growth curveat pH 5.5 was almost identical to that of cells with E. coli FoF1 at pH7.5 (Fig. 2C).
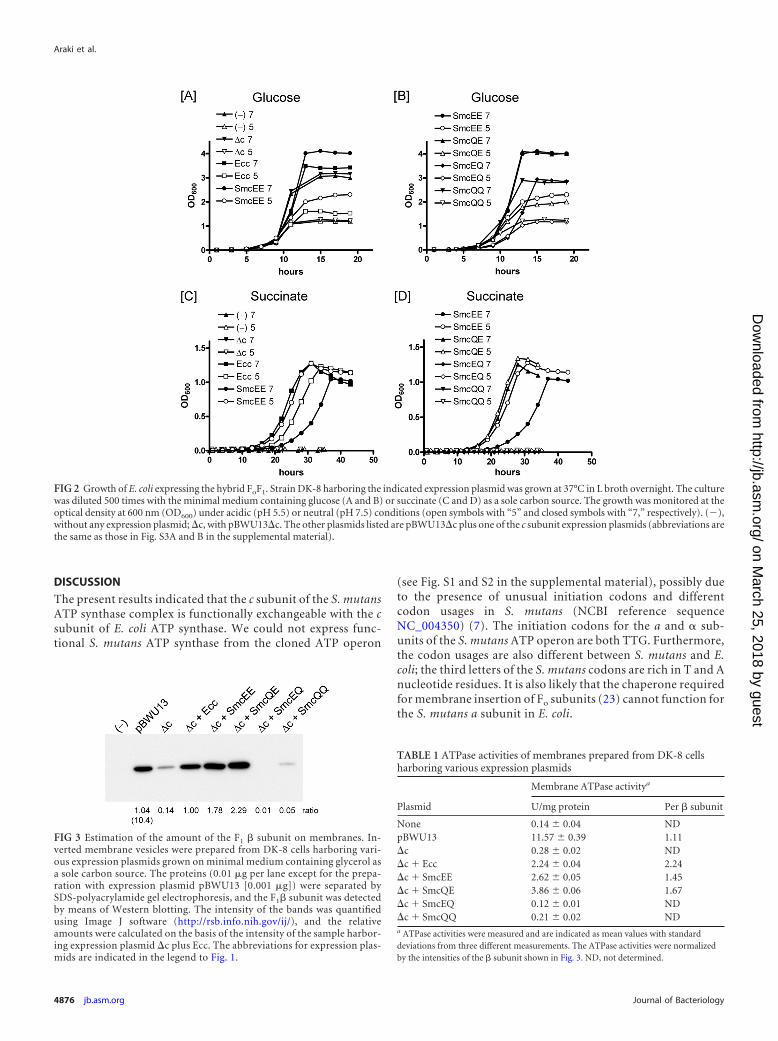
Detection of the � subunit on the membranes. The functionalsubstitution of the c subunits of E. coli and S. mutans caused us tobe interested in how chimeric Fo transports H� and affects thecatalytic activity of F1. We first determined F1 assembly on themembranes using an antibody for the � subunit. As shown in Fig.3, the � subunit could be detected in the positive control (lane 2).Without expression of the c subunit, a significantly small amountof the � subunit was recovered from the membrane (lane 3). Thenegative control without FoF1 did not show any background signal(lane 1). Upon expression of the E. coli c subunit, as much as a7-times higher level of the � subunit was detected on the mem-branes (lane 4) compared to that on the membranes without the csubunit (lane 3). Interestingly, much greater amounts of the �subunit were found with the S. mutans c subunit (lane 5). Theseresults indicate that the F1 binding site could be similarly formedwith the S. mutans c subunit together with the subunits a and b ofthe E. coli Fo.
Araki et al.
4874 jb.asm.org Journal of Bacteriology
on March 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

We further assayed the ATPase activity of F1 to confirm thefunctional assembly of F1 on Fo. The membrane F1-ATPase activ-ities of E. coli FoF1 and the hybrid FoF1 were ten and twenty timeshigher than those in negative controls without the c subunit orwithout FoF1, respectively (Table 1) The F1-ATPase activity ex-pressed from the E. coli ATP operon of pBWU13 was much higherthan that from the c subunit gene plus the rest of the subunit genesfrom pBWU13�c. When the ATPase activities per amount of �subunit were compared (Table 1, right), the calculated values forE. coli FoF1 and the hybrid enzyme (2.24 and 1.45, respectively)were not significantly different, although the value for the latter isslightly lower.
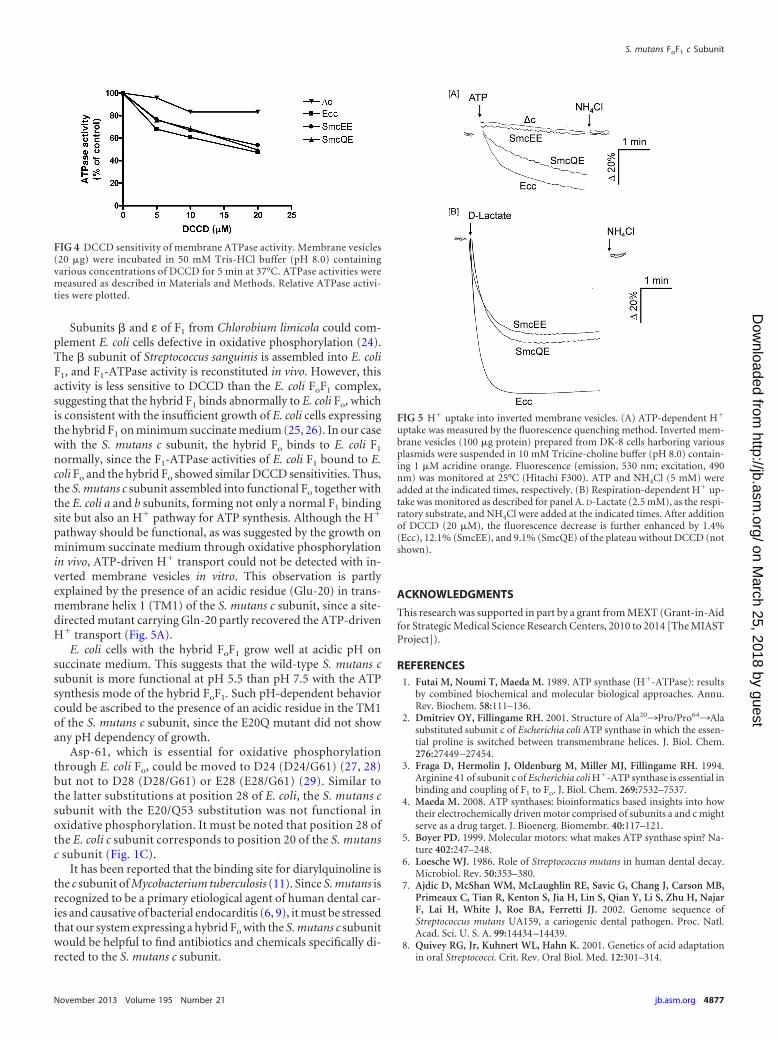
DCCD binding to an H�-transporting acidic residue of theFo c subunit affects the catalytic activity of F1 through a stalkregion (22). As expected from the functional substitution of the
S. mutans c subunit, the F1-ATPase activity of the hybrid FoF1
on membranes was DCCD sensitive, similar to that of E. coliFoF1 (Fig. 4). At a concentration of 20 �M DCCD, 50% of theactivities were inhibited, whereas the lower level of ATPaseactivity without the c subunit was not sensitive to DCCD. Theseresults suggest that F1 on membranes interacted normally withthe hybrid Fo composed of the S. mutans c subunit and the E.coli a and b subunits.
H� transport activities of inverted membrane vesicles. It wasinteresting to determine whether or not the hybrid FoF1 couldtransport H� coupled with ATP hydrolysis, since F1 interacts withFo with the S. mutans c subunit normally. The ability of the hybridFoF1 to catalyze in vivo oxidative phosphorylation in succinatemedium strongly indicated that H� transport through Fo fromoutside to inside the plasma membrane is coupled with ATP syn-thesis at the catalytic sites of F1. Although ATP-dependent H�
transport was detectable using membrane vesicles with E. coliFoF1, the activity could not be detected with the hybrid FoF1 (Fig.5A). This negative result was not due to the H�-related leakinessof the membranes, since a respiratory chain-dependent H� gra-dient was formed in the presence of D-lactate (Fig. 5B) and theformation of the gradient was only slightly increased in the pres-ence of DCCD (Fig. 5). The H� transport of E. coli FoF1 decreasedwith reduction of the pH of the assay buffer from 8.0 to 5.5. Fur-thermore, H� transport of the hybrid FoF1 could not be detectedin this pH range (data not shown).
Properties of the hybrid FoF1 with the mutated S. mutans csubunit. The properties of the hybrid FoF1 could be due to thepresence of an acidic residue in the first transmembrane helix (Fig.1C). Thus, we mutated Glu-20 of the S. mutans c subunit toGln-20 or Ala-20 (E20Q [SmcQE] and E20A [SmcAE] mutants,respectively) and expressed it instead of the wild-type S. mutans csubunit. Both mutants grew on succinate plates (Fig. 1B). TheE20Q mutant grew as well on succinate plates as the wild-type S.mutans c subunit, unlike the E20A mutant. The difference ingrowth between neutral and acidic pH in the liquid succinate me-dium observed with the hybrid enzyme containing the wild-typeS. mutans c subunit was almost completely abolished with theE20Q mutation (Fig. 2D, closed and open triangles, respectively).The growth behavior on glucose medium was similar for the hy-brid FoF1 with the wild-type (SmcEE) and mutant (SmcQE) csubunits (Fig. 2B, closed and open circles and closed and opentriangles, respectively). The membrane ATPase activity andDCCD sensitivity of the hybrid FoF1 with SmcEE and SmcQE wereessentially the same (Table 1 and Fig. 4). Interestingly, ATP-de-pendent H� transport into the membranes was detectable withthe hybrid FoF1 with the E20Q mutation (Fig. 5A).
We further replaced the conserved amino acid residue (Glu-53) with Gln or Ala together with the Glu-20 residue (E53Q, E53A,E20Q�E53A, E20A�E53Q, E20Q�E53Q, and E20A�E53A).None of these mutant c subunits was active in in vivo oxidativephosphorylation, since E. coli cells expressing either of these csubunits could not grow on succinate plates or liquid medium(Fig. 1B and 2D). Furthermore, the F1 did not assemble on themembranes of the cells with E53Q and E20Q�E53Q mutations(Fig. 3, lanes 7 and 8). Thus, the Glu-53 residue of the S. mutans csubunit is essential for the formation of functional FoF1 as well asH� transport. Further, it is interesting to know whether the mu-tated c subunits are assembled in Fo or not.
FIG 1 Complementation of the S. mutans ATP synthase c subunit with thatof E. coli. (A) A c subunit expression plasmid (pSmcEE or pEcc) and a �cplasmid (pBWU13�c) were introduced into E. coli strain DK-8, fromwhich all ATP synthase subunit genes had been deleted. Transformantswere cultured on agar plates with the minimal medium containing a carbonsource (glucose or succinate) at 37°C. No. 1, negative control without anyexpression plasmids; no. 2, positive control with pBWU13; no. 3, �c(pBWU13�c); no. 4, Ecc (expression plasmid for E. coli c subunit); no. 5,�c and Ecc; no. 6, SmcEE (expression plasmid for S. mutans c subunit withglutamic acids at positions 20 and 53, respectively); no. 7, �c and SmcEE.(B) Effects of the mutations on the S. mutans c subunit were determined.Two glutamic acid residues (SmcEE) in transmembrane helices of the S.mutans c subunit were replaced with glutamine or alanine. Growth of themutants was examined as described for panel A. No. 1, negative controlwithout any expression plasmids; no. 2, positive control with pBWU13; no.3, �c; no. 4, Ecc; no. 5, �c and Ecc; no. 6, �c and SmcEE; no. 7, �c andSmcQE; no. 8, �c and SmcAE; no. 9, �c and SmcEQ; no. 10, �c and SmcEA;no. 11, �c and SmcAQ; no. 12, �c and SmcQA; no. 13, �c and SmcQQ; no.14, �c and SmcAA. (C) The primary sequences for various bacterial Fo csubunits were aligned. Two transmembrane helices are underlined and aredenoted as TM1 and TM2. The essential acidic residues for H� transportare indicated by the asterisk. Amino acid residues are denoted by singleletters, and the residue numbers are also shown. The sequences of S. mu-tans, E. coli, S. sanguinis, and S. salivarius are from Entrez Gene numbers1028775, 948253, 4806586, and 10972708, respectively.
S. mutans FoF1 c Subunit
November 2013 Volume 195 Number 21 jb.asm.org 4875
on March 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
DISCUSSION
The present results indicated that the c subunit of the S. mutansATP synthase complex is functionally exchangeable with the csubunit of E. coli ATP synthase. We could not express func-tional S. mutans ATP synthase from the cloned ATP operon
(see Fig. S1 and S2 in the supplemental material), possibly dueto the presence of unusual initiation codons and differentcodon usages in S. mutans (NCBI reference sequenceNC_004350) (7). The initiation codons for the a and � sub-units of the S. mutans ATP operon are both TTG. Furthermore,the codon usages are also different between S. mutans and E.coli; the third letters of the S. mutans codons are rich in T and Anucleotide residues. It is also likely that the chaperone requiredfor membrane insertion of Fo subunits (23) cannot function forthe S. mutans a subunit in E. coli.
FIG 2 Growth of E. coli expressing the hybrid FoF1. Strain DK-8 harboring the indicated expression plasmid was grown at 37°C in L broth overnight. The culturewas diluted 500 times with the minimal medium containing glucose (A and B) or succinate (C and D) as a sole carbon source. The growth was monitored at theoptical density at 600 nm (OD600) under acidic (pH 5.5) or neutral (pH 7.5) conditions (open symbols with “5” and closed symbols with “7,” respectively). (),without any expression plasmid; �c, with pBWU13�c. The other plasmids listed are pBWU13�c plus one of the c subunit expression plasmids (abbreviations arethe same as those in Fig. S3A and B in the supplemental material).
FIG 3 Estimation of the amount of the F1 � subunit on membranes. In-verted membrane vesicles were prepared from DK-8 cells harboring vari-ous expression plasmids grown on minimal medium containing glycerol asa sole carbon source. The proteins (0.01 �g per lane except for the prepa-ration with expression plasmid pBWU13 [0.001 �g]) were separated bySDS-polyacrylamide gel electrophoresis, and the F1� subunit was detectedby means of Western blotting. The intensity of the bands was quantifiedusing Image J software (http://rsb.info.nih.gov/ij/), and the relativeamounts were calculated on the basis of the intensity of the sample harbor-ing expression plasmid �c plus Ecc. The abbreviations for expression plas-mids are indicated in the legend to Fig. 1.
TABLE 1 ATPase activities of membranes prepared from DK-8 cellsharboring various expression plasmids
Plasmid
Membrane ATPase activitya
U/mg protein Per � subunit
None 0.14 0.04 NDpBWU13 11.57 0.39 1.11�c 0.28 0.02 ND�c � Ecc 2.24 0.04 2.24�c � SmcEE 2.62 0.05 1.45�c � SmcQE 3.86 0.06 1.67�c � SmcEQ 0.12 0.01 ND�c � SmcQQ 0.21 0.02 NDa ATPase activities were measured and are indicated as mean values with standarddeviations from three different measurements. The ATPase activities were normalizedby the intensities of the � subunit shown in Fig. 3. ND, not determined.
Araki et al.
4876 jb.asm.org Journal of Bacteriology
on March 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
Subunits � and ε of F1 from Chlorobium limicola could com-plement E. coli cells defective in oxidative phosphorylation (24).The � subunit of Streptococcus sanguinis is assembled into E. coliF1, and F1-ATPase activity is reconstituted in vivo. However, thisactivity is less sensitive to DCCD than the E. coli FoF1 complex,suggesting that the hybrid F1 binds abnormally to E. coli Fo, whichis consistent with the insufficient growth of E. coli cells expressingthe hybrid F1 on minimum succinate medium (25, 26). In our casewith the S. mutans c subunit, the hybrid Fo binds to E. coli F1
normally, since the F1-ATPase activities of E. coli F1 bound to E.coli Fo and the hybrid Fo showed similar DCCD sensitivities. Thus,the S. mutans c subunit assembled into functional Fo together withthe E. coli a and b subunits, forming not only a normal F1 bindingsite but also an H� pathway for ATP synthesis. Although the H�
pathway should be functional, as was suggested by the growth onminimum succinate medium through oxidative phosphorylationin vivo, ATP-driven H� transport could not be detected with in-verted membrane vesicles in vitro. This observation is partlyexplained by the presence of an acidic residue (Glu-20) in trans-membrane helix 1 (TM1) of the S. mutans c subunit, since a site-directed mutant carrying Gln-20 partly recovered the ATP-drivenH� transport (Fig. 5A).
E. coli cells with the hybrid FoF1 grow well at acidic pH onsuccinate medium. This suggests that the wild-type S. mutans csubunit is more functional at pH 5.5 than pH 7.5 with the ATPsynthesis mode of the hybrid FoF1. Such pH-dependent behaviorcould be ascribed to the presence of an acidic residue in the TM1of the S. mutans c subunit, since the E20Q mutant did not showany pH dependency of growth.
Asp-61, which is essential for oxidative phosphorylationthrough E. coli Fo, could be moved to D24 (D24/G61) (27, 28)but not to D28 (D28/G61) or E28 (E28/G61) (29). Similar tothe latter substitutions at position 28 of E. coli, the S. mutans csubunit with the E20/Q53 substitution was not functional inoxidative phosphorylation. It must be noted that position 28 ofthe E. coli c subunit corresponds to position 20 of the S. mutansc subunit (Fig. 1C).
It has been reported that the binding site for diarylquinoline isthe c subunit of Mycobacterium tuberculosis (11). Since S. mutans isrecognized to be a primary etiological agent of human dental car-ies and causative of bacterial endocarditis (6, 9), it must be stressedthat our system expressing a hybrid Fo with the S. mutans c subunitwould be helpful to find antibiotics and chemicals specifically di-rected to the S. mutans c subunit.
ACKNOWLEDGMENTS
This research was supported in part by a grant from MEXT (Grant-in-Aidfor Strategic Medical Science Research Centers, 2010 to 2014 [The MIASTProject]).
REFERENCES1. Futai M, Noumi T, Maeda M. 1989. ATP synthase (H�-ATPase): results
by combined biochemical and molecular biological approaches. Annu.Rev. Biochem. 58:111–136.
2. Dmitriev OY, Fillingame RH. 2001. Structure of Ala20¡Pro/Pro64¡Alasubstituted subunit c of Escherichia coli ATP synthase in which the essen-tial proline is switched between transmembrane helices. J. Biol. Chem.276:27449 –27454.
3. Fraga D, Hermolin J, Oldenburg M, Miller MJ, Fillingame RH. 1994.Arginine 41 of subunit c of Escherichia coli H�-ATP synthase is essential inbinding and coupling of F1 to Fo. J. Biol. Chem. 269:7532–7537.
4. Maeda M. 2008. ATP synthases: bioinformatics based insights into howtheir electrochemically driven motor comprised of subunits a and c mightserve as a drug target. J. Bioenerg. Biomembr. 40:117–121.
5. Boyer PD. 1999. Molecular motors: what makes ATP synthase spin? Na-ture 402:247–248.
6. Loesche WJ. 1986. Role of Streptococcus mutans in human dental decay.Microbiol. Rev. 50:353–380.
7. Ajdic D, McShan WM, McLaughlin RE, Savic G, Chang J, Carson MB,Primeaux C, Tian R, Kenton S, Jia H, Lin S, Qian Y, Li S, Zhu H, NajarF, Lai H, White J, Roe BA, Ferretti JJ. 2002. Genome sequence ofStreptococcus mutans UA159, a cariogenic dental pathogen. Proc. Natl.Acad. Sci. U. S. A. 99:14434 –14439.
8. Quivey RG, Jr, Kuhnert WL, Hahn K. 2001. Genetics of acid adaptationin oral Streptococci. Crit. Rev. Oral Biol. Med. 12:301–314.
FIG 5 H� uptake into inverted membrane vesicles. (A) ATP-dependent H�
uptake was measured by the fluorescence quenching method. Inverted mem-brane vesicles (100 �g protein) prepared from DK-8 cells harboring variousplasmids were suspended in 10 mM Tricine-choline buffer (pH 8.0) contain-ing 1 �M acridine orange. Fluorescence (emission, 530 nm; excitation, 490nm) was monitored at 25°C (Hitachi F300). ATP and NH4Cl (5 mM) wereadded at the indicated times, respectively. (B) Respiration-dependent H� up-take was monitored as described for panel A. D-Lactate (2.5 mM), as the respi-ratory substrate, and NH4Cl were added at the indicated times. After additionof DCCD (20 �M), the fluorescence decrease is further enhanced by 1.4%(Ecc), 12.1% (SmcEE), and 9.1% (SmcQE) of the plateau without DCCD (notshown).
FIG 4 DCCD sensitivity of membrane ATPase activity. Membrane vesicles(20 �g) were incubated in 50 mM Tris-HCl buffer (pH 8.0) containingvarious concentrations of DCCD for 5 min at 37°C. ATPase activities weremeasured as described in Materials and Methods. Relative ATPase activi-ties were plotted.
S. mutans FoF1 c Subunit
November 2013 Volume 195 Number 21 jb.asm.org 4877
on March 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

9. Nomura R, Nakano K, Nemoto H, Fujita K, Inagaki S, Takahashi T,Taniguchi K, Takeda M, Yoshioka H, Amano A, Ooshima T. 2006.Isolation and characterization of Streptococcus mutans in heart valve anddental plaque specimens from a patient with infective endocarditis. J.Med. Microbiol. 55:1135–1140.
10. Andries K, Verhasselt P, Guillemont J, Göhlmann HWH, Neefs J-M,Winkler H, Gestel JV, Timmerman P, Zhu M, Lee E, Williams P, deChaffoy D, Huitric E, Hoffner S, Cambau E, Truffot-Pernot C, LounisN, Jarlier V. 2005. A diarylquinoline drug active on the ATP synthase ofMycobacterium tuberculosis. Science 307:223–227.
11. Koul A, Dendouga N, Vergauwen K, Molenberghs B, Vranskx L, Wil-lebrords R, Ristic Z, Lill H, Dorange I, Guillemont J, Bald D, AndriesK. 2007. Diarylquinolines target subunit c of mycobacterial ATP synthase.Nat. Chem. Biol. 3:323–324.
12. Klionsky DJ, Brusilow WSA, Simoni RD. 1984. In vivo evidence for therole of the epsilon subunit as an inhibitor of the proton-translocatingATPase of Escherichia coli. J. Bacteriol. 160:1055–1060.
13. Iwamoto A, Omote H, Hanada H, Tomioka N, Itai A, Maeda M, FutaiM. 1991. Mutation in Ser174 and the glycine-rich sequence (Gly149, Gly150,and Thr156) in the � subunit of Escherichia coli H�-ATPase. J. Biol. Chem.266:16350 –16355.
14. Sanger F, Coulson AR, Barrell BG, Smith AJH, Roe BA. 1980. Cloningin single-stranded bacteriophage as an aid to rapid DNA sequencing. J.Mol. Biol. 143:161–178.
15. Sambrook J, Fritsch EF, Maniatis T. 1989. Molecular cloning: a labora-tory manual, 2nd ed. Cold Spring Harbor Laboratory Press, Cold SpringHarbor, NY.
16. Takeyama M, Ihara K, Moriyama Y, Noumi T, Ida K, Tomioka N, ItaiA, Maeda M, Futai M. 1990. The glycine-rich sequence of the beta sub-unit of Escherichia coli H�-ATPase is important for activity. J. Biol. Chem.265:21279 –21284.
17. Futai M, Sternweiss PC, Heppel LA. 1974. Purification and properties ofreconstitutively active and inactive adenosinetriphosphatase from Esche-richia coli. Proc. Natl. Acad. Sci. U. S. A. 71:2725–2729.
18. Moriyama Y, Iwamoto A, Hanada H, Maeda M, Futai M. 1991. One-step purification of Escherichia coli H�-ATPase (FoF1) and its reconstitu-
tion into liposomes with neurotransmitter transporters. J. Biol. Chem.266:22141–22146.
19. Bradford MM. 1976. A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein-dyebinding. Anal. Biochem. 72:248 –254.
20. Laemmli UK. 1970. Cleavage of structural proteins during the assembly ofthe bacteriophage T4. Nature 227:680 – 685.
21. Towbin H, Staehelin T, Gordon J. 1979. Electrophoretic transfer ofproteins from polyacrylamide gels to nitrocellulose sheets: procedure andsome applications. Proc. Natl. Acad. Sci. U. S. A. 76:4350 – 4354.
22. Girvin ME, Fillingame RH. 1994. Hairpin folding of subunit c of F1Fo
ATP synthase: 1H distance measurements to nitroxide-derivatized aspar-tyl-61. Biochemistry 33:665– 674.
23. Yi L, Jiang F, Chen M, Cain B, Bolhuis A, Dalbey RE. 2003. YidC isstrictly required for membrane insertion of subunits a and c of theF1FoATP synthase and SecE of the SecYEG translocase. Biochemistry 42:10537–10544.
24. Xie D-L, Lill H, Hauska G, Maeda M, Futai M, Nelson N. 1993. The atp2operon of the green bacterium. Chlorobium limicola. Biochim. Biophys.Acta 1172:267–273.
25. Kuhnert WL, Quivey RG, Jr., 2003. Genetic and biochemical character-ization of the F-ATPase operon from Streptococcus sanguis 10904. J. Bac-teriol. 185:1525–1533.
26. Trüper HG, De’Clari L. 1997. Necessary correction of specific epithetsformed as substantives (nouns) “in apposition.” Int. J. Syst. Bacteriol.47:908 –909.
27. Zhang Y, Fillingame RH. 1994. Essential aspartate in subunit c of F1Fo
ATP syntase: effect of position 61 substitutions in helix-2 on function ofAsp24 in helix-1. J. Biol. Chem. 269:5473–5479.
28. Miller JH, Oldenburg M, Fillingame RH. 1990. The essential carboxylgroup in subunit c of the F1F0 ATP synthase can be moved and H(�)-translocating function retained. Proc. Natl. Acad. Sci. U. S. A. 87:4900 –4904.
29. Jones PC. 2001. Introduction of a carboxyl group in the first transmem-brane helix of Escherichia coli F1Fo ATPase subunit c and cytoplasmic pHregulation. J. Bacteriol. 183:1524 –1530.
Araki et al.
4878 jb.asm.org Journal of Bacteriology
on March 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from


















