
Tip60 is regulated by circadian transcription factor Clock ... · Tip60 is regulated by circadian...
27
Tip60 is regulated by circadian transcription factor Clock and is involved in cisplatin resistance Naoya Miyamoto 1,2 , Hiroto Izumi 1 , Takako Noguchi 3 , Yoshihiro Nakajima 3 , Yoshihiro Ohmiya 3,4 , Masaki Shiota 1 , Akihiko Kidani 1 , Akihiko Tawara 2 and Kimitoshi Kohno 1 Department of 1 Molecular Biology and 2 Ophthalmology, School of Medicine, University of Occupational and Environmental Health, Kitakyushu, 807-8555 3 National Institute of Advanced Industrial Science and Technology, Osaka, 563-8577 4 Department of Photobiology, Hokkaido University Graduate School of Medicine, Sapporo, 060-8633, Japan Running title: Histone acetyltransferase and cisplatin resistance Correspondence: Kimitoshi Kohno M.D., Ph.D., Department of Molecular Biology, School of Medicine, University of Occupational and Environmental Health, Japan. Tel: +81-93-691-7423; Fax: +81-93-692-2766; E-mail: [email protected] Histone modification is important for maintaining chromatin structure and function. Recently, histone acetylation has been shown to have a critical regulatory role in both transcription and DNA repair. We report here that expression of HAT genes is associated with cisplatin resistance. We found that Tip60 is overexpressed in cisplatin-resistant cells. The expression of two other HAT genes, HAT1 and MYST1, did not differ between drug-sensitive and -resistant cells. Knock-down of Tip60 expression rendered cells sensitive to cisplatin but not to oxaliplatin, vincristine and etoposide. Tip60 expression is significantly correlated with cisplatin sensitivity in human lung cancer cell lines. Interestingly, the promoter region of the Tip60 gene contains several E boxes, and its expression was regulated by the E-box-binding circadian transcription factor Clock, but not by other E-box-binding transcription factors such as c-Myc, Twist and USF1. Hyperacetylation of H3K14 and H4K16 was found in cisplatin-resistant cells. The microarray study reveals that several genes for DNA repair are downregulated by the knockdown of Tip60 expression. Our data show that HAT gene expression is required for cisplatin resistance and suggest that Clock and Tip60 regulate not only transcription, but also DNA repair, through periodic histone acetylation. INTRODUCTION Our research has focused on factors affecting cellular sensitivity of solid tumors to anticancer agents, and investigation of promising molecular targets for cancer treatment (1, 2). Among many drugs, cis-diamminechloroplatinum (II) (cisplatin) plays a crucial role in the treatment of various solid tumors (3, 4). Cisplatin has 1 http://www.jbc.org/cgi/doi/10.1074/jbc.M802332200 The latest version is at JBC Papers in Press. Published on May 5, 2008 as Manuscript M802332200 Copyright 2008 by The American Society for Biochemistry and Molecular Biology, Inc. by guest on June 30, 2018 http://www.jbc.org/ Downloaded from
Transcript of Tip60 is regulated by circadian transcription factor Clock ... · Tip60 is regulated by circadian...

Tip60 is regulated by circadian transcription factor Clock and is involved in cisplatin resistance
Naoya Miyamoto1,2, Hiroto Izumi1, Takako Noguchi3, Yoshihiro Nakajima3, Yoshihiro Ohmiya3,4, Masaki Shiota1, Akihiko Kidani1, Akihiko Tawara2 and Kimitoshi Kohno1
Department of 1Molecular Biology and 2Ophthalmology, School of Medicine, University of Occupational and Environmental Health, Kitakyushu, 807-8555
3National Institute of Advanced Industrial Science and Technology, Osaka, 563-8577 4Department of Photobiology, Hokkaido University Graduate School of Medicine, Sapporo,
060-8633, Japan Running title: Histone acetyltransferase and cisplatin resistance Correspondence: Kimitoshi Kohno M.D., Ph.D., Department of Molecular Biology, School of Medicine, University of Occupational and Environmental Health, Japan. Tel: +81-93-691-7423; Fax: +81-93-692-2766; E-mail: [email protected]
Histone modification is important for maintaining chromatin structure and function. Recently, histone acetylation has been shown to have a critical regulatory role in both transcription and DNA repair. We report here that expression of HAT genes is associated with cisplatin resistance. We found that Tip60 is overexpressed in cisplatin-resistant cells. The expression of two other HAT genes, HAT1 and MYST1, did not differ between drug-sensitive and -resistant cells. Knock-down of Tip60 expression rendered cells sensitive to cisplatin but not to oxaliplatin, vincristine and etoposide. Tip60 expression is significantly correlated with cisplatin sensitivity in human lung cancer cell lines. Interestingly, the promoter region of the Tip60 gene contains several E boxes, and its expression was regulated by the E-box-binding circadian transcription factor Clock, but not by other
E-box-binding transcription factors such as c-Myc, Twist and USF1. Hyperacetylation of H3K14 and H4K16 was found in cisplatin-resistant cells. The microarray study reveals that several genes for DNA repair are downregulated by the knockdown of Tip60 expression. Our data show that HAT gene expression is required for cisplatin resistance and suggest that Clock and Tip60 regulate not only transcription, but also DNA repair, through periodic histone acetylation.
INTRODUCTION Our research has focused on factors
affecting cellular sensitivity of solid tumors to anticancer agents, and investigation of promising molecular targets for cancer treatment (1, 2). Among many drugs, cis-diamminechloroplatinum (II) (cisplatin) plays a crucial role in the treatment of various solid tumors (3, 4). Cisplatin has
1
http://www.jbc.org/cgi/doi/10.1074/jbc.M802332200The latest version is at JBC Papers in Press. Published on May 5, 2008 as Manuscript M802332200
Copyright 2008 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest on June 30, 2018http://w
ww
.jbc.org/D
ownloaded from

been shown to form a cross-link between adjacent purines in genomic DNA, and can cause DNA-damaging signals to induce apoptosis (1, 2). Cisplatin treatment also induces oxidative and endoplasmic reticulum stresses (5). Thus, the nature of cellular sensitivity to cisplatin is highly complex.
The development of cisplatin resistance is a major clinical limitation in cancer chemotherapy. Cisplatin resistance is influenced by many factors, which affect intracellular drug accumulation (6-8), increased activity of intracellular thiol production (9, 10) and DNA repair (11, 12). However, little is known about the molecular mechanisms involved in drug resistance. Cellular factors involved in transcription contribute to the induction of apoptosis, or transient or acquired resistance. We have tried to identify the cisplatin-inducible transcription factors and transcription-related factors which are highly expressed in drug-resistant cells (1, 2, 13). We have previously shown that expression of activating transcription factor 4 (ATF4) is inducible by cisplatin treatment, and is high in cisplatin-resistant cells (14). The cellular level of ATF4 expression correlates with cisplatin sensitivity in human lung cancer cell lines. DNA microarray analysis has revealed that ATF4 regulates genes involved in glutathione biosynthesis and conjugation (15). Interestingly, we have found that ATF4 gene expression is regulated by the circadian transcription factor Clock, which is also overexpressed in cisplatin-resistant cells (16).
It has recently been shown that histones are periodically acetylated (17) and that Clock is a member of the histone acetyltransferases (18). Clock seems to have a highly specific emzymatic activity and acetylates histone 3 and histone 4 (19). Acetylation of histone 4 has been shown to be required for repair of DNA double-strand breaks. The yeast HAT Esa1 mutant is defective for NHEJ (non-homologous end joining) and is markedly hypersensitive to DNA-damaging agents, such as camptothecin-11 and methylmethanesulphonate (20). The acetyl-CoA binding motif is found in Clock and shows high sequence similarity to yeast Esa1 and other MYST members. One of the MYST family members, Tip60, has been shown to be involved in DNA repair and apoptosis (21). Through our studies seeking to determine the role of HAT in cisplatin resistance, we found that Tip60, but not HAT1 and MYST1, was overexpressed in cisplatin-resistant cells. Silencing of Tip60 expression effectively sensitized both parental and resistant cells to cisplatin, but not to oxaliplatin, vincristine and etoposide. These results can open new aspect for cisplatin resistance.
EXPERIMENTAL PROCEDURES Cell culture.
Human epidermoid cancer KB cells and human prostate cancer PC3 cells were cultured in Eagle’s Minimal Essential Medium. This medium was purchased from
2
by guest on June 30, 2018http://w
ww
.jbc.org/D
ownloaded from

Nissui Seiyaku (Tokyo, Japan) and contained 10% fetal bovine serum. The cisplatin-resistant KB/CP4 and P/CDP6 cells were derived from KB and PC3 cells as described previously (13), and found to be 23–63-fold more resistant to cisplatin than their parental cells (6). Eleven lung cancer cell lines were cultured as described previously (14). Cell lines were maintained in
a 5% CO2 atmosphere at 37°C.
Antibodies and drugs. Antibodies against c-Myc (sc-764),
Clock (sc-6927, sc-25361X), USF1 (sc-8983), Tip60 (sc-5727), PCNA (sc-56), HAT1 (sc-8752), MYST1 (sc-13677), Twist (sc-15393), H4K16 (sc-8662), and ERCC1 (sc-10785) were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Anti-H3K9 (31388) and anti-H3K14 (30020) antibodies were purchased from Upstate (Charlottesville, VA, USA). Anti-H2AK5 antibody (#2576), anti-APE1 antibody (#4128) was purchased from Cell Signaling (Danvers, MA, USA). Anti-β-actin antibody (AC-15) was purchased from Sigma. Cisplatin, vincristine and etoposide were purchased from Sigma (St. Louis, MO, USA). Oxaliplatin was kindly provided from Yakult (Tokyo, Japan).
Plasmid construction.
To obtain the full-length complementary DNA (cDNAs) for human Clock and human Bmal1, PCR was carried out on a SuperScript cDNA library
(Invitrogen Life Technologies, CA, USA) using the following primer pairs (double underlining indicates the start codons): ATGTTGTTTACCGTAAGCTGTAG and CTACTGTGGTTGAACCTTGGAAG for clock; and ATGGCAGACCAGAGAATGGAC and TTACAGCGGCCATGGCAAGTC for Bmal1. These PCR products were cloned into the pGEM-T easy vector (Promega, Madison, WI). To construct a pcDNA3-Bmal1, the EcoRI fragment including Bmal1 cDNA was ligated into a pcDNA3 vector. (Invitrogen). To construct a plasmid expressing 3xFlag-HA tagged Clock, pcDNA3-Flag vector was prepared by ligation of BamHI-EcoRI fragment including 3 times repeat Flag and HA into a pcDNA3 vector (Invitrogen). pcDNA3-3xFlag-HA Clock was obtained by ligation of NotI fragment including Clock cDNA into a pcDNA3-3xFlag-HA vector. The core promoter and partial first exon (–63 to +233) of the wild-type Tip60 gene was amplified by PCR using placenta DNA and the following primer pair: 5’-AGATCTCACGTGACCCGCTCCGCATACACGTG-3’ and 5’-AAGCTTCCGGCCCCTCTGGGAGACGTCAC -3’. PCR was also performed to introduce the E-box mutations into the Tip60 promoter, using the following primer pairs: 5’-AGATCTAAAGTGACCCGCTCCGCATAAAAGTG -3’ and 5’-AAGCTTCCGGCCCCTCTGGGAGACGTCAC’ -3’ for MT. Single and double underlined nucleotides indicate wild-type
3
by guest on June 30, 2018http://w
ww
.jbc.org/D
ownloaded from

E-boxes and mutated E-boxes, respectively. To prepare Tip60-WT-Luc containing wild-type E-boxes and Tip60-MT-Luc containg mutated E-boxes, these PCR products were cloned and ligated into the BglII-HindIII sites of the pGL3-basic vector (Promega).
Western blot analysis. Preparation of whole-cell lysates
and whole-nuclear lysates have been described previously (16, 22). Briefly, to prepare whole-nuclear lysates, isolated nuclei were directly sonicated for 10 s. The indicated amounts of whole-cell lysates or whole-nuclear lysates were separated by SDS-PAGE (10 or 15% gel). The proteins were transferred to PVDF (polyvinylidenedifluoride) microporous membranes (Millipore, Bedford, MA, USA) using a semi-dry blotter. The blotted membranes were treated with 5% (w/v) skimmed milk in 10 mM Tris, 150 mM NaCl, 0.2% (v/v) Tween 20, and incubated for 2 h at room temperature with a 1:5000 dilution of anti-Clock (sc-6927), USF1, c-Myc, and β-actin antibodies; a 1:2500 dilution of anti-Tip60, HAT1, MYST1, H2AK5 and H4K16 antibodies; a 1:2000 dilution of anti-PCNA, H3K9, and H3K14 antibodies; a 1:1000 dilution of anti-ERCC1 antibody; and a 1:500 dilution of anti-Twist, and APE1 antibodies. The membrane was then incubated with a peroxidase-conjugated secondary antibody for 40 min at room temperature, and was developed using a
chemiluminescence protocol (Amersham Biosciences, Piscataway, NJ, USA). Gel staining with Coomassie brilliant blue (CBB) was used as a internal control.
Transient transfection and luciferase assay. Transient transfection and a
luciferase assay were performed as described previously (16, 22). The indicated amounts of Tip60 reporter plasmid and the expression plasmid were co-transfected using Superfect reagent (Qiagen, Hilden, Germany), according to the manufacturer’s instructions. After transfection, the cells were cultured for 48 h. Luciferase activity was detected by a Picagene kit (Toyoinki, Tokyo, Japan), and the light intensity was measured with a luminometer (Luminescencer JNII RAB-2300; ATTO, Japan), according to the manufacturer’s instructions. The results shown are normalized to β-galactosidase activity, and are representative of at least three independent experiments.
Knockdown analysis using small interfering RNAs.
The following double-stranded RNA 25-base-pair oligonucleotides were commercially generated (Invitrogen Life Technologies, Carlsbad, CA, USA): Clock small interfering RNA (siRNA), 5’-UAAAGUCUGUUGUUGUAUCAUGUGC-3’ (sense) and 5’-GCACAUGAUACAACAACAGACUUUA-3’ (antisense); Tip60 siRNA,
5’-
4
by guest on June 30, 2018http://w
ww
.jbc.org/D
ownloaded from

AUAGUACAGUGUCUUAUGGUCAAGG -3’ (sense) and 5’--CCUUGACCAUAAGACACUGUACUAU -3’ (antisense); HAT1 siRNA,
5’- UAUGAACUGUUUCAAGAAGUUGAGC -3’ (sense) and 5’- GCUCAACUUCUUGAAACAGUUCAUA -3’ (antisense); MYST1 siRNA, 5’- UUGGGCUGUUUCCCAUAGUCUUCGG -3’ (sense) and 5’- CCGAAGACUAUGGGAAACAGCCCAA -3’ (antisense); Twist siRNA, 5’- UUGAGGGUCUGAAUCUUGCUCAGCU -3’ (sense) and 5’- AGCUGAGCAAGAUUCAGACCCUCAA -3’ (antisense);. c-Myc siRNA, 5’- UUUGUGUUUCAACUGUUCUCGUCGU -3’ (sense) and 5’-GCACAUGAUACAACAACAGACUUUA-3’ (antisense); USF1 siRNA,
5’- UCACAAAGAAUUGACCAGUGCCAGG -3’ (sense) and
5’-CCUUGACCAUAAGACACUGUACUAU -3’ (antisense). siRNA transfections were performed according to the manufacturer’s instructions, with modification (Invitrogen). Ten microliters lipofectamine transfection reagent was
diluted in 250 μl Opti-MEM I medium and incubated for 5 min at room temperature. Next, 500 pmol or 250 pmol Clock, Tip60, HAT1, MYST1, Twist, c-Myc, USF1, and inverted control duplex Stealth RNA
(Invitrogen) diluted in 250 μl Opti-MEM I were added gently, and incubated for 20 min at room temperature. Oligomer–lipofectamine complexes and
aliquots of 1×106 PC3 cells in 500 μl culture medium were combined and incubated for 10 min at room temperature. Aliquots of 4×102 PC3 cells or 6×102 P/CDP6 cells were used for a colony-formation assay as described below. The remaining cells were seeded in 100 mm dishes with 10 ml culture medium and harvested after 96 h culture for Western blotting, as described above.
Chromatin immunoprecipitation assay (ChIP).
The ChIP assay was performed as described previously (16, 22). Briefly, soluble chromatin from 1 x 106 PC3 or PCDP6 cells was incubated with 2 μg of anti-Clock antibody (sc-25361X), anti-H3K14 anti-body, anti-H4K16 antibody, and normal goat or rabbit immunoglobulin G (IgG). For transfection experiment, PC3 cells were transiently transfected with 3xFlag-HA vector or 3xFlag-HA Clock plasmid, and cultured for 48 hr as described above. Soluble chromatin from 1 x 106 cells was incubated with 2 μg of anti-Flag (M2) affinity gel or normal mouse IgG.
5
by guest on June 30, 2018http://w
ww
.jbc.org/D
ownloaded from

Immunoprecipitaed DNAs with protein A/G agarose were purified and dissolved in 20 μl of dH2O. Each 2 μl of DNA was used for PCR analysis with the following primer pairs; Tip60 #1 (–2586 to –2073), 5’-ATTACAGGAGCGAGCCACTGCGCAC-3’ (forward) and 5’-GGTAACTTCCTGCCGTTGCCATGGC-3’ (reverse); Tip60 #2 (–1336 to –944), 5’-CCCTTGAGCACCTATGCTCAGCCGG-3’ (forward) and 5’-GGCGCGGTGGCTCACGCCTATAATC-3’ (reverse); Tip60 #3 (–467 to +233),
5’- AGATCTAGACCATCCTGGCCAACATGG -3’ (forward) and 5’- AAGCTTCCGGCCCCTCTGGGAGACGTCAC -3’ (reverse); Tip60 #4 (+823 to +1212),
5’- TTCTGGGCCTGAGGTGGGGCATCAG -3’ (forward) and 5’- AGGACTAGCTACTGGACTCTGGGAGC -3’ (reverse). The PCR products were separated by electrophoresis on 2% agarose gels and stained with ethidium bromide.
Cytotoxicity assay. For the colony-formation assay,
4×102 PC3 cells or 6×102 P/CDP6 cells transfected with siRNA were seeded in 35 mm dishes with 2 ml culture medium. The following day, the cells were treated with indicated concentrations of cisplatin,
oxaliplatin, etoposide and vincristine. After 7 days, the number of colonies was counted. For correlation analysis, we calculated 50% inhibitory concentration (IC ) from the concentration–response curves.
50
RNA isolation. Total RNA was extracted using Qiagen™ RNeasy Mini Kits (Qiagen, Valencia, CA) following the RNeasy mini protocols for isolation from animal cells (I. spin protocol). RNA quality and integrity were determined using the Eukaryote Total RNA Nano 6000 assay (Agilent RNA 6000 Nano LabChip Kit) on the Agilent Technologies 2100 Bioanalyzer and quantified by measuring A260nm on a UV/Vis spectrophotometer (ND-1000, NanoDrop Technologies, Rockland, DE).
Microarray studies and data analysis. High-quality RNA samples (200 ng each) prepared from PC3 cells 72hr after transfection with control or Tip60 siRNA were amplified and labeled with Cy5-and Cy3-CTP (Amersham Biosciences) to produce labeled cRNA using Agilent low RNA input fluorescent linear amplification kits following the manufacturers protocol. After the amplification and labeling, the dye-incorporation ratio was determined using a Nanodrop spectrophotometer and the ratios were within 10 to 20 pmol per μg cRNA, the range the manufacturer suggests prior to hybridization. For hybridization, 750 ng of
6
by guest on June 30, 2018http://w
ww
.jbc.org/D
ownloaded from

cyanine 3- and 750 ng of cyanine 5-labeled cRNA were fragmented and hybridized to an Agilent Technologies Human 1A (V2) Gene Expression Microarray, using the Agilent Gene Expression hybridization kit as described in the Two-Color Microarray-Based Gene Expression Analysis v4.0 manual. Following hybridization, all microarrays were washed as described in the manual and scanned using Agilent's dual-laser DNA microarray scanner. The scans were converted to data files with Agilent's Feature Extraction software (Version 8.5). Data were analyzed using Microsoft Access and Spotfire. The arrays were scanned by the Agilent's dual-laser DNA microarray scanner using SureScan technology, extracted by feature Extraction software, and analyzed by Rosetta ResolverR software. An average of three replicate samples was used for each experiment.
Statistical analysis. Expression levels of protein assayed by western blotting were assessed numerically with the NIH image program (NIH, Bethesda, MD). The Pearson correlation was used for statistical analysis, and significance was set at the 5% level.
RESULTS Tip60 is overexpressed in cisplatin-resistant cells Human histone acetyltransferases (HATs) are categorized into three groups on the basis of their catalytic domains (23). The Gcn5
N-acetyltransferase (GNAT) family includes GCN5 and HAT1. The MYST family is named for the members of HAT family: MOZ, Ybf2, Sas2 and Tip60. Other HATs include p300/CBP and one of the nuclear receptor co-activators (24). We first investigated the expression levels of three human HAT genes: Tip60, MYST1, and HAT1 in cisplatin-sensitive and -resistant cells. As shown in Fig. 1, Tip60 was overexpressed in cisplatin-resistant cells when compared to sensitive cells. On the other hand, cellular expression of two HAT genes, MYST1 and HAT1, was almost similar in drug-sensitive and -resistant cells. Although we also examined other proteins, β-actin and PCNA, there was no difference in their expression in drug-sensitive and -resistant cells.
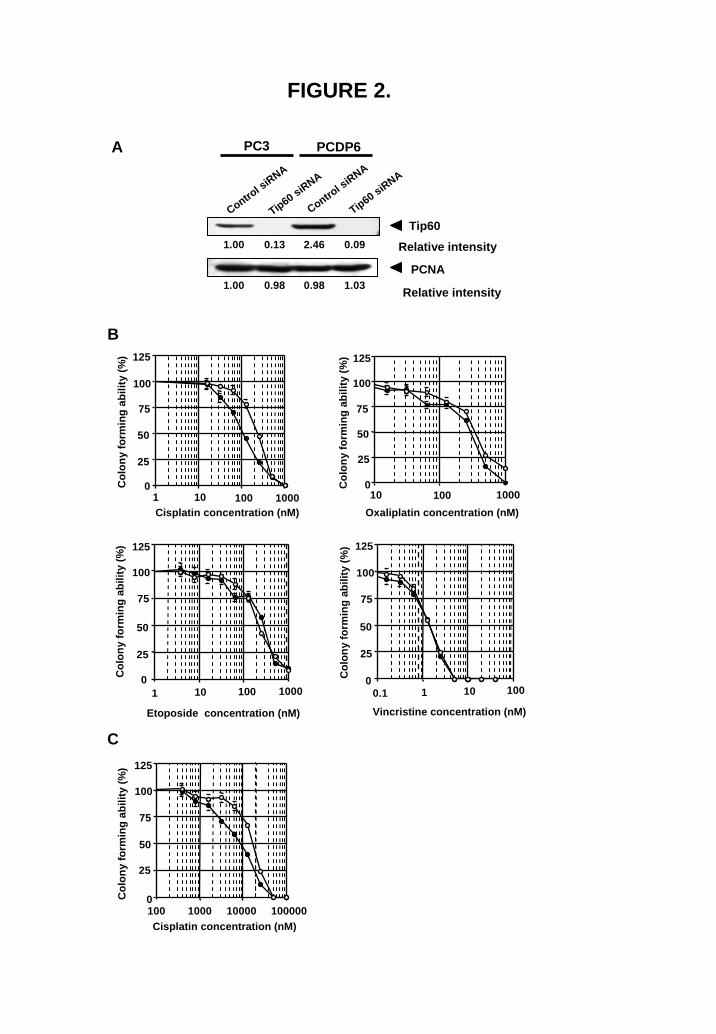
Tip60 silencing effectively sensitizes cells to cisplatin To examine whether Tip60 is involved in drug resistance, an siRNA strategy was employed (Fig. 2A). Cisplatin sensitivity was significantly enhanced by the knockdown of Tip60 expression (Fig. 2B). Knockdown of Tip60 expression did not induce cellular sensitivity to vincristine, etoposide and oxaliplatin (Fig. 2B). We next investigated whether down-regulation of Tip60 expression partially overcomes cisplatin resistance in cisplatin-resistant cell line P/CDP6. As shown in Fig. 2C, down-regulation of Tip60 expression partially overcame cisplatin resistance. We also confirmed that knockdown of other HAT
7
by guest on June 30, 2018http://w
ww
.jbc.org/D
ownloaded from

genes, MYST1 and HAT1, did not induce cellular sensitivity to cisplatin (data not shown). Circadian transcription factor Clock regulates Tip60 gene expression To explore the reason why Tip60 was overexpressed in cisplatin-resistant cells, we analyzed the function of the promoter of the Tip60 gene. There are two proximal E-boxes in the core promoter region of the Tip60 gene (Fig. 4A). We have previously shown that the E-box-binding transcription factor Clock is overexpressed in cisplatin-resistant cells (16). This prompted us to examine whether circadian transcription factor Clock is involved in Tip60 gene expression. Knockdown of Clock expression markedly reduced the expression of Tip60 in two human cancer PC3 and KB cell lines (Fig. 3A). On the other hand, the expression of other E-box-binding factors, c-Myc, Twist and USF1, did not influence the expression of Tip60 (Fig. 3B). The heterodimeric transcription factors Clock/Bmal1 that bind to E-box plays an indispensable role in driving the transcription of target genes. We have introduced the mutation in the two proximal E-boxes of Tip60 promoter as shown in Fig. 4A. The reporter assay showed that cotransfection of both Clock and Bmal1 expression plasmids displayed a 8-fold higher levels of luciferase activity. However, no transactivation was observed when the Tip60-MT-Luc was used (Fig. 4B). Further, silencing of Clock expression reduced the
promoter activity of the Tip60 gene (Fig. 4C).
Differences in histone acetylation in cisplatin-sensitive and -resistant cells It has been shown that Clock is an HAT (18) and is overexpressed in cisplatin-resistant cells (16). To determine whether overexpression of Clock and Tip60 leads to enhanced histone acetylation in cisplatin-resistant cells, we performed Western blotting using four acetylated histone-specific antibodies. Hyperacetylation of histones was clearly observed in cisplatin-resistant cells (Fig. 5). Both H3K14 and H4K16 were significantly hyperacetylated in cisplatin-resistant cells in comparison with parental cells. On the other hand, the acetylation of both H2AK5 and H3K9 did not alter between drug-sensitive and -resistant cells. Specificity of both Clock and Tip60 enzymatic activity was investigated by using specific siRNA for Clock and Tip60. Knockdown of Clock expression reduced the acetylation of H3K14 and H4K16, and knockdown of Tip60 expression reduced the acetylation of H4K16 in both cisplatin-sensitive and –resistant cells. The acetylation of both H2AK5 and H3K9 did not alter in cisplatin-sensitive and –resistant cells by using specific siRNA either for Clock or Tip60. Our results indicate that Clock acetylates H3K14 and Tip60 acetylates H4K16 in drug-sensitive and -resistant cells (Fig. 6).
8
by guest on June 30, 2018http://w
ww
.jbc.org/D
ownloaded from

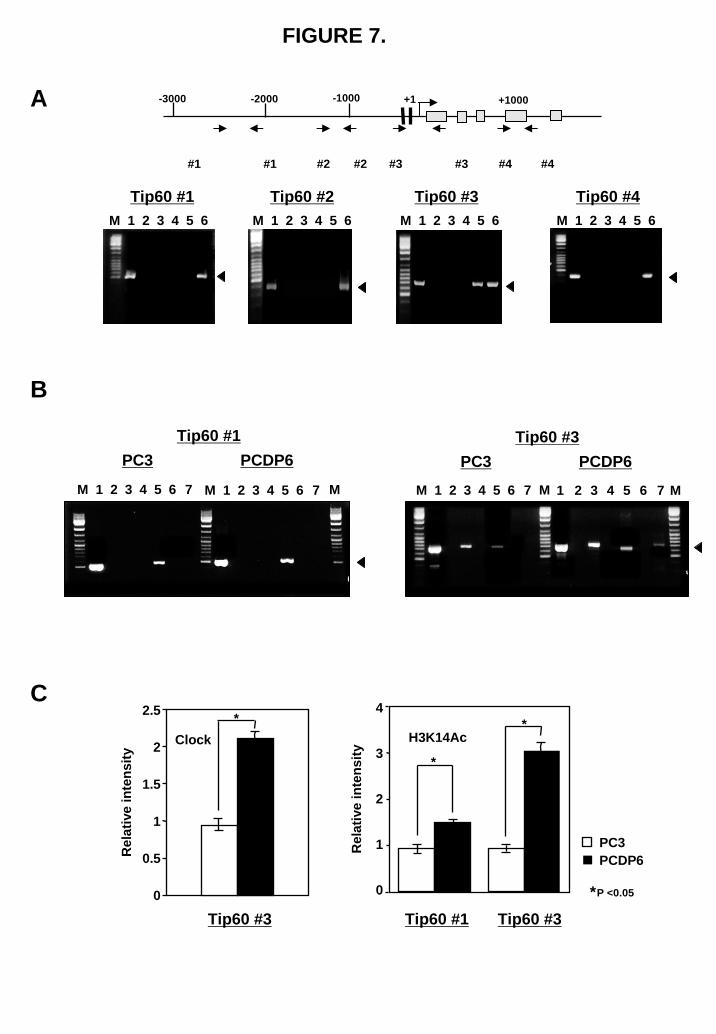
Clock and acetylated histone interaction with the Tip60 promoter To determine if the transactivation of Clock/Bmal1 on the Tip60 promoter was a result of direct recruitment of Clock/Bmal1 to the promoter, we carried out chromatin immunoprecipitation (ChIP) assays covering different segments of the promoter and 5’ end of the Tip60 gene. Initially, transient transfection of PC3 cells with a 3xFlag-HA-tagged Clock expression plasmid was used. Forty-eight hours after transfection, a ChIP assay was done with an anti-Flag antibody. As shown in Fig. 7A, we found that Clock was only present in one segment containing E-boxes. Fig. 7B shows the endogenous nuclear Clock interaction with the Tip60 promoter element was verified using two selected primer pairs (Tip60#1, #3). Anti-Clock antibody specifically coprecipitated the Tip60 promoter fragment containing E-boxes from both PC3 and P/CDP6 cells (Fig. 7C, right panel). We have also investigated the status of H3K14 and H4K16 acetylation associated with the Tip60 promoter. Both PC3 and P/CDP6 cells were subjected to ChIPs with antibodies against acetylated H3K14 and H4K16. Interestingly, it was observed that acetylated H3K14 was markedly bound to the Tip60 promoter when both Tip60 #1 and #3 were used (Fig. 7B, lane 5). On the other hand, H4K16 acetylation was less evident overall (Fig. 7B lane 7). Quantification of the band densities after normalization shows that drug resistant P/CDP6 cells have significant increase in
Clock and acetylated H3K14 binding to the Tip60 promoter compared with PC3 cells (Fig. 7C).
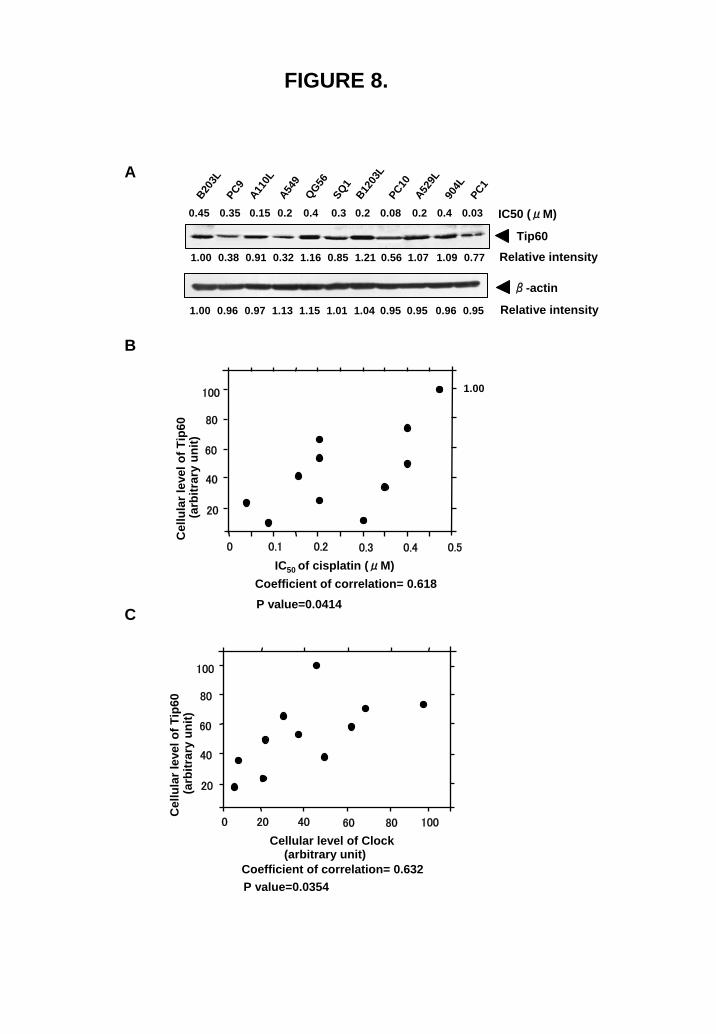
Cellular expression of Tip60 correlates with cisplatin sensitivity We examined the correlation between Tip60 expression and cisplatin sensitivity in 11 lung cancer cell lines (Fig. 8). Expression of β-actin was also examined as a control (Fig. 8A), because β-actin expression in cisplatin-resistant cells is almost similar to that in parental cells (Fig. 1). Tip60 expression significantly correlated with cisplatin sensitivity (Fig. 8B), and Clock expression (Fig. 8C). We have previously shown that Clock expression correlated with cisplatin sensitivity (16).
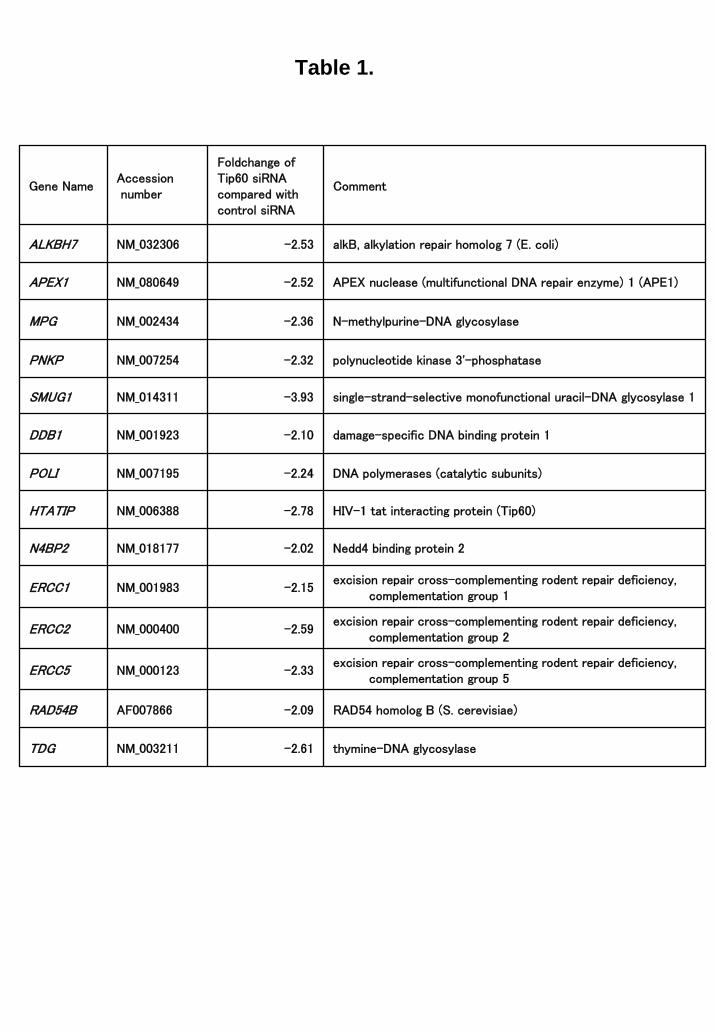
Microarray analysis of Tip60-regulated genes Because the available information regarding the Tip60-target genes is limited, we used microarray technology. To investigate the transcriptional changes by the Tip60 siRNA, DNA microarray study was carried out in PC3 cells treated with control or Tip60 siRNA. Data analysis identified 1091 genes, which were down regulated more than 2- fold and 1150 genes which were upregulated (data not shown). Among down-regulated genes, several genes involved in DNA repair was identified (Table 1). It was reported that ERCC1 was one of the key molecules to determine the cellular sensitivity of cisplatin (25, 26). To evaluate the microarray study, we selected two genes,
9
by guest on June 30, 2018http://w
ww
.jbc.org/D
ownloaded from

and examine these expression by western blot analysis. Both ERCC1 and APE1 (27) were significantly down regulated by the Tip60 siRNA (Fig. 9A). Further, the expression of these genes was significantly upregulated in cisplatin-resistant cells (Fig. 9B).
DISCUSSION Alterations in transcription are thought to play a major role in tumorigenesis, as well as in tumor response and resistance to anticancer agents (1, 2). Our objective was to identify transcription-related factors whose expression was up-regulated and involved in cisplatin resistance (1, 2, 13). Recently, we have reported that the circadian transcription factor Clock is overexpressed in cisplatin-resistant cells (16). Clock has recently been found to have intrinsic HAT activity (18). The HAT activity is required to activate Clock/Bmal1-dependent transcription. Thus, Clock may promote Clock/Bmal1-dependent transcription by forming active chromatin and increasing the accessibility of transcriptional cofactors to the promoter region.
Tip60 was originally identified as a cellular acetyltransferase that interacts with HIV-1 Tat protein (28). Furthermore, Tip60 exists as a multimeric protein complex on chromatin, and has been shown to be involved in DNA damage response and repair (20, 29). These findings prompted us to test the hypothesis that HAT gene expression might affect cellular sensitivity to DNA-damaging agents including cisplatin. In
the present study, we investigated which HATs are involved in modifying cellular sensitivity to anticancer agents and drug resistance. We found that Tip60 was overexpressed in cisplatin-resistant cells, but MYST1 and HAT1 were not (Fig. 1). We have previously shown that down-regulation of the cellular expression of the Clock protein conferred cisplatin and etoposide sensitivity to human lung cancer cell lines (16). Here, we also found that Tip60 expression correlated well with cisplatin sensitivity (Fig. 8B). However, down-regulation of Tip60 expression conferred only cisplatin sensitivity (Fig. 2B), which suggests a different mechanism through which either Clock or Tip60 modifies drug sensitivity. Several mechanisms, are involved in the acquisition of cisplatin resistance including decreased cisplatin accumulation, increased levels of cellular glutathione and increased DNA repair activity. These supported that the knockdown of Tip60 expression by Tip60 siRNA showed only a small effect on the cellular sensitivity of cisplatin (Fig. 2B, C). We have previously shown that ATF4 is a target of Clock, and the Clock and ATF4 transcription system plays an important role in multidrug resistance, through a glutathione-dependent redox system (16). The knockdown of Tip60 expression also showed a similar effect on the cellular sensitivity in drug sensitive cell. DNA microarray analysis showed several DNA repair genes are regulated by the Tip60 expression (Table 1). As shown in Fig. 8,
10
by guest on June 30, 2018http://w
ww
.jbc.org/D
ownloaded from

both ERCC1 and APE1 gene are regulated by Tip60.
To investigate the molecular mechanism of the enhanced expression of Tip60 in cisplatin-resistant cells, we surveyed the transcription factor binding sites in the promoter region using the human genome database. There are two proximal E-boxes in the core promoter region of the Tip60 gene (Fig. 4A). We found that the Tip60 gene is a target of Clock, but not of the other E-box-binding factors, such as Twist, c-Myc and USF1 (Fig. 3B). The ChIP assay also support these results (Fig. 7). Our results indicate that Tip60 might also be involved in rhythmic histone acetylation. Histone acetylation in cisplatin-resistant cells is supposed to be regulated by Clock and Tip60. We found that both H3K14 and H4K16 were highly acetylated in cisplatin-resistant cells (Fig. 5). It has been shown that Clock can acetylate H3K14 well (19), and that Tip60 acetylates H2A and H4 (23, 29), which indicates that H4K16 may be catalyzed by Tip60. Our results using siRNA for Clock and Tip60 are consistent with those in previous studies (Fig. 6). The acetylation of H3K14 has been shown to be involved in transcriptional activation (19). It has been shown that the acetylation of H4K16 may contribute to the decondensation of chromatin (30). As shown in Fig. 7, the ChIP assays demonstrate evidence of high Clock binding and H3K9 acetylation at the Tip60 promoter in drug resistant cells, implicating a more open state of chromatin primed for
transcription. Based on these data, Clock overexpression might be involved in maintaining transcription-permissive chromatin structure through H3K14 acetylation, and Tip60 might be involved in establishing DNA-repair-permissive chromatin structure in cisplatin-resistant cells. Thus, our results stress the positive role that the HAT function of both Clock and Tip60 contribute to drug resistance.
In conclusion, we established that both Clock and Tip60 are required for cisplatin resistance. Our data also suggest that Tip60 may play a role in circadian clock function. Our study begins to offer molecular insight into the circadian regulation of genomic response against anticancer agents, and drug sensitivity through histone acetylation.
11
by guest on June 30, 2018http://w
ww
.jbc.org/D
ownloaded from

REFERENCES 1. Kohno, K., Uchiumi, T., Niina, I., Wakasugi, T., Igarashi, T., Momii, Y., Yoshida, T.,
Matsuo, K., Miyamoto, N., Izumi, H. (2005). Eur J Cancer. 41, 2577-2586. 2. Torigoe, T., Izumi, H., Ishiguchi, H.,Yoshida, Y., Tanabe, M., Yoshida, T., Igarashi, T.,
Niina, I., Wakasugi, T.,Imaizumi, T., Momii, Y., Kuwano, M., and Kohno, K. (2005).
Curr Med Chem Anticancer Agents. 5, 15-27. 3. Zamble, D. B., and Lippard, S. J. (1995). Trends Biochem Sci. 20, 435-439. 4. Cohen, S. M., and Lippard, S. J. (2001). Prog Nucleic Acid Res Mol Biol. 67, 93-130. 5. Wada, T., and Penninger, J. M. (2004). Oncogene. 23, 2838-2849. 6. Fujii, R., Mutou, M., Niwa, K., Yamada, K., Akitou, T., Nakagawa, M., Kuwano, M., and
Akiyama, S. (1994). Jpn J Cancer Res. 85, 426-433. 7. Komatsu, M., Sumizawa, T., Mutou, M., Chen, Z. S., Terada, K., Furukawa, T. Yang, X. L.,
Gao, H., Miura, N., Sugiyama, T., and Akiyama, S. (2000). Cancer Res. 60, 1312-1316. 8. Nakayama, K., Kanzaki, A., Ogawa, K., Miyazaki, K., Neamati, N., and Takebayashi, Y.
(2002). Int J Cancer. 101, 488-495. 9. Lai, G. M., Ozols, R. F., Young, R. C., and Hamilton, T. C. (1989). J Natl Cancer Inst. 81,
535-539.
10. Tew KD. (1994). Cancer Res. 54, 4313-4320. 11. Chaney, S. G., and Sancar, A. (1996). J Natl Cancer Inst. 88, 1346-1360. 12. Husain, A., He, G., Venkatraman, E. S., and Spriggs, D. R. (1998). Cancer Res. 58,
1120-1123. 13. Murakami, T., Shibuya, I., Ise, T., Chen, Z. S., Akiyama, S., Nakagawa, M., Izumi, H.,
Nakamura, T., Matsuo, K., Yamada, Y., and Kohno, K. (2001). Int J Cancer. 93, 869-874. 14. Tanabe, M., Izumi, H., Ise, T., Chen, S. J., Higuchi, S., Yamori, T., Yasumoto, K., and
Kohno, K. (2003). Cancer Res. 63, 8592-8595. 15. Harding, H. P., Zhang, Y., Zeng, H., Novoa, I., Lu, P. D., Calfon, M., Sadri, N., Yun, C.,
Popko, B., Stojdl, D. F., Bell, J. C., Hettmann, T., Leiden, J. M., and Ron, D. (2003). Mol
Cell. 11, 619-633. 16. Igarashi, T., Izumi, H., Uchiumi, T., Nishio, K., Arao, T., Tanabe, M., Uramoto, H., Sugio,
K., Yasumoto, K., Sasaguri, Y., Wang, K. W., Otsuji, Y., and Kohno, K. (2007).
Oncogene. 26, 4749-4760. 17. Etchegaray, J. P., Lee, C., Wada, P. A., and Reppert, S. M. (2003). Nature. 421, 177-182. 18. Doi, M., Hirayama, J., and Sassone-Corsi, P. (2006). Cell. 125, 497-508. 19. Nakahata, Y., Grimaldi, B., Sahar, S., Hirayama, J., Sassone-Corsi, P. (2007). Curr Opin Cell
Biol. 19, 230-237.
12
by guest on June 30, 2018http://w
ww
.jbc.org/D
ownloaded from

20. Bird, A. W., Yu, D. Y., Pray-Grant, M. G., Qiu, Q., Harmon, K. E., Megee, P. C., Grant, P.
A. , Smith, M. M., and Christman, M. F. (2002). Nature. 419, 411-415. 21. Ikura, T., ogryzko, V. V., Grigoriev, M., Groisman, R., Wang, J., Horikoshi, M., Scully, R.,
Qin, J., and Nakatani, Y. (2000). Cell. 102, 463-473. 22. Wakasugi, T., izumi, H., Uchiumi, T., Suzuki, H., Arao, T., Nishio, K., and Kohno, K.
(2007). Oncogene. 26, 5194-5203. 23. Lee, K. K., and Workman, J. L. (2007). Nat Rev Mol Cell Biol. 8, 284-95. 24. Kimura, A., Matsubara, K., and Horikoshi, M. (2005). J. Biochem (Tokyo). 138, 647-662. 25. Olaussen, K. A., Dunant, A., Fouret, P., Brambilla, E., Andre, F., Haddad, V., Taranchon, E.,
Filipits, M., Pirker, R., Popper, H. H., Stahel, R., Sabatier, L., Pignon, J. P., Tursz, T., Le
Chevalier, T., Soria, J. C. (2006). N Engl J Med. 355, 983-991. 26. Takenaka, T., Yoshino, I., Kouso, H., Ohba, T., Yohena, T., Osoegawa, A., Shoji, F., and
Maehara, Y. (2007). Int. J. Cancer. 121, 895-900. 27. Daniel, R. M. and David, M. W. (2007). Mol. Cancer. Res. 5, 61-70. 28. Kamine, J., Elangovan, B., Subramanian, T., Coleman, D., and Chinnadurai, G. (1996).
Virology. 216, 357-366. 29. Squatrito, M., Gorrini, C., and Amati, B. (2006). Trends Cell Biol. 16, 433-442. 30. Shogren-Knaak, M., Ishii, H., Sun, J. M., Pazin, M. J., Davie, J. R., and Peterson, C. L.
(2006). Science. 311, 844-847.
FOOTNOTES *Grant support: the Ministry of Education, Culture, Sports, Science, and Technology of Japan (Mext), Kakenhi (13218132 and 18590307), Kobayashi Institute for Innovative Cancer Chemotherapy, and a Grant-in-Aid for Cancer Reserch from the Fukuoka Cancer Society,Japan. We thank Satoko Takazaki and Yukiko Yoshiura for their technical assistance.
The abbreviations used are: Tip60, HIV-1-tat interactive protein; HAT, histone acetyltransferase; CBB, Coomassie Brilliant Blue; SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gel electrophoresis; DTT, dithiothreitol; PCR, polymerase chain reaction; ERCC1, excision repair cross-complementation group 1; APE1, apurinic/apyrimidinic exonuclease 1; Clock, circadian locomoter output cycles kaput; Bmal1, brain and muscle ARNT-like protein 1; ATF4, activating transcription factor 4; USF1, upstream stimulatory factor 1; MYST1, MYST histone acetyltransferase 1; siRNA, small interference RNA; PCNA, proliferating cell nuclear antigen; ChIP, chromatin immunoprecipitation.
FIGURE LEGENDS FIGURE 1.
13
by guest on June 30, 2018http://w
ww
.jbc.org/D
ownloaded from

Tip60 is overexpressed in cisplatin resistant cells. Fifty μg of whole-nuclear lysates of PC3 and KB cells and their cisplatin-resistant P/CDP6 and KB/CP4 derivatives were subjected to SDS-PAGE, and Western blotting was performed with the indicated antibodies. Relative intensity is shown under each blot.
FIGURE 2. Silencing Tip60 sensitizes the cells to cisplatin. (A) Control siRNA (100 pmol) or Tip60 siRNA (100 pmol) were transfected into PC3 cells or P/CDP6 cells, and whole-nuclear lysates (50 μg) were subjected to SDS-PAGE. Transferred membrane was blotted with anti-Tip60 and anti-PCNA antibodies. (B) PC3 cells were treated with 100 pmol control siRNA (open circles) or 100 pmol Tip60 siRNA (closed circles) for 24 h, and exposed to various concentrations of cisplatin, oxaliplatin, etoposide, and vincristine for 7 days. The colony number in the absence of drug corresponded to 100%. All values were the mean ±SD of at least three independent experiments. (C) P/CDP6 cells were treated with 100 pmol control siRNA (open circles) or 100 pmol Tip60 siRNA (closed circles) for 24 h, and exposed to various concentrations of cisplatin for 7 days. The colony number in the absence of drug corresponded to 100%. All values are the mean±SD of at least three independent experiments.
FIGURE 3. Tip60 gene expression is regulated through the E-boxes located in the promoter region by Clock but not by c-Myc, Twist and USF1. (A) Control siRNA (100 pmol) or Clock siRNA (100 pmol) were transfected into PC3 cells and KB cells. Fifty μg of whole-nuclear lysates were subjected to SDS-PAGE, and Western blotting was performed with the indicated antibodies. Relative intensity is shown under each blot. (B) Indicated siRNA (100 pmol) were transfected into PC3 cells and KB cells. Whole-nuclear lysates (50 μg for c-Myc and USF1, and 100 μg for Twist) were subjected to SDS-PAGE, and Western blotting was performed with the indicated antibodies. Relative intensity is shown under each blot.
FIGURE 4. Clock regulates Tip60 expression through binding to the E-boxes in the promoter region. (A) Schematic representation of the promoter region of the Tip60 gene. Tip60-WT-Luc and Tip60-MT-Luc used in Fig. 4B are also shown. +1 indicates transcription start site. (B) Indicated amounts of Clock and/or Bmal1 expression plasmids were transiently co-transfected with Tip60-WT-Luc or Tip60-MT-Luc into PC3 cells. pGL3-B.V. indicates pGL3 basic vector in which luciferase gene is not driven by any promoters. The results were normalized to β-galactosidase activity and are representative of at least three independent experiments. Bars
represent ±SD. The luciferase activity of Tip60-WT-Luc was set to 1. (C) PC3 cells were transfected with control siRNA (100 pmol) or Clock siRNAs (100 pmol). The following day, they were transfected with the indicated reporter plasmids. The results were normalized to
14
by guest on June 30, 2018http://w
ww
.jbc.org/D
ownloaded from

β-galactosidase activity and pGL3 promoter vector. All values are the mean of at least three independent experiments. pGL3-P.V. indicates the pGL3 promoter vector in which luciferase
gene is driven by SV-40 promoter and pGL3-B.V. was described above. Bars represent ±SD. FIGURE 5. Histone acetylation is upregulated in cisplatin resistant cells. Whole-cell extracts (25 μg for H3K9, H3K14, and 50 μg for H2AK5, H4K16) of PC3 and KB cells and their cisplatin-resistant PC/DP6 and K/CP4 derivatives were subjected to SDS-PAGE, and Western blotting was performed with the indicated antibodies. Relative intensity is shown under each blot.
FIGURE 6. Silencing Clock and Tip60 reduces histone acetylation. PC3 and PC/DP6 cells were transfected with control siRNA (100 pmol), Clock siRNA (100 pmol) and Tip60 siRNA (100 pmol). Whole-nuclear lysates (50 μg for Clock and Tip60) and whole-cell extracts (25 μg for H3K9, H3K14, and 50 μg for H2AK5, H4K16) were subjected to SDS-PAGE, and Western blotting was performed with the indicated antibodies. Relative intensity is shown under each blot.
FIGURE 7. Clock and acetylated histone interaction with the Tip60 promoter. (A) Schematic representation of the promoter region and 5’ end of Tip60 gene. Black box and gray box indicate E-box (CACGTG) and exon, respectively. Chromatin immunoprecipitation assays using PC3 cells transiently transfected with 3xFlag vector (lane 1-3) or 3xFlag-HA-Clock plasmid (lane 4-6) was done with an anti-Flag (M2) antibody or normal mouse IgG. Immunoprecipitated DNAs with normal mouse IgG (lane 2 and 4) or anti-Flag antibody (lane 3 and 5) and soluble chromatin (lane 1 and 6) were amplified by PCR with specific primer pairs for the Tip60 promoter (#1, -2586 to -2073 bp; #2, -1336 to -944 bp; #3, -467 to +233 bp; #4, +823 to +1212 bp). Lane M, DNA size marker; arrowhead, PCR products (514 bp for Tip60 #1; 393 bp for Tip60 #2; 700 bp for Tip60 #3; 390 bp for Tip60 #4). (B) A ChIP assay of PC3 and PCDP6 cells was performed with normal goat IgG (lane 2 and 6), normal rabbit IgG (lane 4), anti-Clock antibody (lane 3), anti-H3K14 antibody (lane 5) or anti-H4K16 antibody (lane 7), and soluble chromatin (lane 1). Immunoprecipitated DNAs were amplified by PCR with specific primer pairs for the Tip60 promoter (#1, -2586 to -2073 bp) or (#3, -467 to +233 bp). Lane M, DNA size marker; arrowhead, PCR products (514 bp for Tip60 #1). (C) Densitometric quantification of PCR products of (B). All values are the mean of three independent experiments. *P<0.05 versus control. FIGURE 8. Tip60 expression correlates with cisplatin resistance and Clock expression. (A) Western blotting was performed with whole-nuclear lysates (100 μg) for Tip60 and β-actin expression in 11 lung cancer cell lines. (B, C) Expression levels of Tip60 and Clock (whole band) were determined by
15
by guest on June 30, 2018http://w
ww
.jbc.org/D
ownloaded from

NIH imaging program using Fig. 7A, and normalized by each β-actin expression level. The maximum expression levels of Tip60 and Clock were set to 100, and the IC50 of each cell line was calculated from the concentration–response curves for cisplatin (14). FIGURE 9. Tip60 regulates ERCC1 and APE1 expression. (A) PC3 cells and KB cells were treated with 100 pmol control siRNA or Tip60 siRNA for 72 hr, and whole-nuclear lysates (100 μg) were subjected by SDS-PAGE, and Western blotting was performed with the indicated antibodies. Relative intensity is shown under each blot. (B) Whole-nuclear lysates (100 μg) of PC3 and KB cells and their cisplatin-resistant P/CDP6 and KC/P4 were subjected by SDS-PAGE. Western blotting was performed with the indicated antibodies. Relative intensity is shown under each blot.
Table 1. List of DNA repair genes regulated by Tip60. DNA microarray analysis of Tip60-regulated genes was investigated. Oligonucleotide microarray study was carried out in PC3 cells treated with Tip60 siRNA (100 pmol) or control siRNA (100 pmol). Data analysis identified 1091 genes, which were down-regulated more than 2.0-fold. The subset of DNA repair genes was further selected if fold change marked > 2.0 between averaged Tip60 siRNA and control siRNA samples.
16
by guest on June 30, 2018http://w
ww
.jbc.org/D
ownloaded from

FIGURE 1.
CBB
P/CDP6
KB/CP4
KB
PC3Tip60
MYST1
HAT1
PCNA
β-actin
Relative intensity
Relative intensity
Relative intensity
Relative intensity
Relative intensity
1.00 3.22 1.00 3.13
1.00 1.02 1.00 1.03
1.00 0.98 1.00 1.00
1.00 0.99 1.00 1.01
1.00 1.00 1.00 1.01
by guest on June 30, 2018http://w
ww
.jbc.org/D
ownloaded from
0
25
50
75
100
125
1 10 100 10000
25
50
75
100
125
0.1 1 10 100
0
25
50
75
100
125
1 10 100 10000
25
50
75
100
125
10 100 1000
Col
ony
form
ing
abili
ty(%
)C
olon
y fo
rmin
g ab
ility
(%)
Col
ony
form
ing
abili
ty(%
)C
olon
y fo
rmin
g ab
ility
(%)
Cisplatin concentration (nM)
Etoposide concentration (nM)
Oxaliplatin concentration (nM)
Vincristine concentration (nM)
B
Cisplatin concentration (nM)
Col
ony
form
ing
abili
ty(%
)
A
FIGURE 2.
0
25
50
75
100
125
100 1000 10000 100000
C
Control siRNA
Tip60 siRNA
PC3 PCDP6
Control siRNA
Tip60 siRNA
Tip60
Relative intensity
PCNA
Relative intensity
1.00 0.13 2.46 0.09
1.00 0.98 0.98 1.03
by guest on June 30, 2018http://w
ww
.jbc.org/D
ownloaded from
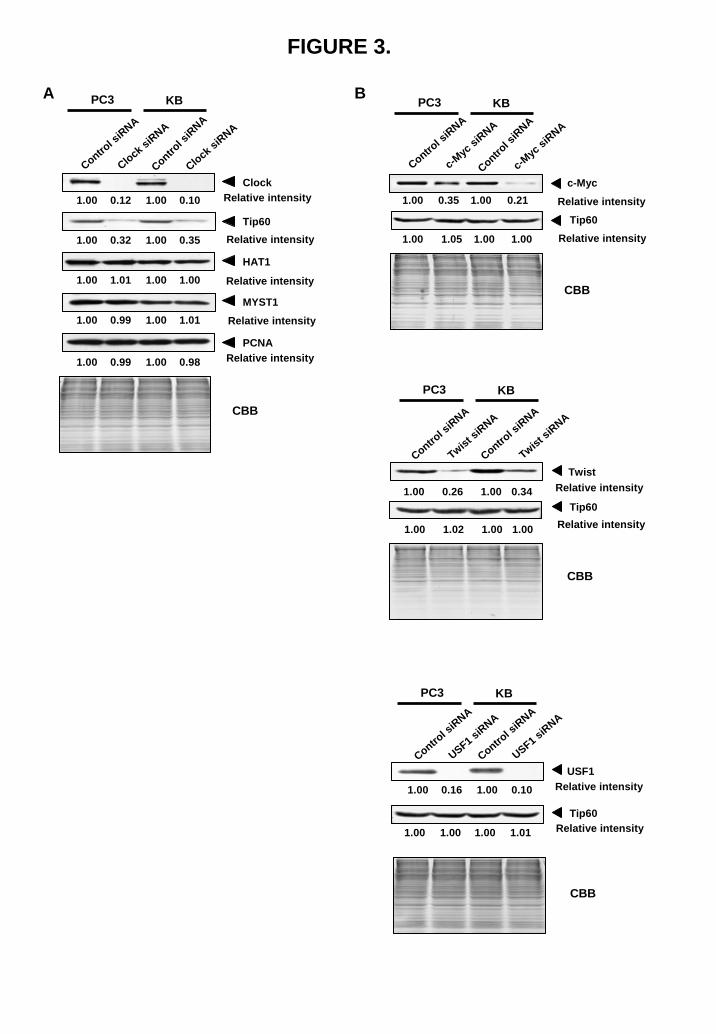
FIGURE 3.
B
c-Myc
CBB
Control s
iRNA
c-Myc
siRNA
Tip60
Control s
iRNA
c-Myc
siRNA
Relative intensity
Relative intensity1.00 1.05 1.00 1.00
1.00 0.35 1.00 0.21
PC3 KB
CBB
Control s
iRNA
Twist siRNA
Tip60
TwistContro
l siRNA
Twist siRNA
Relative intensity
Relative intensity
1.00 0.26 1.00 0.34
1.00 1.02 1.00 1.00
PC3 KB
USF1
CBB
Control s
iRNA
USF1 siRNA
Tip60
Control s
iRNA
USF1 siRNA
Relative intensity
Relative intensity
1.00 0.16 1.00 0.10
1.00 1.00 1.00 1.01
PC3 KB
A
CBB
Relative intensityClock
PCNA
HAT1
MYST1
Tip60Relative intensity
Relative intensity
Relative intensity
Relative intensity
PC3 KB
1.00 0.99 1.00 0.98
1.00 0.99 1.00 1.01
1.00 1.01 1.00 1.00
1.00 0.32 1.00 0.35
1.00 0.12 1.00 0.10
Control s
iRNA
Clock siRNA
Control s
iRNA
Clock siRNA
by guest on June 30, 2018http://w
ww
.jbc.org/D
ownloaded from
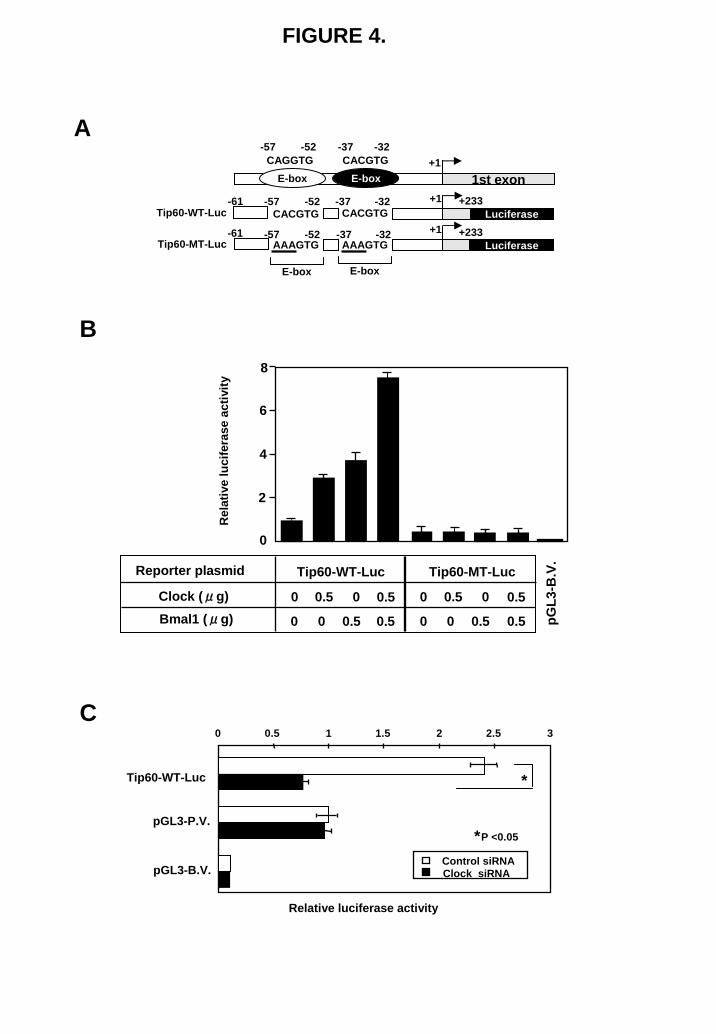
FIGURE 4.
A
C
B
0
2
4
6
8
pGL3
-B.V
.Tip60-WT-LucReporter plasmid
0.5
0.50.5
00.5
0
0
0
Tip60-MT-Luc
0.5
0.50.5
00.5
0
0
0
Clock (μg)Bmal1 (μg)
Rel
ativ
e lu
cife
rase
activ
ity
Relative luciferase activity
0 0.5 1 1.5 2 2.5 3
Clock siRNAControl siRNA
*P <0.05
*
pGL3-P.V.
Tip60-WT-Luc
pGL3-B.V.
Tip60-WT-Luc
Tip60-MT-Luc
-37 -32-57 -52
E-box 1st exonE-boxCACGTGCAGGTG
CACGTG Luciferase+233
AAAGTG Luciferase+233
CACGTG
AAAGTG
E-boxE-box
-37 -32-57 -52
-37 -32-57 -52
-61
-61
+1
+1
+1
by guest on June 30, 2018http://w
ww
.jbc.org/D
ownloaded from
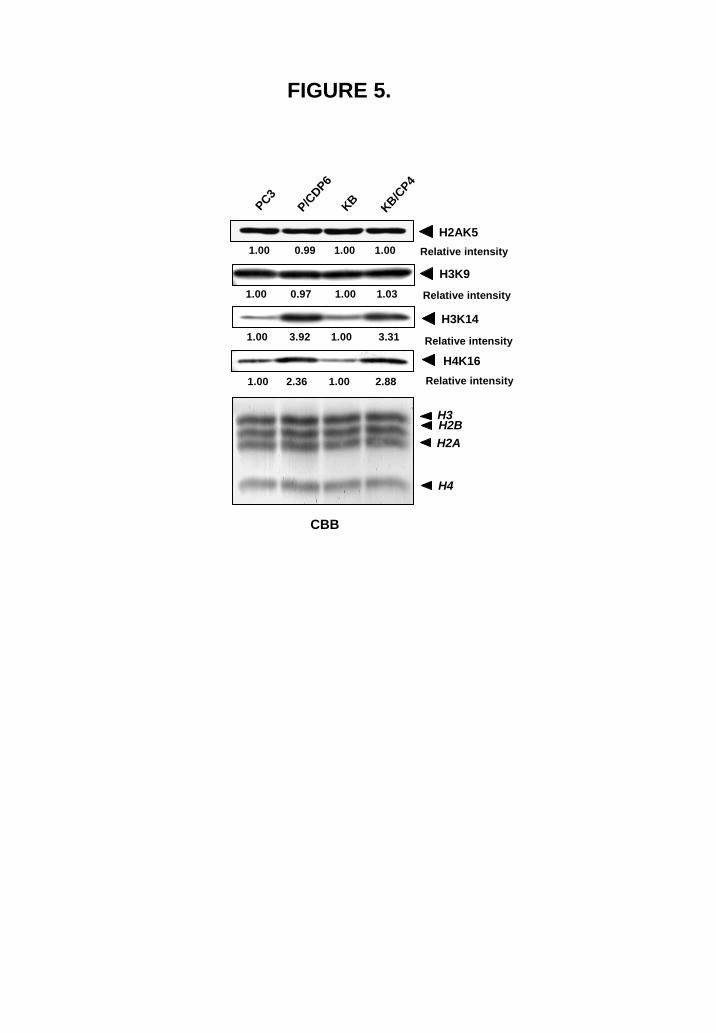
FIGURE 5.
P/CDP6
KB/CP4
CBB
KBPC3
H2AK5
H3K9
H3K14
H4K16
H3
H2A
H4
H2B
Relative intensity
Relative intensity
Relative intensity
Relative intensity
1.00 0.99 1.00 1.00
1.00 0.97 1.00 1.03
1.00 3.92 1.00 3.31
1.00 2.36 1.00 2.88
by guest on June 30, 2018http://w
ww
.jbc.org/D
ownloaded from
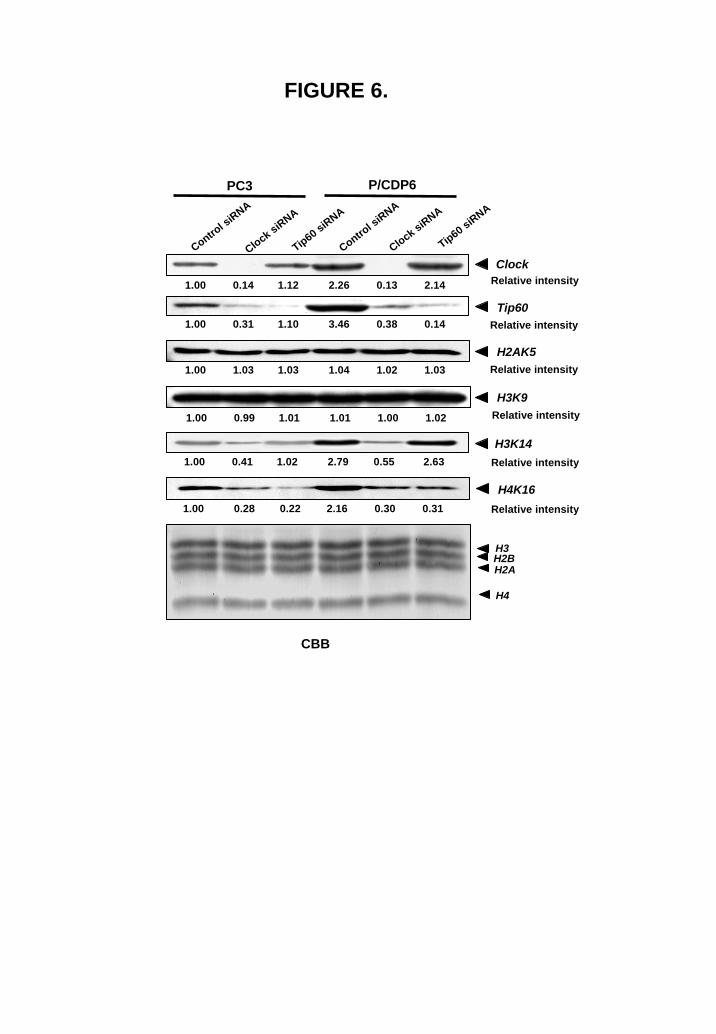
FIGURE 6.
H3K14
PC3 P/CDP6
Clock
Tip60
H4K16
Control s
iRNA
Clock siRNA
Tip60 siRNA
Tip60 siRNA
Clock siRNA
Control s
iRNA
H3
H2A
H4
H2B
Relative intensity
Relative intensity
Relative intensity
Relative intensity
CBB
H2AK5Relative intensity
H3K9Relative intensity
1.00 0.14 1.12 2.26 2.140.13
1.00 0.31 1.10 3.46 0.140.38
1.00 1.03 1.03 1.04 1.031.02
1.00 0.99 1.01 1.01 1.021.00
1.00 0.41 1.02 2.79 2.630.55
1.00 0.28 0.22 2.16 0.310.30
by guest on June 30, 2018http://w
ww
.jbc.org/D
ownloaded from
#1 #1 #2 #2 #3 #3 #4 #4
+1 +1000-1000-2000-3000
Tip60 #3 Tip60 #4Tip60 #2Tip60 #1M 1 2 3 4 5 6 M 1 2 3 4 5 6 M 1 2 3 4 5 6 M 1 2 3 4 5 6
FIGURE 7.
A
B
Tip60 #1
M 1 2 3 4 5 6 7 M 1 2 3 4 5 6 7
PC3 PCDP6
M M 1 432 5 6 7M 1 2 3 4 5 6 7
Tip60 #3PC3 PCDP6
M
C
0
1
2
3
4
Clock H3K14Ac
PCDP6PC3
0
0.5
1
1.5
2
2.5
Rel
ativ
e in
tens
ity
Rel
ativ
e in
tens
ity
Tip60 #1Tip60 #3 Tip60 #3
*
*
*
*P <0.05
by guest on June 30, 2018http://w
ww
.jbc.org/D
ownloaded from
B
C
A
FIGURE 8.
Coefficient of correlation= 0.618
20
40
60
80
100
0 0.1 0.2 0.3 0.4 0.5
IC50 of cisplatin (μM)
P value=0.0414
Cel
lula
r lev
el o
f Tip
60(a
rbitr
ary
unit)
1.00
20
40
60
80
100
0 20 40 60 80 100
Cellular level of Clock
Coefficient of correlation= 0.632P value=0.0354
Cel
lula
r lev
el o
f Tip
60(a
rbitr
ary
unit)
(arbitrary unit)
B203L
B1203
L
A110L
A549
QG56SQ1
PC10A52
9L90
4LPC1
PC9Tip60
β-actin
1.00 0.38 0.91 0.32 1.16 0.85 1.21 0.56 1.07 1.09 0.77
1.00 0.96 0.97 1.13 1.15 1.01 0.950.951.04 0.96 0.95
Relative intensity
Relative intensity
0.45 0.35 0.15 0.2 0.4 0.3 0.2 0.08 0.2 0.4 0.03 IC50 (μM)
by guest on June 30, 2018http://w
ww
.jbc.org/D
ownloaded from
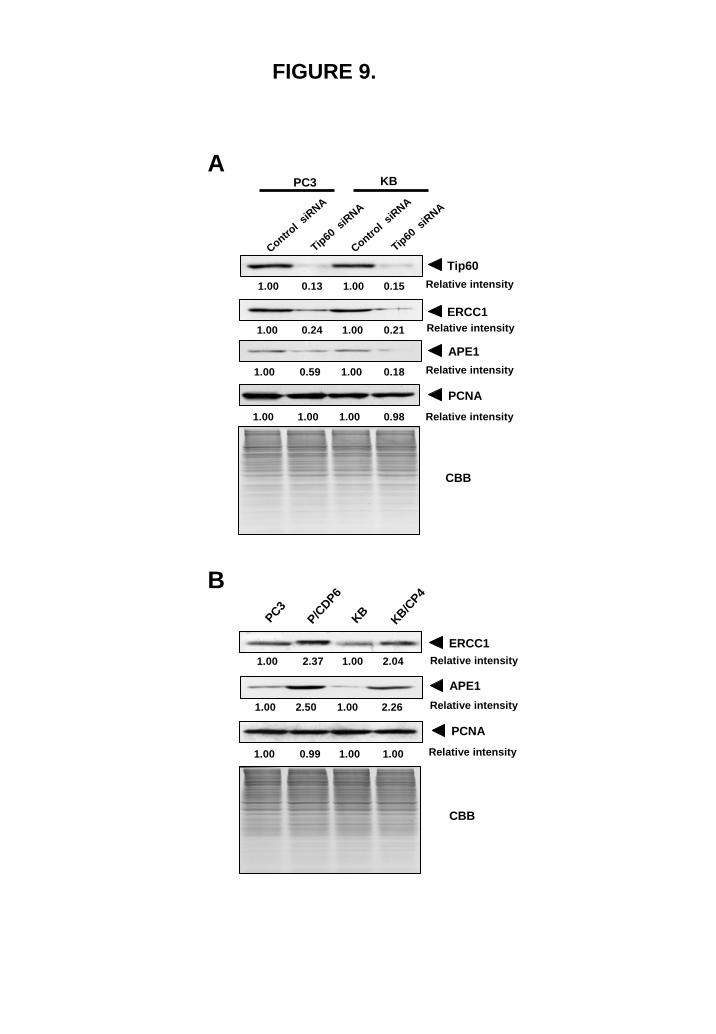
FIGURE 9.
P/CDP6
KB/CP4
KBPC3
ERCC1
APE1
PCNA
Relative intensity
Relative intensity
Relative intensity
B
1.00 2.37 1.00 2.04
1.00 2.50 1.00 2.26
1.00 0.99 1.00 1.00
Control s
iRNA
Tip60 siRNA
Control s
iRNA
Tip60 siRNA
PC3 KB
APE1
PCNA
Relative intensity
Relative intensity
A
1.00 0.13 1.00 0.15
1.00 0.24 1.00 0.21
1.00 0.59 1.00 0.18
1.00 1.00 1.00 0.98
Tip60Relative intensity
Relative intensityERCC1
CBB
CBB
by guest on June 30, 2018http://w
ww
.jbc.org/D
ownloaded from
Gene NameAccession number
Foldchange ofTip60 siRNAcompared withcontrol siRNA
Comment
ALKBH7 NM_032306 -2.53 alkB, alkylation repair homolog 7 (E. coli)
APEX1 NM_080649 -2.52 APEX nuclease (multifunctional DNA repair enzyme) 1 (APE1)
MPG NM_002434 -2.36 N-methylpurine-DNA glycosylase
PNKP NM_007254 -2.32 polynucleotide kinase 3'-phosphatase
SMUG1 NM_014311 -3.93 single-strand-selective monofunctional uracil-DNA glycosylase 1
DDB1 NM_001923 -2.10 damage-specific DNA binding protein 1
POLI NM_007195 -2.24 DNA polymerases (catalytic subunits)
HTATIP NM_006388 -2.78 HIV-1 tat interacting protein (Tip60)
N4BP2 NM_018177 -2.02 Nedd4 binding protein 2
ERCC1 NM_001983 -2.15excision repair cross-complementing rodent repair deficiency,
complementation group 1
ERCC2 NM_000400 -2.59excision repair cross-complementing rodent repair deficiency,
complementation group 2
ERCC5 NM_000123 -2.33excision repair cross-complementing rodent repair deficiency,
complementation group 5
RAD54B AF007866 -2.09 RAD54 homolog B (S. cerevisiae)
TDG NM_003211 -2.61 thymine-DNA glycosylase
Table 1.
by guest on June 30, 2018http://w
ww
.jbc.org/D
ownloaded from

Ohmiya, Masaki Shiota, Akihiko Kidani, Akihiko Tawara and Kimitoshi KohnoNaoya Miyamoto, Hiroto Izumi, Takako Noguchi, Yoshihiro Nakajima, Yoshihiro
cisplatin resistanceTip60 is regulated by circadian transcription factor clock and is involved in
published online May 5, 2008J. Biol. Chem.
10.1074/jbc.M802332200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on June 30, 2018http://w
ww
.jbc.org/D
ownloaded from


















