
Time 3 - Forsiden - Universitetet i Oslo · 2 Hvorfor bevare mangfoldet? • Mange arter avhenger...
42
Time 3 Biodiversitet og ”innvandring”
Transcript of Time 3 - Forsiden - Universitetet i Oslo · 2 Hvorfor bevare mangfoldet? • Mange arter avhenger...

Time 3
Biodiversitet og ”innvandring”

2
Hvorfor bevare mangfoldet?
•
Mange arter avhenger av hverandre
•
Artsrike økosystemer er mer robuste ved
endringer i miljøet
•
Arter representerer fremtidig ukjente verdier
•
Etisk begrunnelse
•
Mangfold kontra sjeldne arter

3
Biologisk mangfold Figur: Ekstam
& Forshed
1996
Gen-, art-, biotop-
og
landskapsmangfold

4
StorblåfjærFoto: Anders Bryn, Beitevoll, Vega, Nordland 2003
Genetisk mangfold

5
Artsmangfold

6

7
Det er ”farlig å
være få”
•
Små
populasjoner
har
begrenset
levetid
•
De er lettere ofre for tilfeldigheter; f. eks. tilfeldige svinginger i populasjonsstørrelsen (og tilfeldige veldig lave tettheter)
populasjonstørrelse
Årlig sannsynlighet
for å
dø
ut
0%
100%Ødeleggelse av
habitat
•
De er utsatt for innavlAB x AA
AB x AB
BB
AB x AA
AB x AB
BB
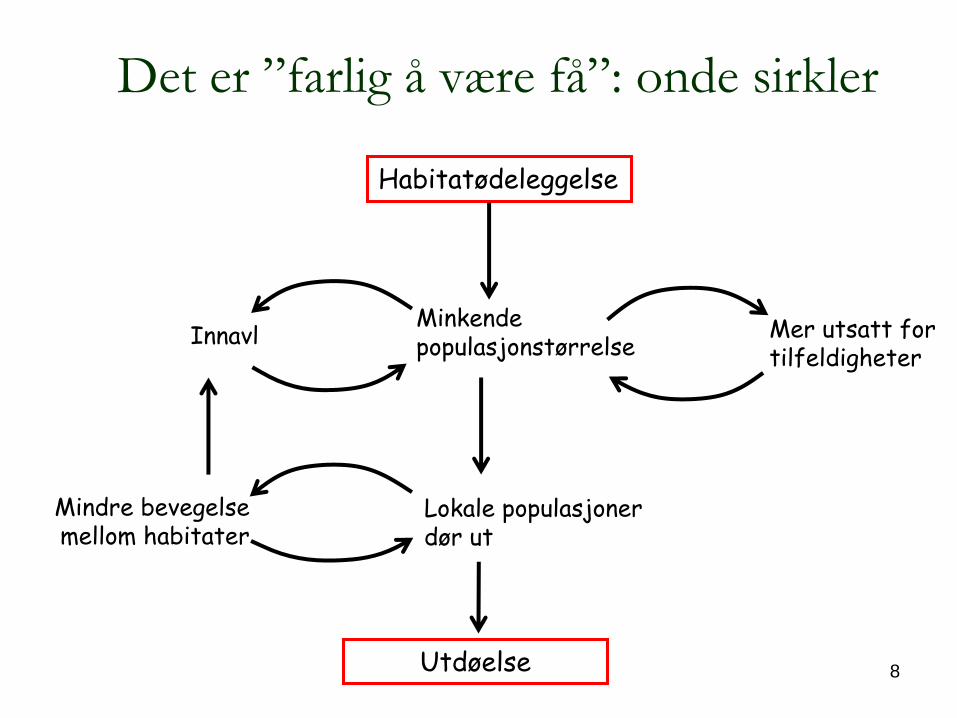
8
Det er ”farlig å
være få”: onde sirkler
Minkende populasjonstørrelseInnavl
Lokale populasjoner dør ut
Mindre bevegelse mellom habitater
Habitatødeleggelse
Utdøelse
Mer utsatt for tilfeldigheter

9
Biologisk mangfold –
tid. Vi så
på
rom i går.
•
Langt tidsperspektiv:–
eks platetektonikk / istider / migrasjon
–
eks evolusjon
•
Historisk tidsperspektiv–
eks kontinuitet / source
-
sink
–
eks suksesjon
•
Kort tidsperspektiv–
Eks caruosel
/ naturlig dynamikk
–
Meta-populasjonsdynamikk

10
Norge - biodiversitet

11
Norge arter
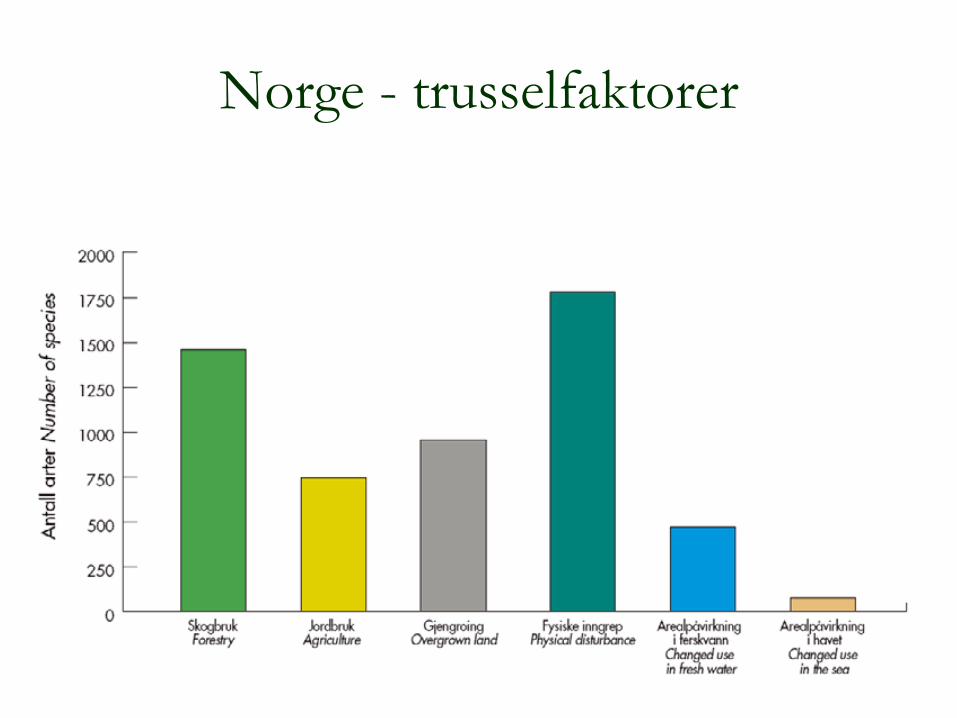
12
Norge - trusselfaktorer

13
Landskapsvariasjon = mange arter
•
Resultater fra 20 3Q flater på
Østlandet
Antall fuglearter per flate
Heterogenitetsindeks
(Hix)
R2= 0.63
0
5
10
15
20
25
30
35
0 0.2 0.4 0.6 0.8
Antall
karplantearter
per flate
Heterogenitetsindeks
(Hix)
R2 = 0.52
0
20
40
60
80
100
120
0 0.2 0.4 0.6 0.8

14
Gjengroing og landskapsvariasjon
•
Gjengroing:–
mer vanlige biotoper
–
færre uvanlige biotoper
•
Gjengroing gir lavere landskapsvariasjon

15
Gjengroingsgrad (tid)
α-diversitet
Gjengroingsgrad (tid)
β-diversitet
Gjengroingsgrad (tid)
γ-diversitet
Gjengroing og artsvariasjon

16

17
Verden – terrestrisk biodiversitet

18
Verden – hot spotsEndemiske arter viktig
Trusselbildet er klart…

19
Verden – vann biodiversitet

20
Utdøelsesgjeld
Antall
arter
1000
500
År
etter
habitatødeleggelse
0 10 50 100
Hab
itatø
deleg
gelse
Utdøelsesgjeld
• Artene dør ikke ut umiddelbart etter at arealet reduseres
• Dette kalles utdøelsesgjeld
(extinction
debt) En del av artene vi observerer i dag er ”dødsdømte”
–
de vil
ikke greie seg i landskapet over tid

Innvandring etter istiden
•
Tabula
rasa?
•
Relikter som overlevde istiden på
nunataker?
•
Endemismer
•
Unisentriske
arter
•
Genetiske markører

22
Sør-Norsk
bjørn har mt-DNA
som ligner det i Pyrenéene. Nordlige bjørners
mt-DNA
likner de finske og russiske. Grensen går ved Storsjøen i Sverige.
Kilde: http://www.bearpr
oject.info/norsk/t
he%20brown%20b
ears.htm
og
http://no.wikipedi
a.org/wiki/Brunbj
%C3%B8rn
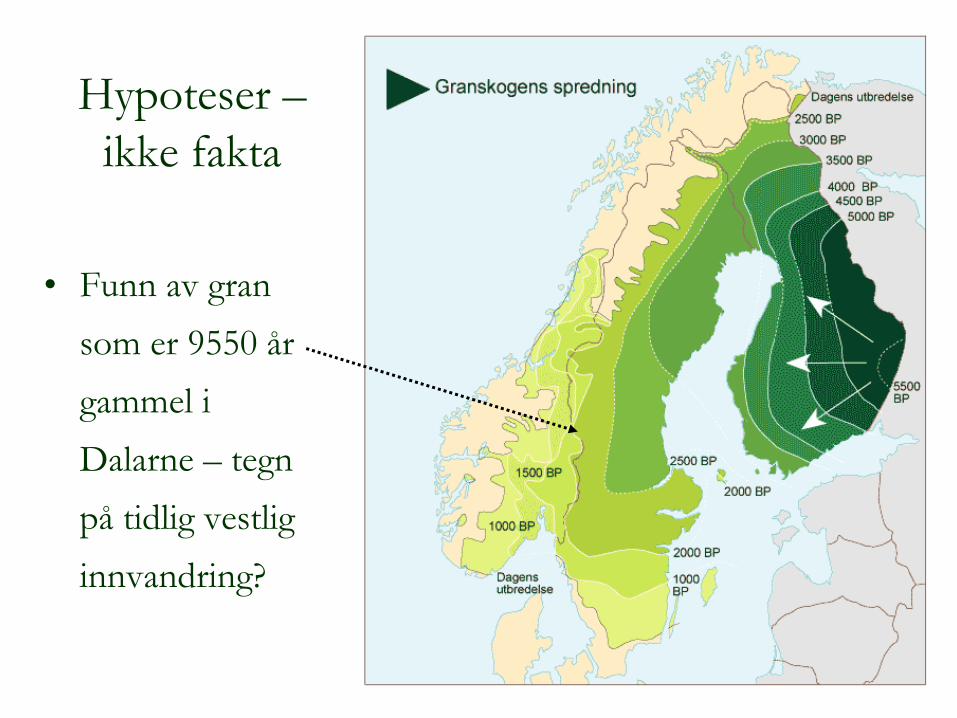
Hypoteser – ikke fakta
•
Funn av gran som er 9550 år gammel i Dalarne
–
tegn
på
tidlig vestlig innvandring?
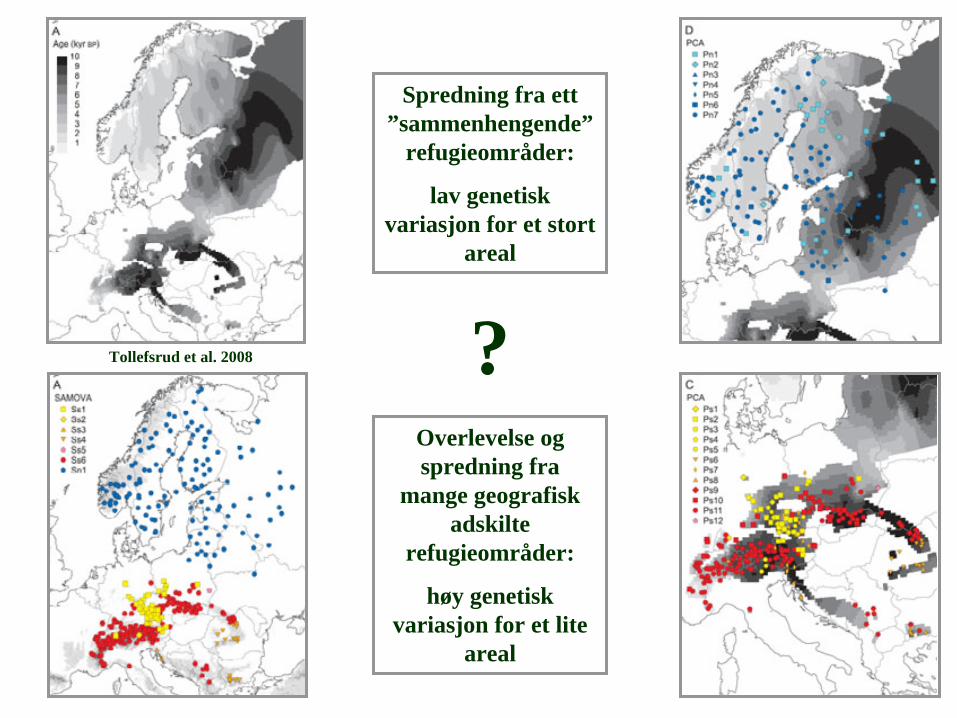
Spredning fra ett ”sammenhengende”
refugieområder:
lav genetisk variasjon for et stort
areal
Overlevelse og spredning fra
mange geografisk adskilte
refugieområder:
høy genetisk variasjon for et lite
areal
?Tollefsrud et al. 2008

Time 4Klimaendringer –
skoggrenser i Norge
som eksempel
26
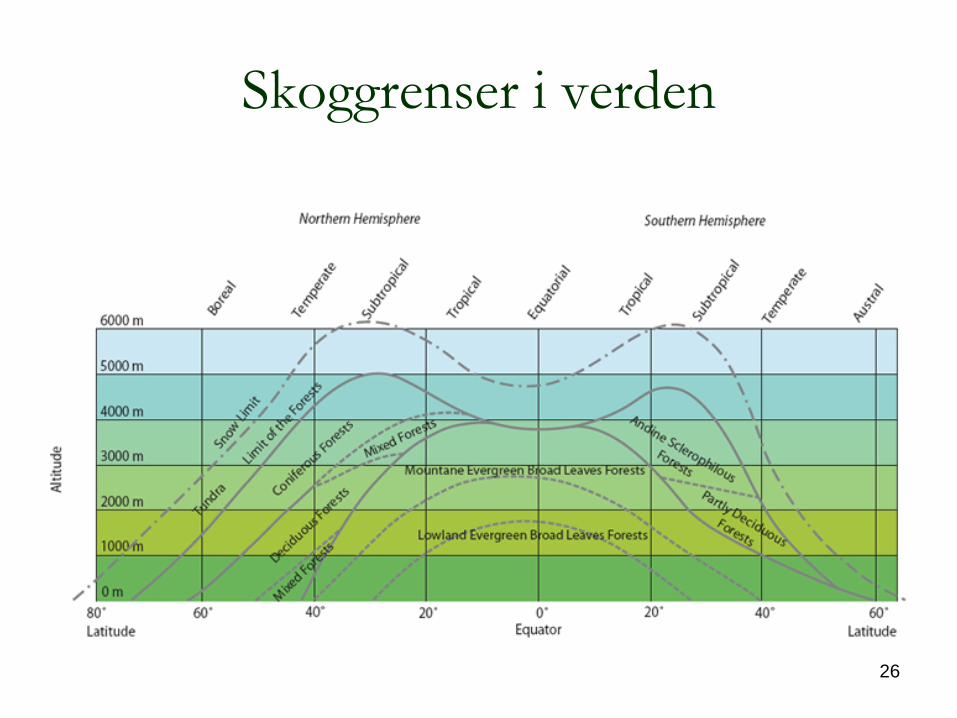
Skoggrenser i verden

Skoggrensa i Norge•
Grenser: –
artsgrense
–
tregrense–
Skoggrense
•
Skoggrenser:–
topografisk
–
klimatisk–
antropogen
–
edafisk–
biotisk
Kilde: B. Aas 1964

28Figur: Børre Aas 1969

29
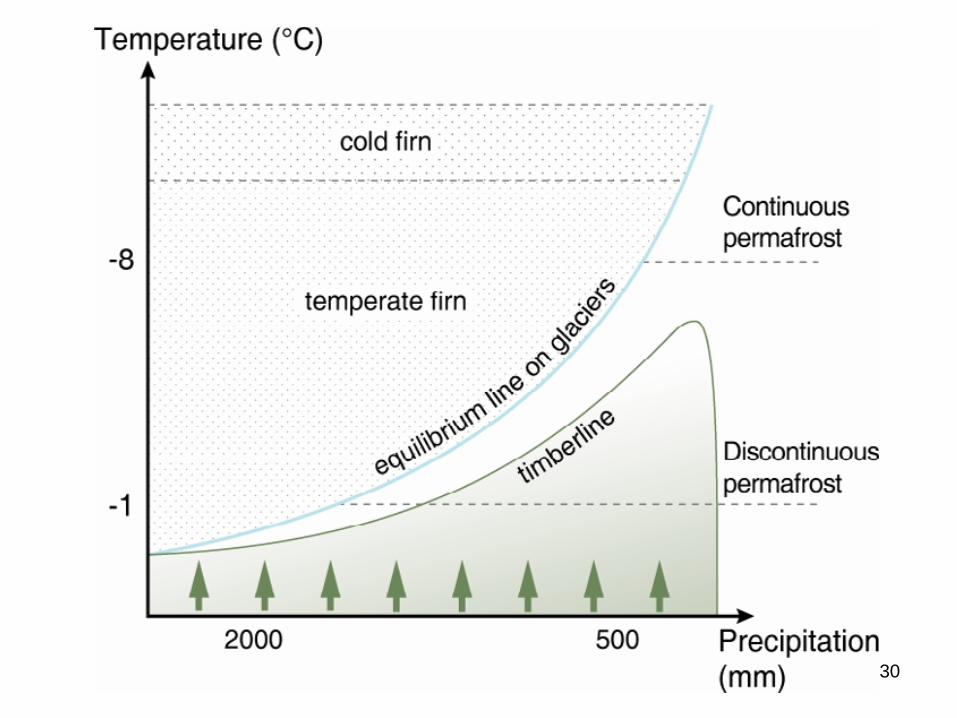
Potensielle skoggrenser
Figur: Kullmann 1990Figur: Aas & Faarlund 1988
30

31
Studieområde og mål
•
Eksemplifisere:
–
Habitat modellering
–
Skoggrenseendringer
•
Case-studies:
–
Fjellområde
i Sørøst-Norge (161 km2)
–
Sør-Norge
(210 000 km2)


33

34
Third example – caterpillar outbreakPhotos: Anders Bryn 2008

35
Skoggrense og seterdrift• Effekten av seter- og gardsdrift på skogutbredelsen er klar
• Store arealer er avskoget – mange tolker dette som ”fjellområder”
• Ny skog i høyden tolkes derfor ofte som klimabedring
1944-1959 1986-2001

36

37
Ergo så langt
•
> 97% av endringene mellom 1959 -
2001 synes å
skyldes gjengroing alene, men gjør den det?
•
Hva med det potensielle klimatiske bidraget til
gjengroing i perioden 1959 -
2001?
•
Medførte høye temperaturer raskere gjengroing?
•
Hva med de siste somrenes ekstreme temperaturer?

38
55,052,550,047,545,0Temperature sums in degrees C (6 years)
120
110
100
90
80
70
60
50
Hei
ght g
row
th s
ums
in c
m (6
yea
rs)
R2 = 0,409 P<0,001 F=54,7 Y=3,577x-91,381 D.F.=79
Meget sterk positiv sammenheng mellom tilvekst og triterm / tetraterm
400375350325300275Precipitation sums in mm (6 years)
120
110
100
90
80
70
60
50
Hei
ght g
row
th s
ums
in c
m (6
yea
rs)
R2=0,215 P<0,001 F=21,7 Y=-0,215x+158,437 D.F.=79
Negativ sammenheng mellom tilvekst og nedbør triterm / tetraterm
1995-2006 N=20
1975-1994 N=23
1960-1974 N=20
1936-1959 N=18
Growth periods
52
50
48
Ave
rage
tetr
ater
m s
ums
for 6
yea
rs
Overall mean=49,68 N=81 Error bars=95% C.I.
Ingen indikasjon på klimatisk bidrag til gjengroing 1959 - 1994
1995-2006 N=20
1975-1994 N=23
1960-1974 N=20
1936-1959 N=18
Growth periods
100
95
90
85
80
75
70
65
Mea
n he
ight
gro
wth
for 6
yea
rs in
cm
Overall mean=86,33 N=81 Error bars=95% C.I.
Svakere tilvekst 1960 - 1994 enn tidligere, øker fra 1995 - 2006

39
Ergo litt lenger
•
Sommertemperatur (tri/tetra) er viktigste ”naturlige”
forklaringsvariabel for skoggrenser og skogvekst i denne
høydesonen
•
Skoggrenseendringene fra 1959 –
1994 skyldes i all hovedsak
gjengroing
•
Indikasjoner fra skoggrenser, temperaturmålinger og tilvekst-
målinger
tyder på
økt klimatisk endringseffekt de siste 10 årene

40
Generaliserbarhet
•
Konservativt estimat av
potensielle gjengroings-
områder
i Oppland
•
Konservativt estimat av
potensielle gjengroings-
områder
i Sør-Norge

41
Oppsummering 1
•
Gjengroing etter nedlagt utmarksbruk er viktigste faktor for endringer i seter-
og fjellregionen mellom 1959 –
1995
•
Klimaendringer bidro ikke til økt gjenvekst eller hevede potensielle skoggrenser i denne perioden
•
Nylige klimaendringer antyder svakt økende gjengroingshastighet og svakt hevede skoggrenser etter særlig år 2000

42
Oppsummering 2
•
Modellering av potensiell skog viser at 15-18% av
utmarka i Sør-Norge har et potensial for skog
•
I tillegg kommer 5% med dyrka mark
•
To regioner er spesielt utsatt for gjengroing med skog:
–
Kystregionene –
50% av regionen kan gro igjen!
–
Seterregionene –
50% av all gjengroing kommer her!


















