
Relationship between body colour, feeding, and reproductive arrest under short-day development in...
7
Relationship between body colour, feeding, and reproductive arrest under short-day development in Tetranychus pueraricola (Acari: Tetranychidae) Katsura Ito • Tatsuya Fukuda • Hiroshi Hayakawa • Ryo Arakawa • Yutaka Saito Received: 16 May 2012 / Accepted: 30 January 2013 / Published online: 19 February 2013 Ó Springer Science+Business Media Dordrecht 2013 Abstract In Tetranychus spider mites (Acari: Tetranychidae), diapausing females have a conspicuous orange body colour, which is used as an indicator of diapause induction in many laboratory studies. However, to which extent body colour reflects reproductive activity is scarcely investigated. In this study, we investigated the relationship between body colour, reproductive arrest, and food intake in the inbred strain of T. pueraricola individually reared at 20 °C with a 10:14 h light: dark photoperiod. Our results showed that (1) body colour is a good indicator of reproductive arrest 11 days after adult emer- gence but does not completely reflect reproductive status at an earlier age; (2) even orange females intermittently feed, and the arrest of feeding comes after the change in body colour; and (3) reproducing females have a higher risk of death than non-reproducing females. These results suggest that measurement of diapause incidence by body colour alone may miss the variation in reproductive status in early adult life. Keywords Acari Á Diapause incidence Á Environmental variation Á Spider mite Á Tetranychidae Introduction In Tetranychus spider mites, the body colour of the females developed under short-day and low-temperature conditions becomes conspicuous orange. This is a phenotype involved in the adult diapause of spider mites (reviewed in Veerman 1985), and thus has been used as an indicator of diapause induction in many experimental studies (e.g. Veerman 1977; K. Ito (&) Á T. Fukuda Á R. Arakawa Faculty of Agriculture, Kochi University, Monobeotsu 200, Nankoku, Kochi, Japan e-mail: [email protected] H. Hayakawa National Institute for Agro-Environmental Science, 3-1-3 Kanondai, Tsukuba, Japan Y. Saito Graduate school of Agriculture, Hokkaido University, 9 nishi 9 kita, kita-ku, Sapporo, Japan 123 Exp Appl Acarol (2013) 60:471–477 DOI 10.1007/s10493-013-9660-3
-
Upload
yutaka-saito -
Category
Documents
-
view
212 -
download
0
Transcript of Relationship between body colour, feeding, and reproductive arrest under short-day development in...

Relationship between body colour, feeding,and reproductive arrest under short-day developmentin Tetranychus pueraricola (Acari: Tetranychidae)
Katsura Ito • Tatsuya Fukuda • Hiroshi Hayakawa • Ryo Arakawa •
Yutaka Saito
Received: 16 May 2012 / Accepted: 30 January 2013 / Published online: 19 February 2013� Springer Science+Business Media Dordrecht 2013
Abstract In Tetranychus spider mites (Acari: Tetranychidae), diapausing females have a
conspicuous orange body colour, which is used as an indicator of diapause induction in
many laboratory studies. However, to which extent body colour reflects reproductive
activity is scarcely investigated. In this study, we investigated the relationship between
body colour, reproductive arrest, and food intake in the inbred strain of T. pueraricola
individually reared at 20 �C with a 10:14 h light: dark photoperiod. Our results showed
that (1) body colour is a good indicator of reproductive arrest 11 days after adult emer-
gence but does not completely reflect reproductive status at an earlier age; (2) even orange
females intermittently feed, and the arrest of feeding comes after the change in body
colour; and (3) reproducing females have a higher risk of death than non-reproducing
females. These results suggest that measurement of diapause incidence by body colour
alone may miss the variation in reproductive status in early adult life.
Keywords Acari � Diapause incidence � Environmental variation � Spider mite �Tetranychidae
Introduction
In Tetranychus spider mites, the body colour of the females developed under short-day and
low-temperature conditions becomes conspicuous orange. This is a phenotype involved in
the adult diapause of spider mites (reviewed in Veerman 1985), and thus has been used as
an indicator of diapause induction in many experimental studies (e.g. Veerman 1977;
K. Ito (&) � T. Fukuda � R. ArakawaFaculty of Agriculture, Kochi University, Monobeotsu 200, Nankoku, Kochi, Japane-mail: [email protected]
H. HayakawaNational Institute for Agro-Environmental Science, 3-1-3 Kanondai, Tsukuba, Japan
Y. SaitoGraduate school of Agriculture, Hokkaido University, 9 nishi 9 kita, kita-ku, Sapporo, Japan
123
Exp Appl Acarol (2013) 60:471–477DOI 10.1007/s10493-013-9660-3

Ignatowicz and Helle 1986; Goka and Takafuji 1990, 1991; Takafuji et al. 1991; Kroon
et al. 1998; Koveos et al. 1999; Ito 2003; Kawakami et al. 2009; Suzuki and Takeda 2009).
However, what ‘diapause’ means in them is often unclear, because the change of colour
is only one of the physiological attributes in diapause. The most important character in the
ecological context is reproductive arrest. Nevertheless, to which extent the colour change
reflects the reproductive activity has scarcely been investigated. As Saito et al. (2005)
pointed out, the most reliable criterion to determine reproductive diapause is the pattern of
reproduction itself, and the body colour is useful only after this is verified to be parallel to
the cessation of reproduction. Recent studies suggest that the arrest of egg deposition is not
accompanied by a change in pigmentation in Stigmaeopsis species (Saito et al. 2002,
2005). Following field observation showed that the change in pigmentation occurs
[1 month after the cessation of egg production in S. longus in southwestern Japan (Ito
unpublished data). These results suggest that body pigmentation and reproductive arrest
might generally be governed by separate physiological mechanisms. Thus, the use of body
colour to infer reproductive status needs to be re-examined.
The aim of this study is to clarify the relationship between body colour, reproductive
arrest, and feeding under constant day length and temperature conditions by using the spider
mite Tetranychus pueraricola Ehara and Gotoh as a model species. This species is considered
to be closely related to the red form of T. urticae (Suwa and Gotoh 2006); moreover, it is
widely distributed throughout Japan and infest kudzu vine (Pueraria lobata [Willd.]) in the
wild, on which it makes conspicuous red scars on the leaf surface due to mass feeding (Gotoh
and Tokioka 1996; Gotoh et al. 2004). This species shows geographic variation in the ten-
dency to ‘diapause’, and crossing experiments with simple Mendelian inheritance have
shown that ‘non-diapause’ is a dominant character over ‘diapause’ (Suwa and Gotoh 2006).
However, these conclusions are based on the body colour, so that the correlation between
body colour and reproduction is unknown. To enable the clear assessment of environmental
effects, we used a genetically homogeneous inbred line established from 1 female.
Materials and methods
Mites
The strain of T. pueraricola mite used in this study was derived from one female obtained
from a kudzu-infesting population, which was collected from Hitsuzan Park, Hitsuzan,
Kochi, Japan (N 33.551, E 133.536, WGS84) on 7 December 2010. The species was
identified primarily from the characteristic shape of the knob on the aedeagus in males, the
body colour of adults, and the red scars along leaf veins of the host plant (Ehara and Gotoh
2009). The isofemale strain was established by repeated full-sib (brother-sister) mating for
[12 generations on 2 9 2-cm leaf squares of kidney bean (Phaseolus vulgaris L.)
maintained at 25 �C with a 16:8 h light: dark (L:D) photoperiod. Thereafter, individuals
were allowed to mate freely and were kept at 20 �C with a 16:8 h L:D photoperiod. All
rearing and experimental procedures used the ordinary leaf-disk method with water-soaked
cotton pads (Ehara and Gotoh 2009).
Diapause induction
Twenty adult females were placed on a bean leaf and allowed to oviposit for 1 day while
being maintained at 25 �C with a 16:8 h L:D photoperiod. After the removal of females,
472 Exp Appl Acarol (2013) 60:471–477
123

the culture was kept at 20 �C with a 10:14 h L:D photoperiod, which reflected the October
(autumn) field conditions at the collection site of this population.
On days 13 and 14 each, 54 newly emerged adult females were individually isolated on
a 1 9 1-cm leaf square. The survival rate and the day of first oviposition were checked
every day until day 28. Every 7 days, mites were transferred to new leaf squares, and the
number of deposited eggs was recorded. If mites were in reproductive arrest, the presence
or absence of leaf scars (made by feeding) was recorded (on days 14, 21, and 28). Body
colour was checked every day, and the day at which mites became orange was recorded.
Statistical analysis
Survival curves were plotted for all adult females and each colour morph by using the
Kaplan–Meier method. A log-rank test was used to compare the curves of each morph. A
two-way ANOVA was used to test the effects of body colour and week on the egg
production, and Ryan’s method for multiple comparisons of proportions (Ryan 1960) was
used to compare the proportion of feeding females between different weeks. All statistical
analyses were conducted using R version 2.13.0 (R Development Core Team 2011).
Results
30 out of 108 females underwent a change in body colour to orange at some point during
the 28-days observation period, whereas the others remained non-pigmented within the
observation period. The change in body colour began after 7 days of adult life (days
20–21) and become conspicuous until 11 days (days 25–26). The observed orange colour
was distinct from the red colour that starved females exhibit (this is typically much weaker
than the colour of diapausing).
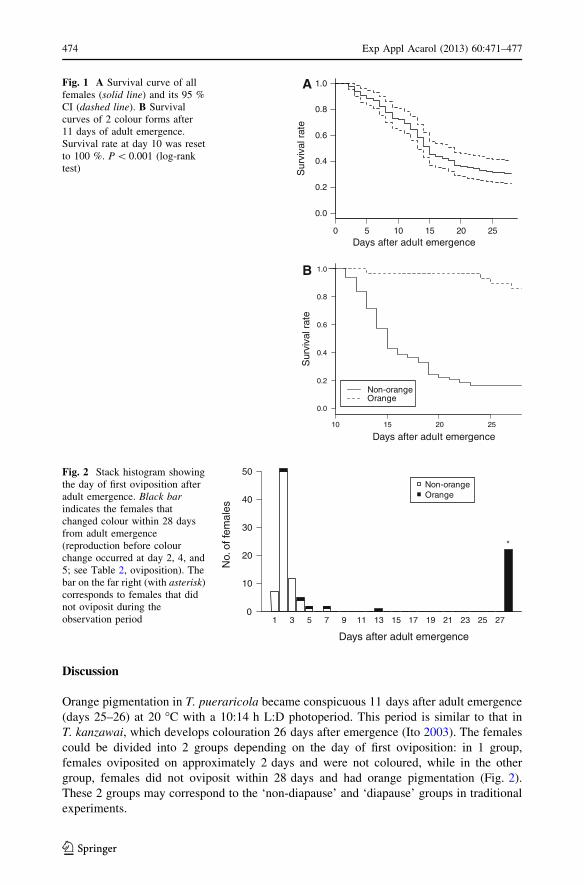
The survival curves of all females and of each colour morph (‘orange’ and ‘non-
orange’) 11 days after adult emergence are shown in Fig. 1. Orange females had a sig-
nificantly lower risk of death than non-orange females (log-rank test, P \ 0.001). By day
11, 33 non-orange females had died, 29 of which produced C1 egg(s) in 2.103 ± 0.143
(mean ± SE) days after adult emergence.
Figure 2 shows the histogram of adult age of first reproduction. Distribution of the day
was bimodal. One group, which mostly included non-orange females, was distributed
around day 2. The other group, which included only orange females, did not oviposit
within 28 days (rightmost bar in Fig. 2). There were 3 intermediate individuals that started
oviposition on days 2–5 but later arrested oviposition and became orange until day 28
(Table 2, oviposition).
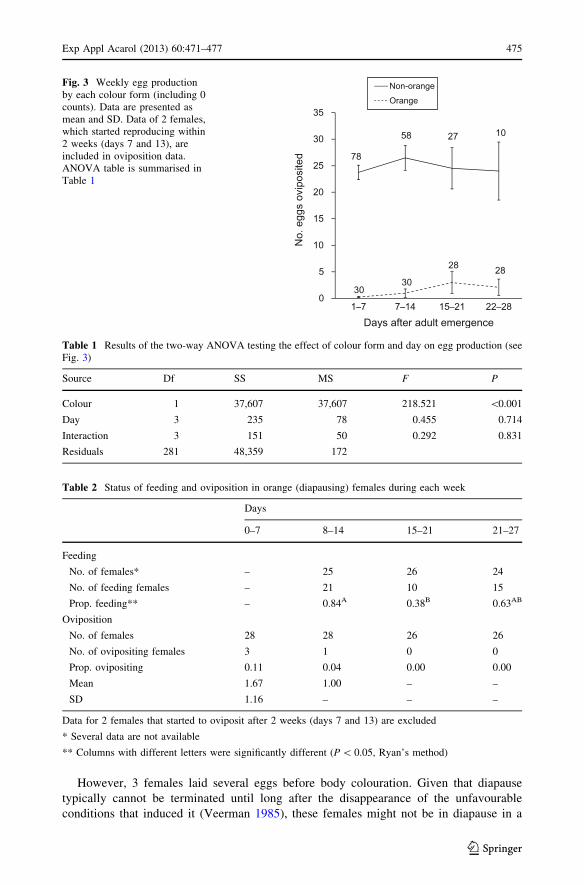
The weekly mean number of eggs produced by each colour morph (including 0 counts)
is presented in Fig. 3. The summary of ANOVA is presented in Table 1, showing a
significant effect of only body colour. Non-orange females constantly produced an average
of ca. 25 eggs, whereas orange females produced \5 eggs; A slight increase in egg
production after day 15 was attributed to 2 females that began oviposition within 15 days
(Fig. 2, day 7 and 13 of orange females). Data excluding these females is summarised in
Table 2 (oviposition).
All 23 orange females for which a complete set of feeding data are available were feeding
at some time during the observation period, and of these 4 females fed every week. The
proportion of females feeding significantly decreased during 15–21 days when compared
with the previous week (Table 2, feeding). All non-orange females (n = 78) were feeding.
Exp Appl Acarol (2013) 60:471–477 473
123
Discussion
Orange pigmentation in T. pueraricola became conspicuous 11 days after adult emergence
(days 25–26) at 20 �C with a 10:14 h L:D photoperiod. This period is similar to that in
T. kanzawai, which develops colouration 26 days after emergence (Ito 2003). The females
could be divided into 2 groups depending on the day of first oviposition: in 1 group,
females oviposited on approximately 2 days and were not coloured, while in the other
group, females did not oviposit within 28 days and had orange pigmentation (Fig. 2).
These 2 groups may correspond to the ‘non-diapause’ and ‘diapause’ groups in traditional
experiments.
0 5 10 15 20 25
0.0
0.2
0.4
0.6
0.8
1.0
Days after adult emergence
Sur
viva
l rat
e
10 15 20 25
0.0
0.2
0.4
0.6
0.8
1.0
Days after adult emergence
Sur
viva
l rat
e
Non-orangeOrange
A
B
Fig. 1 A Survival curve of allfemales (solid line) and its 95 %CI (dashed line). B Survivalcurves of 2 colour forms after11 days of adult emergence.Survival rate at day 10 was resetto 100 %. P \ 0.001 (log-ranktest)
1 3 5 7 9 11 13 15 17 19 21 23 25 27
Days after adult emergence
No.
of f
emal
es
0
10
20
30
40
50Non-orangeOrange
*
Fig. 2 Stack histogram showingthe day of first oviposition afteradult emergence. Black barindicates the females thatchanged colour within 28 daysfrom adult emergence(reproduction before colourchange occurred at day 2, 4, and5; see Table 2, oviposition). Thebar on the far right (with asterisk)corresponds to females that didnot oviposit during theobservation period
474 Exp Appl Acarol (2013) 60:471–477
123
However, 3 females laid several eggs before body colouration. Given that diapause
typically cannot be terminated until long after the disappearance of the unfavourable
conditions that induced it (Veerman 1985), these females might not be in diapause in a
Fig. 3 Weekly egg productionby each colour form (including 0counts). Data are presented asmean and SD. Data of 2 females,which started reproducing within2 weeks (days 7 and 13), areincluded in oviposition data.ANOVA table is summarised inTable 1
Table 1 Results of the two-way ANOVA testing the effect of colour form and day on egg production (seeFig. 3)
Source Df SS MS F P
Colour 1 37,607 37,607 218.521 \0.001
Day 3 235 78 0.455 0.714
Interaction 3 151 50 0.292 0.831
Residuals 281 48,359 172
Table 2 Status of feeding and oviposition in orange (diapausing) females during each week
Days
0–7 8–14 15–21 21–27
Feeding
No. of females* – 25 26 24
No. of feeding females – 21 10 15
Prop. feeding** – 0.84A 0.38B 0.63AB
Oviposition
No. of females 28 28 26 26
No. of ovipositing females 3 1 0 0
Prop. ovipositing 0.11 0.04 0.00 0.00
Mean 1.67 1.00 – –
SD 1.16 – – –
Data for 2 females that started to oviposit after 2 weeks (days 7 and 13) are excluded
* Several data are not available
** Columns with different letters were significantly different (P \ 0.05, Ryan’s method)
Exp Appl Acarol (2013) 60:471–477 475
123

strict sense (or quite shallow diapause). In any case, this result indicates that body colour
reflects the reproductive status in the later adult life but does not necessarily reflect that in
the early life. In the congener T. kanzawai, reproductive arrest and a change in body
colouration occurred if day length conditions were changed from long to short at the time
of adult emergence (Saito et al. 2005). The present study indicates that this change can
occur even under constant day length conditions in T. pueraricola. In addition, because
most of the females dying before pigmentation were producing eggs, exclusion of dead
females from experimental count may underestimate the degree of real reproductive
diapause.
Notably, even orange females frequently feed after the development of colouration.
Orange females isolated in cages and preserved in cool conditions tend to desiccate in
T. telarius (McEnroe 1961) and often have lower fecundity following diapause in
T. kanzawai (Ito 2004). Thus, feeding activity might compensate for the loss of body water,
allowing physiological conditions for reproduction to be maintained until diapause has
ended. The relationship between body colour, reproductive arrest, and feeding, as observed
in this study, is shown in Fig. 4.
In mass-rearing experiments, females are allowed to lay eggs for several days and are
maintained under short-day conditions until 1–2 weeks after adult emergence, following
which the proportion of orange coloured females is assumed to reflect the incidence of
diapause. Although such an assumption may be appropriate when evaluating the incidence of
reproductive arrest, it could ignore the variation in reproductive status during early adult life.
Acknowledgments We appreciate Dr. T. Gotoh for kindly providing information on T. pueraricola thisspecies. We thank Dr. H. Numata and Dr. S.G. Goto for providing useful information on insect diapause. Wethank Mr. Y. Che, Mr. Y. Paku, and Ms. K. Tamura for assistance with experiments.
References
Development Core Team R (2011) R: a language and environment for statistical computing. R Foundationfor Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0, URL http://www.R-project.org/
Ehara S, Gotoh T (2009) Colored guide to the plant mites of Japan. Zenkoku Noson Kyoiku Kyokai, Tokyo(in Japanese)
Goka K, Takafuji A (1990) Genetical studies on the diapause of the two-spotted spider mite, Tetranychusurticae Koch (1). Appl Entomol Zool 25:119–125
Goka K, Takafuji A (1991) Genetical studies on the diapause of the two-spotted spider mite Tetranychusurticae Koch (2). Appl Entomol Zool 26:77–84
Gotoh T, Tokioka T (1996) Genetic compatibility among diapausing red, non-diapausing red and diapausinggreen forms of the two-spotted spider mite, Tetranychus urticae Koch (Acari: Tetranychidae). Jpn JEntomol 64:215–225
Pigmentation
Reproductivearrest
Cessation offeeding
Fig. 4 Venn diagram aboutrelationship between diapausecharacteristics
476 Exp Appl Acarol (2013) 60:471–477
123

Gotoh T, Suwa A, Kitashima Y, Rezk HA (2004) Developmental and reproductive performance of Tetr-anychus pueraricola Ehara and Gotoh (Acari: Tetranychidae) at four constant temperatures. ApplEntomol Zool 39:675–682
Ignatowicz S, Helle W (1986) Genetics of diapause suppression in the two-spotted spider mite, Tetranychusurticae Koch. Exp Appl Acarol 2:161–172
Ito K (2003) Effect of leaf condition on diapause induction of a Kanzawa spider mite Tetranychus kanzawaiKishida (Acari: Tetranychidae) population on tea plants. Appl Entomol Zool 38:559–563
Ito K (2004) Deteriorating effects of diapause duration on postdiapause life history traits in the Kanzawaspider mite. Physiol Entomol 29:453–457
Kawakami Y, Numata H, Ito K, Goto SG (2009) Dominant and recessive inheritance patterns of diapause inthe two-spotted spider mite Tetranychus urticae. J Hered 101:20–25
Koveos DS, Veerman A, Broufas GD, Exarhou A (1999) Altitudinal and latitudinal variation in diapausecharacteristics in the spider mite Tetranychus urticae Koch. Entomol Sci 2:607–613
Kroon A, Veenendaal RL, Veerman A (1998) Response to photoperiod during diapause development in thespider mite Tetranychus urticae. J Insect Physiol 44:271–277
McEnroe WD (1961) The control of water loss by the two-spotted spider mite (Tetranychus telarius). AnnEntomol Soc Am 54:883–887
Ryan TA (1960) Significance tests for multiple comparison of proportions, variances and other statistics.Psychol Bull 57:318–328
Saito Y, Sakagami T, Sahara K (2002) Differences in diapause attributes between two clinal forms dis-tinguished by male-to-male aggression in a subsocial spider mite, Schizotetranychus miscanthi Saito.Ecol Res 17:645–653
Saito Y, Ito K, Sakagami T (2005) Imaginal induction of diapause in several adult-diapausing spider mites.Physiol Entomol 30:96–101
Suwa A, Gotoh T (2006) Geographic variation in diapause induction and mode of diapause inheritance inTetranychus pueraricola. J Appl Entomol 130:329–335
Suzuki T, Takeda M (2009) Diapause-inducing signals prolong nymphal development in the two-spottedspider mite Tetranychus urticae. Physiol Entomol 34:278–283
Takafuji A, So PM, Tsuno N (1991) Inter- and intra-population variations in diapause attribute of the two-spotted spider mite, Tetranychus urticae Koch, in Japan. Res Popul Ecol 33:331–344
Veerman A (1977) Aspects of the induction of diapause in a laboratory strain of the mite Tetranychusurticae. J Insect Physiol 23:703–711
Veerman A (1985) Diapause. In: Helle W, Sabelis MW (eds). Spider mites. Their biology, natural enemiesand control, 1A. Elsevier, Amsterdam, pp 279–316
Exp Appl Acarol (2013) 60:471–477 477
123


















