
Programmed cell death. Apoptose - Københavns … kapitel 18... · 18_18_sculpts_digits.jpg...
46
Programmed cell death. Apoptose (betyder at falde af på græsk) Alberts. Essential Cell Biology. Kap.18, side 625
Transcript of Programmed cell death. Apoptose - Københavns … kapitel 18... · 18_18_sculpts_digits.jpg...

Programmed cell death.
Apoptose (betyder at falde af på græsk)
Alberts. Essential Cell Biology.
Kap.18, side 625
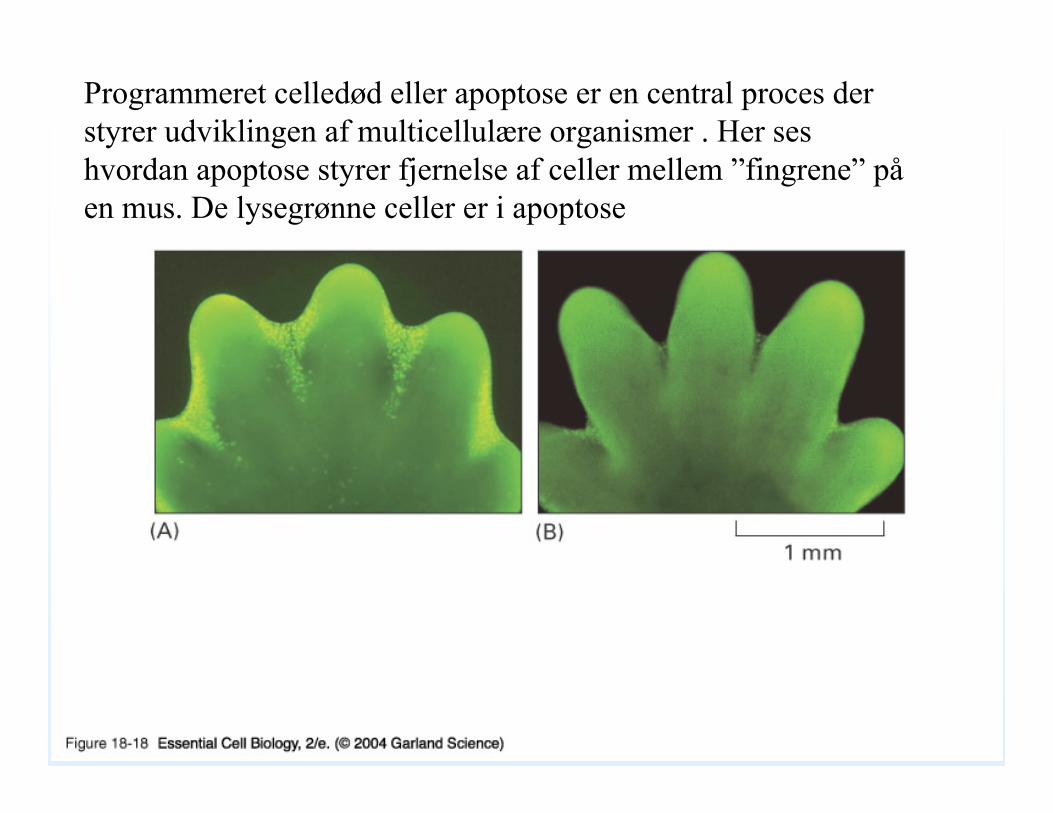
18_18_sculpts_digits.jpgProgrammeret celledød eller apoptose er en central proces der styrer udviklingen af multicellulære organismer . Her ses hvordan apoptose styrer fjernelse af celler mellem ”fingrene” påen mus. De lysegrønne celler er i apoptose
Downloaded from: StudentConsult (on 8 October 2005 02:42 PM)© 2005 Elsevier
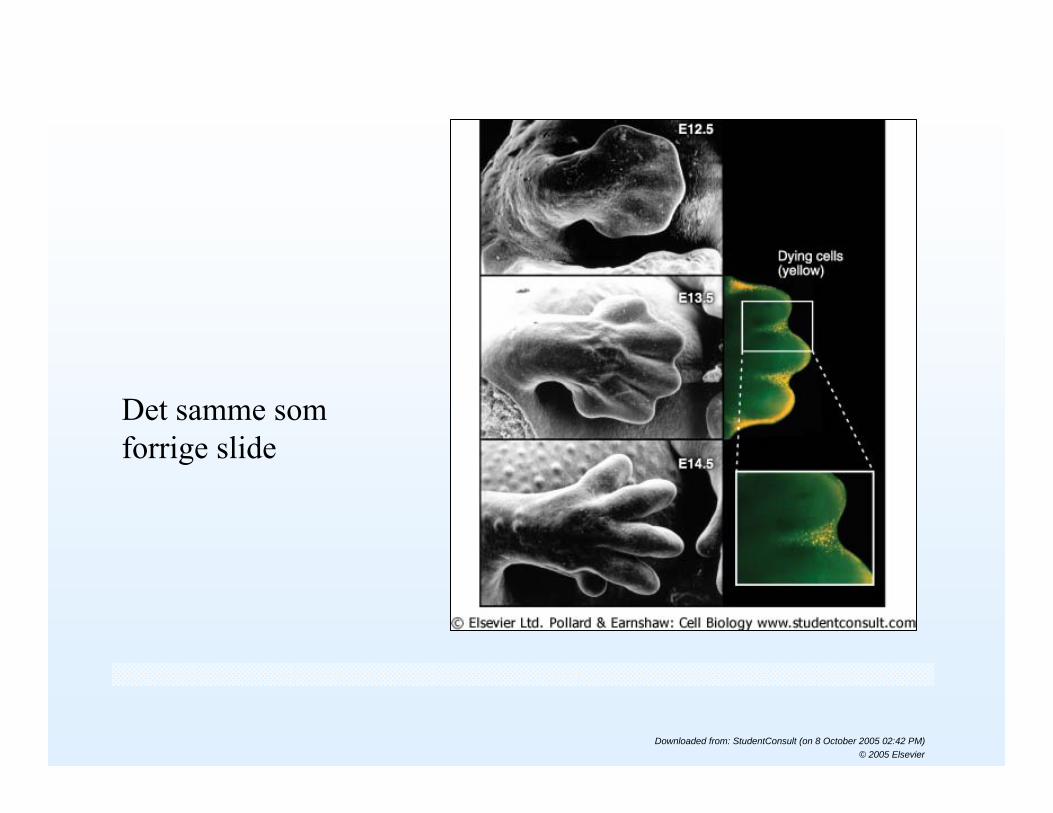
Det samme som forrige slide

Utrolig mange celler dør (ved apoptose) under udviklingen. Omkring halvdelen af alle nerveceller dør næsten med det samme. Nogle celler dør fordi de ikke længere skal bruges som f.eks haletudsens hale. I nervesystemet under udvikling er det for at tilpasse antallet af nerveceller til de celler der skal have nervekontakt. I den voksne organisme sørger apoptosen for at holde antallet af celler konstant så celledeling og apoptose matches præcist. Hvis ikke ville vævene vokse eller mindskes.
Fra Haletudse til frø
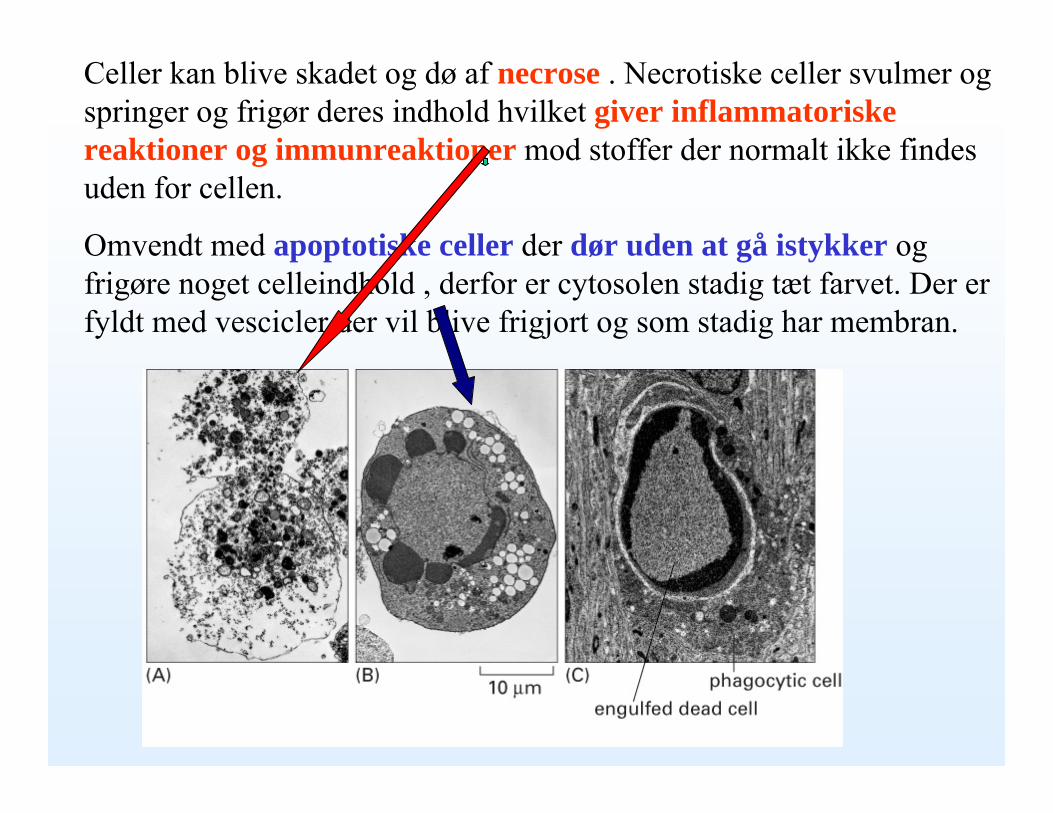
Celler kan blive skadet og dø af necrose . Necrotiske celler svulmer og springer og frigør deres indhold hvilket giver inflammatoriske reaktioner og immunreaktioner mod stoffer der normalt ikke findes uden for cellen.
Omvendt med apoptotiske celler der dør uden at gå istykker og frigøre noget celleindhóld , derfor er cytosolen stadig tæt farvet. Der er fyldt med vescicler der vil blive frigjort og som stadig har membran.
Downloaded from: StudentConsult (on 8 October 2005 02:42 PM)© 2005 Elsevier
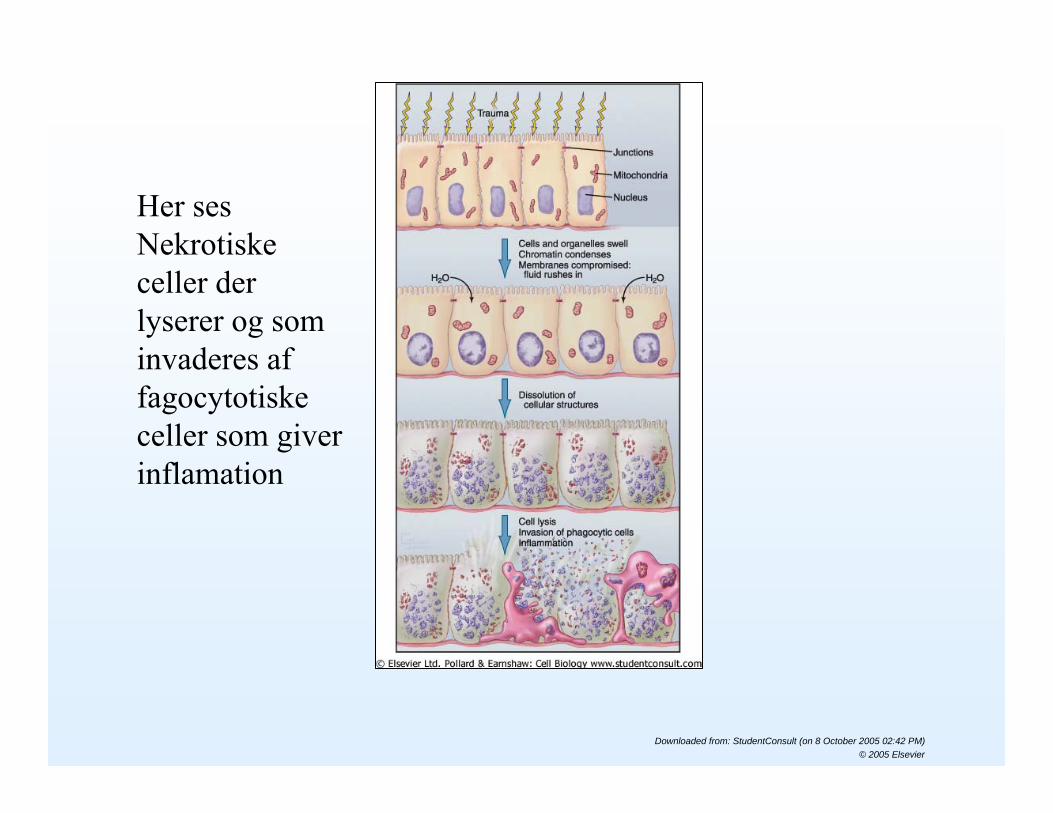
Her ses Nekrotiske celler der lyserer og som invaderes af fagocytotiskeceller som giver inflamation
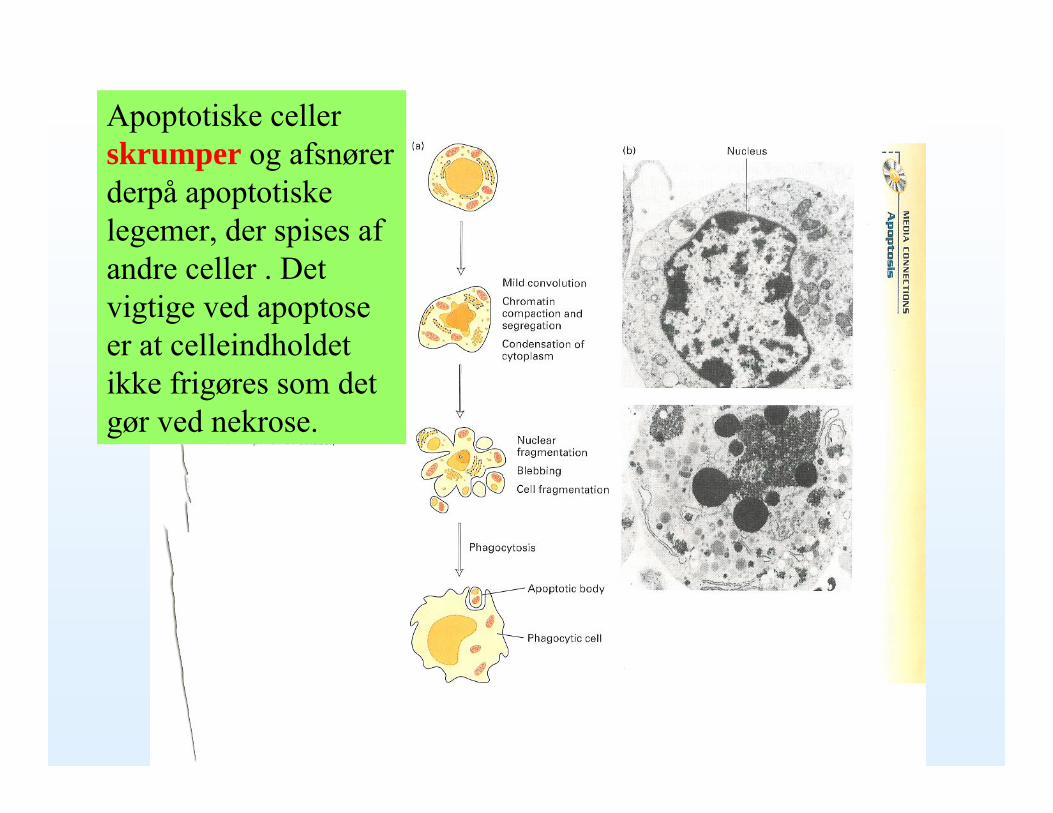
Apoptotiske celler skrumper og afsnører derpå apoptotiske legemer, der spises af andre celler . Det vigtige ved apoptose er at celleindholdet ikke frigøres som det gør ved nekrose.
Downloaded from: StudentConsult (on 8 October 2005 02:42 PM)© 2005 Elsevier
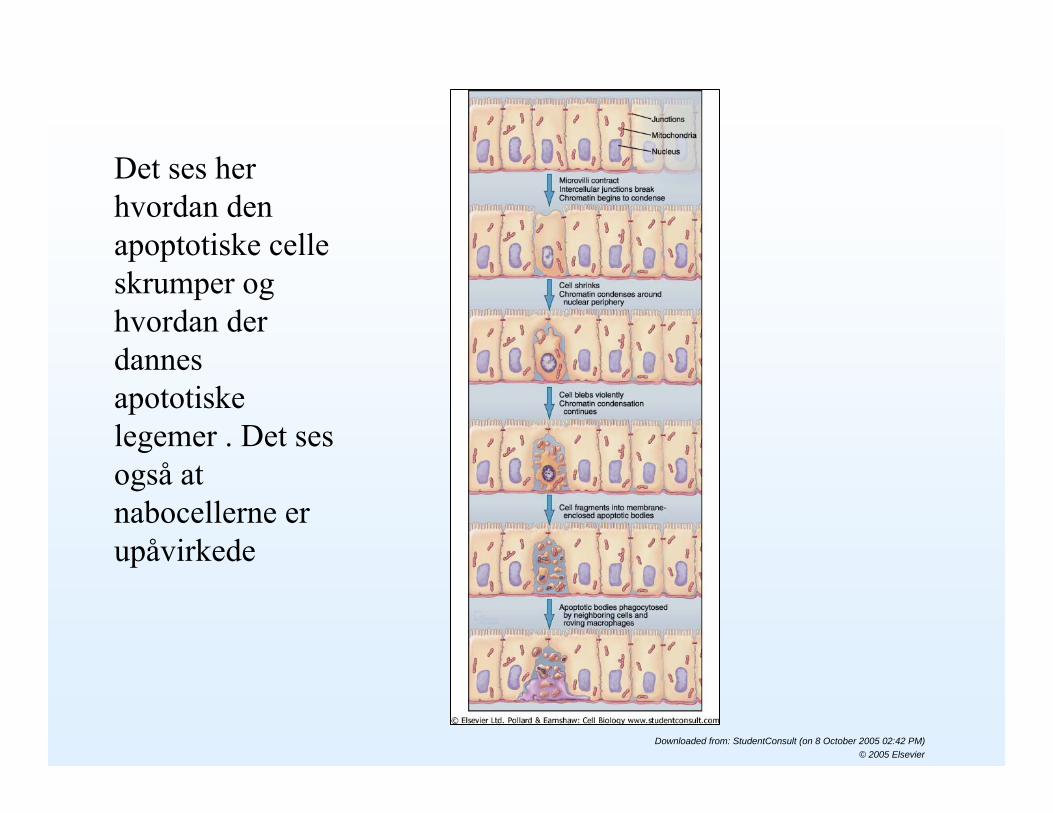
Det ses her hvordan den apoptotiske celle skrumper og hvordan der dannes apototiskelegemer . Det ses også at nabocellerne er upåvirkede
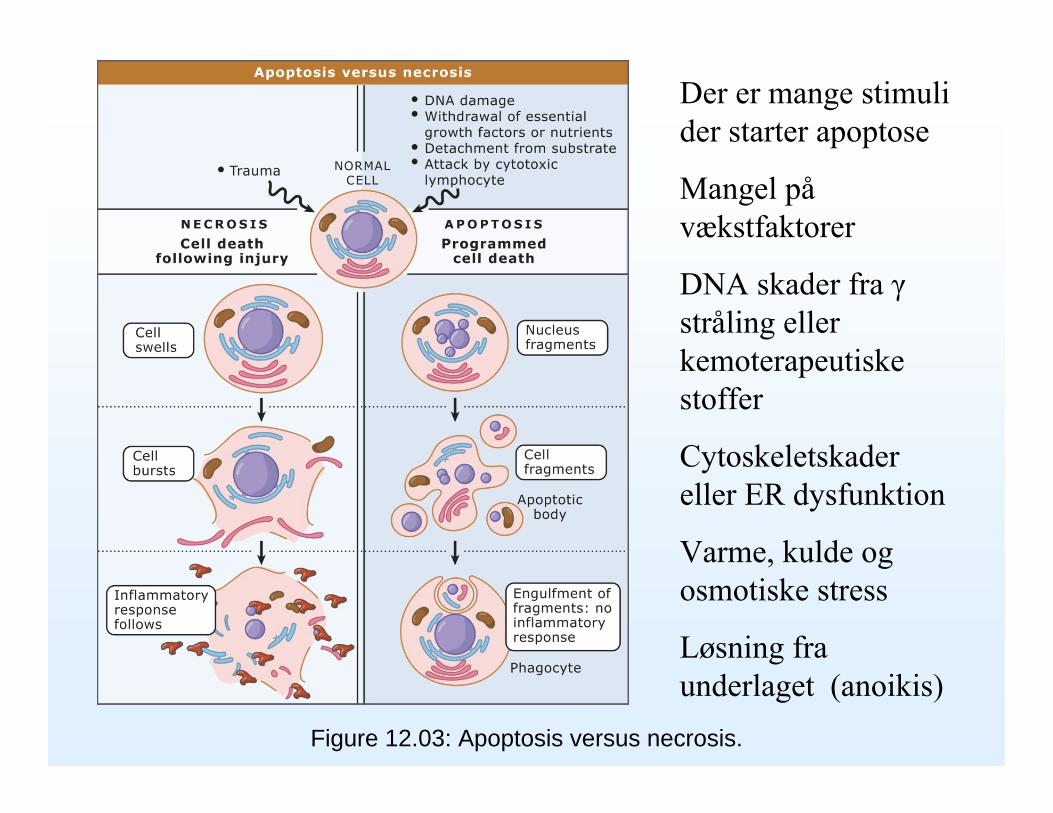
Figure 12.03: Apoptosis versus necrosis.
Der er mange stimuli der starter apoptose
Mangel påvækstfaktorer
DNA skader fra γstråling eller kemoterapeutiske stoffer
Cytoskeletskadereller ER dysfunktion
Varme, kulde og osmotiske stress
Løsning fra underlaget (anoikis)
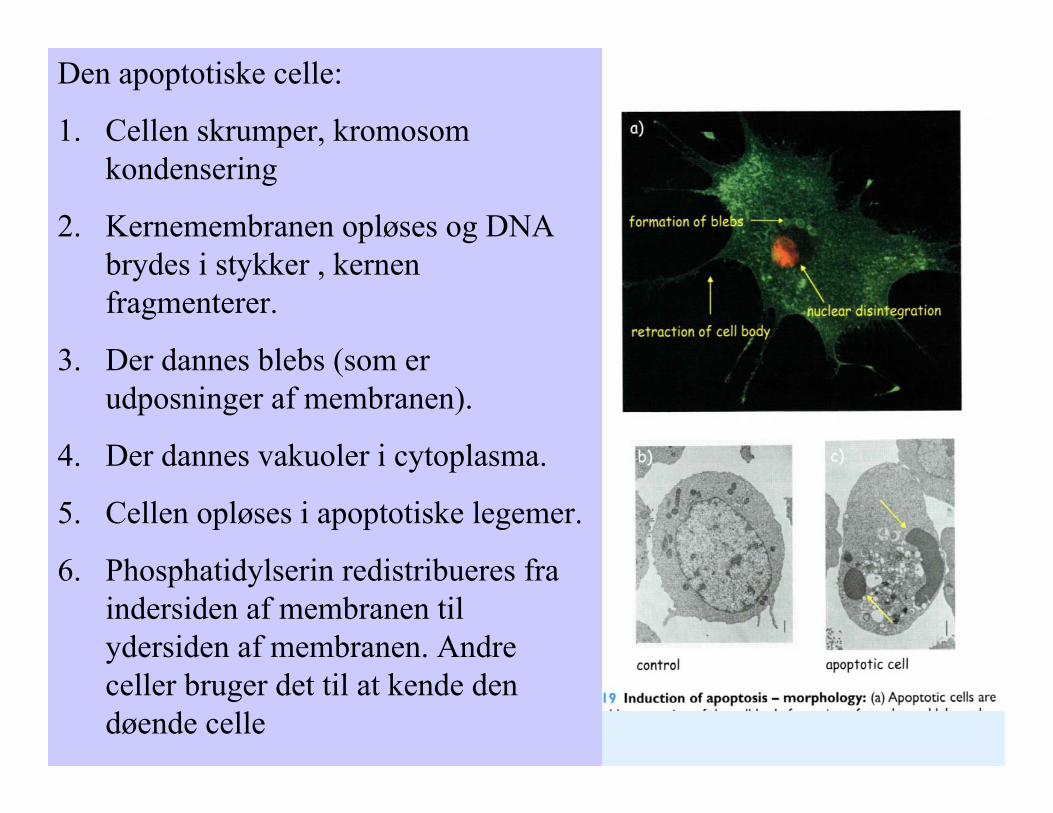
Den apoptotiske celle:
1. Cellen skrumper, kromosom kondensering
2. Kernemembranen opløses og DNA brydes i stykker , kernen fragmenterer.
3. Der dannes blebs (som er udposninger af membranen).
4. Der dannes vakuoler i cytoplasma.
5. Cellen opløses i apoptotiske legemer.
6. Phosphatidylserin redistribueres fra indersiden af membranen til ydersiden af membranen. Andre celler bruger det til at kende den døende celle
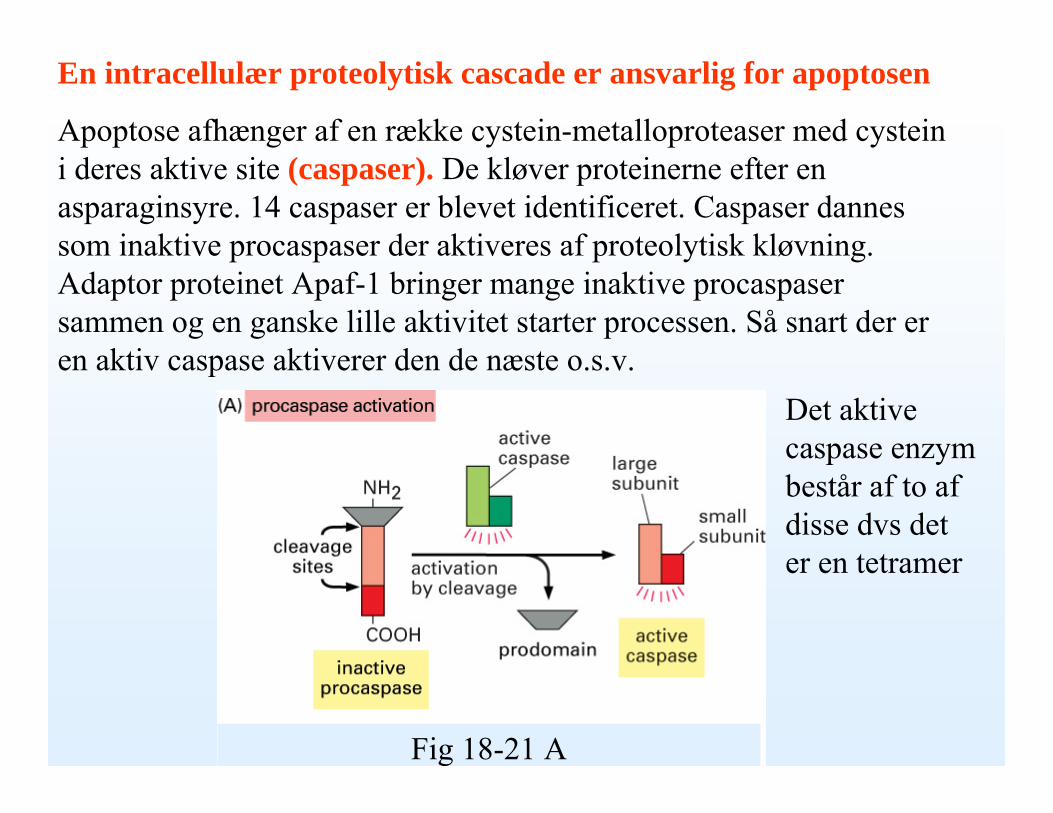
En intracellulær proteolytisk cascade er ansvarlig for apoptosen
Apoptose afhænger af en række cystein-metalloproteaser med cysteini deres aktive site (caspaser). De kløver proteinerne efter en asparaginsyre. 14 caspaser er blevet identificeret. Caspaser dannes som inaktive procaspaser der aktiveres af proteolytisk kløvning. Adaptor proteinet Apaf-1 bringer mange inaktive procaspasersammen og en ganske lille aktivitet starter processen. Så snart der er en aktiv caspase aktiverer den de næste o.s.v.
Det aktive caspase enzym består af to af disse dvs det er en tetramer
Fig 18-21 A
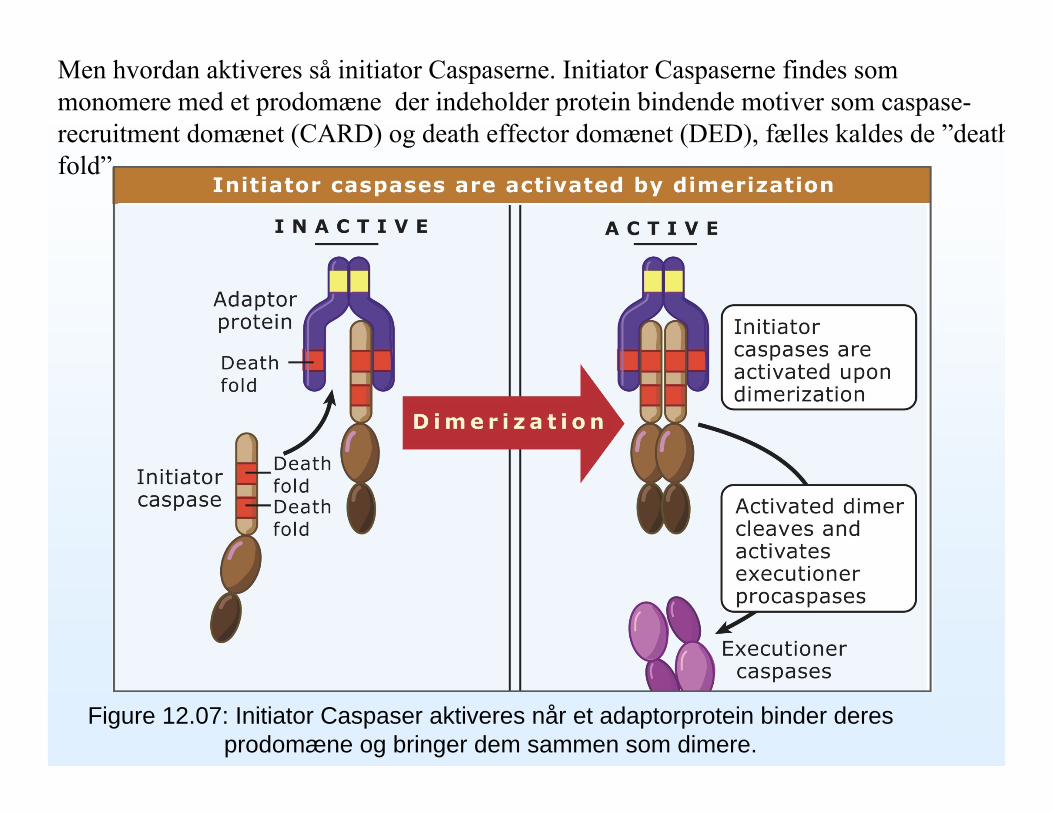
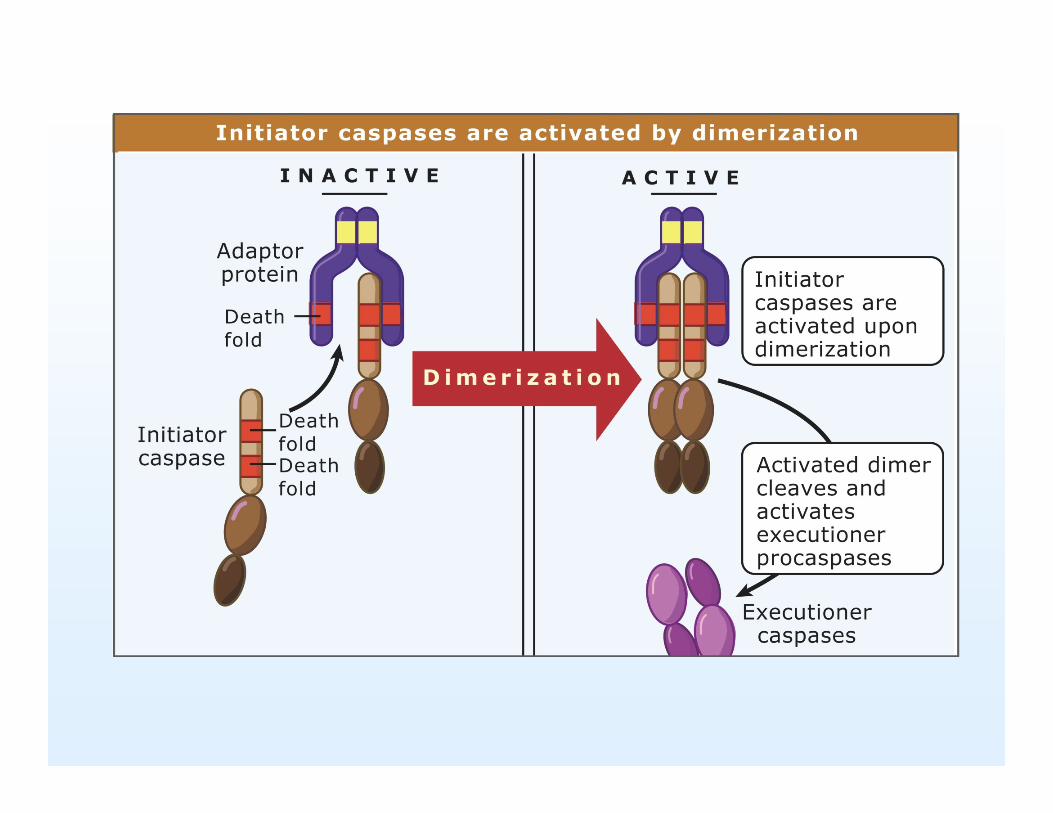
Figure 12.07: Initiator Caspaser aktiveres når et adaptorprotein binder deresprodomæne og bringer dem sammen som dimere.
Men hvordan aktiveres så initiator Caspaserne. Initiator Caspaserne findes som monomere med et prodomæne der indeholder protein bindende motiver som caspase-recruitment domænet (CARD) og death effector domænet (DED), fælles kaldes de ”deathfold”
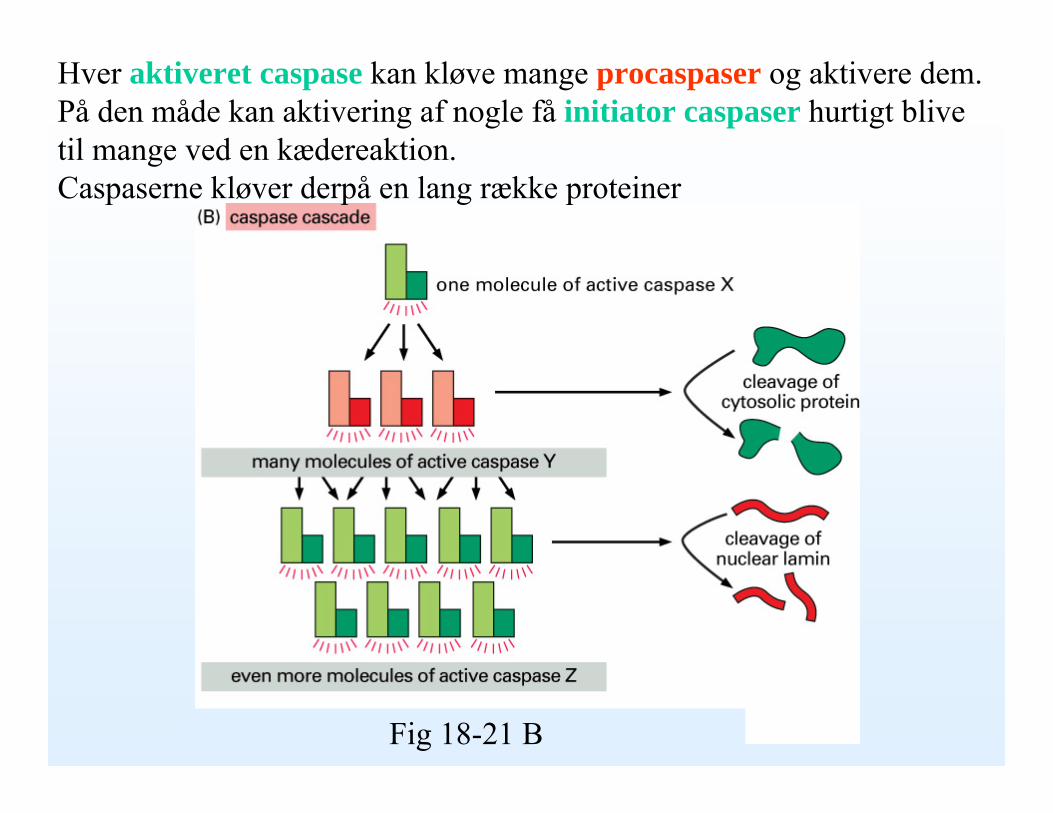
Hver aktiveret caspase kan kløve mange procaspaser og aktivere dem. På den måde kan aktivering af nogle få initiator caspaser hurtigt blive til mange ved en kædereaktion. Caspaserne kløver derpå en lang række proteiner
Fig 18-21 B
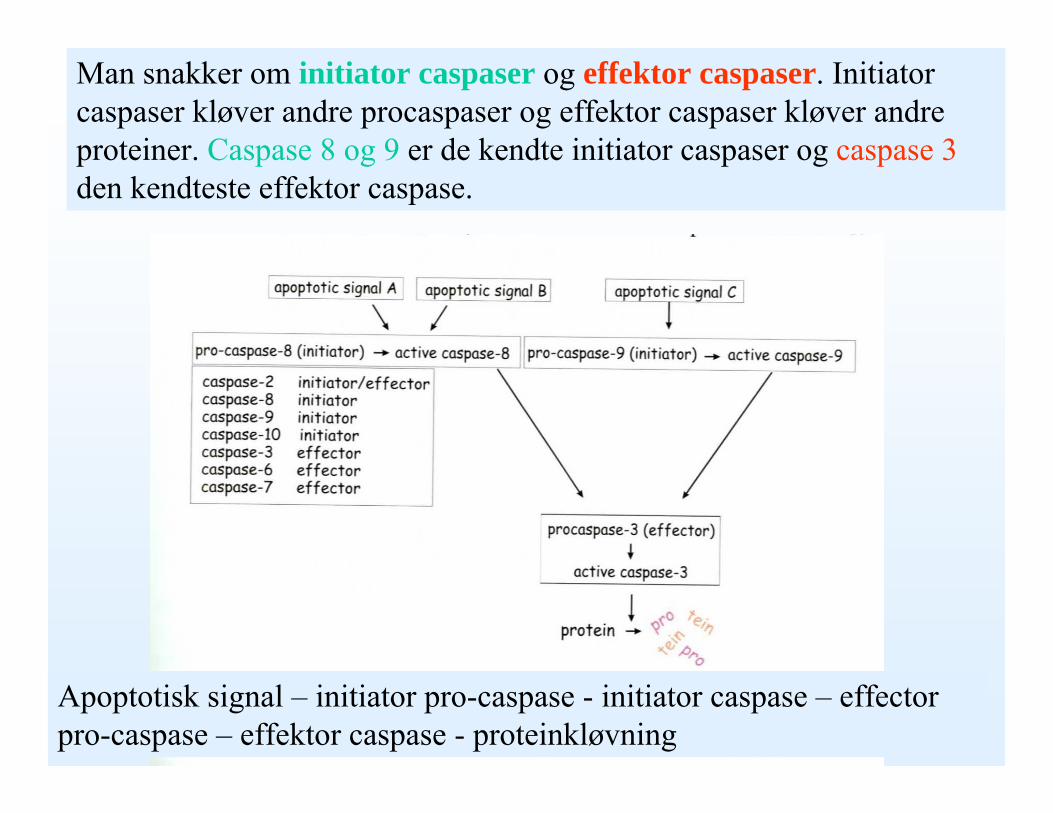
Man snakker om initiator caspaser og effektor caspaser. Initiator caspaser kløver andre procaspaser og effektor caspaser kløver andre proteiner. Caspase 8 og 9 er de kendte initiator caspaser og caspase 3den kendteste effektor caspase.
Apoptotisk signal – initiator pro-caspase - initiator caspase – effector pro-caspase – effektor caspase - proteinkløvning

Proteiner af Bcl-2 familien
• Proteiner af Bcl-2 familien er centrale i apoptosen. Der er proapoptotiske og antiapoptotiskemedlemmer af familien.
• Bcl-2 er antiapoptotisk• Bad inaktiverer Bcl-2 og er proapoptotisk• Bax og Bak som laver porer i mitochondrierne er
proapoptotiske• Bid som aktiverer Bax og Bak er proapoptotisk
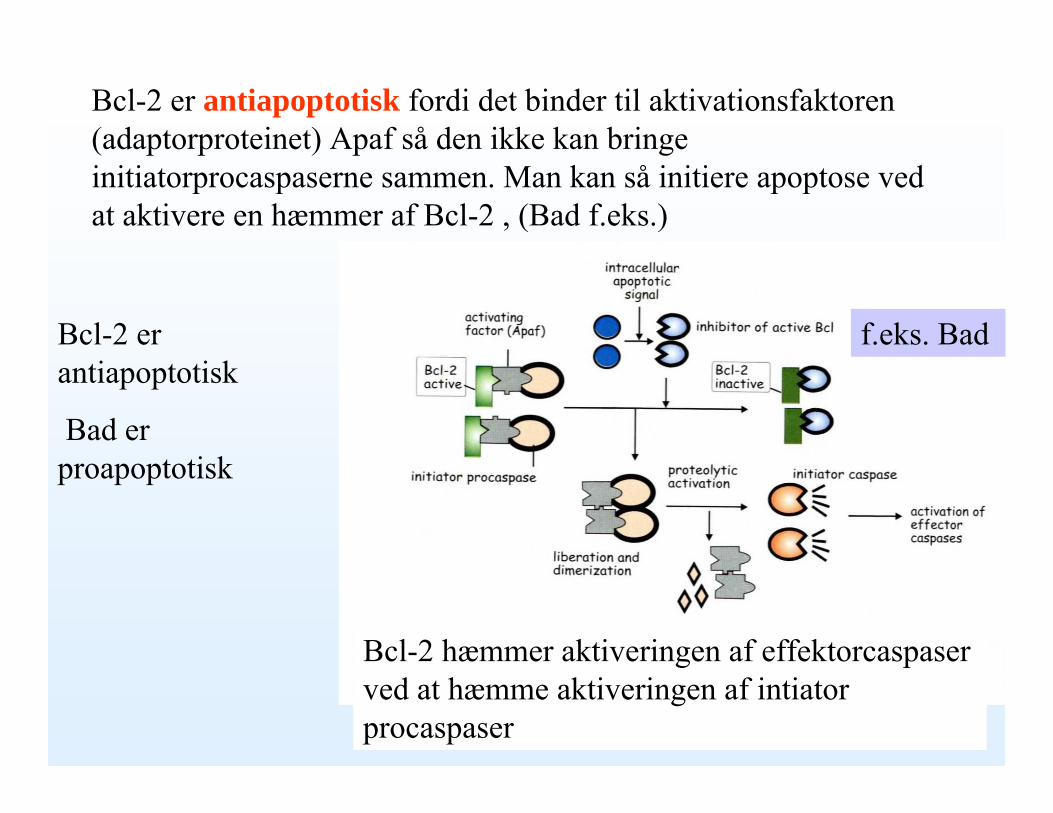
Bcl-2 er antiapoptotisk fordi det binder til aktivationsfaktoren(adaptorproteinet) Apaf så den ikke kan bringe initiatorprocaspaserne sammen. Man kan så initiere apoptose ved at aktivere en hæmmer af Bcl-2 , (Bad f.eks.)
f.eks. BadBcl-2 er antiapoptotisk
Bad er proapoptotisk
Bcl-2 hæmmer aktiveringen af effektorcaspaserved at hæmme aktiveringen af intiatorprocaspaser
18_22_Bcl_2_family.jpg
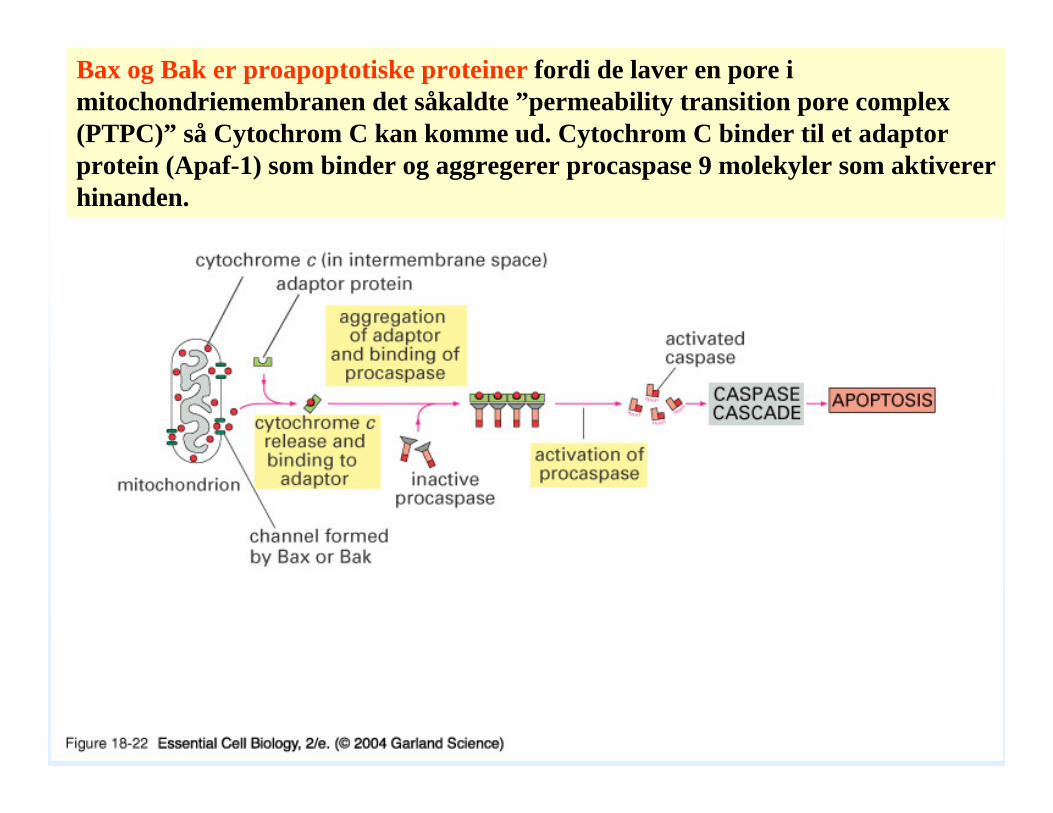
Bax og Bak er proapoptotiske proteiner fordi de laver en pore i mitochondriemembranen det såkaldte ”permeability transition pore complex(PTPC)” så Cytochrom C kan komme ud. Cytochrom C binder til et adaptorprotein (Apaf-1) som binder og aggregerer procaspase 9 molekyler som aktiverer hinanden.
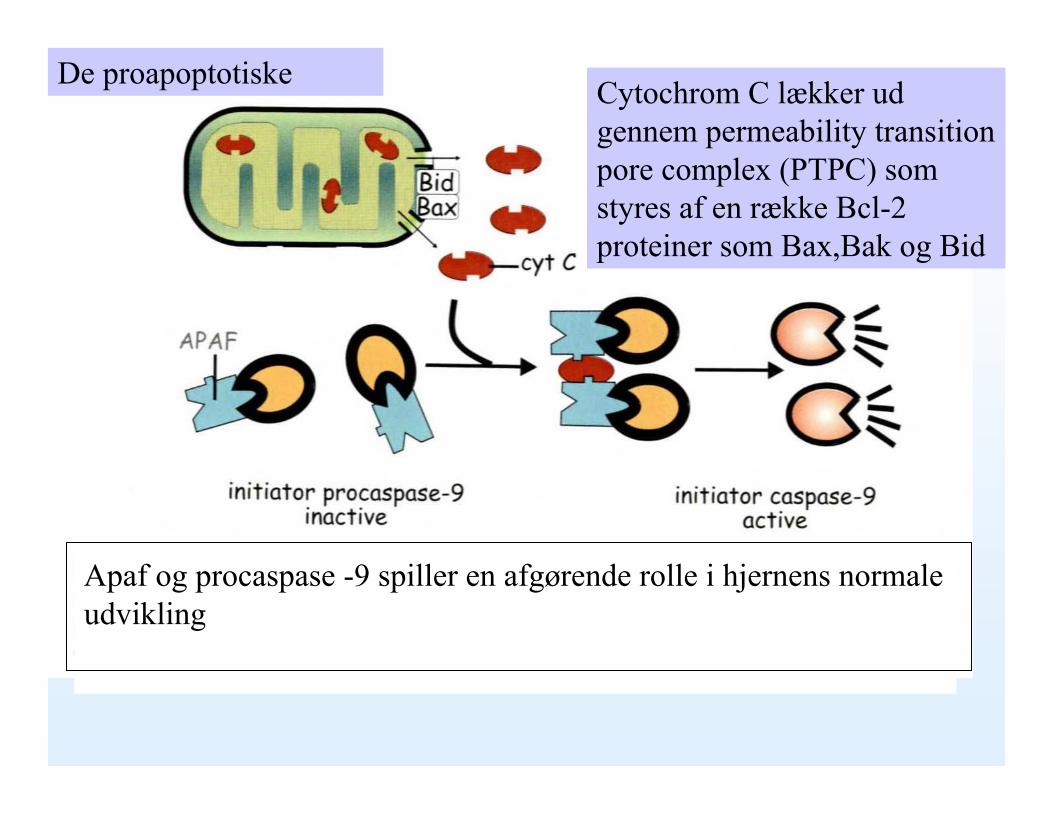
Cytochrom C lækker ud gennem permeability transition pore complex (PTPC) som styres af en række Bcl-2 proteiner som Bax,Bak og Bid
Apaf og procaspase -9 spiller en afgørende rolle i hjernens normale udvikling
De proapoptotiske
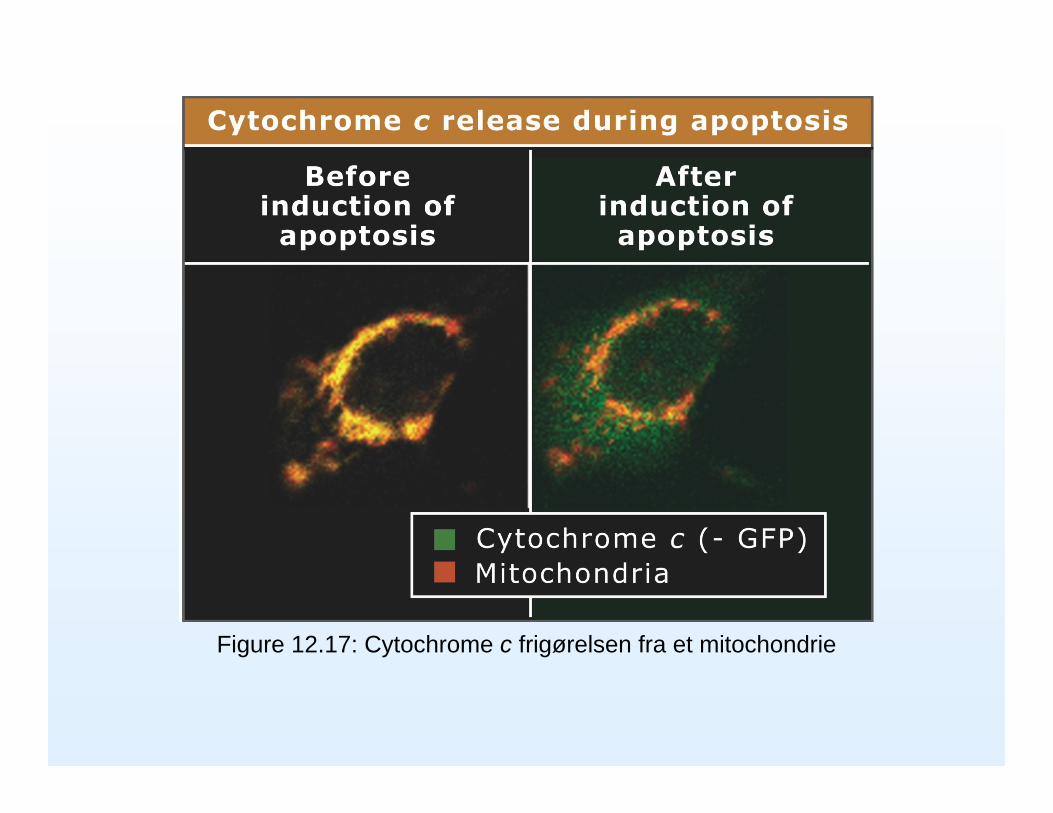
Figure 12.17: Cytochrome c frigørelsen fra et mitochondrie

Ekstracellulær kontrol med celledeling, cellevækst og apoptose
s. 628• Et befrugtet museæg og et befrugtet menneskeæg
har samme størrelse alligevel ender menneskeægget med at blive til et individ af meget større størrelse, hvad betinger det?
• Hvad bestemmer at elefanten får en lang snabel?• Hvorfor bliver leveren ikke 10 gange så stor?
• Det hele bestemmes af forholdet mellem cellevækst og celledeling på den ene side og programmeret celledød på den anden side.

De ekstracellulære signaler der stimulerer celledeling, cellevækst og celleoverlevelse• Mitogener der stimulerer celledeling
(celleproliferation)• Vækstfaktorer der stimulerer cellevækst
(forøgelse af cellemasse)• Overlevelsesfaktorer der undertrykker apoptose
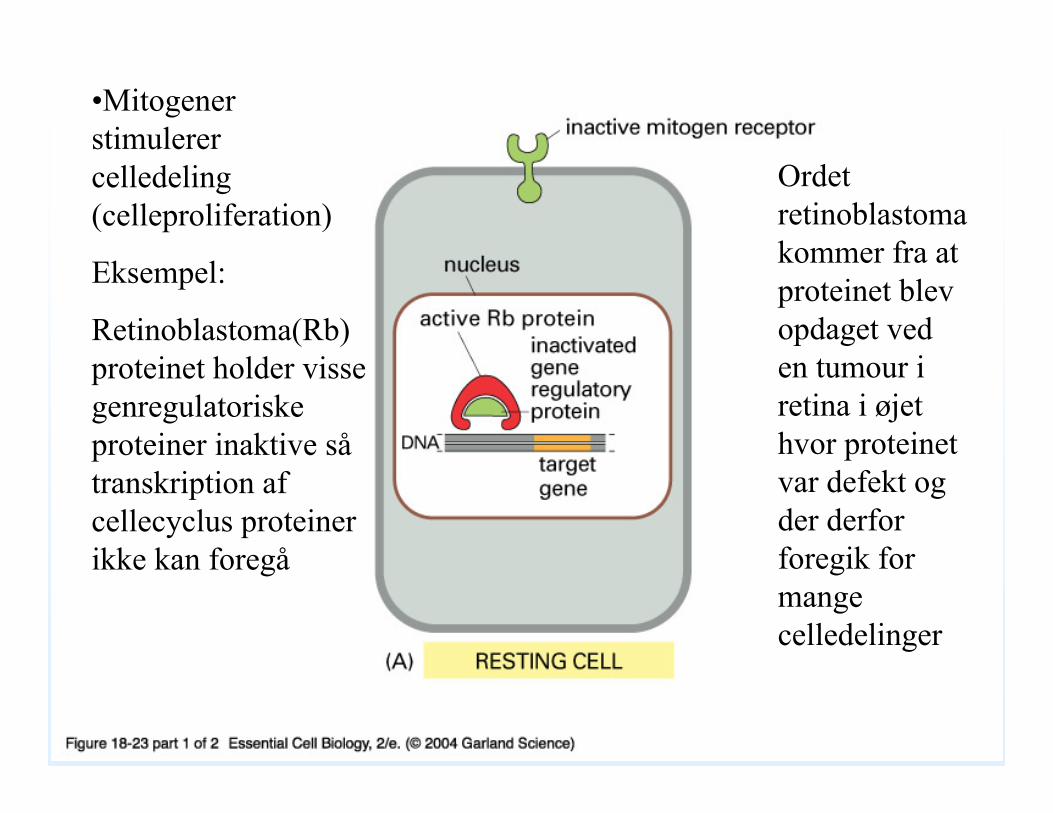
18_23_01_mitogens.jpg•Mitogenerstimulerer celledeling (celleproliferation)
Eksempel:
Retinoblastoma(Rb) proteinet holder visse genregulatoriskeproteiner inaktive såtranskription af cellecyclus proteiner ikke kan foregå
Ordet retinoblastomakommer fra at proteinet blev opdaget ved en tumour i retina i øjet hvor proteinet var defekt og der derfor foregik for mange celledelinger
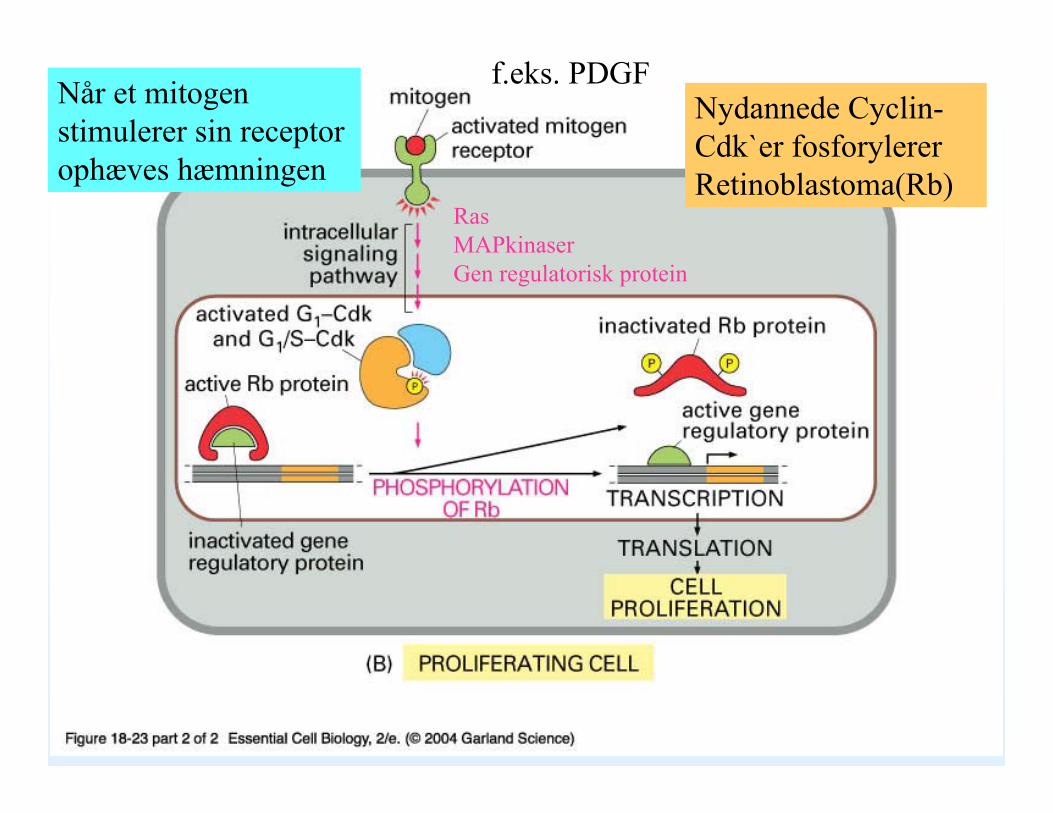
18_23_02_mitogens.jpgNår et mitogen stimulerer sin receptor ophæves hæmningen
Ras MAPkinaserGen regulatorisk protein
Nydannede Cyclin-Cdk`er fosforylererRetinoblastoma(Rb)
f.eks. PDGF
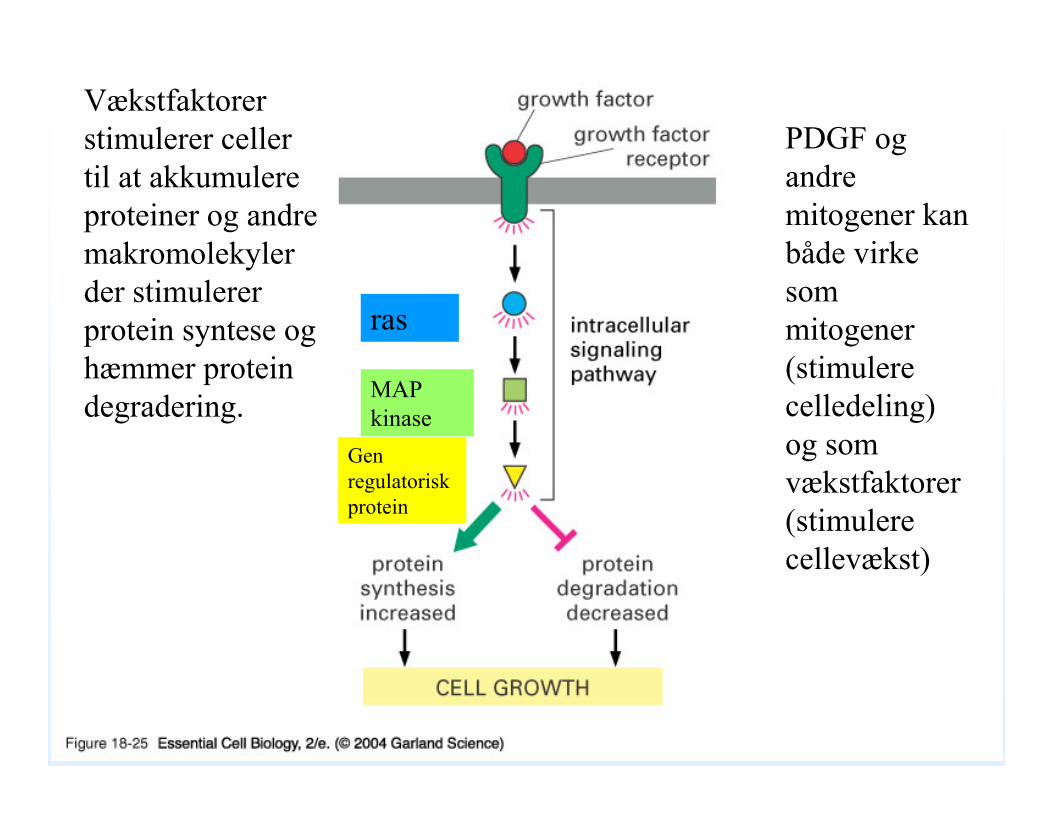
18_25_growth_factors.jpgVækstfaktorer stimulerer celler til at akkumulere proteiner og andre makromolekyler der stimulerer protein syntese og hæmmer protein degradering.
PDGF og andre mitogener kan både virke som mitogener(stimulere celledeling) og som vækstfaktorer (stimulere cellevækst)
ras
MAP kinase
Gen regulatoriskprotein

18_24_SEM_fibroblast.jpg
En fibroblast cellekultur i vækst . Der er tilsat serum som indeholder både vækstfaktorer og mutagener så cellerne både gror og deler sig
Celler i deling

Apoptose startes udefra på to måder
• Mangel på overlevelsesfaktor • Aktivering af dødsreceptorer

Naboceller konkurerer om overlevelsesfaktorer. Dvs at ved stor celletæthed induceres apoptose
Dette er vigtigt for at forhindre proliferation udover en ønsket tæthed, kaldes ”density-dependent inhibition of cell division”.
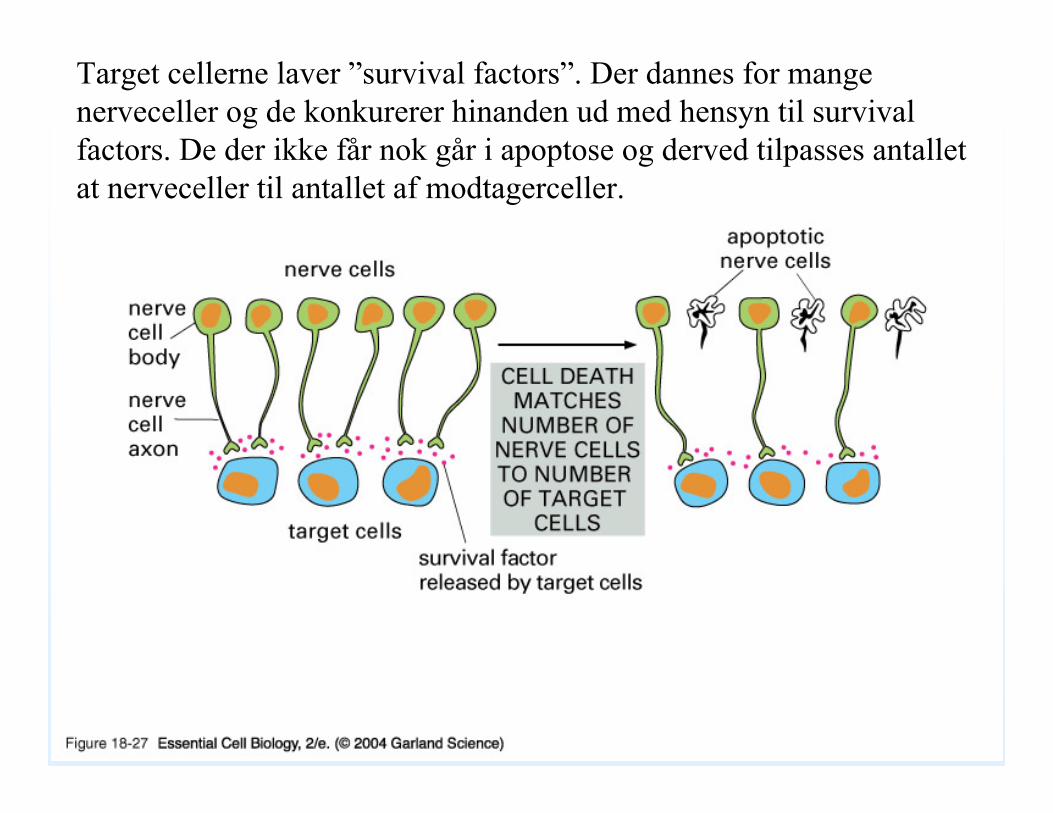
18_27_nerve_target.jpg
Target cellerne laver ”survival factors”. Der dannes for mange nerveceller og de konkurerer hinanden ud med hensyn til survivalfactors. De der ikke får nok går i apoptose og derved tilpasses antallet at nerveceller til antallet af modtagerceller.
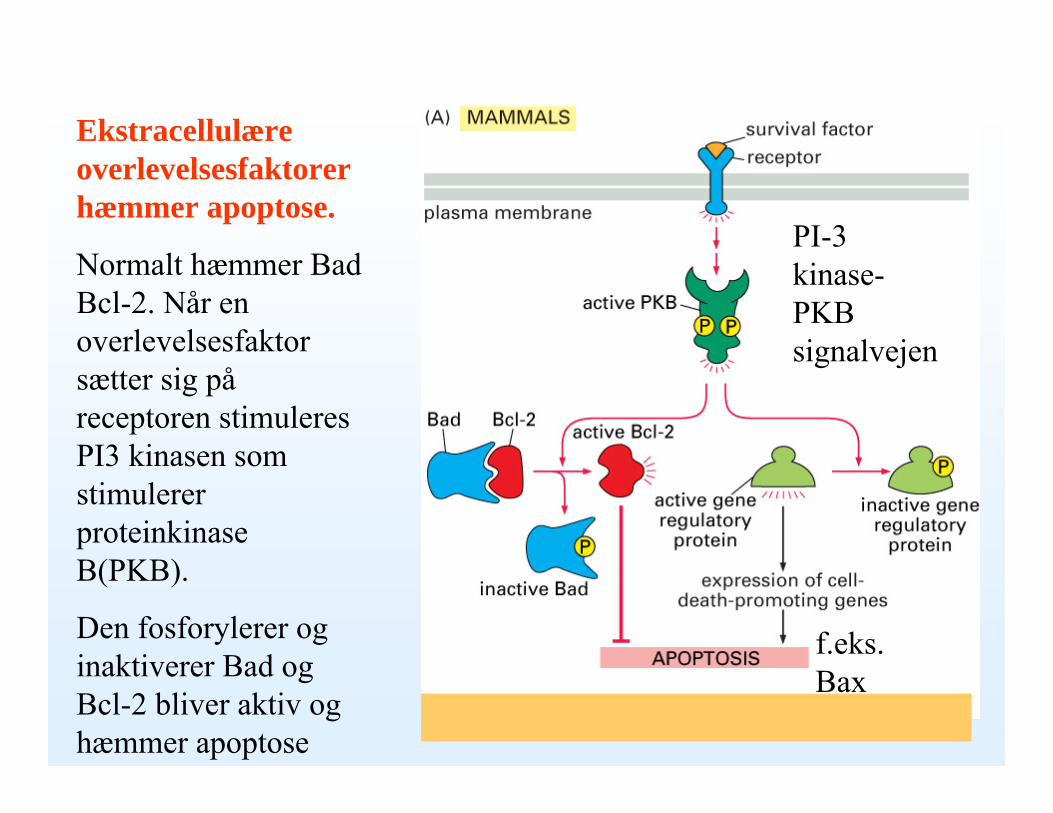
Ekstracellulære overlevelsesfaktorer hæmmer apoptose.
Normalt hæmmer Bad Bcl-2. Når en overlevelsesfaktor sætter sig påreceptoren stimuleres PI3 kinasen som stimulerer proteinkinaseB(PKB).
Den fosforylerer og inaktiverer Bad og Bcl-2 bliver aktiv og hæmmer apoptose
PI-3 kinase-PKB signalvejen
f.eks. Bax
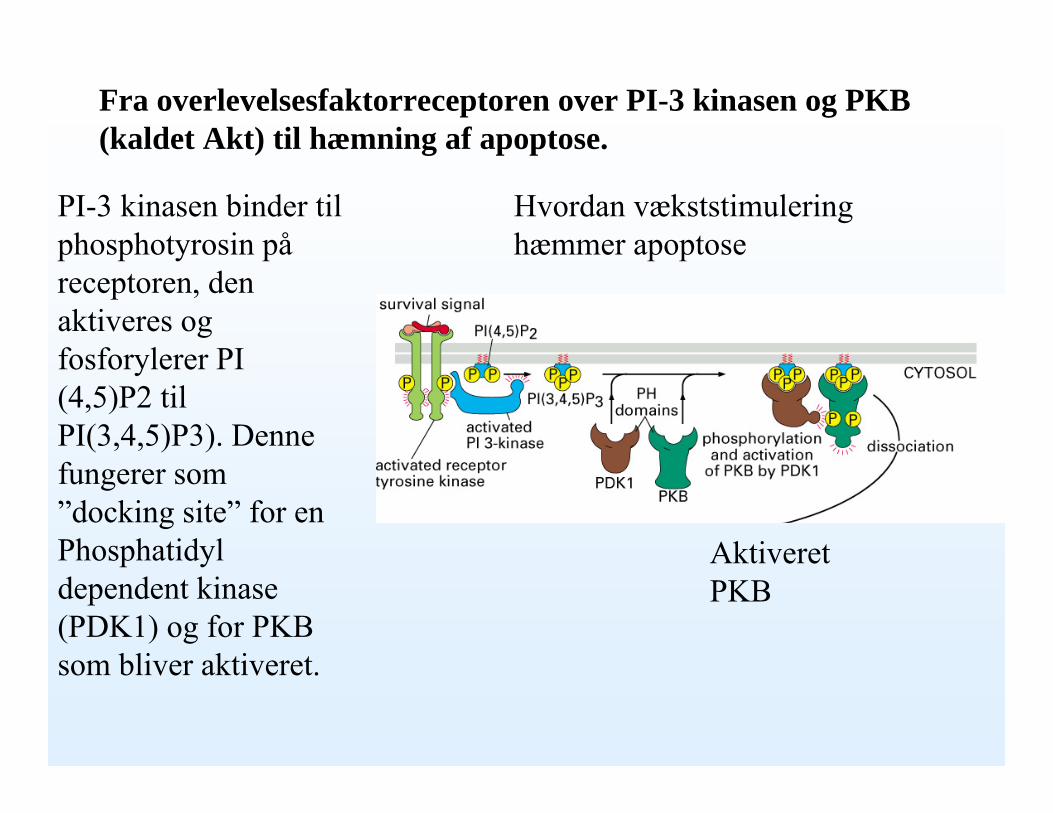
Fra overlevelsesfaktorreceptoren over PI-3 kinasen og PKB (kaldet Akt) til hæmning af apoptose.
PI-3 kinasen binder til phosphotyrosin påreceptoren, den aktiveres og fosforylerer PI (4,5)P2 til PI(3,4,5)P3). Denne fungerer som ”docking site” for en Phosphatidyldependent kinase (PDK1) og for PKB som bliver aktiveret.
Hvordan vækststimulering hæmmer apoptose
Aktiveret PKB
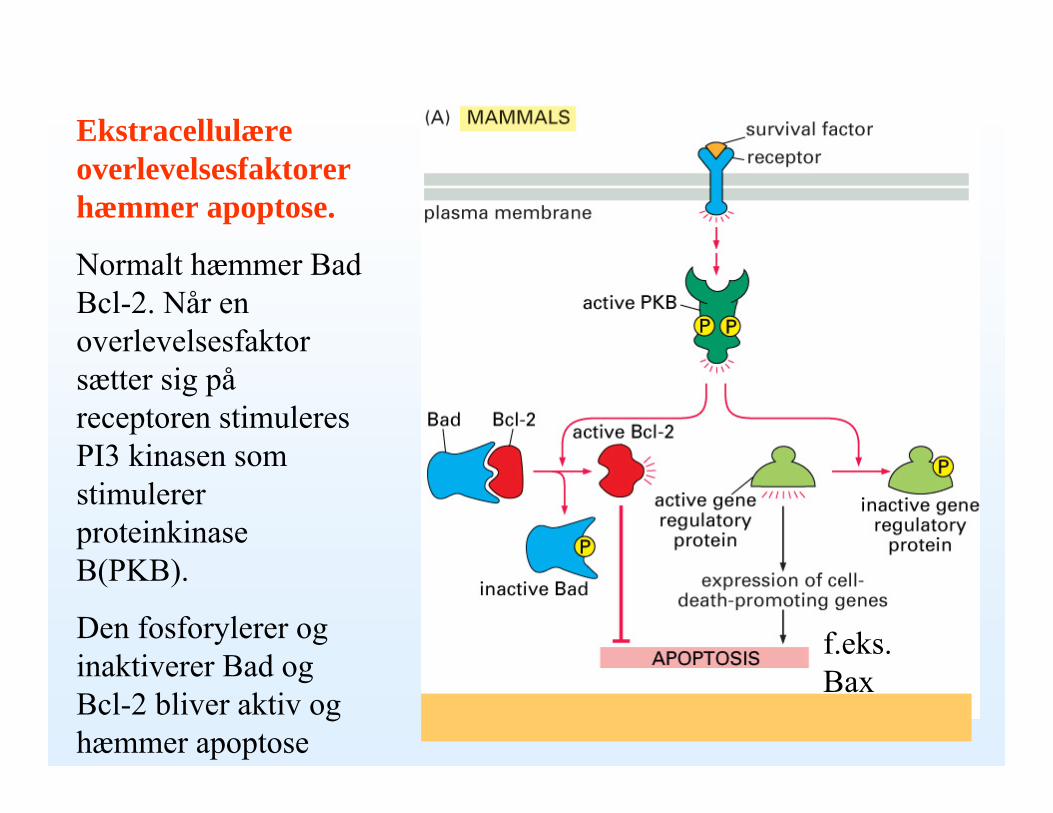
Ekstracellulære overlevelsesfaktorer hæmmer apoptose.
Normalt hæmmer Bad Bcl-2. Når en overlevelsesfaktor sætter sig påreceptoren stimuleres PI3 kinasen som stimulerer proteinkinaseB(PKB).
Den fosforylerer og inaktiverer Bad og Bcl-2 bliver aktiv og hæmmer apoptose
f.eks. Bax
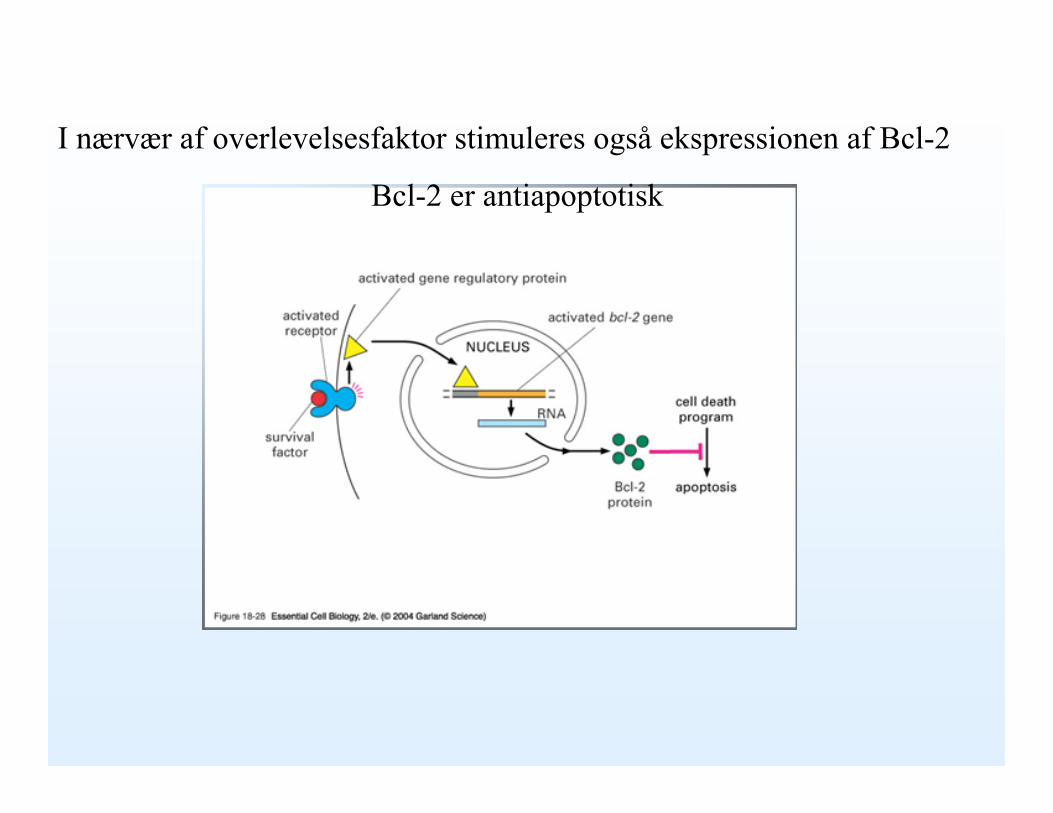
I nærvær af overlevelsesfaktor stimuleres også ekspressionen af Bcl-2
Bcl-2 er antiapoptotisk
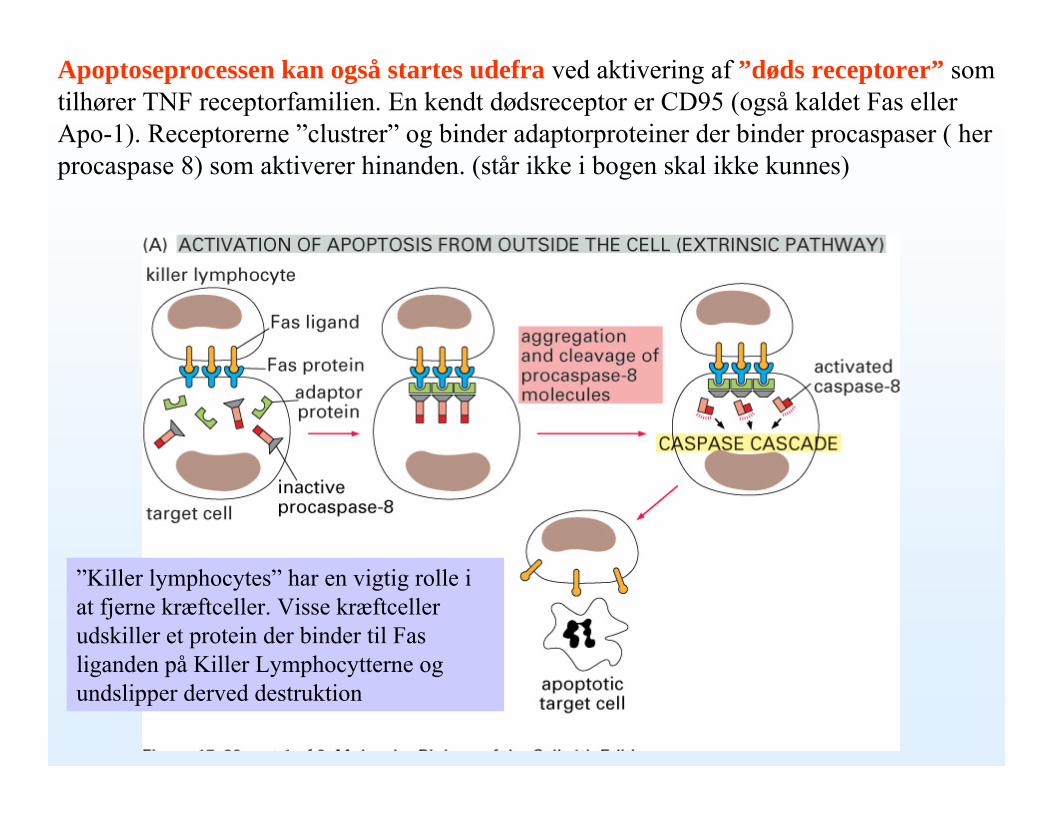
Apoptoseprocessen kan også startes udefra ved aktivering af ”døds receptorer” som tilhører TNF receptorfamilien. En kendt dødsreceptor er CD95 (også kaldet Fas eller Apo-1). Receptorerne ”clustrer” og binder adaptorproteiner der binder procaspaser ( her procaspase 8) som aktiverer hinanden. (står ikke i bogen skal ikke kunnes)
”Killer lymphocytes” har en vigtig rolle i at fjerne kræftceller. Visse kræftceller udskiller et protein der binder til Fas liganden på Killer Lymphocytterne og undslipper derved destruktion

18_29_myostatin_gene.jpg
Der er andre væksthæmmere end FAS, det er agonister som hæmmer vækst og proliferation men i modsætning til FAS ikke laver apoptose. I muskler findes f.eks. Myostatin et signalprotein der stopper vækst uden at lave apoptose. Hvis det muteres fås meget store muskler som her i en ”Belgian Blue ” ko.

Celleskrumpning er et af de karakteristiske tegn på apoptose men
er celleskrumpningen et signal til apoptose??
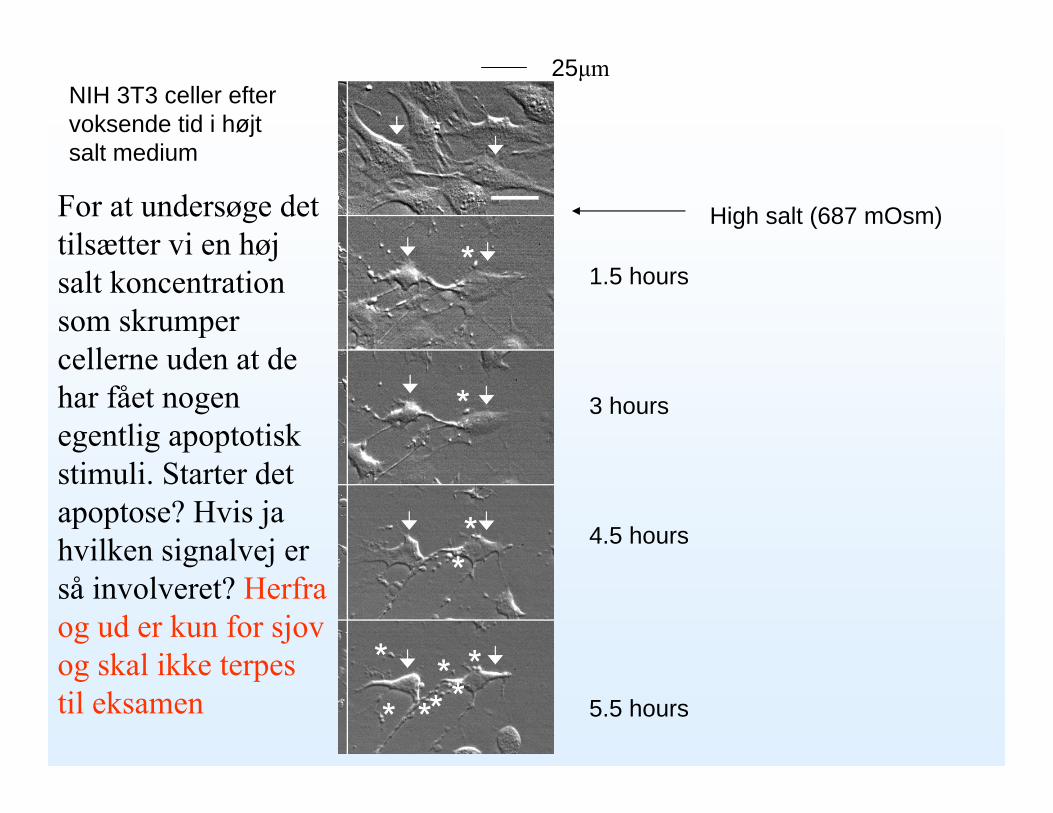
NIH 3T3 celler efter voksende tid i højt salt medium
High salt (687 mOsm)
1.5 hours
3 hours
4.5 hours
5.5 hours
25μm
For at undersøge det tilsætter vi en høj salt koncentration som skrumper cellerne uden at de har fået nogen egentlig apoptotiskstimuli. Starter det apoptose? Hvis ja hvilken signalvej er så involveret? Herfra og ud er kun for sjov og skal ikke terpes til eksamen
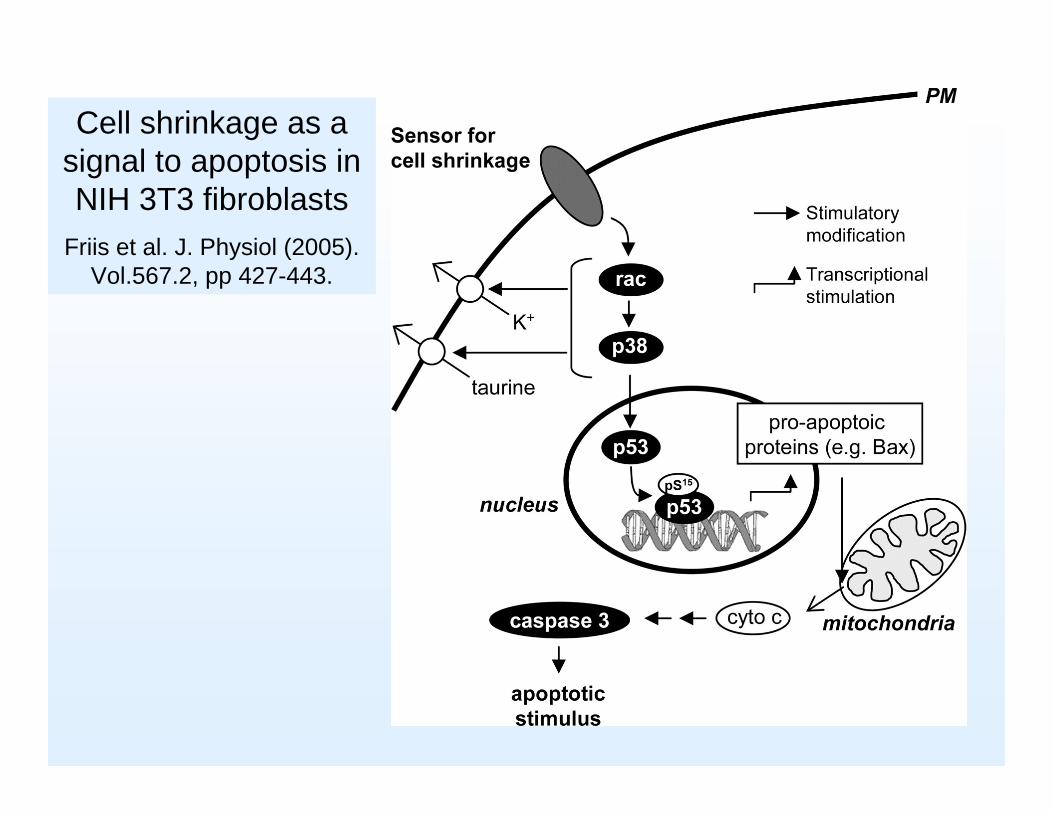
Cell shrinkage as a signal to apoptosis in NIH 3T3 fibroblasts
Friis et al. J. Physiol (2005). Vol.567.2, pp 427-443.
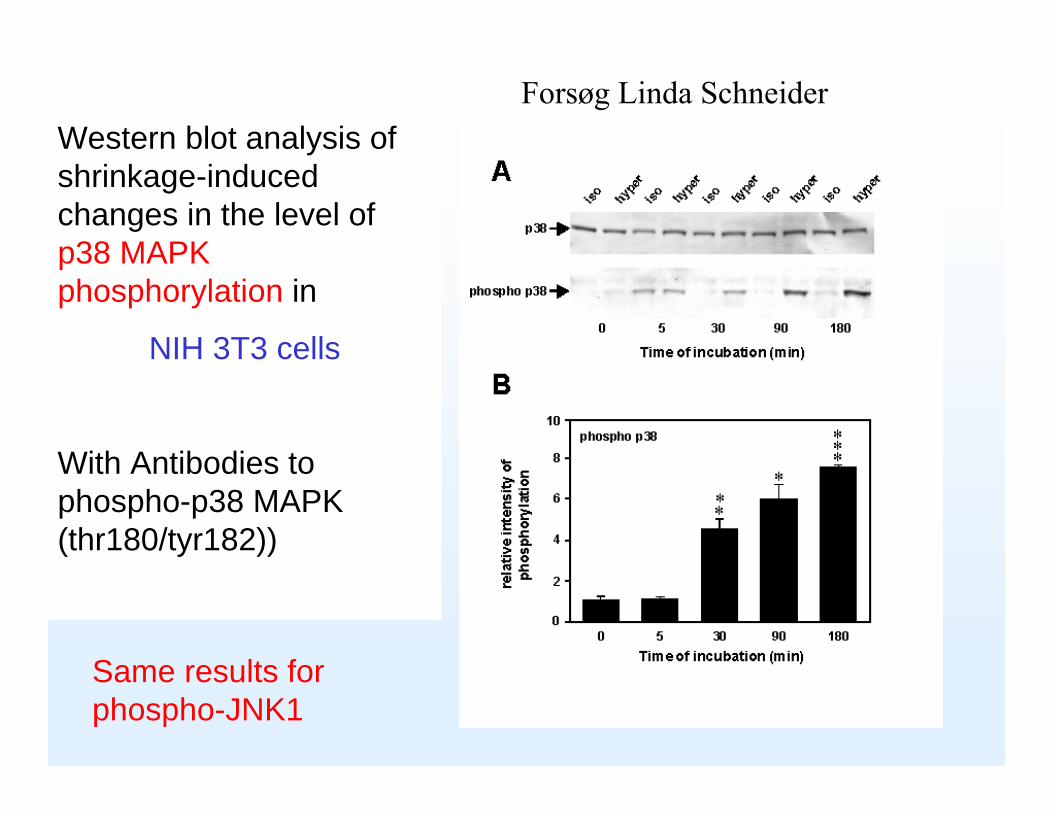
Western blot analysis of shrinkage-induced changes in the level of p38 MAPK phosphorylation in
NIH 3T3 cells
With Antibodies to phospho-p38 MAPK (thr180/tyr182))
Same results for phospho-JNK1
Forsøg Linda Schneider
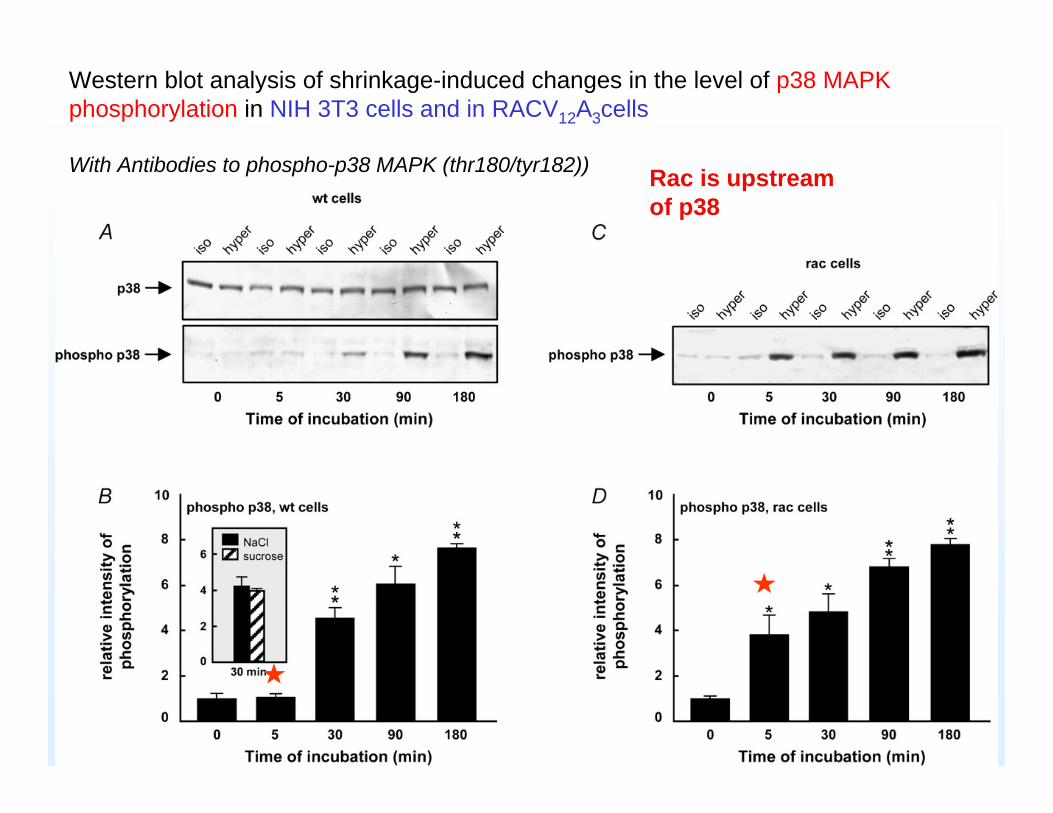
Western blot analysis of shrinkage-induced changes in the level of p38 MAPK phosphorylation in NIH 3T3 cells and in RACV12A3cells
With Antibodies to phospho-p38 MAPK (thr180/tyr182)) Rac is upstreamof p38

P53 er en essentiel nødbremse i cellecyklus når DNA skader opstår, hvilket bevises af at p53 genet er muteret i op til halvdelen af alle kræfttilfælde
P53 ”genomets vogter”Den cellulære concentration af p53 stiger når der forekommer DNA skader, kontrollen er post-transkriptionel idet translationen af eksisterende mRNA øges. Samtidig stabiliseres p53 også og holder længere. Ilt mangel kan også inducere p53 ligesom celleskrumpning kan
Når det forhøjede p53 niveau har stoppet cellens cyklus får cellen tid til at rette skaderne , niveauet af p53 falder igen og cyklus fortsætter. Er skaderne for store startes apoptose (programmeret celledød)
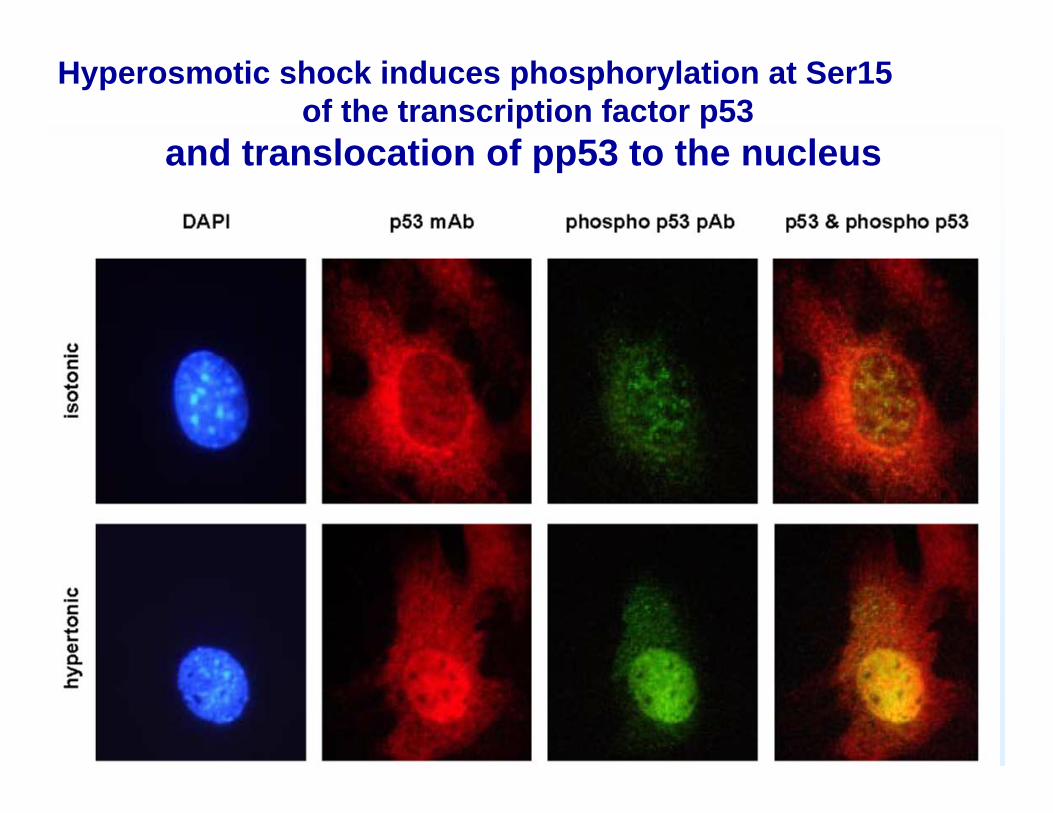
Hyperosmotic shock induces phosphorylation at Ser15of the transcription factor p53
and translocation of pp53 to the nucleus
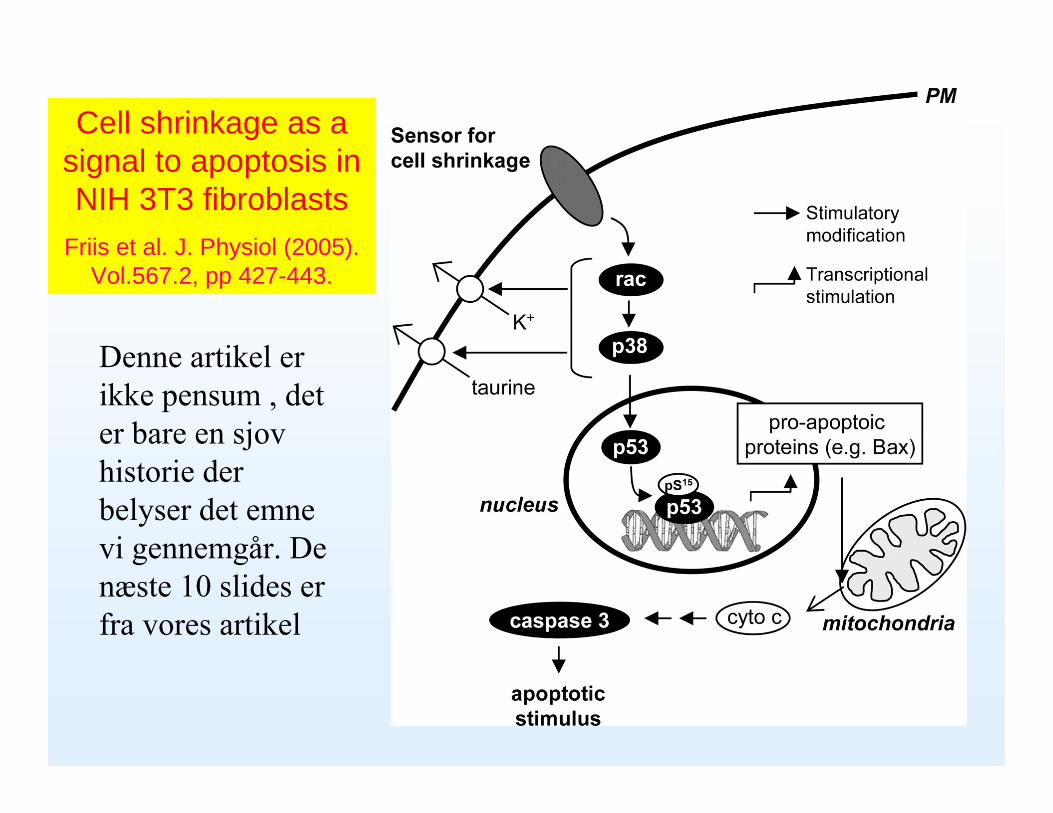
Cell shrinkage as a signal to apoptosis in NIH 3T3 fibroblasts
Friis et al. J. Physiol (2005). Vol.567.2, pp 427-443.
Denne artikel er ikke pensum , det er bare en sjov historie der belyser det emne vi gennemgår. De næste 10 slides er fra vores artikel
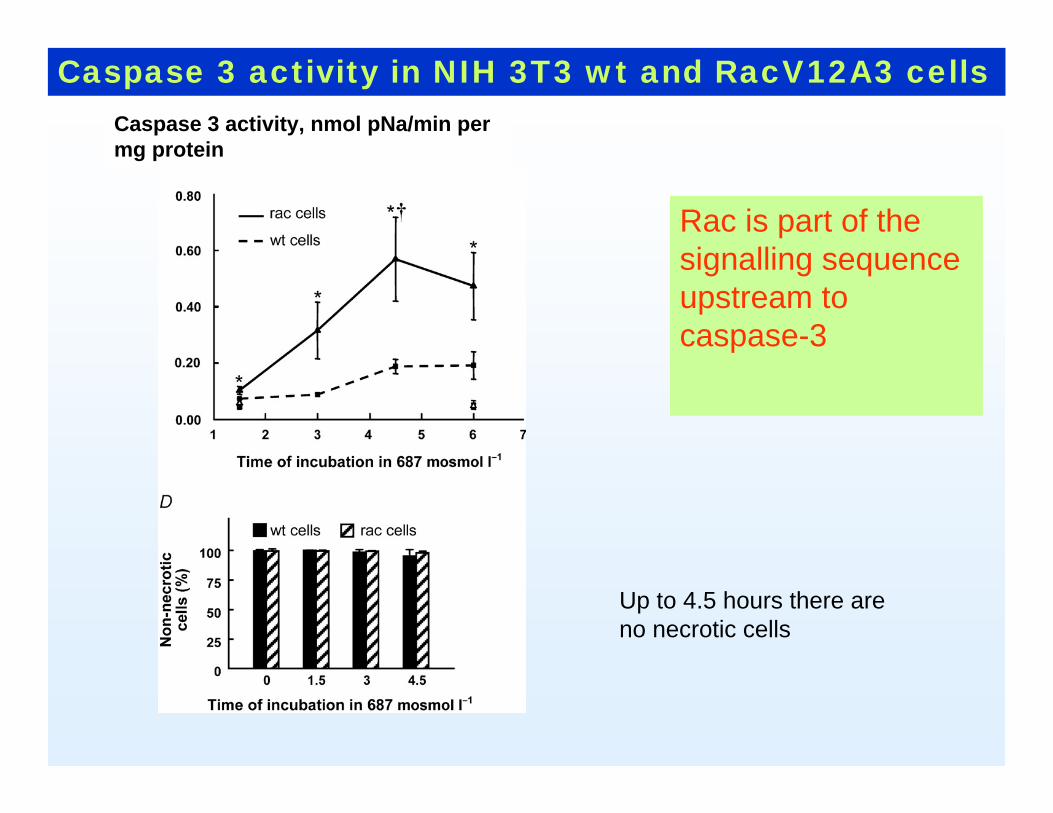
Caspase 3 activity in NIH 3T3 wt and RacV12A3 cells
Rac is part of the signalling sequence upstream to caspase-3
Caspase 3 activity, nmol pNa/min per mg protein
Up to 4.5 hours there areno necrotic cells
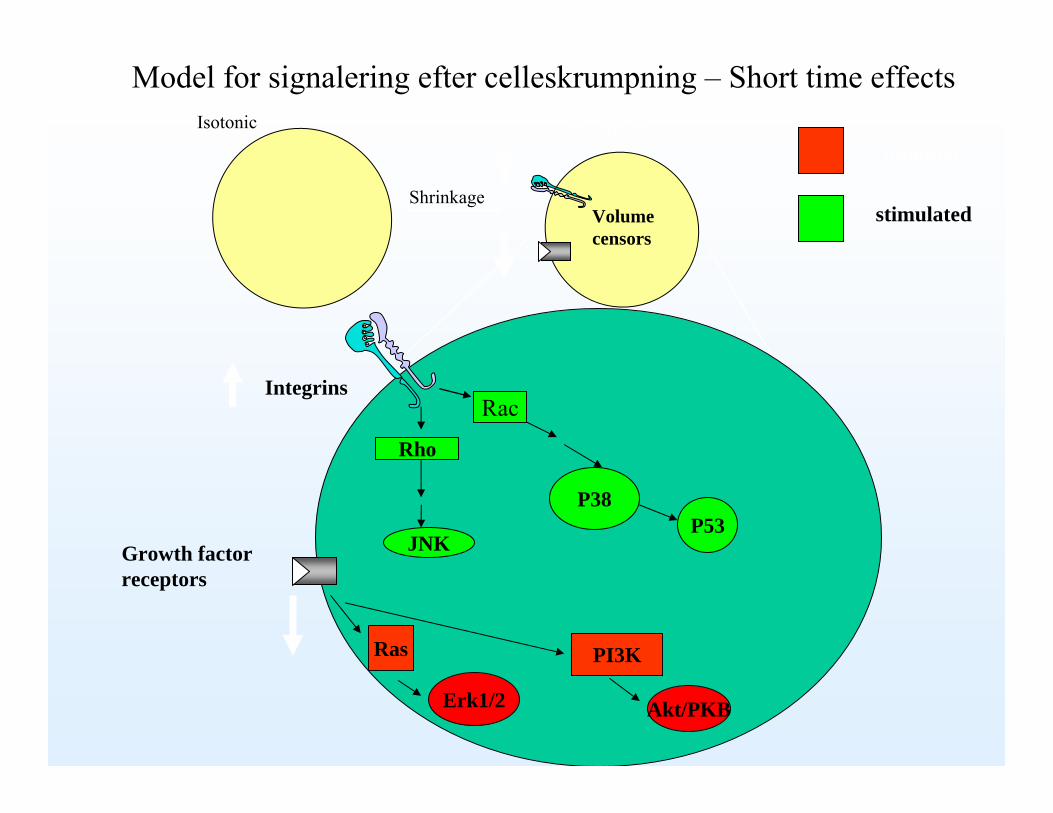
Isotonic Hypertonic
Shrinkage
P38
Rac
Rho
Integrins
PI3KRas
Growth factorreceptors
Model for signalering efter celleskrumpning – Short time effects
Volumecensors
JNK
Erk1/2
inhibited
stimulated
Akt/PKB
P53
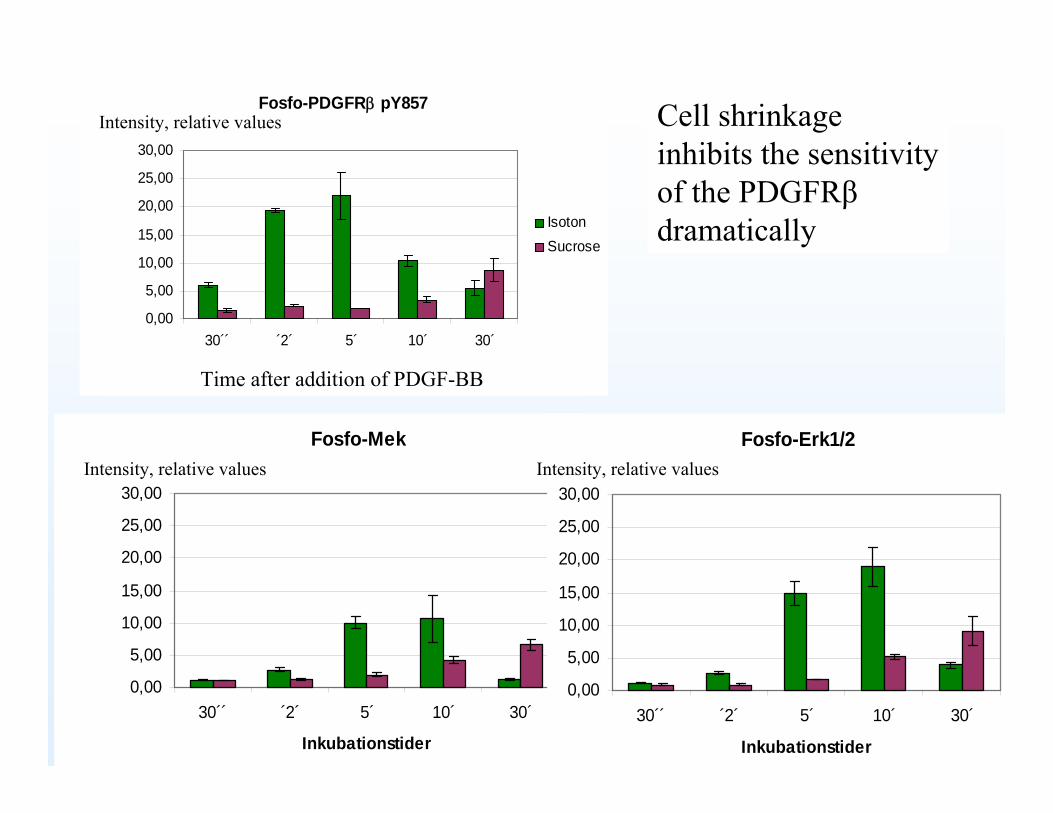
Fosfo-Mek
0,00
5,00
10,00
15,00
20,00
25,00
30,00
30´´ ´2´ 5´ 10´ 30´
Inkubationstider
Inte
nsite
tsfo
rhol
d +/
- PD
GF-
BB
Fosfo-PDGFRβ pY857
0,00
5,00
10,00
15,00
20,00
25,00
30,00
30´´ ´2´ 5´ 10´ 30´
Inkubationstider
Inte
nsite
tsfo
rhol
d +/
- PDG
F-BB
IsotonSucrose
Fosfo-Erk1/2
0,00
5,00
10,00
15,00
20,00
25,00
30,00
30´´ ´2´ 5´ 10´ 30´
Inkubationstider
Time after addition of PDGF-BB
Intensity, relative values
Intensity, relative values Intensity, relative values
Cell shrinkageinhibits the sensitivityof the PDGFRβdramatically

![PCD (2) 04.07.2016.ppt [Kompatibilit tsmodus]) · 3 Cytochrom c-Freisetzung Durch Caspase-Aktivierung Aktivierung Apoptose-spezifischer Endonucleasen Molekulare Merkmale der Apoptose](https://static.fdocument.pub/doc/165x107/5d49631588c9933f6b8b87b4/pcd-2-04072016ppt-kompatibilit-tsmodus-3-cytochrom-c-freisetzung-durch.jpg)















{kind=link}