Nuevos datos sobre algunas especies de Nemopteridae y ... · Nemopteridae eta Crocidae familietako...
33
Heteropterus Revista de Entomología Heteropterus Rev. Entomol. 2008 8(1): 1-33 ISSN: 1579-0681 Nuevos datos sobre algunas especies de Nemopteridae y Crocidae (Insecta: Neuroptera) V. J. MONSERRAT Departamento de Zoología y Antropología Física; Facultad de Biología; Universidad Complutense; E-28040 Madrid; Spain; E-mail: [email protected] Resumen Se aportan nuevos datos sobre la taxonomía, distribución, biología, morfología y/o estadios preimaginales de 16 especies de Nemopteridae y Crocidae. Se propone Pterocroce Withycombe, 1923 como nueva sinonimia de Necrophylus Roux, 1833 y Pterocroce capillaris (Klug, 1836) como nueva sinonimia de Necrophylus arenarius Roux, 1833. Se propone la designación de neotipo para Necrophylus arenarius Roux, 1833. Se describe el huevo de Lertha extensa (Olivier, 1811), L. sheppardi (Kirby, 1904) y Dielocroce ephemera (Gerstaecker, 1894). Se describen los estadios lar- varios de Lertha extensa y L. sheppardi y se aportan nuevos datos biológicos y morfológicos sobre las larvas de Croce schmidti (Navás, 1927), Necrophylus arenarius Roux, 1833, Laurhervasia setacea (Klug, 1836) y Dielocroce hebraea Hölzel, 1975. Se comentan algunos aspectos sobre el comportamiento, biología, preferencias alimentarias y desarrollo en las larvas de Lertha. Se aporta una clave de géneros de los estadios juveniles conocidos en Nemopteridae y Crocidae y, con base en los datos conocidos sobre sus estadios larvarios, se propone una nueva clasificación taxonómica y sistemática en Crocidae, incluyendo Necrophylini trib. nov. Por las diferentes características de sus larvas, se reconsidera válido el género Thysanocroce Withycombe, 1923 y no como sinónimo de Laurhervasia Navás, 1914. Se realiza un análisis biogeográfico que puede explicar la posible evolución y la actual distribución geográfica de Crocidae. Palabras clave: Faunística, taxonomía, sistemática, Neuroptera, Nemopteridae, Crocidae, huevo, larva, desa- rrollo, comportamiento. Laburpena Datu berriak Nemopteridae eta Crocidae-ren espezie batzuei buruz (Insecta: Neuroptera) Nemopteridae eta Crocidae familietako 16 espezieren taxonomia, banaketa, biologia, morfologia edota fase preimaginalei buruzko datu berriak ematen dira. Pterocroce Withycombe, 1923 generoa Necrophylus Roux, 1833 generoaren sinonimotzat proposatzen da, eta Pterocroce capillaris (Klug, 1836) espeziea Necrophylus arenarius Roux, 1833 espeziearena. Neotipoa proposatzen da Necrophylus arenarius Roux, 1833 espeziearentzat. Lertha extensa (Oli- vier, 1811), L. sheppardi (Kirby, 1904) eta Dielocroce ephemera (Gerstaecker, 1894) espezieen arrautzak deskribatzen dira. Lertha extensa eta L. sheppardiren larba-estadioak deskribatzen dira eta Croce schmidti (Navás, 1927), Necrophylus arenarius Roux, 1833, Laurhervasia setacea (Klug, 1836) eta Dielocroce hebraea Hölzel, 1975 espezieen larbei buruzko datu biologiko eta morfologiko berri batzuk ematen dira. Lertharen larben portaeraren, biologiaren, elika-prefe- rentzien eta garapenaren inguruko zenbait alderdi komentatzen dira. Nemopteridae eta Crocidae-ren generoetarako ezagunak diren gazte-estadioen klabe bat aurkezten da eta, larba-es- tadioei buruzko datu ezagunak aintzat hartuta, Crocidae-ren sailkapen taxonomiko eta sistematiko berri bat propo- satzen da, Necrophylini trib. nov. ere barnean delarik. Haien larben ezaugarri desberdinak direla eta, Thysanocroce Withycombe, 1923 ontzat berrartzen da eta ez Laurhervasiaren Navás, 1914 sinonimotzat. Crocidae-ren litekeen ebo- luzioa eta gaur egungo banaketa geografikoa azaltzeko balio dezakeelakoan, analisi biogeografikoa ere garatzen da. Gako-hitzak: Faunistika, taxonomia, sistematika, Neuroptera, Nemopteridae, Crocidae, arrautza, larba, gara- pena, portaera. Abstract New data on some species of Nemopteridae and Crocidae (Insecta: Neuroptera) New data on the taxonomy, distribution, biology, morphology and/or preimaginal stages of 16 spoon-winged
Transcript of Nuevos datos sobre algunas especies de Nemopteridae y ... · Nemopteridae eta Crocidae familietako...

Heteropterus Revista de Entomología
Heteropterus Rev. Entomol.2008 8(1): 1-33
ISSN: 1579-0681
Nuevos datos sobre algunas especies de
Nemopteridae y Crocidae (Insecta: Neuroptera)
V. J. MONSERRAT
Departamento de Zoología y Antropología Física; Facultad de Biología; Universidad Complutense;
E-28040 Madrid; Spain; E-mail: [email protected]
Resumen
Se aportan nuevos datos sobre la taxonomía, distribución, biología, morfología y/o estadios preimaginales de16 especies de Nemopteridae y Crocidae. Se propone Pterocroce Withycombe, 1923 como nueva sinonimia deNecrophylus Roux, 1833 y Pterocroce capillaris (Klug, 1836) como nueva sinonimia de Necrophylus arenarius Roux, 1833.Se propone la designación de neotipo para Necrophylus arenarius Roux, 1833. Se describe el huevo de Lertha extensa
(Olivier, 1811), L. sheppardi (Kirby, 1904) y Dielocroce ephemera (Gerstaecker, 1894). Se describen los estadios lar-varios de Lertha extensa y L. sheppardi y se aportan nuevos datos biológicos y morfológicos sobre las larvas de Croce
schmidti (Navás, 1927), Necrophylus arenarius Roux, 1833, Laurhervasia setacea (Klug, 1836) y Dielocroce hebraea Hölzel,1975. Se comentan algunos aspectos sobre el comportamiento, biología, preferencias alimentarias y desarrollo enlas larvas de Lertha.
Se aporta una clave de géneros de los estadios juveniles conocidos en Nemopteridae y Crocidae y, con base enlos datos conocidos sobre sus estadios larvarios, se propone una nueva clasificación taxonómica y sistemática enCrocidae, incluyendo Necrophylini trib. nov. Por las diferentes características de sus larvas, se reconsidera válidoel género Thysanocroce Withycombe, 1923 y no como sinónimo de Laurhervasia Navás, 1914. Se realiza un análisisbiogeográfico que puede explicar la posible evolución y la actual distribución geográfica de Crocidae.
Palabras clave: Faunística, taxonomía, sistemática, Neuroptera, Nemopteridae, Crocidae, huevo, larva, desa-rrollo, comportamiento.
Laburpena
Datu berriak Nemopteridae eta Crocidae-ren espezie batzuei buruz (Insecta: Neuroptera)
Nemopteridae eta Crocidae familietako 16 espezieren taxonomia, banaketa, biologia, morfologia edota fasepreimaginalei buruzko datu berriak ematen dira. Pterocroce Withycombe, 1923 generoa Necrophylus Roux, 1833generoaren sinonimotzat proposatzen da, eta Pterocroce capillaris (Klug, 1836) espeziea Necrophylus arenarius Roux,1833 espeziearena. Neotipoa proposatzen da Necrophylus arenarius Roux, 1833 espeziearentzat. Lertha extensa (Oli-vier, 1811), L. sheppardi (Kirby, 1904) eta Dielocroce ephemera (Gerstaecker, 1894) espezieen arrautzak deskribatzendira. Lertha extensa eta L. sheppardiren larba-estadioak deskribatzen dira eta Croce schmidti (Navás, 1927), Necrophylus
arenarius Roux, 1833, Laurhervasia setacea (Klug, 1836) eta Dielocroce hebraea Hölzel, 1975 espezieen larbei buruzkodatu biologiko eta morfologiko berri batzuk ematen dira. Lertharen larben portaeraren, biologiaren, elika-prefe-rentzien eta garapenaren inguruko zenbait alderdi komentatzen dira.
Nemopteridae eta Crocidae-ren generoetarako ezagunak diren gazte-estadioen klabe bat aurkezten da eta, larba-es-tadioei buruzko datu ezagunak aintzat hartuta, Crocidae-ren sailkapen taxonomiko eta sistematiko berri bat propo-satzen da, Necrophylini trib. nov. ere barnean delarik. Haien larben ezaugarri desberdinak direla eta, Thysanocroce
Withycombe, 1923 ontzat berrartzen da eta ez Laurhervasiaren Navás, 1914 sinonimotzat. Crocidae-ren litekeen ebo-luzioa eta gaur egungo banaketa geografikoa azaltzeko balio dezakeelakoan, analisi biogeografikoa ere garatzen da.
Gako-hitzak: Faunistika, taxonomia, sistematika, Neuroptera, Nemopteridae, Crocidae, arrautza, larba, gara-pena, portaera.
Abstract
New data on some species of Nemopteridae and Crocidae (Insecta: Neuroptera)
New data on the taxonomy, distribution, biology, morphology and/or preimaginal stages of 16 spoon-winged

Introducción
Bien sean consideradas a nivel de subfamilia (Nemop-terinae y Crocinae), bien a nivel de familia (Nemop-teridae y Crocidae), este interesante grupo de neu-rópteros incluye respectivamente 19 y 17 génerosvivientes distribuidos por las zonas áridas y subde-sérticas de las Regiones Paleártica occidental, Afro-tropical, de Oriente Medio, Neotropical, Australianay Sur de la Región Oriental y, en su conjunto, inclu-yen 151 especies, en ocasiones muy espectacularesy a veces de gran tamaño y de vistosos colores.Son habitantes de espacios abiertos en el caso deNemopteridae (101 especies) y frágiles, delicadas yhabitantes de oquedades, cuevas y habitaciones hu-manas en el de Crocidae (50 especies), con curiosasadaptaciones y estrategias de supervivencia a losáridos e inhóspitos medios donde habitualmenteviven (Hölzel, 1975a; Aspöck et al., 1980, 2001; Man-sell, 1992, 1996).
Estas familias habían sido revisadas por Navás (1910,1912) y no adquirieron un suficiente nivel de conoci-miento hasta la obra de Tjeder (1967) pero, a diferen-cia de lo que ocurre recientemente en otras familiasde neurópteros, especialmente en aquellas de interésaplicado, se ha venido publicando un proporcional-mente escaso número de datos sobre estas familias;esto es probablemente debido, en Nemopteridae, ala mayor dificultad en su recolección por el caráctermonovoltino y por la breve temporalidad en la emer-gencia de sus imagos, generalmente restringidos a
hábitats específicos y, en Crocidae, a las especialescondiciones ambientales, mayoritariamente troglo-bias, donde se desarrollan, siendo realmente escasala información de que disponemos sobre la distri-bución real de muchas especies. Aunque los estadioslarvarios en Crocidae son bastante bien conocidosen la mayoría de sus géneros, en Nemopteridae soncasi desconocidos (ver referencias en cada caso).
En la presente contribución anotamos nuevos datossobre la distribución, biología, morfología, y/o esta-dios preimaginales de 16 especies pertenecientes aestas dos familias.
Material y método
La mayoría de los ejemplares que se citan pertenecena la colección del autor; sin embargo, otra parte delmaterial estudiado pertenece a diferentes coleccio-nes. Listamos todas ellas a continuación, anotandolas siglas empleadas para indicar la pertenencia delmaterial que se cita:
(DBDU): Department of Biology, Dicle University(Diyarbakir, Turquía).
(JB): Colección J. Blasco (Zaragoza, España).
(MG): Museo Civico di Storia Naturale «G. Doria»(Génova, Italia).
MONSERRAT: Nuevos datos sobre Nemopteridae y Crocidae (Neuroptera)2
and thread-winged lace-wing species (Nemopteridae and Crocidae) are given. The genus Pterocroce Withycombe,1923 is proposed as a new synonym of Necrophylus Roux, 1833, and Pterocroce capillaris (Klug, 1836) as a newsynonym of Necrophylus arenarius Roux, 1833. A neotype designation for Necrophylus arenarius Roux, 1833 is pro-posed. The eggs of Lertha extensa (Olivier, 1811), L. sheppardi (Kirby, 1904) and Dielocroce ephemera (Gerstaecker,1894) are described. The larval stages of Lertha extensa and L. sheppardi are described and new data are provided onthe larval morphology and biology of Croce schmidti (Navás, 1927), Necrophylus arenarius Roux, 1833, Laurhervasia
setacea (Klug, 1836) and Dielocroce hebraea Hölzel, 1975. Some new data on the behaviour, biology, feeding prefe-rences and development of Lertha larvae are commented.
A key to the genera of the known mature larvae of Nemopteridae and Crocidae is included and the known larvalcharacters of the different genera are commented in order to contribute to the systematics of these families. Onthe basis of the larval characteristics a new taxonomic and systematic classification within Crocidae is provided,including Necrophylini trib. nov., and the genus Thysanocroce Withycombe, 1923 is reconsidered as a valid genusand not as a synonym of Laurhervasia Navás, 1914. A biogeographical analysis is also made on Crocidae in orderto explain its possible evolution and its current geographical distribution.
Key words: Faunistics, taxonomy, systematics, Neuroptera, Nemopteridae, Crocidae, egg, larva, development,behaviour.

(MZB): Museu de Zoologia (Barcelona, España).
(NHM): The Natural History Museum (Londres,Reino Unido).
(NMB): Naturhistorisches Museum (Basel, Suiza).
(TAMU): Department of Entomology, Texas A. & M.University (College Station, USA).
(UCM): Departamento de Zoología y AntropologíaFísica, Universidad Complutense (Madrid,España).
(VM): Colección V. J. Monserrat (Madrid, España).
Las especies que se mencionan siguen la ordenacióngeneral dada por Aspöck et al. (1980, 2001). De cadauna de las 16 especies citadas en esta contribuciónanotamos una breve sinopsis de su estado actual deconocimiento, de su distribución general previamenteconocida, así como de la información reciente mássignificativa que merezca citarse en relación con losdatos aportados.
Para la exposición del material estudiado, éste se citaordenado alfabética- y cronológicamente siguiendola siguiente secuencia: País, región-provincia-esta-do-localidad de captura, fecha de captura, númerode %%, && o larvas estudiadas, datos de biología,recolector e institución a la que dicho material per-tenece (entre paréntesis y según las siglas anotadas).Para no reiterar en esta exposición datos de país,provincia, localidad o fecha de captura, se separanpor comas o puntos según se haya citado o no partede la información en los ejemplares mencionadosinmediatamente antes de la que se expone.
Para acceder a sus estructuras y genitalia internas, elabdomen de los imagos era reblandecido en agua ytratado con ácido láctico templado. Las fotografíasse han realizado a través de microscopio OlympusBH-2 y lupa binocular Jaelsa adaptada a cámaraPresentco CCD 3630V con tarjeta digitalizadoraGeforce 3 TI 200.
Las características, condiciones y medios utilizadospara el mantenimiento de los estadios larvarios cul-tivados en el laboratorio con el fin de obtener datossobre su morfología, desarrollo y biología se expo-nen en cada caso.
Resultados y discusión
1 . N e m o p t e r i d a e
MATERIAL ESTUDIADO
Nemoptera bipennis (Illiger, 1812)
Especie de distribución atlanto-mediterránea, cono-cida de España, Portugal, Francia y Marruecos.
Material estudiado:
ESPAÑA: Almería , Cabo de Gata , Los Escul los,9.VII.2005, 1 % avistado, V.J. Monserrat (VM), La Serena,Bédar, 17.VII.2007, 1 &, V.J. Monserrat (VM). Ávila, Cebre-ros, VI.1987, 1 &, M.C. Ochoa (UCM), 1 &, J. Gómez (UCM),La Horcajada, 21.VI.1998, 1 &, A. Arillo (VM), Peguerinos,V.1988, 1 &, M.E. Durán (UCM). Cuenca, Cañete, VI.1987,1 %, P. Bosanta (UCM). Guadalajara, Bolarque, 12.IV.1986,1 %, Anza (UCM). Madrid, Aranjuez, 20.VI.1982, 1 &,E. Ortega (UCM), Cercedilla, Pantano de Navalmedio,27.VI.1980, 2 %%, R. Outerelo (UCM), Cotos, 4.V.1987,1 %, leg.? (UCM), Galapagar, 25.V.1988, 1 %, R.J. Felipe(UCM), Hoyo de Manzanares, 22.VI.1987, 1 %, A. Fer-nández (UCM), 12.III.1996, 1 larva ex ovo a partir de una &capturada el V.1994, V.J. Monserrat (VM), 9.VII.2004, 1 &,11.VI.2005, 4 %%, 1 &, V.J. Monserrat (TAMU), esta última &realizó una puesta de 33 huevos, La Berzosa, 1.V.1988, 1 %,H. Rambla (UCM), La Pedriza, 18.VI.1987, 1 %, F. Silvestre(UCM), Madrid, 2.V.1987, 1 &, C. Grande (UCM), VI.1987,1 %, M. Sánchez (UCM), 17.V.1990, 1 %, M.J. López (UCM),Manzanares, VII.1977, 1 &, J.L. Delgado (UCM), Navas delRey, 11.VI.1988, 1 %, I. Sánchez (UCM), Pelayos, 5.VI.1988,1 %, Fresan (UCM), Pinto, 7.V.1982, 1 %, T. Matrán (UCM),5.V.1987, 1 %, A. Martín (UCM), San Martín de la Vega,13.VI.1984, 1 %, M.B. Chavans (UCM). Murcia, Mazarrón,VII.1984, 1 &, M. Pérez (UCM). Sevilla, Constantina, V.1988,1 &, M.A. Fillol (UCM). Toledo, Talavera, 8.VIII.1987, 1 &,A. Díaz (UCM). Zaragoza, Pina de Ebro, 30TYL29, 350 m,22.V-6.VIII.1991-1993, numerosos ejemplares, frecuentemen-te posados sobre Santolina chamaecyparissus L., J. Blasco (JB).
La morfología y estructura del huevo de esta especiefue descrita por Monserrat (1985) y ahora se anotannuevos elementos para su comparación con los hue-vos de Lertha spp. (Fig. 2). También se aportan nuevosdatos sobre la larva para su comparación con otrasespecies (Figs. 7 y 8), datos que se comentan conjun-tamente más adelante.
Heteropterus Rev. Entomol. (2008) 8(1): 1-33 3

Nemoptera sinuata Olivier, 1811
Especie ponto-mediterránea, conocida de Bulgaria,Grecia, Macedonia, Turquía, Georgia, Armenia,Azerbaiyán, Siria e Irán.
Material estudiado:
BULGARIA: Struma Valley, Kresna, 3-6.VI.1892, 1 %, 1 &,Z. Mészáros (UCM). TURQUÍA: Nurdagi gecidi, Gazian-tep, 27.VI.1971, 1 &, B.G. Osella (MG).
Nemoptera coa (Linnaeus, 1758)
Especie ponto-mediterránea, conocida de Grecia,Cáucaso y Turquía.
Material estudiado:
GRECIA: Peloponeso, Githion, 1.VI.1990, 1 &, Hert(UCM). Hellas mer., Lakonia, Monemvasia, 14.V.1979, 1 &,Gozmány (UCM).
Halter halteratus (Forskal, 1775)
Especie erémica conocida desde Arabia Saudita,Yemen y Kuwait a Sudán, Egipto, Túnez, Argelia yMarruecos.
Material estudiado:
ARABIA SAUDITA: Medina, 20.IV.1977, 1 %, J. Gómez(UCM).
Lertha ledereri (Selys Longchamps, 1866)
Especie erémica conocida de Anatolia y Grecia (Kos).
Material estudiado:
TURQUÍA: Akschehir, 16.VI.1930, 1 %, 1 &, Wagner (NMB).
Lertha sofiae Monserrat, 1988
Especie endémica del sureste árido de la PenínsulaIbérica. Para su comparación con otras especies, ano-tamos algunos elementos del huevo y de los estadiosjuveniles (Figs. 2l y 8d) a partir del material obtenidoy utilizado por Monserrat (1988, 1996), y que discu-timos conjuntamente con las especies siguientes deeste género.
La biología y comportamiento de los imagos en estafamilia había sido recopilada por Monserrat (1996) y,entre otros datos, cita en algunas especies de esta fa-milia la utilización de sus alas posteriores para otrasfunciones al margen del vuelo (disrupción, cripsis,señalización visual, defensa, etc.) y en la especie quenos ocupa menciona su utilización, en situaciones depeligro, para alejar las partes más vitales del cuerpo dela atención y efectos del posible ataque de un depre-dador, hecho que evidencia la Fig. 1. Datos sobre lautilización de las alas en esta familia ya fueron anota-dos por Mansell (1986, 1996), Leon y Picker (1990),Picker y Leon (1990) y Picker et al. (1991, 1992).
Material estudiado:
Huevos y larvas (ex ovo) obtenidos a partir de && recolec-tadas en ESPAÑA: Almería, Balanegra, 20.VII.1991,V.J. Monserrat (VM).
Lertha extensa (Olivier, 1811)
Especie irano-erémica conocida de Anatolia, Jorda-nia, Irak e Irán.
Material estudiado:
Se han estudiado 11 huevos y larvas neonatas de ellosemergidas, obtenidos a partir de hembras grávidasrecolectadas por A. Satar en Turquía, sin habernosespecificado más datos pero que corresponden almaterial anotado por Satar y Özbay (2004a, 2004b).
El aspecto y ultraestructura del huevo de esta espe-cie se ilustran en las Figs. 2d-f y 2j y ahora se anotannuevos elementos para su comparación con los hue-vos de Nemoptera y Lertha (Fig. 2), que se comentanconjuntamente más adelante. La eclosión y ruptura
MONSERRAT: Nuevos datos sobre Nemopteridae y Crocidae (Neuroptera)4
FIGURA 1. Comportamiento defensivo del imago de Lerthasofiae (fotografía de Fernando Rodrigo).
FIGURE 1. Defensive behaviour of the imago of Lertha sofiae(photography by Fernando Rodrigo).

del huevo corresponde a lo descrito por Monserrat(1996) para este género.
Al recibir este material, la mayoría de las larvas yahabían nacido y se hallaban muertas por inaniciónen la muestra, pero a partir de uno de los huevos,se obtuvo una larva viva que se trató de cultivarsiguiendo el método de Monserrat y Martínez (1995)y Monserrat (1996). Las condiciones de cultivo co-rresponden a las de laboratorio en penumbra/oscu-ridad según fotoperiodo natural, humedad relativaambiente y temperaturas máxima y mínima segúnse anota (en ºC): VII.2003 (31/25), VIII (34/24). Lalarva fue aislada en una caja de cultivo de 2 x 4,5 x4,5 cm parcialmente rellena de arena fina, donde seenterró espontáneamente y fue observada periódica-mente cada 10-15 días para seguir su desarrollo yanotar los datos que se consideraran oportunos.Conocidas las preferencias alimentarias en esta fa-milia (Monserrat y Martínez, 1995; Monserrat, 1996),la larva fue alimentada con larvas de Crematogaster
scutellaris (Olivier, 1792) (Hymenoptera: Formicidae),consiguiendo que creciera y prosperara entre el18.VII.2003 y el 28.VIII.2003, fecha en la que fuehallada muerta con las mandíbulas atrapadas por ellíquido desecado de una de su presas.
La larva neonata es de color pardo muy pálido. Trasunas horas de vida fijan su coloración, que va siendomás contrastada conforme madura (Figs. 6g-i). La cáp-sula cefálica es parda oscura dorsalmente, más pálidaen la zona de inserción de las antenas y levementesobre la sutura media occipital (Figs. 6g-h); ventral-mente es parda oscura (Fig. 6i). Antenas, maxilas ypalpos pardos pálidos, al igual que las mandíbulas,cuyo ápice es pardo rojizo más oscuro. Estemas ne-gros. Tegumento torácico y abdominal pardo pálido,con setas y dolichaster pardos oscuros. Pronoto condos manchas circulares pardas más oscuras y dospequeñas líneas longitudinales sobre los dos últimossegmentos, tegumento ventral pardo pálido. Pataspardas, algo más oscuras las P1 y las tibias-tarsos deP2 y P3 (Fig. 6g-i).
Con 40 días de vida, la larva mantiene el estadio y lacoloración cefálica anteriormente anotada, pero noasí la tegumentaria torácica y abdominal, donde seaprecian dorsalmente sendas llamativas bandas pardasa uno y otro lado de la línea media, otra más frag-mentada y tenue hacia el exterior de cada una de ellasy aún otra apenas marcada en la región más externa.Todas ellas están más contrastadas en los segmentostorácicos. Ventralmente la larva mantiene su colora-ción parda pálida citada (Figs. 6j-k).
Discutimos estos datos conjuntamente con la espe-cie siguiente.
Lertha sheppardi (Kirby, 1904)
Especie erémica conocida de Anatolia.
Material estudiado:
Se han estudiado numerosos huevos obtenidos apartir de hembras grávidas recolectadas por A. Sataren Turquía, sin especificarnos más datos, pero quecorresponden al material anotado por Satar y Özbay(2004a, 2004b). A partir de ellos se han obtenido nu-merosas larvas neonatas que han sido cultivadas paraconocer diversos aspectos de su morfología y biolo-gía. Dada la escasa información de que disponemossobre los estadios preimaginales de esta familia, algunade las larvas fue aislada para obtener su desarrollocompleto y describir la larva madura, mientras queotras fueron utilizadas para realizar diversas expe-riencias con el fin de aportar nuevos datos sobre subiología y desarrollo.
Para su cultivo se siguió el método de Monserrat yMartínez (1995) y Monserrat (1996). Las larvas fue-ron bien aisladas o bien agrupados varios ejemplaresen cajas de cultivo de 2 x 4,5 x 4,5 cm, parcialmenterellenas de arena fina, donde las larvas se enterraronespontáneamente. Tras sufrir la segunda muda fuerontrasladadas a cajas cilíndricas de cultivo de 4,5 cm dealto y 7 cm de diámetro, también parcialmente rellenasde arena fina, y fueron observadas periódicamentecada 10-15 días para seguir su desarrollo y anotar losdatos que se consideraran oportunos. Las larvas empe-zaron a eclosionar un día después de recibir el envío,a partir del 18.VI.2003, por lo que, a diferencia de laespecie anterior, dispusimos de numerosos ejemplarespara su cultivo y obtuvimos diferentes resultados ensu desarrollo, en función de las experiencias realiza-das sobre ellas.
Conocidas sus preferencias alimentarias (Monserraty Martínez, 1995; Monserrat, 1996), los ejemplaresfueron alimentados con larvas y/o pupas de Cremato-
gaster scutellaris (Olivier, 1792), Tetramorium forte Forel,1904, Messor structor (Latreille, 1798), Messor barbarus
(Linnaeus, 1767), Monomorium salomonis (Linnaeus,1758), Tapinoma nigerrima (Nylander, 1856), Formica py-
renaea Bondroit, 1918, Lasius alienus (Foerster, 1850),Aphaenogaster senilis Mayr, 1853 y Camponotus cruentatus
(Latreille, 1802) (Hymenoptera: Formicidae). Paraestudiar la incidencia en su desarrollo, en ocasiones
Heteropterus Rev. Entomol. (2008) 8(1): 1-33 5

sólo se les proporcionaron larvas, en otros casossólo pupas o ambos tipos de potenciales presas.
Las condiciones de cultivo de estas larvas corres-ponden a las de laboratorio en penumbra/oscuridadsegún fotoperiodo natural, humedad relativa am-biente y temperaturas máxima y mínima según se
anota (en ºC): VII.2003 (31/25), VIII.2003 (34/24),IX.2003 (34/21), X.2003 (22/15), XI.2003 (24/15),XII.2003 (21/12), I.2004 (20/12), II.2004 (24/14),III.2004 (24/14), IV.2004 (23/15), V.2004 (25/16),VI.2004 (32/25), VII.2004 (33/25), VIII.2004 (32/24),IX.2004 (29/23), X.2004 (26/17), XI.2004 (23/16),
MONSERRAT: Nuevos datos sobre Nemopteridae y Crocidae (Neuroptera)6
FIGURA 2. Huevos de: (a)-(c) Nemoptera bipennis: (a) Aspecto general; (b) Aspecto de su micropilo; (c) Aspecto del corion;(d-f) Lertha extensa, ídem; (g)-(i) L. sheppardi, ídem; (j)-(l) Detalle del corion de: (j) L. extensa; (k) L. sheppardi; (l) L. sofiae.
FIGURE 2. Eggs of: (a)-(c) Nemoptera bipennis: (a) General aspect; (b) Aspect of its micropyle; (c) Aspect of the chorion; (d-f) Lerthaextensa, ditto; (g)-(i) L. sheppardi, ditto; (j)-(l) Detail of the chorion of: (j) L. extensa; (k) L. sheppardi; (l) L. sofiae.
(a)
(d)
(g)
(j)
(b)
(e)
(h)
(k)
(c)
(f)
(i)
(l)

XII.2004 (22/15), I.2005 (22/14), II.2005 (22/14),III.2005 (24/15), IV.2005 (23/16), V.2005 (27/20),VI.2005 (32/26), VII.2005 (32/27), VIII.2005 (31/25),IX.2005 (30/21), X.2005 (23/19), XI.2005 (23/17),XII.2005 (22/16), I.2006 (22/16), II.2006 (23/17),III.2006 (23/14).
Citadas las condiciones de cultivo, pasamos a la des-cripción de sus estadios preimaginales.
Los huevos no viables permanecen de color azul-verdoso, los viables muestran las bandas metaméri-cas/segmentarias y los estemas y setas pardos oscurosvisibles por transparencia. La eclosión y ruptura delhuevo corresponde a lo anotado por Monserrat(1996) para este género; se ayudan empujando conel vértex y el pronoto. El ovirruptor es pardo y elcorion abandonado es blanco intenso. El aspecto yultraestructura del huevo de esta especie se anota enlas Figs. 2g-i y 2k. Estos datos se discuten, más ade-lante, conjuntamente con los de las especies anteriores.
Las larvas neonatas son de coloración general blan-quecina con setas negras. Tras unas horas de vidafijan su coloración (Figs. 6a-c). En la región dorsalde la cabeza poseen una mancha parda oscura entrela base de las mandíbulas, que se continúa con unamancha circular a cada lado de la línea media, y losmárgenes laterales y el margen occipital son pardososcuros, siendo llamativo el extremo de las mandí-bulas de color pardo rojizo. Antenas y palpos pardospálidos. Estemas negros. El tórax porta dos conspi-cuas manchas pardas sobre el pronoto y dos hilerasde manchas irregulares pardo-rojizas sobre los otrosdos segmentos, que se continúan fragmentadamentesobre el abdomen. El tórax y el abdomen conservanuna banda media interna de color verdoso, similar al
del vitelo. Patas pardo-grisáceas, especialmente desdela mitad distal del fémur. Esta coloración general sehace más patente y se fija conforme las larvas crecen(Figs. 7k-l), añadiéndose nuevas hileras longitudi-nales de manchas pardo-rojizas hacia la zona lateraldel abdomen. La banda más próxima a la línea mediaes más continuada, la intermedia más fragmentada yla más externa más tenue (Figs. 6a-c).
Las larvas maduras (Figs. 7e-h) poseen la cápsula ce-fálica dorsalmente parda oscura con estemas negros,setas pardo-rojizas, antenas y maxilas pálidas y man-díbulas pardo-rojizas; ventralmente la cápsula cefálicaes uniformemente parda. Patas pardas pálidas, másoscuros los tarsos. Tórax y abdomen con tegumentopardo pálido con setas pardas oscuras-negras; en laregión dorsal del tórax se disponen dos bandas lon-gitudinales pardas en zigzag que se continúan sobreel abdomen, donde, a ambos lados de ellas, se dis-pone una banda media discontinua. Hacia la zonamedia las bandas laterales se bifurcan; la banda máscentral es ancha y la más externa forma manchasarqueadas que rodean los estigmas y portan largassetas. Ventralmente a los estigmas existen similaresmanchas arqueadas mucho más pálidas. Últimos seg-mentos parduscos con setas rojizas (Figs. 7g-h).
Algunos aspectos de su biología han sido compren-didos a partir de las diversas experiencias y condi-ciones a las que hemos sometido a estas larvasdurante los tres años en los que las hemos mantenidocriándolas en el laboratorio. Estos resultados, juntocon los datos de morfología y ultraestructura delhuevo y morfología de las larvas, se comentan, másadelante, conjuntamente con alguna de las especiesanteriormente citadas.
Heteropterus Rev. Entomol. (2008) 8(1): 1-33 7
(a) (b) (c)
FIGURA 3. Aspectos del huevo de Carausius morosus (Brunner, 1907) (Insecta: Phasmida).
FIGURE 3. Aspects of the egg of Carausius morosus (Brunner, 1907) (Insecta: Phasmida).

DISCUSIÓN SOBRE LA BIOLOGÍA Y MORFO-LOGÍA DE LOS ESTADIOS PREIMAGINALES
EN NEMOPTERIDAE
Respecto a la biología de su desarrollo preimaginaldebe indicarse que, a pesar del tamaño y vistosidadde sus imagos, el estado de conocimiento sobre losestadios preimaginales de esta familia es, sorpren-dentemente, muy escaso e incompleto. Tjeder (1967)compiló los datos existentes hasta entonces y poste-riormente Monserrat (1985, 1996) y Tröger (1993)revisan el estado de conocimiento hasta esa fechasobre este particular y Monserrat y Martínez (1995)demuestran las preferencias alimentarias (larvas dehormigas) y sugieren una posible mirmecofilia en suciclo biológico, hecho que no parecía concordar conlo anotado por Tröger (1993) para Nemoptera coa, peroque es sugerido por Popov (1998, 2002); asimismo,
el relativo éxito en el cultivo de estos ejemplarescon larvas/pupas de hormigas que aportamos ahoraparece seguir corroborándolo (Tabla 1). Es cierto queninguno de los ejemplares ahora criados consiguiópupar, y sólo uno inició, sin completarlo con éxito,este proceso (Tabla 1: C5) por lo que podría sugerirsela mirmecofilia sólo en los primeros estadios de suciclo biológico, hecho que sí parece corroborar loanotado por Tröger (1993) para Nemoptera coa, y qui-zás otro tipo de presa/alimento deba añadirse en sudesarrollo. En cualquier caso, citamos algunas consi-deraciones sobre su biología que merecen anotarse yotras que han sido obtenidas a partir de ciertas expe-riencias realizadas sobre las larvas en desarrollo yque quedan reflejadas en la Tabla 1.
• Las larvas neonatas permanecen unos minutos enreposo antes de comenzar su actividad, siendo des-pués muy activas, y tienden a enterrarse activamente
MONSERRAT: Nuevos datos sobre Nemopteridae y Crocidae (Neuroptera)8
(e)
(f)
(g)
(h) (i) (j)
(a) (b)
(c) (d)
FIGURA 4. Aspectos de la biología ycomportamiento de Lertha sheppardi:(a)-(b) Corion abandonado; (c)-(d) Ca-nibalismo en L1; (e)-(j) Comporta-miento de voltearse para recuperar laposición normal en L3.
FIGURE 4.Aspects of the biology and be-haviour of Lertha sheppardi: (a)-(b) Aban-doned chorion; (c)-(d) Cannibalism inL1; (e)-(j) Behaviour to turn around torecover the normal position in L3.

abriéndose paso con las mandíbulas y ayudándoseespecialmente del primer par de patas, como se haanotado en otras especies (Monserrat, 1985, 1996;Monserrat y Martínez, 1995).
• Los datos en el modo de alimentación, comporta-miento, tanatosis, tigmotactismo y forma de reali-zar la ecdisis (incluida la fijación en el substrato delápice abdominal para favorecerla) coinciden con loanotado por Tröger (1993), Monserrat y Martínez(1995) y Monserrat (1996) para otras especies deNemoptera y Lertha. Su comportamiento y capacidadde poder voltearse para recuperar la posición nor-mal se ilustra en las Figs. 4e-j.
• Al margen de ciertas experiencias a las que hemossometido a algunos ejemplares (Tabla 1: A, B), laslarvas alimentadas adecuadamente (Tabla 1: C) sufrendos mudas y tres estadios larvarios que requierencasi dos años para completar su desarrollo (Tabla 1:
C5) y que, de forma general, coincide con lo anotadopor Monserrat y Martínez (1995) y Monserrat(1996) y Popov (1998) para otras especies ibéricasy griegas de Nemoptera y Lertha, aunque, quizás poradaptación a una mayor xericidad o a un más breveperiodo de bonanza en los recursos alimentariosoriginales, en estas especies turcas parece adelan-tarse la primera muda (a los 30-45 días frente a45-60/72 días respecto a las especies ibéricas/grie-gas), así como la segunda muda (a los 15-35 díasfrente a 2-10 meses en las especies ibéricas segúnlo anotado por Monserrat y Martínez (1995) y Mon-serrat (1996) ), hechos que refleja la Tabla 1: C yque podría sugerir en estas especies la posibilidadde poseer un ciclo biológico más corto.
• Las larvas a las que se les proporcionó exclusiva-mente pupas de hormigas como alimento no consi-guieron mudar, muriendo a las pocas semanas (Ta-
Heteropterus Rev. Entomol. (2008) 8(1): 1-33 9
TABLA 1. Tiempos de desarrollo (....) hasta su muerte (+) o hasta el inicio de su pupación (P) y mudas (M1, M2) sufridas por algunaslarvas de Lertha sheppardi (A1, B1-5, C1-5) sometidas a diferentes tipos de alimentación: Grupo A: alimentada sólo con pupas de hor-migas; Grupo B: alimentadas con pupas y larvas de hormigas; Grupo C: alimentadas sólo con larvas de hormigas. En abscisas, eltiempo en meses (a diferente escala según 1º ó 2º-4º año). Para la larva C3 la flecha (ò) indica el momento en el que se le dejó deproporcionar alimento (explicación en el texto).
TABLE 1. Times of development (....) before its death (+) or the beginning of the pupation (P) and moultings (M1, M2) under-gone by some larvae of Lertha sheppardi (A1, B1-5, C1-5) breaded under different types of feeding: Group A: fed only with pupae ofants; Group B: fed with pupae and larvae of ants; Group C: fed only with larvae of ants. In abscissas the time in months (diffe-rent scales according to 1st or 2nd-4th year). For the C3 larva the arrow (ò) indicates the moment at which feeding was stopped(explanation in the text).

bla 1: A). Las larvas a las que se les proporcionaroncomo alimento pupas y larvas de hormigas mez-cladas (Tabla 1: B) o no consiguieron mudar o sóloconsiguieron mudar una vez o bien las mudas mues-tran cierta irregularidad temporal respecto a las quefueron exclusivamente alimentadas con larvas de hor-migas, siendo en cualquier caso mucho más brevesu periodo de vida respecto a las larvas a las que seles ofreció exclusivamente larvas de hormigas (Ta-bla 1: C), las cuales aumentan significativamente sulongevidad y consiguen un mayor desarrollo res-pecto a ellas, que no prosperaron y/o permanecie-ron más retrasadas (Tabla 1: B).
• Una de las larvas en normal desarrollo (Tabla 1: C3)fue sometida a completa inanición, consiguiendo so-brevivir 15 meses sin alimentarse. De esto se deduceuna enorme capacidad de ayuno en los estadios lar-varios de esta familia, ya sugerido (9 meses) porPopov (1998) para Nemoptera sinuata.
• Las larvas neonatas colocadas juntas (de dos endos) en la misma caja de cultivo presentan un ele-vado grado de canibalismo, al menos en esta faseinicial, y especialmente en el caso de aquellas a lasque se les ofreció exclusivamente pupas como ali-mento (Tabla 1: A). Este comportamiento fue indi-cado por Monserrat (1996) para larvas de Nemoptera
bipennis sometidas a inanición y se muestra en lasFigs. 4c-d.
• En función de las disponibilidades alimentarias, eldiferente tiempo entre mudas y en el desarrollo delas larvas (Tabla 1) permite, sin duda, el flujo gené-tico entre generaciones, hecho ya sugerido porMonserrat (1996).
• En algunos casos, en el interior de las cajas de cul-tivo, los imagos de hormigas que fueron emergiendoa partir de las pupas ofrecidas como alimento, reor-ganizaban su nuevo nido a partir de los materialesexistentes en ella y se observó que transportaban nosólo sus larvas, sino también las larvas de Lertha,de lo que se deducen mecanismos de inhibición oadquisición del olor de la colonia, hecho ya sugeridopor Monserrat y Martínez (1995).
• La citada muerte del ejemplar de Lertha extensa fuecausada por secreciones de la larva de la hormigasobre la que depredaba y corresponde a uno de losmecanismos de defensa habitual en larvas de hor-migas y de Crematogaster en particular (Leclercq etal., 1997; Hölldobler y Wilson, 1990), hecho que,junto con las conocidas adaptaciones para evitar elataque de las hormigas mediante un particular tipo
de puesta/morfología del huevo o mediante el com-portamiento de las larvas neonatas (bien conocidoen Ascalaphidae, Nymphidae, Berothidae, Chryso-pidae o Mantispidae), no es el único caso descritoentre las reacciones de las hormigas ante la depre-dación de neurópteros o viceversa (Halversar et al.,1976; Hölldobler y Wilson, 1990; Tedders et al.,1990; Gatti y Farju-Brener, 2002).
Con respecto al huevo de estas especies de esta fami-lia, debemos indicar que, al margen de algunos datosa veces erróneos, dudosos o muy vagos y en ocasionesreiterativos que Monserrat (1996) recopila, dentro deesta familia sólo existen datos sobre el huevo de losgéneros Nemoptera, Knersvlaktia, Palmipenna y Lertha.Lo que se conocía del huevo de Nemoptera bipennis
(Figs. 2a-c) fue recopilado por Monserrat (1985), elde Nemoptera sinuata fue descrito por Popov (1963,1973, 1998, 2002) y Candan et al. (2005), el deKnersvlaktia nigroptera fue descrito por Picker (1984), elde Palmipenna aeoleoptera fue descrito por Picker (1987)y el de Lertha sofiae (Fig. 2l) fue descrito por Monse-rrat y Martínez (1995) y Monserrat (1996), anotandoahora nuevos elementos para su comparación con losde otras especies (Fig. 2l); por último, Satar y Özbay(2004a) dan cuenta del aspecto externo de los hue-vos de Lertha extensa y Lertha sheppardi.
Los huevos de Palmipenna y Knersvlaktia son adhe-sivos, muy fusiformes y alargados, con corion sinestructuras hexagonales y con micropilo de aspectofungiforme (Picker, 1984, 1987), mientras que en Ler-
tha o Nemoptera no son adhesivos y son más esféricosy redondeados, con corion portando estructurashexagonales y con micropilo de aspecto de chapa obotón (Figs. 2b, 2e y 2h), siendo los de Lertha bas-tante más elipsoides que en Nemoptera (Fig. 2a frentea 2d), que posee huevos más esféricos y donde lasceldillas hexagonales del corion son menos convexas,menos regulares y de límites más flexuosos que enLertha (Fig. 2c frente a 2f y 2i). Dentro del géneroLertha anotamos marcadas diferencias entre las tresespecies de las que hasta ahora conocemos o hemosdescrito: En Lertha extensa la formación que delimitalas celdillas hexagonales está profusamente fragmen-tada y éstas poseen una superficie fuertemente escul-pida (Fig. 2j), mientras que en Lertha sheppardi y enLertha sofiae esta formación marginal es continua y deaspecto subcilíndrico (Figs. 2k y 2l), diferenciándoseuna y otra en una mayor complejidad en las esculturasde la superficie de las celdillas hexagonales en Lertha
sofiae respecto a Lertha sheppardi (Fig. 2l frente a 2k).
MONSERRAT: Nuevos datos sobre Nemopteridae y Crocidae (Neuroptera)10

Parece evidente que existe cierta similitud entre loshuevos tanto de Nemopteridae (Figs. 2a-b, 2d-e, 2g-h)como de otros insectos mirmecófilos como losinsectos palo (Fig. 3) y el eleosoma (elaiosoma) deciertas semillas de plantas (Fig. 5) que utilizan la mir-mecocoria para completar su ciclo biológico o dis-persar sus semillas (Hanzawa et al., 1988; Huges yWestoby, 1992; Gullan y Cranston, 1996; Handel yBeattie, 1998). Este hecho podría corroborar la mir-mecofilia sugerida para las larvas de esta familia porMonserrat y Martínez (1995).
Acabado el desarrollo embrionario, en las especiesde Lertha ahora estudiadas, la larva también realizauna ruptura incompleta del corion para emerger, lacual, a diferencia de Knersvlaktia, que la efectúa anivel del ápice micropilar (Picker, 1984), en Lertha serealiza a nivel de 1/3 de su longitud, como ha sidodescrito habitualmente en esta familia (Withycombe,1925; Tjeder, 1967; Monserrat, 1996; Popov, 2002;Candan et al., 2005) y el corion abandonado es decolor blanco intenso y a él queda adherido el oscuroovirruptor (Figs. 4a-b).
En relación con las larvas de esta familia, sorprenden-temente existen muy pocos datos, a veces erróneos,dudosos o muy vagos, y en ocasiones reiterativos,que Monserrat (1996) recopila. Circunscribiéndonosa los datos más recientes, se conocen las larvas de losgéneros Nemoptera (Popov, 1963, 1973, 1998, 2002;Tröger, 1993; Monserrat y Martínez, 1995; Monse-rrat, 1996; Satar y Özbay, 2004a; Candan et al., 2005),Chasmoptera (Mathews, 1947; Tjeder, 1967), Derhyn-
chia (Mansell, 1973), Semirhynchia (Mansell, 1996) yLertha (Monserrat, 1996; Satar y Özbay, 2004a), que,en general, muestran un elevado grado de uniformi-dad morfológica (Figs. 7 y 8), muy distinto a lo queveremos en Crocidae (Fig. 11). Por otra parte, la ma-yoría de las descripciones ofrecen el aspecto generalde las larvas, sin descender a mayor detalle ni ultraes-tructura que pudiera darnos datos sobre nuevas dife-rencias dentro de su aparente uniformidad. Las larvasneonatas del género Lertha son de una gran unifor-midad morfológica y de coloración (Fig. 6) y, ya queésta se fija tras unas horas de vida, puede existircierta variabilidad entre los ejemplares estudiados enfunción de su edad (ver Satar y Özbay, 2004a).
Referencias sobre las pupas sólo son conocidas deforma vaga y general (Popov, 1973; Sengonça, 1981)y para Derhynchia (Mansell, 1973) y Nemoptera (Trö-ger, 1993).
Aunque Tröger (1993), Mansell (1996) y Monserrat
(1996) comentan elementos distintivos entre deter-minadas larvas de esta familia, podemos aportarahora, con los elementos de que disponemos, unaclave de larvas de los géneros hasta el momentoconocidos.
CLAVES DE IDENTIFICACIÓN DE LARVASCONOCIDAS DE NEMOPTERIDAE
En primer lugar, presentamos una clave de génerospara las larvas desarrolladas de Nemopteridae cono-cidas.
Por otra parte, y aunque sólo son conocidas las larvasde 4 de las 6 especies del género Nemoptera y 3 de las12 especies de Lertha, podemos aportar unas prime-ras claves (las tres claves siguientes) de sus estadiospreimaginales, basadas en lo ahora descrito.
Debe insistirse en que se trata de claves para aquellosgéneros y especies en que se conocen sus larvas.
Heteropterus Rev. Entomol. (2008) 8(1): 1-33 11
FIGURA 5. Semillas que utilizan la mirmecocoria para su dis-persión (de Handel y Beattie, 1998).
FIGURE 5. Seeds that use myrmechocory for its dispersion(from Handel and Beattie, 1998).

MONSERRAT: Nuevos datos sobre Nemopteridae y Crocidae (Neuroptera)12
Clave de géneros para larvas desarrolladas de Nemopteridae
(1) Cabeza circular y redondeada en su margen posterior, ciega, sin estemas. Mandíbula progresiva- y unifor-memente adelgazada hacia su extremo (Fig. 8a) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Derhynchia
- Cabeza cuadrada y acodada en su margen posterior (Figs. 7a, 7e y 8b), no ciega, con 7 estemas (Figs. 7a y 7e).Mandíbula irregular- y súbitamente adelgazada hacia su extremo (Figs. 7a, 7e y 8b) . . . . . . . . . . . . . . . . . . . . 2
(2) Mandíbulas rectas sólo con el ápice curvado (Fig. 8b) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Semirhynchia
- Mandíbulas marcadamente curvas (Figs. 7a, 7e y 8c) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3
(3) Mandíbulas progresivamente adelgazadas hacia su extremo (Fig. 8c) . . . . . . . . . . . . . . . . . . . . Chasmoptera
- Mandíbulas progresivamente adelgazadas, pero de forma muy remarcada en su extremo (Figs. 7a y 7e) . . . . 4
(4) Estemas dispuestos en dos arcos dorsales de 3 unidades sobre un estema mayor de posición ventral oanterior. Último segmento antenal marcadamente subsegmentado (Fig. 7m-B) . . . . . . . . . . . . . . . . Nemoptera
- Estemas dispuestos en un arco dorsal de 5 unidades rodeando uno central sobre otro estema más ventral.Último segmento antenal muy tenuemente subsegmentado (Fig. 7m-A) . . . . . . . . . . . . . . . . . . . . . . . . Lertha
Clave de especies para larvas neonatas de Lertha
(1) Cápsula cefálica dorsal mayoritariamente parda oscura salvo la zona próxima a la base de las mandíbulasy sobre la sutura media occipital, que son más pálidas (Figs. 6g-h) . . . . . . . . . . . . . . . . . . . . . . . . . . . L. extensa
- Cápsula cefálica en la región dorsal pálida con la zona próxima a las dos ramas de la sutura epicraneal («Y»)y márgenes laterales y posterior de color oscuro (Figs. 6a-b y 6d-e) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2
(2) Manchas del pronoto marcada-llamativamente pardas. Setas cefálicas, torácicas y abdominales de aspectonormal, mucho más abundantes que los dolichaster (Figs. 6a-c) . . . . . . . . . . . . . . . . . . . . . . . . . . . L. sheppardi
- Manchas del pronoto poco destacadas del resto del tegumento. Setas cefálicas, torácicas y abdominales deaspecto normal, mucho menos abundantes que los dolichaster (Figs. 6d-f) . . . . . . . . . . . . . . . . . . . . . L. sofiae
Clave de especies para larvas maduras de Lertha
(sólo conocida hasta final de L1 en L. extensa)
(1) Bandas pardas laterales a ambos lados de la línea media dorsal sobre el tórax y el abdomen marcadamenteinterrumpidas en manchas aisladas (Fig. 8d) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . L. sofiae
- Bandas pardas laterales a ambos lados de la línea media dorsal sobre el tórax y el abdomen más o menoscontinuadas, sin formar manchas marcadamente aisladas (Figs. 7i y 7k) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2
(2) Manchas pardas sobre el pronoto apenas marcadas. Bandas pardas laterales a ambos lados de la líneamedia dorsal sobre el tórax y el abdomen irregulares e interrumpidas entre los segmentos (Fig. 7i). Regiónventral con dos bandas pardas longitudinales bien marcadas (Fig. 7j) . . . . . . . . . . . . . . . . . . . . . . . . . L. extensa
- Manchas pardas sobre el pronoto fuertemente marcadas. Bandas pardas laterales a ambos lados de la líneamedia dorsal sobre el tórax y el abdomen regulares y continuas, no interrumpidas entre los segmentos (Figs.7f y 7k). Región ventral sin dos bandas pardas longitudinales bien marcadas (Figs. 7g y 7l) . . . . . L. sheppardi
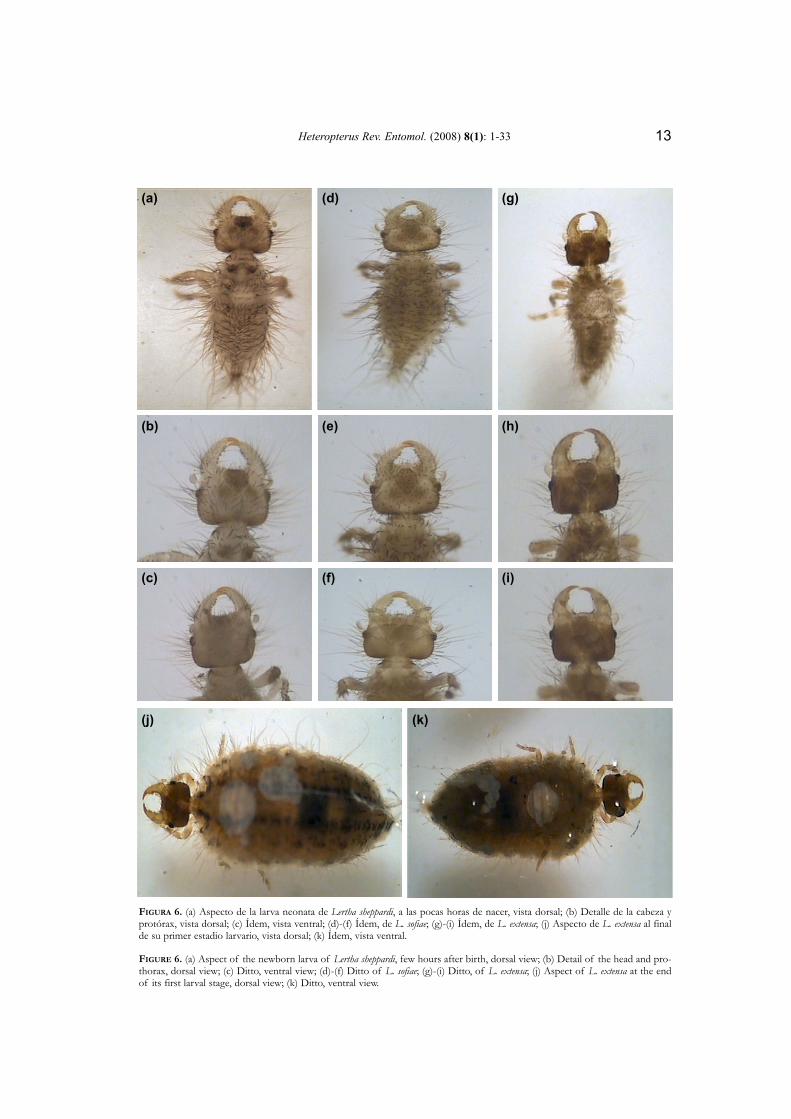
Heteropterus Rev. Entomol. (2008) 8(1): 1-33 13
(a)
(b)
(c)
(d)
(e)
(f)
(g)
(h)
(i)
FIGURA 6. (a) Aspecto de la larva neonata de Lertha sheppardi, a las pocas horas de nacer, vista dorsal; (b) Detalle de la cabeza yprotórax, vista dorsal; (c) Ídem, vista ventral; (d)-(f) Ídem, de L. sofiae; (g)-(i) Ídem, de L. extensa; (j) Aspecto de L. extensa al finalde su primer estadio larvario, vista dorsal; (k) Ídem, vista ventral.
FIGURE 6. (a) Aspect of the newborn larva of Lertha sheppardi, few hours after birth, dorsal view; (b) Detail of the head and pro-thorax, dorsal view; (c) Ditto, ventral view; (d)-(f) Ditto of L. sofiae; (g)-(i) Ditto, of L. extensa; (j) Aspect of L. extensa at the endof its first larval stage, dorsal view; (k) Ditto, ventral view.
(j) (k)

MONSERRAT: Nuevos datos sobre Nemopteridae y Crocidae (Neuroptera)14
FIGURA 7. (a) Larva madura de Nemoptera bipennis, cabeza y protórax, vista dorsal; (b) Ídem, aspecto general, vista dorsal; (c) Ídem,vista ventral; (d) Ídem, últimos segmentos abdominales, vista ventral; (e)-(h) Ídem de Lertha sheppardi; (i) Aspecto de L. extensa alfinal de su primer estadio larvario, vista dorsal; (j) Ídem, vista ventral; (k) Aspecto de L. sheppardi al final de su primer estadio lar-vario, vista dorsal; (l) Ídem, vista ventral; (m) Antena de larva de Lertha sofiae (A) y de Nemoptera bipennis (B) (de Monserrat, 1996).
FIGURE 7. (a) Mature larva of Nemoptera bipennis, head and prothorax, dorsal view; (b) Ditto, general aspect, dorsal view; (c) Ditto,ventral view; (d) Ditto, last abdominal segments, ventral view; (e)-(h) Ditto of Lertha sheppardi; (i) Aspect of L. extensa at the endof its first larval stage, dorsal view; (j) Ditto, ventral view; (k) Aspect of L. sheppardi at the end of its first larval stage, dorsal view;(l) Ditto, ventral view; (m) Antenna of the larva of Lertha sofiae (A) and Nemoptera bipennis (B) (from Monserrat, 1996).
(a)
(b)
(c)
(e)
(f)
(g)
(i)
(j) (l)
(d) (h)
(k)
(m)
A B
mm

Heteropterus Rev. Entomol. (2008) 8(1): 1-33 15
Clave de especies para larvas neonatas/maduras de Nemoptera
(1) Bandas longitudinales pardas sobre la región dorsal y ventral del tórax y abdomen bien marcadas y continuas.Cabeza con mancha parda dorsal en «T» invertida. Márgenes cefálicos laterales y caudal negros (Figs. 7a-d) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . N. bipennis
- Bandas longitudinales pardas sobre la región dorsal y ventral del tórax y abdomen tenues y fragmentadas.Cabeza dorsalmente parda. Márgenes laterales y caudal de la cabeza pardos . . . . . . . . . . . . . . . . . . . . . . . . . . 2
(2) Cabeza dorsalmente parda . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . N. coa
- Cabeza parda con una mancha dorsal marcadamente más oscura . . . . . . . . . . . . . . . . . . . . . . . . N. sinuata
FIGURA 8. (a) Aspecto de larva madura de Derhynchia, vista dorsal (de Mansell, 1973); (b) Aspecto de la larva madura deSemirhynchia, vista dorsal (de Mansell, 1996); (c) Aspecto de la larva, probablemente inmadura, de Chasmoptera, vista dorsal (deMathews, 1947); (d) Aspecto de la larva madura de Lertha sofiae, vista dorsal (de Monserrat, 1996).
FIGURE 8. (a) Aspect of the mature larva of Derhynchia, dorsal view (from Mansell, 1973); (b) Aspect of the mature larva ofSemirhynchia, dorsal view (from Mansell, 1996); (c) Aspect of the larva, probably immature, of Chasmoptera, dorsal view (fromMathews, 1947); (d) Aspect of the mature larva of Lertha sofiae, dorsal view (from Monserrat, 1996).
(a) (b) (d)(c)

2 . C r o c i d a e
MATERIAL ESTUDIADO
Josandreva sazi Navás, 1906
El género Josandreva es conocido del sureste de laPenínsula Ibérica, Chad e isla de Sokotra (Yemen) yla especie que citamos es marcadamente erémica, detendencia antropófila, frecuentemente citada de habi-taciones humanas (Monserrat, 1979, 1983a; Monserratet al., 1999) y hasta ahora sólo era conocida del sur-este y centro de la Península, aunque Aspöck et al.
(2001) la dan como probable en el norte de África,hecho que ahora confirmamos.
El ejemplar que citamos de Marruecos es una &,capturada en el interior de una vivienda humana, yposee una morfología externa, especialmente la lon-gitud del rostro y del pronoto, muy similar al restodel material ibérico estudiado y a lo citado para estaespecie (Hölzel, 1975a); su tegumento es más pálidoy sus alas son de mayor envergadura (longitud del alaanterior: 13,2 mm) que en ejemplares ibéricos delmismo sexo (longitud del ala anterior: 10,4 mm) yposeen una mayor tendencia a presentar celdillaspoligonales en el campo radial (Figs. 9g-h).
Aunque pudiera pertenecer a una nueva especie,dado que existe otra especie de esta familia (Pterocroce
capillaris) que también habita en el sureste de laPenínsula Ibérica y está presente a ambos lados delEstrecho de Gibraltar, mientras no dispongamos de%% que nos demuestren lo contrario y antes de des-cribir una nueva especie potencialmente sinónima,preferimos citar este ejemplar como perteneciente ala mencionada especie ibérica, que es nueva paraMarruecos y para el continente africano.
La larva descrita por Pierre (1952) de Argelia y porél asociada a Berlandus saharae Pierre, 1952 = Dielo-
croce berlandi (Navás, 1936) no corresponde a lo queconocemos de los estadios larvarios en Dielocroce
(Fig. 11m-A), tanto por la escasa longitud del protó-rax, como por la posesión de dentículos (uno espe-cialmente desarrollado) en el margen interno de lasmandíbulas, hechos que, por el contrario, coincidencon lo que conocemos del género Josandreva (Fig. 11a)que ahora citamos de Marruecos, por lo que podría-mos sugerir una mayor extensión del género en estazona septentrional del continente africano.
Material estudiado:
ESPAÑA: Alicante, Orihuela, VII.1905, 1 %, VII.1911, 3 %%,VI.1934, 3 ej., Andreu (MZB), de esta localidad, y bastantedeteriorados, hemos estudiado 3 ejemplares (NHM).Almería, Las Negras, 22.VII.1998, 1 % volando al atardeceralrededor de oquedad en pared de rambla, V.J. Monserrat(VM), Ocaña, 26.VII.1991, 4 %% en cueva con suelo are-noso, V.J. Monserrat (VM), Tabernas, 25.VII.1991, 18 %%,4 && en casa en ruinas, V.J. Monserrat (VM). Murcia, SierraEspuña, El Azaraque, 17.VIII.1984, 3 %%, E. Martínez Or-tega (VM), 31.VIII.1984, 2 %% en cueva con suelo pedre-goso, E. Martínez Ortega (VM). MARRUECOS: Azilal,Ait-Bouguemez Douar Rit-Zizj, 22-26.VIII.1989, 1 & entrampa adhesiva en el interior de vivienda, E. MartínezOrtega (VM).
Croce schmidti (Navás, 1927)
Especie irano-erémica, conocida de Pakistán, Afga-nistán, Irán, Irak, Yemen, Arabia Saudita e Israel, queahora citamos para Anatolia (Turquía), ampliandosignificativamente el límite occidental de su distri-bución.
Los ejemplares que ahora citamos poseen una mor-fología y genitalia que coincide con lo conocido paraesta especie (Hölzel, 1975a), siendo algo menor laenvergadura alar en los %% (longitud del ala anterior:9 mm, frente a 11 mm habitualmente registrado eneste sexo).
El aspecto de su larva neonata se anota por primeravez en la Fig. 9f.
Material estudiado:
TURQUÍA: Derik province, Mardin city, 14.VIII.2004,4 %%, 3 &&, A. Satar et al. (VM). Conozco la existencia deotros 16 %%, 27 && con iguales datos de captura (DBDU).
Laurhervasia setacea (Klug, 1836)
Especie conocida de Sudáfrica y Namibia.
La larva de esta especie fue descrita por Mansell(1976, 1980) y aportamos su aspecto general en lasFigs. 9j-l. Aun siendo una especie cuyas larvas pare-cen poseer una morfología y pigmentación variable(Mansell, 1976, 1980), coincidimos con él en losargumentos que esgrime para no considerar válida lasinonimia entre el género Laurhervasia Navás, 1914 yThysanocroce Withycombe, 1923 que propuso Tjeder(1967) y mantiene Hölzel (1975a) y que, entre otros
MONSERRAT: Nuevos datos sobre Nemopteridae y Crocidae (Neuroptera)16

caracteres de venación y genitalia masculina, difierenen las muy diferentes morfología (Figs. 9j-l y 11hfrente a 11f) y biología de sus larvas (Mansell, 1980).
Material estudiado:
REPÚBLICA DE SUDÁFRICA: Northern Cape Province,along road east of Upington, 12.II.1988, 1 larva, P. Duelli(VM).
Dielocroce ephemera (Gerstaecker, 1894)
Especie irano-erémica conocida de Anatolia, Irak eIrán.
A partir de material capturado en Turquía (SE Ana-tolia, Batman-BeSiri, 1020 m, 3.VII.2002, 3 %%, 7 &&)y asignado a esta especie por A. Satar (Satar y Özbay,2004b) se obtuvo un huevo de esta especie (Figs. 9a-e)que nos ha permitido estudiar y describir su morfo-logía y ultraestructura. Sólo se conocía la morfologíadel huevo en este género para D. baudii (Griffini,1895), especie irano-erémica conocida de Chipre,Israel, Anatolia, Irak y Afganistán, recientementedescrito por Suludere et al. (2006).
El huevo es de aspecto elipsoide, típico en esta fa-milia, con el corion sin aparentes impresiones hexa-gonales (Fig. 9a) y sin aeropilos bien delimitados yaislados como ha sido descrito en otros géneros(Mansell, 1977, 1980, 1981a, 1981b; Monserrat, 1983b,1985), sino que es portador de numerosos aeropilosirregulares de aspecto esponjoso (Fig. 9c-d), aparen-temente mucho más abundantes y esponjosos queen D. baudii (Suludere et al., 2006), y el micropilo essemiesférico y compacto (Fig. 9b).
Dielocroce baudii (Griffini, 1895)
Especie irano-erémica conocida de Chipre, Israel,Anatolia, Irak y Afganistán.
Material estudiado:
TURQUÍA: Adiyaman City, Nemrut Mountain, 5.VIII.2005,vista alguna larva obtenida (ex ovo) a partir de 9 %%, 12 &&,A. Satar et al. (DBDU).
Dielocroce berlandi (Navás, 1936)
Especie afro-sirio-erémica conocida de Argelia,Egipto, Sudán, Kenia, Arabia Saudita y Yemen.
El ejemplar que ahora citamos posee una morfología
y genitalia que coincide con lo conocido para estaespecie (Hölzel, 1975a).
Material estudiado:
EGIPTO: Luxor, 12.VIII.1988, 1 % en pequeña oquedadde un cortado arenoso en la ribera del río Nilo, V.J. Mon-serrat (VM).
Dielocroce hebraea Hölzel, 1975
Especie sirio-erémica, conocida de la Península delSinaí y que ahora citamos de Jordania y Marruecos,por lo que puede presumirse una amplia distribuciónnorteafricana, aunque es cierto que el nivel de cono-cimiento de las especies de este género es, en ciertomodo, problemático (Hölzel, 1975a) y hace, a veces,subjetiva la identificación de alguna de sus especies.
Los imagos de esta especie, obtenidos a partir de lar-vas capturadas en su medio natural y posteriormentecultivadas en el laboratorio, nos permiten la descrip-ción morfológica de sus larvas y anotar ciertos datossobre su biología que hasta ahora eran desconocidos(Fig. 10).
Hasta su pupación, estas larvas fueron cultivadas enplacas de Petri de 11,5 cm de diámetro y 1,5 cm defondo, rellenas hasta 1 cm de arena de grano muyfino, sobre la que las larvas mantuvieron su actividad,habiendo sido alimentadas con pequeños insectosvivos, principalmente áfidos. Las condiciones decultivo fueron realizadas en el laboratorio en pe-numbra/oscuridad según fotoperiodo natural y atemperaturas máxima y mínima según se anota(en ºC): VII.2003 (31/25), VIII.2003 (34/24), IX.2003(34/21), X.2003 (22/15).
Las larvas mantuvieron una actividad normal, cazan-do las presas y manteniéndose sobre la arena osemienterradas en ella, de la forma habitual como seha citado en otras especies, y no se ha observadoningún caso de canibalismo. Tras su pupación y parafavorecer la correcta emergencia de los imagos, lascámaras pupales eran trasladadas a tubos de 10 cmde alto y 2,5 cm de diámetro, manteniéndose ente-rradas bajo 1-2 cm de arena sobre la que se habíandispuesto algunas pequeñas ramitas para favorecer latrepa de los imagos recién emergidos.
Las larvas maduras poseen una coloración tegumen-taria, especialmente cefálica, algo variable (Figs. 10a-d).En las condiciones citadas y por los datos obtenidosen la cría de sus larvas, una vez realizada la pupación(Fig. 10f) parece que requieren 31, 37, 38 y 38 días
Heteropterus Rev. Entomol. (2008) 8(1): 1-33 17

en su fase de pupa antes de emerger como imagos(Figs. 10e y 10k). La cámaras pupales son del tipocomún en Myrmeleontoidea, forradas interiormentecon una brillante seda a la que quedan adheridosexteriormente los granos de arena, formando unaesfera de 4-5 mm de diámetro (Fig. 10e), como fuedescrito por Ghosh (1910), Imms (1911), Hafez yEl-Moursy (1965), Mansell (1976, 1977) y Monserrat(1983a, 1983b) para otras especies y muy diferentes alo que para esta familia expuso Pierre (1952) y que hasido posteriormente reproducido por Tjeder (1967).
El imago emerge por el orificio practicado por lapupa (Figs. 10e y 10k), cuya exuvia (Figs. 10i-j) quedaretenida en este orificio (Fig. 10e), tal como Ghosh(1910), Imms (1911), Hafez y El-Moursy (1965), Man-sell (1976, 1977) y Monserrat (1983a, 1983b) anotanpara otras especies. En el interior de la cámara pupalqueda abandonada la última exuvia larvaria (Figs.10g-h). En esta fase de emergencia se produce unarelativamente alta mortalidad (hasta un 15%). La ex-pulsión del meconio (Fig. 10l) se realiza durante otras la fase de estabilización tegumentaria del imagoy éstos son de aspecto, grosor y tamaño algo dife-rentes, según los ejemplares.
Material estudiado:
JORDANIA: Petra, 25.VII.2003, 1 % en cueva, V.J. Mon-serrat (VM), s.f., 1 & en tela de araña en cueva, V.J. Mon-serrat (VM), 14.IX.2003, 1 & ex l., capturada el 25.VII.2003en oquedad de rambla, pupó el 8.VIII.2003, V.J. Monserrat(VM), 17.IX.2003, 1 % ex l., capturada el 25.VII.2003 enoquedad de rambla, pupó el 10.VIII.2003, V.J. Monserrat(VM), 18.IX.2003, 1 % ex l., capturada el 25.VII.2003 enoquedad de rambla, pupó el 11.VIII.2003, V.J. Monserrat(VM), 2.X.2003, 1 & ex l., capturada el 25.VII.2003 en oque-dad de rambla, V.J. Monserrat (VM). Wadi Rum, 26.VII.2003,varias cámaras de pupación abandonadas en oquedadde roca, V.J. Monserrat (VM). MARRUECOS: Tirhboula,5.VII.1986, 1 % ex l., capturada en hueco de rambla el25.V.1987, pupó el 4.VI.1987, V.J. Monserrat (VM).
Pterocroce capillaris (Klug, 1836)
Especie de amplia distribución erémica, conocida dellímite meridional de la Región Paleártica occidental ycitada desde el sureste de la Península Ibérica, Ma-rruecos, Argelia, Túnez, Chad, Egipto y Chipre aArabia Saudita e Irán.
En relación con esta especie, quizás es el momentode aclarar definitivamente una cuestión taxonómicaque está aún sin resolver desde que Roux (1833)«describió» una larva (aportando una figura) de,
según él, un «insecto áptero», al que denominó Ne-
crophylus arenarius Roux, 1833, basándose en materialrecolectado en las cuevas y tumbas de las proxi-midades de las pirámides de Giseh, en El Cairo(Egipto), y que resultó ser la primera descripción deuna larva de Crocidae. Comentamos la historia deesta especie.
Mientras se iba poniendo en solfa la sistemática y lataxonomía de esta familia (Westwood, 1841, 1842),esta larva fue reproducida, si bien algo más tarde,en diversas obras por Sharp (1895) y otros (ver El-tringham, 1923). Schaum (1857) cita numerosos ejem-plares de esta especie recolectados en las tumbas deBeni-Hassan, sur de El Cairo (Egipto), y aporta unafigura que es posteriormente reproducida por Navás(1910, 1912). Otras referencias o figuras de esta es-pecie (Necrophylus arenarius Roux, 1833) fueron dadaspor Hagen (1886), Imms (1911), Navás (1924) yTröger (1993).
Por otra parte, larvas muy similares son posterior-mente descritas por Withycombe (1923a, 1923b) yEltringhan (1923) sobre la base de material recolec-tado por Williams, Buxton y Nevison en 1922 encuevas del Wadi Digla, sureste de El Cairo (Egipto),y Withycombe (1923a, 1923b, 1925), basándose enestas larvas y también en imagos, describe Pterocroce
storeyi Withycombe, 1923, que reproduce Wheeler(1929) y una fotografía de cuya larva recogen Navás(1924) y Saz (1925). Esta especie fue consideradasinónima de P. capillaris por Hölzel (1975a). Igual-mente Pierre (1952) describe otra larva muy similarque asigna a una nueva especie, Pterocroce troglophilus
Pierre, 1952, de Béni Abbès (Sahara noroccidental)y la asocia a las larvas anteriormente citadas porWithycombe (1923a, 1923b) y Eltringhan (1923). Encualquier caso, también esta especie fue consideradasinónima de la que tratamos por Hölzel (1975a),siendo pues Pterocroce capillaris la única especie cono-cida de este género monotípico. Al margen de todaesta información sobre los estadios preimaginales deesta especie (descritos bien como pertenecientes a Pte-
rocroce sp., P. storeyi, P. troglophilus o P. capillaris), otrosartículos dan nuevos datos sobre su huevo, larva, oreproducen los ya conocidos en la bibliografía (Pau-lian, 1950; Abonnec y Rioux, 1961; Tjeder, 1967;Hölzel, 1975a; Monserrat, 1983b, 1985; Hölzel, 1999)y datos sobre su biología (como Pterocroce arenaria)son aportados por Hafez y El-Moursy (1959). De sularva, aportamos ahora una imagen del aspecto ge-neral (Fig. 9i).
Expuesta la situación de conocimiento actual, debe-mos indicar que Necrophylus arenarius Roux, 1833 fue
MONSERRAT: Nuevos datos sobre Nemopteridae y Crocidae (Neuroptera)18

inicialmente asignada de forma subjetiva (pie de notade Férussac en: Roux, 1833) a ciertos neurópteros deprotórax largo (Mantispidae o Raphidiidae) y fueronWestwood (1840), Schaum (1857) y Hagen (1866,1886) los primeros que sugirieron la asociación de
este insecto con una larva de neuróptero nemoptérido.Más tarde, Hagen (1886) la asoció con Nemoptera oli-
vieri (Rambur, 1842) y Navás (1910, 1912) la asocióprovisionalmente con Stenorrhachus costatus (Klug,1836), ambas sinónimas de Savigniella costata (Klug,
Heteropterus Rev. Entomol. (2008) 8(1): 1-33 19
FIGURA 9. (a)-(d) Aspectos del huevo de Dielocroce ephemera: (a) Aspecto general; (b)-(d) Aspectos del corion y sus aeropilos;(e) Micropilo; (f) Larva neonata de Croce schmidti (omitidas las patas), vista dorsal (fotografía de A. Satar); (g) Ala anterior deJosandreva sazi, ejemplar de Tabernas, Almería (España); (h) Ídem, ejemplar asignado inicialmente a esta especie, de Azilal, Ait-Bouguemez Douar Rit-Zizj (Marruecos); (i) Aspecto de la larva madura de Necrophylus arenarius, vista dorsal, ejemplar de RamblaCarboneras, Almería (España); (j)-(l) Aspectos de la larva, probablemente madura, de Laurhervasia setacea.
FIGURE 9. (a)-(d) Aspects of the egg of Dielocroce ephemera: (a) General aspect; (b)-(d) Aspects of the chorion and its aeropyles;(e) Micropyle; (f) Newborn larva of Croce schmidti (legs left out), dorsal view (photograph by A. Satar); (g) Front wing of Josandrevasazi, specimen from Tabernas, Almeria (Spain); (h) Ditto, specimen assigned initially to this species, from Azilal, Ait-BouguemezDouar Rit-Zizj (Morocco); (i) Aspect of the mature larva of Necrophylus arenarius, dorsal view, specimen from Rambla Carboneras,Almeria (Spain); (j)-(l) Aspects of the larva, probably mature, of Laurhervasia setacea.
(a)
(d)
(g)
(j)
(b)
(e)
(h)
(k)
(c)
(f)
(i)
(l)
mm mm

1836), y ambos hechos son descartables pues se tratade Insecta: Nemopteridae nec Crocidae. Aún mástarde, Wheeler (1929) la asocia con Pterocroce storeyi yCowley (1941) mantiene una postura algo más escép-tica ante el desconocimiento que entonces se teníasobre las larvas de Crocidae. Tjeder (1967) recopilaparte de esta información y asocia esta larva a Ptero-
croce o a Dielocroce y ahora el segundo supuesto debeser rechazado con base en la longitud del protórax deuno y otro géneros (Figs. 9i, 11l-m). También Aspocket al. (2001) indican que la asociación entre Necro-
phylus y Pterocroce no puede realizarse con seguridad alno considerar sus larvas idénticas y que su asociacióncon otras especies egipcias de Dielocroce no puederealizarse con seguridad. El primer hecho es com-prensible dados el momento y circunstancias en elque la larva fue descrita por Roux (1833) (exagerandola longitud del pronoto como elemento más llama-tivo) y el segundo hecho ahora sí lo consideramosfactible por lo anteriormente citado.
La asociación entre Necrophylus arenarius y Pterocroce
capillaris ha sido anotada por Navás (1926), Wheeler(1929), Cowley (1941), Hafez y El-Moursy (1959),Monserrat (1983b) y Tröger (1993), quienes más omenos abiertamente las sugieren como sinónimas,y bajo Pterocroce arenaria (Roux, 1833) ha sido citada(Navás, 1926; Hafez y El-Moursy, 1959). El caso esque la especie ha venido siendo considerada comonomen dubium/ nomen nudum (Aspöck et al., 2001; Os-wald, 2003).
Desde el punto de vista nomenclatural debe indicar-se que el nombre genérico Necrophylus, así citado enel texto de la descripción y como Necrophilus en el piede la figura (Roux, 1833), ha sido citado como preo-cupado por Necrophilus Latreille, 1829 (Coleoptera:Polyphaga: Agyrtidae). Basándose en esta supuestahomonimia, Navás (1926) relega a homonimia elgénero Necrophilus (no menciona en ningún momentoNecrophylus) y genera la nueva combinación Pterocroce
arenaria (Roux, 1833), al considerarla sinónima dePterocroce storeyi Withycombe, 1923. La primera utili-zación original de este género (texto de Roux, 1833)fue como Necrophylus y la actuación de Navás (1926)no puede considerarse, ya que no cita ambos nom-bres genéricos, ni cita ambas posibles combinaciones,ni elige la preferencia de una sobre otra (Artículo 24bdel CINZ). La correcta designación de Necrophylus
(y no Necrophilus) fue fijada por Wheeler (1929: 313)actuando como primer revisor y así lo recogen Os-wald y Penny (1991). Esta presunta homonimia yafue cuestionada por Cowley (1941) y queda descar-tada por las normas del Código Internacional de
Nomenclatura Zoológica. Por ello, debe citarse yconsiderarse como Necrophylus.
Desconocemos si existía un solo tipo o varios sintiposde Necrophylus arenarius Roux, 1833, aunque quizássea sólo uno (habla de un único ejemplar: «he encon-
trado un pequeño animal.... Lo he encontrado corriendo sobre
las arenas que cubren el interior de las tumbas excavadas en
las rocas alrededor de la Pirámides de Giza»), que podríahaber sido depositado en el Muséum d’HistoireNaturelle de Marsella, donde Roux era conservador.No lo/s hemos encontrado en el citado museo; yaCowley (1941) dudaba de su existencia.
Ante ello, designamos como neotipo de Necrophylus
arenarius Roux, 1833 un ejemplar macho recolectadoen la localidad tipo y en los medios originalmentecitados (cuevas arenosas en los alrededores de las pirá-mides de Giza): EGIPTO: El Cairo, Giza, 28.VII.1989,1 % ex l., obtenido de una larva, similar a la descritapor Roux (1833), capturada en oquedades y cuevas desuelo arenoso el 14.VIII.1988, que, criada en el labo-ratorio, pupó el 10.VII.1989, emergiendo el imagoen la fecha citada, V.J. Monserrat (VM).
Por la morfología y la pigmentación tegumentariaanotada en la descripción original (Roux, 1833) haypocas dudas en asociar este ejemplar con Pterocroce
capillaris. Además hemos estudiado material original delas larvas asignadas por Withycombe (1923a, 1923b)y Eltringhan (1923) a Pterocroce storeyi Withycombe,1923, así como nuevo material recolectado en lalocalidad tipo y alrededores (Egipto, El Cairo: Giza ySaqqara, ver material estudiado) y todos estos ejem-plares pertenecen a lo que conocemos como Ptero-
croce capillaris. Por otra parte, hemos citado Pterocroce
Withycombe, 1923 como un género monotípico conuna única especie válida, Pterocroce capillaris, de ampliadistribución geográfica, por lo que no hay opciónni duda en la relación entre las especies de ambosgéneros.
Ante todo esto, proponemos formalmente las si-guientes sinonimias, ya sugeridas por Wheeler (1929),Cowley (1941), Monserrat (1983b) y Tröger (1993).
Necrophylus Roux, 1833: 76
= Pterocroce Withycombe, 1923a: 141 (redefinido enWithycombe, 1923b: 276) n. sin.
Necrophylus arenarius Roux, 1833: 76
= Pterocroce capillaris (Klug, 1836) (descrita comoNemoptera capillaris Klug, 1836: 96) n. sin.
= Pterocroce storeyi Withycombe, 1923= Pterocroce troglophilus Pierre, 1952
MONSERRAT: Nuevos datos sobre Nemopteridae y Crocidae (Neuroptera)20

Heteropterus Rev. Entomol. (2008) 8(1): 1-33 21
FIGURA 10. Estadios preimaginales de Dielocroce hebraea: (a)-(d) Variabilidad en la pigmentación dorsal de la cápsula cefálica en lalarva madura; (e) Cámaras de pupación con exuvias pupales asociadas; (f) Pupa; (g)-(h) Exuvias de la larva madura; (i)-(j) Exuviaspupales; (k) Imago emergiendo de la cámara de pupación; (l) Meconio.
FIGURE 10. Preimaginal stages of Dielocroce hebraea: (a)-(d) Variability of dorsal pigmentation of the cephalic capsule in the maturelarva; (e) Pupal cocoon with associated pupal exuviae; (f) Pupa; (g)-(h) Exuviae of the mature larva; (i)-(j) Pupal exuviae; (k) Imagoemerging from the pupal cocoon; (l) Meconium.
(a)
(d)
(g)
(j)
(b)
(e)
(h)
(k)
(c)
(f)
(i)
(l)

La morfología de los estadios larvarios de esta espe-cie (Figs. 9i, 11l y 11m-B y C), así como su biología,son suficientemente conocidas y estos datos puedenrecabarse en la bibliografía anotada. No obstante, ycomo hemos citado en algunas especies de Lertha delas que disponíamos de abundante material, hemosrealizado algunos experimentos con numerosas lar-vas (11) recolectadas en cuevas y cavidades de paredesde ramblas en ESPAÑA: La Serena, Bédar, 28.IV.2007,a las que hemos mantenido en el laboratorio hastasu pupación. Estas larvas fueron cultivadas en dosplacas de Petri de 11,5 cm de diámetro y 1,5 cm defondo, rellenas hasta 1 cm de arena de grano muyfino, sobre la que las larvas mantuvieron su actividad,habiendo sido alimentadas con pequeños insectosvivos, principalmente áfidos y hormigas. Las condi-ciones de cultivo corresponden a las de laboratorio enpenumbra/oscuridad según fotoperiodo natural, ahumedad relativa ambiente y a temperaturas máximay mínima según se anota (en ºC): IV.2007 (23/16),V.2007 (26/19), VI.2007 (19/39), VII.2007 (25/41).
Estas larvas mantuvieron una actividad normal,cazando las presas y manteniéndose sobre la arena osemienterradas en ella, de la forma habitual como seha citado en otras especies, y no se ha observadoningún caso de canibalismo. Tras haber completadosu desarrollo, los ejemplares iniciaron su pupacióncon casi un mes de diferencia entre unos y otros(entre 26.VI-21.VII.2007), hecho que repercutiríaen el tiempo en el que se mantiene activa la pobla-ción de imagos. Practicaron la característica cámarade pupación (como Fig. 10e), que quedaba enterrada osemienterrada y casi siempre visible desde el exterior.Estas cámaras pupales fueron trasladadas individual-mente a viales de cristal de 10 cm de alto y 2,5 cm dediámetro, siendo experimentalmente enterradas bajoarena fina a profundidades de 1, 5, 2, 2, 5 y 3 cm,colocando sobre la superficie briznas, ramitas o tirasde papel para favorecer la trepa y posterior emer-gencia del imago. Ninguno de estos frágiles imagosconsiguió superar el peso de la arena situada sobreellos a las profundidades citadas, recuperándose mástarde los imagos muertos durante el intento de libe-rarse de la exuvia pupal (como Fig. 10k). Esto hacesuponer que la mortalidad en esta fase (también ano-tada anteriormente para Dielocroce hebraea Hölzel, 1975)puede ser debida a factores experimentales (enterra-das a más de 1 cm) y esta alta mortalidad observadapodría no tener tanta repercusión de forma espontá-nea en la naturaleza.
Material estudiado:
EGIPTO: Cairo, Giza, 28.VII.1989, 1 % ex l., capturada el14.VIII.1988, pupó el 10.VII.1989, V.J. Monserrat (VM),neotipo de Necrophylus arenarius Roux, 1833, 10-19.VIII.1994,4 && ex l., capturadas el 1.V.1994, puparon el 3.VII.1994,V.J. Monserrat (VM), Cairo, Wadi Digla, XII.1922, 1 larva,P.A. Buxton (NHM), 2 larvas, B.G. Nevinson (NHM),VIII.1922, 1 %, 1 & bread from larva, C.B. Williams (NHM).Saqqara, 14.VIII.1988, 1 larva en cavidad de ruina, V.J. Mon-serrat (VM), 1.V.1994, 3 larvas en cueva, V.J. Monserrat(VM). ESPAÑA: Almería, Alcandique, 2.VI.1986, 4 larvasen cueva de rambla, V.J. Monserrat (VM), Cuevas de Al-manzora, 16.VII.2007, 8 %%, 5 && en cueva antropizada,V.J. Monserrat (VM), 4 %%, 1 & en cueva antropizada, V.J.Monserrat (TAMU), La Serena, Bédar, 28.IV.2007, nume-rosas larvas (11) en cuevas y cavidades de paredes de ram-blas, V.J. Monserrat (VM), Níjar, 8.VII.1996, 1 larva encueva antropizada, V.J. Monserrat (VM), Rambla Carbone-ras, 17.VIII.2001, numerosas larvas en cavidades de pare-des de ramblas, V.J. Monserrat (VM), Rambla de Tabernas,17.VII.1988, 1 %, E. Martínez Ortega (VM), Tabernas,25.VII.1991, 1 % en ruinas de casa, V.J. Monserrat (VM).Murcia, Rambla de la Murta, 16.VIII.1984, 6 && en cuevacon suelo polvoriento, E. Martínez Ortega (VM), Rambladel Puerto de la Cadena, 29.VIII.1984, 1 % en huecos cali-zos de pared de rambla, E. Martínez Ortega (VM), Siscar,16.VIII.1984, 1 & en huecos calizos de pared de rambla,E. Martínez Ortega (VM).
CONSIDERACIONES SOBRE LOS ESTADIOSLARVARIOS EN CROCIDAE
Una vez aportados los datos que se han anotadosobre los estadios larvarios de alguna de las especiesde esta familia, recopilamos toda la informaciónexistente sobre este particular con el fin de confec-cionar una clave de géneros basada en sus estadioslarvarios maduros y, también de acuerdo con ello,agrupar los géneros por su aparente afinidad o seme-janza respecto a sus estadios larvarios y que, obvia-mente, debería corresponder a una mayor afinidaden la morfología de los imagos.
Es cierto que en la mayoría de la abundante infor-mación que poseemos sobre los estadios juveniles deesta familia sólo se describe el aspecto general exter-no, y hay mucha menos información sobre el detallede las estructuras, por lo que, de momento, sólotenemos datos sobre aspectos generales. Tambiénparece que algunos elementos de morfología y colo-
MONSERRAT: Nuevos datos sobre Nemopteridae y Crocidae (Neuroptera)22

ración, y especialmente las proporciones en las larvasde esta familia, pueden cambiar levemente en funcióndel autor que los describe, de las poblaciones y delestadio de desarrollo en el que se encuentre (Withy-combe, 1923b; Mansell, 1976, 1980, 1996; y materialahora descrito), pero la mayoría de los caracteres semantienen dentro de cada género.
Claves basadas en los estadios larvarios de algunosgéneros y especies han sido previamente aportadospor Mansell (1981b) y Miller y Stange (1989) y parala realización de esta clave contamos con la infor-mación existente sobre los estadios larvarios de estafamilia y que, al margen de ciertas referencias biblio-graficas que sugieren tratarse de esta familia recopi-ladas por Tjeder (1967) y Monserrat (1996), es lasiguiente: Croce = Klugina (Ghosh, 1910; Lefroy, 1910;Imms, 1911; Withycombe, 1923b, 1925; Hafez yEl-Moursi, 1965; Tjeder, 1967; Sengonça, 1981),Necrophylus (= Pterocroce Roux, 1833) (Westwood, 1840;Schaum, 1857; Navás, 1910, 1912, 1924; Eltringhan,1923; Withycombe, 1923b; Saz, 1925; Pierre, 1952;Abonnec y Rioux, 1961; Tjeder, 1967; Hölzel, 1975a;Monserrat, 1983b, 1985; Tröger, 1993; Hölzel, 1999),
Josandreva (Monserrat, 1983a), Dielocroce (Withycombe,1923b; Tjeder, 1967; Suludere et al., 2006), Laurher-
vasia (Mansell, 1976, 1980), Tjederia (Mansell, 1977,1981b), Thysanocroce (Mansell, 1980), Concroce (Mansell,1981a), Moranida (Mansell, 1983a; Miller y Stange,1989), Amerocroce (Mansell, 1983a), Veurise (Mansell,1983a), Austrocroce (Mansell, 1983b) y Carnarviana
(Mansell, 1983b).
Como se ha indicado, la larva descrita por Pierre(1952) de Argelia y por él asociada a Berlandus saharae
Pierre, 1952 = Dielocroce berlandi (Navás, 1936) nopuede asignarse al género Dielocroce (Fig. 11m-A), ymás parece pertenecer a una fase no madura delgénero Josandreva (Fig. 11a), que ahora hemos citadocomo nuevo para el norte de África.
CLAVE DE GÉNEROS PARA LARVASMADURAS DE CROCIDAE CONOCIDAS
Se presenta a continuación.
Heteropterus Rev. Entomol. (2008) 8(1): 1-33 23
Clave de géneros para larvas maduras de Crocidae
(1) Pronoto + cuello de menor longitud que el doble de la longitud de la cápsula cefálica (Figs. 11a-g) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2
- Pronoto + cuello de similar o mayor longitud que el doble de la longitud de la cápsula cefálica (Figs. 11h-m) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8
(2) Mandíbulas sin dolichaster/dentículos evidentes (Fig. 11g) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Veurise
- Mandíbulas con dolichaster/dentículos evidentes (Figs. 11a-f) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3
(3) Mandíbulas con dentículos evidentes sólo sobre su margen interno (Figs. 11a-b) . . . . . . . . . . . . . . . . . . 4
- Mandíbulas con dentículos evidentes sobre el margen interno, externo y superficie dorsal (Figs. 11c-f) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5
(4) Cabeza trapecial. Mandíbulas con dolichaster/dentículos del margen interno abundantes, dispuestos sobresus 2/3 basales y sin que destaque ninguno de ellos sobre los demás. Primer segmento del palpo labial subci-líndrico (Figs. 9f y 11b) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Croce
- Cabeza cuadrada. Mandíbulas con dolichaster/dentículos del margen interno escasos, dispuestos sobresu 1/2 basal y destacando uno de mayor tamaño sobre los demás. Primer segmento del palpo labial poliédrico(Fig. 11a) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Josandreva

MONSERRAT: Nuevos datos sobre Nemopteridae y Crocidae (Neuroptera)24
(5) Dolichaster de la cápsula cefálica, pronoto y resto del tegumento muy abundantes y notorios; los dolichas-ter/dentículos de las mandíbulas formando 4 hileras sobre los 2/3 basales sin alcanzar la zona basal (Fig. 11c) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Austrocroce
- Dolichaster de la cápsula cefálica, pronoto y especialmente del resto del tegumento muy escasos y menosnotorios; los dolichaster/dentículos de las mandíbulas formando 2-3 hileras sobre toda la 1/2 basal (Figs. 11d-f) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6
(6) Metatórax y terguitos abdominales surcados por 3 hileras de manchas que los recorren longitudinal-mente (Fig. 11f) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Thysanocroce
- Sin esas 3 hileras longitudinales (Figs. 11d-e) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 7
(7) Región dorsal de la cabeza, tórax y abdomen con numerosas y llamativas manchas pardas (Fig. 11e) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Concroce
- Región dorsal de la cabeza, tórax y abdomen con tenues y escasas manchas pardas (Fig. 11d) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Carnarviana
(8) Pronoto + cuello de similar o algo mayor longitud que el doble de la longitud de la cápsula cefálica (Figs.11h-i) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 9
- Pronoto + cuello sobradamente de mayor longitud que el triple de la longitud de la cápsula cefálica (Figs.11j-m) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10
(9) Pronoto + cuello de similar o algo mayor longitud que el doble de la longitud de la cápsula cefálica.Cápsula cefálica sin manchas oscuras en forma de «V» (Figs. 9l y 11h) . . . . . . . . . . . . . . . . . . . . . Laurhervasia
- Pronoto + cuello de similar o algo menor longitud que el triple de la longitud de la cápsula cefálica.Cápsula cefálica con una o dos manchas oscuras en forma de «V» (Fig. 11i) . . . . . . . . . . . . . . . . . . . . . Tjederia
(10) Mandíbulas recurvadas desde su inicio. Longitud del pronoto + cuello entre 3,2 y 3,8 veces la longitudde la cabeza. Metanoto portando una llamativa mancha parda en «V» (Figs. 11j-k) . . . . . . . . . . . . . . . . . . . . 11
- Mandíbulas rectas en su tercio basal. Longitud del pronoto + cuello entre 3,4 y 4,3 veces la longitud dela cabeza. Metanoto sin una llamativa mancha parda en «V» (Figs. 9i y 11l-m) . . . . . . . . . . . . . . . . . . . . . . . 12
(11) Cabeza más triangular que trapecial. Longitud del pronoto + cuello 3,2 veces la longitud de la cabeza (Fig. 11k) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Moranida
- Cabeza más trapecial que triangular. Longitud del pronoto + cuello 3,8 veces la longitud de la cabeza (Fig. 11j) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Amerocroce
(12) Longitud del pronoto + cuello 3,4 veces la longitud de la cabeza (Fig. 11m-A) . . . . . . . . . . . . Dielocroce
- Longitud del pronoto + cuello 4,3 veces la longitud de la cabeza (Figs. 9i, 11l y 11m-B y C) . . . Necrophylus
FIGURA 11. Aspecto externo de las larvas de Crocidae, vista dorsal: (a) Josandreva (de Monserrat, 1983a); (b) Croce (de Imms, 1911);(c) Austrocroce (de Mansell, 1983b); (d) Carnarviana (de Mansell, 1983b); (e) Concroce (de Mansell, 1981a); (f) Thysanocroce (de Mansell,1980); (g) Veurise (de Mansell, 1983a); (h) Laurhervasia (de Mansell, 1980); (i) Tjederia (de Mansell, 1977); (j) Amerocroce (de Mansell,1983a); (k) Moranida (de Mansell, 1983a); (l) Necrophylus = (Pterocroce) (de Monserrat, 1983b); (m) Cabeza y protórax de Dielocroce (A)y Necrophylus = (Pterocroce) (B y C) (de Withycombe, 1923b).
FIGURE 11. External aspect of the larvae of Crocidae, dorsal view: (a) Josandreva (from Monserrat, 1983a); (b) Croce (from Imms,1911); (c) Austrocroce (from Mansell, 1983b); (d) Carnarviana (from Mansell, 1983b); (e) Concroce (from Mansell, 1981a); (f) Thysa-nocroce (from Mansell, 1980); (g) Veurise (from Mansell, 1983a); (h) Laurhervasia (from Mansell, 1980); (i) Tjederia (from Mansell,1977); (j) Amerocroce (from Mansell, 1983a); (k) Moranida (from Mansell, 1983a); (l) Necrophylus = (Pterocroce) (from Monserrat,1983b); (m) Head and prothorax of Dielocroce (A) and Necrophylus = (Pterocroce) (B and C) (from Withycombe, 1923b).

Heteropterus Rev. Entomol. (2008) 8(1): 1-33 25
(m)
A B C
(a) (b)
(c)(d)
(e)(f)
(g)(h)
(i)
(j) (k) (l)

CONSIDERACIONES SOBRE LA SISTEMÁTICADE CROCIDAE CON BASE EN SUS
LARVAS MADURAS
Es cierto que aún no conocemos la morfología pre-imaginal de varios géneros (Pastranaia, Apocroce, Afga-
nocroce y Anacroce) pero, al menos dentro del orden delos neurópteros, los Crocidae representan la familiamejor estudiada y conocida en este particular(Mansell, 1986) y el nivel de conocimiento sobre susestadios larvarios, con datos de 13 de sus 17 génerosconocidos, es bastante excepcional en este orden ynos permite ciertas garantías para aplicar estos datosa la sistemática general de la familia.
También es cierto, y ya lo hemos mencionado, quese ha citado una cierta variabilidad en la pigmenta-ción (Figs. 10a-d) y proporciones de las larvas segúnla edad y las poblaciones en algunas especies de estafamilia (Withycombe, 1923b; Mansell, 1976, 1980,1996), aunque este último particular debería revisarsecon mayor profundidad, y que su morfología y sucomportamiento están condicionados por el mediodonde se desarrollan (generalmente sobre o semien-terradas en la fina arena y sedimentos en cuevas yoquedades, en especies de protórax largo, y entredetritus vegetales en grietas y sobre polvo y sedi-mentos en cuevas, oquedades y viviendas humanas,en especies de protórax corto) y que este modo devida troglobio ha podido generar convergencias yespecíficas adaptaciones que les ha permitido colo-nizar medios marcadamente xéricos e inhóspitos(Mansell, 1996). Sin embargo, no es menos ciertoque una mayor similitud entre la morfología de losestadios juveniles permitiría contribuir con nuevoscaracteres, ayudando a asociar los géneros dentro dela sistemática general de la familia, donde sus géne-ros se han descrito sobre la base de caracteres mor-fológicos de sus imagos, por otra parte de una granuniformidad morfológica externa y genital (Hölzel,1975a; Mansell, 1986, 1996).
Con base en los caracteres conocidos sobre la mor-fología de las larvas, se segregan dos grandes gruposde géneros. Aquellos con larvas de protórax corto ymandíbulas frecuentemente armadas de dolichasterdenticulados, a veces formando auténticos dentículos(Figs. 11a-g) y las de protórax largo y mandíbulas sindolichaster denticulados (Figs. 11h-m). En el primergrupo podrían asociarse los géneros Josandreva y Croce,con fuertes dentículos/dolichaster en el margen inter-no de las mandíbulas (Figs. 11a-b), Austrocroce, Car-
narviana, Thysanocroce y Concroce, con dentículos/doli-
chaster más tenues, muy abundantes sobre ambosmárgenes de la mandíbula (Figs. 11c-f), y de formaaislada, Veurise, que carece de ellos (Fig. 11g). Dentrodel grupo con géneros de protórax largo, y que pare-ce más evolucionado y mucho más uniforme que elanterior, parecen más próximos entre sí los génerosLaurhervasia y Tjederia, con protórax moderadamentelargo (Figs. 11h-i), frente a los demás, con protóraxextremadamente largo (Figs. 11j-m) y, entre ellos,los géneros Necrophylus y Dielocroce parecen más afinesentre sí (Figs. 11l-m), como ocurre entre Moranida yAmerocroce (Figs. 11j-k).
La sistemática de la familia parte del tratamiento conrango de tribu Crocini Navás (1910, 1912) con gé-nero tipo Croce McLachlan, 1885, que pasó a serconsiderada subfamilia Crocinae por Tillyard (1925),categoría que ha sido comúnmente aceptada desdeentonces y que Monserrat (1996) eleva a la categoríade familia (Crocidae). Sólo la citada tribu Crocini (queOrfila (1955) se asigna pero que ya había sido designa-da por Navás, 1910) y la tribu Pastranaiini Orfila,1955, por el autor creada para su género Pastranaia (yque es su género tipo) y en el que también incluyóa Josandreva Navás, 1906, son las únicas categoríassupragenéricas descritas. La obra de Hölzel (1975a)asocia los géneros en dos grupos (grupo Dielocroce ygrupo Laurhervasia) con base en ciertas similitudesexistentes y, sobre todo, la obra de Mansell (1986)aporta excelente información sobre la relación ysimilitud entre los diferentes géneros, pero no haymás categorías supragenéricas descritas.
No es de sorprender que la vinculación y relaciónentre los diferentes géneros de Crocidae basadas enla morfología y genitalia de los imagos que ofrecenHölzel (1975a) y sobre todo Mansell (1986) tengan,mayoritariamente, su correspondencia con las queacabamos de mencionar basadas en caracteres de suslarvas, y que sea la similitud y proximidad entre susestadios juveniles una herramienta más en la siste-mática del grupo para apoyar y/o discutir la relaciónentre sus diferentes géneros.
Hemos citado la tribu Pastranaiini Orfila, 1955, cre-ada para el género Pastranaia (del que desconocemossu & y sus estadios juveniles) y en la que tambiénse incluyó Josandreva Navás, 1906 (Orfila, 1955).Estamos de acuerdo con Hölzel (1975a) y Mansell(1986) en que la asociación entre los géneros Josan-
dreva y Pastranaia parece poco sostenible y que la pre-sencia de venas Cup y A1 independientes y no fu-sionadas en las alas anteriores presentes en ambosgéneros y que definen la tribu como único carácter
MONSERRAT: Nuevos datos sobre Nemopteridae y Crocidae (Neuroptera)26

(Orfila, 1955) parece tratarse de una elemento ple-siomorfo ancestral en la familia Crocidae y que éstacomparte con Nemopteridae (Mansell, 1986). Por elcontrario, la vinculación entre los imagos de Josan-
dreva y Croce ha sido suficientemente argumentadapor Hölzel (1975a) y particularmente por Mansell(1986) y esta vinculación se apoya en otros elemen-tos que se comentarán más adelante.
Por lo anteriormente indicado, y ya que Croce es elgénero tipo de la citada tribu Crocini (Navás, 1910),parece más adecuado transferir y ubicar el géneroJosandreva en esta tribu Crocini y dejar, de momento,el género Pastranaia como género único dentro de latribu Pastranaiini, que sería la más primitiva y que,por otra parte, habría que redefinir, pues el únicocarácter (plesiomórfico) que la sustentaba carece deinterés sistemático diagnóstico y por ello la hacíainsostenible (Mansell, 1986). Esta tribu podría rede-finirse a partir de la suma de estos caracteres: Alasanteriores grandes (longitud 28-29 mm), con venasA1 y A2 independientes y no fusionadas con Cup,venas apicales y venas marginales de los camposradial, medial y cubital mayoritariamente bifurcadasy ectoprocto en los machos muy largo y digitiforme.Podría aventurarse/asegurarse, con estrecho margende error, que la desconocida larva de Pastranaia debede tener el cuello corto y abundantes dentículos o almenos numerosos dolichaster en el margen internode sus mandíbulas.
A la tribu Pastranaiini, que parece ser la más primitiva,le seguiría, en siguiente grado de mayor evolución, latribu Crocini, por poseer fuertes dentículos/doli-chaster en el margen interno de las mandíbulas y elcuello (protórax) corto (Figs. 11a-g), y donde se ini-cia ya, en la mayoría de sus géneros (con excepciónde Josandreva), la fusión de las venas Cup y A1, ade-más de los caracteres 9-10 y 16-17 de Mansell (1986),que bien comparten o que son exclusivos de algunosgéneros y que avalan su particularidad o la fuertevinculación entre algunos de ellos, como es el casode los géneros Croce y Josandreva, cuya vinculaciónentre sus imagos ha sido suficientemente argumen-tada por Hölzel (1975a) y particularmente porMansell (1986), según se ha citado. Esta vinculaciónqueda reforzada por la afín morfología de sus larvas(Figs. 11a-b), así como por compartir ciertos elementosde su biología (ambas especies citadas de habitacio-nes humanas por: Andreu Rubio, 1909; Lefroy, 1910;Saz, 1925; Monserrat, 1979, 1983a; Monserrat et al.,1999); todo ello avala aún más esta fuerte relación.
Podrían asociarse a esta tribu Crocini, y en orden de
menor a mayor evolución, los géneros Austrocroce,Carnarviana, Thysanocroce y Concroce (Figs. 11c-f), con laprogresiva pérdida de fuertes dentículos/dolichasteren el margen interno de las mandíbulas, manteniendotenues dentículos/dolichaster muy abundantes sobreambos márgenes de la mandíbula y mucho mástenues y muy abundantes dolichaster sobre el tegu-mento cefálico; y por último, y de forma aislada, elaparentemente más evolucionado género Veurise, quelos pierde y prácticamente carece de ellos (Fig. 11g).Salvo la ubicación que ahora proponemos de Thysa-
nocroce dentro de este grupo de géneros, estos datoscorroboran la fuerte asociación entre los génerosAustrocroce, Carnarviana y Concroce como un grupomonofilético dada por Mansell (1986) con base encaracteres de los imagos, así como la posición inde-pendiente y algo aislada de Veurise, que, en principio,mantenemos dentro de esta tribu.
Los demás géneros cuyas larvas son conocidas com-parten caracteres más evolucionados, como haberperdido dentículos/dolichaster en las mandíbulas yposeer mucho más tenues y menos aparentes los doli-chaster sobre el tegumento cefálico, así como poseerel característico y largo protórax (Figs. 11h-m), ytanto por los caracteres de sus imagos, como el queahora citamos de sus larvas, deberían agruparse den-tro de una nueva categoría supragenérica de la quehablaremos a continuación. Estos caracteres apare-cen de forma inicial en los géneros más primitivosLaurhervasia y Tjederia, con protórax moderadamentelargo (Figs. 9j-k y 11h-i), frente a los géneros másevolucionados Dielocroce, Necrophylus, Moranida y Ame-
rocroce, donde aparecen de forma completamentedesarrollada (Figs. 11j-m). Salvo la ubicación de Thysa-
nocroce, que anteriormente hemos ubicado en Crocini,no es de sorprender que estos datos ahora aportadoscoincidan plenamente con lo anotado por Mansell(1986) para los imagos. Corroboran la fuerte asocia-ción entre los géneros Laurhervasia y Tjederia comoun grupo monofilético dada por Mansell (1986) conbase en caracteres de los imagos, y respecto a losdemás géneros y, aunque en alguno de ellos sus larvasno son conocidas (Apocroce, Afganocroce y Anacroce), esde suponer que sean similares a las del otro grangrupo citado de Dielocroce, Necrophylus, Moranida yAmerocroce. Los caracteres citados para sus estadiosjuveniles, así como los caracteres 11-15 y 18-20 deMansell (1986), definen este conjunto de géneros y,aunque se ha citado Dielocroce como el género proba-blemente más evolucionado y el más exitoso dentro deeste grupo (Mansell, 1986), consideramos a Necrophylus
Heteropterus Rev. Entomol. (2008) 8(1): 1-33 27

el más representativo y al que parece corresponderleun mayor grado de evolución en los estadios larvariosde todos estos géneros, géneros que asociamos enuna nueva tribu para la que designamos este géneroNecrophylus como género tipo.
Desde los taxones (tribus-géneros) aparentementemenos evolucionados a los aparentemente más evolu-cionados, la familia quedaría, pues, sistemáticamenteorganizada según se expone:
Familia C r o c i d a e
Tribu Pastranaiini Orfila, 1955 (partim)Género tipo: Pastranaia Orfila, 1955
Pastranaia Orfila, 1955
Tribu Crocini Navás, 1910Género tipo: Croce McLachlan, 1885
Josandreva Navás, 1906
Croce McLachlan, 1885
Austrocroce Tjeder, 1974
Carnarviana Mansell, 1983
Concroce Tjeder, 1967
Thysanocroce Withycombe, 1923
Veurise Navás, 1927
Tribu Necrophylini Monserrat, 2008 trib. nov.Género tipo: Necrophylus (Roux, 1833)
Laurhervasia Navás, 1914
Tjederia Mansell, 1977
Amerocroce Mansell, 1983
Moranida Mansell, 1983
Apocroce Tjeder, 1974 (sensu Tjeder, 1975)
Anacroce Hölzel, 1975
Afganocroce Hölzel, 1968
Dielocroce Cowley, 1941
Necrophylus (Roux, 1833) = (Pterocroce Withy-combe, 1923a, redefinido en Withycombe,1923b) n. sin.
CONSIDERACIONES BIOGEOGRÁFICASSOBRE LA FAMILIA CROCIDAE
La efímera existencia y la escasa capacidad de vuelo ydispersión de los imagos, así como la estrecha vincu-lación y adaptación de las larvas al medio troglobioy a los hábitats xéricos donde prosperan, hacen quelos miembros de esta familia posean una muy escasacapacidad de dispersión.
Por sus enormes vínculos morfológicos es seguroque Nemopteridae y Crocidae tuvieron antecesorescomunes antes de la disgregación de la Pangea y suactual distribución y la presencia de géneros fósilesde Nemopteridae (Marquettia Navás, 1913 y Olivierina
Navás, 1910 = Lertha Navás, 1910 del Oligoceno deNorteamérica) lo sugieren. Por lo demás, ambas fa-milias tienen una distribución actual muy similar (Tje-der, 1967). Con respecto a Crocidae, Mansell (1986)ya explica la distribución y el origen gondwánico deesta familia (presente en los continentes meridiona-les y extendida por las regiones térmicas y áridas delPaleártico occidental y ausente en Norteamérica ycentro y norte de la Región Paleártica), anotando unprimer aislamiento de taxones habitantes del conti-nente australiano (separado de África y Sudaméricaa principios del Jurásico, hace 180 millones de años)y que una posterior fragmentación de África, Suda-mérica y Laurasia (finales del Cretácico, hace 65 mi-llones de años), así como los cambios en la extensióndel Mediterráneo y la barrera húmeda tropical enÁfrica (también en el sureste de Asia) limitaría einfluiría sobre la radiación y distribución del resto desus taxones (Hölzel, 1968, 1975a, 1975b, 1999; Po-pov, 1970; Tjeder, 1974a, 1974b, 1975; Aspöck et al.,1980, 2001; Mansell, 1983a, 1983b, 1986).
En función de los datos ahora expuestos sobre lasistemática de la familia basada en sus larvas, nuevosdatos contribuyen a reforzar esta hipótesis (Mansell,1986).
La primitiva tribu Pastranaiini, y su único géneroPastranaia, ha quedado confinada en Sudamérica,aunque no se descarta la posibilidad de que puedanaparecer nuevos representantes en otros continentesde origen gondwánico, como ocurre en otras familiasde neurópteros (Dilaridae, Hemerobiidae, Chrysopi-dae, Berothidae, etc.).
La tribu Crocini, algo más evolucionada, posee unadistribución típicamente gondwánica con elementosaustralianos (Austrocroce y Carnarviana), sudamerica-nos (Veurise) y sudafricanos (Concroce y Thysanocroce)que han quedado aislados por las citadas barreras
MONSERRAT: Nuevos datos sobre Nemopteridae y Crocidae (Neuroptera)28

continentales y climáticas. Otros elementos de estatribu (Josandreva y Croce), que poseen un parentescoevidente, han alcanzado una mayor dispersión, perosu origen gondwánico también parece obvio y puedeexplicarse adecuadamente. Así, parece evidente queCroce colonizó el Paleártico occidental (OrienteMedio, Oriente Próximo y norte de África) desdeel subcontinente indio, donde es bien conocido, yJosandreva colonizó el Paleártico occidental (Marrue-cos, Argelia y sureste de la Península Ibérica) yRegión Afrotropical asiática (isla de Sokotra, enYemen) a partir de su ancestral dispersión africananorecuatorial (de la que permanece su presencia enChad como testigo). Parece que esta expansión occi-dental de Croce hacia el Paleártico occidental ha debi-do de estar potenciada y ser paralela al progresivoaumento reciente de la xericidad del clima con laretirada de los bosques tropicales y el avance de lassabanas y posterior desertificación de estas zonas apartir del finales del Terciario y especialmente en elCuaternario. Y en el caso de Josandreva, y dada la uni-formidad entre sus especies, el paso al continenteeuropeo ha debido de ser mucho más reciente enesta expansión intercontinental que el contacto entreambos continentes admitido durante el Mesozoico(hace 225-65 millones de años) entre África y Eu-ropa y deberse al puente y contacto intercontinentalacaecido durante el Eoceno (hace 40-50 millones deaños) y con mucha mayor probabilidad durante elque se produjo en el Mioceno (hace aprox. 6 millo-nes de años).
Similar análisis puede hacerse para los taxones de lamás evolucionada tribu Necrophylini trib. nov., queparece ser de origen más reciente, ya que la ausenciade elementos australianos induce a sugerir un origenposterior a la separación de Australia de Áfricay Sudamérica (a principios del Jurásico, hace 180millones de años) y su ausencia en el subcontinenteindio avalan esta hipótesis, permaneciendo, no obs-tante, elementos sudamericanos (Moranida y Amero-
croce) y sudafricanos (Laurhervasia y Tjederia), que pos-teriormente han quedado aislados por las citadasbarreras continentales, climáticas y biológicas. Otroselementos de esta tribu han quedado aislados dentrode los límites de la Región Afrotropical (Apocroce enla isla de Sokotra, de Yemen) y otros (Dielocroce y Ne-
crophylus) mantienen su distribución africana originalcon representantes testigo en Chad, Sudán o Keniay poseen una amplia expansión posterior por elPaleártico occidental árido (norte de África, OrientePróximo y Medio) y en el caso de Necrophylus su
colonización al continente europeo (suroeste deEspaña y Chipre) ha debido de seguir las pautasmencionadas para Josandreva. Por último, el resto delos elementos parecen presentar una dispersión ydiversificación mucho más reciente, ya que han per-dido su representación subsahariana en el continenteafricano o, incluso, su vinculación afrotropical, dedonde sin duda parecen proceder ancestralmente,dispersándose y diversificándose por el sur de laRegión Paleártica occidental según el reciente cam-bio en las condiciones climáticas y aumento de laxericidad anteriormente citadas, siendo más limitaday restringida su dispersión: Anacroce en la Penínsulade Sinaí (Hölzel, 1975b) y Afganocroce en Afganistáne Irán (Hölzel, 1968, 1975a; Meinander, 1980).
Como conclusión, podemos decir que la similitudmorfológica entre las larvas de los diferentes géne-ros que hemos citado coincide con la dispersión y/oaislamiento de sus diversos taxones y avala el origeny patrón de distribución de los diversos géneros quehemos mencionado.
Agradecimiento
Deseamos manifestar nuestro más sincero agradeci-miento a Mª Dolores Martínez del Departamento deZoología y Antropología Física de la Facultad deBiología de la Universidad Complutense de Madridpor la identificación de las hormigas que se hancitado. A Eduardo Ruiz de este Departamento y alServicio de Microscopía Electrónica de esta univer-sidad por su ayuda en la realización y tratamiento delas imágenes, a Jaime Aguilar Ruiz por su ayuda en laconfección de la gráfica y en textos en inglés y aDavid Papenberg por su apoyo en el alemán. Tam-bién a Fernando Rodrigo por cederme alguna foto-grafía, así como a las instituciones y personas quenos han permitido intentar localizar o estudiar elmaterial citado, en especial a Sylvie Pichard del Mu-séum d’Histoire Naturelle de Marseille por su infor-mación y a Ali Satar de la Art and Science Facultyde la Dicle University en Diyarbakir por enviarnosmaterial biológico y fotográfico que nos ha permitidoalgunas de las descripciones aquí anotadas.
Heteropterus Rev. Entomol. (2008) 8(1): 1-33 29

Bibliografía
ABONNENC E, RIOUX JA. 1961. Contribution a l’étu-de des phlébotomes (Diptera-Psychodidae) du Nord-Tchad (Pp.: 30-52). En: Mission Epidemiologique au
Nord-Tchad. Ed. Prohuza. Paris.
ANDREU RUBIO J. 1909. Neurópteros de los alrede-dores de Orihuela. En: Actas y Memorias del Primer
Congreso de Naturalistas Españoles (octubre 1908): 159-162. Zaragoza.
ASPÖCK H, ASPÖCK U, HÖLZEL H. 1980. Die Neu-
ropteren Europas (vol. 1; vol. 2). Goecke & Evers.Krefeld.
ASPÖCK H, HÖLZEL H, ASPÖCK U. 2001. Kommen-tierter Katalog der Neuropterida (Insecta: Raphidiop-tera, Megaloptera, Neuroptera) der Westpaläarktis.Denisia 2: 5-606.
CANDAN S, SULUDERE Z, AÇIKGÖZ F, HASBENLI A.2005. Ultrastructure of the egg chorion of Nemop-
tera sinuata Olivier 1811 (Neuroptera: Nemopteridae)from Turkey. Entomological News 116(1): 1-10.
COWLEY J. 1941. Two new generic names in theNemopteridae (Neuroptera). Annals and Magazine of
Natural History (11) 7: 121-126.
ELTRINGHAM H (con notas adicionales de WILLMER
EN y WILLIAMS CB). 1923. On the larva of Pterocroce
storeyi, With. (Nemopteridae). Transactions of the Ento-
mological Society of London 1923: 263-268.
GATTI MG, FARJU-BRENER AG. 2002. Low densityof ant lion larva (Myrmeleon crudelis) in ant-acacia clea-rings: Hight predation risk or inadequate substrate?Biotropica 34(4): 458-462.
GHOSH CC. 1910. Croce filipennis, Westw. Journal of the
Bombay Natural History Society 20: 530-532.
GULLAN PJ, CRANSTON PS. 1996. The insects: An outline
of entomology. Chapman & Hall. London.
HAFEZ M, EL-MOURSY AA. 1959. Storage excretionin some insect larvae with blocked hind gut. Bulletin
de la Société Royale Entomologique d’Egypte 43: 85-88.
HAFEZ M, EL-MOURSY AA. 1965. On the biology ofKlugina aristata Klug (Neuroptera: Nemopteridae).Bulletin de la Société Royale Entomologique d’Egypte 48:183-191.
HAGEN HA. 1866. Hemerobidarum Synopsis syno-nymica. Stettiner Entomologische Zeitung 27: 369-462.
HAGEN HA. 1886. Monograph of the Hemerobidae.Part I. Nemopteridae. Proceedings of the Boston Society
of Natural History 23: 250-269.
HALVERSAR DD, WHEELER J, WHEELER GC. 1976.
Natural history of the sandhill ant, Formica bradlei
(Hymenoptera: Formicidae). Journal of the Kansas En-
tomological Society 49(2): 280-303.
HANDEL SN, BEATTIE AJ. 1998. Semillas dispersadaspor hormigas. Investigación y Ciencia, Serie Temas 14:36-43.
HANZAWA FM, BEATTIE AJ, CULVER DC. 1988.Demographic analysis of an ant-seed mutualism.American Naturalist 131(1): 1-13.
HÖLLDOBLER B, WILSON EO. 1990. The ants. Har-vard University Press. Harvard.
HÖLZEL H. 1968. Die Neuropteren VorderasiensIII. Nemopteridae. Beiträge zur Naturkundlichen Fors-
chung in Südwestdeutschland 27: 37-47.
HÖLZEL H. 1975a. Revision der Netzflügler-Unterfa-milie Crocinae (Neuroptera: Nemopteridae). Entomo-
logica Germanica 2: 44-97.
HÖLZEL H. 1975b. Eine neue Gattung und Art derCrocinae aus Sinai (Planipennia, Nemopteridae). Ento-
mologische Zeitschrift, Frankfurt am Main 85: 21-23.
HÖLZEL H. 1999. Die Nemopteriden (Fadenhafte)Arabiens. Ein Beitrag zur Kenntnis der Neuropteridader Arabischen Halbinsel (Neuropterida: Neuroptera:Nemopteridae). Stapfia 60: 129-146.
HUGES L, WESTOBY M. 1992. Capitula on stickinsect eggs and elaiosomes on seeds: convergentadaptations for burial by ants. Functional Ecology 6:642-648.
IMMS AD. 1911. Contributions to the knowledge ofthe structure and biology of some Indian insects. I.On the life-history of Croce filipennis, Westw. (OrderNeuroptera, Fam. Hemerobiidae). Transactions of the
Linnean Society of London, Zoology (2) 11: 151-160.
LECLERCQ S, BRAEKMAN JC, KAISIN JC, DALOZE D,DETRAIN C, BISEAU JC DE, PASTEELS JM. 1997.Venom constituents of three species of Crematogaster
ants from Papua New Guinea. Journal of Natural
Products 60: 1143-1147.
LEFROY M. 1910. The Indian nemopterid and itsfood. Journal of the Bombay Natural History Society 19:1005-1007.
LEON B, PICKER MD. 1990. Function of the hind-wings of Palmipenna aeoleoptera Picker (Insecta: Neu-roptera: Nemopteridae). En: Mansell MW, AspöckH (Eds.). Advances in Neuropterology. Proceedings of
the Third International Symposium on Neuropterology: 89-94. Pretoria.
MANSELL MW. 1973. The first record of a larvalnemopterid from southern Africa (Neuroptera: Ne-
MONSERRAT: Nuevos datos sobre Nemopteridae y Crocidae (Neuroptera)30

mopteridae: Nemopterinae). Journal of the Entomolo-
gical Society of Southern Africa 36: 133-137.
MANSELL MW. 1976. The larva of Laurhervasia setacea
(Klug), (Neuroptera: Nemopteridae: Crocinae) fromsouthern Africa. Journal of the Entomological Society of
Southern Africa 39: 153-158.
MANSELL MW. 1977. A new genus and species in theCrocinae (Neuroptera: Nemopteridae) from southernAfrica. Journal of the Entomological Society of Southern
Africa 40: 195-203.
MANSELL MW. 1980. The Crocinae of southernAfrica (Neuroptera: Nemopteridae). 1. The generaLaurhervasia Navás and Thysanocroce Withycombe.Journal of the Entomological Society of Southern Africa 43:341-365.
MANSELL MW. 1981a. The Crocinae of southernAfrica (Neuroptera: Nemopteridae). 2. The genusConcroce Tjeder. Journal of the Entomological Society of
Southern Africa 44: 91-106.
MANSELL MW. 1981b. The Crocinae of southernAfrica (Neuroptera: Nemopteridae). 3. The genusTjederia Mansell, with keys to the southern AfricanCrocinae. Journal of the Entomological Society of Southern
Africa 44: 245-257.
MANSELL MW. 1983a. New Crocinae (Neuroptera:Nemopteridae) from South America, with descrip-tions of larvae. Journal of the Entomological Society of
Southern Africa 46: 115-130.
MANSELL MW. 1983b. A revision of the AustralianCrocinae (Neuroptera: Nemopteridae). Australian
Journal of Zoology 31: 607-627.
MANSELL MW. 1986. Biogeography and phylogeny ofthe Crocinae (Neuroptera: Nemopteridae). En: Gepp J,Aspöck H, Hölzel H (Eds.). Recent Research inNeuropterology. Proceedings of the Second International
Symposium on Neuropterology: 77-85. Graz.
MANSELL MW. 1992. The systematic position of theNemopteridae (Insecta: Neuroptera: Myrmeleonti-dae). En: Canard M, Aspöck H, Mansell MW (Eds.).Current Research in Neuropterology. Proceedings of the
Fourth International Symposium on Neuropterology: 233-241. Toulouse.
MANSELL MW. 1996. Unique morphological and bio-logical attributes: the keys to success in Nemopteri-dae (Insecta: Neuroptera:). En: Canard M, Aspöck H,Mansell MW (Eds.). Pure and Applied Research inNeuropterology. Proceedings of the Fifth International
Symposium on Neuropterology: 171-180. Toulouse.
MATHEWS WH. 1947. Some notes on the spoon-
winged lacewing (Chasmoptera hutti). Western Austra-
lian Naturalist 1: 42-44.
MEINANDER M. 1980. Insects of Saudi Arabia. Neu-roptera: Fam. Nemopteridae. Fauna of Saudi Arabia 2:174-178.
MILLER RB, STANGE LA. 1989. A new species ofMoranida Mansell from Venezuela (Neuroptera: Ne-mopteridae). Insecta Mundi 3: 65-70.
MONSERRAT VJ. 1979. Sobre los neurópteros ibé-ricos (II); (Neuroptera, Planipennia). Boletín de la
Asociacion Española de Entomología 3: 17-21.
MONSERRAT VJ. 1983a. Estadios larvarios de losneurópteros ibéricos I: Josandreva sazi (Neur. Plan.,Nemopteridae). Speleon 26/27: 39-51.
MONSERRAT VJ. 1983b. Pterocroce capillaris (Klug, 1836)en Europa (Neur., Plan., Nemopteridae). Neuroptera
International 2: 109-128.
MONSERRAT VJ. 1985. Morfología del huevo en losnemoptéridos ibéricos (Neur., Plan.: Nemopteridae).Boletim da Sociedade Portuguesa de Entomologia, Suple-mento 1, 2: 463-474.
MONSERRAT VJ. 1988. Revisión de las especies deLertha del Mediterraneo occidental (Neuropteroidea,Planipennia: Nemopteridae). Annali del Museo Civico
di Storia Naturale Giacomo Doria 87: 85-113.
MONSERRAT VJ. 1996. Larval stages of EuropeanNemopteridae, with systematic consideration on thefamily Nemopteridae (Insecta, Neuroptera). Deutsche
Entomologische Zeitschrift 43(1): 99-121.
MONSERRAT VJ, MARTÍNEZ MD. 1995. On the possi-ble myrmecophily of Nemopterinae larvae (Neurop-tera, Nemopteridae). Sociobiology 26: 55-68.
MONSERRAT VJ, GAMARRA P, DERETSKY Z. 1999.The neuropterological fauna of the houses of thecity of Madrid. Journal of Neuropterology 2: 35-44.
NAVÁS L. 1910. Monografía de los nemoptéridos(insectos neurópteros). Memorias de la Real Academia
de Ciencias y Artes de Barcelona (3) 8: 341-408.
NAVÁS L. 1912. Family Nemopteridae. Neuroptera.Genera Insectorum 136: 1-23.
NAVÁS L. 1924. Comunicaciones. Boletín de la Sociedad
Ibérica de Ciencias Naturales 23(6): 65-67.
NAVÁS L. 1926. Névroptères d’Egypte et de Palestine.3me partie. Bulletin de la Société Royale Entomologique
d’Egypte 10: 192-216.
ORFILA RN. 1955. Un nuevo Nemopteridae (Neu-ropt.) americano con una sinopsis de la familia.Revista de la Sociedad Entomológica Argentina 17: 29-32.
Heteropterus Rev. Entomol. (2008) 8(1): 1-33 31

OSWALD JD. 2003. Index to the Neuropterida species of
the world. Disponible en línea en: http://entowww.ta-mu.edu/research/neuropterida/neur_sp_index/ins_search.html.
OSWALD JD, PENNY ND. 1991. Genus-group namesof the Neuroptera, Megaloptera and Raphidiopteraof the world. Occasional Papers of the California Aca-
demy of Sciences 147: 1-94.
PAULIAN R. 1950. La vie larvaire des insectes. R. Tho-mas. París.
PICKER MD. 1984. A new genus and species ofspoon-wing lacewing (Neuroptera: Nemopteridae)from Namaqualand, South Africa. Journal of the Ento-
mological Society of Southern Africa 47: 259-268.
PICKER MD. 1987. An unusual species of spoon-wing lacewing (Neuroptera: Nemopteridae) fromSouth Africa, with notes on its biology. Systematic
Entomology 12: 239-248.
PICKER MD, LEON B. 1990. Population biology ofPalmipenna aeoleoptera Picker with reference to thepossible semaphore function of the hindwings (In-secta: Neuroptera: Nemopteridae). En: Mansell MW,Aspöck H (Eds.). Advances in Neuropterology. Procee-
dings of the Third International Symposium on Neuropte-
rology: 95-99. Pretoria.
PICKER MD, LEON B, LONDT JGH. 1991. The hyper-trophied hindwings of Palmipenna aeoleoptera Picker,1987 (Neuroptera, Nemopteridae) reduce attack byrobber flies by increasing apparent body size. Animal
Behaviour 42: 821-825.
PICKER M, LEON B, LONDT JGH. 1992. Influence ofhindwing size in nemopterids (Insecta: Neuroptera:Nemopteridae) on predation by robber flies (Dip-tera: Asilidae). En: Canard M, Aspöck H, MansellMW (Eds.). Current Research in Neuropterology.Proceedings of the Fourth International Symposium on
Neuropterology: 313-318. Toulouse.
PIERRE F. 1952. Morphologie, milieu biologique etcomportement de trois Crocini nouveaux du SaharaNord-Occidental (Planipennes Nemopteridae).Annales de la Société Entomologique de France 119: 1-22.
POPOV A. 1963. An interesting neuropterous insectin Bulgaria, Nemoptera sinuata Olivier. Priroda [Na-
ture], Sofía 12(3): 90-93. (en ruso)
POPOV A. 1970. Verbreitung der europaischen Ne-mopteriden-Arten (Neuroptera). Izvestiya na Zoologiches-
kiya Institut s Muzei (Bulgarska Akademiya na Naukite,
Sofia) 32: 5-31.
POPOV A. 1973. Über die präimaginalen Stadien pa-läarktischer Vertreter der Ordnung Neuroptera und
Versuch einer neuen systematischen Gruppierung derFamilien mit Rücksicht auf ihre morphologischenund ökologischen Besonderheiten. Bulletin de l’Institut
de Zoologie et Musée, Academie Bulgare des Sciences, Section
de Biology 37: 79-101.
POPOV A. 1998. Bionomy and development of Ne-
moptera sinuata Olivier (Neuroptera, Nemopteridae).Acta Zoologica Fennica 209: 215-216.
POPOV A. 2002. Autecology and biology of Nemop-
tera sinuata Olivier (Neuroptera: Nemopteridae). Acta
Zoologica Academiae Scientiarum Hungaricae 48(Suppl. 2):293-299.
ROUX P. 1833. Lettre relative à divers coquilles, crus-tacés, insectes, reptiles et oiseaux, observés en Égyp-te; adressée par M. Roux à M. le baron de Férussac.Annales des Sciences Naturelles, Zoologie (1) 28: 72-78.
SATAR A, ÖZBAY C. 2004a. Eggs, first instar larvaeand distribution of the neuropterids Lertha extensa
and L. shappardi (Neuroptera: Nemopteridae) in south-eastern Turkey. Zoology in the Middle East 32: 5-11.
SATAR A, ÖZBAY C. 2004b. Remarks on Neuropteraof southeastern Turkey. Entomologica Fennica 15: 1-6.
SAZ E. 1925. La Josandreva sazi Nav. Ibérica 562-563:67-70.
SCHAUM H. 1857. Necrophylus arenarius Roux, diemuthmassliche Larve von Nemoptera. Berliner Ento-
mologische Zeitschrift 1: 1-9.
SENGONÇA C. 1981. Taxonomische Untersuchungenuber de Nemopteridenfauna (Insecta: Neuroptera)der Turkei. 1. Allgemeine Grundrisse der Familie.Turkiye Bitki Koruma Dergisi 5: 91-99.
SHARP D. 1895. Neuroptera Planipennia (Pp.: 444-472). En: Harmer SF, Shipley AE (Eds.). The Cam-
bridge Natural History. Vol. 5. MacMillian and Co.London.
SULUDERE Z, SATAR A, CANDAN S, CANBULAT S.2006. Morphology and surface structures of eggsand first instar larvae of Dielocroce baudii (Neurop-tera: Nemopteridae) from Turkey. Entomological News
117(5): 521-530.
TEDDERS WL, REILLY CC, WOOD BW, MORRISON RK,LOFGREN CS. 1990. Behavior of Solenopsis invicta
(Hymenoptera: Formicidae) in Pecan orchards (Chry-
soperla rufilabris). Environmental Entomology 9: 44-53.
TILLYARD RJ. 1925. A new species of spoon-wingedlacewing (family Nemopteridae, order NeuropteraPlanipennia) from western Australia. Journal [and Pro-
ceedings] of the Royal Society of Western Australia 12: 1-4.
TJEDER B. 1967. Neuroptera-Planipennia. The lace-
MONSERRAT: Nuevos datos sobre Nemopteridae y Crocidae (Neuroptera)32

wings of Southern Africa. 6. Family Nemopteridae(Pp.: 290-501). En: Hanström B, Brinck P, Rudebec G(Eds.). South African Animal Life, Vol. 13. SwedishNatural Science Research Council. Stockholm.
TJEDER B. 1974a. Nemopteridae from the Island ofSocotra (Neuroptera) with descriptions of two genera.Entomologica Scandinavica 5: 291-299.
TJEDER B. 1974b. Taxonomic notes on species of theCrocini (Neuroptera, Nemopteridae). Entomologica
Scandinavica 5: 300-304.
TJEDER B. 1975. A new species name in the Cro-cinae (Neuroptera: Nemopteridae). Entomologica
Scandinavica 6: 133.
TRÖGER EJ. 1993. Die Larve von Nemoptera coa (Lin-naeus, 1758) (Neuropteroidea, Planipennia). Deutsche
Entomologische Zeitschrift, Berlin (N.F.) 40: 357-368.
WESTWOOD JO. 1840. Families Myrmeleonidae, He-merobiidae, Sialidae, Raphidiidae and Mantispidae(Pp.: 41-59). En: An introduction to the modern classifica-
tion of insects. Vol. 2. London.
WESTWOOD JO. 1841. A monograph on the genusNematoptera. Proceedings of the Zoological Society of
London 9: 9-14.
WESTWOOD JO. 1842. A monograph on the genus
Nematoptera. Annals and Magazine of Natural History 8:376-380.
WHEELER WM. 1929. Is Necrophylus arenarius Rouxthe larva of Pterocroce storeyi Withycombe? Psyche 36:313-320.
WITHYCOMBE CL. 1923a. A new Egyptian Nemop-terid (Neuroptera). The Entomologist 56(721): 141.
WITHYCOMBE CL. 1923b. Systematic notes on theCrocini (Nemopteridae), with descriptions of newgenera and species. Transactions of the Entomological
Society of London 1923: 269-287.
WITHYCOMBE CL. 1925. Some aspects of the biolo-gy and morphology of the Neuroptera. With specialreference to the immature stages and their possiblephylogenetic significance. Transactions of the Entomo-
logical Society of London 1924: 303-411.
Heteropterus Rev. Entomol. (2008) 8(1): 1-33 33
Recibido / Hartua / Received: 17/07/2007
Aceptado / Onartua / Accepted: 18/02/2008
Publicado / Argitaratua / Published: 21/06/2008