
La barrera hematoencefálica en patologías del sistema nervioso … · La barrera...
44
La barrera hematoencefálica en patologías del sistema nervioso central: ¿una barrera impenetrable? Ignacio (Nacho) A. Romero Department of Life Sciences The Open University Milton Keynes, UK
Transcript of La barrera hematoencefálica en patologías del sistema nervioso … · La barrera...

La barrera hematoencefálica en patologías del sistema nervioso
central: ¿una barrera impenetrable?
Ignacio (Nacho) A. RomeroDepartment of Life Sciences
The Open UniversityMilton Keynes, UK

Outline of the talkThe blood-brain barrier (BBB) in health:
•
Overview of BBB structure and function
•
Establishment of an in vitro human BBB model
•
Role of the BBB in CNS availability of drugs
The blood-brain barrier (BBB) in disease:
•
Understanding the molecular mechanisms of BBB dysfunction in CNS pathologies

The Blood-brain barrier (BBB) in health:
Structure and function
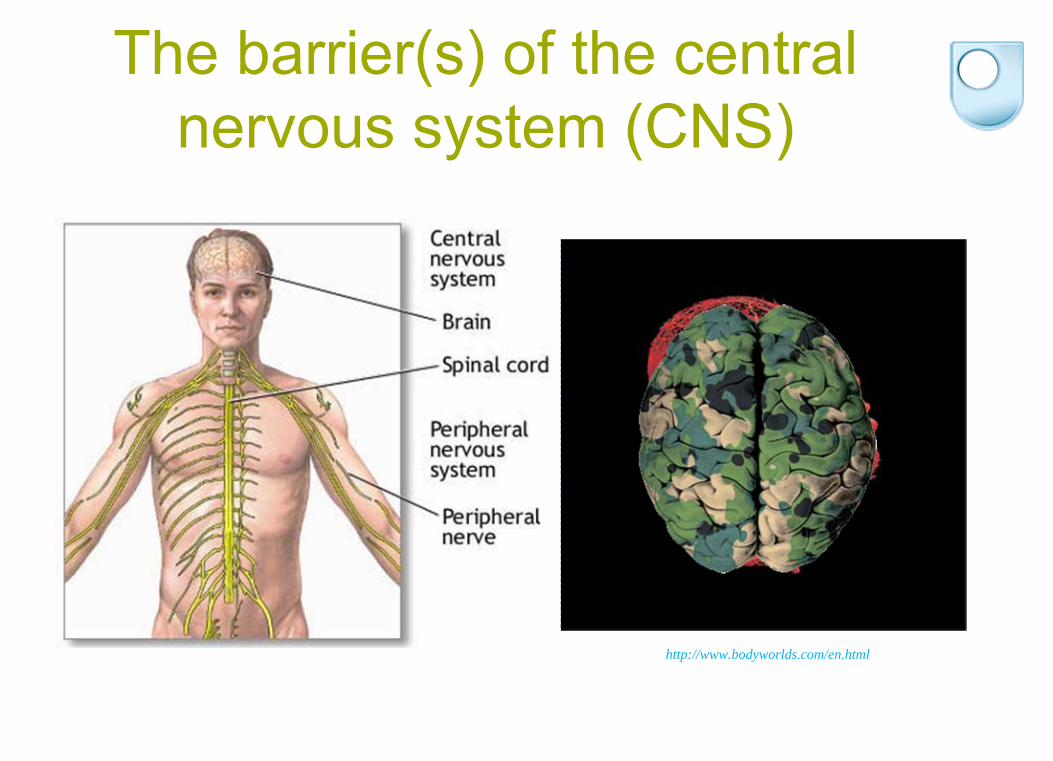
The barrier(s) of the central nervous system (CNS)
http://www.bodyworlds.com/en.html

SITES OF CNS BARRIERS
1.
Brain parenchyma capillary endothelium
2.
Arachnoid epithelium
3.
Choroid plexus epithelium1
2
3Exceptions:Circumventricular
organs (CVO)
Abbott NJ et al.
(2006) Astrocyte–endothelial interactions at the blood–brain barrier
Nat. Rev. Neuro.
7:
41–53 doi:10.1038/nrn1824

1. Control molecular traffic → supply nutrients, remove waste products & keep out toxins
2. Ion homeostasis → optimal neural signalling
3. Low protein environment → limit proliferation
4. Separate CNS and peripheral neurotransmitter pools → allows non-synaptic signalling
5. Immune surveillance with minimal inflammation or cell damage
FUNCTIONS OF CNS BARRIERS
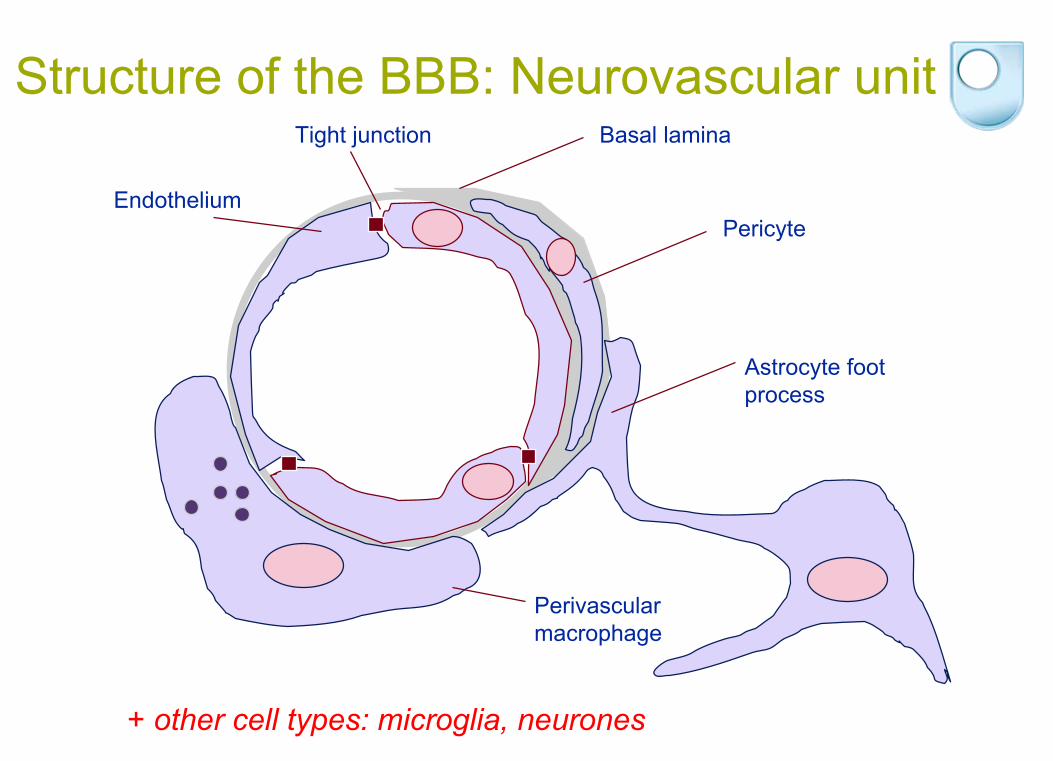
Structure of the BBB: Neurovascular unit
+ other cell types: microglia, neurones
Basal laminaTight junction
EndotheliumPericyte
Astrocyte foot process
Perivascular macrophage
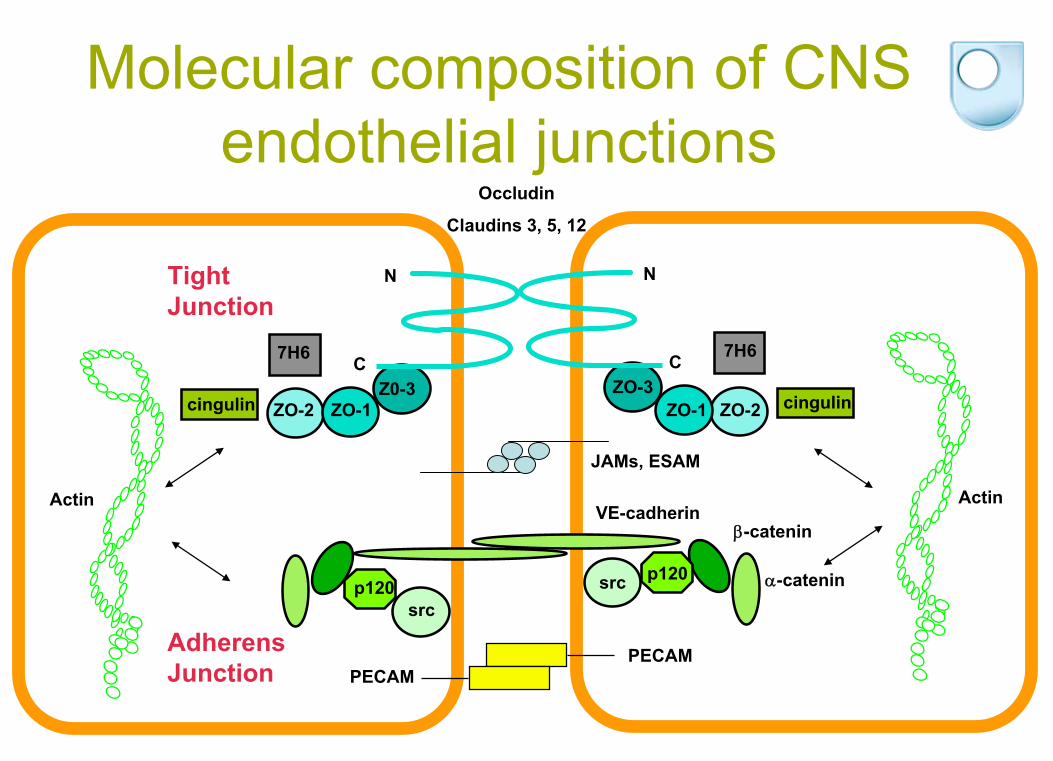
Molecular composition of CNS endothelial junctions
ZO-1ZO-2Z0-3
7H6 C
N
cingulin
srcp120
Adherens Junction
-catenin
-catenin
Actin
ZO-1 ZO-2ZO-3
7H6C
N
cingulin
src p120
VE-cadherinActin
Occludin
Claudins 3, 5, 12
PECAMPECAM
JAMs, ESAM
Tight Junction

The BBB in health
• Physical Barrier
• Transport Barrier
• Metabolic Barrier

The Blood-brain barrier (BBB) in health:
In vitro human BBB model

Development of an in vitro human blood- brain barrier model
Brain endothelial cellsAstrocytes
Primary cultures
Didier et al., Cytokines and the BBB, J. Neurochem, 2003
Maru et al., Chemokines and glial tumours, JNI, 2008
Subileau et al., Chemokines and the BBB, JNEN, 2009
Tai et al., P-glycoprotein and the BBB, Brain Research, 2009
Endothelial cells
Astrocytes
• Individuals with intractable epilepsy →
temporal lobectomy(King’s College Hospital)
• Individuals with multiple sclerosis →
post-mortem cortical grey matter (UK MS Tissue Bank)

Immortalization of primary human adult brain microvascular endothelial cells
Lentiviral hTERT
supernatants
Primary human brain endothelium
~ 3 weeks post isolation
Lentiviral SV40 large T supernatants
5 days
5 days
Isolation of clones by limited dilution
~ 3-4 weeks post infection
~100 clones, one of which was selected and termed hCMEC/D3
Weksler et al., FASEB J, 2005
US Patent no. WO0066149
hCMEC/D3 brain endothelial cell line
Distributed to >120 research groups> 30 research articlesP.O. Couraud, ICGM, Paris B.B. Weksler, Cornell Uni, NY

PECAM-1 catenin
Expression of tight and adherens junction-associated
endothelial markers by hCMEC/D3 cells
-cateninVE-cadherin F-actin ZO1
Claudin-5 JAM-1
Adherens Junctions
Tight Junctions

Permeability to paracellular tracers: Comparison between hCMEC/D3 cells and other in vitro BBB models
Permeability Coefficient (x10-3
cm/min)
sucrose inulin FITC-dextran 4 kDa FITC-dextran 70 kDa Refs
Bovine primary
+astrocytes 0.8 0.4 Cecchelli et al., 1999
primary 0.5 Mar&Davis, 2002
Porcine primary 0.3 Franke et al., 1999
Rat RBE4 4.8 1.3 Rist et al, 1997
GPNT 7.4 2.6 1.0 0.05 Romero et al, 2003
Mouse b.END3 1.2 Omidi et al. 2003
Human primary 0.3 0.02 Unpublished results
+astrocytes
hCMEC/D3 1.7 0.4 0.3 0.01 Weksler et al. 2005
HUVEC 4.2 0.12 Unpublished results
CN
S m
icro
vasc
ular
EC
NON-CNS

The Blood-brain barrier (BBB) in health:
Transcriptional control of TJ proteins

Minimal promoter (endothelium) 1505-1853
Activity of minimal occludin promoter-GFP reporter in endothelia
1 400 800 1200 1600 2000
Lung endotheliumBrain endothelium
>1,000 putative binding sites in the occludin promoter (TESS)
Sade H, et al. (2009) Biochim Biophys
Acta
1789(3):175

Sp-3 binds to occludin minimal promoter in hCMEC/D3 cells (EMSA)
Cold probe inhibition
Antibody supershift
1 -400 -800 -1200 -1600 2000
F8

Mithramycin (Sp-inhibitor) decreases occludin
promoter-GFP reporter expression in hCMEC/D3 cells
0
20
40
60
80
100
120
Occludin Claudin 5 Z01
ControlMM treated
Med
ian
fluor
esce
nce
Control
Occ
Cl-5
ZO-1
200nM Mithramycin
*
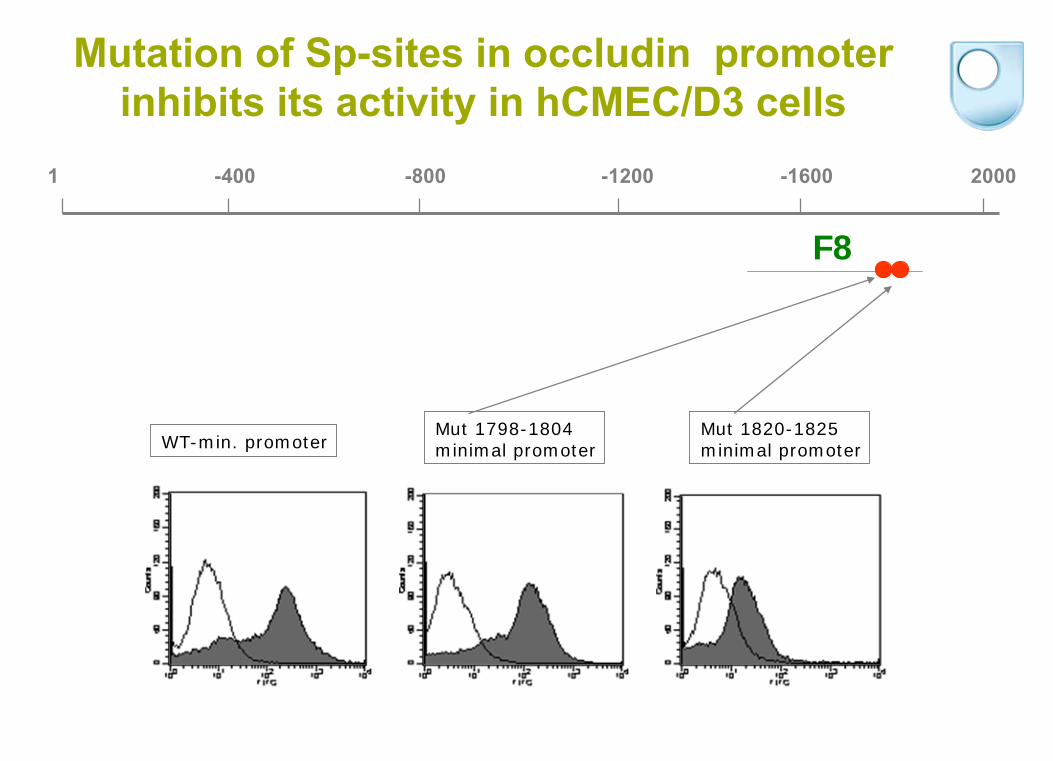
F8
1 -400 -800 -1200 -1600 2000
WT-min. promoterMut 1798-1804minimal promoter
Mut 1820-1825minimal promoter
Mutation of Sp-sites in occludin promoter inhibits its activity in hCMEC/D3 cells

The Blood-brain barrier (BBB) in health:
Role of the BBB in CNS availability of drugs

CNS penetration of drugs: Diffusion across BBBParameters limiting diffusion:
↓lipophilicity
H-bonds>8 MW>700 Da
vinblastine
vincristine
Abbott NJ, Romero IA.Mol Med Today. 1996

1 2 3 4TJ TJ
Rc +ve
(a) Passivediffusion
Lipidsolublenon-polarmolecules
(b) ABCTransporterEfflux
Lipidsolublenon-polarmoleculesandconjugates
(c) SoluteCarriersSLC
GlucoseAmino acidsNucleosidesMonocarboxylatesSmall peptidesFFAsOrganic anionsOrganic cations
(d) TranscytosisReceptor-mediatedAdsorptive-mediated
RMTTransferrinMelanotransferrinLipoproteinsAmyloid βGlycosylated
proteins
IgGInsulinLeptinTNFEGF
(e) MononuclearCellMigration
AMTCationised
albumin
HistoneAvidinTATSynB1(Cell penetrating peptides)
PgpBCRP
MRP1-5*
MRP1-5*
RMT AMT
Molecular and Cellular traffic Across the BBB

Correlation between in vivo (adult rats) and in vitro (hCMEC/D3 cells) permeability of several standard drugs
vincristine
colchicine
R = 0.938
0.01
0.1
1
10
100
0 1 2 3 4
Pe
(10-3 cm·min-1)
Kin
(10-3
ml·s
-1·g
-1)
Morphine-6--d- glucuronide
prazosin
diazepam
Weksler et al., FASEB J, 2005
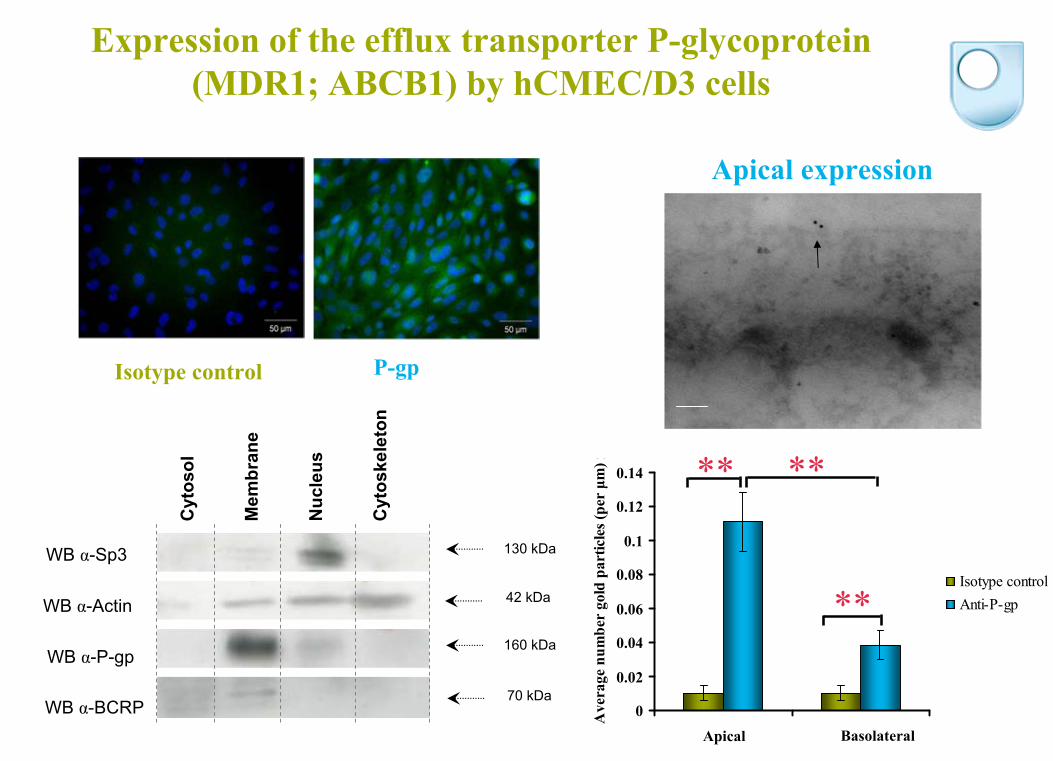
WB α-BCRP
Expression of the efflux transporter P-glycoprotein (MDR1; ABCB1) by hCMEC/D3 cells
WB α-Sp3
WB α-Actin
WB α-P-gp
130 kDa
42 kDa
160 kDa
70 kDa
Cyt
osol
Mem
bran
e
Nuc
leus
Cyt
oske
leto
n
Apical expression
0
0.02
0.04
0.06
0.08
0.1
0.12
0.14
Luminal Abluminal
Ave
rage
num
ber
gold
par
ticle
s (pe
r μm
) rIsotype controlAnti-P-gp
** **
**
Apical Basolateral
P-gp Isotype control
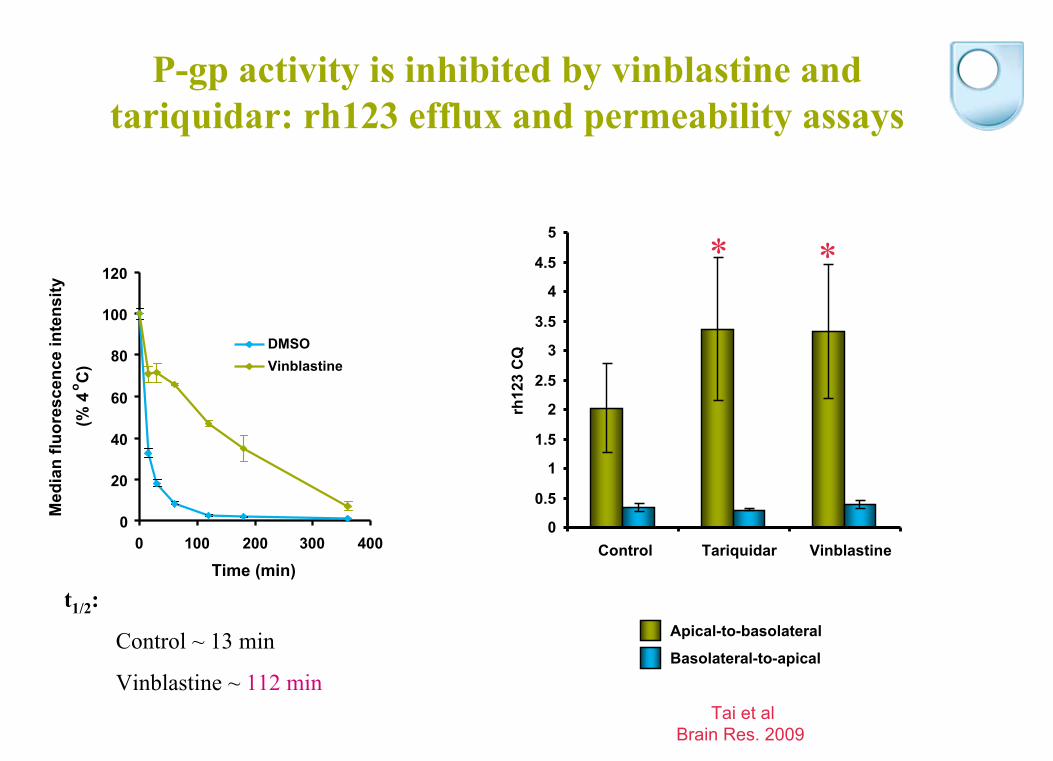
P-gp activity is inhibited by vinblastine and tariquidar: rh123 efflux and permeability assays
t1/2
:
Control ~ 13 min
Vinblastine ~ 112 min
0
20
40
60
80
100
120
0 100 200 300 400
Time (min)
Med
ian
fluor
esce
nce
inte
nsity
(%
4o C
)
DMSOVinblastine
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
5
Control Tariquidar Vinblastine
rh12
3 C
Q
Apical-to-basolateral
Basolateral-to-apical
* *
Tai et alBrain Res. 2009

0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
Control Tariquidar Vinblastine FTC
Aβ
clea
ranc
e qu
otie
nt
The apical-to-basolateral permeability of 125I Aβ 1-40
is increased by P-gp and BCRP inhibition in BECs
* **
P-gp inhibitors BCRP inhibitor
Basolateral to apical Apical to basolateral
Tai et al (2009) J Cereb Blood Flow Metab
29(6):1079-83.
Basolateral
Apical
Aβ 1-40
or inulin
0.1 nM 125I Aβ 1-40 30 min
Aβ CQ = 125IAP
/ 125I total14CAP
/14Ctotal

The Blood-brain barrier (BBB) in disease
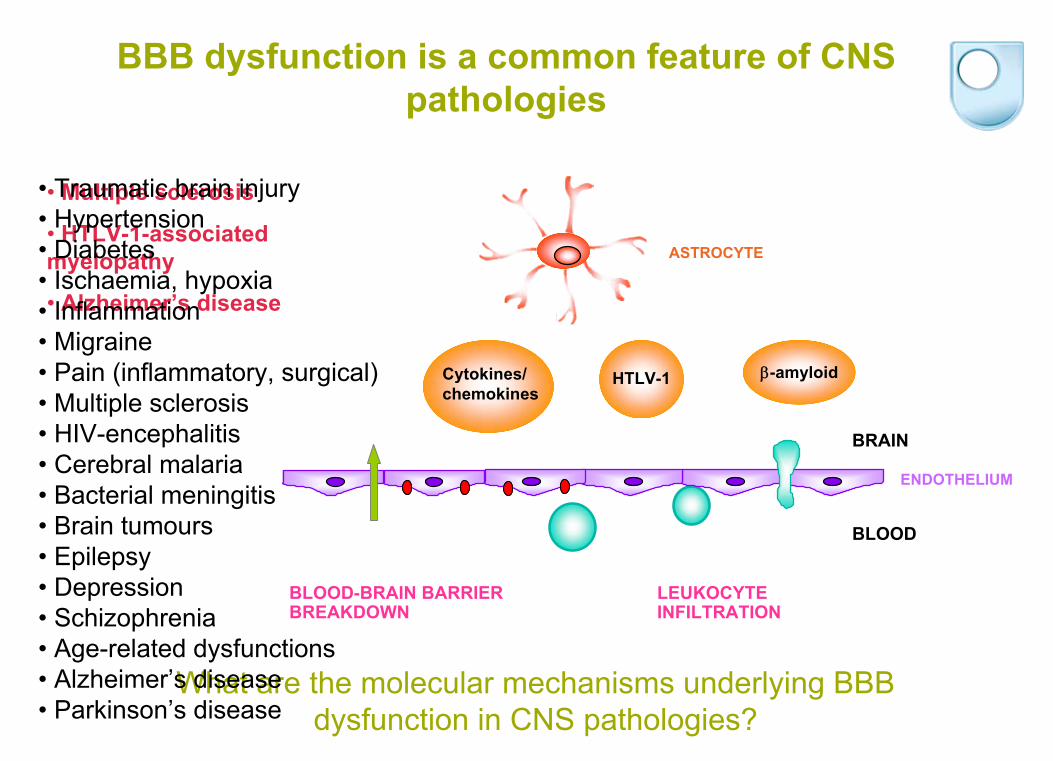
BBB dysfunction is a common feature of CNS pathologies
Cytokines/chemokines
-amyloidHTLV-1
BLOOD-BRAIN BARRIER BREAKDOWN
LEUKOCYTE INFILTRATION
ASTROCYTE
ENDOTHELIUM
BLOOD
BRAIN
What are the molecular mechanisms underlying BBB dysfunction in CNS pathologies?
• Multiple sclerosis
• HTLV-1-associated
myelopathy
• Alzheimer’s disease
• Traumatic brain injury• Hypertension• Diabetes• Ischaemia, hypoxia• Inflammation• Migraine• Pain (inflammatory, surgical)• Multiple sclerosis• HIV-encephalitis • Cerebral malaria• Bacterial meningitis• Brain tumours• Epilepsy• Depression• Schizophrenia• Age-related dysfunctions• Alzheimer’s disease• Parkinson’s disease

Consequences of BBB opening
If mild or transient → BBB resealing → protection against further stress If severe or chronic → BBB dysfunction → brain oedema and persistent CNS pathology
• Removes protective/homeostatic actions of barrier• Allows CNS access of peripheral transmitters/
neuroactive agents and vice versa
• Plasma albumin toxic → glial proliferation and scar
formation• Leukocyte infiltration → CNS inflammation

The Blood-brain barrier (BBB) in disease:
REGULATION OF TIGHT JUNCTIONAL PERMEABILITY

BBB breakdown in the spinal cord of HTLV-1-infected HAM/TSP patients
NON-HAM/TSP
HAM/TSP
-ZO1
-OCLN
NON-HAM/TSP HAM/TSP
-fibrinogen Afonso et al. (2007) J Immunol 179, 2576

MOG and spinal cord immunization induces BBB breakdown in mice
Fabis et al. (2007)
PNAS 104, 5656- 5661
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
2
Control Acute G1 Acute G4 Recovery Remission Chronic
Com
para
tive
expr
essi
on (
2-??
CT)
occludin
claudin-5

0
1
2
3
4
5
6
7
8
sAβ 1-40 Aβ 1-40 sAβ 1-42 Aβ 1-42
Pe (c
m/m
in x
10-5
)
Extracellular pathogenic stimuli increases hCMEC/D3 paracellular permeability
D3CEMHUT102C81-66
0 5 10 15 200.0
2.5
5.0
7.5
t (h)Afonso et al. (2007) J Immunol
179, 2576
HTLV-1-infected lymphocytes*
0
2
4
6
8
10
12
14
Cont
rol
TNF2
hTN
AFan
dIFN2
h
TNF6
hTN
AFan
dIFN6
h
TNF2
4hTN
AFan
dIFN2
4h
Pe
(1x1
0-5
cm/m
in)
TNF-αIFN-γ
- +- -
++
+-++ +- ++
2 h 6 h 24 h
Cytokines
Amyloid-
Tai et al. (2010) J Cell Mol Med 14(5):1101

TNFα and IL-1
are necessary for HTLV-1-
induced BBB breakdown
- IL1 TNF IL1+TNF0.0
2.5
5.0
7.5
10.0
0.0
2.5
5.0
7.5
Afonso et al. (2007) J Immunol 179, 2576

Mechanisms involved in disruption of tight junctions
• Redistribution of transmembrane and adaptor proteins from cell-cell junctions: transient BBB opening
– Alterations in actin cytoskeleton– Posttranslational modifications of AJ & TJ proteins– Proteolytic
degradation of AJ & TJ proteins
• Changes in expression of AJ & TJ proteins: chronic BBB opening

Afonso et al. (2007) J Immunol 179, 2576
Actin: FITC-phalloidin
Control HTLV-infected
Pathogenic stimuli induce reorganization of
actin cytoskeleton and TJ proteins
hCMEC/D3+PBL sAβ 1-40 Aβ
1-40
50 μm
B
F
A
C
Claudin-5
ZO-1
Occludin
50 μm
Amyloid-
Afonso et al. (2007) J Immunol
179, 2576
Tai et al. (2010) J Cell Mol Med 14(5):1101

00.20.40.60.8
11.21.41.61.8
mR
NA
rela
tive
leve
ls
0
0.2
0.4
0.6
0.8
1
1.2
Expression of occludin and claudinExpression of occludin and claudin--5 protein and mRNA is 5 protein and mRNA is decreased by combination of cytokines in hCMEC/D3 cellsdecreased by combination of cytokines in hCMEC/D3 cells
2
Occludin
Time hTNFα+IFNγ
6 24 2 6 24
Control
Actin
2 6 24Time h
% o
f con
trol
0
0.2
0.4
0.6
0.8
1
1.2
2
TNFα+IFNγ
6 24 2 6 24
Control
2 6 24Time h
% o
f con
trol
Claudin-5
Actin
1 2 3 4 5 6 12 24
Time (h)
occludinclaudin-5
TNF-α+IFN-γ

Signalling at the TJ
• INTERACTIONS WITH G PROTEINS: heterotrimeric
G proteins (e.g. Gi2) and small G proteins (e.g. Rho)
• INTERACTIONS WITH PROTEIN KINASES: PKC, PKA
• PHOSPHORYLATION OF TJ COMPONENTS: ZAK1, Casein kinase II, MLCK

Time (h) 0 1 2 3 4
5 6
Afonso et al. (2007) J Immunol 179, 2576
Identification of signalling pathways mediating BBB breakdown by pathogenic stimuli: HTLV-1-infected
T lymphocytes
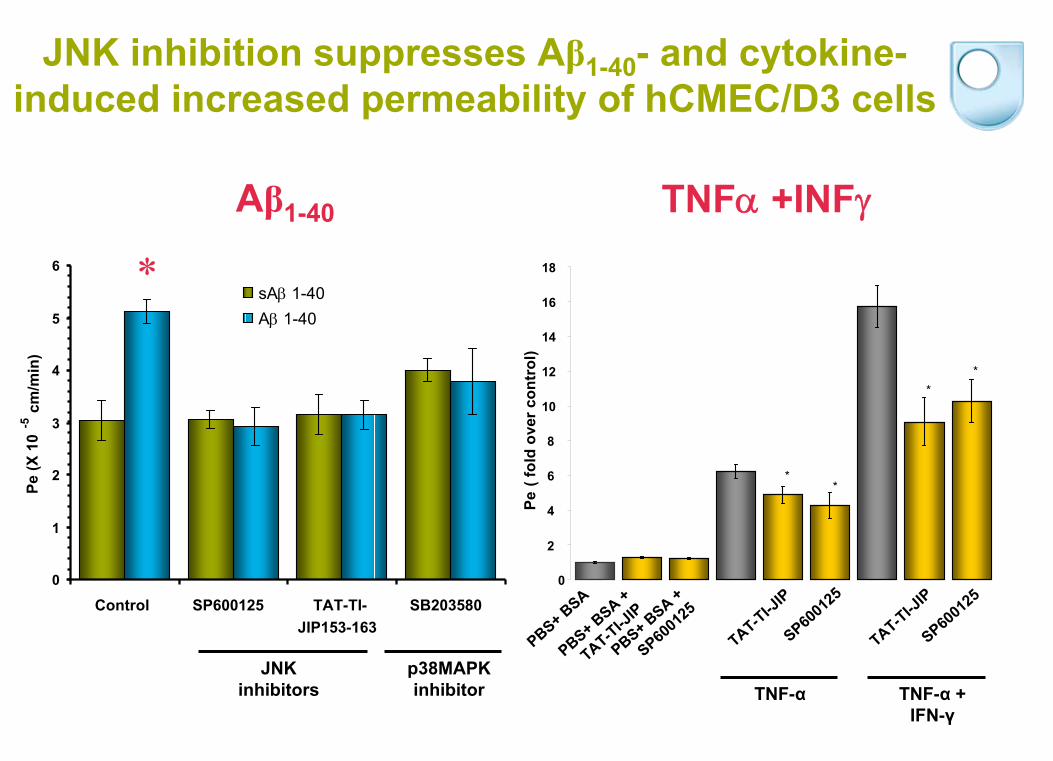
JNK inhibition suppresses Aβ1-40 - and cytokine-
induced increased permeability of hCMEC/D3 cells
*
Aβ1-40
0
1
2
3
4
5
6
Control SP600125 TAT-TI-JIP153-163
SB203580
Pe(X
10
-5cm
/min
)
sA 1-40
A 1-40
TNF +INF
Pe( f
old
over
con
trol
)
PBS+ BSA
PBS+ BSA +
SP6001
25
PBS+ BSA +
TAT-TI-JIP
SP6001
25
TAT-TI-JIP
SP6001
25
TAT-TI-JIP
TNF-α TNF-α +
IFN-γ
0
2
4
6
8
10
12
14
16
18
**
**
JNK inhibitors
p38MAPK inhibitor

Regulation of claudin-5 and occludin levels at the BBB by cytokines and amyloid-
TNF
Claudin-5 Occludin
amyloid
JNK
IFN
p38PKC Rho
ROCK
Regulation of TJ protein expression
Actin-myosin contraction
TJ protein re- distribution
Apoptosis
MLCK

• The hCMEC/D3 cell line is a suitable in vitro human
model to study BBB function in health and CNS pathologies
• Understanding the molecular basis of BBB function may
provide a rationale for targeted delivery to the CNS
• Understanding common molecular mechanisms mediating
BBB dysfunction in CNS pathologies may provide tools for targeted prophylaxis and therapies to BBB
Summary

Acknowledgments:
• Institut
Cochin
Pierre-Olivier Couraud
Paris, France
Babette Weksler
• Institut
Pasteur
Simona Ozden
Paris, France
Pierre-Emmanuel Ceccaldi
• National Research Council
Arsalan
Haqqani
Ottawa, Canada
Danica Stanimirovic
• QMUL, London, UK
David Baker
Greg Michael
• University of Sheffield, UK
Basil Sharrack
• The Open University
David Male
Milton Keynes, UK
Jane Loughlin Mark Hirst
Karen Logan Evanne
Subileau
Hadassah Sade Leon
Tai
Alejandro Lopez-Ramirez Camilla Cerutti

THE BLOOD-BRAIN BARRIER
3-year Post-Doctoral Research Associate:
microRNAs in the cerebral vasculature and multiple sclerosis
Deadline: 30th November


















