
Kvadrupleksne strukture DNKmafija.fmf.uni-lj.si/seminar/files/2009_2010/... · 2010-04-07 ·...
19
Seminar – 4. letnik Kvadrupleksne strukture DNK Avtor: Tinkara Troha Mentor: prof. dr. Irena Drevenšek-Olenik Ljubljana, marec 2010 Povzetek V seminarju sem predstavila kvadrupleksne strukture DNK in njihovo uporabo. Opisala sem nastanek molekulskih orbital, katerih razumevanje je ključno pri razlagi nalaganja baz v kvadrupleksih. Nalaganje baz v kvadrupleksih je opisano s preprostim modelom π- π interakcij. Izpostavila sem tudi pomen kovinskih ionov za nastanek in stabilizacijo kvadrupleksov, na koncu pa sem navedla še nekaj možnih načinov zlaganja kvadrupleksov. Kot primer uporabe sem opisala G-žice. 1
Transcript of Kvadrupleksne strukture DNKmafija.fmf.uni-lj.si/seminar/files/2009_2010/... · 2010-04-07 ·...

Seminar – 4. letnik
Kvadrupleksne strukture DNK
Avtor: Tinkara TrohaMentor: prof. dr. Irena Drevenšek-Olenik
Ljubljana, marec 2010
Povzetek
V seminarju sem predstavila kvadrupleksne strukture DNK in njihovo uporabo. Opisala sem nastanek molekulskih orbital, katerih razumevanje je ključno pri razlagi nalaganja baz v kvadrupleksih. Nalaganje baz v kvadrupleksih je opisano s preprostim modelom π-π interakcij. Izpostavila sem tudi pomen kovinskih ionov za nastanek in stabilizacijo kvadrupleksov, na koncu pa sem navedla še nekaj možnih načinov zlaganja kvadrupleksov. Kot primer uporabe sem opisala G-žice.
1

Kazalo
1 Uvod 2
2 Zgodovina in osnove kvadrupleksov 3 3 Osnovni gradniki DNK in RNK 4 4 Teorija molekulskih orbital 6
4.1 Hibridizacija . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 84.2 Delokalizirane orbitale . . . . . . . . . . . . . . . . . . . . . . . . . . 9
5 Model π-π interakcij 9
6 Koordinacija kovinskih ionov 12
7 Zlaganje in topologija kvadrupleksov 12
8 Dosežki Nobelovih nagrajencev 148.1 Povezava med telomeri in rakom . . . . . . . . . . . . . . . . . . . . . 16
9 G-žice 16
10 Zaključek 18
1 Uvod
Molekula DNK lahko poleg dobro znane desnosučne dvojne vijačnice (oblika B) tvori še celo vrsto drugih oblik. Med drugim lahko molekule DNK tvorijo strukture višjega reda znane pod imenom kvadrupleksi. Slednji so bili prvič opaženi okoli leta 1960 pri študijah DNK zaporedij bogatih z gvaninskimi nukleotidi. Izkazalo se je, da gvaninski polinukleotidi tvorijo strukture, sestavljene iz štirih pramenov, katerih osnovni strukturni element je G-kvartet, ki ga sestaljajo štirje z vodikovimi vezmi povezani gvaninski nukleotidi [1]. Zanimanje za G- kvadruplekse se je v zadnjem času močno povečalo, saj se je izkazalo, da sekvence omenjenih oblik najdemo na koncih kromosomov, v t.i. telomeričnem območju in v transkripcijskih regulacijskih območjih v večih pomembnih onkogenih. Ker so vzdrževalni mehanizmi telomer in regulacija transkripcije onkogena pomembni pri načrtovanju zdravil, predstavljajo G-kvadrupleksi pomembno področje raziskovanja v onkologiji oz. Farmaciji [1, 6].
2

2 Zgodovina in osnove kvadrupleksov
Spontano povezovanje gvaninov v vodnih raztopinah v milimolarnih koncentracijah je bilo opaženo že v 19. stoletju. Leta 1960 je Gellert s kristalografskimi metodami določil, da se gvaninske baze združujejo v tetramerično ureditev, ki jo je poimenoval G-kvartet (slika1). Posebna lastnost molekule gvanozina, ki omogoča tvorbo teh G-kvartetov, je da vsebuje donorske in akceptorske skupine za tvorbo vodikove vezi. Poleg tega pa ima gvanin tudi polarizabilno aromatsko površino. Z razvojem kemijske sinteze poligvaninskih oligonukleotidov pa so bile opažene tudi nekatere druge oblike povezovanja gvaninov. Meritve cirkularnega dikroizma (CD) in infrardeča (IR) spektroskopija sta potrdili ureditev gvaninov v G-kvartete, medtem ko so raziskave s sipanjem žarkov X pokazale štiri pramenski motiv s tetradami naloženimi v sklade, poimenovanih kvadrupleksi. Zgradba kvadrupleksov je zelo podobna dvojni vijačnici DNK in jo opišemo s predpisanim porastom višine in zasukom med sosednjimi tetradami. Vijačna struktura je desnosučna. V kvadrupleksih fosftna glavna veriga, ki povezuje nukleozide, generira štiri žlebiče različnih širin, namesto dveh, ki sta značilna za dvojno vijačnico DNK [1].
Slika 1: Shema G-kvarteta. Črtkane črte predstavljajo vodikove vezi med Hoogsteenovimi in Watson-Crick-ovimi povezovalnimi mesti. V središču G-kvarteta je kalijev ion [1].
Zanimanje za strukturno ureditev G-kvadrupleksov se je močno povečalo okoli leta 1990 z odkritjem z gvaninom bogatih ponavljajočih se sekvenc na koncu kromosomov in encima, ki je udeležen pri vzdrževanju te ponavljajoče se telomerne DNK. Zasluge za prelom v razumevanju prepisovanja kromosomov imajo trije Američani (Elizabeth Blackbourn, Carol Greider in Jack Szostak ), ki so leta 2009 za svoje delo tudi prejeli nobelovo nagrado za medicino [9].
3

a) b)
Dupleksna B-DNK Kvadrupleksna DNK
Slika 2: (a) En spiralen obrat vijačnice B-DNK; (b) Dva naložena paralelna G-kvadrupleksa [1].
3 Osnovni gradniki DNK in RNK
Nukleinske kisline so polimeri, katerih osnovna enota je nukleotid. Vsak nukleotid pa je sestavljen iz treh komponent: dušikove baze, sladkorja in fosfatne skupine. Pri DNK je sladkor deoksiriboza, pri RNK pa riboza. Dušikove baze v DNK vključujejo adenin, timin, gvanin in citozin, medtem ko pri RNK namesto timina nastopa uracil. Baza in sladkor skupaj tvorita nukleozid, fosfatna skupina pa je pripeta na 5´ stran nukleozida in služi kot povezovalni element med nukleozidi. Polimeri, ki so sestavljeni iz teh treh komponent, imajo značilnosti, ki so idealne za dolgotrajno skladiščenje genetskih informacij v živih celicah. Nukleotidi so kemično stabilni in posamezne molekule se lahko med seboj združijo preko komplementarnih baz in tvorijo dvojno vijačnico. Takšni polimeri so sposobni hraniti komplementarne kopije genetskih informacij v kompaktni obliki, ki se lahko razdruži in zopet združi [1, 10].Dušikove baze so ključne komponente, zaradi katerih je DNK/RNK tako kemično raznolika. Baze vsebujejo komplementarne donorje in akceptorje vodikove vezi, ki povzročajo specifične vezi med bazami. Obstajata dva načina tvorbe vodikovih vezi, preko Watson-Crickovih in Hoogstenovih vezavnih mest. Parjenje ponavadi poteka med purini (gvanin, adenin) in pirimidini (timin, citozin in uracil) preko Watson-Crickovega pravila baznih parov. Možni pa so tudi drugi načini parjenja med bazami, in sicer z vključitvijo Hoogsteenove interakcije. Slednji način je pomemben pri tvorbi in stabilizaciji tetrad.
4
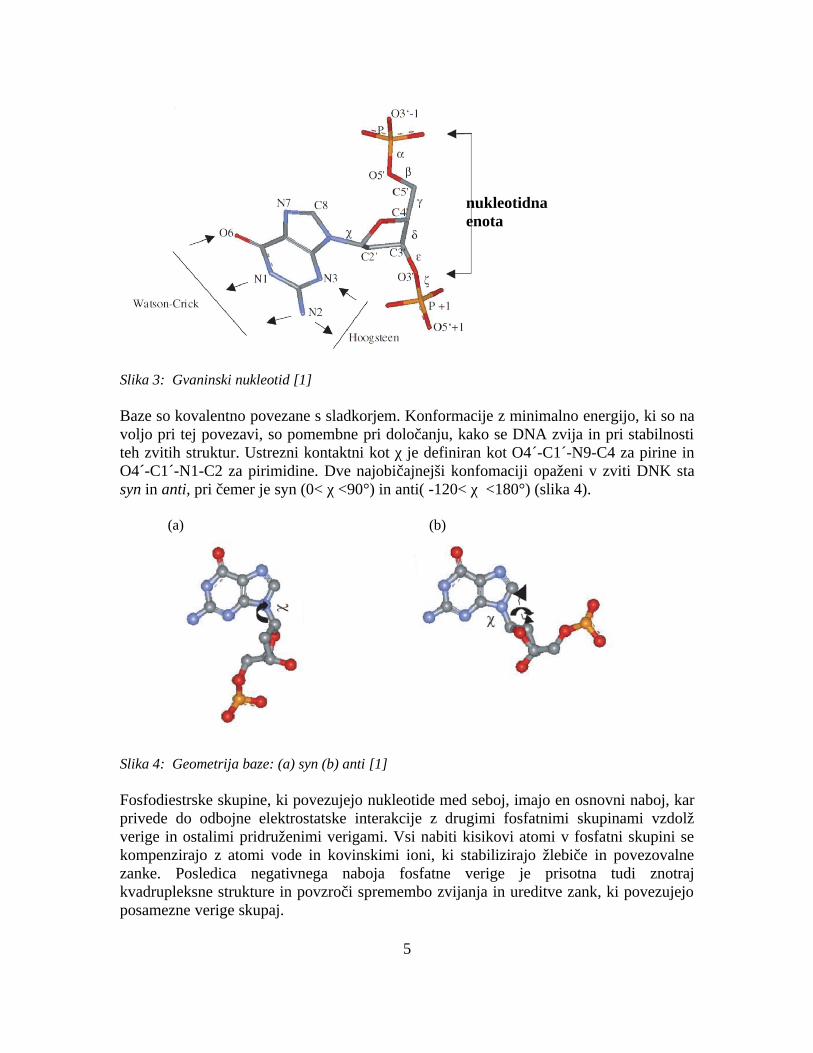
Slika 3: Gvaninski nukleotid [1]
Baze so kovalentno povezane s sladkorjem. Konformacije z minimalno energijo, ki so na voljo pri tej povezavi, so pomembne pri določanju, kako se DNA zvija in pri stabilnosti teh zvitih struktur. Ustrezni kontaktni kot χ je definiran kot O4´-C1´-N9-C4 za pirine in O4´-C1´-N1-C2 za pirimidine. Dve najobičajnejši konfomaciji opaženi v zviti DNK sta syn in anti, pri čemer je syn (0< χ <90°) in anti( -120< χ <180°) (slika 4).
(a) (b)
Slika 4: Geometrija baze: (a) syn (b) anti [1]
Fosfodiestrske skupine, ki povezujejo nukleotide med seboj, imajo en osnovni naboj, kar privede do odbojne elektrostatske interakcije z drugimi fosfatnimi skupinami vzdolž verige in ostalimi pridruženimi verigami. Vsi nabiti kisikovi atomi v fosfatni skupini se kompenzirajo z atomi vode in kovinskimi ioni, ki stabilizirajo žlebiče in povezovalne zanke. Posledica negativnega naboja fosfatne verige je prisotna tudi znotraj kvadrupleksne strukture in povzroči spremembo zvijanja in ureditve zank, ki povezujejo posamezne verige skupaj.
5
nukleotidna enota

3.1 Nalaganje baz
Baze v DNK so nepolarne in ne interagirajo s polarnimi topili. Baze, ki nastopajo v parih, se bodo združile in naložile ena na drugo, da bi zmanjšale prostor izpostavljen topilu. K stabilnosti DNK torej prispeva nalaganje baz, ki vključuje kombinacijo hidrofobnih, elektrostatskih in Van der Waalsovih sil. V kanonični B-DNA so vezavne energije, povezane z nalaganjem v sklade, ocenjene na približno od -9.5 in -13.2 kcal/mol za korak GC baznega para, medtem ko ima korak AT baznega para manjšo energijo, okoli -5.4 kcal/mol. Te naložene baze ne morejo direktno »sesti« ena na drugo, ampak so zvite okoli spiralne osi. Razdalja med baznimi pari je definirana kot višina oz. hod vijačnice, rotacija okoli spiralne osi pa je definirana kot zasuk. Za popoln opis pozicije ene baze glede na drugo so potrebni še drugi parametri, kot so vrtenje, nagib in premik.Natančnejša razlaga, zakaj se baze sploh nalagajo druga na drugo, je prikazana z modelom π- π interakcij [2]. Za lažje razumevanje tega modela bom najprej na kratko opisala nastanek molekulskih orbital.
4 Teorija molekulskih orbital
Po teoriji molekulskih orbital iz atomskih orbital dveh atomov nastanejo molekulske orbitale, ko se atomska jedra nahajajo na določeni razdalji (ravnotežna razdalja). V nastalih skupnih molekulskih orbitalah se potem nahajajo elektroni spojenih atomov. Kadar so elektroni pretežno bližje enemu ali drugemu jedru, potem je obnašanje elektrona v molekulski orbitali skoraj enako kot v njegovi atomski orbitali. Torej kadar je elektron v bližini jedra A, mora biti molekulska orbitala (MO) podobna atomski orbitali nevezanega atoma- ψA (enako velja za elektron, ki se nahaja v bližini jedra B). Takšna molekulska orbitala se imenuje enoelektronska molekulska orbitala. Skupna molekulska orbitala ψMO mora imeti karakteristike atomskih orbital ψA in ψB. Iz dveh atomskih orbital ψA in ψB nastaneta z linearno kombinacijo dve enoelektronski molekulski orbitali:
ψMO = ψA + ψB (1)
ψMO* = ψA – ψB (2)
Dve atomski orbitali, iz katerih s prekrivanjem nastanejo molekulske orbitale, vsebujejo štiri elektrone. Prav tako tudi dve novonastali molekulski orbitali vsebujeta štiri elektrone. Medsebojnemu povezovanju atomov (zaradi povečanja amplitude vala) odgovarja le funkcija ψMO. Molekulska valovna funkcija za elektron 1 je:
ψMO(1)= ψA(1) + ψB(1) (3)in za elektron 2:
ψMO(2) = ψA(2)+ ψB(2) (4)
Za dvoelektronski sistem (npr. molekula H2) je skupna valovna funkcija zmnožek molekulskih valovnih funkcij za vsaki atom:
6

ψ = ψMO(1) ψMO(2) = [ ψA(1) + ψB(1)] [ψA(2)+ ψB(2)] (5)
Verjetnost elektronske gostote dobimo s kvadriranjem izrazov za ψMO in ψMO*:
ψMO2 = ψA
2 + 2 ψA ψB + ψB2 (6)
ψMO*2= ψA
2 - 2 ψA ψB + ψB2 (7)
ψMO2 in ψMO
*2 so elektronske gostote med atomoma A in B. ψA2 in ψB
2 predstavljata elektronsko gostoto nespojenih atomov A in B, njuna vsota ψA
2 + ψB2 pa predstavlja
vsoto elektronske gostote med nespojenima atomoma A in B. Torej velikost 2 ψA ψB
določa elektronsko gostoto med jedroma spojenih atomov A in B (stopnja prekrivanja). Če je ta količina pozitivna, potem je gostota med spojenima atomoma A in B večja kot med nespojenima. Večja prostornina, v kateri se elektron giblje, zmanjšuje njegovo energijo (medsebojno odbijanje elektrona). Iz tega sledi, da je energija molekulske orbitale ψMO manjša od vsote energij posameznih atomskih orbital ψA + ψB. Bolj kot se atomske orbitale prekrivajo, kar pomeni večjo gostoto elektronov med jedroma spojenih atomov, nižja je energija molekulske orbitale in stabilnejša je nastala vez. Zaradi tega se valovna funkcija ψMO imenuje vezna orbitala, ker omogoča povezovanje atoma v molekulo. Po drugi strani pa se zaradi istih razlogov valovna funkcija ψMO
* imenuje protivezna orbitala.
(a) (b)
Slika 5: Verjetnostne gostote za elektron za (a) vezno orbitalo in (b) nevezno orbitalo [7]
Atomska orbitala, ki je podana z valovno funkcijo ψ, je lahko pozitivno ali negativno predznačena. Valovi elektrona v atomu so stoječi valovi, zato predstavlja atomska orbitala stoječi val, ki ima pozitivni predznak, če prestavlja vrh in negativni predznak, če predstavlja dolino vala. S prekrivanjem atomskih orbital istega algebrskega predznaka nastane vezna molekulska orbitala, s prekrivanjem atomskih orbital negativnega algebrskega predznaka pa nastanejo protvezne molekulske orbitale. Poznamo dve vrsti veznih molekulskih orbital:
− σ orbitale, ki nastanejo s prekrivanjem s-orbitale s s- ali p- ali d-orbitalo
7

− π orbitale, ki nastanejo z bočnim prekrivanjem p-orbital s p-orbitalami ali d-orbital z d-orbitalami
Slika 6: nastanek σ- orbitale oz. σ- vezi [8]
Slika 7: nastanek π-orbitale oz. Π-vezi [8]
4.1 Hibridizacija
Hibridizacija je linearna kombinacija atomskih orbital približno enake energije, pri čemer se tvorijo hibridne oz. hibridizirane orbitale, ki imajo enake energijske nivoje.
Za primer sem opisala nastanek hibridnih orbital v benzenu zaradi podobnosti z dušikovimi bazami, ki tako kot benzen vsebujejo aromatski obroč. Ogljik ima elektronsko konfiguracijo 1s2 2s2 2p2 , zato bi pričakovali, da bo tvoril dve kovalentni vezi. Vendar pa ogljik v molekuli benzena tvori štiri vezi. Tri hibridne orbitale nastanejo z linearno kombinacijo 2s, 2px in 2py orbital, 2pz orbitala pa ostane nespremenjena.
Hibridizirane orbitale:
Ψ1 = (Ψ2s + Ψ2px + Ψ2py) (8)
Ψ2 = (Ψ2s - Ψ2px - Ψ2py ) (9)
Ψ3 = (Ψ2s + Ψ2px - Ψ2py ) (10)
Gostote verjetnosti imajo obliko ročk v ravnini, med katerimi je kot 120°.
8

4.2 Delokalizirane orbitale
Po teoriji molekulskih orbital se lahko molekulske orbitale raztezajo preko celotne molekule in jih lahko smatramo kot delokalizirane, ker povezujejo več atomskih jeder. Nekatere molekulske strukture je mogoče opisati le na ta način, da predpostavimo, da so nekateri elektroni delokalizirani. Ti elektroni se nahajajo v delokaliziranih orbitalah in povezujejo več atomskih jeder. Za primer vzemimo molekulo benzena C6H6. Z medsebojnim prekrivanjem sp2 hibridnih orbital šestih C-atomov in njihovim prekrivanjem s s-orbitalami šestih H-atomov nastane dvanajst lokaliziranih σ-vezi, ki ležijo v ravnini molekule. Preostalih šest 2p-orbital C-atomov, ki so vertikalne na ravnino molekule, pa z medsebojnim prekrivanjem ustvari šest molekulskih π-orbital. Od tega so tri vezne in tri protivezne molekulske orbitale. Šest elektronov se nahaja v treh veznih molekulskih orbitalah, tri protivezne orbitale pa so prazne. Na ta način je šest elektronov v molekulskih π-orbitalah delokalizirano, a so zaradi medsebojnega odbijanja enakomerno razporejeni preko celotne strukture šestih C-atomov. Molekula benzena torej sestoji iz ogrodja lokaliziranih σ-vezi v eni ravnini, nad in pod tem ogrodjem pa se nahajajo delokalizirane π-vezi. Njihovi elektronski oblaki imajo obliko koluta z malo večjo gostoto na mestu C-atomov [3, 7].
5 Model π- π interakcij
Preprost model porazdelitve naboja v planarnih molekulah z delokaliziranimi elektroni (π-sistem) je uporaben za razlago geometrije intermolekularnih interakcij med aromatskimi molekulami, prav tako pa je primeren za opis nalaganja baz v nukleinskih kislinah.
Interakcijsko energijo med dvema molekulama zapišemo:
Etotal = Eelectrostatic + Einduction + Edispersion + Erepulsion , (8)
kjer Eelectrostatic označuje elektrostatsko energijo med stalnimi multipoli, Einduction
predstavlja interakcijo med stalnim multipolom na eni molekuli z induciranim multipolom na drugi molekuli. Edispersion je interakcija med trenutnimi multipoli molekul, ki nimajo stalnega multipolnega momenta, Erepulsion pa je interakcija, ki preprečuje kolaps molekul in prevladuje, ko se molekule približujejo ena drugi.K interakcijski energiji največ prispevajo elektrostatske in van der Waalsove komponente (Edispersion ), indukcijsko interakcijo in sile kratkega dosega zanemarimo. Očitno je, da interakcijska energija med dvema molekulama v raztopini vključuje združenje dveh molekul in premik molekul topila. Vezavna energija in energija raztapljanja pa sta povezani z van der Waalsovimi interakcijami.Van der Waalsova interakcija med dvema molekulama se izračuna po enačbi:
Evan der Waals = ∑ {Aij exp(-αij rij) – Cij/rij6} , (9)
9

kjer je rij razdalja med atomom i v prvi molekuli in atomom j v drugi, koeficienti Cij, Aij
in αij pa so standardni parametri. Van der Waalsovo energijo moramo oceniti, da bi dobili kvantitativnmo vrednost magnitude interakcije.Elektrostatska interakcija: Z oceno elektrostatske energije za interakcijo dveh π sistemov lahko predvidimo geometrijo π –π interakcije. Preprost model π-sistema je ilustriran na sliki 8a: sestavljajo ga pozitivno nabito σ-ogrodje med dvema negativno nabitima π-elektronskima oblakoma. Interakcija med dvema takima π-sistemoma je odbojna. Ko je razdalja med dvema π-sistemoma primerljiva z njuno debelino, pa moramo π-elektrone obravnavati ločeno od σ-ogrodja. Za predstavitev porazdelitve elektrostatskega naboja uporabimo niz točkastih nabojev (slika 8b) in izračunamo elektrostatsko interakcijo kot vsoto interakcij med vsemi pari nabojev dveh π-sistemov. Ta približek je osnova metode DMA (distributed multipole approach) za izračun intermolekularnih elektrostatskih potencialov.
(a) (b)
Slika 8: (a) interakcija med dvema zrcalnima π-sistemoma. (b) Model atoma, ki prispeva en elektron k π-sistemu [2].
V najpreprostejšem primeru uporabimo v π-sistemu za vsak C-atom naboj +1 v jedru atoma in dva naboja -1/2 na razdalji δ nad in pod nivojem π-sistema. Parameter δ je določen z eksperimentalno vrednostjo kvadrupolnega momenta benzena in njegova optimalna vrednost je δ=0.47A. To vrednost uporabimo kot približek za δ v drugih π-sistemih. Za prikaz idealiziranega π-sistema oz. π-atoma torej uporabimo tri naboje. Na sliki je prikazano, kako se interakcija med dvema takšnima π-sistemoma spreminja kot funkcija njune orientacije. Na y-osi je kot rotacije okoli centralnega pozitivnega naboja zgornjega π-atoma, na x-osi pa je premik zgornjega π-atoma glede na spodnjega. Geometrija dveh π-sistemov, pri katerem sistema nista zasukana niti premaknjena drug na drugega (zrcalna geometrija), je v odbojnem območju. Slika 9 prikazuje dve limitni geometriji, ki vodita
10

do ugodne elektrostatske interakcije ((a) zgornji π-atom je zavrten za kot 90º glede na drugega; (b) zgornji π-atom je zamaknjen glede na drugega). Pri omenjenih geometrijah je σ-π privlak dominantna interakcija, medtem ko pri zrcalni geometriji prevladuje π-π odboj. Za napoved geometrije celotne molekule moramo sešteti elektrostatske interakcije preko vseh atomov. Poleg tega je treba upoštevati še prisotnost polariziranih atomov. Če imamo dva π-sistema v zrcalni geometriji, kjer je prevladujoča interakcija odbojna, potem bo atom, ki prispeva delokalizirane elektrone, stabiliziral interakcijo z zmanjšanjem odboja. Nasprotno pa bo atom, ki ne prispevadelokaliziranih elektronov, še bolj destabiliziral interakcijo.
Slika 9: Interakcija med dvema idealiziranima π-atomoma kot funkcija orientacije med njima [2].
V DNK so π -π interakcije prisotne med sosednjimi nukleotidi in prispevajo k stabilnosti molekularne strukture. Dušikove baze vsebujejo aromatske obroče, ki so pravokotni na dolžino DNK pramena. Ravnine aromatskih obročev so urejene paralelno ena na drugo, kar bazam omogoča medsebojne aromatske interakcije. Iz opisanega modela se da napovedati tudi geometrijo naloženih tetrad. Izkaže se, da je najugodnejša konfiguracija takšna, pri kateri so sosednji G-kvarteti zasukani za 30°, razdalja med njimi pa je 3.3 Ǻ [2, 4].
11

6 Koordinacija kovinskih ionov
Odvisnost kvadrupleksne formacije od koordinacije kovinskih ionov je bila znana od samega odkritja kvadrupleksnih struktur. K nestabilnosti kvadrupleksov prispeva ureditev gvaninske O6 karbonilne skupine v centru G-kvarteta. O6 atomi tvorijo kvadratno planarno zgradbo za vsak kvartet z zasukom 30° in višino 3.3Ǻ med posamezno tetrado. Te negativno nabite luknje med G-tetradami morajo biti stabilizirane s kationi. Množica enovalenčnih in dvovalenčnih kationov je dejansko sposobna stabilizirati kvadrupleksne formacije različnih stopenj. Ioni povečujejo stabilnost strukture, kar se najbolj jasno vidi pri zvijanju kvadrupleksov v raztopini s K+ in Na+ ioni. Povečanje koncentracije enovalentnih kationov ima velik prispevek pri stopnji zgibanja. Dodajanje kalijevih ionov ima večji učinek kot dodajanje natrijevih, kar se kaže v stabilnosti kvadrupleksa. Karakteristike kalijevih ionov so idealne za učinkovito koordinacijo karboksilnih kisikovih atomov, ki so za 2.73 A oddaljeni od vsakega O6 atoma. Idejo o kationih, ki stabilizirajo en kvartet preko drugega, sta prvič opisala Sen in Gilbert kot Na+ -K+
zamenjavo. Ta zamenjava je biološko pomembna, saj je koncentracija K+ znotraj celice velika v primerjavi s koncentracijo Na+, medtem ko velja ravno obratno za območje izven celice. Dodajanje Mg2+ ionov in drugih multivalentnih kationov pomaga k združitvi kvadrupleksov, lahko pa tudi zmanjša stabilnost že zvitega kvadrupleksa.
Slika 10: Koordinacija kovinskih ionov med tetradami. Prikazan je zasuk med bazami. Kalijev ion se nahaja med osmimi karbonilnimi kisikovimi atomi s povprečno razdaljo 2.73 Å med njimi [1].
7 Zlaganje in topologija kvadrupleksov
Za nastanek kvadrupleksne strukture so potrebne štiri gvaninske baze, ki se med seboj združijo. Najenostavnejša zgradba je oblikovana pri združitvi štirih DNK pramenov v raztopini, ki vsebuje kratke gvaninske nize, na primer:
Xn Gp Xn ,
12

kjer je Xn katerikoli nukleotid dolžine n in Gp je neko število gvaninov vpletenih v tetradno strukturo dolžine p. Strukture, ki vsebujejo takšne kratke gvanozinske nize, se imenujejo intermolekularni oz. tetramolekularni kvadrupleksi.Obstajajo štiri načini združevanja pramenov, kot je vidno na sliki 11(a-d). V enomernih raztopinah, ki vsebujejo posamezne G-nize, so bile opažene samo vzporedne ureditve, kjer fosfatna veriga teče v enaki smeri, in kjer so vse baze v antiglikozidni orientaciji [slika 11(a)].
Slika 11: Slike prikazujejo polarnost pramenov in z njimi povezane glikozidne torzijske kote intermolekularnih kvadrupleksov. (a,b) Paralelni kvadrupleks s samimi anti/syn glikozidnimi koti; (c,d) antiparalelni kvadrupleksi s pripadajočimi anti in syn konformacijami [1].
Kompleksnejše strukture in topologije se lahko tvorijo iz pramenov, ki vsebujejo dve gvaninski ponovitvi, ločeni z negvaninskimi nukleotidi, na primer:
Xn Go Xp Go Xn ,
kjer je Xn katerikoli negvaninski nukleotid dolžine n, Go je število gvaninov udeleženih v nastanek tetrade dolžine o, Xp pa je katerikoli nukleotid dolžine p udeležen pri nastanku zanke.Če združimo dva od teh pramenov, dobimo štiri pramenske kvadruplekse, imenovane dimerni kvadrupleksi. Topologija teh kvadrupleksov je odvisna od tipa pramenov, ki se povežejo med seboj. Element, ki povezuje pramene, je lahko diagonalen, obstranski ali zunanji glede na kvadrupleks. Povezovalni nukleotidi so kritični pri določanju kvadrupleksne stabilnosti v odvisnosti od dolžine in bazne sekvence.
Intramolekularni kvadrupleksi pa se združujejo iz naslednjih splošnih sekvenc:
Xn Go Xp Go Xp Go Xp Go Xn.
Topologije, ki jih lahko zavzemajo intramolekularni kvadrupleksi, so različne in kompleksnejše kot tiste, ki so na voljo intermolekularnim ali bimolekularnim
13

kvadrupleksom, kar je posledica dodatnih povezujočih nukleotidov.
Slika 12: Razporeditev zank in pripadajočih topologij v bimolekularnih kvadrupleksih [1].
Izmed naštetih kvadrupleksov so termično najstabilnejši tetramolekularni kvadrupleksi. Sekvence, ki vsebujejo štiri ali več G-bogatih ponovitev, so ponavadi povezane s telomernimi regijami nahajajočimi se na koncu kromosomov. Organizmi z linearnimi kromosomi varujejo DNK z G-bogatimi ponavljajočimi se sekvencami vsebovanimi znotraj dvojne vijačnice DNK, ki se konča z enopramenskim 3’ previsom. Le te 3’ razširjene enopramenske G-ponovitve so na voljo za nastanek kvadrupleksa brez oviranja od komplementarnega 5’ C-bogatega pramena. Ponavljajoče se G-bogate sekvence prav tako najdemo v genomu, povezanim s promotorskimi območji.
8 Glavni dosežki nobelovih nagrajencev
Zanimanje za telomere se je začelo leta 1930, ko sta raziskovalca B. McClintock in H. Mueller opazila, da so konci kromosomov zaščiteni. Takrat struktura kromosomov ni bila znana. Raziskovalci so vedeli, da kromosomi nosijo genetski material, ki ga ščitijo telomere, ni pa bilo znano, na kakšen način ta zaščita deluje. Telomeri so ponavljajoče DNK sekvence na koncih kromosomov, ki imajo pomembno varovalno vlogo v preprečevanju razgradnje kromosomskih koncev. Pri vsaki delitvi somatskih celic se izgubi del telomerov. Delitev celic brez telomerov bi povzročila izgubo informacij na koncu kromosoma. Večina telomerov je zgrajena iz kratkih ponavljajočih se oligonukleotidnih zaporedij. Telomeri sesalcev imajo 5 do 25 kilobaz dolge ponovitve šestih nukleotidov TTAGGG v smeri 5' -> 3' proti koncu kromosoma. To pomeni, da so mesta bogata z gvanini na tisti verigi dvojne vijačnice, ki predstavlja 3'-konec kromosoma. Nasprotna veriga vijačnice (5'-konec) pa je bogata s citozini. S C
14

bogata veriga je nekoliko krajša od verige bogate z G, zaradi česar nastane enoverižni 3'-previs. Telomeri se ne končajo linearno, ampak tvorijo zankaste strukture z G-bogatimi in s C-bogatimi verigami (slika 14). Nastanek in vzdrževanje te zanke pa olajšujejo proteini, ki se na koncu vsakega kromosoma vežejo na DNA.
Slika 13: Sladkorni obroč z označenimi ogljikovimi atomi. 5´ konec DNK označuje tisti pramen molekule DNK, ki ima 5´ C-atom v sladkornem obroču deoksiriboze [13].
Replikacija DNK se začne na sredini polinukleotidne verige. Podvaja jo encim DNA-polimeraza, ki vedno potuje od 5'- proti 3'-koncu. Verigo, ki poteka od 3' -> 5', polimeraza popolnoma prepiše, saj jo lahko prepisuje v smeri 5' -> 3'. Komplementarna veriga, ki pa poteka v smeri 5' -> 3', pa se podvaja v krajših ločenih Okazakijevih
Slika 14: Človeške telomerne sekvence in podvajanje DNK [11]
15
Človeška telomerna DNK Problem replikacijskega konca
telomere
Linearni kromosom
Ponavljajoča sekvenca TTAGGG in3'-previs
T- zanka
Replikacijske vilice
Sinteza glavnega pramena
Sinteza zaostaja-jočega pramena
Odstranitev RNA začetnika privede do vrzeli
Starševska DNKGlavni pramenZaostajajoči pramenRNK začetnik

fragmentih. Da polimeraza lahko začne prepisovati, potrebuje začetni del, ki se imenuje RNK začetnik. Ko pridejo replikacijske vilice proti koncu kromosoma, ni več osnovne verige, ki bi služila kot matrica za sintezo Okazakijevih fragmentov. To povzroči, da se vsak 5'-konec nove DNA ne more v celoti podvojiti in da ima vsak hčerinski kromosom krajšo telomerno regijo. Telomerne sekvence so za celico nepomembne. Obstajajo zato, ker se genom ob vsaki celični delitvi krajša in ne bi bilo dobro, da bi se krajšal na račun genov, ki kodirajo proteine. 8.1 Povezava med telomeri in rakom Pri vsaki delitvi navadnih celic se torej izgubi del telomer. Čez čas, ko teh telomer zmanjka, se celica ne more več deliti in vstopi v celično apoptozo. Tako imajo normalne somatske celice omejeno podvojevalno obdobje, ki traja približno petdeset delitev. Rakave in tumorske celice pa imajo veliko količino encima telomeraza, ki podaljšuje telomere, tako da jih nikoli ne zmanjka. Zato jim pravimo nesmrtne celice. Znanstveniki si prizadevajo, da bi aktivirali produkcijo telomeraze v navadnih celicah, kar bi preprečevalo bolezni, ki so povezane s staranjem ter inhibirali telomerazo v rakavih celicah, kar bi povzročilo njihovo apoptozo.Zdravila, ki se direktno vežejo na telomere in stabilizirajo DNA strukture kot so G-kvadrupleksi, so hkrati tudi inhibitorji telomeraze in zmotijo telomerno zgradbo.
9 G-žice
V 70. in 80. letih prejšnjega stoletja je bilo doseženo pomembno odkritje in sicer, da se G-kvarteti v prisotnosti različnih alkalnih kationov samouredijo v večje strukture in sicer stabilne dolge 1D agregate, ki so bolj znani pod imenom G-žičke. Gvanin in njegovi derivati so izmed ostalih baz DNK najprimernejši za tvorbo visoko stabilnih molekulskih struktur. G-žice lahko nastanejo iz posameznih gvaninskih nukleotidov ali pa iz enega, dveh ali štirih G-bogatih DNK pramenov. Gvanin se od ostalih baz razlikuje po nizkem ionizacijskem potencialu, zaradi česar ima pomembno vlogo pri električni prevodnosti materialov iz DNK. Prevodnost G-žic omogočajo gibljivi kationi med posameznimi tetradami, ki tvorijo ionski kanal [5].Tvorba G-žičk v razredčenih raztopinah je precej dobro raziskana [12]. G-žicke imajo premer 2.5 nm, kar se sklada s premerom G-kvadrupleksa. Tvorba G-žic v raztopini je odvisna od prisotnosti tipa kationov (K, Na, Mg) in njihove koncentracije. Ko se enkrat žice formirajo, so odporne na denaturacijo. Te strukture se formirajo v minutah oz. urah tudi pri zmernih koncentracijah. Formacija G-žick je odvisna tudi od temperature (hitreje nastanejo pri višjih temperaturah). Manj raziskano je področje samourejenaja z gvanozinom bogatih DNK sekvenc v G-žice na površinah. Glavni namen raziskav je pridobiti ponovljive G-žice in nadzorovati njihove dimenzije.Stabilnost G-žic: pri meritvah z mikroskopom na atomsko silo opazimo, da je višina kvadrupleksne DNK (≈1.2nm) skoraj dvakrat večja kot višina dupleksne DNK (≈0.6nm), kljub temu da je njun premer približno enak (≈2.3nm in ≈2.1nm) . Manjša deformacija
16

G4-DNK v primerjavi z dupleksno DNK je posledica večje odpornosti G4-žic. Možna razlaga za to je, da se pri adsorpciji dupleksna DNK vijačnica razvije in ne zavzema več spiralne oblike. Poleg tega G4-žice sestavljajo štirje prameni, v središču pa se nahaja kation, ki še dodatno stabilizira strukturo (slika 15).
Slika 15: Dupleksna DNK in kvadrupleksna DNK na sljudi [12]
Na oddelku za kompleksne snovi inštituta J. Stefan raziskujemo tvorbo G4 žic na vzorcih z različnimi oligonukleotidnimi sekvencami. S pufrom razredčeni vzorci (20µM) so bili nanešeni na sveže razcepljeno sljudo. Površina razcepljene sljude vsebuje plast kalijevih ionov, ki vplivajo na rast G-žic. S spiranjem sljude z MgCl2 je doseženo, da se učinek kalijevih ionov zmanjša. Strukturo analiziramo z mikroskopom na atomsko silo [14]. Na slikah 16 in 17 so prikazane meritve na sekvenci GCGGAGGCG.
Substrat: razcepljena sljudaa) b)
Slika 16: a) tvorba dolgih G-žic; b) G-zanke in krajše G-žice [15]
17

Substrat: razcepljena sljuda sprana z MgCl2 in H2O
Slika 17: Zamenjava ionov povzroči večjo tvorbo G-žic z ožjo porazdelitvijo dolžine žic. G-žice so visoke približno 1.7 nm v kvadrupleksih, stabiliziranih z Na+ ioni in 2.2 nm v kvadrupleksih, stabiliziranih z K+ in Mg+ ioni. Višina žic kaže na to, da G-kvadrupleksi ležijo vodoravno na površini [15].
10 Zaključek
V seminarju sem predstavila nekaj osnovnih značilnosti kvadrupleksnih struktur DNK. Najbolj pomembna vloga kvadrupleksov je prav gotovo povezana z njihovim posredovanjem pri ključnih procesih znotraj celic, še posebno tistih povezanih s telomerno strukturo in funkcijo. Telomeraza je ključna tarča proti raku. Razvoj kvadrupleksnih zdravil za zaviranje telomeraze se bo prav gotovo nadaljeval. G-kvadrupleksi pa so zanimivi tudi iz drugih vidikov. Z uporabo nekovalentnih interakcij lahko pridobimo uporabne supermolekularne strukture, ki predstavljajo pomembno vlogo pri razvoju senzorjev in nanomaterialov. S kombinacijo navdiha iz narave in človeške domišljije bodo znanstveniki na področju kvadrupleksov nedvomno odkrili še marsikaj zanimivega.
18

Literatura
[1] Stephen Neidle, Shankar Balasubramanian, Quadruplex Nucleic Acids, The Royal Society of Chemistry, 2006
[2] Christopher A. Hunter, Jeremy K. M. Sanders, The Nature of π -π Interactions, 1990
[3] Ivan Filipovič, Stjepan Lipanovič, Opča I anorganska kemija, Zagreb, 1991
[4] J. M. Ducere, L. Cavallo, Parametrization of an Empirical Correction Term to Density Functional Theory for an Accurate Description of π – Stacking Interactions in Nucleic Acids, J. Phys. Chem. B, 2007
[5] M. S. Kaucher, W. A. Harrell, J. T. Davis, A Unimolecular G-Quadruplex that Functions as a Synthetic Transmembrane Na+ Transporter, J. Am. Chem. Soc, 2006
[6] http://www.nature.com/nrc/journal/v2/n3/abs/nrc749.html (10. 3. 2010)
[7] http://www.fiz.uni-lj.si/~tine/fizikaII.html (10. 3. 2010)
[8] http://www.chem.umass.edu/people/botch/Chem121F06/Chapters/Ch16 (10. 3. 2010)
[9] http://nobelprize.org/nobel_prizes/medicine/laureates/2009 (10. 3. 2010)
[10] V. A. Bloomfield, D. M. Crothers, I. Tinoco Jr., Nucleic Acids, University Science Books, 2000
[11] http://novebiologije.wikia.com/wiki/Telomeri (10. 3. 2010)
[12] Thomas C. Marsh, James Vesenka, Eric Henderson, A new DNA nanostructure, the G- wire, imaged by scanning probe microscopy, Nucleic Acid Research, 1995
[13] http://en.wikipedia.org/wiki/Directionality%28molecular_biology%29 (10. 3. 2010)
[14] K. Kunstelj, F. Federiconi, L. Spindler, I. Drevenšek-Olenik, Self-organization of guanosine 5'- monophosphate on mica, Science Direct, 2007
[15] Slike z inštituta J. Stefan
19


















