
マイクロサテライトDNAマーカーによるサクラマス...
8
マイクロサテライトDNAマーカーによるサクラマスの遺伝的多 様性と集団の遺伝的分化 誌名 誌名 水産増殖 = The aquiculture ISSN ISSN 03714217 巻/号 巻/号 554 掲載ページ 掲載ページ p. 521-527 発行年月 発行年月 2007年12月 農林水産省 農林水産技術会議事務局筑波産学連携支援センター Tsukuba Business-Academia Cooperation Support Center, Agriculture, Forestry and Fisheries Research Council Secretariat
Transcript of マイクロサテライトDNAマーカーによるサクラマス...

マイクロサテライトDNAマーカーによるサクラマスの遺伝的多様性と集団の遺伝的分化
誌名誌名 水産増殖 = The aquiculture
ISSNISSN 03714217
巻/号巻/号 554
掲載ページ掲載ページ p. 521-527
発行年月発行年月 2007年12月
農林水産省 農林水産技術会議事務局筑波産学連携支援センターTsukuba Business-Academia Cooperation Support Center, Agriculture, Forestry and Fisheries Research CouncilSecretariat

水産増殖 (AquacultureSci.) 55 (4), 521 -527 (2007)
マイクロサテライト DNAマーカーによる
サクラマスの遺伝的多様性と集団の遺伝的分化
野口大毅1,2.谷口順彦1,2
Studies on the Genetic Diversity of Wild Populations of Masu Salmon, Oncorhynchus mαsou masou, by Microsatellite D NA Markers
Daiki NOGUCHI1,2 and Nobuhiko TANIGUCHI1,2
Abstract: A large number of hatchery masu salmon, Oncorhynchus masou masou, have been
released into the rivers in ]apan in order to enhance the fisheries resources of this species.
Unconscious genetic changes that may occur during artificial seed production should be prevented
江weare to perform effective and responsible resource enhancement. In this pape乙thegenetic
variabi1ity and population structure of masu sa1mon were estimated using five microsate1lite DNA
markers (msDNA). Masu salmon showed a high level of genetic variabi1ity at the msDNA loci.
Average allele numbers ranged between 10.40 and 15.00, average numbers of allele richness
between 9.74 and 11.72, and expected heterozygosities between 0.824 and 0.874. Genetic
differentiation was not observed between different year c1asses. Meanwhile, genetic differentiation
was detected among captured locations, but distinct correlation between genetic distance and
geographic distance was not observed. Finally, methods of brood stock management in artificial
propagation to prevent genetic change and loss of variation for conservation of genetic diversity in
wild populations are discussed.
Key words: Oncorhynchus masou; Microsate11ite DNA; Genetic variabi1ity; Population structure
降海型サクラマスは重要な漁業資源であるが,近年
漁獲量が低下してきているため,大規模な種苗放流事
業が進められている(鈴木ら 2000;さけ ・ます資源管
理センター 2005)。産卵親魚の回帰数の減少と少数親
魚からの種苗の作出は,種苗集団の遺伝的多様性の低
下をまねき,そのような種苗集団の放流は野生集団の
遺伝的構成に何らかの影響をもたらすこ とが予測され
るO このため,遺伝的多様性を的確な方法で評価し,
経年的に追跡調査をする必要がある(谷口 1999)。
サクラマスは,母川回帰性が強く,河川毎に相対的
な独立性を備える繁殖集団が形成され易いことが予測
2007年3月16日受付 :2007年8月30日受理.
される。Okazaki(1986)は,アイソザイム分析を行
うことによって,河川聞に独立性が見られ, 日本海側
グループと太平洋側グループの 2つに分けられること
を示唆した。これに対し,鈴木ら (2000)は,ミ トコ
ンドリア DNAD-Iρop領域の RFLP分析により,河
川聞の分化は小さく地理的なグループ構造を示してい
ないと報告している。一方,Edpalina et a1.は, DNA
D-Loop領域の塩基配列比較を行い, 日本海側グルー
プと太平洋側グループの 2つに分かれるこ とを示唆
し, Okazaki (1986)の結果を支持した *1。これらの
研究では,使用したマーカーの感度(変異性),採集
I東北大学大学院農学研究科 (GraduateSchool of Agricultural Science, Tohoku University, Sendai, Miyagi 981-8555, ]apan). 2現所属 .福 山大学内海生物資源研究所(百leResearch Institute of Marine Bioresources, Fukuyama University,
Onomichi, Hiroshima 722-2101, ]apan) . *1 Edpalina, R.R., S. Sato, S. Urawa, V. Brykov, A. Urano and S. Abe (2005) Genetic variation and population structure of masu sa¥mon based on mitochondrial DNA sequence analysis.平成17年度日本水産学会大会.

522 野口・谷口
した標本集団などが異なるため,集団構造に関して異
なった結論が得られた可能性が考えられる。本研究で
は,十分な感度を備え,集団の独立性,異集団の混合
に関する評価判定が可能である核 DNAのマイクロサ
テライト領域多型を採用し,サクラマスの分集団構造
を解明することを試みた。
サクラマスと同様に母川回帰する魚種では,カラフ
トマス等で偶数年級群と奇数年級群で遺伝的分化が観
察されており (AspinwaI11974),このような年級群間
分化が存在している場合は,地域個体群のみならず,
年級群も保全の対象として考慮に入れなくてはならな
い。サクラマスの標本採集に関しては,年級群聞の遺
伝的分化がおきている可能性に配慮して標本採集を進
めた。
以上の観点から,本研究では,マイクロサテライト
DNAマーカーを利用して 1)集団内の遺伝的多様
性の レベルの評価, 2)漁獲年級群聞の遺伝的分化の
有無, 3)採集水域聞の分化, の3点を明らかにする
ことを目的とした。
Shokanbetsu River(SK)
Ara rive(AR) Miomote River (MM)
材料および方法
供試魚
富山県神通川 (2001年, 2002年),新潟県魚野川,
荒川, 三面川,北海道尻別川,暑寒別川,標津川,
岩手県安家川 (以上2001年),宮城県気仙沼沖 (2003
年) の 9 水域 (10標本集団) でそれぞれ17~48個体を
採集し,計333個体を標本として供した (Fig.1)。こ
のうち,神通川, 三面川,暑寒別川, 魚野川, 荒川,
安家川の標本は,河川へ遡上してきた降海型サクラマ
スである。標津川,尻別川の標本は,河川残留型サク
ラマスである。宮城県気仙沼沖の標本は,海域で漁獲
され市場に流通していた降海型サクラマスである。
遺伝マーカー
遺伝マーカーとして, 0ぉ520,OmaOl, Oma02,
Oma3ke, Oma4myの5マイクロサテライト DNAマー
カ一 (野口ら 2003) を用いた。DNAの抽出,およ
-}!
Shibetsu River(SB)
Akka River(AK)
4一一一KesennumaOffshore(KS)
Fig. 1. Sampling locations of the masu salmon, Oncorhynchus masou masou. JZ2001 and JZ2002 (36045'N 137013'E),
UN (37056'N 139003'E), AR (38009'N 139024'E), MM (38014'N 139026'E), SR (42052'N 140021'E), SK (43051'N
141030'E), SB (43040'N 145007'E), AK (40003'N 141051'E), KS (38051'N 141048'E).

サクラマスの遺伝的多様性と集団の分化 523
び,マイクロサテラ イト 座の PCR反応は,野口ら
(2003)に従った。増幅断片の検出およびア リルサイ
ズの決定には,ABI PRISMR 377 DNA Sequencerおよ
び377GeneScanTM (Applied Biosystems製)を用い
た。
遺伝的多様性と集団の分化
遺伝的多様性を測る指標として,ア リル数 (NA),
アリルリ ッチネス (AR),各マーカーにおけるヘテロ
接合度の期待値 (he) と観察値 (ho),平均ヘテロ接
合度の期待値 (He) と観察値 (Ho),ハーデイーワイ
ンベルグ平衡からの逸脱に関する 界valueを求めた。
集団聞に遺伝的分化があるか否かを調べるために,
各標本集団聞の F'5['を求め AMOVA分析 (Weirand
Cockerham 1984; Excoffier et al. 1992) を行こった。ま
た,河口聞の海上における最短距離を計測し,遺伝
的距離 (F':{r)と地理的距離に相関があるかを調べる
ため,相関係数の算出および Mantel検定を行った。
魚野川の地理的位置は本流である信濃川の河口で代用
した。さらに,集団に構造があるか否かを検討するた
めに,Cavalli-Sforza and Edwardsの遺伝距離(Cavalli-
Sforza and Edwards 1967)を求め,UPGMA法と NJ
法 (Saitouand Nei 1987)によ ってデンドログラムを
作成した。なお,遺伝距離の算出とデンドログラムの
作成の際は, 1000回のブー トス トラップを行った。
凡4とARは,コンピューターソフ トウェアFstatver.
2.9.3 (Goudet 2001)を用いて算出したo he, ho,
He,' Ho, 1主value,AMOVA分析各標本集団聞の F訂
の算出には,コンピューターソフ トウェア, Arlequin
ver. 3.0 (Excoffier 2005)を使用した。遺伝距離と
NJ法は,コンピューターソフ トウェアPopulations
ver.1.2.29 (Langella 1999)によ って実行した。デンド
ログラムの表示は,TreeExplore (Tamura 1999)を
使用した。
結果
遺伝的多様性
アリル数 (NA),アリ Jレリ ッチネス (AR),各マー
カーにおけるヘテロ接合度の期待値 (he) と観察
値 (ho),平均ヘテロ接合度の期待値 (He) と観察値
(Ho),ハーディーワインベルグ平衡からの逸脱に関
する P-valueをTable1に示した。各地点における,ア
リル数の平均値 (5マーカ一座)は,三面川標本集団
と安家川標本集団の10.40から気仙沼標本集団の15.00
までの間にあり ,他魚種に比べても平均的な値を示し
た (Taniguchiet al. 2003)。アリル リッチネスの平均
値 (5マーカ一座)は安家川標本集団の9.74から荒川
標本集団の11.72までの値を示し,水域間でのア リル
リッチネスの値の差はそれほど大きいものではなかっ
た。平均ヘテロ接合度の期待値 (He)(5マーカ 一座)
は気仙沼沖標本集団の0.824から荒川標本集団の0.874
の間にあり, 他魚種に比べても比較的高い値であった
(Taniguchi et al. 2003)。ハーディーワインベルグ平衡
に関しては,荒川標本集団は,全てのマーカ一座に
おいて逸脱は確認されず,それ以外の標本集団では,
1 ~ 2 マーカ一座でハーデイーワ イ ンベルグ平衡から
の逸脱が観察された。 しかし,全てのマーカ一座で逸
脱している集団はなかった。
集団の分化
AMOVA分析を行ったところ,全標本集団の変異量
のうち,集団間の変異は3.18%を占め,有意な値を示
した (P= 0.00)。各標本集団聞の地理的距離と遺伝的
距離 (F,釘、)を求めたところ,Table 2および Fig.2の
値を示した。神通川における2001年度と2002年度の
標本集団間では異質性が観察されなかったが,それ
以外の全ての組み合わせでは異質性が観察された。水
域間の F'5['値は,0.015から0.060までの値を示した。
水域聞の地理的距離は,本ナ|‘旧 本海側群,本州太平
洋側群, 北海道日本海側群と分けたとき,それぞれの
群内の水域間の距離は, 10.72 kmから256.71kmであ
り,異なる群に属する水域聞の距離は555.84kmから
1191.79 kmであった。遺伝的距離 (F'5[') と地理的距
離の 2変数で Mantel検定を行ったところ有意な相聞
が検出され (P= 0.00),相関係数は0.588を示した。
また,地理的距離は,256.71 km以下の群と555.84km
以上の群に分かれたため,両群を,遺伝的距離 (F.訂)
の値によ って Mann-WhitneyのU検定を行ったとこ
ろ,有意な違いが観察された (P= 0.00)。それぞれ
の群内で遺伝的距離 (F'5[') と地理的距離の 2変数で
相関係数を求め Mantel検定を行った。256.71km以
下の群では,相関係数は-0.615, Mantel検定では,
0.05の危険率で相関は観察されなかった (P=0.975)。
555.84 km以上の群では,相関係数は0.381,0.05の危
険率で相闘が観察された (P= 0.014) 0 Cavalli-Sforza
and Edwardsの遺伝距離を求め,UPGMA法と NJ法
とによ ってデンドログラムを作成したところ, Fig.3
の樹形を示した。異質性が観察されなかった神通川の
2001年度と2002年度の標本群は, UPGMA法と NJ法
で共にクラスターを形成し,ブー トス トラップ値も極
めて大きく,信頼性が高かった。しかし,それ以外の
地点では, UPGMA法と NJ法で共通のクラスターは
なく ,ブー トス トラップ値も小さく ,それぞれのクラ
スターの再現性が低かった。
524 野口・ 谷口
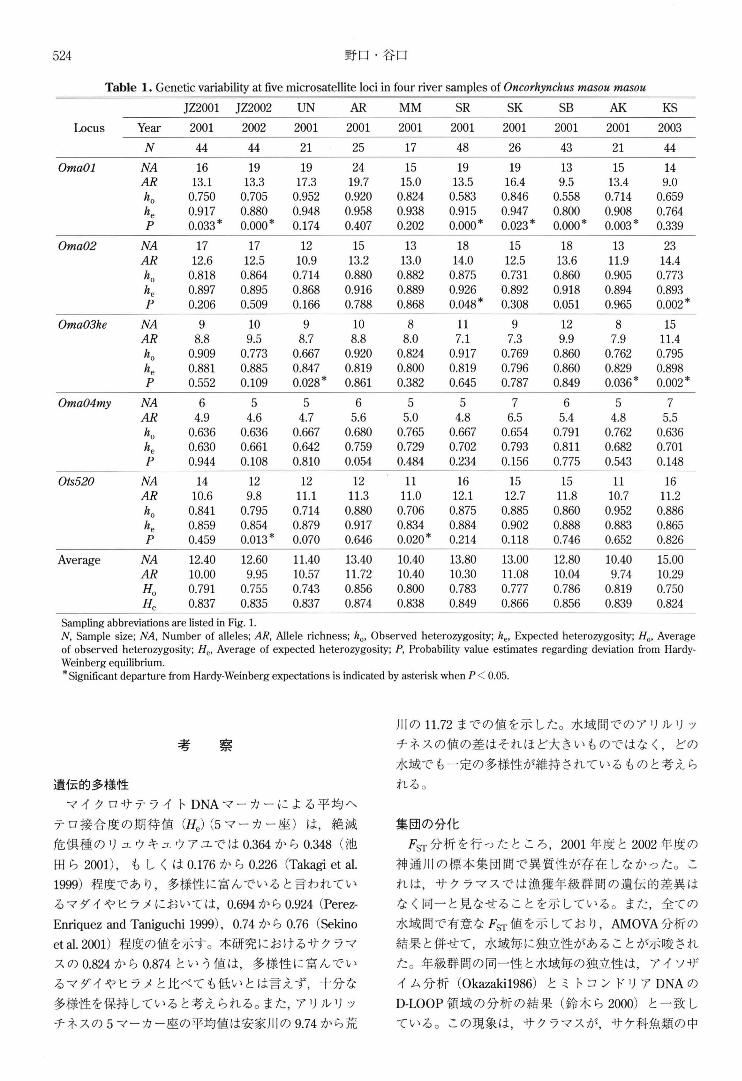
Table 1. Genetic variability at :five microsatellite ¥oci in four river samp¥es of Oncorhynchus masou masou
1ρcus
OmaOl
加
一N一M
M九九P一M
M九九P一M
H九九P一M
M九九P一M
H九九P一MMH04
Oma02
Oma03ke
Oma04my
Ots520
Average
JZ2001 JZ2002 UN
*
1A-
itnU7sつd一
、】且U司tnhv一
Qd1A9-FbnUAUZ
、31iQdQd一nunU1A司i
加一HH
一日
配
周
到
瓜
一口
引
訓
mm一9MM別
∞
m周一6ωmmM一UH山
Mum部
品
一μωnm
n4一
1JAUAUnu一
可JAUAUAり一
日UAUnununUAU-
-JAUAりnu一司i唱AAUAU
*
也
卒
、
己
》
》
FHUAUAU-3刈
AτFhun汐一
内JRUQU一
nhv唱inxu一
Fhda宮内J-nUFHUFHUFhd
側一叫
一回
収
川
路
加
一日
山
崩
mm別
一mm刀∞
mM一5ω泊
節
目
一ロ
mmmmm一ωω万
mm
m4
1JAVAVAV-JAUnUAU一
AVAunu一
ハununu一
AUAUAU一唱Anunu
則一目
一mm能
川
一ロ
mm出
一9幻
路
側
一5
U閉
山
山
一ロ
山
mmm一
MMm附
一1
7
0
8
7
一
J
0
6
8
一
0
9
1
0
9
4
一
1
0
7
6
一0
2
6
4
組
一∞
一お
一MVU白
目
的
一日
山
部
旧
市
一mMm別
加
一6M槌
拓
郎
一U
U槌
訓
M一uumu
」
一2
1
&
a
o
-
-仏ハ札口比
一
十
o
o
o
-
-
o
a仏
一
1仏
0
0一工
1
0
0
*
則一則
一口
一目
印
刷
叩
仰
一日
削
減
問
一8
M邸
側
一5日
間
問
叩
一日
山
印
刷
問
一MM州
側
*
*
t
A
-
〉。drDAU一
、.JFD£UOO一
司iQdEd一
円iヮ“
AUz-
-LPb泊品
ZA守一AUAりつdQd
涼
一肌
一必
一日即日品川訓加
一日則的訓
mmM一日
日
別
訓
Mm一5川
日以
mmn一日
山
Mωmm一日
ω河川訓
-n41Jnununu-
-JnUAUAU一
ハunUAU一
ハUnunu-
-JnUAVAU--itムハUAU
*
一
一1
-
l
6
7
3
一
-3128一
9
6
7
一
4
3
6
一
7
5
2
8
一0
8
7
6
E一
O一6一9
J
4
4
2
一5
1
3
9
0
一
3
6
9
8
一
5
5
9
5
一5
J
8
0
1
一0
0
7
6
g一ゆ
一2
一1
凶
89AM--
ロ1
8
3
一9
1
1
1
1
一7氏弘
β
y
l
一1円凶
Q』
Qadi
-a
L
y
ob
nJ,“一
1Jnununu-
-JAuhunU一
ハUAUAV一
向UAUAU-
-JAUAVAV-'A1iAunv
*
1一
8
0
0
一
、3
O
RUI
n
v
0
9
一
1
1
5
一
、3
0
8
6一0
4
6
6
3J一一一一
つd一qdRURdハunu一白U4、氏U14Ed一9bQd氏U氏ud--、3A生
QJ1ムヴd一巳d《,A£U良UAT一良unuau巳d
qd一ゆ
一4
一1Q458AM--β059AM--
Q山
8
8
8
一E
5
7
8
Y
一ーは
oA8y一Z
O
Y
8
-nd
h
U
A
U
A
U
一
1JAUAUAU一
nunUAU-
nununu-
-JハUnunU一唱i唱inUAU
民
一則
一目
一日
品
協
一日
ω邸
側
一8日
間
協
一5品
出
一日
町
邸
側
一沈
滞
*
*
一吋3一
QUATQd一
自室内JqJヮ“
itFDQUヮ“
一
戸01AOO一
〉dpOFDPO一AVQdnVA坐
円b一γ山一4合一4含ハUFbpoqd一3J,A74QdAU--、Jdし
QdQdnu一F
Fりつdova--nD,JQUρbヮ“
一ハUつム「Dヮ“
K一伽
一必
一L
弘
6
7
3
一Z
4
7
8
0
一山
1
7
8
0
一7
広
6
7
1
一山
1
8
8
8
一広
臥
7
8
114一2一
!
o
o
o
-
-
o
o
o
-
-
o
o
o
-
-4
0
o
o-
-
0
0
0
一b
b
o
o
Sampling abbreviations are listed in Fig. 1. N, Sample size; NA, Number of alleles; AR, Allele richness; ho' Observed heterozygosity; he, Expected heterozygosity; Ho, Average of observed heterozygosity; He, Average of expected heterozygosity; P, Probability value estimates regarding deviation from Hardy-Weinberg equilibrium. * Significant departure from Hardy-Weinberg expectations is indicated by asterisk when P < 0.05
考 察
遺伝的多様性
マイク ロサテラ イト DNAマーカ ーによる平均ヘ
テロ接合度の期待値 (He)(5マーカ一座) は,絶滅
危倶種の リュウキュウアユでは0.364から 0.348(池
田ら 2001),も しくは 0.176から 0.226(Takagi et al.
1999)程度であり ,多様性に富んでいると言われてい
るマダイやヒラメにおいては,0.694から 0.924(Perez-
Enriquez and Taniguchi 1999), 0.74から 0.76(Sekino
et al. 2001)程度の値を示す。本研究におけるサクラマ
スの 0.824から 0.874という値は,多様性に富んでい
るマダイやヒ ラメと比べても低いとは言えず,十分な
多様性を保持していると考えられる。また,ア リルリ ッ
チネスの 5マーカ一座の平均値は安家川の9.74から荒
川の 11.72までの値を示した。水域間でのア リルリ ッ
チネスの値の差はそれほど大きいものではなく,どの
水域で、も一定の多様性が維持されているものと考えら
れる。
集団の分化
F':IT分析を行ったところ, 2001年度と 2002年度の
神通川の標本集団間で異質性が存在しなかった。こ
れは,サクラマスでは漁獲年級群問の遺伝的差異は
なく 同ーと見なせることを示している。また,全ての
水域間で、有意な F':IT値を示しており, AMOVA分析の
結果と併せて,水域毎に独立性があることが示唆され
た。年級群聞の同一性と水域毎の独立性は,アイソザ
イム分析(Okazaki1986) とミ トコン ドリ アDNAの
D-LOOP領域の分析の結果(鈴木ら 2000)と一致し
ている。この現象は, サクラマスが,サケ科魚類の中

サクラマスの遺伝的多様性と集団の分化 525
Table 2. Pairwise comparison of Fsr values (lower) and geographical distances (upper) between samples
JZ2001 JZ2002 UN AR MM SR SK SB AK KS
JZ2001 0.00 210.41 247.80 256.71 758.79 901.07 1191.79 888.87 1048.05
JZ2002 0.002 210.41 247.80 256.71 758.79 901.07 1191.79 888.87 1048.05
UN 0.020* 0.027* 37.66 46.89 588.47 730.56 1021.21 718.31 877.43
AR 0.015* 0.023キ 0.032* 10.72 562.67 705.26 996.18 692.73 852.44
MM 0.016キ 0.021 * 0.023* 0.026* 555.84 698.07 988.98 685.91 845.16
SR 0.019* 0.026* 0.031本 0.022* 0.031 * 169.26 736.45 591.94 751.58
SK 0.031 * 0.043 * 0.040* 0.024 * 0.038* 0.022* 606.81 734.60 893.79
SB 0.045 * 0.057* 0.052* 0.043* 0.047* 0.035* 0.034 * 568.82 694.79
AK 0.027* 0.028* 0.045* 0.033 * 0.025* 0.030* 0.047* 0.060* 171.65
KS 0.030* 0.037* 0.042* 0.039* 0.026* 0.031 * 0.041 * 0.029* 0.031 *
Sampling abbreviations are listed in Fig. 1. * Significant differentiation is indicated by asterisk when P < 0.01.
0.08
0.06十 • • • • • • 圃 • '‘ . • • • • • • ..・• • • .・ • , r=一0.615,p= 0.975 (・) r = 0.381, P= 0.014 (・)
0.00
O 200 400 600 800 1000 1200
Geographical Distance (km)
Fig. 2. Relationship between Fsr and geographical distance. Solid square: The geographical distances ranged from 0.00 to
256.71km, Solid circle:τbe geographical distances ranged企om555.84 to 1191.79 km.
でも特に強い母川回帰性を有しているため,水域毎に
強い遺伝的独立性を備えたことが示唆される。
地理的な距離が 256.71km以下の群と 555.84km以上
の群,それぞれの群内で遺伝的距離 (Fsr)と地理的
距離の 2変数の相関を求めたところ, 555.84km以上
の群では正の相闘が観察された。256.71km以下の群
では,負の相関係数を示しその絶対値は 555.84km以
上の群より大きかったが, Mantel検定では有意な相
闘が観察されなかった。また, 256.71krn以下の群と
555.84km以上の両群において遺伝的距離 (Fsr)の値
によ ってMann-WhitneyのU検定を行ったところ,有
意な違いが観察された。これは,海峡をまたぐような
長距離の回遊においては 海峡をまたがない回遊に比
べ迷入が少ないことを示唆している可能性がある。
UPGMA法と NJ法でデンドログラムを描いたとこ
ろ,異質性が観察されなかった2001年度と2002年度の
標本群は,両方法で同ーのクラスターを形成し,ブー
トストラップ値も極めて大きかったが,それ以外では,
両方法で樹形が異なっており,両方法ともにブートス
トラップ値が小さくクラスターの信頼性は低かった。
従って,それぞれのグループ性に関しては系統的意味
を見出すことは出来ないと判断された。
地理的な距離と遺伝的距離およびクラスターで一様
な相聞がない原因として,以下の四点が考えられた。
一つ目は,異なる河川に由来する種苗が放流され,遺
伝的撹乱がおきている可能性があること。二つ目は,
近年集団サイズが急激に減少して小集団化し,遺伝的
浮動による無方向への分化が加速している可能性があ
ること。三つ目は,現在でも一定の遺伝子交流がおき
ており,分集団問の遺伝的分化が中断したため分化の

526
a)
野口 ・谷口
AK
SK
SR
0.05 SB
JZ2001 100
JZ2002
AR MM SR
UN
AK SB
KS
SK
0.05
Fig. 3. Unrooted neighbor-joining dendrogram(a) and UPGMA dendrogram(b), both of them based on Cavalli-Sforza's chord distance, estimated from microsatellite allele frequencies(percentage value refers to the bootstrap consensus value after 1000 replications).
程度が小さくなった可能性があること。四つ目は,サ
クラマスの回遊経路が地理的な最短距離と異なってい
る可能性があることである。
今後,サクラマスの栽培漁業を継続していくにあた
り,現在の集団構造と遺伝的多様性を維持していくこ
とが重要である。各水域ごとに分化がおきていること
から,対象河川に回帰した親魚から種苗を生産して放
流することにより,地域集団の遺伝的撹乱を防がなく
てはならない。また,各河川で種苗を作出する際,十
分な親魚数を確保することにより遺伝的多様性の低下
を防ぎ,遺伝資源を保全する必要がある。
要約
日本列島周辺の水域に生息する 9水域10標本集団
のサクラマスを,マイクロサテライ トDNAマーカー
5座を用いて集団遺伝学的解析を行った。その結果,
日本列島周辺に生息するサクラマス群は,十分な多
様性を保持していることが明らかになった。また,同
一水域で、の漁獲年級群聞には異質性が検知されなかっ
たが,異なる水域間では集団の分化が観察された。一
方,これらの分化には,地理的な距離との一様な相聞
は観察されなかった。今後,種苗放流を行う際には,
それぞれの河川を管理単位とすることが妥当と考えら
れた。
謝辞
本研究を行うにあたり, 工藤飛雄馬 (岩手県内水面
水産技術センター),佐藤 将 (新潟県内水面水産試

サクラマスの遺伝的多様性と集団の分化 527
験場),田子泰彦(富山県水産試験場),安藤大成(北
海道立水産僻化場),市村政樹 (標津サーモン科学館),
伊藤 博(宮城県気仙沼水産試験場)の各氏から, サ
ンプルの御提供を頂いた。また,各関係漁業組合の方々
の御協力を頂いた。ここに記して御礼申し上げる。
文 献
Aspinwall, N. (1974) Genetic examination of North American
populations of the pink salmon, Oncorhynchus gorbuscha,
possible evidence for the neutral mutation random dri丘
hypothesis. Evolution, 28, 295-305.
Cavalli引 orza,L. L. and A. W. Edwards (1967) Phylogenetic
analysis. Models and estimation procedures. Am. ]. Hum.
Genet., 19, Suppl19, 233+.
Excoffier, L., P. E. Smouse and]. M. Quattro (1992) Analysis of
molecular variance inferred from metric distances among
DNA haplotypes: application to human rnitochondrial DNA
restriction data. Genetics, 131,479-491.
Excoffier, L., G. Laval and S. Schneider (2005) Arlequin ver. 3.0:
An integrated software package for population genetics
data analysis. Evolutio仰 ryBioinformatics Online, 1,47-50.
Goudet,]. (2001) FSTAT, a program to estimate and test gene
diversities and fixation indices (version 2.9.3). http:/ /
www.unil.ch/izea/so立wares/fstat.h凶11.
池田 実・立原一憲 ・誠布川・ 谷口l順彦 (2001)マイク
ロサテライト DNAおよびミ トコ ンド リアDNAPCR-
RFLP分析によるリュウキュウアユ導入集団の遺伝的評
価.水産育種, 31, 33-37.
Langella, O. (1999) Populations, 1.2.29. http://www.cnrs-gif.fr/
pge/bioinfo/ debian/.
野口大毅・池田 実・中島正道 ・谷口}11ii:彦 (2003)サクラマ
ス集団解析のためのマイクロサテライ トDNAマーカー
の開発とその特性について.水産育種, 33, 61-66.
Okaz心a,T (1986) Genetic variation and population structure in
masu salmon, Oncorhynchus masou of ]apan. Bull. Japa.仰.
Soc. Sci. Fish., 52, 1365-1376
Perez-Enriquez, R. and N. Taniguchi (1999) Genetic structure
of red sea bream (Pagrus major) population of ]apan and
the southwest Pacific using microsatellite DNA markers.
Fisheries Science, 65, 23-30.
Saitou, N. and M. Nei (1987) The neighbor-joining method: a
new method for reconstructing phylogenetic trees. Mol.
Biol. Evol., 4, 406-425
さけ ・ます資源管理センター (2005)道府県別の事業実績
http・//www.salmon.affrc.go.jp/zousyoku/result/result.htm.
Sekino, M. and M. Hara (2001) Application of microsatellite
markers to population genetics studies of]apanese flounder
Paralichthys olivaceus. Mar. Biotechnol., 3, 572-589.
鈴木研一 ・小林敬典 ・松石 陵 ・沼知健一 (2000)ミトコ
ンド リアDNAの制限酵素切断型多型解析から見た北海
道内におけるサクラマスの遺伝的変異性.日水誌,66,
639-646.
Takagi, M., E. Shoji and N. Taniguchi (1999) Microsatellite
DNA polymorphism to reveal genetic divergence in Ayu,
Plecoglossus altivelis. Fisheries Science, 65, 507-512
Tamura, K. (1999) TreeExplore, Version 2.12. Metro University,
Tokyo, http://evolgen.biol.me仕o-u.ac.jp/TE/
谷口順彦(1999)魚介類の遺伝的多様性とその評価法.海
洋と生物, 123, 28Cト289.
Taniguchi, N, R. Perez-Enriquez and N. Estu (2003) DNA
markers as a tool for genetic management of broodstock
for aquaculture. In “Aquatic Genornics" (ed. By N. Shirnizu,
T Aoki, 1. Hirono and F. Takashima), Springer-Verlag,
Tokyo,pp.41下429.
Weir, B. S. and C. C. Cockerham (1984) Estimating F-statistics
for the analysis of population structure. Evolution 38,
1358-1370.


















