
骨格筋における虚血・再濯流障害 - JSVS · 2019. 7. 22. · 1992年10月 XOH :iantinn...
5
骨格筋における虚血・再濯流障害 一特に活性酸素産生源について 根本 日普 森瀬 昌樹 杉野三千男 今井 潔 金城 正佳 島津 盛一 柿田 章 索引用語:虚血・再灌流障害,活性酸素,顆粒球 骨格筋の再潅流に際して,活性酸素が関与すること はすでに発表してきたが,間接的証明にとどまらず, 活性酸素の産生源を直接明らかにすることが,再濯流 障害を解明する上で必要と考え,実験的に検討した. 対象および方法 北里大学医学部外科 〒228 相模原市北里卜15-1 受付:1992年8月10日 35 要 旨:骨格筋の虚血・再濯流時の活性酸素産生源については,まだ十分明らかではな い.これを解明することが,再濯流障害を明らかにする基本となるので実験的に検討を加 えた. ラットの薄筋を用い,動脈遮断は60分とし,再潅流後120分までCLA依存化学発光強 度を,虚血・再濯流群,抗好中球抗体投与群, liposomal SOD十カタラーゼ投与群, PAF (血小板活性化因子)桔抗剤投与群について測定した.また,イヌ薄筋を用い,6時間虚 血,再潅流120分まで,虚血・再濯流群,抗好中球抗体投与群, PAF措抗剤投与群につい て,血中ミオグロビン値,各種筋逸脱酵素値を測定した. ラット虚血・再濯流群では,筋への好中球浸潤を認め,特異的発光,すなわちoiの上昇 は再濯流後45分でピークに達し,95分で非特異的発光強度レベルに戻った.これに対し て,虚血前抗好中球抗体投与群では,再潅流時のOiの産生はほとんど認められず, liposomal SOD十カタラーゼ投与群でも再濯流時の相対的発光強度は,非特異的発光強度 と有意差は認められなかった.これはPAF措抗剤投与群でもほぼ同様であった.またイヌ では抗好中球抗体およびPAF措抗剤投与群で虚血・再漕流後のミオグロビン,各種逸脱酵 素値の上昇抑制が認められた. 以上より,骨格筋における虚血・再濯流障害時の活性酸素産生源としては,穎粒球にお けるNADPH oxidase系が強く示唆された.(日血外会誌1 : 35-39, 1992) るcaudal femomal arteryおよびveinのみを残した.動 脈遮断は60分とし,再濯流後120分まで高橋,中野ら の方法に準じてウミホクル・ルシフェリン誘導体であ るCLA (2-melhyl-6-phenyl-3,7-dihydroimidazo[1,2 -a]pyrazin-3-one)に依存する化学発光強度を測定し た.なお, photoraultiplierおよびsingletphoton count- Wistar系雄性ラットの薄筋を用い,これに出入りす ing apparatusは,フランスCherpentier社製およびス ウェーデンA. G. Woltemett社製のを用いた.また,遮 断前抗好中球抗体投与群, liposomal SOD とカタラー ゼ投与群および血小板活性化因子(以下PAF)の措抗
Transcript of 骨格筋における虚血・再濯流障害 - JSVS · 2019. 7. 22. · 1992年10月 XOH :iantinn...

骨格筋における虚血・再濯流障害
一特に活性酸素産生源について
根本 日普 森瀬 昌樹 杉野三千男 今井 潔
金城 正佳 島津 盛一 柿田 章
索引用語:虚血・再灌流障害,活性酸素,顆粒球
骨格筋の再潅流に際して,活性酸素が関与すること
はすでに発表してきたが,間接的証明にとどまらず,
活性酸素の産生源を直接明らかにすることが,再濯流
障害を解明する上で必要と考え,実験的に検討した.
対象および方法
北里大学医学部外科
〒228 相模原市北里卜15-1
受付:1992年8月10日
35
要 旨:骨格筋の虚血・再濯流時の活性酸素産生源については,まだ十分明らかではな
い.これを解明することが,再濯流障害を明らかにする基本となるので実験的に検討を加
えた.
ラットの薄筋を用い,動脈遮断は60分とし,再潅流後120分までCLA依存化学発光強
度を,虚血・再濯流群,抗好中球抗体投与群, liposomal SOD十カタラーゼ投与群, PAF
(血小板活性化因子)桔抗剤投与群について測定した.また,イヌ薄筋を用い,6時間虚
血,再潅流120分まで,虚血・再濯流群,抗好中球抗体投与群, PAF措抗剤投与群につい
て,血中ミオグロビン値,各種筋逸脱酵素値を測定した.
ラット虚血・再濯流群では,筋への好中球浸潤を認め,特異的発光,すなわちoiの上昇
は再濯流後45分でピークに達し,95分で非特異的発光強度レベルに戻った.これに対し
て,虚血前抗好中球抗体投与群では,再潅流時のOiの産生はほとんど認められず,
liposomal SOD十カタラーゼ投与群でも再濯流時の相対的発光強度は,非特異的発光強度
と有意差は認められなかった.これはPAF措抗剤投与群でもほぼ同様であった.またイヌ
では抗好中球抗体およびPAF措抗剤投与群で虚血・再漕流後のミオグロビン,各種逸脱酵
素値の上昇抑制が認められた.
以上より,骨格筋における虚血・再濯流障害時の活性酸素産生源としては,穎粒球にお
けるNADPH oxidase系が強く示唆された.(日血外会誌1 : 35-39, 1992)
るcaudal femomal arteryおよびveinのみを残した.動
脈遮断は60分とし,再濯流後120分まで高橋,中野ら
の方法に準じてウミホクル・ルシフェリン誘導体であ
るCLA (2-melhyl-6-phenyl-3,7-dihydroimidazo[1,2
-a]pyrazin-3-one)に依存する化学発光強度を測定し
た.なお, photoraultiplierおよびsingletphoton count-
Wistar系雄性ラットの薄筋を用い,これに出入りす ing apparatusは,フランスCherpentier社製およびス
ウェーデンA. G. Woltemett社製のを用いた.また,遮
断前抗好中球抗体投与群, liposomal SOD とカタラー
ゼ投与群および血小板活性化因子(以下PAF)の措抗
36
ミオグロピン
ng/mj
12
10
8
6
4
2
0
1。6 1.4
1‐2
相対的発光強度
1.0
0.8
1.2
1.0
0.8
動脈遮断再潅流
0
再漢流
0
特異的発光 1.2
30 60 90
1-SOD+catalase投与群
4゛
`
b-
30
1201nin
マ竺e
,----?..--.
非特異的発光
60 90 1201nin
1.4
1.2
1.0
0.8
抗好中球抗体投与群
日血外会誌 1巻1号
抗好中球抗体投与群
iOmin
0 30 60 90 120min
動脈遮断 再濯流
60min
0 30 60 90 120min
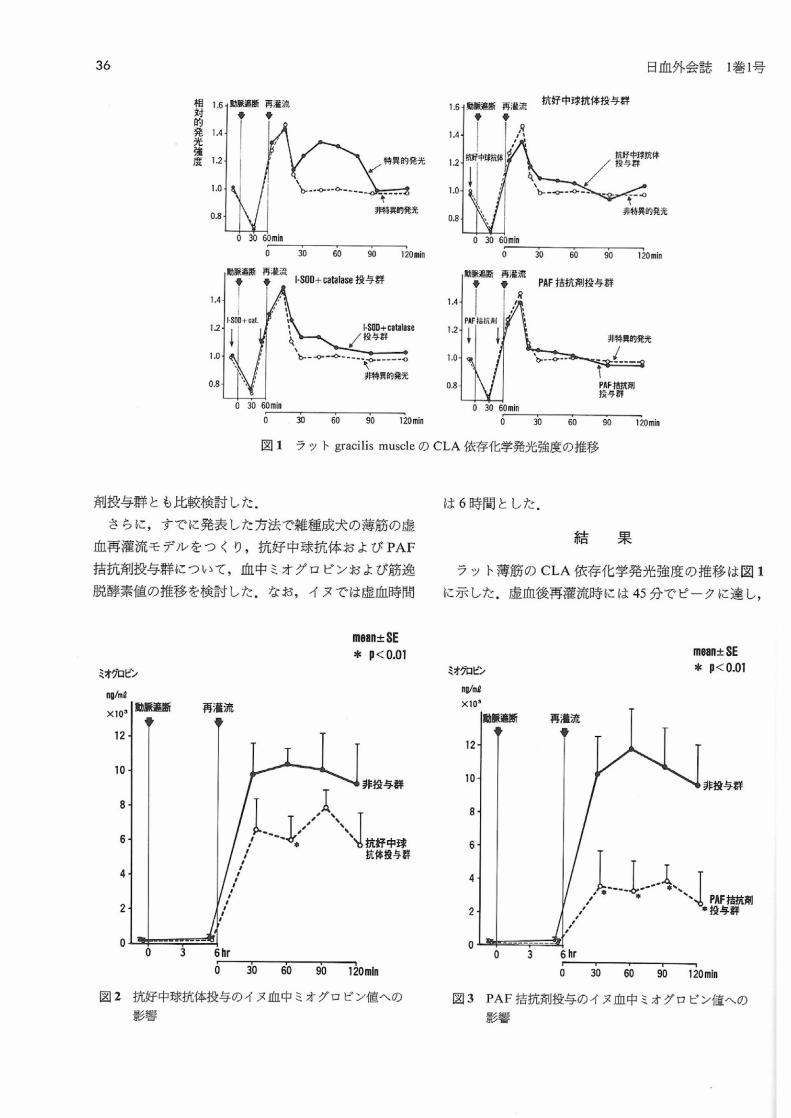
図1 ラットgracilis muscleのCLA依存化学発光強度の推移
mean土SE
*p<0.01
は6時間とした.
結 果
ラット薄筋のCLA依存化学発光強度の推移は図1
に示した.虚血後再濯流時には45分でピークに達し,
ミオグロビン
ns/ni
XIO*
1 1
抗体投与群
8
6
4
2
0
0 3
maan士SE
*p<0.01
非投与群
\リレ
-
U
゛゛ヽ
I
PAF桔抗剤/ *投与群
6h「
0 30 60 90 1 20mill
図3 PAF措抗剤投与のイヌ血中ミオグロビン値への
影響
・―
j 。4
1
|・SO【】
↓
十cat.
0 30 60niin
剤投与群とも比較検討した.
さらに,すでに発表した方法で雑種成犬の薄筋の虚
血再濯流モデルをつくり,抗好中球抗体およびPAF
措抗剤投与群について,血中ミオグロビンおよび筋逸
脱酵素値の推移を検討した.なお,イヌでは虚血時間
1 √゛`非投与群
レじ
゛`
Jj/’
l
゛゛
゛
1
抗好中球
6h「
0 30 60 90 120min
図2 抗好中球抗体投与のイヌ血中ミオグロビン値への
影響

1992年10月
XOH : iantinn dehydrogensse
根本ほか:骨格筋における虚血・再濯流障害
XOD:lanUiina oiidase
図4 骨格筋再濯流障害における微小循環系の血管内皮一穎粒球相互反応
95分で非特異的発光強度レベルに戻るパターンを示
すのに対して,抗好中球抗体投与群, liposomal SOD と
カタラーゼ投与群およびPAF桔抗剤投与群では,そ
の発光強度は非特異的発光強度と類似しており,再濯
流時のOiの産生抑制パターンを示した.
一方,抗好中球抗体投与によるイヌ血中ミオグロピ
ン値への影響をみると,図2のように非投与群に比べ
て上昇は,特に再濯流60分後で有意に抑制された.
また,イヌ血中ミオグロビンのPAF桔抗剤投与で
も,図3に示すように非投与群に比べて有意の低値を
示した.
イヌ薄筋虚血再濯流モデルでの抗好中球抗体投与時
のCPK,LDH,GOT,アルドラーゼの推移をみると,
これらいずれの筋逸脱酵素値においても,有意の上昇
抑制が認められた.
また, PAF措抗剤投与群では,非投与群と比べて,
イヌ筋枝静脈血中のCPK, LDH, GOT,アルドラー
ゼは有意の低値を示した.
考 察
McCord, Grangerらの報告以来,再潅流時の活性酸
素産生源としては, hypoxanthine-xanthine oxidase系
が主として考えられてきたが,高分子であるSODが
細胞内のこの系に作用するとは考え難いこと,肝では
xanthine dehydrogenaseからxanthine oxidaseへの変
換に時間を要すること,また, xanthine oxidase活性は
37
動物種,臓器種による差が著しいことなどより,他の
活性酸素産生源が検討されてきた.骨格筋では脳,心
臓とともに,ヒト,イヌでxanthine oxidase活性はほ
とんど認められない.
今回の実験から,骨格筋の再濯流時には,ラットで
はCLA依存化学発光で示されたOiが抗好中球抗体
あるいはPAF措抗剤で抑制され,またイヌでは再濯
流時に上昇するミオグロピンあるいは各種筋逸脱酵素
がやはり抗好中球抗体あるいはPAF措抗剤で抑制さ
れることが明らかとなった.
この実験結果と諸家の報告とから,現段階では骨格
筋の再濯流障害については,以下のように考えている
(図4).
すなわち,骨格筋微小循環系における穎粒球の
NADPH oxidase系が主な活性酸素産生源で,細胞質
内のNADPHより電子を得て,細胞膜外で02分子は
すべてOiとなり,さらに不均化反応によりH202を
生ずる.これはアズール穎粒の脱穎粒によるミ土ロペ
ルオキシダーゼを介してC10-に変換される.
これ自身,強力な細胞障害および組織障害を示すが,
また不活性型の蛋白分解酵素を活性型とし,障害はさ
らに広範に及び,さらにpluggingによるno reflow現
象も加わり,再濯流障害が発現するものと考えられる.
この際,穎粒球の活性化および血管内皮へのadhesion
に関与するのがPAFではないかと推測される.前述
の通り, xanthine oxidasc 活性は,骨格筋細胞では認め

38
られないが,微小血管内皮細胞内には普遍的に存在し
ているわけで,そこのhypoxanthine-xanthine oxidase
系から生じたH202がphospholipase A2に働き, lyso
PAFを経てPAFが生ずるとの報告もあり,このよう
な微小循環系における血管内皮と頴粒球の相互反応が
重要と考えられる.
なお,ミエロペルオキシダーゼやPAF以外にも,細
胞・組織障害に関与する未知のchemical mediator が存
在する可能性もある.いずれにしても,活性酸素は再
濯流障害のinitiatorの役をしているといえよう.
まとめ
1)ラット薄筋でのCLA依存化学発光測定では,
日血外会誌 I巻1号
抗好中球抗体,SODとカタラーゼあるいはPAF措抗
剤投与により,非投与時に認められる再濯流時のOi
産生は抑制された.
2)イヌ薄筋における再潅流時の血中ミオグロピン
値あるいは各種筋逸脱酵素値は,抗好中球抗体あるい
はPAF措抗剤投与群で,非投与群に比べ有意の上昇
抑制を示した.
3)以上より,骨格筋における虚血後再濯流時の主
な活性酸素産生源としては,穎粒球におけるNADPH
oxidase系が考えられる.

1992年10月 根本ほか:骨格筋における虚血・再濯流障害
Abstract
Ischemia Reperfusion Injury of the Skeletal Muscle I
Generation of Active Oxygens
Hiromu Nemoto, Masaki Morise, Michio Sugino, Kiyoshi Imai,
Masayoshi Kinjou, SeiichiShimazu and Akira Kakita
Department of Surgery, School of Medicine, Kitasato University
Key words: Ischemia reperfusioninjury, Active oxigens,Granulocyte
39
Ischemia reperfusion of the skeletal muscle in relation to the generation of active oxygens stillhas not
been made sufficiently clear. In making this clear, the basis of reperfusion injury was experimentaly
examined.
The gracilis muscle of rats with ischemia at 60 minutes and reperfusion at 120 minutes were divided
into groups in which the in situ O2 generation was measured. They were the ischemia reperfusion only
group, monoclonal neutrophil antibody prescribed group, liposomal SOD + catalase prescribed group and
the platelet activating factor (PAF) antagonist prescribed group.
Also the gracilis muscle of mongrel dogs with ischemia at 6 hours and reperfusion at 120 minutes were
divided into the ischemia reperfusion only group, monoclonal neutrophil antibody prescribed group and
the PAF antagonist prescribed group where the serum myoglobin, CPK, LDII, GOT and aldolase were
measured.
In the rat ischemia reperfusion only group, neutrophil infiltration into the muscle was seen along with
an elevation of Oi at the time of reperfusion and then 95 minutes later changing to a non-specific level.
On the other hand, the monoclonal neutrophil antibody prescribed group, liposomal SOD + catalase
prescribed group and the PAF antagonist prescribed group showed no signs of O2 generation at the time
of reperfusion.
Also unlike the ischemia perfusion only group, in the mongrel dogs, the monoclonal antibody
prescribed group and the PAF antagonist prescribed group showed a rise in myoglobin, CPK, LDH, GOT
and aldolase.
It can be seen from the above data that at the time of the reperfusion injury, the skeletal muslce's active
oxygens generation was granulocyte. (Jpn. J. Vase. Surg., 1: 35-39, 1992)


















