
A kettős peroxisoma proliferator receptor aktiválta (PPAR alfa/gamma) agonisták

De rol van Peroxisome Proliferator Activated Receptor
Alfa in leversteatotische muizen
Sofie DAELEMANS
Verhandeling ingediend tot
het verkrijgen van de graad van
Master in de Biomedische Wetenschappen
Promotor: Prof. Dr. Geerts A.
Co-promotor: Dr. Van Steenkiste C.
Vakgroep Hepatologie
Academiejaar 2013-2014

“De auteur en de promotor geven de toelating deze masterproef voor consultatie beschikbaar
te stellen en delen ervan te kopiëren voor persoonlijk gebruik. Elk ander gebruik valt onder
de beperkingen van het auteursrecht, in het bijzonder met betrekking tot de verplichting
uitdrukkelijk de bron te vermelden bij het aanhalen van resultaten uit deze masterproef.”
14/05/2014
Sofie Daelemans Prof. Dr. Anja Geerts

Voorwoord
Via deze weg had ik graag enkele mensen bedankt die hun steentje hebben bijgedragen aan
dit project en ook aan mijn verdere carrière. Mijn promotor prof. dr. Anja Geerts bedank ik
voor haar steun en vertrouwen. Een deel van het praktisch werk werd uitgevoerd in de
laboratoria van de Université de Louvain là Nèuve in Brussel. Daar had ik graag prof. dr.
Isabelle Leclercq erg hard bedankt voor de kans die ze het project en mij gaf. Verder was er
Vanessa Legry die met ongelooflijke toewijding zich stortte op het project terwijl ze zelf de
handen vol had. Haar wetenschappelijk inzicht heeft de basis van deze masterproef tot een
hoger niveau gebracht. Overigens dank ik eveneens de doctoraatsstudenten, postdocs en
laboranten die altijd bereid waren om me te helpen.
De verdere analyses werden in het laboratorium in Gent uitgevoerd. Daar kon ik altijd
rekenen op de hulp van Anja Van den Bussche, Eliene Bogaerts, Lindsey Devisscher en
Debby Laukens waarvoor mijn dank. Als laatste maar niet de minste wil ik absoluut mijn co-
promotor, dr. Christophe Van Steenkiste, bedanken voor zijn genialiteit. Het was zijn idee en
volharding die ervoor zorgde dat dit project tot stand gekomen is. Afgelopen jaar heb ik goed
met hem kennis gemaakt en hoop dat we in de toekomst zo of zelfs nog beter kunnen
samenwerken.
Door deze masterscriptie ben ik in aanraking gekomen met de onderzoekswereld en het
maakte de zin om te participeren alleen maar groter. Deze masterproef is slecht een klein
voorsmaakje van het werk dat moet gebeuren maar het opende wel veel deuren.

Inhoudstafel
Samenvatting .............................................................................................................................. 1
Summary .................................................................................................................................... 2
1. Inleiding .............................................................................................................................. 3
1.1. Probleemstelling .......................................................................................................... 3
1.2. Proefdiermodel: foz/foz ................................................................................................ 4
1.3. Probiotica en leverziekten ............................................................................................ 5
1.4. Saccharomyces cerevisiae ............................................................................................ 6
1.5. Peroxisome proliferator activated receptor α ............................................................... 6
1.5.1. Rol van PPAR-α in de intestinale vetzuuroxidatie ................................................ 8
1.5.2. PPAR-α reguleert de intestinale cholesterol flux .................................................. 8
1.5.3. PPAR-α reguleert het intestinale nutriëntentransport en metabolisme .................. 8
1.5.4. PPAR-α geeft verminderde effecten van oxidatieve stress ................................... 8
1.5.5. Andere effecten van PPAR-α ................................................................................ 9
1.6. Experimentele NASH (foz/foz muismodel) en PPAR-α .............................................. 9
1.7. Hypothese .................................................................................................................... 9
2. Materiaal en methode ....................................................................................................... 10
2.1. Pilotstudie .................................................................................................................. 10
2.2. Dieet-geïnduceerd en genetisch gemanipuleerd model ............................................. 10
2.3. Behandeling met Saccharomyces cerevisiae ............................................................. 10
2.4. Groepen ...................................................................................................................... 11
2.5. Algemene parametersmeting ..................................................................................... 12
2.6. Orale glucose tolerantie test ....................................................................................... 12
2.7. Permeabiliteitstest ...................................................................................................... 12
2.8. Bacteriele translocatie ................................................................................................ 13
2.9. Weefsel verzameling .................................................................................................. 13
2.10. Microscopie ............................................................................................................. 14
2.11. RT- qPCR ................................................................................................................ 14
2.12. Statistiek .................................................................................................................. 15

3. Resultaten ......................................................................................................................... 16
3.1. Pilotstudie .................................................................................................................. 16
3.2. Algemene parameters ................................................................................................ 16
3.2.1. Voedselinname .................................................................................................... 16
3.2.2. Lichaamsgewicht ................................................................................................. 17
3.2.3. Glycemie .............................................................................................................. 19
3.3. Orale glucose tolerantietest ........................................................................................ 21
3.4. Weefselanalyse .......................................................................................................... 22
3.4.1. Histologie ............................................................................................................ 25
3.4.2. RT-qPCR ............................................................................................................. 27
3.5. FITC dextran permeabiliteitstest ............................................................................... 29
3.6. Bacteriële translocatie ................................................................................................ 29
4. Bespreking ........................................................................................................................ 30
4.1. Steatose muismodel ................................................................................................... 30
4.2. Metabole parameters .................................................................................................. 30
4.3. Cerevisiae behandeling en microbiota ....................................................................... 32
4.4. Effect van cerevisiae op PPAR-α afhankelijke genen in de darm ............................. 32
4.5. PPAR-α en extra-intestinale effecten ......................................................................... 34
4.6. NAFLD en intestinale permeabiliteit ......................................................................... 34
5. Kritische bemerkingen en slotconclusies ......................................................................... 36
6. Referentielijst .................................................................................................................... 37

1
Samenvatting
Niet alcoholische steatohepatitis (NASH) is één van de oorzaken van chronisch leverlijden in
België. Bij deze leveraandoening wordt leversteatose geïnfiltreerd door inflammatoire cellen
en kan het een voorbode zijn van fibrose, cirrose en hepatocellulair carcinoom. Onderzoek
heeft aangetoond dat met behulp van een peroxisome proliferator activated receptor α
(PPAR-α) agonist steatose kan verholpen worden. Deze receptor en transcriptiefactor is
betrokken bij het vetzuur- en glucosemetabolisme en heeft een anti-inflammatoire werking.
De gist Saccharomyces cerevisiae heeft volgens Desmeureux et al. een PPAR-α agonistische
werking op de darm. In onze studie werd er een profylactisch experiment opgesteld met als
doelstelling de leversteatose te voorkomen. Zes weken lang werd er dagelijks cerevisiae
toegediend via orale gavage aan een steatose muismodel (foz/foz) en wild type (WT) muizen
om later de effecten van PPAR-α te testen. Het steatose model werd bevestigd a.d.h.v.
leverhistologie en vetverhouding. Hiertoe was er zowel in placebo als cerevisiae behandelde
foz/foz muizen met een high fat dieet (HFD) histologisch meer steatose en een grotere
vetverhouding gevonden t.o.v. alle WT muizen en foz/foz muizen met een normaal dieet
(ND). In de vijfde week van de behandeling werden significant verhoogde glycemiewaarden
gevonden voor cerevisiae behandelde foz/foz muizen met een HFD t.o.v. hun respectievelijke
placebo’s. Het lijkt erop dat deze muizen door de cerevisiae behandeling eerder metabole
veranderingen als glucose-gevoeligheid en insulineresistentie ondergaan. Dat fenomeen
wordt bevestigd met een orale glucose tolerantietest. Er zijn echter vele mechanismen
betrokken bij de interactie van glucosehomeostase en PPAR-α. Het PPAR-α afhankelijke
intestinale barrière gen Slc27a4 vertoonde een significant hogere expressie t.h.v. het jejunum
bij WT muizen en t.h.v. het ileum bij foz/foz muizen. Cerevisiae voert dus een lokaal effect
uit op de darm en mogelijks ook op de microbiota. Dit laatste bleek uit de verandering van
het caecum gewicht, gezien dit een indirecte maat is voor de veranderde microbiota. Ondanks
het lokale effect van het PPAR-α afhankelijke intestinale barrière gen Slc27a4 is er geen
behandelingseffect gevonden voor de intestinale permeabiliteit en bacteriële translocatie. Tot
slot kan geconcludeerd worden dat het voedingssupplement Saccharomyces cerevisiae in
combinatie met een vetrijk dieet de aangeboren genetische voorbestemming tot ontwikkeling
van obesitas en insulineresistentie van foz/foz muizen beïnvloedt door stimulatie van de
metabole stoornis. De translatie van een muismodel naar de humane setting dient hier met
zeer veel voorzichtigheid uitgevoerd te worden.

2
Summary
Background: Non-alcoholic steatohepatitis (NASH) is a severe and common disease. It can
lead to fibrosis, cirrhosis and finally hepatocellular carcinoma. An peroxisome proliferator
activated receptor α (PPAR-α) agonist has an treatment effect on steatosis. Saccharomyces
cerevisiae is a yeast that upregulates PPAR-α.
Method: An prophylactic study was initiated to test cerevisiae’s PPAR-α upregulation to
prevent NASH. WT and foz/foz mice were fed an HFD or ND under pair feeding conditions
and given cerevisiae or placebo (PBS) for 6 week. Food intake, body weight and glycemia
was checked on a regular basis. In the last week of treatment an OGTT was performed.
PPAR-α expression was tested with RT-qPCR by PPAR-α dependent barrier protein genes
(Abcd3, Cyp4f16, Slc27a4)
Results: Liver steatosis was present in all foz/foz with HFD treated with placebo and
cerevisiae. Glycemia levels were significant higher at week 5 in treated foz/foz mice with
HFD when compared to placebo. OGTT showed also an higher glycemia level after 15 min in
treated foz/foz mice. PPAR-α expression of Slc27a4 was significant elevated in jejunum of
treated WT mice and in ileum of treated foz/foz mice when compared to placebo.
Conclusion: It seems that the yeast Saccharomyces cerevisiae stimulates the metabolic
disorder in foz/foz mice to develop obesity and insulin resistance when a HFD is fed. More
research is needed to discover mechanism between glucose homeostasis and PPAR-α.

3
1. Inleiding
1.1. Probleemstelling
De voornaamste oorzaken van chronisch leverlijden in België en de wereld zijn alcohol
abusus, een overvloedig Westers dieet, virale hepatitiden (hepatitis B en C), leversteatose en
niet alcoholische vette leverziekte (NAFLD). Laatstgenoemde is een leveraandoening die in
fasen verloopt. Zo omvat NAFLD twee onderdelen. Enerzijds is er de goedaardige
leververvetting of benigne steatose, ook wel ‘non-alcoholic fatty liver’ (NAFL) genoemd.
Anderzijds is er een agressieve variant, niet-alcoholische steatohepatitis of ‘non-alcoholic
steatohepatitis’ (NASH). Vooreerst ontstaat er leversteatose door accumulatie van vet in de
lever waarbij naast leververvetting ook een chronische inflammatie van de lever ontstaat, die
naarmate de ziekte vordert, kan leiden tot leverfibrose, cirrose en zelfs hepatocellulair
carcinoom. In 20% tot 30% van de gevallen evolueert steatose naar NASH. De pathogenese
van NASH is een multifactorieel proces waarbij het tot op heden nog onduidelijk is welke
factoren oorzakelijk zijn en welke factoren tot een verdere evolutie in de ziekte leiden. Het
huidige concept bestaat uit een twee hit model waarbij een initieel metabolische verandering,
zoals abdominale obesitas, dyslipidemie, hypertensie en insuline resistentie met of zonder
hyperglycemie, steatose zou veroorzaken en een tweede stimulus, zoals oxidatieve stress,
bacteriële toxines en inflammatoire cytokines (TNF-α), doet evolueren tot steatohepatitis
(Figuur 1) (1–3).
Naast directe effecten van een vetrijk dieet, zoals accumulatie van vet in de lever, hebben
maaltijden met een hoge hoeveelheid vrije vetzuren ook effect op de genen. Zo spelen genen
als de peroxisome proliferator activated receptoren α en γ een belangrijke rol in dit onderzoek
en zijn ze van belang bij de ontwikkeling van een tweede hit (1). Recente inzichten geven
overigens aan dat de darmflora evenzeer aanleiding kan geven tot een tweede hit via
aangeboren immuun mechanismen of overvloedige endogene alcoholproductie (4).
Er zijn momenteel nog geen specifieke behandelingen voor NASH. Enkel levensstijl-
veranderingen zoals een dieet, beweging en gewichtsverlies kunnen voor een verbetering
zorgen (1).

4
Figuur 1: De twee hit hypothese voor de pathogenese van NASH. Hepatische steatose is een initiële hit die vrij algemeen plaatsvindt. Eenmaal dat steatose ontwikkeld is, wordt de lever kwetsbaar voor de tweede hit. Het induceert verder oxidatieve stress. Dit laatste kan eventueel leiden tot celdood en inflammatie en tot slot steatohepatitis (5).
1.2. Proefdiermodel: foz/foz
Onderzoek naar NASH wordt gelimiteerd door het gebrek aan een goed proefdiermodel.
Veelgebruikte diermodellen voor NASH, zijn genetische modellen (zoals ob/ob muizen,) of
dieet geïnduceerde modellen zoals (choline/methionine deficiëntie dieet of ‘high fat’ dieet)
(5). Vele obese en diabetische muismodellen ontwikkelen steatosis en slechts enkele daarvan
ontwikkelen een pathologie dat overeenkomt met NASH. Sinds 2006 is er een nieuw obees
muismodel voorgesteld door de Australische onderzoeksgroep van Arsov et al., namelijk Fat
Aussie muis. Deze stam heeft insuline resistentie en diabetes type 2 dat bovendien een
transitie meemaakt van eenvoudige steatose tot NASH wanneer een high fat dieet (HFD)
wordt gegeven. De aussie muis ontstaat door een spontane mutatie (foz), een 11-bp deletie in
exon 8 van het Alms1 gen. Dit gen is verantwoordelijk voor het Alström syndroom (6). Dit is
een autosomale recessieve ziekte van kinderobesitas, blindheid, gehoorverlies, diabetes type
2, dyslipidemia, lage levels van het groeihormoon, hypothyroidie, infertiliteit en frequent
abnormaliteiten van nier, hart en lever. Dit laatste heeft betrekking tot steatose, fibrose en
cirrose. De foz/foz muis wordt obees op een leeftijd van 100 tot 120 dagen wanneer een HFD
gegeven wordt en ze ontwikkelt later een hepatomegalie en macroscopische steatose. De
biochemische basis voor de transitie van steatosis naar ernstige fibrotische steatohepatitis is
een falen van het opreguleren van pathways in het hepatische vetzuur katabolisme en dit leidt
tot massieve vet-accumulatie (6–8). Vanaf het spenen eten de foz/foz muizen meer dan hun
wild-type (WT) nestgenoten. Hiertoe stijgt hun lichaamsgewicht eveneens meer. Volgens
Arsov et al. is het gemiddelde lichaamsgewicht van muizen met een normaal dieet (ND) op
een leeftijd van 200 dagen: 54 ± 1,1g foz/foz tegenover 30 ± 1,1g WT (p<0,0001) en met een

5
HFD: 61 ± 1,9g foz/foz tegenover 36 ± 1,2g wild type muizen (P < 0,001). De foz/foz muizen
met een ND en de wild type muizen met een HFD vertonen simpele steatosis op een leeftijd
van 300 dagen. De foz/foz muizen die een HFD aten kregen daarentegen steatohepatitis. De
pathologie vertoont kenmerken als een hogere hoeveelheid hepatocyten, ernstige ballooning1
degeneratie, extensieve lobulaire inflammatie met polymorfe en mononucleaire cellen en
pericellulaire en pericentrale fibrose (7).
1.3. Probiotica en leverziekten
De Wereldgezondheidsorganisatie definieert probiotica als levende microbiële culturen die
wanneer toegediend in voldoende aantallen, een gezondheid bevorderend effect kunnen
uitoefenen op de gastheer. De menselijke darmflora bestaat uit 1013 – 1014 micro-organismen.
Het collectieve genoom van probiotica bevat op zijn minst 100 maal meer genen als dat van
onszelf. Dit representeert 500-1000 species in het totaal. Ze worden gebruikt om de darmflora
te moduleren en werden reeds voorgesteld om mogelijks leverfuncties te verbeteren (9). Dieet
en levensstijlveranderingen zijn momenteel de beste behandelingen voor het controleren van
NAFLD bij de patiënten (10).
De lever en de darmen hebben dezelfde embryologische foregut. Bijkomstig krijgt de
lever bloed afkomstig van de darm door het portale systeem. Hiertoe is duidelijk dat de
darmen en lever een nauwe relatie hebben. Een dieet met hoog vetgehalte is gerelateerd aan
een verandering in de compositie van de intestinale bacteriën en dit is mogelijk geassocieerd
met NAFLD. De darmflora is mogelijks ook betrokken bij het ontstaan ervan. Dit door de
intestinale permeabiliteit te vergroten, activatie van inflammatoire cytokines door vrijlating
van lipopolysaccharide (LPS) en absorptie van endotoxines (10). Endotoxinen activeren
Kupffercellen in de lever die een verhoogde productie van TNF-α en IL-6 veroorzaken.
Samen met het complex mechanisme van extensieve lipide accumulatie, systemische
inflammatie, oxidatieve stress en insulineresistentie wordt een hepatopathologie veroorzaakt.
NAFLD in mensen is geassocieerd met een stijging van de intestinale permeabiliteit en dit is
abnormaal gerelateerd aan de stijgende aanwezigheid van intestinale bacteriële overgroei.
Probiotica kunnen de proliferatie van schadelijke bacteriën inhiberen, de bacteriële overgroei
reduceren, de gastro-intestinale barrière functie van epitheliale cellen herstellen, de intestinale
permeabiliteit verminderen en het immuunsysteem moduleren (10). Darmflora spelen dus een 1 Ballooning: Hepatocellulaire ballooning is een typisch kenmerk voor NASH. Dit is microscopisch te zien op een hematoxyline en eosine gekleurde coupes. Het is een grote vetdruppel binnenin een cel met een dun cytoplasma (30).

6
belangrijke rol in de gezondheid van de gastheer. Allereerst helpt het bij de vertering en
absorptie van complexe koolhydraten. Als tweede produceren de darmbacteriën daarbij
essentiële substanties als vitamine K. Ze helpen ten derde bij het behoud van de integriteit
van de epitheelbarrière door de productie van korte keten vetzuren. Darmflora helpen bij de
maturatie van het immuunsysteem van de gastheer en beschermen tegen pathogene micro-
organismen door competitie ervan (4).
1.4. Saccharomyces cerevisiae
Het probiotica Cerevisia wordt momenteel als supplement voorgeschreven voor mensen met
een prikkelbare darm of irritable bowel syndroom (IBS). Het verbetert abdominale pijn en
verteringsdiscomfort. Door zijn anti-inflammatoire activiteit in experimentele colitis muizen,
visceraal analgetische effecten in een ratmodel van colon hypersensitiviteit en een
antagonistische werking op E. Coli O157:H7is het een interessante gist om verder onderzoek
op uit te voeren (11–13).
In het onderzoekscentrum in Lille ontdekten Desreumaux en zijn collega’s dat de gist
Saccharomyces cerevisiae CNCM I-3856 PPAR-α tot expressie brengt in de darmen van de
muis en rat. In vitro werd dit gezien op epitheelcellen en bij een in vivo model voor
colorectale distensie bij ratten. Deze data werden gepubliceerd in abstract vorm in 2010, maar
de volledige data zijn tot op heden nog niet gepubliceerd. Uit een gerandomiseerde placebo-
controleerde studie bij patiënten met IBS blijkt dat Saccharomyces cerevisiae CNCM 1-3856
een goed tolereerbare gist is dat o.a. een visceraal analgetisch effect (11). Verschillende
stammen van de gist Saccharomyces cerevisiae worden reeds lange tijd gebruikt bij het
maken van brood, yoghurt, wijn en bier. Ze zijn algemeen bekend voor hun invloed op de
spijsvertering en de darmtransit (14).
1.5. Peroxisome proliferator activated receptor α
De peroxisome proliferator activated receptors zijn delen van de humane nucleaire receptoren
die functioneren als ligand-afhankelijke transcriptiefactoren. Ze worden geactiveerd door
binding met verschillende endogene vetzuren, hun metabolieten en synthetische liganden.
Eenmaal geactiveerd door hun ligand vormen ze een heterodimeer met de nucleaire retinoid
X receptor (RXR). Het dimeer bindt met de cis-element van een promotor van een doelgen.
Peroxisome proliferator activated receptor α is een van de drie subtypes en komt tot expressie
in verschillende organen waaronder de lever en darmen (15).
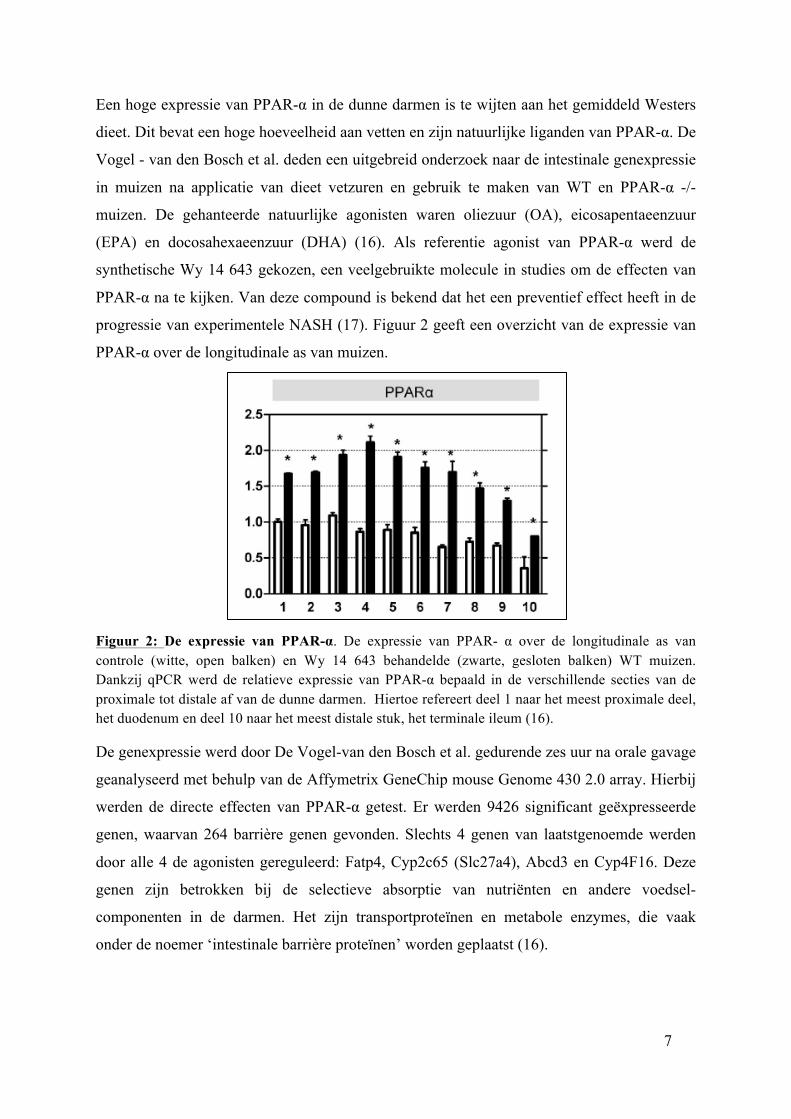
7
Een hoge expressie van PPAR-α in de dunne darmen is te wijten aan het gemiddeld Westers
dieet. Dit bevat een hoge hoeveelheid aan vetten en zijn natuurlijke liganden van PPAR-α. De
Vogel - van den Bosch et al. deden een uitgebreid onderzoek naar de intestinale genexpressie
in muizen na applicatie van dieet vetzuren en gebruik te maken van WT en PPAR-α -/-
muizen. De gehanteerde natuurlijke agonisten waren oliezuur (OA), eicosapentaeenzuur
(EPA) en docosahexaeenzuur (DHA) (16). Als referentie agonist van PPAR-α werd de
synthetische Wy 14 643 gekozen, een veelgebruikte molecule in studies om de effecten van
PPAR-α na te kijken. Van deze compound is bekend dat het een preventief effect heeft in de
progressie van experimentele NASH (17). Figuur 2 geeft een overzicht van de expressie van
PPAR-α over de longitudinale as van muizen.
Figuur 2: De expressie van PPAR-α. De expressie van PPAR- α over de longitudinale as van controle (witte, open balken) en Wy 14 643 behandelde (zwarte, gesloten balken) WT muizen. Dankzij qPCR werd de relatieve expressie van PPAR-α bepaald in de verschillende secties van de proximale tot distale af van de dunne darmen. Hiertoe refereert deel 1 naar het meest proximale deel, het duodenum en deel 10 naar het meest distale stuk, het terminale ileum (16).
De genexpressie werd door De Vogel-van den Bosch et al. gedurende zes uur na orale gavage
geanalyseerd met behulp van de Affymetrix GeneChip mouse Genome 430 2.0 array. Hierbij
werden de directe effecten van PPAR-α getest. Er werden 9426 significant geëxpresseerde
genen, waarvan 264 barrière genen gevonden. Slechts 4 genen van laatstgenoemde werden
door alle 4 de agonisten gereguleerd: Fatp4, Cyp2c65 (Slc27a4), Abcd3 en Cyp4F16. Deze
genen zijn betrokken bij de selectieve absorptie van nutriënten en andere voedsel-
componenten in de darmen. Het zijn transportproteïnen en metabole enzymes, die vaak
onder de noemer ‘intestinale barrière proteïnen’ worden geplaatst (16).

8
1.5.1. Rol van PPAR-α in de intestinale vetzuuroxidatie
De peroxisome proliferator activated receptor α is een hoofdzakelijke regulator van het
vetzuurkatabolisme. Vele transporters en fase I enzymes, die betrokken zijn bij de vetzuur
opname en oxidatie, zijn PPAR-α afhankelijk. Lange ketenvetzuur opname, mitochondriale
en peroxisomale β-oxidatie, ω-oxidatie en het energiemetabolisme als de glycogenolysis en
Krebs cyclus vinden hun regulatie bij o.a. PPAR-α (16).
1.5.2. PPAR-α reguleert de intestinale cholesterol flux
De expressie van de apicale cholesterol opname transporter Npc1l1 was PPAR-α afhankelijk
onderdrukt na een behandeling met Wy 14 643, EPA en DHA. Het PPAR-α afhankelijke
Abca1 gen promoot de cholesterol efflux op het basolaterle membraan naar Apo-AI voor de
vorming van HDL. Dit was verhoogd na een behandeling met Wy 14 643 en EPA. Dit wil
zeggen dat er minder cholesterol wordt geabsorbeerd van het lumen en meer cholesterol
wordt getransfereerd naar Apo-A1 waardoor er een verminderde intracellulaire cholesterol
gehalte ontstaat in de enterocyten (16).
1.5.3. PPAR-α reguleert het intestinale nutriëntentransport en metabolisme
Na de behandeling met de agonisten werd gezien dat de apicale glucose opname transporter
(Sglt1) en de basale glucose transporter (Glut2) bedwongen werd. Hiertoe werd gesuggereerd
dat PPAR-α activatie belangrijk is in het transport van glucose door de intestinale wand. Er
werden eveneens verscheidene transporters betrokken bij het aminozuurmetabolisme PPAR-α
afhankelijk onderdrukt (16).
1.5.4. PPAR-α geeft verminderde effecten van oxidatieve stress
Oxidatieve stress is het resultaat van een onbalans tussen de vorming en afbraak van pro-
oxidanten of dalende cellulaire antioxidanten protectie. Dit alles kan leiden tot celschade en
apoptose. Vele barrière genen zijn betrokken bij de verdedigen tegen oxidatieve stress.
Volgens de data van De Vogel-Van den Bosch et al. wordt dit bekomen door de inhibitie van
genen die betrokken zijn bij de apoptose in de dunne darmen na behandeling met Wy 14 643
(16).

9
1.5.5. Andere effecten van PPAR-α
Bijkomstig heeft PPAR-α een anti-inflammatoir effect. Activatie van de receptor leidt tot
inhibitie van NF κβ, een centrale mediator van ontsteking, en andere pro-inflammatoire
genen. Inflammatory bowel disease (IBD) onderzoek heeft aangetoond dat PPAR-α activiteit
betrokken is in de neutrofiele infiltratie door opregulatie van ICAM1 (18).
1.6. Experimentele NASH (foz/foz muismodel) en PPAR-α
Het nieuwe foz/foz model biedt de mogelijkheid om het pathologisch mechanisme van
hepatische vetverdeling tijdens de transitie van steatosis naar steatohepatitis te bestuderen.
De onderzoekers in Australie deden hele interessante bevindingen. Wanneer HFD gevoede
foz/foz muizen de peroxisome proliferator activated receptor α agonist, Wy 14 643, voor 10
of 20 dagen toedienden, verbeterde de metabolische indexen, steatose, ballooning en
leverinflammatie in diabetische muizen met NASH. De behandeling had bovendien een
gunstig effect op de insuline gevoeligheid en induceerde een verminderd lichaamsgewicht.
Adiponectine levels waren evenzeer lager in de foz/foz muizen met de behandeling ten
opzichte van de WT muizen (19). Adiponectine is een beschermend hormoon vrijgesteld uit
vetweefsel door o.a. insuline en zorgt voor een controle van het energiemetabolisme (20).
Een laag gehalte aan adiponectine, bijvoorbeeld bij obesitas, is een risico factor voor het
ontwikkelen van het metabolisch syndroom en diabetes mellitus. Hypoadiponectinema levert
een eerste mogelijke verklaring waarom hepatische PPAR-α gemedieerde effecten
verminderd bij foz/foz muizen met een HFD en uiteindelijk steatohepatitis (7). De opvallende
biochemische verschillen tussen de levers van “HFD gevoede foz/foz muizen met
steatohepatitis” versus “ de foz/foz muizen met een ND of WT muizen met een HFD met
simpele steatosis” is te wijten aan het falen van de opregulatie van PPAR-α en zijn lager
gelegen genen die betrokken zijn met vetzuuroxidatie en triglyceride vrijlating uit de lever
(7).
1.7. Hypothese
Met bovengenoemde gegevens waaruit blijkt dat PPAR-α een rol speelt in het
levermetabolisme werd beslist om Saccharomyces cerevisiae (met PPAR-α effect) toe te
dienen aan foz/foz proefdieren. Er werd een profylactische studie opgesteld met als
doelstelling de leversteatose in de dieren te voorkomen. De effecten op verschillende
metabole parameters, darmpermeabiliteit en bacteriële translocatie werden bestudeerd.

10
2. Materiaal en methode
2.1. Pilotstudie
Praktische handelingen zoals orale gavage, glycemiemeting, lichaamsgewichtmeting en
gewenning met de muizen werd eerst geoefend op 14 mannelijke WT muizen. Hiertoe werd
aan 7 muizen een cerevisiae behandeling gegeven via orale gavage en aan de andere 7 een
placebo behandeling met (PBS 1x). De toegediende oplossing cerevisiae werd eerst uitgetest
met een aantal verdunningen met PBS 1x waarna de ideale toedieningsverdunning en
hoeveelheid gezocht werd.
2.2. Dieet-geïnduceerd en genetisch gemanipuleerd model
Mannelijke foz/foz muizen afkomstig van laboratorium Gastro-Enterologie in het UCL van
kregen een HFD met een calorische waarde van 5,24 kcal/g en 60% vet of een ND met een
calorische waarde van 2,83 kcal/g. WT muizen afkomstig van hetzelfde laboratorium kregen
eveneens een HFD of een ND. De groepen werden onder de vorm van pair feeding gevoed en
kregen water ad libitum toegediend gedurende 6 weken. De muizen werden ondergebracht in
kooien waar zowel luchtvochtigheid als temperatuur zo constant mogelijk gehouden wordt
met een 12 uur gecontroleerde dag/nacht cyclus om de invloed van deze parameters op het
experiment te beperken. Omwille van de pair feeding condities zijn de proefdieren
individueel ondergebracht in afzonderlijke kooien.
2.3. Behandeling met Saccharomyces cerevisiae
Één capsule Cerevisia geregistreerd bij de Franse Nationale Collectie van Microbiële
Culturen (CNCM) I-3856 (Trenker) bevat 4x109 kolonie vormende eenheden (k.v.e.) van
Saccharomyces cerevisiae. Voor dagelijks versbereide orale gavage dient de inhoud van de
capsules opgelost te worden in Phosfate Buffered Saline (PBS) met de verhouding 1:1,6ml.
Na schudden gedurende 2 minuten werd een lopende bruine oplossing bekomen. Met behulp
van een gavage spuit werd aan de proefdieren een minimum 5x108 k.v.e. cerevisiae
toegediend. Saccharomyces cerevisiae is een niet-koloniserende gist maar bereikt een hoge
concentratie in de darm en blijft aanwezig onder een levende vorm in hoge concentraties na
applicatie voor 12 uur (14). De energetische waarde van Cerevisia is 340 kcal/100g. Hierop
gebaseerd wordt verwacht dat één capsule Cerevisia 1,700 kcal bevat. Elke muis ontving
daaraan sluitend 0,2125 kcal/dag. De energetische waarde toegediend door de Cerevisia
applicatie kan dus als verwaarloosbaar worden beschouwd. Als placebo werd PBS gebruikt.

11
2.4. Groepen
Groep één omvatte bij start van de behandeling 13 mannelijke foz/foz muizen. Er werden
koppels gevormd aan de hand van hun lichaamsgewicht en glycemie van op dag 1. Hiertoe
werden ze onderverdeeld in drie subgroepen zodat er een combinatie kan worden gemaakt
van HFD met cerevisiae behandeling, HFD met placebo (PBS) behandeling en ND met
placebo behandeling. Het gemiddelde lichaamsgewicht was respectievelijk 25,20 ± 1,64 g,
24,80 ± 1,92 g en 23,67 ± 3,06 g. De gemiddelde glycemie op dag 1 was respectievelijk:
157,80 ± 6,38 mg/dl, 153,00 ± 6,16 mg/dl en 160,33 ± 33,29 mg/dl. Deze groep werd
gekozen voor de uitvoering van algemene parameters meting, orale glucose tolerantietest
(OGTT), weefselanalyse zoals histologie en real-time poly chain reaction (RT q-PCR).
Groep twee werd samengesteld voor het uitvoeren van de algemene parameters,
permeabiliteitstest en de bacteriële translocatie. De groep bestond uit 8 mannelijke foz/foz
muizen. Op basis van het lichaamsgewicht en glycemie werden koppels en twee subgroepen
gevormd. Beide subgroepen kregen een HFD met een cerevisiae behandeling of placebo. Het
gemiddelde lichaamsgewicht op dag 1 was respectievelijk 29,50 ± 1,29 g en 29,75 ± 2,97 g.
De gemiddelde glycemie op dag 1 was respectievelijk: 153,75 ± 24,55 mg/dl en 156,75 ±
16,21 mg/dl.
De controle muizen van groep één werden verdeeld in groep drie. Deze groep omvatte 9
WT muizen. Het werd samengesteld als controle voor weefselanalyse zoals histologie en RT
q-PCR. Net zoals de andere groepen werd hier een indeling met koppels gemaakt volgens het
lichaamsgewicht en glycemie van op dag 1. Het gemiddelde lichaamsgewicht op dag 1 was
voor de behandelingssubgroep 24,80 ± 2,86 g en voor de placebosubgroep 24,50 ± 1,29 g. De
gemiddelde glycemie op dag 1 was respectievelijk: 147,60 ± 11,46 mg/dl en 156,50 ± 23,95
mg/dl. De bacteriële translocatie test en permeabiliteitstest werden vergeleken in groep vier
die bestond uit 6 WT muizen met een gemiddelde lichaamsgewicht op dag 1 van 24,00 ± 2,00
g voor de subgroep die cerevisiae kreeg en 23,67 ± 3,51 g voor de subgroep die de placebo
kreeg. De gemiddelde glycemie op dag 1 was 142,67 ± 1,11 mg/dl en 145,33 ± 19,73 mg/dl.
Groep vijf en zes omvat elk 4 WT muizen. Er werden 2 subgroepen gevormd voor elke
groep waardoor telkens 2 muizen de cerevisiae behandeling kreeg en de andere 2 de placebo
behandeling. Het gemiddelde lichaamsgewicht voor groep vijf was respectievelijk 23,00 ±
1,41 g en 15,00 ± 5,66 g en voor groep zes 20,50 ± 2,12 g en 17,5 ± 0,71g. De gemiddelde
glycemie waarden waren op dag 1 van het experiment voor groep vijf respectievelijk 161,00
± 2,83 mg/dl en 180,00 ± 7,07 mg/dl en voor groep 6 177,5 ± 27,58 mg/dl en 186,00 ± 22,63
mg/dl.

12
2.5. Algemene parametersmeting
Lichaamsgewichtmeting gebeurde op steeds dezelfde balans om de foutmarge zo klein
mogelijk te houden. Dit om de dag uitgevoerd. De glycemie wordt gemeten met behulp van
een glucose meter (Accu-chek Active, Roche diagnostics, Basel, Zwitselrand) die
routinematig in het laboratorium wordt gebruikt. Voedselinname werd dagelijks gemeten en
bijgehouden op dezelfde balans.
2.6. Orale glucose tolerantie test
De orale glucose tolerantietest (OGTT) werd in de vijfde week van de behandeling toegepast.
De dieren werden gedurende 4 uur nuchter gehouden door het verwijderen van eten en
verversen van de kooien. Vóór tijdstip 0 werden de glycemie en lichaamsgewicht gemeten
met de Accu-chek via een druppel bloed uit de staartvene, onmiddellijk gevolgd door
toediening van 6,25 µl/g lichaamsgewicht 20% glucose, met een pH van 4,4 en 1110
mosmol/l. Na 15, 30, 45, 60, 90, 120 en 180 minuten werd de glycemie telkens gemeten via
bovengenoemde methode.
2.7. Permeabiliteitstest
Uitvoeren van de permeabiliteitstest werd gedaan aan de hand van een flucorescente
dextraanmolecule, namelijk Fluorescein-isothiocyanaat (FITC) dextran (Sigma). Om een
oplossing met een concentratie van 125 mg/ml te bekomen werd 300 mg poeder opgelost in
2,4 ml PBS (1x) en afgeschermd van licht met behulp van aluminiumfolie. Na de bereiding
werd 600 mg/kg via orale gavage toegediend aan 4 uur nuchtere muizen. Na exact 60
minuten werd een bloedafname via intra-cardiale punctie uitgevoerd. Na het collecteren van
bloedstalen en bewaring op ijs werden de stalen gecentrifugeerd gedurende 5 minuten op
10000 rpm. Serum werd vervolgens opgevangen in een epje per muis en bewaard op -4°C.
Op eenzelfde wijze werd serum verzameld van 7 wild type muizen zonder FITC dextran
toediening voor het bereiden van een verdunningsreeks. Dit laatste werd 1/3 verdund met
PBS (1x). Een verdunningsreeks van FITC dextran met serum, van 1 mg/dl tot 0,00015 mg/dl
werd opgesteld en in duplo verdeeld over een 96 well plaat. Alle andere bekomen
serumstalen met FITC dextran werd eveneens 1/3 verdund met PBS (1x) en eveneens in
duplo verdeeld over de 96 well plaat. De plaat werd bewaard op -4°C met aluminiumfolie om
de volgende dag geanalyseerd te kunnen worden. Analyse werd uitgevoerd met Perkin Elmer
met een excitatie golflengte van 485 nm en emissie golflengte van 535 nm.

13
2.8. Bacteriele translocatie
Muizen werden verdoofd met een Ketamine/Xylazine oplossing 10 µl/g. Na het openen van
de buikholte werd er een steriele isolatie van een viscerale lymfeknoop, gelegen in het
peritoneale vet bij groep 2 en 4 uitgevoerd. Dit gebeurde vóór de intra-cardiale punctie voor
de FITC dextran permeabiliteitstest. In de buurt van een vlam werd de lymfeknoop
mechanisch verkleind door middel van spuitjes en opgelost in 0,300 ml PBS 1x. Hiervan
werd 1 druppel opgevangen op een dekglaasje en het overige uitgestreken volgens de 5-
hoeksmethode over een LP groeibodem in petrischaaltjes. Na het uitstrijken werden de
petrischaaltjes geïncubeerd op 37°C voor 72u. De glaasjes daarentegen werden direct via de
onderkant verwarmd in de vlam en overnacht gedroogd. Tot slot werd een gramkleuring
verricht volgens volgend schema: 1 minuut Kristal violet en spoeling met water, 1 minuut
lugol met spoeling met water, 20 seconden alcohol 96% gevolgd door spoeling met water, 1
minuut fuchsine gevolgd door spoeling met water. Na drogen, werden de coupes bekeken met
een olie-immersie onder de microscoop.
2.9. Weefsel verzameling
Na 4 uur vasten werden de muizen verdoofd met een Ketamine/Xylazine oplossing (10 µl/g).
Wanneer de controle van de bewusteloosheid en weging werd de buikholte geopend om de
bloedafname via intra-cardiale punctie uit te voeren. Bloedstalen werden bewaard op ijs in
een lithiumspuit. Na de sacrificaties werden ze voor 5 minuten gecentrifugeerd op 10000
omwentelingen per minuut (rpm). Serum werd opgevangen en bewaard op -80°C.
Aansluitend op de bloedafname werd lever, milt, jejunum, ileum, caecum, faeces, visceraal
en epidydimiaal vetweefsel gepreleveerd. De bewaring werd op verschillende manieren
uitgevoerd, afhankelijk van de doelanalyse.
• In formol gedurende 24h van trapezevormige leverlob, jejunum, ileum, caecum, milt
en visceraal vetweefsel.
• In 0,600µl RNA later (Qiagen, Venlo, Nederland) van lever, jejunum, ileum en
caecum voor enkele minuten en aansluitend een snelle bevriezing in vloeibare
stikstof. De langdurige bewaring vindt plaats op -80°C.
• Een snelle bevriezing in vloeibare stikstof en voor de langdurige bewaring op -80°C.

14
2.10. Microscopie
Na 24 uur fixatie in formol werden de weefselstukjes overgebracht in PBS. De stalen werden
op een routinematige manier in stijgende ethanolbaden gebracht en gespoeld met xyleen door
de afdeling anatoompathologie van het UCL. Hierna werden de stalen gepositioneerd en
ingebed in paraffine. Voor histologische en immunohistochemische kleuringen werden uit
deze paraffineblokjes met behulp van de microtoom, coupes met een dikte van 5 µm
gesneden.
Vooraleer een kleuring uitgevoerd kon worden, werden de gefixeerde coupes
gedeparaffineerd door ze achtereenvolgens in xyleen, 99% ethanol, 96% ethanol, 70%
ethanol en gedestilleerd water te brengen. De kleuring werd uitgevoerd door ze eerst in
hematoxyline en na spoeling in eosine te onderdompelen voor 3 minuten. De coupes werden
opnieuw gedehydrateerd door ze in omgekeerde volgorde door de ethanol en xyleenbaden te
doorlopen. Montering werd uitgevoerd met DpX.
2.11. RT- qPCR
Een eerste stap in het RT-qPCR proces was RNA isolatie. RNA lysaten werden gemaakt met
behulp van de Qiagen RNeasy Mini Kit volgens het protocol van de fabrikant. Na een
kwaliteitscontrole met de Eppendorf BioPhotometer van alle stalen werden ze op een gelijke
concentratie van 100 ng/µl gebracht en bewaard op -80°C. De volgende stap was de
conversie van RNA naar cDNA. Dit werd aan de hand van oligoDT primers en met behulp
van de Biorad iScript cDNA Synthesis kit uitgevoerd. Het cDNA werd bewaard op -20°C.
De RNA expressie van de verschillende relevante genen werd bepaald met RT-qPCR.
Dit werd uitgevoerd met de Lightcycler 480 Green Master Mix (Roche) en de primers
volgens een standaard gebruikt protocol in het laboratorium. Gebruikte genen,
overeenkomstige primersets en functie worden weergegeven in onderstaande tabel (Tabel 1).
Tabel 1: Gebruikte genen, overeenkomstige primersets en functie
Afkorting Eiwit Forward primer Reversed primer Merker
HMBS Hydroxymethyl bilanesynthase
AAGGGCTTTTCTGAGGCACC
AGTTGCCCATCTTTCATCACTG
Huishoudgen
SDHA Succinaatdehydrogenasecomlex
CTTGAATGAGGCTGACTGTG
ATCACATAAGCTGGTCCTGT
Huishoudgen
HPRT Hypoxanthine – Guanine Phosfo-ribosyltransferase
GTTAAGCAGTACAGCCCCAAA
AGGGCATATCCAACAACAAACTT
Huishoudgen

15
ABCD3 ATP-binding cassette, sub-family D, member 3
TATTGCGGTTATGCTGGTATCTC
TTGCTGCTGCGACCAATGAT
PPAR-α expressie
CYP4F16 Cytochrome P450, family
4, subfamily f, polypeptide 16
CCGCCTCAGTTGTTTCCCTC
TGCCCAAGTGACCTGAAAACC
PPAR-α expressie
FATP4 (SLC27A4)
solute carrier family 27, member 4
ACTGTTCTCCAAGCTAGTGCT
GATGAAGACCCGGATGAAACG
PPAR-α expressie
CYP2C6 cytochrome P450, family
2, subfamily c, polypeptide 65
TCAGAGAAGAAACCTTCCTCCTG
CATATCCATGTAACACCACAGCA
PPAR-α expressie
2.12. Statistiek
Om het effect van de cerevisiae behandeling statistisch te staven wordt gebruik gemaakt van
het programma SPSS-Statistics 22. De muizen werden verdeeld in koppels waarvan één de
cerevisiae en één de placebo behandeling kreeg. Dankzij de pair feeding condities werd
ervoor gezorgd dat de voedselinname overeenstemde binnen het koppel. De verschillende
parameters worden per koppel vergeleken met de test “mixed models”. Hiertoe wordt er
gecorrigeerd voor de correlatie binnen de paren zoals familiale afkomst en voedselinname als
variabele. Het model gaat het verschil tussen de behandeling met cerevisiae en placebo na
voor één variabele. Als random factor werd hierbij het paar gekozen. De paren worden
tegelijkertijd binnenin en onderling met elkaar vergeleken. Een gepaarde test kon niet worden
uitgevoerd omwille van data die verloren zou gaan dan.
Om de verschillende muismodellen en diëten met elkaar te vergelijken wordt gebruik
gemaakt van de ‘one way anova’ test. Hiertoe wordt er eerst de homogeniteitstest uitgevoerd.
Indien deze niet significant is (p>0,05), kunnen de groepen onder elkaar vergeleken worden
met de ‘post hoc Tukey’ test. Indien de homogeniteitstest wel significant is (p<0,05), wordt
een ‘Welch’ test uitgevoerd. Bij een significantie van die test wordt er via de ‘post hoc
Gemes Howell’ test gekeken waar het verschil binnen de groep zit. Als de ‘Welch’ test niet
significant blijkt te zijn, is het ware verschil tussen de gemiddelde van de groepen gelijk aan
nul.

16
3. Resultaten
3.1. Pilotstudie
Uit de voorafgaande pilot studie blijkt dat muizen behandeld met cerevisiae minder aten dan
placebo behandelde dieren: 2,17 ± 0,18 g en 2,39 ± 0,45 g. Vooral gedurende eerste dagen
van het experiment aten de dieren met een cerevisiae behandeling minder. De dagelijkse
supplementatie van een dosis cerevisiae zorgt voor een aanvulling van het dieet met 0,2125
kcal. Dit is slechts een fractie indien dit vergeleken werd met de totale dagelijkse calorie-
inname. De maximale hoeveelheid cerevisiae in oplossing (PBS 1x) dat gegeven kon worden
was 200 ml. De maximale verdunning die bekomen kon worden was 1/1,600ml. Dit bevat
5x108 k.v.e. Saccharomyces cerevisiae.
3.2. Algemene parameters
3.2.1. Voedselinname
Dankzij de ‘pair feeding’ methode werd er gecorrigeerd voor voedselinname. Zo werd er
gezorgd dat zowel cerevisiae behandelde als placebo behandelde muizen quasi evenveel
voedsel innamen. In onderstaande figuur staat de gemiddelde voedsel inname per dag per
groep weergegeven (Figuur 3). Dit is voor wild type muizen met een ND 3,39 ± 0,38 g en
9,58 ± 1,07 kcal voor de placebo behandelde groep en 3,07 ± 0,31 g en 8,7 ± 0,87 kcal voor
de cerevisiae behandelde groep. Wild type muizen met een HFD aten 2,32 ± 0,41 g voor de
placebo en 2,52 ± 0,36 g voor de cerevisiae behandelde groep. Een HFD bevat echter een
hogere calorische inhoud dan een ND waardoor 12,1 ± 2,19 kcal en 13,23 ± 1,90 kcal
respectievelijk werden opgenomen. Foz/foz muizen met een ND aten gemiddeld gezien iets
minder dan WT muizen met een high fat van dezelfde leeftijd. Dit is 3,88 ± 0,36 g en 10,97 ±
1,04 kcal. De foz/foz muizen met een HFD aten meer dan hun soortgenoten met een ND en
WT tegenhangers met een HFD. Dit was 2,86 ± 0,37 g en 14,99 ± 1,92 kcal voor placebo
behandelde en 3,14 ± 0,37 g en 16,46 en 1,93 kcal voor cerevisiae behandelde muizen.

17
Figuur 3: Gemiddelde voedselinname. Gemiddelde voedselinname weergegeven in gram en kcal voor wild type (WT) muizen met normaal dieet (ND) of high fat dieet (HFD) en foz/foz muizen met een ND of HFD. Figuur geeft zowel hoeveelheid voor placebo behandelde dieren (PBS) als voor cerevisiae behandelde dieren (CER). Hoogste calorie-inname is voor foz/foz muizen met een HFD.
3.2.2. Lichaamsgewicht
Lichaamsgewicht is een belangrijke parameter voor het metabool foz/foz proefdiermodel. In
onderstaande figuur 4 is de relatieve lichaamsgewicht toename weergegeven. Het
lichaamsgewicht is toegenomen in alle groepen zoals verwacht. Het lichaamsgewicht van de
WT muizen met een ND op het einde van de behandeling is toegenomen met een relatieve
stijging van 16,23 ± 4,30 % voor placebo behandelde en 5,67 ± 4,13% cerevisiae behandelde
muizen. Cerevisiae behandelde dieren hadden een lager lichaamsgewicht op het einde van het
experiment maar het verschil was niet significant (p=0,507). De WT muizen met een HFD
hebben geen significant verschillend lichaamsgewicht indien ze een behandeling kregen of
niet. Op dag 40 is er een relatieve lichaamsgewicht stijging van 8,92 ± 5,29 % voor placebo
behandelde en 5,06 ± 8,60 % voor cerevisiae behandelde muizen (p=0.766). Dag 45 is dit
respectievelijk 8,36 ± 5,11% en 6,97 ± 10,1% (p=0.78).
Bij foz/foz muizen met een HFD is dit de grootste relatieve lichaamsgewichtstijging te
vinden. Hiertoe is er een relatieve lichaamsgewicht stijging van en 38,58 ± 21,76% voor
placebo behandelde en 54,16 ± 36,17% voor cerevisiae behandelde muizen geregistreerd. Er
is echter geen significant verschil waargenomen tussen foz/foz muizen met een HFD met en
zonder behandeling van cerevisiae op dag 40 (p=0.281) en dag 45 (p=0.325).

18
Figuur 4: Relatieve lichaamsgewichttoename. Dit gewichtstoename is ten opzichte van dag 1 voor alle wild type (WT) en foz/foz muizen met normaal dieet (ND) of high fat dieet (HFD). Foz/foz proefdieren met een HFD en cerevisiae behandeling (CER) hebben hoogste gewichtstoename. Placebo (PBS) behandelde foz/foz muizen met een HFD hebben een niet significant verschillende gewichtstoenames ten opzichte van CER behandeling (p>0,05).
In figuur 5 wordt het absolute lichaamsgewicht op het einde van het experiment
weergegeven. Hier is te zien dat WT muizen met een ND of HFD en foz/foz muizen met een
ND een gelijkaardig lichaamsgewicht hebben. Het absoluut lichaamsgewicht op het einde van
de behandeling is 23,0 ± 2,58 g voor placebo behandelde en 24,25 ± 4,95 g voor cerevisiae
behandelde WT muizen met een ND. De placebo behandelde en cerevisiae behandelde WT
muizen met een HFD hebben een absoluut lichaamsgewicht van respectievelijk 26,14 g ±
2,54 en 26,33 g ± 3,27. Ondanks de behandeling (placebo vs. cerevisiae) en het dieet (ND vs.
HFD) is geen statistisch significant verschil te vinden tussen de WT muizen (p=0,217).
Het lichaamsgewicht van de foz/foz muizen met een ND verschilt zelfs niet significant
met het lichaamsgewicht van WT muizen met hetzelfde dieet (p=0,637). Deze dieren hebben
een absoluut lichaamsgewicht van 28,0 ± 4,36 op het einde van het experiment. Dit laatste is
gezien de mutatie van foz/foz muizen toch eerder onverwacht. Foz/foz met een ND en WT
met een HFD hebben eveneens een gelijkwaardig lichaamsgewicht (p=0,798). De foz/foz
muizen met een HFD hebben duidelijk een groter lichaamsgewicht dan hun tegenhangers. Dit
is voor placebo groep 37,33 ± 7,18 g en behandelde 41,43 ± 8,9 g. Zo verschilt het

19
lichaamsgewicht van foz/foz muizen met een HFD met foz/foz muizen met een ND zoals
verwacht (p=0,04). Het lichaamsgewicht van foz/foz muizen met een HFD verschillen
eveneens significant met het lichaamsgewicht van WT muizen met een HFD (p<0,001) en
met het lichaamsgewicht van WT muizen met een ND (p<0,001)
Figuur 5: Absoluut lichaamsgewicht. Voor wild type (WT) muizen met een normaal dieet is dit na 5 weken behandeling met placebo (PBS) of cerevisiae (CER). Voor WT en foz/foz muizen is dit na 6 weken behandeling met PBS of CER. Foz/foz muizen met een HFD hebben een significant hoger lichaamsgewicht dan foz/foz muizen met een ND (p=0,04), dan WT muizen met een HFD (p<0,001) en dan WT muizen met een ND (p<0,001).
3.2.3. Glycemie
De glycemie werd wekelijks bijgehouden en op gelijkaardige tijdstippen gemeten. Figuur 6
toont de relatieve glycemietoename t.o.v. dag 1. Na vijf weken is er een relatieve daling
waargenomen van 25,17 ± 3,98 % bij placebo behandelde en 18,13 ± 9,89 % bij cerevisiae
behandelde WT muizen met een ND. Dit verschil van 7,04% tijdens de laatste week van hun
behandeling is echter niet statistisch significant verschillend. (p=0,900). De absolute
glycemiewaarde op het einde van de behandeling is 136,50 ± 1,41 mg/dl voor de placebo en
138,00 ± 16,25 mg/dl voor de cerevisiae behandelde muizen. Bij WT muizen met een HFD is
er respectievelijk een relatieve glycemiestijging en absolute glycemie van 24,6 ± 22,0 % en
189,71 ± 47, 76 mg/dl voor placebo en 37,58 ± 41,89 % en 202,50 ± 61,87 mg/dl voor
cerevisiae behandelde dieren waar te nemen op het einde van de behandeling. Hiertoe is er
geen significant verschil voor de twee behandelingen op week 5 (p=0,180) en week 6
(p=0,806) gevonden. Voor WT muizen is er eveneens geen significant verschil gevonden
tussen de twee diëten (p=0,543).
Foz/foz muizen met een ND hebben een relatieve glycemiestijging van 7,48 ± 5,14 %
doorgemaakt tot het einde van de placebo behandeling. De absolute glycemie is 172,33 ±

20
35,53 mg/dl. De glycemie van foz/foz muizen verschilden niet significant met zowel de
glycemiën van WT muizen met een ND (p=0,382) als die van WT muizen met een HFD
(p=0,868).
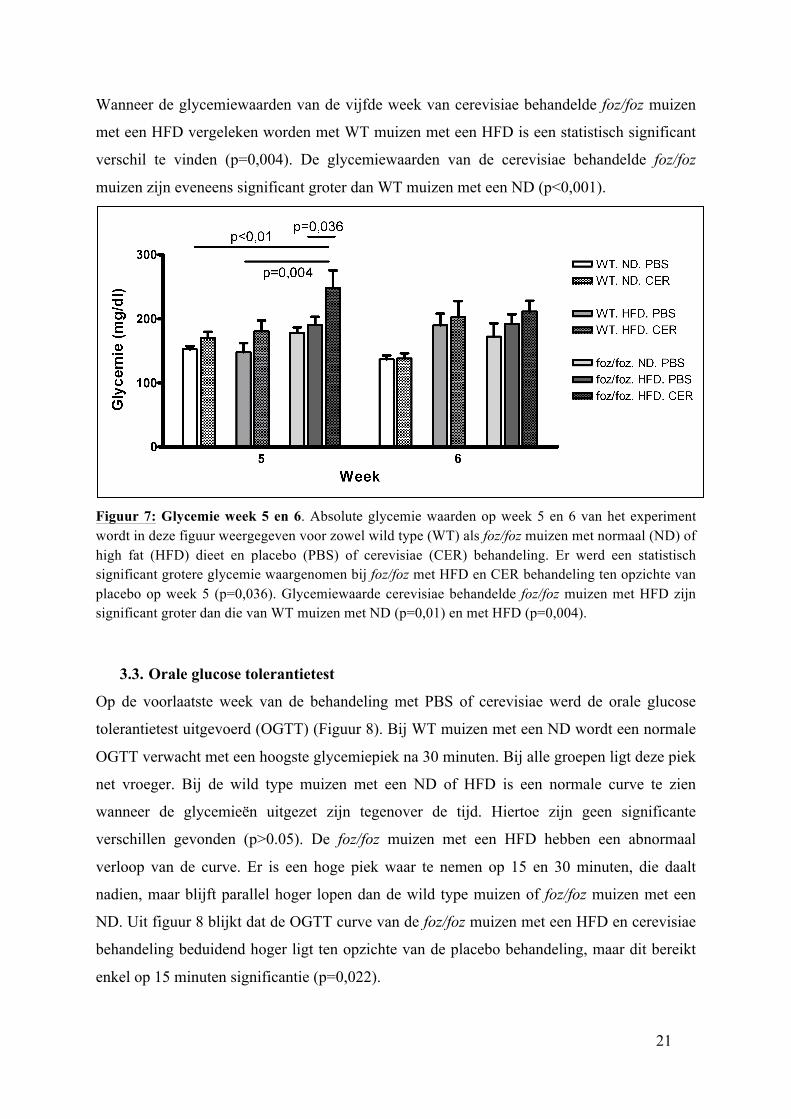
Figuur 6: Relatieve glycemietoename. Glycemietoename procentueel gezien ten opzichte van dag 1 wordt in deze figuur weergegeven voor zowel wild type (WT) als foz/foz muizen met een normaal (ND) of high fat (HFD) dieet en placebo (PBS) of cerevisiae (CER) behandeling. Er werd een statistisch significant grotere glycemie waargenomen bij foz/foz met een HFD en CER behandeling ten opzichte van placebo op week 5 (p=0,036).
Bij foz/foz muizen met een HFD is er echter wel een significant verschil gevonden op week
vijf tussen placebo en cerevisiae behandelde dieren (p=0,036). Hierbij is de glycemie (Figuur
7) en relatieve glycemie-toename (Figuur 6) van cerevisiae behandelde foz/foz muizen met
een HFD 248,43 ± 71,47 mg/dl en 65,51 ± 46,88 %. Laatst genoemde is significant groter
dan de glycemie 190,78 ± 36,94 mg/dl en relatieve glycemie-toename 24,12 ± 25,88 % bij
placebo behandelde foz/foz muizen met eenzelfde dieet. Dit verschil is niet meer te zien op
week 6 (p=0,401). Hierbij is de glycemie en relatieve glycemie toename 192,11 ± 44,41
mg/dl en 24,82 ± 30,22 % voor de placebogroep en 211,57 ± 43,73 mg/dl en 35,62 ± 27,03 %
voor cerevisia behandelde muizen.
21
Wanneer de glycemiewaarden van de vijfde week van cerevisiae behandelde foz/foz muizen
met een HFD vergeleken worden met WT muizen met een HFD is een statistisch significant
verschil te vinden (p=0,004). De glycemiewaarden van de cerevisiae behandelde foz/foz
muizen zijn eveneens significant groter dan WT muizen met een ND (p<0,001).
Figuur 7: Glycemie week 5 en 6. Absolute glycemie waarden op week 5 en 6 van het experiment wordt in deze figuur weergegeven voor zowel wild type (WT) als foz/foz muizen met normaal (ND) of high fat (HFD) dieet en placebo (PBS) of cerevisiae (CER) behandeling. Er werd een statistisch significant grotere glycemie waargenomen bij foz/foz met HFD en CER behandeling ten opzichte van placebo op week 5 (p=0,036). Glycemiewaarde cerevisiae behandelde foz/foz muizen met HFD zijn significant groter dan die van WT muizen met ND (p=0,01) en met HFD (p=0,004).
3.3. Orale glucose tolerantietest
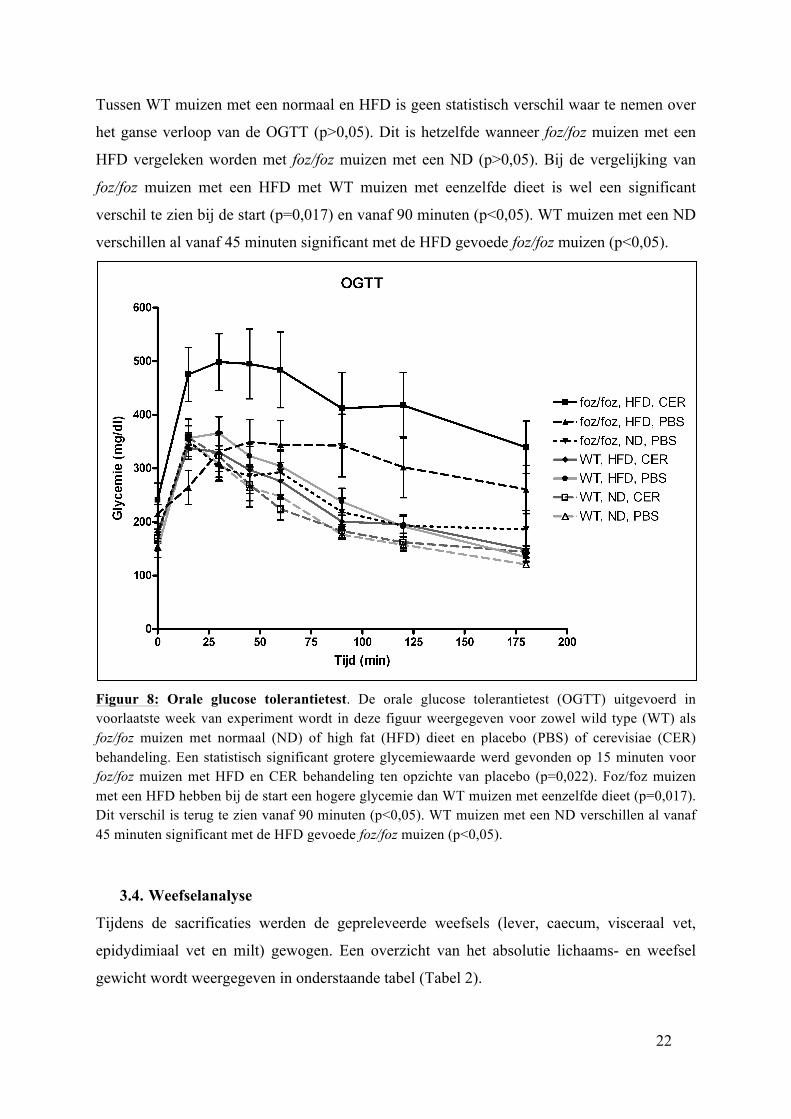
Op de voorlaatste week van de behandeling met PBS of cerevisiae werd de orale glucose
tolerantietest uitgevoerd (OGTT) (Figuur 8). Bij WT muizen met een ND wordt een normale
OGTT verwacht met een hoogste glycemiepiek na 30 minuten. Bij alle groepen ligt deze piek
net vroeger. Bij de wild type muizen met een ND of HFD is een normale curve te zien
wanneer de glycemieën uitgezet zijn tegenover de tijd. Hiertoe zijn geen significante
verschillen gevonden (p>0.05). De foz/foz muizen met een HFD hebben een abnormaal
verloop van de curve. Er is een hoge piek waar te nemen op 15 en 30 minuten, die daalt
nadien, maar blijft parallel hoger lopen dan de wild type muizen of foz/foz muizen met een
ND. Uit figuur 8 blijkt dat de OGTT curve van de foz/foz muizen met een HFD en cerevisiae
behandeling beduidend hoger ligt ten opzichte van de placebo behandeling, maar dit bereikt
enkel op 15 minuten significantie (p=0,022).
22
Tussen WT muizen met een normaal en HFD is geen statistisch verschil waar te nemen over
het ganse verloop van de OGTT (p>0,05). Dit is hetzelfde wanneer foz/foz muizen met een
HFD vergeleken worden met foz/foz muizen met een ND (p>0,05). Bij de vergelijking van
foz/foz muizen met een HFD met WT muizen met eenzelfde dieet is wel een significant
verschil te zien bij de start (p=0,017) en vanaf 90 minuten (p<0,05). WT muizen met een ND
verschillen al vanaf 45 minuten significant met de HFD gevoede foz/foz muizen (p<0,05).
Figuur 8: Orale glucose tolerantietest. De orale glucose tolerantietest (OGTT) uitgevoerd in voorlaatste week van experiment wordt in deze figuur weergegeven voor zowel wild type (WT) als foz/foz muizen met normaal (ND) of high fat (HFD) dieet en placebo (PBS) of cerevisiae (CER) behandeling. Een statistisch significant grotere glycemiewaarde werd gevonden op 15 minuten voor foz/foz muizen met HFD en CER behandeling ten opzichte van placebo (p=0,022). Foz/foz muizen met een HFD hebben bij de start een hogere glycemie dan WT muizen met eenzelfde dieet (p=0,017). Dit verschil is terug te zien vanaf 90 minuten (p<0,05). WT muizen met een ND verschillen al vanaf 45 minuten significant met de HFD gevoede foz/foz muizen (p<0,05).
3.4. Weefselanalyse
Tijdens de sacrificaties werden de gepreleveerde weefsels (lever, caecum, visceraal vet,
epidydimiaal vet en milt) gewogen. Een overzicht van het absolutie lichaams- en weefsel
gewicht wordt weergegeven in onderstaande tabel (Tabel 2).

23
Tabel 2 Overzicht absoluut lichaams- en weefselgewicht. Het absolute weefselgewicht, gewogen tijdens sacrificaties wordt in deze tabel weergegeven voor zowel wild type (WT) als foz/foz muizen met normaal (ND) of high fat (HFD) dieet en placebo (PBS) of cerevisiae (CER) behandeling.
WT, ND, PBS
WT, ND, CER
WT, HFD, PBS
WT, HFD, CER
Foz/foz, ND, PBS
Foz/foz, HFD, PBS
Foz/foz, HFD, CER
Absoluut lichaams-gewicht
21,38 ± 0,25 g
22,20 ±2,40 g
27,25 ± 2,64 g
27,49 ± 2,78 g
27,68 ± 4,26 g
36,74 ± 6,37 g
39,77 ± 5,52 g
Lever 0,83 ± 0,23 g
0,87 ± 0,29 g
1,09 ± 0,10 g
1,06 ± 0,11 g
1,32 ± 0,29 g
1,37 ± 0,22 g
1,66 ± 0,21 g
Caecum 0,15 ± 0,01 g
0,19 ± 0,06 g
0,06 ± 0,01 g
0,11 ± 0,02 g
0,31 ± 0,06 g
0,16 ± 0,05 g
0,23 ± 0,05 g
Visceraal vet 0,22 ± 0,06 g
0,31 ± 0,12 g
0,48 ± 0,11 g
0,51 ± 0,10 g
0,39 ± 0,08 g
0,77 ± 0,26 g
0,78 ± 0,14 g
Epidydimiaal vet
0,32 ± 0,08 g
0,36 ± 0,17 g
1,15 ± 0,27 g
0,95 ± 0,30 g
0,79 ± 0,34 g
2,42 ± 1,07 g
2,51 ± 0,43 g
Milt 0,07 ± 0,01 g
0,06 ± 0,01 g
0,06 ± 0,009 g
0,09 ± 0,02 g
0,08 ± 0,01 g
0,23 ± 0,005 g
0,1 ± 0,001 g
Er is bij het eerste orgaan, de lever, geen significant behandelingseffect waar te nemen bij het
vergelijken van de procentuele verhouding van de lever op het lichaamsgewicht van WT
muizen met een ND, WT muizen met een HFD en foz/foz muizen met een HFD
(respectievelijk: p=0,720; p=0,601 en p=0,102). Er is toch een neiging tot hoger levergewicht
bij de HFD gevoede foz/foz muizen. Tussen de WT en foz/foz muizen met hun verschillende
diëten zijn er eveneens geen constateringen te doen ondanks de gegeven behandeling (Figuur
9). De levers van WT muizen met een HFD en ND verschillen niet significant van elkaar
(p=0,993). Wanneer deze, leververhouding van WT muizen met ND of HFD, vergeleken
worden met de ratio lever op lichaamsgewicht van foz/foz muizen met een ND is er nog altijd
geen significantie te vinden (p>0,05). De ratio van lever op lichaamsgewicht van de foz/foz
muizen met een HFD is even groot als deze van WT muizen met een HFD (p=1,00) en ook
helemaal niet verschillend met deze van WT muizen met een ND (p=0,990).
Wanneer de verhouding caecumgewicht op lichaamsgewicht wordt vergeleken tussen
placebo behandelde en cerevisiae behandelde WT muizen en een ND is er slechts een klein
niet statistische significant verschil te zien (p=0,118). Bij WT muizen met een HFD is dit wel
het geval. De verhouding caecum/lichaamsgewicht van placebo behandelde muizen is kleiner
dat die van cerevisiae behandelde muizen (p=0,015). De behandeling met cerevisiae had geen

24
significant effect op het gewicht van het caecum bij foz/foz muizen met een HFD t.o.v. de
placebo behandeling (p=0,352). Tussen WT muizen met een ND of HFD en foz/foz muizen
met een ND of HFD zijn er onderling geen significante verschillen waar te nemen (p>0,05).
Visceraal en epidydimiaal vetweefsel werd afgenomen en gewogen om zo een beeld te
kunnen krijgen van het vetpercentage in het lichaam van de muizen. De behandeling met
cerevisiae of PBS had geen statistisch effect op zowel het visceraal als epidydimiaal
vetweefsel. Dit is te zien zowel bij WT muizen met een ND (p=0,406 en p=0,827) of HFD
(p=0,114 en p=0,186) als bij foz/foz muizen met HFD (p=0,697 en p=0,934).
Omdat er geen effect is te zien van de behandeling kan er naar het effect van het dieet
en muismodel gekeken worden. WT muizen met een ND hebben zoals verwacht een lagere
vetverhouding, voor zowel visceraal als epidydimiaal vet, dan WT muizen met een HFD
(p=0,049 en p<0,001). Eenzelfde waarneming kan gedaan worden bij foz/foz muizen. Hiertoe
zorgt het HFD t.o.v. een ND voor een hoger epidydimiaal vetgehalte (p=0,009) maar niet
voor een hoger visceraal vetgehalte (p=0,298). Zo zijn er ook verschillen te zien tussen
foz/foz muizen met een HFD en WT muizen met een ND (p=0,01 en p<0,001). Opmerkelijk
is dat er tussen foz/foz muizen met een ND en WT muizen met een HFD geen statistisch
significant verschil te zien is voor zowel de hoeveelheid visceraal als epidydimiaal vetweefsel
(p=0,577 en p=0,450). Er is bijkomstig ook geen hogere hoeveelheid vetmassa te zien bij
foz/foz muizen met een ND en WT muizen met een hetzelfde dieet (p=0,893 en p=0,280).
Hoewel WT muizen met een verschillend dieet wel een verschillend vetmassa hebben.
Foz/foz muizen met een HFD hebben geen hogere hoeveelheid visceraal vetweefsel
(p=0,890) dan WT muizen met een HFD maar wel een hogere epidydimiale vetmassa
(p=0,007).
Bij de vergelijking van het gewicht van de milt met het lichaamsgewicht tussen de placebo
behandelde en cerevisiae behandelde WT muizen met een ND is er geen significant verschil
te vinden (p=0,233). Hetzelfde is te zien bij foz/foz muizen met een HFD (p=0,422). Dit is
echter niet het geval voor WT muizen met een HFD. Hiertoe is de verhouding van milt op
lichaamsgewicht bij cerevisiae behandelde muizen significant groter dan placebo behandelde
muizen (p=0,017). Als er gekeken wordt naar het effect van het dieet en diermodel is er geen
verschil te zien tussen de verhouding milt op lichaamsgewicht bij zowel WT muizen met een
ND of HFD en foz/foz muizen met een ND of HFD (p>0,05).

25
Figuur 9: Procentuele weefselverhouding. De procentuele verhouding van weefsels ten opzichte van absoluut lichaamsgewicht tijden de sacrificaties wordt in deze figuur weergegeven voor zowel wild type (WT) als foz/foz muizen met normaal (ND) of high fat (HFD) dieet en placebo (PBS) of cerevisiae (CER) behandeling. Een significant groter caecum werd gevonden bij WT muizen met een CER behandeling en HFD ten opzichte van placebo (p=0,015). Verhouding visceraal vetweefsel is significant groter bij WT muizen met een HFD t.o.v. een ND (p=0,49) en bij foz/foz muizen met een HFD t.o.v. WT muizen met een ND (p=0,01). Verhouding epidydimiaal vet is groter bij WT muizen met een HFD t.o.v. een ND (p<0,001), bij foz/foz muizen met een HFD t.o.v. WT muizen met een ND (p<0,001), bij foz/foz muizen met een HFD t.o.v. WT muizen met een ND (p=0,007) en bij foz/foz muizen met een HFD t.o.v. een ND (p=0,009)
3.4.1. Histologie
Voor de histologie werd gebruik gemaakt van een hematoxyline en eosine kleuring (Figuur
10). Foz/foz muizen met een HFD en behandeld met zowel placebo als cerevisiae tonen een
beeld van lichte steatose met milde ballooning of cytoplasmatische steatose. De andere
groepen, WT muizen met een ND en HFD en foz/foz muizen met een ND vertonen geen van
steatose. Deze bevindingen zijn preliminair en dienen nog door een leverpatholoog te worden
geverifieerd.

26
Figuur 10: Leverhistologie. Hematoxyline Eosine leverkleuring, (vergroting 200x) Waarbij A= WT, ND, PBS met geen aanwezige steatose en ballooning; B= WT, ND, CER met geen aanwezige steatose en ballooning; C= WT, HFD, PBS met geen aanwezige steatose en ballooning; D= WT, HFD, CER met geen aanwezige steatose en ballooning; E= foz/foz, ND, PBS met geen aanwezige steatose en ballooning; F= foz/foz, HFD, PBS met lichte steatose aanwezig en milde ballooning; G= foz/foz, HFD, CER met cytoplasmatische steatose aanwezig
A B
C D
E F
G
27
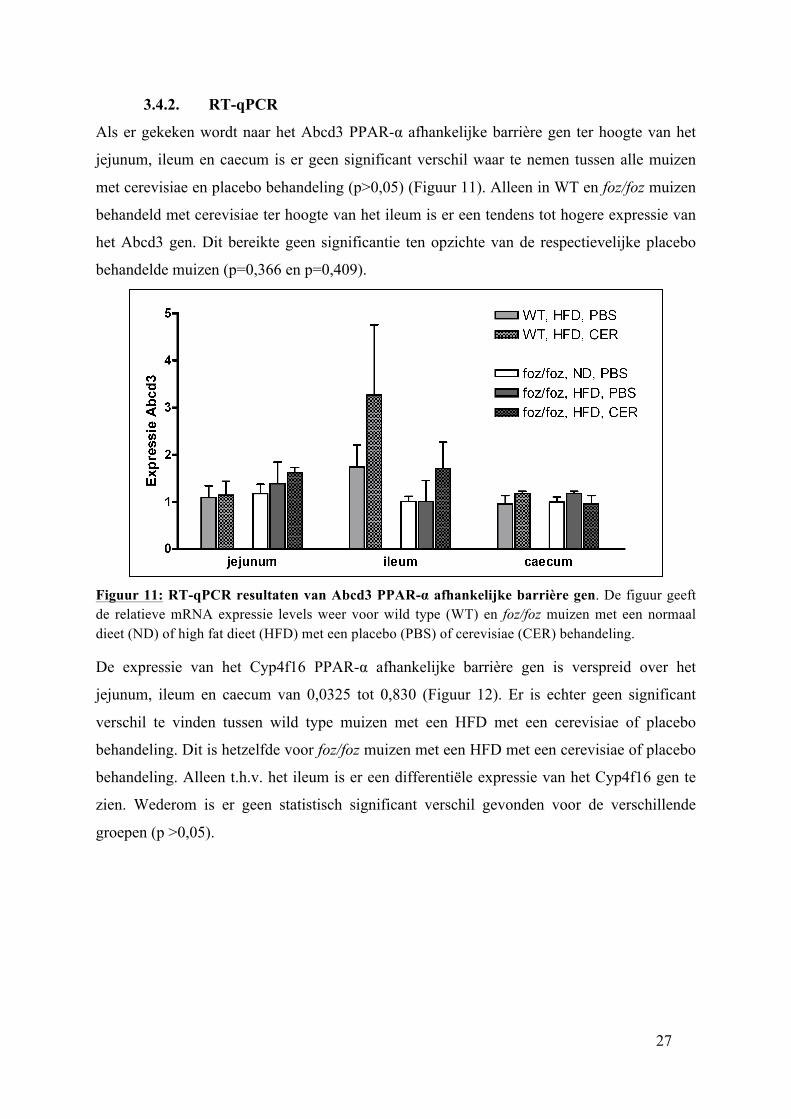
3.4.2. RT-qPCR
Als er gekeken wordt naar het Abcd3 PPAR-α afhankelijke barrière gen ter hoogte van het
jejunum, ileum en caecum is er geen significant verschil waar te nemen tussen alle muizen
met cerevisiae en placebo behandeling (p>0,05) (Figuur 11). Alleen in WT en foz/foz muizen
behandeld met cerevisiae ter hoogte van het ileum is er een tendens tot hogere expressie van
het Abcd3 gen. Dit bereikte geen significantie ten opzichte van de respectievelijke placebo
behandelde muizen (p=0,366 en p=0,409).
Figuur 11: RT-qPCR resultaten van Abcd3 PPAR-α afhankelijke barrière gen. De figuur geeft de relatieve mRNA expressie levels weer voor wild type (WT) en foz/foz muizen met een normaal dieet (ND) of high fat dieet (HFD) met een placebo (PBS) of cerevisiae (CER) behandeling.
De expressie van het Cyp4f16 PPAR-α afhankelijke barrière gen is verspreid over het
jejunum, ileum en caecum van 0,0325 tot 0,830 (Figuur 12). Er is echter geen significant
verschil te vinden tussen wild type muizen met een HFD met een cerevisiae of placebo
behandeling. Dit is hetzelfde voor foz/foz muizen met een HFD met een cerevisiae of placebo
behandeling. Alleen t.h.v. het ileum is er een differentiële expressie van het Cyp4f16 gen te
zien. Wederom is er geen statistisch significant verschil gevonden voor de verschillende
groepen (p >0,05).

28
Figuur 12: RT-qPCR resultaten van Cyp4f16 PPAR-α afhankelijke barrière gen. De figuur geeft de relatieve mRNA expressie levels weer voor wild type (WT) en foz/foz muizen met een normaal dieet (ND) of high fat dieet (HFD) met een placebo (PBS) of cerevisiae (CER) behandeling.
Op onderstaande figuur is de expressie van het Slc27a4, een PPAR-α afhankelijk barrière
gen, weergegeven ter hoogte van het jejunum, ileum en caecum (Figuur 13). Wanneer
gekeken wordt naar het ileum is een ideale verhouding van expressie van het Slc27a4 gen te
zien tussen de verschillende muismodellen, diëten en behandelingen. Er is een significant
grotere expressie van het Slc27a4 gen te zien bij foz/foz muizen met een HFD en cerevisiae
behandeling in vergelijking met foz/foz muizen met een placebo behandeling en een HFD
(p=0,015). Eenzelfde niet significante trend in het ileum wordt gezien bij de WT muizen met
een HFD behandeld met placebo of cerevisiae (p=0,608). Er wordt wel een significant
behandelingseffect waargenomen bij WT muizen met een HFD ter hoogte van het jejunum
(p=0,009). Ter hoogte van het caecum werd voor WT muizen met een HFD een bijna
statistisch significante grotere expressie te zien bij de placebo behandelde dieren in
vergelijking met de cerevisiae behandelden (p=0,066).

29
Figuur 13 RT-qPCR resultaten van Slc27a4 PPAR-α afhankelijke barrière gen. De figuur geeft de relatieve mRNA expressie levels weer voor wild type (WT) en foz/foz muizen met een normaal dieet (ND) of high fat dieet (HFD) met een placebo (PBS) of cerevisiae (CER) behandeling. Er is een significant hogere expressie van het Slc27a4 gen te zien ter hoogte van het jejunum bij de cerevisiae behandelde WT muizen met een HFD t.o.v. placebo (p=0,009). Bij foz/foz muizen met een HFD en cerevisiae behandeld is een significant hogere expressie van het Slc27a4 gen te zien t.o.v. placebo (p=0,015)
3.5. FITC dextran permeabiliteitstest
Het resultaat van de permeabiliteitstest is voor WT muizen met een HFD en
placebobehandeling 0,002793 ± 0,00069 mg/dl en voor cerevisiae behandeling 0,007464
mg/dl. Bij foz/foz muizen met een HFD is dit 0,004469 ± 0,00051 mg/dl voor de
placebobehandeling en 0,003858 ± 0,00177 mg/dl voor de cerevisiae behandeling. Hoewel
minder FITC dextran werd gemeten bij cerevisiae behandelde foz/foz muizen is geen verschil
te zien tussen de placebo behandelde en cerevisiae behandelde dieren.
3.6. Bacteriële translocatie
Twaalf uur na het uitplaten van de viscerale lymfeknoop werd op geen enkel lp-plaat een
kolonie waargenomen. Na 72 uur in de incubator op 37°C werden voor WT muizen met een
HFD gemiddeld 14 ± 22,54 kolonies voor de placebo en 1 kolonie voor de cerevisiae
behandelde muizen waargenomen. Bij de foz/foz muizen met een HFD werden gemiddeld 1,5
± 2,38 kolonies voor de placebo en 4,33 ± 4,04 kolonies voor de cerevisiae behandelde
muizen gevonden. Hiertoe zijn er geen significante verschillen waar te nemen (p>0,05). Bij
de gramkleuring werden oninterpreteerbare resultaten gevonden.

30
4. Bespreking
4.1. Steatose muismodel
In deze studie werd een steatose muismodel opgezet door een HFD te geven aan foz/foz
muizen. Histologisch gezien volgens een H&E kleuring op leverweefsel is er meer vet
aanwezig bij het foz/foz muismodel dan alle WT muizen en foz/foz muizen met een ND. De
viscerale vetverhouding is daarentegen niet groter bij foz/foz muizen t.o.v. WT muizen met
hetzelfde HFD. Bij de epidydimiale vetverhouding is dit wel te zien. Hiertoe is er duidelijk
significantie naar een hogere vetverhouding bij het foz/foz muismodel (figuur 9). De levers
van foz/foz muizen met een HFD en cerevisiae behandeling zijn niet significant zwaarder dan
die van hun complementaire placebo’s (p=0,102). Hoewel verwacht werd dat deze lichter
zouden zijn. Er is dus een steatose model te zien bij de placebo en cerevisiae behandelde
foz/foz muizen met een HFD. De leverhistologie en het levergewicht laten niet duidelijk
blijken of er een PPAR-α effect heeft plaatsgevonden ter hoogte van de lever. Histologisch
gezien lijkt het erop dat er meer ballooning te zien is in de levers van de foz/foz muizen en
HFD met de placebo behandeling. Bij foz/foz muizen en HFD en cerevisiae behandeling lijkt
er meer een cytoplasmatisch vet aanwezig te zijn. Deze bevindingen zijn preliminair en
dienen nog door een leverpatholoog te worden geverifieerd. Het effect van cerevisiae op de
lobulaire inflammatie en de hepatocytaire ballooning dient nog te worden bekeken.
4.2. Metabole parameters
Lichaamsgewicht is een uitstekende parameter voor controleren van het welzijn van de
proefdieren. In het metabool proefdiermodel foz/foz is dit bijkomstig een belangrijke factor
om de metabole veranderingen bij te houden. Zoals verwacht is het lichaamsgewicht van
foz/foz muizen met HFD groter dan foz/foz en WT muizen met een ND of HFD. Wat echter
opviel is de gewichtstoename van de muizen met de cerevisiae behandeling. Deze lag hoger
dan hun complementaire placebo’s maar dit verschil is echter niet significant. Belangrijk is
dat het kleine visuele verschil niet kan door de voedselinname omwille van de pair feeding
condities.
Hoe ouder foz/foz muizen worden hoe meer ze toenemen in lichaamsmassa en op een
leeftijd van 100 tot 120 dagen zijn ze obees. Bovendien zijn ze vanaf dit moment
insulineresistent en zijn er hogere glucosewaarden in het bloed waarneembaar (6). In de
huidige studie zijn er tijdens de voorlaatste week van de behandeling met cerevisiae zijn er
significant hogere glycemiewaarden gevonden bij de foz/foz muizen met een HFD in

31
vergelijking met placebo behandelde muizen. De muizen zijn op dat moment bijna 100 dagen
oud. Het lijkt er op dat de foz/foz muizen met een HFD en cerevisiae behandeling eerder dan
hun complementaire placebo’s metabole veranderingen ondergaan. Dit fenomeen wordt
bevestigd met de orale glucose tolerantietest. Bij normale muizen met een ND wordt een
normale OGTT verwacht met een hoogste glycemiepiek na 30 minuten. Bij alle groepen ligt
deze piek net vroeger. Een hoogste piek wordt waargenomen bij foz/foz muizen met een HFD
en een cerevisiae behandeling. Het verschil is enkel op 15 minuten significant na de orale
glucose belasting. In het verder verloop van de test is een merkbaar niet significant verschil
tussen de foz/foz muizen met een placebobehandeling en cerevisiae behandeling (Figuur 8).
In een experiment uitgevoerd bij de validering van de foz/foz muis werd eveneens een orale
glucose tolerantietest uitgevoerd (6). Net zoals in deze huidige studie wordt een verminderde
glucose tolerantie gevonden bij prediabetische foz/foz muizen in vergelijking met WT
muizen. Wanneer onze gegeven worden bekeken kan er een bijkomend effect worden
genoteerd. Heeft de behandeling met cerevisiae een bijkomstig effect op de glucose
gevoeligheid en/of insulineresistentie waardoor de muizen sneller een verstoorde glycemie en
een insulineresistentie ontwikkelden dan hun respectievelijke placebo’s? Hiervoor zijn
uitgebreide analyses en onderzoeken nodig. Verschillende mechanismen zijn mogelijk, maar
glucoseopname wordt immers ook door o.a. PPAR-α gereguleerd. Zo weten we uit de
literatuur dat de expressie van de apicale glucose opname transporter (Sglt1) en de basale
glucose transporter (Glut2) beiden verminderd worden door PPAR-α inductie. In deze
huidige studie werden echter hogere bloedglucosewaarden teruggevonden bij foz/foz muizen
die een cerevisiae behandeling en dus PPAR-α opregulatie ondergingen, ondanks dat er een
lokaal PPAR-α effect is (Slc27a4 proteïnen kwamen meer tot expressie). Echter vele andere
mechanismen zijn betrokken in de PPAR-α inductie en glucosehomeostase zoals o.a. de
PPAR-α interactie met inflammatoire mediatoren (TNF-α, IL-6) (21), de link van PPAR-α
met galzoutenmetabolisme (22) en het effect van probiotica op insuline sensitiviteit via
transcriptie van glucosetransporters (4). Dit zal het onderwerp van een volgende masterthesis
worden.
Een andere opmerkelijke bevinding, in deze huidige studie, is dat de veranderde
glucosehuishouding alleen optreedt in foz/foz muizen met een HFD en cerevisiae behandeling
terwijl de respectievelijke WT muizen dit niet vertonen. Dit zou er kunnen op wijzen dat de
genetische achtergrond van de muizen een noodzakelijke factor is om de glycemie stijging tot
stand te brengen. De specifieke pathogenese van Alms1 deficiëntie bij foz/foz muizen is nog

32
niet volledig opgehelderd. Preliminaire data uit het lab van Prof. dr. Lerclercq doen ook een
veranderde intestinale brush border functie vermoeden die bv. mogelijks door de darmflora
kan worden gemoduleerd (23).
4.3. Cerevisiae behandeling en microbiota
Een belangrijk element in deze studie is de vraag of de cerevisiae behandeling inderdaad de
microbiota in de darm heeft veranderd. Om dit na te gaan zal een high-throughput methode
voor de darmflora karakterisatie moeten gebruikt worden (zoals pyrosequentie analyse). Een
indirecte maat voor de microbiota is het gewicht van het caecum. Het feit dat de
gistbehandeling het caecumgewicht verhoogt bij de WT muizen met een HFD, betekent
inderdaad dat de behandeling de microbiota heeft veranderd. Wetende dat het HFD normaal
het caecum gewicht drastisch verminderd in vergelijking met een ND, maakt het effect van
cerevisiae op het caecum des te opmerkelijker. Het caecumgewicht van alle muizen met een
cerevisiae behandeling is groter dan de complementaire placebo’s ervan. Bij foz/foz muizen
met een HFD en cerevisiae behandeling is het caecum zelf zwaarder dan de foz/foz muizen
met een ND en placebo behandeling. Hieruit kan zeker besloten worden dat de cerevisiae de
darmflora van de muizen beïnvloed heeft (24).
4.4. Effect van cerevisiae op PPAR-α afhankelijke genen in de darm
Het effect van de gist werd nagekeken via RT-qPCR. Er is geweten uit het werk van Prof. dr.
Desmeuraux et al. dat cerevisiae een potentieel PPAR-α effect heeft (gepubliceerd in
abstractvorm). De PPAR-α afhankelijke intestinale barrière proteïnen (Abcd3, Cyp4f16 en
Slc27a4) werden daarom bestudeerd. Deze genen zijn betrokken bij de selectieve absorptie
van nutriënten en andere voedselcomponenten in de darmen (16). Enkel Slc27a4 loste het
verwachtingspatroon duidelijk in tussen de behandelde en placebo-behandelde WT en foz/foz
muizen ter hoogte van het jejunum en ileum (Figuur 14). In de studie van de Vogel-van den
Bosch werd eveneens de intestinale expressie van de Slc27a4, ook Fatp4 genaamd, nagegaan
na toediening van een van de PPAR-α agonisten (WY 14 643, OA, EPA en DHA) ten
opzichte van de basale activatie t.h.v. het ileum. Figuur 15 geeft een voor elk van de
agonisten de relatieve mRNA levels weer waarbij de dunne darm in 10 stukken is verdeeld.
Ten eerste wordt er gezien dat de Slc27a4 niet homogeen verspreid is doorheen de darm.
Bovendien zijn de Slc27a4 mRNA levels het hoogst in het jejunum na PPAR-α agonisme
volgens onderstaande figuur. In deze huidige studie werd naar analogie met figuur 15 een
hoogste expressie van Slc27a4 verwacht ter hoogte van het jejunum voor foz/foz muizen.
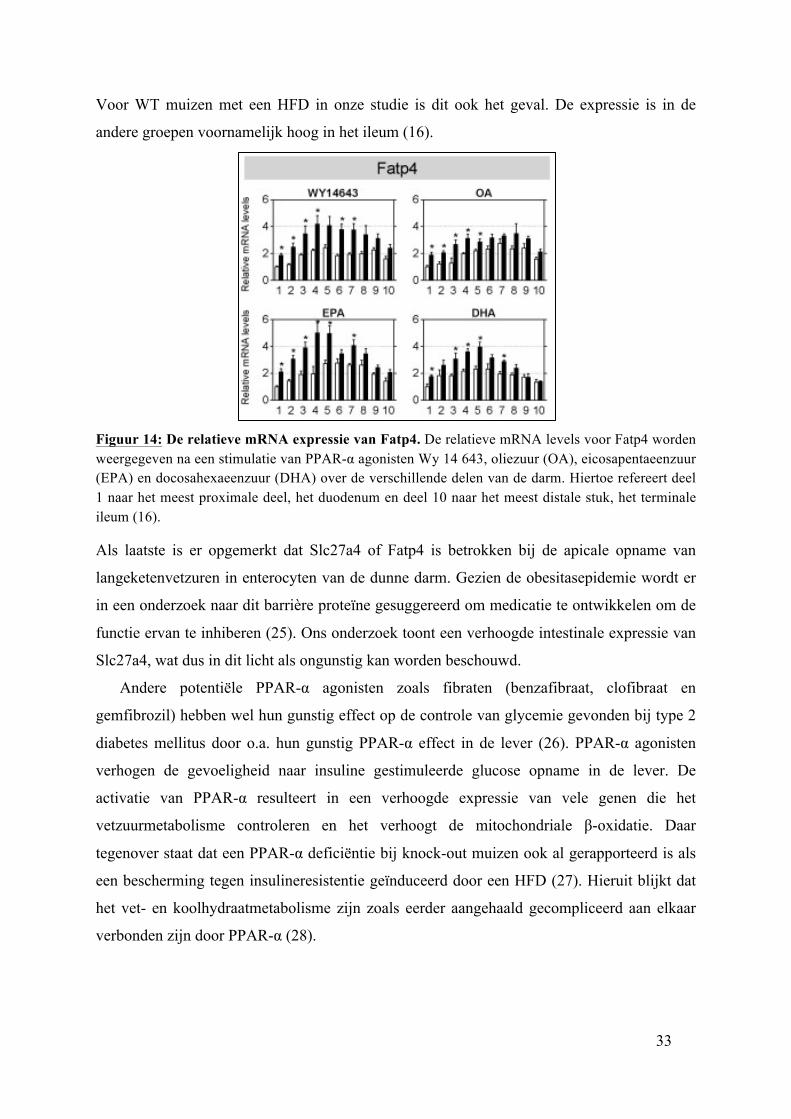
33
Voor WT muizen met een HFD in onze studie is dit ook het geval. De expressie is in de
andere groepen voornamelijk hoog in het ileum (16).
Figuur 14: De relatieve mRNA expressie van Fatp4. De relatieve mRNA levels voor Fatp4 worden weergegeven na een stimulatie van PPAR-α agonisten Wy 14 643, oliezuur (OA), eicosapentaeenzuur (EPA) en docosahexaeenzuur (DHA) over de verschillende delen van de darm. Hiertoe refereert deel 1 naar het meest proximale deel, het duodenum en deel 10 naar het meest distale stuk, het terminale ileum (16).
Als laatste is er opgemerkt dat Slc27a4 of Fatp4 is betrokken bij de apicale opname van
langeketenvetzuren in enterocyten van de dunne darm. Gezien de obesitasepidemie wordt er
in een onderzoek naar dit barrière proteïne gesuggereerd om medicatie te ontwikkelen om de
functie ervan te inhiberen (25). Ons onderzoek toont een verhoogde intestinale expressie van
Slc27a4, wat dus in dit licht als ongunstig kan worden beschouwd.
Andere potentiële PPAR-α agonisten zoals fibraten (benzafibraat, clofibraat en
gemfibrozil) hebben wel hun gunstig effect op de controle van glycemie gevonden bij type 2
diabetes mellitus door o.a. hun gunstig PPAR-α effect in de lever (26). PPAR-α agonisten
verhogen de gevoeligheid naar insuline gestimuleerde glucose opname in de lever. De
activatie van PPAR-α resulteert in een verhoogde expressie van vele genen die het
vetzuurmetabolisme controleren en het verhoogt de mitochondriale β-oxidatie. Daar
tegenover staat dat een PPAR-α deficiëntie bij knock-out muizen ook al gerapporteerd is als
een bescherming tegen insulineresistentie geïnduceerd door een HFD (27). Hieruit blijkt dat
het vet- en koolhydraatmetabolisme zijn zoals eerder aangehaald gecompliceerd aan elkaar
verbonden zijn door PPAR-α (28).

34
4.5. PPAR-α en extra-intestinale effecten
Preliminaire data van onze studie tonen een effect op de leverhistologie (cfr. supra). In een
bijkomende andere studie met een PPAR-α agonist (WY 14 643) werd ook een gunstig effect
op de levers van foz/foz muizen gevonden. Het verbeterde de metabolische indexen, steatose,
ballooning en leverinflammatie in diabetische muizen met NASH. De behandeling had
bovendien een gunstig effect op de insuline gevoeligheid en induceerde een verminderd
lichaamsgewicht (19). Een kritische bemerking is het bestaan van een mogelijks effect op
genetische pathways (bv. PPAR-α) secundair aan het gebruik van het genetisch foz/foz
muismodel. Het is namelijk gekend dat bij een genetisch gemanipuleerd muismodel soms
andere genen door de deletie niet meer kunnen functioneren.
Bovendien is het opmerkelijk dat orale toediening van een gist met een lokaal intestinaal
effect ook invloed kan uitoefenen op een verder gelegen orgaan. Het is echter reeds gekend
dat een probioticum de lever, skeletspieren of vetweefsel kan beïnvloeden (9).
Misschien is er ook een dosis-effect te verwachten. In deze huidige studie werd een dosis
van 5 x 108 k.v.e. per dag gegeven. Desmeuraux et al. voerde een dosis-respons experiment
uit waarbij een 106 k.v.e. per dag toediening een nociceptief effect en PPAR-α inductie
vertoonde (29).
4.6. NAFLD en intestinale permeabiliteit
Niet alcoholische vette leverziekte is gekoppeld aan een gestegen intestinale permeabiliteit.
Dit wordt veroorzaakt door de onderbreking van intercellulaire tight junctions in de darmen.
Wild type en foz/foz muizen met een HFD hebben een grotere intestinale permeabiliteit t.o.v.
WT en foz/foz muizen met een ND, zoals blijkt uit ongepubliceerd doctoraatswerk van
Laurance Poekes uit het UCL laboratorium. Zij voerde een permeabiliteitstest met FITC
dextran uit bij foz/foz muizen en wild type muizen. Dit is weergegeven in figuur 15, waarbij
er verschil te zien is tussen muizen met een ND en een HFD. In onze huidige studie is er geen
verschil in permeabiliteit te zien tussen de placebobehandeling en de cerevisiae behandeling.
Aan de hand van FITC dextran kon er dus geen verschil worden vastgesteld, alhoewel er wel
een verschil in barrière genen (Slc27a4 expressie) te zien was ter hoogte van het ileum bij
foz/foz muizen en ter hoogte van het jejunum bij WT muizen.
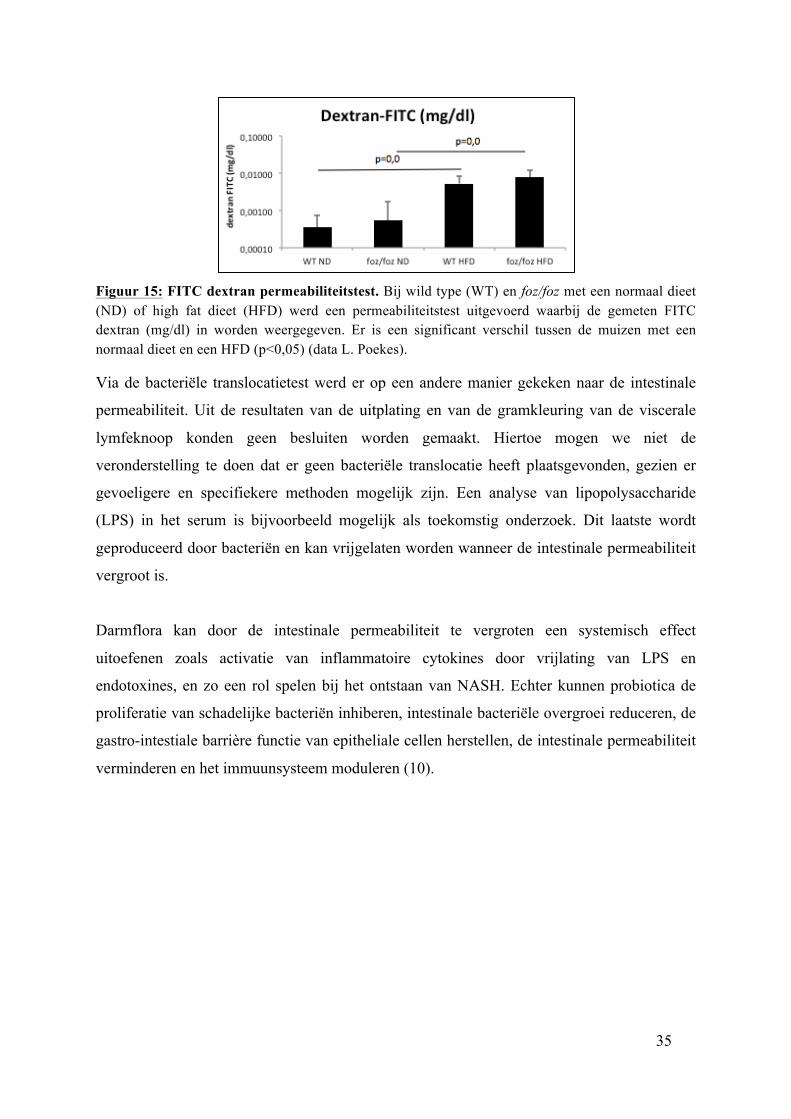
35
Figuur 15: FITC dextran permeabiliteitstest. Bij wild type (WT) en foz/foz met een normaal dieet (ND) of high fat dieet (HFD) werd een permeabiliteitstest uitgevoerd waarbij de gemeten FITC dextran (mg/dl) in worden weergegeven. Er is een significant verschil tussen de muizen met een normaal dieet en een HFD (p<0,05) (data L. Poekes).
Via de bacteriële translocatietest werd er op een andere manier gekeken naar de intestinale
permeabiliteit. Uit de resultaten van de uitplating en van de gramkleuring van de viscerale
lymfeknoop konden geen besluiten worden gemaakt. Hiertoe mogen we niet de
veronderstelling te doen dat er geen bacteriële translocatie heeft plaatsgevonden, gezien er
gevoeligere en specifiekere methoden mogelijk zijn. Een analyse van lipopolysaccharide
(LPS) in het serum is bijvoorbeeld mogelijk als toekomstig onderzoek. Dit laatste wordt
geproduceerd door bacteriën en kan vrijgelaten worden wanneer de intestinale permeabiliteit
vergroot is.
Darmflora kan door de intestinale permeabiliteit te vergroten een systemisch effect
uitoefenen zoals activatie van inflammatoire cytokines door vrijlating van LPS en
endotoxines, en zo een rol spelen bij het ontstaan van NASH. Echter kunnen probiotica de
proliferatie van schadelijke bacteriën inhiberen, intestinale bacteriële overgroei reduceren, de
gastro-intestiale barrière functie van epitheliale cellen herstellen, de intestinale permeabiliteit
verminderen en het immuunsysteem moduleren (10).

36
5. Kritische bemerkingen en slotconclusies
Het probioticum Cerevisia wordt momenteel als supplement voorgeschreven voor mensen
met een prikkelbare darm of irritable bowel syndroom (IBS). Het verbetert abdominale pijn
en verteringsdiscomfort. Door zijn anti-inflammatoire activiteit in experimentele colitis
muizen, visceraal analgetische effecten in een ratmodel van colon hypersensitiviteit en een
antagonistische werking op E. Coli O157:H7 is het een interessante gist om onderzoek op uit
te voeren (11–13).
Over de bijwerkingen van dit voedingssupplement is echter niets te vinden. Om een hoge
concentratie in de darm te blijven houden, is het nodig om tweemaal per dag een capsule in te
nemen volgens een onderzoek naar de overleving naar Saccharomyces cerevisiae CNCM I-
3856 in het gastro-intestinale stelsel. Bovendien is er door voedsel inname een vertraging in
de gistvrijlating (14). Probiotica kunnen slechts gunstige effecten uitoefenen als ze inderdaad
levend aanwezig zijn in het verteringsstelsel. Die overleving is afhankelijk van de
toedieningswijze, de dosis en het dieet (4). In onze studie speelt dit laatstgenoemde een
belangrijke rol. Naast het vetrijke dieet en de aangeboren genetische voorbestemming tot het
ontwikkelen van obesitas en insuline resistentie, blijkt de inname van dagelijks
Saccharomyces cerevisiae de metabolische stoornis van de foz/foz muizen te stimuleren. Uit
de resultaten van onze studie lijkt het daarom ook geen goed optie om de gist te geven als
supplement aan mensen met leverproblemen door een Westers dieet, alhoewel natuurlijk
translatie van een muismodel naar de humane setting met zeer veel voorzichtigheid dient te
worden uitgevoerd.

37
6. Referentielijst
1. Day C. (2002). Non-alcoholic steatohepatitis (NASH): where are we now and where are we going? Gastroenterology 50:585–8.
2. Green RM. (2003). NASH--hepatic metabolism and not simply the metabolic syndrome. Hepatology 38(1):14–7.
3. Sheth SG, Gordon FD, Chopra S. (1997). Nonalcoholic Steatohepatitis. Annals of Internal Medicine 126c;137–46.
4. Goel A, Gupta M, Aggarwal R. (2014). Gut microbiota and liver disease. Journal of Gastroenterology and Hepatology 1–30.
5. Koteish A, Diehl AM. (2002). Animal models of steatohepatitis. Best Practice and Research Clinical Gastroenteroly 16(5):679–90.
6. Arsov T, Silva DG, O’Bryan MK, Sainsbury A, Lee NJ, Kennedy C, et al. (2006). Fat aussie--a new Alström syndrome mouse showing a critical role for ALMS1 in obesity, diabetes, and spermatogenesis. Molecular Endocrinology 20(7):1610–22.
7. Arsov T, Larter CZ, Nolan CJ, Petrovsky N, Goodnow CC, Teoh NC, et al. (2006). Adaptive failure to high-fat diet characterizes steatohepatitis in Alms1 mutant mice. Biochemical and Biophysical research communications 342(4):1152–9.
8. Teoh NC, Williams J, Hartley J, Yu J, Mccuskey RS, Farrell GC. (2010). Short-term therapy with peroxisome proliferation-activator receptor-α agonist Wy-14,643 protects murine fatty liver against ischemia–reperfusion injury. Hepatology 51(3):996–1006.
9. Kelishadi R, Farajian S, Mirlohi M. (2013). Probiotics as a novel treatment for non-alcoholic Fatty liver disease; a systematic review on the current evidences. Hepatitis Monthly 13(4):e7233.
10. Ma Y-Y, Li L, Yu C-H, Shen Z, Chen L-H, Li Y-M. (2013). Effects of probiotics on nonalcoholic fatty liver disease: a meta-analysis. World Journal of Gastroenterology 19(40):6911–8.
11. Desreumaux P, Neut C, Cazaubiel M, Matthieu P, Fanny P. (2010). Saccharomyces Cerevisiae CNCM 1-3856 Reduces Digestive Discomfort and Abdominal Pain in Subjects With irritable Bowel Syndrome: A Randomized Double-Blinded Placebo-Controlled Clinical Trial. Gastroenterology 138.
12. Generoso S V, Viana M, Santos R, Martins FS, Cardoso VN. (2010). Saccharomyces cerevisiae strain UFMG 905 protects against bacterial translocation, preserves gut barrier integrity and stimulates the immune system in a murine intestinal obstruction model. Clinical Gastroenterology 477–84.
13. Zanello G, Berri M, Dupont J, Sizaret P-Y, D’Inca R, Salmon H, et al. (2011). Saccharomyces cerevisiae modulates immune gene expressions and inhibits ETEC-mediated ERK1/2 and p38 signaling pathways in intestinal epithelial cells. PLoS One 6(4):e18573.
14. Blanquet-Diot S, Denis S, Chalancon S, Chaira F, Cardot J-M, Alric M. (2012). Use of artificial digestive systems to investigate the biopharmaceutical factors influencing the survival of probiotic yeast during gastrointestinal transit in humans. Pharmaceutical Research 29(6):1444–53.
15. Cavalieri D, Calura E, Romualdi C, Marchi E, Radonjic M, Ommen B Van, et al. (2009). Filling gaps in PPAR-alpha signaling through comparative nutrigenomics analysis. BMC Genomics 16:1–16.

38
16. De Vogel-van den Bosch HM, Bünger M, de Groot PJ, Bosch-Vermeulen H, Hooiveld GJEJ, Müller M. (2008). PPARalpha-mediated effects of dietary lipids on intestinal barrier gene expression. BMC Genomics 9:231.
17. Ban S, Kasuga J, Nakagome I, Nobusada H, Takayama F. (2011). Bioorganic & Medicinal Chemistry ( NASH) -preventive effect of phenylpropanoic acid peroxisome proliferator-activated receptor ( PPAR ) a -selective agonists. Bioorganic and Medicinal Chemistry 19(10):3183–91.
18. Cuzzocrea S, Di Paola R, Mazzon E, Genovese T, Muià C, Centorrino T, et al. (2004). Role of endogenous and exogenous ligands for the peroxisome proliferators activated receptors alpha (PPAR-alpha) in the development of inflammatory bowel disease in mice. Laboratory Investigation 84(12): 1643–54.
19. Larter CZ, Yeh MM, Rooyen DM Van, Brooling J, Ghatora K, Farrell GC. (2012). Wy 14 643 , improves metabolic indices, steatosis and ballooning in diabetic mice with non-alcoholic steatohepatitis. Journal of Gastroenterology and Hepatology 27:341–50.
20. Nedvídková J, Smitka K, Kopský V, Hainer V. (2005). Adiponectin, an adipocyte-derived protein. Physiological Research 54(2):133–40.
21. Delerive P, Gervois P, Fruchart JC, Staels B. (2000). Induction of IκBα expression as a mechanism contributing to the anti-inflammatory activities of peroxisome proliferator-activated receptor-alpha activators. The Journal of Biological Chemistry 275(47):36703–7.
22. Mandard S, Müller M, Kersten S. (2004). Peroxisome proliferator-activated receptor alpha target genes. Cellular and Molecular Life Sciences 61(4):393–416.
23. Poekes L, Legry V, Farrell GC, Leclercq IA (2014). Role of ciliary dysfunction in a new model of obesity and non-alcoholic steatohepatitis: the foz/foz mice. Abstract.
24. Zhou M, Pu C, Xia L, Yu X, Zhu B, Cheng R, et al. (2014). Salecan diet increases short chain fatty acids and enriches beneficial microbiota in the mouse cecum. Carbohydrate Polymers 102:772–9.
25. Stahl A, Hirsch DJ, Gimeno RE, Punreddy S, Ge P, Watson N, et al. (1999). Identification of the major intestinal fatty acid transport protein. Molecular Cell 4(3):299–308.
26. Chen X, Matthews J, Zhou L, Pelton P, Liang Y, Xu J, et al. (2008). Improvement of dyslipidemia, insulin sensitivity, and energy balance by a peroxisome proliferator-activated receptor alpha agonist. Metabolism 57(11):1516–25.
27. Haluzik M, Gavrilova O, LeRoith D. (2004). Peroxisome proliferator-activated receptor-alpha deficiency does not alter insulin sensitivity in mice maintained on regular or high-fat diet: hyperinsulinemic-euglycemic clamp studies. Endocrinology 145(4):1662–7.
28. Black RN, Ennis CN, Young IS, Hunter SJ, Atkinson B, Bell PM. (2014) The peroxisome proliferator-activated receptor alpha agonist fenofibrate has no effect on insulin sensitivity compared to atorvastatin in type 2 diabetes mellitus; a randomised, double-blind controlled trial. Journal of Diabetes Complications 28(3):323–7.
29. Rousseaux C, Bouguen G, Dubuquoy C, Dubuquoy L, Vandekerckove P, Desreumaux P. Saccharomyces cerevisiae CNCM I-3856 decreases intestinal pain through PPAR alpha activation in the gut. S51 274R.
30. Caldwell S, Ikura Y, Dias D, Isomoto K, Yabu A, Moskaluk C, et al. (2010). Hepatocellular ballooning in NASH. Journal of Hepatology 53(4):719–23.


















