Characterization of Met95 mutants of a heme-regulated phosphodiesterase from Escherichia coli :...
9
Characterization of Met95 mutants of a heme-regulated phosphodiesterase from Escherichia coli Optical absorption, magnetic circular dichroism, circular dichroism, and redox potentials Satoshi Hirata, Toshitaka Matsui, Yukie Sasakura, Shunpei Sugiyama, Tokiko Yoshimura, Ikuko Sagami and Toru Shimizu Institute of Multidisciplinary Research for Advanced Materials, Tohoku University, Sendai, Japan On the basis of amino acid sequences and crystal structures of similar enzymes, it is proposed that Met95 of the heme- regulated phosphodiesterase from Escherichia coli (Ec DOS) acts as a heme axial ligand. In accordance with this proposal, the Soret and visible optical absorption and magnetic circular dichroism spectra of the Fe(II) complexes of the Met95Ala and Met95Leu mutant proteins indicate that these complexes are five-coordinated high-spin, suggesting that Met95 is an axial ligand for the Fe(II) complex. How- ever, the Fe(III) complexes of these mutants are six-coordi- nated low-spin, like the wild-type enzyme. The latter spectral findings are inconsistent with the proposal that the axial ligand to the Fe(III) heme is Met95. To determine the possibility of a redox-dependent ligand switch in Ec DOS, we further analyzed Soret CD spectra and redox potentials, which provide direct evidence on the environmental structure of the heme protein. CD spectra of Fe(III) Met95 mutants were all different from those of the wild-type protein, suggesting indirect coordination of Met95 to the Fe(III) wild-type heme. The redox potentials of the Met95Leu, Met95Ala and Met95His mutants were consid- erably lower than that of the wild-type enzyme (+70 mV) at )1, )26, and )122 mV vs. SHE, respectively. Thus, it is reasonable to speculate that water (or hydroxy anion) interacting with Met95, rather than Met95 itself, is the axial ligand to the Fe(III) heme. Keywords: heme sensor; magnetic circular dichroism; optical absorption; phosphodiesterase; redox potential. Heme-regulated phosphodiesterase from Escherichia coli (Ec DOS) has been cloned, and its structure–function relationships have been partially characterized by both our group and the Kitagawa group [1,2]. Interestingly, phosphodiesterase (PDE) is active only when the heme iron is in the Fe(II) redox state. Consistent with this finding, the PDE activity of Ec DOS was dramatically inhibited by CO and NO, which display strong affinities for the Fe(II) complex. Therefore, Ec DOS is likely to be a heme-based sensor, in which the redox state of the heme iron appears to control protein conformational changes, which in turn transmit signals to other domains to initiate the PDE catalytic function. The heme-bound N-terminal portion of Ec DOS has been identified as a PAS domain, based on the sequence and tertiary structure. PAS is an acronym formed from the names of proteins in which imperfect repeat sequences were initially recognized specifically: Drosophila period clock protein (PER) [3], vertebrate aryl hydrocarbon receptor nuclear translocator (ARNT) [4], and Drosophila single- minded protein (SIM) [5]. Site-directed mutagenesis studies reveal that one of the two axial ligation sites of the heme iron is occupied by His77 [1,2]. The Gilles-Gonzalez group reported on the physicochemical properties of the isolated N-terminal heme-bound PAS domain of Ec DOS and suggested that the heme axial ligand trans to His77 is Met95 on the basis of sequence homology data and the crystal structures of other heme-bound PAS proteins [6,7]. How- ever, resonance Raman spectra of Met95Ala and Met95His mutants of Ec DOS suggest that a different molecule occupies this axial position of the Fe(III) complex [2]. Therefore, it is of interest to determine the characteristics of Met95 mutants of Ec DOS using other spectroscopic techniques and kinetic analyses. Optical absorption [8–10], magnetic circular dichroism (MCD) [10–12], and CD [13,14] spectroscopy are useful tools for examining the heme coordination structure, protein folding and the environment of aromatic amino acid residues. Here, we present an initial report on the optical absorption, MCD and CD spectra of Met95 mutants of both full-length Ec DOS and the isolated PAS Correspondence to T. Shimizu, Institute of Multidisciplinary Research for Advanced Materials, Tohoku University, 2-1-1 Katahira, Aoba-ku, Sendai 980-8577, Japan. Fax: + 81 22 217 5604, 5605, 5390, Tel.: + 81 22 217 5604, 5605, E-mail: [email protected] Abbreviations: Ec DOS, heme-regulated phosphodiesterase obtained from Escherichia coli; PDE, phosphodiesterase; PAS, acronym formed from the names Drosophila period clock protein (PER), vertebrate aryl hydrocarbon receptor nuclear translocator (ARNT), and Drosophila single-minded protein (SIM); MCD, magnetic circular dichroism; FixL, oxygen sensor heme protein from Rhizobium meliloti. (Received 19 June 2003, revised 12 October 2003, accepted 16 October 2003) Eur. J. Biochem. 270, 4771–4779 (2003) ȑ FEBS 2003 doi:10.1046/j.1432-1033.2003.03879.x
-
Upload
satoshi-hirata -
Category
Documents
-
view
212 -
download
0
Transcript of Characterization of Met95 mutants of a heme-regulated phosphodiesterase from Escherichia coli :...

Characterization of Met95 mutants of a heme-regulatedphosphodiesterase from Escherichia coliOptical absorption, magnetic circular dichroism, circular dichroism, and redoxpotentials
Satoshi Hirata, Toshitaka Matsui, Yukie Sasakura, Shunpei Sugiyama, Tokiko Yoshimura,Ikuko Sagami and Toru Shimizu
Institute of Multidisciplinary Research for Advanced Materials, Tohoku University, Sendai, Japan
On the basis of amino acid sequences and crystal structuresof similar enzymes, it is proposed that Met95 of the heme-regulatedphosphodiesterase fromEscherichia coli (Ec DOS)acts as a heme axial ligand. In accordancewith this proposal,the Soret and visible optical absorption and magneticcircular dichroism spectra of the Fe(II) complexes of theMet95Ala and Met95Leu mutant proteins indicate thatthese complexes are five-coordinated high-spin, suggestingthat Met95 is an axial ligand for the Fe(II) complex. How-ever, the Fe(III) complexes of these mutants are six-coordi-nated low-spin, like thewild-type enzyme. The latter spectralfindings are inconsistent with the proposal that the axialligand to the Fe(III) heme is Met95. To determine thepossibility of a redox-dependent ligand switch in Ec DOS,we further analyzed Soret CD spectra and redox potentials,
which provide direct evidence on the environmentalstructure of the heme protein. CD spectra of Fe(III) Met95mutants were all different from those of the wild-typeprotein, suggesting indirect coordination of Met95 to theFe(III) wild-type heme. The redox potentials of theMet95Leu, Met95Ala and Met95His mutants were consid-erably lower than that of the wild-type enzyme (+70 mV)at )1, )26, and )122 mV vs. SHE, respectively. Thus, it isreasonable to speculate that water (or hydroxy anion)interacting with Met95, rather thanMet95 itself, is the axialligand to the Fe(III) heme.
Keywords: heme sensor;magnetic circular dichroism; opticalabsorption; phosphodiesterase; redox potential.
Heme-regulated phosphodiesterase from Escherichia coli(Ec DOS) has been cloned, and its structure–functionrelationships have been partially characterized by bothour group and the Kitagawa group [1,2]. Interestingly,phosphodiesterase (PDE) is active only when the heme ironis in the Fe(II) redox state. Consistent with this finding, thePDE activity of Ec DOS was dramatically inhibited byCO and NO, which display strong affinities for the Fe(II)complex. Therefore, Ec DOS is likely to be a heme-basedsensor, in which the redox state of the heme iron appears tocontrol protein conformational changes, which in turntransmit signals to other domains to initiate the PDEcatalytic function.
The heme-bound N-terminal portion of Ec DOS hasbeen identified as a PAS domain, based on the sequence andtertiary structure. PAS is an acronym formed from thenames of proteins in which imperfect repeat sequences wereinitially recognized specifically: Drosophila period clockprotein (PER) [3], vertebrate aryl hydrocarbon receptornuclear translocator (ARNT) [4], and Drosophila single-minded protein (SIM) [5]. Site-directed mutagenesis studiesreveal that one of the two axial ligation sites of the hemeiron is occupied by His77 [1,2]. The Gilles-Gonzalez groupreported on the physicochemical properties of the isolatedN-terminal heme-bound PAS domain of Ec DOS andsuggested that the heme axial ligand trans to His77 isMet95on the basis of sequence homology data and the crystalstructures of other heme-bound PAS proteins [6,7]. How-ever, resonance Raman spectra of Met95Ala andMet95Hismutants of Ec DOS suggest that a different moleculeoccupies this axial position of the Fe(III) complex [2].Therefore, it is of interest to determine the characteristics ofMet95 mutants of Ec DOS using other spectroscopictechniques and kinetic analyses.
Optical absorption [8–10], magnetic circular dichroism(MCD) [10–12], and CD [13,14] spectroscopy are usefultools for examining the heme coordination structure,protein folding and the environment of aromatic aminoacid residues. Here, we present an initial report on theoptical absorption, MCD and CD spectra of Met95mutants of both full-length Ec DOS and the isolated PAS
Correspondence to T. Shimizu, Institute of Multidisciplinary Research
for Advanced Materials, Tohoku University, 2-1-1 Katahira,
Aoba-ku, Sendai 980-8577, Japan.
Fax: + 81 22 217 5604, 5605, 5390, Tel.: + 81 22 217 5604, 5605,
E-mail: [email protected]
Abbreviations: Ec DOS, heme-regulated phosphodiesterase obtained
fromEscherichia coli; PDE, phosphodiesterase; PAS, acronym formed
from the namesDrosophila period clock protein (PER), vertebrate aryl
hydrocarbon receptor nuclear translocator (ARNT), and Drosophila
single-minded protein (SIM); MCD, magnetic circular dichroism;
FixL, oxygen sensor heme protein from Rhizobium meliloti.
(Received 19 June 2003, revised 12 October 2003,
accepted 16 October 2003)
Eur. J. Biochem. 270, 4771–4779 (2003) � FEBS 2003 doi:10.1046/j.1432-1033.2003.03879.x

domain. Mutations at Met95 did not alter the spin statefrom six-coordinated low spin to five-coordinated high spinin the Fe(III) complex in Soret and visible absorption andMCD spectra, but caused significant changes in the SoretCD peak position, and dramatically decreased the redoxpotentials. We discuss the heme coordination structure ofthis redox-sensitive heme sensor, Ec DOS, in view of ourspectral findings.
Experimental procedures
Materials and proteins
The fluorescent substrate, 2¢-o-anthraniloyl adenosine 3¢,5¢-cyclic monophosphate (ant-cAMP), was purchased fromCalbiochem (La Jolla, CA, USA). Calf intestine alkalinephosphatase was purchased from Takara Syuzo Co (Otsu,Japan). DEAE-Sephadex and Sephadex G25 were obtainedfrom Amersham Biosciences (Uppsala, Sweden). Otherchemicals were acquired from Wako Pure Chemicals(Osaka, Japan).
Cloning, mutations, expression in E. coli and purificationof full-length Ec DOS (amino acids 1–807) and the isolatedheme-bound PAS domain (amino acids 1–133) wereperformed as described previously [1,2]. Site-directed muta-genesis was performed using the PCR-based strategy withthe ODA-LA PCR kit from Takara Shuzo. The sequencewas confirmed by Sanger’s method using a DSQ-2000 Lautomatic sequencer (ShimadzuCo.).Ec DOS-PASmutantproteins were expressed in BL21 cells. Purified full-length Ec DOS and isolated PAS domain were morethan 95% homogeneous, as confirmed by SDS/PAGE.Molar absorption coefficients of the wild-type Fe(III) andFe(II) complexes were 129 cm)1ÆmM
)1 (at 416 nm) and175 cm)1ÆmM
)1 (at 427 nm), respectively, as determined bythe pyridine–hemochromogen method [18,19]. The coeffi-cients of the Met95Ala (at 414 nm), Met95Leu (at 414 nm)and Met95His (at 415 nm) mutant Fe(III) complexes were124, 124 and 126 cm)1ÆmM
)1, respectively.
Optical absorption, MCD, CD and fluorescence spectra
Optical absorption experiments were performed on Shim-adzu UV-1650, UV-2500 and Hitachi U-2010 spectropho-tometers maintained at 25 �C by a temperature controller.MCD spectra were obtained on a Jasco J-500 spectro-polarimeter equippedwith an electromagnetwhichproducesa longitudinal magnetic field in the sample (up to 1.53 T).CD spectra were obtained with Jasco J-720 and JascoJ-500 CD spectropolarimeters. Fluorescence spectra wereobtained using a Shimadzu RF-5300PC spectrofluorimeter.To ensure the appropriate temperature of the solution, thereaction mixture was equilibrated for 10 min before eachspectroscopic measurement.
Redox potentials
Anaerobic spectral experiments were performed on aShimadzu UV-160A spectrophotometer in a glove box.Redox potentials were obtained on the same spectro-meter in the glove box. 2,3,5,6-Tetramethylphenylenedi-amine (10 lM), N-ethylphenazonium ethosulfate (10 lM)
and 2-hydroxy-1,4-naphthoquinone (10 lM) were added asmediators to the wild-type protein solution before titration[15]. For Met95Ala and Met95Leu mutants, the dyeconcentrations were changed to 5 lM. For the Met95Hismutant, 5 lM anthraquinone-2,6-disulfonate was added tothe solution. The heme protein concentration used was15 lM. Spectral changes in intensity at 563 nm for wild-type, 560 nm for Met95Ala, 557 nm for Met95Leu and563 nm forMet95His accompanying the redox change weremonitored, as dye absorption hampers the detection ofSoret spectral changes. To ensure that the appropriatetemperature of the solution was maintained, the reactionmixture was incubated for 10 min before spectroscopicmeasurements. For reduction titration, the sample wasinitially fully oxidized with potassium ferricyanide. Aftereach addition of sodium dithionite and allowing equilibriumand stabilization, spectra were recorded. Titration experi-ments were repeated at least three times for each complex.
Enzymatic assays
PDE activities were measured using a fluorescence bandof the product, as described previously [1,19,20]. Briefly,Ec DOS was incubated at 37 �C with ant-cAMP in a500 lL reactionmixture comprising 50 mMTris/HCl buffer(pH 8.5), 2 mM MgCl2 and 1 mM dithiothreitol. Toterminate the reaction, Ec DOS was removed using anUltrafree-MC centrifugal filter (Millipore Co., Bedford,MA, USA). Then 2 U alkaline phosphatase was added tothe mixture and incubated for 1 h at 37 �C. Next, thereaction mixture was applied to a DEAE-Sephadex columnand washed with water. Finally, the fluorescence intensity(excitation at 330–350 nm, emission at 410 nm) of theeluted fraction was measured to determine the amount ofproduct [1]. At least four experiments were conducted toobtain each value. Experimental errors were less than 20%.
The enzyme was reduced with 10 mM sodium dithionite.Excess sodium dithionite was removed using a gel-filtrationcolumn (Sephadex G25) in a glove box under nitrogenatmosphere with an O2 concentration of less than 50 p.p.m.[18,19]. The reaction of the Ec DOS Fe(II) complex wasperformed under anaerobic conditions in the glove box.
Results
Optical absorption spectra
Optical absorption spectra (350–650 nm) of the Fe(III),Fe(II), and Fe(II)–CO complexes of wild-type and Met95mutants of the isolated PAS domain are shown in Fig. 1(lower), and summarized in Table 1. The absorptionspectrum (peaks at 416, 530 and 564 nm) of the Fe(III)complex of wild-type Ec DOS is that of a typical six-coordinated low-spin complex. This spectrum closelyresembles that of cytochrome b562 (peaks at 418, 530 and564 nm), but shows less similarity to the spectra of other six-coordinated low-spin heme proteins, including cytochromec and cytochrome b5 [20–24] (Table 1). To identify the axialligand trans to His77, we generated Met95Ala, Met95Leuand Met95His mutants. Soret and visible absorption peaksof the Fe(III) Met95 mutant proteins disclosed six-coordi-nated low-spin complexes essentially similar to that of the
4772 S. Hirata et al. (Eur. J. Biochem. 270) � FEBS 2003

wild-type enzyme (Fig. 1, Table 1). Notably, denaturationof the Met95 mutants occurred occasionally at hightemperatures during expression or as a consequence ofauto-oxidation of the O2-bound Fe(II) complexes duringpurification, often resulting in five-coordinated Fe(III) high-spin complexes.
The close similarities between the absorption spectra ofwild-type Ec DOS and cytochrome b562 are also evident inthe Fe(II) complexes (Table 1). Absorption spectra of Fe(II)complexes of the Met95Ala and Met95Leu mutants werecharacteristic of five-coordinated high-spin complexes sim-ilar to Fe(II) myoglobin, whereas the spectrum of the Fe(II)complex of the Met95His mutant was characteristic of asix-coordinated low-spin complex, analogous to the Fe(II)complex of the wild-type enzyme. Absorption spectra ofFe(II)–CO complexes of the Met95 mutants were compar-able to that of the wild-type enzyme and very similar to thespectrum of the corresponding Fe(II)–CO myoglobincomplex (Table 1). Spectra generated using mutants offull-length Ec DOS were essentially identical with those ofthe isolated PAS domain.
To identify axial ligand(s), the effect ofmodulating pH onabsorption bands was examined using the isolated PAS
domain, as holoenzymes are easily precipitated when pH isvaried. Between pH 3.5 and 9, no spectral changes wereobserved for either the wild-type or Met95His mutant PASdomain. However, Met95Ala and Met95Leu mutants formsix-coordinated high-spin complexes between pH 4 and 7.5.The Met95Leu mutant is likely to have a pKa at� 4.6 (datanot shown), but partial denaturation occurs concomitantlywith this spin-state shift in the acidic region. The Met95Alamutant was easily denatured when pH was varied below 6,and no clear pKa value was obtained.
MCD spectra
Figure 1 (upper) displays the MCD spectra of Fe(III),Fe(II) and Fe(II)–CO complexes of wild-type (A),Met95Ala (B), Met95Leu (C) and Met95His (D) mutantsof Ec DOS.MCD positions and intensities are summarizedin Table 2. The MCD spectrum of the Fe(III) complex ofthe wild-type enzyme contained a peak at 412 nm and atrough at 426 nm in the Soret region, and a small peak at554 nm and a small trough at 569 nm in the visibleregion (solid line in Fig. 1A). TheMCD spectral contour ofthe wild-type enzyme Fe(III) complex is typical of a
Fig. 1. Optical absorption (lower panel) Soret
and visible MCD (upper panel) spectra for the
Fe(III) (—), Fe(II) (- - -), and Fe(II)-CO (…)
complexes of wild-type (A), Met95Ala (B),
Met95Leu (C) and Met95His (D) mutants of
isolated heme-bound PAS domain. MCD
spectra of the wild-type andMet95 mutants of
the full-length enzyme were similar to those of
the isolated heme-bound PAS domain. Spec-
tra were obtained in 0.1 M Tris/HCl (pH 8.0)
buffer. Molar absorption coefficients deter-
mined are described in Experimental pro-
cedures.
� FEBS 2003 Met95 mutants of heme-regulated phosphodiesterase (Eur. J. Biochem. 270) 4773

six-coordinated low-spin complex. The intensity (De/T)of the Soret trough band � 80 M
)1Æcm)1ÆT)1 is also typicalof a low-spin complex [11]. Interestingly, the Met95Ala,Met95Leu and Met95His mutants of Ec DOS displayedsimilar MCD spectra (solid lines in Fig. 1B–D) to the wild-type enzyme, although a specific decrease in Soret MCDintensity was observed for the Met95Ala and Met95Leumutants (Table 2). Note that denaturation occurring duringthe purification procedures often generated five-coordinatedhigh-spin Fe(III) complexes for the Met95 mutants, inclu-ding Met95Phe (not shown).
The Fe(II) complex of thewild-type enzyme had a peak at� 430 nm in the Soret region at a comparable intensity tothe Fe(III) complex (Fig. 1A, broken line). MCD intensitiesrecorded in the visible region for the wild-type enzyme aremore than twofold higher than those in the Soret region,unlike those of the Fe(III) complex. A similarMCD patternwas observed for theMet95Hismutant (Fig. 1D).However,MCDband intensities in the visible spectra of theMet95AlaandMet95Leumutant proteins weremuch lower than thosein the Soret region. The position of theMCDpeak (437 nm)in the Soret region of Met95Ala, Met95Leu and Met95Hismutants was higher than that (430 nm) of the wild-typeprotein. Moreover, the Soret MCD spectra of the wild-typeenzyme and the Met95His mutant started on the plus sideand switched to the minus side (from lower to higherwavelengths), which was reversed for Met95Ala andMet95Leu mutants. The MCD data indicate that the Fe(II)complexes of the wild-type and Met95His mutant arecharacteristic of spectra of the low-spin complexes in thatintensities of the visible MCD bands are much higher thanthose of the Soret MCD bands [10,11]. On the other hand,
the Met95Ala and Met95Leu mutant complexes are high-spin, as the MCD intensities on the plus side in the Soretregion are much higher than those in the visible region[10,11].
The SoretMCDpeaks of the Fe(II)–CO complexes of thewild-type and Met95 mutants were located at � 420 or421 nm, with troughs at � 430 or 431 nm (Fig. 1 andTable 2, dotted lines). The MCD peaks and troughs in thevisible region of these proteins were observed at � 562–564 nm and 579–580 nm, respectively. However, the MCDspectra of the Fe(II)–CO complexes of the wild-type andMet95His mutant proteins differed from those of theMet95Ala and Met95Leu mutants. Specifically, the SoretMCD intensities of the Fe(II)–CO complexes of theMet95Ala and Met95Leu mutants were 20–30% higherthan those of the Fe(II)–CO complexes of the wild-typeenzyme and Met95His mutant (Table 2).
Soret CD spectra
The a-helix and b-sheet contents of the full-length enzymeand isolated PAS domain were determined from the UVregion of the CD spectrum in a previous study by our group[1]. Neither reduction of the heme iron norMet95mutationsaltered the CD spectrum in this region (data not shown).Soret CDbands of the proteins under various conditions areshown in Fig. 2 and summarized in Table 3. The Soret CDband of the Fe(III) wild-type complex was presented at� 421 nm on the plus side with De ¼ 26.2 M)1Æcm)1 (solidline in Fig. 2A). Reduction with sodium dithionite led to ashift in the band position to 431 nm and a slight increase inCD intensity (up to De ¼ 32.3 M
)1Æcm)1: broken line in
Table 1. Optical absorption maxima (nm) of the wild-type andMet95 mutants of isolated Ec DOSPAS domain. Putative coordination structures are
specified in parentheses. Optical absorption wavelengths of full-length enzymes were similar to those of the isolated PAS domain. SwMb, Sperm
whale myoglobin.
Proteins Fe(III) Fe(II) Fe(II)–CO Reference
Wild-type 416, 530, 564
(6c-LS)
427, 532, 563
(6c-LS)
423, 540, 570
(6c-LS)
This work
Met95Ala 414, 534, 563
(6c-LS)
432, 560
(5c-HS)
423, 541, 571
(6c-LS)
This work
Met95Leu 414, 533, 561
(6c-LS)
434, 558
(5c-HS)
423, 541, 570
(6c-LS)
This work
Met95His 415, 533, 564
(6c-LS)
427, 532, 561
(6c-LS)
423, 541, 570
(6c-LS)
This work
His, Met: axial ligands (6c-LS)
Cyt c 408, 530, 695 413, 521, 550 [20]
Cyt c2 416, 550 [21]
Cyt c551 410, 520, 551 416, 520, 551 [22]
Cyt b562 418, 530, 564 427, 531, 562 [23]
His, His: axial ligands (6c-LS)
Cyt c7 408 418, 522, 552 [24]
Cyt b5 413, 540 (br) 423, 526, 556 [23]
His: axial ligand
SwMb
(H2O) 410, 505, 635
(6c-HS)
434, 556
(5c-HS)
423, 542, 579
(6c-LS)
[8]
(OH–) 414, 542, 582
(6c-LS)
[8]
4774 S. Hirata et al. (Eur. J. Biochem. 270) � FEBS 2003

Fig. 2A). The Fe(II)–CO complex displayed a Soret bandat � 425 nm on the plus side with high intensity(De ¼ 58.5 M
)1Æcm)1: dotted line in Fig. 2A). Soret CDbands of the Fe(III) complexes of all the Met95 mutantswere observed at � 414 nm, in contrast with that ofthe wild-type enzyme complex, which was at � 421 nm.The distinct Soret CD positions of the Fe(III) complexes ofthe wild-type enzyme and the Met95 mutants is important,in view of the fact that the optical absorption, MCD, andresonance Raman spectra of the wild-type and mutantproteins are essentially similar [1,2]. The Soret CD bands ofthe Fe(II)Met95Ala andMet95Leumutants were presentedat� 435–437 nm, at a slightly higher position than those ofthe wild-type enzyme and Met95His mutant (430–431 nm).
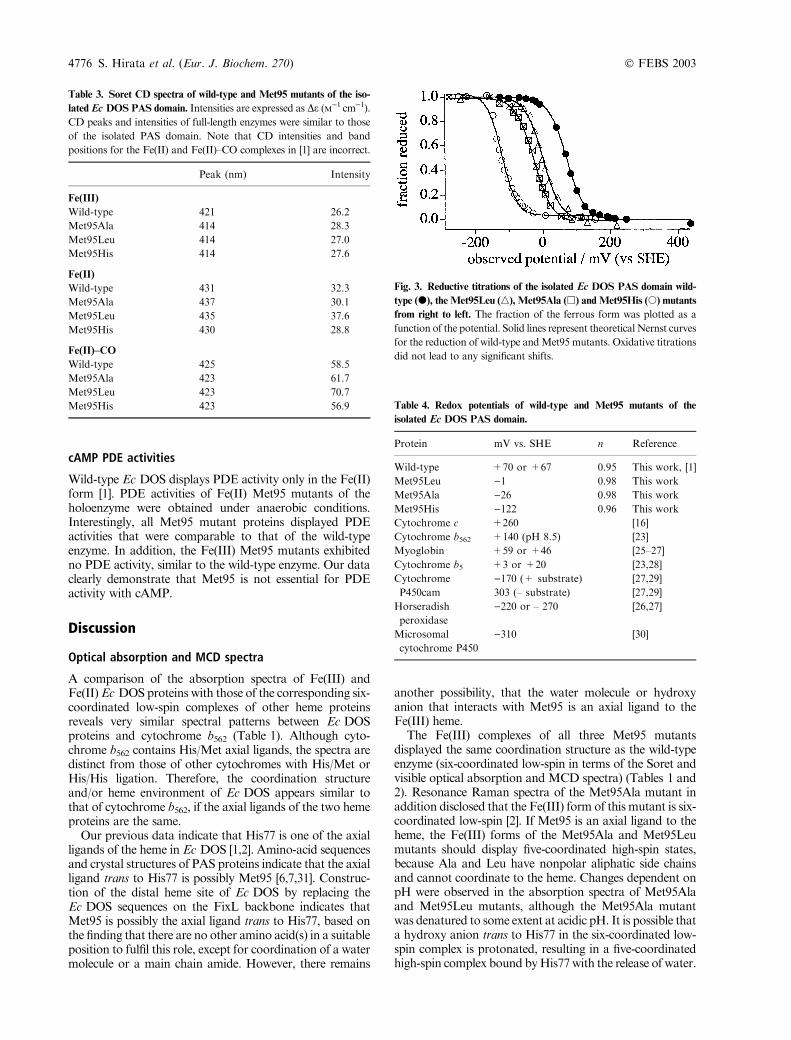
Redox potentials
Electrochemical titrations were conducted for the Met95mutants of the isolated PAS domain, as full-length enzymesprecipitate in the presence of mediators. The one-electronmidpoint potential of the Ec DOS heme was determinedfrom the absorbance change at �560 nm (Fig. 3). Redoxpotentials obtained for the Met95 mutants ()1 to )122 mVvs. SHE) summarized in Table 4 are much lower than thatof the wild-type enzyme (+70 mV). Oxidative titrationswere also conducted for the Met95 mutant proteins todetermine whether a redox-dependent ligand switch occurs.No significant differences were observed between the mid-point potentials of reductive and oxidative titrations.Interestingly, wild-type Ec DOS has a positive potential,similar to heme proteins involved in electron transfer or O2
storage, while Met95 mutants displayed lower, negativepotentials similar to the enzymes involved in O2 or H2O2
activation, such as cytochromes P450 and peroxidases(Table 4).
Table 2. MCD spectra of wild-type and Met95 mutants of isolated
Ec DOS PAS domain. Intensities (indicated in parentheses) are
expressed as De/H (M)1Æcm)1ÆT)1). MCD peaks and intensities of full-
length enzymes were similar to those of the isolated PAS domain.
Soret Visible
Peak Crossover Trough Peak Crossover Trough
Fe(III)
Wild-type 412
(78.9)
420 426
()61.4)554
(12.3)
563 569
()9.8)Met95Ala 410
(68.1)
418 425
()55.2)556
(8.7)
567 580
()7.5)Met95Leu 410
(65.0)
418 425
()48.9)556
(8.9)
571 580
()10.2)Met95His 410
(71.5)
417 425
()67.7)5.54
(9.9)
563 571
()8.3)
Fe(II)
Wild-type 430
(70.9)
558
(176)
561 565
()179)Met95Ala 437
(94.5)
Met95Leu 437
(109)
Met95His 436
(36.1)
555
(95.9)
559 563
()97.7)
Fe(II)–CO
Wild-type 420
(68.9)
427 431
()26.9)564
(18.4)
571 580
()25.1)Met95Ala 421
(88.6)
427 431
()31.7)563
(21.8)
571 579
()24.9)Met95Leu 420
(92.1)
426 430
()36.2)563
(25.0)
570 579
()28.1)Met95His 420
(73.6)
427 431
()27.4)562
(19.2)
570 579
()22.6)
Fig. 2. Soret CD spectra of the Fe(III) (—),
Fe(II) (- - -), and Fe(II)-CO (…) complexes of
wild-type (A), Met95Ala (B), Met95Leu (C)
and Met95His (D) mutants of isolated heme-
bound PAS domain. CD spectra of wild-type
and Met95 mutants of the full-length enzyme
were similar to those of the isolated heme-bound
PAS domain. Concentrations are 7.6 lM for
wild-type, 5.8 lM for the Met95Ala mutant,
4.5 lM for the Met95Leu mutant, and 5.6 lMfor the Met95His mutant in 0.1 M Tris/HCl
(pH 8.0) buffer.
� FEBS 2003 Met95 mutants of heme-regulated phosphodiesterase (Eur. J. Biochem. 270) 4775
cAMP PDE activities
Wild-type Ec DOS displays PDE activity only in the Fe(II)form [1]. PDE activities of Fe(II) Met95 mutants of theholoenzyme were obtained under anaerobic conditions.Interestingly, all Met95 mutant proteins displayed PDEactivities that were comparable to that of the wild-typeenzyme. In addition, the Fe(III) Met95 mutants exhibitedno PDE activity, similar to the wild-type enzyme. Our dataclearly demonstrate that Met95 is not essential for PDEactivity with cAMP.
Discussion
Optical absorption and MCD spectra
A comparison of the absorption spectra of Fe(III) andFe(II)Ec DOS proteins with those of the corresponding six-coordinated low-spin complexes of other heme proteinsreveals very similar spectral patterns between Ec DOSproteins and cytochrome b562 (Table 1). Although cyto-chrome b562 contains His/Met axial ligands, the spectra aredistinct from those of other cytochromes with His/Met orHis/His ligation. Therefore, the coordination structureand/or heme environment of Ec DOS appears similar tothat of cytochrome b562, if the axial ligands of the two hemeproteins are the same.
Our previous data indicate that His77 is one of the axialligands of the heme in Ec DOS [1,2]. Amino-acid sequencesand crystal structures of PAS proteins indicate that the axialligand trans to His77 is possibly Met95 [6,7,31]. Construc-tion of the distal heme site of Ec DOS by replacing theEc DOS sequences on the FixL backbone indicates thatMet95 is possibly the axial ligand trans to His77, based onthe finding that there are no other amino acid(s) in a suitableposition to fulfil this role, except for coordination of a watermolecule or a main chain amide. However, there remains
another possibility, that the water molecule or hydroxyanion that interacts with Met95 is an axial ligand to theFe(III) heme.
The Fe(III) complexes of all three Met95 mutantsdisplayed the same coordination structure as the wild-typeenzyme (six-coordinated low-spin in terms of the Soret andvisible optical absorption and MCD spectra) (Tables 1 and2). Resonance Raman spectra of the Met95Ala mutant inaddition disclosed that the Fe(III) form of this mutant is six-coordinated low-spin [2]. If Met95 is an axial ligand to theheme, the Fe(III) forms of the Met95Ala and Met95Leumutants should display five-coordinated high-spin states,because Ala and Leu have nonpolar aliphatic side chainsand cannot coordinate to the heme. Changes dependent onpH were observed in the absorption spectra of Met95Alaand Met95Leu mutants, although the Met95Ala mutantwas denatured to some extent at acidic pH. It is possible thata hydroxy anion trans to His77 in the six-coordinated low-spin complex is protonated, resulting in a five-coordinatedhigh-spin complex bound byHis77with the release of water.
Table 3. Soret CD spectra of wild-type and Met95 mutants of the iso-
lated Ec DOS PAS domain. Intensities are expressed as De (M)1Æcm)1).
CD peaks and intensities of full-length enzymes were similar to those
of the isolated PAS domain. Note that CD intensities and band
positions for the Fe(II) and Fe(II)–CO complexes in [1] are incorrect.
Peak (nm) Intensity
Fe(III)
Wild-type 421 26.2
Met95Ala 414 28.3
Met95Leu 414 27.0
Met95His 414 27.6
Fe(II)
Wild-type 431 32.3
Met95Ala 437 30.1
Met95Leu 435 37.6
Met95His 430 28.8
Fe(II)–CO
Wild-type 425 58.5
Met95Ala 423 61.7
Met95Leu 423 70.7
Met95His 423 56.9
Fig. 3. Reductive titrations of the isolated Ec DOS PAS domain wild-
type (d), the Met95Leu (n), Met95Ala (h) andMet95His (s) mutants
from right to left. The fraction of the ferrous form was plotted as a
function of the potential. Solid lines represent theoretical Nernst curves
for the reduction of wild-type andMet95 mutants. Oxidative titrations
did not lead to any significant shifts.
Table 4. Redox potentials of wild-type and Met95 mutants of the
isolated Ec DOS PAS domain.
Protein mV vs. SHE n Reference
Wild-type +70 or +67 0.95 This work, [1]
Met95Leu )1 0.98 This work
Met95Ala )26 0.98 This work
Met95His )122 0.96 This work
Cytochrome c +260 [16]
Cytochrome b562 +140 (pH 8.5) [23]
Myoglobin +59 or +46 [25–27]
Cytochrome b5 +3 or +20 [23,28]
Cytochrome
P450cam
)170 (+ substrate) [27,29]
303 (– substrate) [27,29]
Horseradish
peroxidase
)220 or – 270 [26,27]
Microsomal
cytochrome P450
)310 [30]
4776 S. Hirata et al. (Eur. J. Biochem. 270) � FEBS 2003

However, the apparent pKa of 4.6 is unusually low for atransition between an hydroxy anion and water. Thissuggests significant structural differences in the hemeenvironments of the wild-type enzyme and the Met95Alaand Met95Leu mutants. It is possible that an unknownligand, such as water/hydroxylate anion (or an amino-acidmain chain), coordinates to the Fe(III) in Met95Ala andMet95Leu mutants to produce low-spin complexes. Thistype of coordination may occur because of close proximityof the water/hydroxyl to Met95 or flexibility of theenvironment on the heme distal side.
For the Fe(II) complexes, optical absorption spectra ofthe Met95 mutants are consistent with predictions fromthe amino-acid sequences and crystal structures of thePAS proteins. Specifically, the Met95Ala and Met95Leumutants form five-coordinated high-spin complexes,whereas the complex of the Met95His mutant is six-coordinated low-spin, similar to that of the wild-typeenzyme, indicating that His is the sixth axial ligand in theFe(II) Met95His complex.
Optical absorption spectra of the Fe(II)–CO complexesof the wild-type and Met95 mutants are similar, indicatingthe presence a common internal axial ligand, His77, asexpected from the resonance Raman spectra of the proteins[2].
The spin and electronic states determined from theMCDspectra of the Met95 mutants analyzed in this study areconsistent with those obtained from the optical absorptionspectra. The MCD spectra of the Met95 mutants indicatethat substitution with Ala, Leu and His did not alter thespin state of the Fe(III) complex, whereas Met95Ala andMet95Leu mutations changed the spin state of the Fe(II)complex from low spin to high spin. However, the SoretMCD intensities of the Fe(II)–CO complexes of theMet95Ala and Met95Leu mutants were higher than thoseof the wild-type enzyme and Met95His mutant. It appearsthat ligand displacement accompanies theMet95mutations,and Met95 may be the sixth axial ligand in the Fe(II)complex. Therefore, it suggests that redox-dependent axialligand exchange occurs, as indicated by resonance Ramanspectroscopy [2].
Soret CD spectra
The Soret CD band of the Fe(III) form of Ec DOS islocated exclusively on the plus side. This is distinct fromcytochrome b562 [32] and cytochrome c [33] complexes,which display bands on the minus side and both sides,respectively. The Soret CD band of Ec DOS shifted from421 nm to 425 nm and the intensity increased on reduction,suggesting a change in the heme environment. The origin ofthe Soret CD band of the heme protein may be due tointeractions of the p to p* transition of nearby aromatic sidechains with the delocalized p-electron system of the hemeprosthetic group and/or the unique conformation of thepolypeptide surrounding the heme [13,33–35]. In fact, Phemutations markedly altered the Soret CD band shape ofcytochrome c [33]. Importantly, the Soret CD positions ofall the Fe(III) Met95 mutants are 7 nm lower in wavelengththan that of the wild-type enzyme. The Soret CD spectra ofthe Fe(III)Met95mutants thus indicate that substitutions atthis position result in similar and profound conformation
changes near the heme prosthetic group in this particularoxidation state. Therefore, it is reasonable to speculate thatthe Met95 mutations affect the 6th ligand (water/hydroxyion) of the Fe(III) complex at the heme distal side.
From the Soret CD bands of the Fe(II) complexes, itappears that the heme environment structures of the Fe(II)wild-type enzyme and the Met95His mutant are distinctfrom those of the Fe(II) Met95Ala andMet95Leu mutants.These observations appear consistent with the opticalabsorption, MCD and resonance Raman spectra, whichsuggest that the Fe(II) wild-type and Met95His mutant arein the low-spin state, whereas the Fe(II) Met95Ala andMet95Leu mutants are high-spin [1,2]. The Soret CD bandpositions and intensities of the Fe(II)–CO complexes of thewild-type and Met95 mutants are similar.
Redox potentials
If a redox-dependent ligand switch occurs, the value for theoxidative titration would be expected to be different fromthat for the reductive titration. However, no such differencewas observed in the two directions. However, if a redox-dependent ligand switch occurs and one of the axial ligandsin either redox state is a water molecule or hydroxy anion, itis possible that the values for the two directions are similar,because the ligand switching does not need high energy.
The redox potential of the wild-type proteinwas +70 mV vs. SHE, which is within the range ofelectron-transfer hemoproteins, cytochromes, and myoglo-bin (Table 4). On mutation of Met95 to Ala, Leu and His,the redox potential was markedly decreased in all themutants studied (Table 4), suggesting that the mutationsstabilize the Fe(III) form rather than the Fe(II) form.Optical absorption and MCD spectral data are consistentwith the theory that Met coordination favours the Fe(II)form, whereas His coordination favors the Fe(III) form.Again, it is implied that Met95 is not an axial ligand in theFe(III) complex. The redox potentials of theMet95mutantsare negative, similar to cytochrome P450 enzymes andperoxidases. On the other hand, redox potentials ofcytochromes and myoglobin are positive (Table 4). There-fore, Met95 of Ec DOS appears to be an important redoxpotential control residue, preventing the enzyme fromexhibiting peroxidase-like and/or P450-like behavior andavoiding activation of molecular oxygen or H2O2 underaerobic conditions.
Catalytic activities
All Met95 mutants are active only when the heme iron is inthe Fe(II) state, similar to the wild-typeEc DOS enzyme [1].The Fe(II) complexes of Met95Ala and Met95Leu mutantsare five-coordinated high-spin, in contrast with the Fe(II)form of the wild-type enzyme, which is six-coordinated low-spin. From the spectra, it appears that the axial ligand transto His77 is Met95 in the Fe(II) form [2]. Therefore, thechanges in the Fe(II) heme coordination structure andelectronic state caused by the Met95 mutations did notgreatly influence the PDE activity of Ec DOS. Thesestructural changes are induced on the heme distal side,where CO and O2 are expected to bind. Ec DOS activity issensitive to the heme oxidation state. Accordingly, we
� FEBS 2003 Met95 mutants of heme-regulated phosphodiesterase (Eur. J. Biochem. 270) 4777

speculate that the heme proximal site structure (includingthe Fe-His N distance and/or conformation), which altersdepending on the oxidation state, is critical for regulatingthe PDE activity of Ec DOS. Detailed structural studies onthe environment near the heme are required to explain theredox-sensitive PDE activity of this enzyme.
Crystal structure
After we submitted this manuscript, crystal structures of theisolated heme-bound PAS domain of the wild-typeEc DOSprotein (resolution 1.3 A,R ¼ 0.16) were determined in thislaboratory (H. Kurokawa, unpublished results). The struc-tures indicate that the axial ligands of the Fe(III) complexare His77 and water, whereas those of the Fe(II) complexare His77 and Met95. The redox-dependent ligand switchsuggested from the solution and crystallographic studies ofthe enzyme agree well with each other, and this factincreases the value of both approaches. Thus, artifactsresulting from amino-acid substitution, experimental con-ditions, data misinterpretation or the crystallization processcan be ruled out in the 3D structure determination work.
Acknowledgements
We thank Dr Hirofumi Kurokawa for valuable suggestions.
References
1. Sasakura, Y., Hirata, S., Sugiyama, S., Suzuki, S., Taguchi, S.,
Watanabe, M., Matsui, T., Sagami, I. & Shimizu, T. (2002)
Characterization of a direct oxygen sensor heme protein from
Escherichia coli. Effects of the heme redox states and mutations at
the heme-binding site on catalysis and structure. J. Biol. Chem.
277, 23821–23827.
2. Sato, A., Sasakura, Y., Sugiyama, S., Sagami, I., Shimizu, T.,
Mizutani, Y. & Kitagawa, T. (2002) Stationary and time-resolved
resonance Raman spectra of His77 and Met95 mutants of the
isolated heme domain of a direct oxygen sensor from Escherichia
coli. J. Biol. Chem. 277, 32650–32658.
3. Jackson, F.R., Bargiello, T.A., Yun, S.H. & Young, M.W. (1986)
Product of per locus of Drosophila shares homology with pro-
teoglycans. Nature (London) 320, 185–188.
4. Hoffman, E.C., Reyes, H., Chu, F.F., Sander, F., Conley, L.H.,
Brooks, B.A. &Hankinson, O. (1991) Cloning of a factor required
for activity of the Ah (dioxin) receptor. Science 252, 954–958.
5. Nambu, J.R., Lewis, J.O., Wharton, K.A.J. & Crews, S.T. (1991)
The Drosophila single-minded gene encodes a helix-loop-helix
protein that acts as a master regulator of CNS midline develop-
ment. Cell 67, 1157–1167.
6. Tomita, T., Gonzalez, G., Chang, A.L., Ikeda-Saito, M. & Gilles-
Gonzalez,M.-A. (2002) A comparative resonance Raman analysis
of heme-binding PAS domains: heme iron coordination structures
of the BjFixL, AxPDEA1, EcDos, and MtDos proteins.
Biochemistry 41, 4819–4826.
7. Gonzalez, G., Dioum, E.M., Bertolucci, C.M., Tomita, T., Ikeda-
Saito, M., Cheesman, M.R., Watmough, N.J. & Gillez-Gonzalez,
M.-A. (2002) Nature of the displaceable heme-axial residue in the
Ec Dos protein, a heme-based sensor from Escherichia coli.
Biochemistry 41, 8414–8421.
8. Antonini, E. & Brunori, M. (1971) Hemoglobin and Myoglobin in
their Reactions with Ligands. North-Holland, Amsterdam.
9. Eichhorn, G. & Marzilli, L.G., eds. (1988) Advances in Inorganic
Chemistry, Vol. 7 Hemoproteins. Elsevier, New York.
10. Sono, M., Roach, M.P., Coulter, E.D. & Dawson, J.H. (1996)
Heme-containing oxygenases. Chem. Rev. 96, 2841–2887.
11. Dawson, J.H. & Dooly, D.M. (1989) Magnetic circular dichroism
spectroscopy of iron porphyrins and heme proteins. In Iron Por-
phyrins, Part III (Lever, A.B.P. & Gray, H.B., eds), pp. 1–135.
VCH Publishers, NY.
12. Abraham, B.D., Sono, M., Boutaud, O., Shriner, A., Dawson,
J.H., Brash, A.R. & Gaffney, B.J. (2001) Characterization of the
coral allene oxide synthase active site with UV-visible absorption,
magnetic circular dichroism, and electron paramagnetic resonance
spectroscopy: evidence for tyrosinate ligation to the ferric enzyme
heme iron. Biochemistry 40, 2251–2259.
13. Myer, Y. & Pande, A. (1978) Circular dichroism studies of
hemoproteins and heme models. In The Porphyrins, III (Dolphin,
D., ed.), pp. 271–322. Academic Press, NY.
14. Fasman, G.D., ed. (1996) Circular Dichroism and the Conforma-
tional Analysis of Biomolecules. Plenum Press, New York.
15. Dutton, P.L. (1978) Redox potentiometry: determination of
midpoint potentials of oxidation-reduction components of bio-
logical electron-transfer systems.Methods Enzymol. 54, 411–435.
16. Hiratsuka, T. (1982) New fluorescent analogs of cAMP and
cGMP available as substrates for cyclic nucleotide phosphodiest-
erase. J. Biol. Chem. 257, 13354–13358.
17. Kincaid, R.L. & Manganeillo, V.C. (1998) Assay of cyclic
nucleotide phosphodiesterase using radiolabeled and fluorescent
substrates.Methods Enzymol. 159, 457–477.
18. Sagami, I., Daff, S. & Shimizu, T. (2001) Intra-subunit and inter-
subunit electron transfer in neuronal nitric-oxide synthase: effect
of calmodulin on heterodimer catalysis. J. Biol. Chem. 276, 30036–
30042.
19. Rozhkova, E.A., Fujimoto, N., Sagami, I., Daff, S.N. &
Shimizu, T. (2002) Interactions between the isolated oxygenase
and reductase domains of neuronal nitric-oxide synthase: assessing
the role of calmodulin. J. Biol. Chem. 277, 16888–16894.
20. Banci, L. & Assfalg, M. (2001) Mitochodrial cytochrome c. In:
Handbook of Metalloproteins (Messerschmidt, A., Huber, R.,
Poulos, T. &Wieghardt, K., eds), Vol. 1, pp. 33–43. JohnWiley &
Sons, Chicester.
21. Miki, K. & Sogabe, S. (2001) Cytochrome c2. In Handbook of
Metalloproteins (Messerschmidt, A., Huber, R., Poulos, T. &
Wieghardt, K., eds), Vol. 1, pp. 55–68. John Wiley & Sons, Chi-
cester.
22. Cutruzzola, F., Arese, M. & Brunori, M. (2001) Cytochrome c551.
In Handbook of Metalloproteins (Messerschmidt, A., Huber, R.,
Poulos, T. &Wieghardt, K., eds), Vol. 1, pp. 69–79. JohnWiley &
Sons, Chicester.
23. Mathews, F.S. (2001) b-Type cytochrome electron carriers: cyto-
chromes b562 and b5, and flavocytochrome b2. In Handbook of
Metalloproteins (Messerschmidt, A., Huber, R., Poulos, T. &
Wieghardt, K., eds), Vol. 1, pp. 159–171. John Wiley & Sons,
Chicester.
24. Banci, L. & Assfalg, M. (2001) Cytochrome c7. In Handbook of
Metalloproteins (Messerschmidt, A., Huber, R., Poulos, T. &
Wieghardt, K., eds), Vol. 1, pp. 110–118. John Wiley & Sons,
Chicester.
25. Van Dyke, B.R., Saltman, P. & Armstrong, F.A. (1996) Control
of myoglobin electron-transfer rates by the distal (nonbound)
histidine residues. J. Am. Chem. Soc. 118, 3490–3492.
26. Gajhede, M. (2001) Horseradish peroxidase. Handbook of Met-
alloproteins (Messerschmidt, A., Huber, R., Poulos, T. &
Wieghardt, K., eds), Vol. 1, pp. 195–210. John Wiley & Sons,
Chicester.
27. Erman, J.E., Hager, L.P. & Sligar, S.G. (1994) Cytochrome P-450
and peroxidase chemistry. Adv. Inorg. Chem. 10, 71–118.
28. Funk, W.D., Lo, T.P., Mauk, M.R., Brayer, G.D., MacGillivray,
R.T.A. & Mauk, A.G. (1990) Mutagenic, electrochemical, and
4778 S. Hirata et al. (Eur. J. Biochem. 270) � FEBS 2003

crystallographic investigation of the cytochrome b5 oxidation-
reduction equilibrium: involvement of asparagine-57, serine-64,
and heme propionate-7. Biochemistry 29, 5500–5508.
29. Huang, Y.-Y., Hara, T., Sligar, S., Coon, M.J. & Kimura, T.
(1986) Thermodynamic properties of oxidation-reduction reac-
tions of bacterial, microsomal, and mitochondrial cytochromes
P-450: an entropy-enthalpy compensation effect. Biochemistry 25,
1390–1394.
30. Guengerich, F.P. (1983) Oxidation-reduction properties of rat
liver cytochromes P-450 and NADPH-cytochrome p-450 reduc-
tase related to catalysis in reconstituted systems. Biochemistry 22,
2811–2820.
31. Miyatake, H., Mukai, M., Park, S.-Y., Adachi, S., Tamura, K.,
Nakamura, H., Nakamura, K., Tsuchiya, T., Iizuka, T. & Shiro,
Y. (2000) Sensory mechanism of oxygen sensor FixL from Rhi-
zobium meliloti: crystallographic, mutagenesis and resonance
Raman spectroscopic studies. J. Mol. Biol. 301, 415–431.
32. Myer, Y.P. & Bullock, P.A. (1978) Cytochrome b562 from
Escherichia coli: conformational, configurational, and spin-state
characterization. Biochemistry 17, 3723–3729.
33. Pielak, G.J., Oikawa, K., Mauk, A.G., Smith, M. & Kay,
C.M. (1986) Elimination of the negative Soret Cotton effect
of cytochrome c by replacement of the invariant phenylalanine
using site-directed mutagenesis. J. Am. Chem. Soc. 108, 2724–
2727.
34. Hsu,M.-C. &Woody, R.W. (1971) The origin of the heme Cotton
effects inmyoglobin and hemoglobin. J. Am. Chem. Soc. 93, 3515–
3525.
35. Kiefl, C., Sreerama,N., Haddad, R., Sun, L., Jentzen,W., Qiu, Y.,
Shelnut, J.A. & Woody, R.W. (2002) Heme distortions in sperm-
whale carbonmonoxy myoglobin: correlations between rotational
strengths and heme distortions in MD-generated structures.
J. Am. Chem. Soc. 124, 3385–3394.
� FEBS 2003 Met95 mutants of heme-regulated phosphodiesterase (Eur. J. Biochem. 270) 4779