SEMINARIO ESTUDIOS DE POSTGRADO Julio 2012 Ma. Isabel Munita J. Secretaria Ejecutiva Becas Chile.

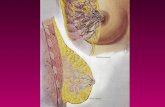
Anatomia e fisiologia dell’apparato mammario

• L’organo della lattazione è costituito dalle mammelle, che sono ghiandole tubulo-alveolari composte di natura ectodermica, poste nella regione pubica nei ruminanti e negli equini, mentre nei suini e nei carnivori sono dislocate dalla regione pubica fino a quella sternale. • Il loro numero è diverso nelle varie specie: due nella giumenta equina e asinina, nella pecora e nella capra; quattro nella vacca; otto nella gatta, da otto a dieci nella cagna e da dieci a dodici nella scrofa.


Funzione biologica della ghiandola mammaria
• Ghiandola esocrina; comune a tutti i mammiferi • Funzione: nutrire il neonato
– Fonte di nutrienti: grasso, proteine, zuccheri, vitamine, minerali, acqua
– Protezione: immunoglobuline

• Ogni ghiandola è munita di un capezzolo; nella vacca, nella pecora e nella capra questo ha un solo orifizio, in cui termina il canale del capezzolo, alla base del quale sta la cisterna del latte o seno galattoforo, che raccoglie il latte veicolato dai canali galattofori. • Nella cavalla, per ogni mammella, gli orifizi e i seni galattofori sono due; nella scrofa, che non ha le cisterne del latte, i canali galattofori sboccano direttamente all’estremità del capezzolo, che ha perciò un numero variabile di orifizi (da 5 a 10).

Struttura della ghiandola mammaria • La mammella è costituita da 2/4 ghiandole separate
– Ogni quarto termina con un capezzolo • Il fondo del capezzolo è chiuso da uno sfintere • Ci possono essere capezzoli sopranumerari, non funzionanti
• Conformazione del capezzolo – Di forma variabile, da conico a cilindrico – I capezzoli anteriori, di norma, sono più corti – Ogni capezzolo dovrebbe essere posto al centro del quarto – Gli sfinteri dei capezzoli dovrebbero essere sufficientemente
stretti da prevenire lo sgocciolamento del latte – I capezzoli sono privi di peli

















Pins
Thurl
Hooks
Short Ribs
Punta ischio (natica)
Punta ileo (anca)
Articolazione coxo-femorale
Vista laterale
Per la vacca da latte il BCS è basato sulla valutazione dell’area pelvica e di quella lombare

Vista da tergo
Pins
Thurl
Hooks
Short Ribs
Tailhead Ligament
Sacral Ligament


Struttura della ghiandola mammaria
• supporto = Stroma (tessuto connettivo)
• Ghiandola; tessuto secernente = Parenchima – Alveoli- cellule epiteliali secernenti
– Lobi e lobuli: clusters di alveoli supportati da tessuto connettivo
– Sistema dei dotti, cellule epiteliali allineate

Quarti anteriore e posteriore di ghiandola mammaria
60% 40%

Medial suspensory
ligament


Anatomia interna • Orifizio del capezzolo
– Contenere il latte all’interno della mammella e tenere i batteri al di fuori
• Cisterna del capezzolo – Dotto interno al capezzolo di capacità pari a 30-45
millilitri (nella vacca) – Separato dall’orifizio del capezzolo da uno strato di
tessuto chiamato rosetta di Furstenberg. • Cisterna del latte
Contiene circa 400 millilitri di latte • Raccoglie il latte proveniente dal sistema dei dotti mammari

Fürstenburg's Rosette - La rosetta di Fürstenburg è localizzata sopra l’orifizio dello sfintere - E’ composta da strati di tessuto che si appiattiscono via via che il latte si accumula nella mammella. - Questo aiuta a bloccare la fuoriuscita di latte fra due mungiture. - Questo strato di tessuto può essere danneggiato da mungiture mal eseguite o da trattamenti sanitari impropri..

Orifizio del capezzolo -chiuso dai muscoli dello sfintere. Ruoli: 1. Prevenire la fuoriuscita di latte tra le mungiture 2. Barriera nei confronti dei batteri e di corpi estranei - La velocità di mungitura è legata alla dimensione
dell’orifizio e alla forza con la quale l’orifizio è chiuso

Cheratina - Le cellule che formano l’orifizio contengono cheratina - La cheratina è una sostanza cerosa. - Questa sostanza aiuta a sigillare il capezzolo tra le mungiture - La cheratina, inoltre, ha proprietà inibenti la crescita batterica. - La formazione di cheratina rappresenta la prima linea di difesa della mammella..

La cisterna del capezzolo - E’ la cavità all’interno della mammella che contiene circa 30 mL di latte (la quantità può variare in funzione delle dimensione della mammella). - Il latte vi si accumula durante la mungitura. - Si riempie in continuo durante la mungitura - Vi si accumula anche il latte tra una mungitura e l’altra (primo latte da rimuovere all’inizio della mungitura)

Seno galattaforo o cisterna mammaria E’ collegato alla cisterna del capezzolo ed è collegato alla base della mammella. Varia in dimensioni ed è il punto di congiunzione che raccoglie il latte dei dotti galattofori. Anch’essa si riempie e si svuota rapidamente durante la mungitura.

Alveoli e sistema dei dotti • Gli alveoli sono l’unità secernente principale
– Sono costituiti da piccoli bulbi con una cavità centrale. – Le cellule epiteliali secernenti sono allineate a formare
l’alveolo • Ogni alveolo è circondato da una rete di capillari e di cellule
mioepiteliali – La contrazione delle cellule mioepiteliali consente l’eiezione
del latte • Gruppi di alveoli che sboccano in un unico dotto formano un
lobulo – Più lobuli formano un lobo – Più lobi sboccano in un dotto galattoforo che a sua volta
sbocca nella cisterna mammaria • I dotti consentono di conservare il latte e di veicolarlo verso
l’esterno.


Struttura dell’alveolo mammario
(Patton, 1969.)

Model alveolus (a) with subtending duct (d) showing blood supply, adipocyte stroma, myoepithelial cells, and plasma cells (PC).

Schema organizzazione ghiandola mammaria

Sezione trasversale di alveolo mammario
An alveolus is composed of a single layer of epithelial cells surrounding a lumen, and surrounded by a basket network of myoepithelial cells. This is surrounded by a basement membrane (made of connective tissue proteins). Blood vessels and capillaries run through the stroma (inter-alveolar spaces). The stroma also contains fibroblasts, leukocytes (plasma cells) and other connective tissue cells.

Circulation • La produzione del latte è sostenuta da un flusso di
sangue che varia da 5 a 9 litri al minuto. In generale sono necessari 500 volumi di sangue per generare un volume di latte.
• Il sangue entra nella mammella attraverso le arterie pudiche esterne
• Il sangue esce dalla mammella dalle vene alla base della mammella, attraverso due vie: – Vene pudiche eserne – Vene addominali subcutanee

Fig 29-6. Blood flow to and from the mammary gland determines milk producing capability of the cow (Courtesy of Iowa State University)

La lattogenesi
Formalmente la lattogenesi è definita come l’inizio della secrezione del latte in conseguenza dell’evento parto.
In realtà il primo stadio della lattazione ha inizio nell’ultima fase della gravidanza, poco prima del parto, quando aumenta il livello plasmatico degli estrogeni che stimolano la secrezione di prolattina, mentre la PGF2α determina la diminuzione del progesterone circolante. Questo fatto riveste un ruolo importante in quanto il progesterone inibisce la sintesi di α-lattoalbumina, una proteina essenziale per la formazione della lattosio sintetasi, enzima limitante per la biosintesi del lattosio.

Curva di lattazione
La produzione del latte nelle varie specie è caratterizzata da una durata differente e da un tipico andamento nel tempo, che vede succedersi più fasi
Durata lattazione (giorni)
Vacca 300-320 Cavalla 180-200 Bufala 280-300 Capra 240-250 Pecora 150-160
Picco di lattazione
Settimane dopo il parto
Fase colostrale
Fase di crescita
Fase di decrescita
Fase di asciutta

La fase colostrale
Il colostro è il primo prodotto di secrezione della ghiandola mammaria subito dopo il parto ed è essenziale per la sopravvivenza del vitello neonato. Il colostro, infatti, fornisce la vitello i nutrienti essenziali per stimolare il suo metabolismo e la sua attività digestiva. Il colostro è anche fonte di protezione immunitaria passiva. La quantità, la qualità e il tempo di somministrazione del colostro sono i fattori principali che determinano l’incidenza di mortalità e morbilità.

Il colostro, rispetto al latte materno, contiene più del doppio di sostanza secca, il triplo di sostanze minerali e cinque volte di più di proteine. Anche il contenuto totale di energia e vitamine è più elevato, in particolare di vitamina A e D, importanti per il neonato in quanto possiede basse riserve di queste sostanze. Il lattosio, al contrario, è contenuto in minori quantità, per evitare che l’iniziale ridotta capacità del vitello di produrre lattasi porti l’instaurarsi di disturbi intestinali.
Colostro
1 2 3 Latte
Sostanza secca (%) 23.9 17.9 14.9 12.9
Proteine (%) 14.0 8.4 5.1 3.1
IgG (mg/ml) 32.0 25.0 15.0 0.6
Grasso 6.7 5.4 3.9 4.0
Lattosio 2.7 3.9 4.4 5.0
Ceneri 1.1 1.0 0.8 0.7
Vitamina A (ug/dl) 295.0 190.0 113.0 34.0

Controllo della lattazione (1) Alla funzionalità della mammella concorrono molti
ormoni:
Ø Prolattina: secreta dalle cellule lattotrofe dell’adenoipofisi, sostiene la lattogenesi e la galattopoiesi che avvia e sostiene i processi di biosintesi e di secrezione del latte.

Controllo della lattazione (2)
Ø Ormone somatotropo: esercita un’azione di controllo sul numero e le dimensioni delle cellule mammarie secernenti durante la lattazione. Le bovine più produttive avrebbero una elevata increzione di GH e questa condizione costituirebbe una caratteristica genetica.
Ø Estrogeni: agiscono sullo sviluppo del tessuto secretivo mammario (accrescimento dei dotti).
Ø Ormoni surrenalici e t iroidei (il Cortisolo, T3: Trioidiotironina e T4 :Tiroxina), necessari al mantenimento della lattazione. I primi sono indispensabili per la formazione della quota minerale e idrica del latte in quanto regolano il ricambio cationico (Na, K, ecc.), mentre i secondi regolano la funzionalità mitocondriale e, quindi, le vie metaboliche energetiche cellulari.

Model of Lactogenesis Pregnancy Lactation
Epithelialcell Laminin-Rich
Basement Membrane
Estrogen, Progesterone
Direct +indirecteffects
TGFa
Growth
ECMSynthesis
Progesterone
InhibitsLactation
Epithelialcell
P4 receptor
Cortisol, Prolactin
Milk Protein,Other Genes
Milk
Laminin-RichBasement Membrane
LowP4 receptor
LowProgesterone


L’unità secernente della ghiandola mammaria è l’alveolo. Ogni alveolo è rivestito di tessuto connettivo a formare dei lobuli che, a loro volta, sono raccolti in lobi. In questo modo avremo un sistema di dotti interalveolari, intralobulari e intralobari o galattofori.. Solo gli alveoli sono rivestiti da cellule epiteliali secernenti, i dotti e le cisterne sono rivestiti di cellule epiteliali non secernenti.
Il sistema dei dotti confluisce in vasi di calibro sempre maggiore che sboccano in una cavità denominata cisterna del latte che comunica con la sottostante cisterna del capezzolo, di forma allungata. Il capezzolo è dotato di un dotto papillare chiuso allo sbocco esterno da uno sfintere.

L’eiezione del latte Il riflesso di eiezione induce la discesa del latte dall’alveolo e dai piccoli dotti delle cisterne per spremitura degli alveoli stessi da parte delle fibre mioepiteliali a canestro, contrattili, che lo avvolgono. La contrazione è dovuta all’ormone ossitocina.
L’ossitocina viene secreta dalla neuro-ipofisi in seguito a stimoli visivi, sonori e meccanici percepiti dall’animale. Il tempo che intercorre fra il rilascio dell’ossitocina e la spremitura degli alveoli è 45-60 ‘’. L’azione dell’ormone ossitocico è di breve durata (4-5 minuti), in tale tempo il latte deve essere estratto, altrimenti, in breve tempo, la pressione mammaria riacquista i valori iniziali.

La rimozione completa e continua del latte dalla ghiandola mammaria è essenziale per il mantenimento della lattazione. L’aumento dell’intervallo di mungitura, infatti, va a detrimento del tasso di secrezione. Per lungo tempo, l’aumento della pressione intramammaria è stata ritenuta la causa di questo fenomeno, più recentemente è stato osservato che altri fattori fisiologici sono da ritenersi più determinanti. Il fattore che sembra essere più importante è l’accumulo di una proteina, genericamente denominata FIL (Feedback Inhibitor of lactation), che, quando si accumula negli alveoli causa un meccanismo di inibizione a feedback della sintesi dei componenti del latte. Per questo motivo l’aumento della frequenza delle mungiture concorre al mantenimento dell’efficienza secretoria della ghiandola mammaria.

Eiezione del latte • Riflesso neuro-‐ormonale • La suzione o il massaggio della mammella s4molano i nervi
mammari che inviano un segnale all’ipofisi posteriore, causando il rilascio dell’ossitocina.
• L’ossitocina induce la contrazione delle cellule mioepiteliali a<orno agli alveoli.
• L’efficienza della mungitura dipende dai seguen4 fa<ori: – S4molazione della mammella almeno 1 minuto prima
dell’inizio della mungitura – Il massimo effe<o dell’ossitocina si realizza nei primi 2-‐3
minu4 di mungitura – Gli stress in pre-‐parto o alla mungitura inibiscono il rilascio
di ossitocina

302520151050-5-10-15-20-250
5
10
15
20
25
30
35Oxytocin Release at Milking
MINUTES
Seru
m O
xyto
cin
(µU
/ml)
Machine onPremilking Stimulus

ORMONI e
METABOLITI
• Clima • Stagione • Mungitura • Management
Latte • Quantità • Qualità
Mammella
Alimentazione Genetica
Tessuti adiposo e muscolare
• Stato salute • Stato benessere (stress)
L’armonizzazione degli effetti ormonali nella vacca da latte è da attribuire al patrimonio genetico, oltre che all’ottimale interazione con i fattori ambientali e, in particolare, la disponibilità di nutrienti per la sintesi del latte.

Acqua
(%)
Residuo secco (%)
Grasso (%)
Proteine (%)
Lattosio (%)
Ceneri (%)
Peso specifico
Energia kcal/kg
Donna 87,57 12,43 3,38 1,64 6,69 0,22 1,0300 682,2
Vacca 87,62 12,38 3,46 3,43 4,71 0,78 1,0321 712,6
Zebù 86,56 13,14 4,90 3,21 4,58 0,75 1,0306 827,7
Asina 90,39 9,61 1,21 1,74 6,23 0,43 1,0336 463,3
Cavalla 90,48 9,52 0,85 2,06 6,26 0,35 1,0356 448,6
Lama 86,55 13,45 3,15 3,90 5,60 0,80 1,0000 745,5
Renna 65,79 34,21 19,73 10,31 2,71 1,46 1,0000 2570,4
Bufala 83,59 16,41 7,16 3,77 4,73 0,75 1,0310 1084,3
Dromedaria 87,83 12,17 4,68 3,31 3,51 0,67 1,0000 773,7
Capra 86,77 13,23 4,62 3,41 4,47 0,73 1,0308 812,1
Pecora 80,48 19,52 7,54 6,17 4,89 0,92 1,0376 1263,6
Scrofa 81,91 18,09 6,16 6,31 4,61 1,01 1,0412 1129,3
Cagna 79,14 20,86 9,21 7,39 3,19 1,07 1,0353 1423,8
Gatta 91,63 18,37 3,33 9,08 4,91 0,58 1,0000 1030,3
Coniglia 69,50 30,50 10,45 15,54 1,95 2,56 1,0493 1956,5
Elefantessa 66,70 33,30 22,07 3,21 7,39 0,63 1,0000 2575,2
Delfina 48,76 51,24 43,71 - - 0,46 1,0000 -
Balena 76,98 33,02 13,42 10,43 1,48 1,15 1,0000 1928,6

Meccanismi di secrezione dei componenti del latte
La cellula mammaria elabora alcuni componenti del latte a partire da precursori ematici cui addiziona acqua e sali minerali per completare la composizione del latte
sangue
+
Cellula mammaria Alveolo mammario
Lattosio
Proteine
Grassi
Acqua
Sali minerali
Fasi fisiche del latte: Lattosio e Sali minerali: soluzione
Proteine: sospensione Grassi: emulsione
Precursori ematici
Latte

Substrate % uptake Utilization (all can be used for energy). Major direct contributions
to milk are in bold Glucose 25 Lactose synthesis
NADPH for fat synthesis Glycerol for triglyceride synthesis Synthesis of glucogenic amino acids Synthesis of fatty acids (non-‐ruminants only) Synthesis of ketogenic amino acids (non-‐ruminants only)
Acetate 56 Fatty acid synthesis Synthesis of ketogenic amino acids
b-‐OH-‐butyrate 40 Fatty acid synthesis Synthesis of ketogenic amino acids
Propionate 45 Glucose synthesis Synthesis of glucogenic amino acids
Triglycerides 58 Milk triglyceride Fatty acid portion -‐ synthesis of ketogenic amino acids Glycerol portion -‐ synthesis of glucose or glucogenic amino acids
Free fatty acids
5 Milk Triglycerides Synthesis of ketogenic amino acids
Amino acids 20-‐60 Most 40-‐50
Protein synthesis Glucogenic amino acids -‐ lactose synthesis Ketogenic amino acids -‐ fat synthesis

Vie anaboliche
MajorOrgan(s)
Input Output Energy Needs
Gluconeogenesis Liver,others Pyruvate, otheracids
Glucose NADH, ATP
Malonyl CoA Most,adipose,mammary
Acetyl CoA Fatty acids NADPH, ATP
ProteinSynthesis
All, Amino acids proteins ATP, GTP
LactoseSynthesis
Mammary Glucose Lactose UTP
Urea Cycle Liver,Others(Mammary)
Amonia Urea ATP

Energia
• Energia riducente – NADH – FADH2 – NADPH
• Energia libera – ATP – GTP, UTP, etc. – Creatine phosphate

Vie cataboliche
Pathway MajorOrgan(s)
Input Output Energy Output
Glycolysis All Glucose Pyruvate, NADH, ATP
Kreb’s Cycle All Acetyl CoA CO2 NADH,FADH2, GTP
PentosePhosphate
Most Glucose CO2, Various sugars NADPH
OxidativePhosphorylation
All O2,NADH,FADH2
H2O ATP
Beta oxidation All Fatty Acids Acetyl CoA ATP
Amino AcidMetabolism
MostLiver
AminoAcids
Amino acids,Carboxylic acids
Various

Conversione dei substrati Blood Secretory Epithelium Milk
Acetateβ-OH Butyrate
Glucogenic amino acids
Propionate
Ketogenic amino acids
Free Fatty Acids
Lipoproteins
Glucose
Triglycerides
Proteins
Lactose
MalonlyCoA Pathway
Fatty Acids
Glycerol
Glucose
LipoproteinLipase
Gluconeogenesis

Fabbisogni energetici Blood Secretory Epithelium Milk
Acetateβ-OH Butyrate
Glucogenic amino acids
Propionate
Ketogenic amino acids
Free Fatty Acids
Lipoproteins
Glucose
Triglycerides
Proteins
Lactose
MalonlyCoA Pathway
Fatty Acids
Glycerol
Glucose
Gluconeogenesis
ATP; NADH
UTP
ATP
ATP; NADPH
ATP, GTP

Fonti di energia
Acetateβ-OH Butyrate
Glucogenic amino acids
Propionate
Ketogenic amino acids
Free Fatty Acids
Lipoproteins
Glucose Glycolysis
Oxidative Phosphorylation
β-Oxidation
Pyruvate
Acetyl CoA
NADHATP
GTPUTP
Glycerol
ATP; NADH
FADH2NADH
KrebsCycle
GTP
OAA
Mitochondria

Ruolo degli organi Bone
GI Tract
Liver
Kidney
AdiposeMuscle
NutrientAbsorption
CirculationGluconeogenesisTransamination
Lipoprotein synthesis
Fat SynthesisStorage
Lipolysis
Ca StorageMobilization Transamination
Protein deposition
Ca Reabsorption
FatProteinLactoseSynthesis

Metabolismo lipidico
GI Tract
Liver
Adipose
AcetatePropionate(Glucose in Non-ruminants)
NEFA
Fat Synthesis
Triglyceride SynthesisLipoprotein assembly
ChylomicronsLiproproteins
Fatty acidSynthesisTriglyceride
Synthesis
MilkFatExport
Inefficient in cows
NEFA uptakeby Udderinefficient

La ghiandola mammaria produce i lipidi partendo da un pool di acidi grassi che per il 40% (in peso) viene sintetizzato “de novo” dalle cellule stesse, a partire dall’acetato e dal β-idrossibutirrato, che contribuisce per il 15%; il rimanente 60% dei lipidi, è costituito da acidi grassi prelevati dal flusso ematico.
I lipidi del latte

rumine
Principi nutritivi
Fermentazioni batteriche ruminali dei
carboidrati
AGV Acetato, Propionato
e Butirrato
1) tessuti
2) Attività digestive post ruminali
Origine dei lipidi nei ruminanti

Cellulosa Amido
Glucosio 1-P Glucosio Glucosio 6-P
Fruttosio 6-P
Glicolisi
Piruvato
Emicellulose Pentosani Pectine
Pentosi
Fruttosio
Via AcetilCoA Via Metil MalonilCoA Via AcetilFosfato
Acetato + metano Acetato e Butirrato Proprionato
Acetato e Butirrato (convertito a βidrossibutirrato a livello della parete ruminale) sono i precursori degli acidi grassi a corta e media catena

Lipidi della dieta dei ruminanti
Intestino
Rumine
Non sono usati come substrati fermentativi, ma gli acidi grassi insaturi vanno incontro ad una massiccia idrogenazione
Sono: Acidi Grassi Liberi (FFA) di origine alimentare o microbica, piccole quantità di fosfolipidi microbici. Nel caso della
somministrazione di grassi protetti si possono avere anche trigliceridi
Lipidi della dieta Acidi grassi Esterificazione tra
glicerina ed acidi grassi
Glucosio
Trigliceridi
Bile (emulsione) Fosfolipasi A2 Via glicerofosfato

Intestino
Lipidi della dieta dei ruminanti
Nelle cellule della mucosa intestinale
Glucosio Glicerina
Esterificazione con gli acidi grassi
Trigliceridi
Assemblaggio in Chilomicroni Tessuti e latte
Fosfolipidi Colesterolo Apoproteine

I chilomicroni vengono “smontati” a livello epatico e gli acidi grassi che ne derivano sono
utilizzati (ossidazione, desaturazione, allungamento, riesterificazione e successivo assemblaggio di lipoproteine a varia densità)
I chilomicroni e le lipoproteine possono raggiungere direttamente i tessuti e la
ghiandola mammaria dove sono utilizzati per la formazione del grasso, senza essere preliminarmente processati nel fegato

glucosio
glicerolo
Acetato
3-idrossibutirrato
Chilomicroni VLDL
LPL
Acil-CoA
TG MFGM
LATTE Ghiandola mammaria
Sangue
Glicerolo 3-fostato
Trasportatore glucosio
glicerolo
ACC FAS Desaturasi
FA Albumina-FA

Il grasso del latte dei ruminati
Cellula secernente
Globulo di grasso formato
Globulo di grasso in formazione
Membrana cellulare
Membrana del globulo
Dimensione dei Globuli di Grasso
Bovini 3.60-5.32 µm Ovini 3.20-4.95 µm
Caprini 3.10-4.89 µm

Neo-formazione mammaria
Dieta e/o mobilizzazione
riserve
SCFA
MCFA (fino a C:14 e parte del C16)
BCFA
UFA (solo i prodotti dell’enzima SCD)
SFA (solo a partire dal C16) TFA
LCFA
MUFA
PUFA

Punti critici del metabolismo lipidico mammario
Afflusso di precursori e acidi grassi preformati dal metabolismo ruminale e dalla dieta: AGV; TFA; LCFA; PUFA; LPL
Allungamento e desaturazione degli acidi grassi a livello mammario: FAS; SCD
Formazione dei trigliceridi: Sn1
Sn2
Sn3
Uguale affinità per LCFA, SFA, UFA.
Maggiore affinità per LCFA, e SCFA, competizione tra C4:0 e C18:0
GPAT (glicerol-3fosfato aciltransferasi)
LPAT (lisofosfatidic acid aciltransferasi)
DGAT (diacilglicerol aciltransferasi)

La posizione Sn2 riveste un ruolo piuttosto importante, in quanto, nell’uomo, le lipasi pancreatiche attaccano
soprattutto le posizioni Sn1 e Sn2, generando 2-monogliceridi assorbiti velocemente ed in via
preferenziale
Affinità per la posizione
Sn2
C16:0
C18:1
C18:3, C18:0, C20:4
C14:0, C12:0, C18:2
Questa caratteristica può essere modificata:
- Aumentando la disponibilità di PUFA a livello della mammella,
si può determinare una loro maggiore inclusione in posizione
Sn2. - Determinando i fattori endogeni di regolazione dell’espressione del gene per la LPAAT (enzima che catalizza l’esterificazione
degli acidi grassi alla posizione Sn2)
La recente localizzazione e caratterizzazione del gene bovino
LLAAT rappresenta un primo passo in questa direzione

Punti critici del metabolismo lipidico mammario
Afflusso di precursori e acidi grassi preformati dal metabolismo ruminale e dalla dieta: AGV; TFA; LCFA; PUFA; LPL
Allungamento e desaturazione degli acidi grassi a livello mammario: FAS; SCD
Formazione dei trigliceridi: Sn1
Sn2
Sn3
Uguale affinità per LCFA, SFA, UFA.
Maggiore affinità per LCFA, e SCFA, competizione tra C4:0 e C18:0
GPAT (glicerol-3fosfato aciltransferasi)
LPAT (lisofosfatidic acid aciltransferasi)
DGAT (diacilglicerol aciltransferasi)

I lipidi del latte si trovano sotto forma di globuli sferici, secreti dalle cellule epiteliali alveolari, tali globuli sono costituiti da un “core” di Trigliceridi, rivestito da una sottile membrana del globulo di grasso

Distribuzione dei lipidi nel latte bovino
Membrana del globulo
Core
0.5-1% lipidi totali del latte
95-98 % lipidi totali del latte
Siero 1.5-4 % lipidi totali
del latte
Costituiti da 21-44% fosfolipidi 56-79% trigliceridi
5.1-10.7% digliceridi 1-5.1% acidi grassi liberi
0.4-1.9% colesterolo
Costituiti da 88-93% trigliceridi 5.2-9.8% digliceridi
1.5-7.3% acidi grassi liberi 0.2-1.4% colesterolo
Costituiti da 30-45% fosfolipidi 71-83% trigliceridi
4.3-10.1% digliceridi 8.7-15.7% acidi grassi liberi
1.2-8.4% colesterolo

I trigliceridi del latte
La distribuzione degli acidi grassi nei trigliceridi non è casuale
• latte ovino: 134 specie di TG (Fontecha et al., 2004)
• latte di capra: 137 specie di TG (Fontecha et al., 2000)
• I più rappresentati nel latte ovino sono: 4:0 – 14:0 – 16:0 (3.5%); (C34) 4:0 – 16:0 – 16:0 (4.2%); (C36) 4:0 – 16:0 – 18:1 (4.5%); (C38) 10:0 – 16:0 – 16:0 (3.5%); (C42) 10:0 – 16:0 – 18:1 (3.5%); (C44) 16:0 – 16:0 – 18:1 (3.0%); (C50) 16:0 – 18:1 – 18:1 (3.0%); (C52) La distribuzione degli acidi grassi nei TG è asimmetrica: il C4:0 esterifica la glicerina in poszione Sn-3 e anche gli altri acidi grassi a corta catena evidenziano una netta preferenza per questo sito
CCH2O
O
CH2O
O
C
CCHO
O
R3FA
R1FA
R2FA
Sn3 Sn2 Sn1
50% trisaturo, 30% monoinsaturo, 20% polinsaturo 51% trisaturo, 31% monoinsaturo, 18% polinsaturo)

Time (min) 7 8 9 10 11
20000
40000
60000
80000
100000
counts
C54
C30 C32
C34
C36
C38 C40
C42 C44
C46 C48
C50
C52
C26 C28
C24
Profilo trigliceridi nel latte ovino
(Mele et al., 2011)

Acido Sn 1 Sn 2 Sn3 C4:0 - - 35.4 C6:0 - 0.9 12.9 C8:0 1.4 0.7 3.6 C10:0 1.9 3.0 6.2 C12:0 4.9 6.2 0.6 C14:0 9.7 17.5 6.4 C15:0 2.0 2.9 1.4 C16:0 34.0 32.3 5.4 C16:1 34.0 3.6 1.4 C17 2.8 1.0 0.1
C18:0 1.3 9.5 1.2 C18:1 10.3 18.9 23.1 C18:2 1.7 3.5 2.3
Distribuzione degli acidi grassi nei trigliceridi Gli acidi grassi
volatili mostrano una netta preferenza per la
posizione sn-3 e competono con il
C18:1 per la posizione Sn-3
La ghiandola mammaria può
modulare la sintesi degli acidi grassi a
corta catena e dell’acido oleico per mantenere costante la fluidità del grasso del
latte

Metabolismo proteico
Liver
Muscle
Amino acidsNon a.a Precursors
Transaminationa.a Synthesis
Protein synthesis
Transaminationa.a. synthesis
Protein Synthesis
Amino acid absorptionTransamination
GI Tract
ProteinSynthesis
MilkProteins

Le proteine del latte
Con il termine “proteina del latte” ci si riferisce in realtà a differenti gruppi di proteine che spesso presentano attributi e proprietà molto eterogenei tra loro.
Alfa-s1
Alfa-s2
Beta
K
Le caseine
Le proteine del siero Beta-lattoglobulina Alfa-lattoalbumina
diventano insolubili a pH 4,6 e, perciò, possono essere determinate tramite precipitazione per aggiustamento del pH
Ciò che rimane in soluzione dopo l’aggregazione delle caseine

La “caseina intera” è in realtà un grosso aggregato di molecole appartenenti a quattro tipi di frazioni
proteiche complesse ovvero tre fosfoproteine calcio-sensibili (αs1-, αs2-, β-caseina) e una
glicofosfoproteina detta k-caseina.
caseina intera 80% delle lattoproteine
αs1-caseina 36% delle caseine αs2-caseina 10% delle caseine β-caseina 34% delle caseine k-caseina 13% delle caseine
γ1-γ2-γ3 caseina 7% delle caseine

LE CASEINE
Le caseine riescono a restare in sospensione nel latte poichè sono organizzate all’interno di una struttura micellare.
L a k - c a s e i n a è u n a p r o t e i n a determinante per il processo di caseificazione.

CASEIN MICELLE
NO
N P
OLA
R PO
LAR
Phenilalanine (AA105) Methionine (AA106)

Chimosin action AA105 AA106
AA105 AA106

Milk coagulation process I – Enzymatic phase
II. Non-enzymatic phase Aggregation Ca++ induced
para K-casein
glyco-macropeptide

The aggregation of casein micelles begins after the 80-90% of K-casein
degradation
Aggregation of casein micelles

LE PROTEINE DEL SIERO • L’α-‐la%oalbumina e la β-‐la%oglobulina sono le principali componen3 della frazione sierosa del la%e.
– L’ α-‐la%oalbumina è un cos4tuente della la<osio-‐sintetasi, l’enzima responsabile della sintesi del la<osio, nell’ul4ma fase del processo dove il glucosio viene legato al gala<osio. Il gene è situato sul cromosoma 5 per una lunghezza di 2023 bp.
– La β-‐la%oglobulina è la principale proteina del siero nel la<e bovino ed il gene che la codifica è situato sul cromosoma 11 per una lunghezza di 4723 bp.

Varianti alleliche in
Bos taurus Altre
varianti Varianti più
frequenti ed in quali razze
CASA1 199aa
A B C D F G H E (yak) Ebali (Banteng)
B (0,75-0,95) C
B in tutte le razze. C predominante sulla alla B in zebu e yak.
CASA2 207aa
A B D B (zebu) C (yak)
A B (B. Indicus 0,34)
A in tutte le razze.
CASB 209aa
A1 A’ A2 B2 A3
H(A4) B F C A5
E G A3mongolia
D (zebu) A4 (Banteng)
A1, A2 B C
A1, A2 quasi tutte le razze. B (Normande e Jersey, 0,3-0,45; altre razze, 0,1-0,25).
CASK 169aa
A H B I C A(1) B2 J E F G
“A dello zebu”
F (yak) G (yak)
A, B C
A prevale in Frisona, Rossa Danese, Ayrshire. B prevale in Jersey e Normande. C si trova in Bruna Italiana
POLIMORFISMO DELLE CASEINE
I 4pi di caseina che influenzano maggiormente le proprietà tecnologiche del la<e sono la k-‐caseina e la β-‐caseina. A queste due proteine sono infaT associa4 notevoli effeT sulle dimensioni delle micelle, sulle cara<eris4che di coagulazione presamica del la<e e sulla stabilità finale del coagulo.

POLIMORFISMO DELLE PROTEINE DEL SIERO DEL LATTE
• Il fenomeno del polimorfismo è stato scoperto nel 1955 analizzando la β-lattoglobulina.
Varianti alleliche in Bos taurus
Altre varianti
Varianti più frequenti ed in quali razze
LGB 162aa
A I B J(X) D Dr W H C
E (yak e Banteng) F (Banteng) G (Banteng)
B A D
B (Shorthorn (0,89); Guersney (0,78); Frisona (0,60); Ayrshire (0,69)) A (in Frisona si trova con frequenza di 0,40)
LALBA 123aa
A B C (Banteng) B A
B (tutte le razze eccetto B. javanicus). A (Comune in B.indicus ma non in B.taurus).

Le caseine sono organizzate in una struttura micellare, in cui la K-caseina è localizzata nella parte più esterna e grazie alla sua idrofilicità ha il ruolo di colloide che protegge e favorisce le micelle in sospensione Le proteine del siero sono assai sensibili al calore a causa dell’elevato contenuto in amminoacidi solforati, mentre, al contrario delle caseine, non precipitano per azione enzimatica. Le immunoglobuline sono la base dell’immunità passiva del colostro.

Le caseine, la lattoalbumina e la β-lattoglobulina sono sintetizzate nel reticolo endoplasmatico rugoso delle cellule alveolari a partire dagli amminoacidi veicolati dal sangue; da qui sono trasferite nell’apparato di Golgi da cui, insieme con lattosio ed elementi minerali, sono secrete nel lume alveolare sotto forma di vescicole. La sieroalbumina e le immunoglobuline, invece, provengono direttamente dal sangue. Le proteine del latte hanno un buon valore biologico; sono da sottolineare in esse gli elevati contenuti di lisina, triptofano, metionina e treonina, che rappresentano gli amminoacidi carenti nelle proteine dei vegetali.

Cellula della ghiandola mammaria e modalità di sintesi e secrezione del latte

Metabolismo del glucosio
Gluconeogenesis
Protein breakdown
Glucogenic amino acids
GI Tract
PropionateGlucose
LactoseSynthesis
MilkLactose
LiverGlycogen
Stores
Fat Synthesis
NADPH forFat SynthesisGlycerol
Fatty Acids
Triglycerides
Glucose is a substratefor fat synthesis inmonogastrics,NOT ruminants.
FromPentosePhosphatePathway
Liver
Muscle

Il lattosio, disaccaride costituito da glucosio e galattosio, è una molecola osmoticamente attiva, che, non essendo in grado di permeare la membrana delle vescicole del Golgi, all’interno delle quali viene secreta nel lume alveolare, richiama all’interno di esse l’acqua necessaria a garantire l’osmolarità. In ultima analisi, pertanto, la quantità di latte prodotta è funzione della quantità di lattosio sintetizzato dalle cellule mammarie.
Lattosio

La sintesi del lattosio è garantita dall’enzima lattosio sintetasi che è formata da due proteine: la UDP-galattosiltransferasi e la alfa-lattoalbumina. In assenza di quest’ultima la sintesi del lattosio non può avvenire. La disponibilità di alfa-lattoalbumina è sotto il controllo della prolattina. Il progesterone esercita sulla lattazione un’azione inibente dovuta all’influenza negativa che ha l’ormone nella formazione della alfa-lattoalbumina.
La disponibilità di glucosio per la ghiandola mammaria è un altro fattore limitante la sintesi del lattosio
Al lattosio è stato riconosciuto un ruolo positivo nel metabolismo del calcio in quanto interviene nell’assorbimento di questo elemento minerale a livello intestinale.

Biosintesi del lattosio
Glucose Glucose-6 PO4 Glucose-1-PO4
UDP-glucose
Glucokinase Isomerase
Uridinediphosphorylglucosephosphorylase
UDP-galactose
UDP-galactose-4-epimerase
Lactose
ATP ADP UTP
2 PO4
Glucose
Lactose SynthaseGalactosyl transferase + α-lactalbumin
Golgi,SecretoryVessicle
UDP
UDP-galactose
Glucose TransportFacilitated, not active
ATPADP
Active Transport ofUDP-galactose

Fonti di glucosio • sangue – la più importante
– Assorbimento intestinale – Gluconeogenesi nel fegato (circa 1/2 glucosio ematico)
• Acido propionico • Aminoacidi glucogenici
– Dalla dieta – Mobilizzazione proteine muscolari
– Mobilizzazione glicogeno del fegato- poco attuata nella specie bovina
• Gluconeogenesi mammaria: minoritaria • Acetato, butirrato, acidi grassi e aminoacidi chetogenici
non foramano glucosio

Regolazione della sintesi del lattosio
Adipose
fat synthesis glucose need
and uptake
Liver
gluconeogenesis
GI Tract
Capacity Intake
VFA, glucose, aa lipid uptake
Glucose availability
PropionateAmino acids
Muscle
Glucose Uptakeand utilization
NADPHProduction(about 1/3) Blood
Flow
LipoproteinsChylomicrons
lipoproteinlipase
NEFAs
Glucose need for fatty acid synthesis
Lactose Synthesis(1/2 - 2/3)
Mammary Gland
α-lactalbumin UDP-galactosetransport

Importanza del lattosio • Controllo del volume di latte (sostanza
osmoticametne attiva) • Fonte energetica per la prole • Il metabolismo lipidico richiede carboidrati
– Mancanza di carboidrati può indurre chetosi – Gli aminoacidi sono convertiti a glucosio
• Evita il consumo di aminoacidi per la gluconeogenesi
• L’uso energetico di proteine è meno efficiente rispetto ai carboidrati

Rappresenta lo 0,7-1% del totale, a seconda della specie, Costituenti
ü Potassio (K): 0.16% ü Calcio (Ca): 0.11 ü Fosforo (P): 0.11%.
Il Ca è presente come fosfato organico ed è più facilmente
disponibile per l’assorbimento del Ca di alcuni alimenti di origine vegetale, che lo contengono come ossalato di Ca o come Fitato di Ca. Infatti, sia i lattanti sia l’uomo adulto non hanno nel loro corredo enzimatico le fitasi necessarie a scindere il legame del Ca fitinico.
Il P è presente in un rapporto ottimale con il Ca (Ca/P>1), mentre questo non avviene negli altri alimenti di largo consumo di origine sia vegetale, sia animale.
La componente minerale del latte

Contenuto di ioni nel sangue, nelle cellule e nel latte
sangue cellula latte
Na (mM) 145 10 25
K (mM) 6 100 35
Cl (mM) 99 10 28

Ioni e acqua
K+
Na+
Cl-
Cl-
Na+
K+
Cl-
Cl-
K+
Na+
K+
Na+
Tight JunctionBlocks ion movement
“leaky” tight junctionIons leak across
K+, lactose Na+, Cl-
Cl-
K+
Na+
K+
Na+
Cl-
Glucose
Glucose
Lactose
H2O
H2O
Tissue Fluid
High NaHigh ClLow K
Milk
High KLow NaLow Cl
Intracellular Fluid
High KLow NaLow Cl

Metabolismo del calcio
Increased CaAbsorption
Decreased Ca Reabsorption------------------Increased CaReabsorption
Calcitonin
Parathormone
IncreasedDeposition---------------IncreasedResorption
Thyroid
Parathyroid
GI Tract
Bones Kidney
Vitamin D

Ca e P nel latte
Blood Ca
Bonereabsorption
GI Tract Absorption
Kidney reabsorption
Parathyroid Hormone Calcitonin
Ca
ATP ADP
Transport Mechanism UnclearSeveral exist, passive and active
Chelation of Ca byPO4 in Casein Micelle
Vitamin D Phosphate
ADP
Electron TransportChain
ATP
ADP
CaseinKinase
Active transport into Golgi
Transport Mechanism UnclearSeveral exist, passive and active
Phosphorylationof Caseins

Paresi postpartum • L’improvviso forte drenaggio di calcio nel
post-partum richiede un rapido adattamento del sistema calcitonina/paratormone
• Se ciò non avviene si può verificare la paresi post-partum (milk fever)
• Prevenzione: evitare eccessi di calcio nella dieta in asciutta.
• Trattamento – Infusione di Sali di calcio, tipicamente calcio-
magnesio gluconato.

Le vitamine del latte
Sono molecole indispensabili per le attività fisiologiche dell’organismo, sono trasferite direttamente dal sangue al latte. Si distinguono in
Vitamine Liposolubili Vitamine Idrosolubili
B2 (Riboflavina o lattoflavina, H (Biotina) PP (Niacina) l’acido Pantotenico l’acido Ascorbico (Vitamina C)
A (Retinolo)
β-carotene
D (Colecalciferolo)
E (Tocoferolo).

GG
GGF
Schema dei meccanismi di secrezione dei componenti del latte da parte di una cellula della ghiandola mammaria (SV = vescicole del Golgi; RER = reticolo endoplasmatico rugoso; N = nucleo; BM = membrana basale; GG = globulo di grasso in formazione; GGF = globulo di grasso formato).