







Languages
Pages
Legal


7/23/2019 Translate EPM
http://slidepdf.com/reader/full/translate-epm 1/20
Jika hal ini tidak terjadi, maka model klasik tidak memiliki utilitas. Kedua
MZ dan DZ kembar yang diperlukan untuk memperkirakan kontribusi relatif genetik
dibandingkan kontributor lingkungan umum. Perbedaan usia antara kelompok MZ dan
Pasangan kembar DZ mungkin menjadi faktor pembaur potensial spesifik untuk analisis
epigenetik [8, !. Dengan demikian, pera"atan harus dilakukan untuk men#o#okkan sedekat
mungkin dalam hal ini.
$eritabilitas adalah perkiraan proporsi total fenotipik %hasil&
'arians dalam populasi yang disebabkan aditif efek genetik. Dalam studi kembar
heritabilitas diperkirakan dengan membandingkan tingkat kesamaan fenotipik antara
kelompok MZ dan DZ kembar, baik sebagai tingkat konkordansi atau korelasi intra(kelas.
Dalam konteks kembar, heritabilitas %h )& ditentukan sebagai dua kali perbedaan antara
MZ dan DZ tarif konkordansi %korelasi* h ) + ) %r MZ ( r DZ&, di mana r adalah konkordansi
atau korelasi antara intra#lass setiap jenis kembar [-!. perkiraan heritabilitas
adalah spesifik populasi ke#uali lingkungan konstan. elain itu, kontribusi
faktor stokastik untuk di'ergensi epigenetik yang sama diasumsikan konstan
seluruh populasi dalam analisis ini.
10.2.2 Studi Twin Mengungkapkan Kumulatif Lingkungan
Kontributor 'epigenetik Drift' Selama Waktu
/fek kumulatif dari 'ariasi lingkungan dan sto#hasti# pada perubahan
profil epigenetik yang pertama digambarkan oleh sebuah studi yang banyak dikutip diperiksa
baik genom 'ariasi metilasi lebar dan lokus spesifik D01 di sejumlah ke#il
muda dan setengah baya MZ kembar [!. edangkan kembar MZ 2 tahun menunjukkan
relatif beberapa perbedaan epigenetik dalam pasangan, mereka yang berusia 3- tahun
menunjukkan #ukup 'ariabilitas dalam pasang, dan ini lebih besar jika si kembar memiliki
gaya hidup yang berbeda. ebuah analisis statistik multi(le'el dilakukan yang menghasilkan
deskriptif tunggal nilai untuk setiap jenis ukuran epigenetik. 4ntuk metilasi D01 ini
termasuk %i& jumlah band hadir pada gel %atau titik pada gel )D& antara indi'idu
1 dan 5 dalam sepasang, dan %ii& persentase mutlak 3(methyl#ytosine hadir
dalam indi'idu. Perbedaan ekspresi gen diukur sebagai jumlah
gen menunjukkan ekspresi diferensial %adjusted p(nilai berikut 10671&
antara saudara kandung. ingkat asetilasi histon diukur sebagai tinggi pun#ak relatif
berikut kromatografi #air kinerja tinggi.

7/23/2019 Translate EPM
http://slidepdf.com/reader/full/translate-epm 2/20
Kesamaan setiap nilai deskriptif dalam pasangan kemudian diperkirakan sebagai
/u#lidean kuadrat jarak dengan mengurangkan nilai masing(masing. 4ntuk
meneliti hubungan antara fenotip 9 data lingkungan dan epigenetik
'ariabel, kategori analisis komponen prinsip itu firstly diterapkan dengan aslinya
kuesioner 'ariabel untuk mengurangi mereka untuk dua komponen berkorelasi disebut
:penuaan: %meliputi umur, berat badan dan tinggi& dan :kesehatan: %semua 'ariabel penyakit
dan pera"atan farmakologis&. Model #ampuran yang digunakan untuk memperkirakan
kontribusi setiap a#ak(efek 'arians dari 'ariabel dependen dan 'arians
Prosedur komponen dinilai dengan 10671. $ubungan nilai tunggal
dihasilkan untuk penuaan dan kesehatan 'ariabel kuesioner yang diturunkan data epigenetik
die'aluasi dengan uji Pearson %di#atat bah"a tipe # spesifik tes tidak dijelaskan&.
Dengan menggunakan pendekatan ini kesimpulan umum adalah bah"a profi le epigenetik
adalah konstan ;Melayang;, meskipun sangat sedikit kembar muda dipelajari dan presisi
statistik adalah rendah [!. elain itu, ini adalah studi #ross se#tional dan tidak menge'aluasi
metilasi dalam indi'idu yang sama dari "aktu ke "aktu. 1khirnya, spesifik # epigenetik
analisis dipekerjakan semua resolusi rendah dalam bah"a mereka tidak memeriksa distribusi
perbedaan epigenetik dalam genom, melainkan memberikan :snapshot: global
kelas yang berbeda dari perbedaan epigenetik dalam pasangan kembar.
Penelitian pertama untuk menguji hanyut epigenetik longitudinal pada kembar yang
terlibat analisis bukal metilasi D01 sel dalam 2 gen di <= MZ dan DZ <3 pada 3 dan
- tahun [!. Perubahan longitudinal dalam indi'idu dihitung dengan
menilai korelasi dalam metilasi pada setiap usia. 4ntuk menilai kontribusi relatif
dari di"ariskan dan lingkungan komponen stokastik 9 ke tingkat metilasi, korelasi
dalam MZ pasangan dibandingkan dengan korelasi dalam pasangan DZ. 1khirnya,
untuk menilai kontribusi relatif dari di"ariskan dan lingkungan 9 sto#hasti#
komponen perubahan metilasi D01 dari "aktu ke "aktu, perubahan intra(indi'idu
kor dihitung dan korelasi dalam pasangan kembar MZ dibandingkan dengan
korelasi dalam pasangan DZ. Penelitian ini adalah yang pertama untuk mengungkapkan
sejauh mana epigenetik kejanggalan pada kembar MZ pada a"al kehidupan dan menyoroti
ketidakstabilan berkelanjutan tingkat metilasi dari "aktu ke "aktu. >ang penting daerah
genom yang berbeda ditemukan menunjukkan berbagai tingkat perbedaan epigenetik dari
"aktu ke "aktu.

7/23/2019 Translate EPM
http://slidepdf.com/reader/full/translate-epm 3/20
Pemeriksaan tingkat metilasi D01 di beberapa situs di beberapa jaringan dari
kembar yang baru lahir kerahasiaan rmed yang hanyut epigenetik antara indi'idu genetik
identik %MZ kembar& dimulai di dalam rahim dan jaringan(spesifik # #ara. Dalam penelitian
ini, koefisien korelasi sien intra#lass %?@@& lebih tinggi pada MZ daripada di DZ kembar [)!
%Aambar. -.& mendukung temuan sebelumnya peran untuk genetik faktor 9 di"ariskan
dalam pembentukan profile epigenetik.
Melayang epigenetik dari "aktu ke "aktu telah diamati untuk lajang dan mungkin
juga bah"a pemeliharaan tanda epigenetik metilasi D01 adalah di ba"ah kontrol genetik
[2!. 0amun, perubahan longitudinal pada tanda epigenetik yang paling mungkin terjadi
dalam subset gen [<!, mungkin pada mereka dengan tingkat metilasi menengah,
daripada mereka yang sepenuhnya unmethylated atau dalam keadaan hypermethylated.
tudi terbatas kini mulai memanfaatkan model kembar untuk memperkirakan relatif
kontribusi faktor genetik dan lingkungan dalam mediasi respon dari
mengembangkan epigenome ke spesifik paparan lingkungan. Pemeriksaan
tingkat metilasi pada 2 independen pen#etakan terkait DMB %K'DMB, P/A
dan $ 9 ?AC)& di 3 pasang kembar %termasuk ) pasang dikandung melalui bayi tabung&
tidak mengungkapkan perbedaan #ant signifikan pada tiga DMBs antara ?7C(dikandung dan
se#ara alami dikandung kembar. 0amun, baik ukuran sampel dan daerah genom
diperiksa yang terlalu ke#il untuk kesimpulan umum yang akan dibuat dan analisis lebih
lanjut di kelompok yang lebih besar dari kembar dibenarkan [3!.
Gambar. 10.1 Distribusi korelasi intra#lass mengungkapkan korelasi sien efisien median
yang lebih tinggi untuk Metilasi D01 pada ?AC) 9 $ lokus di MZ dari pasangan kembar
DZ di lima jaringan dari bayi yang baru lahir. ?@@ mengukur proporsi dari total 'arians
disebabkan dalam 'ariasi pasangan dalam MZ dan kelompok DZ kembar. 1nalisis ?@@. el
mononuklear darah tali @5M@(* manusia $47/@( sel endotel 'ena umbilikalis. %Aambar
diadaptasi dari [)!&
10. !pigenetik dan Model "umbang Twin
10..1 #"um"i $dentita" Genetik
5anyak penelitian telah melaporkan perbedaan fenotipik antara MZ #o(kembar
%4lasan di [=!&. Pasangan MZ sumbang telah terbukti sangat berharga dalammengungkapkan kontribus 'ariasi non(genetik untuk penetrasi penyakit, etiologi, atau efek

7/23/2019 Translate EPM
http://slidepdf.com/reader/full/translate-epm 4/20
dari terapi. 1sumsi yang mendasari dalam semua studi tersebut adalah bah"a MZ kembar
identik se#ara genetik. ?ni telah dibantah langsung dalam beberapa kasus, di mana baik
spesifik titik mutasi [( !, uniparental disomy [)-!, triplet ekspansi ulangi [), ))!,
kromosom mosai#ism [)2! atau heteroplasmi untuk mutasi mitokondria(dikodekan
[)<, )3! atau aneuploidi kromosom %4lasan di [=!& semuanya telah dikaitkan dengan
perbedaan # fenotipik spesifik pada kembar monoEigot. >ang penting, perbedaan
baik jumlah salinan [)=! dan panjang telomer [), )8! telah juga dijelaskan dalam
fenotip sumbang dan sesuai pasangan kembar MZ. 0amun demikian, laporan tersebut
relatif jarang dan kemungkinan bah"a non(genetik %epigenetik& 'ariasi dalam MZ
pasangan kembar mendasari mayoritas diamati kejanggalan fenotipik.
10..2 men%impulkan &en%ebab epigenetik #nali"i" genetik ("
Karena setiap sel dalam indi'idu berasal dari Eigot tunggal dengan satu genom,
tudi genetik umumnya hanya membutuhkan sampel D01 tunggal per indi'idu, diambil dari
setiap jaringan pada usia berapa pun. Penyebab dan efek dapat diandalkan diprediksi, atau
dike#ualikan, sebagai 'ariasi genomik tidak dianggap 'ariabel selama hidup %ke#uali
dalam kaitannya dengan :langka: mutasi somatik&. 1turan tersebut tidak berlaku untuk
sebagian besar epigenetik tudi mana diulang sampel dari lebih dari satu jaringan mungkin
diinginkan. 5ahkan jika biospe#imen dikumpulkan sangat a"al dalam hidup %sebelum
manifestasi fenotipe& mengungkap sebab dan akibat yang bermasalah. Meskipun ini dan
banyak peringatan lain yang terkait dengan menyimpulkan sebab(akibat dalam hubungan
epigenetik Penelitian %dibahas dalam bab(bab lain dari buku ini&, beberapa peneliti telah
berusaha untuk menghubungkan spesifik perubahan epigenetik untuk fenotipe penyakit.
1nehnya beberapa studi telah berusaha untuk memanfaatkan sumbang MZ Model kembar
pasangan, meskipun menunjukkan utilitas dari pendekatan ini dalam mengidentifikasi faktor
spesifik risiko penyakit,
10.. Lo)ali"ed *aria"i epigenetik fenotip
Twin" "umbang
5eberapa studi telah berusaha untuk mengidentifikasi epigenetik :menyebabkan: dari
manusia yang kompleks Aangguan melalui resolusi rendah, pemeriksaan #alon hipotesis gen
'ariasi epigenetik %terutama metilasi D01& di spesifik gen # daerah di fenotip
kembar MZ sumbang %dirangkum dalam abel -.&. Kontribusi
'ariasi epigenetik untuk heritabilitas fenotipe kompleks kompleks dan kemungkinan
diatur di berbagai tingkatan %Aambar. -.)&. e#ara umum, ukuran efek telah
5eberapa #ontoh minimal dan ada replikasi independen diidentifikasi
7/23/2019 Translate EPM
http://slidepdf.com/reader/full/translate-epm 5/20
asosiasi. Dengan demikian, rele'ansi biologis temuan tersebut tetap
dipertanyakan.
10..+ miring ,-Kromo"om $nakti(a"i di fenotip
Twin" "umbang
alah satu fenomena epigenetik yang paling banyak dipelajari pada mamalia
melibatkan inakti'asi mayoritas gen pada satu kromosom F pada "anita. :Dosis ini
kompensasi :eGualises ekspresi sebagian besar gen F(kromosom pada laki(laki
%F>& dan perempuan %FF& dan biasanya terjadi se#ara a#ak, dengan sekitar 3-H
sel pada "anita yang menunjukkan inakti'asi F dari ibu yang diturunkan, sedangkan
3-H menunjukkan inakti'asi lain dari kromosom F dari ayah yang diturunkan. 0amun,
dalam beberapa indi'idu pola miring dari F(inakti'asi jelas, dengan satu orang tua
F, lebih di"akili dalam jaringan atau sel jenis tertentu. Dalam hal demikian, sebaliknya
mutasi resesif dapat memiliki efek samping yang mendalam fenotipik. Dengan demikian,
miring F(inakti'asi, atau lebih tepatnya inakti'asi kesempatan alel fungsional tunggal
dalam jaringan fisiologis yang rele'an, telah dikaitkan dengan kejanggalan untuk
beberapa gangguan termasuk hemofilia [)!, sindrom Cragile(F [)), 2-! dan
Du#henne Mus#ular Dystrophy [2!.
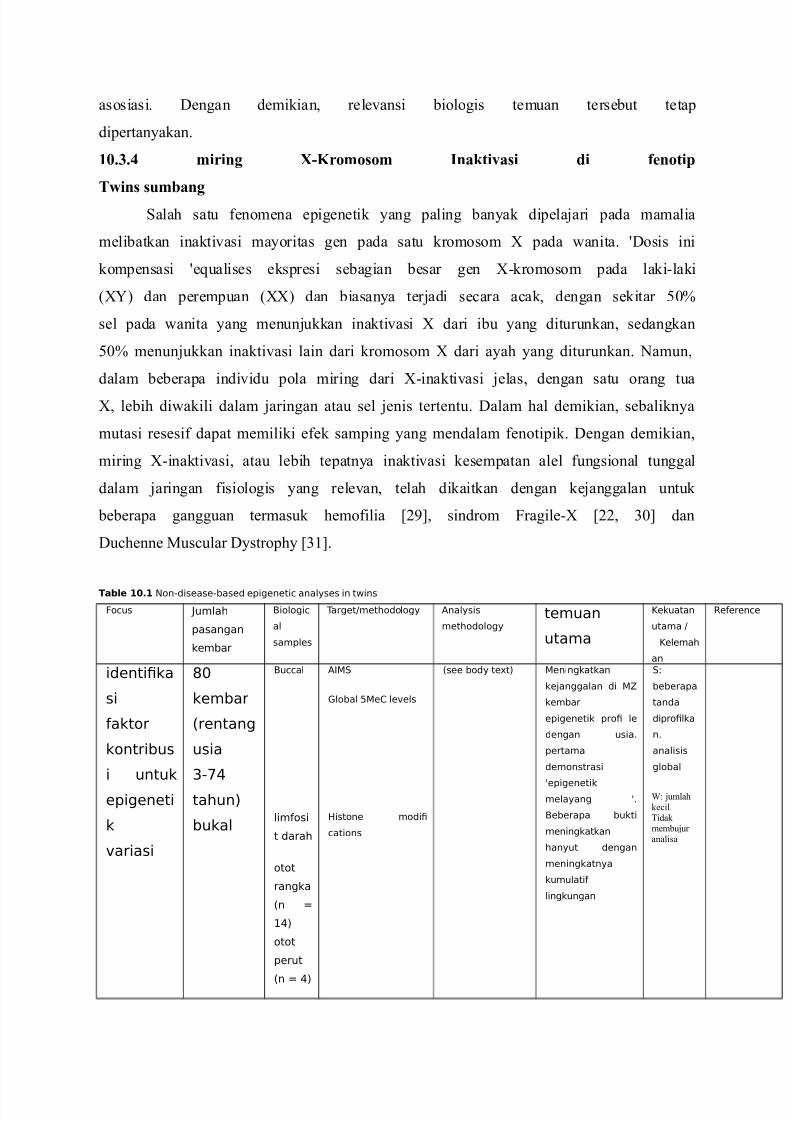
Table 10.1 Non-disease-based epigenetic analyses in twins
Focus Jumlah
pasangan
kembar
Biologic
al
samples
Target/methodology Analysis
methodologytemuan
utama
Kekuatan
utama /
Kelemah
an
Reerence
identi!ka
si
aktor
kontribus
i untuk
epigeneti
k
"ariasi
#$
kembar
%rentang
usia
&-'(
tahun)
bukal
Buccal
limosi
t darah
otot
rangka
%n *
+()
otot
perut
%n * ()
A,.
lobal 0e1 le"els
2istone modi!
cations
%see body te3t) eningkatkan
ke4anggalan di 5
kembar
epigenetik pro! le
dengan usia6
pertama
demonstrasi
7epigenetikmelayang 76
Beberapa bukti
meningkatkan
hanyut dengan
meningkatnya
kumulati
lingkungan
.8
beberapa
tanda
dipro!lka
n6
analisis
global
I jumlahke#il.idak membujur analisa
7/23/2019 Translate EPM
http://slidepdf.com/reader/full/translate-epm 6/20
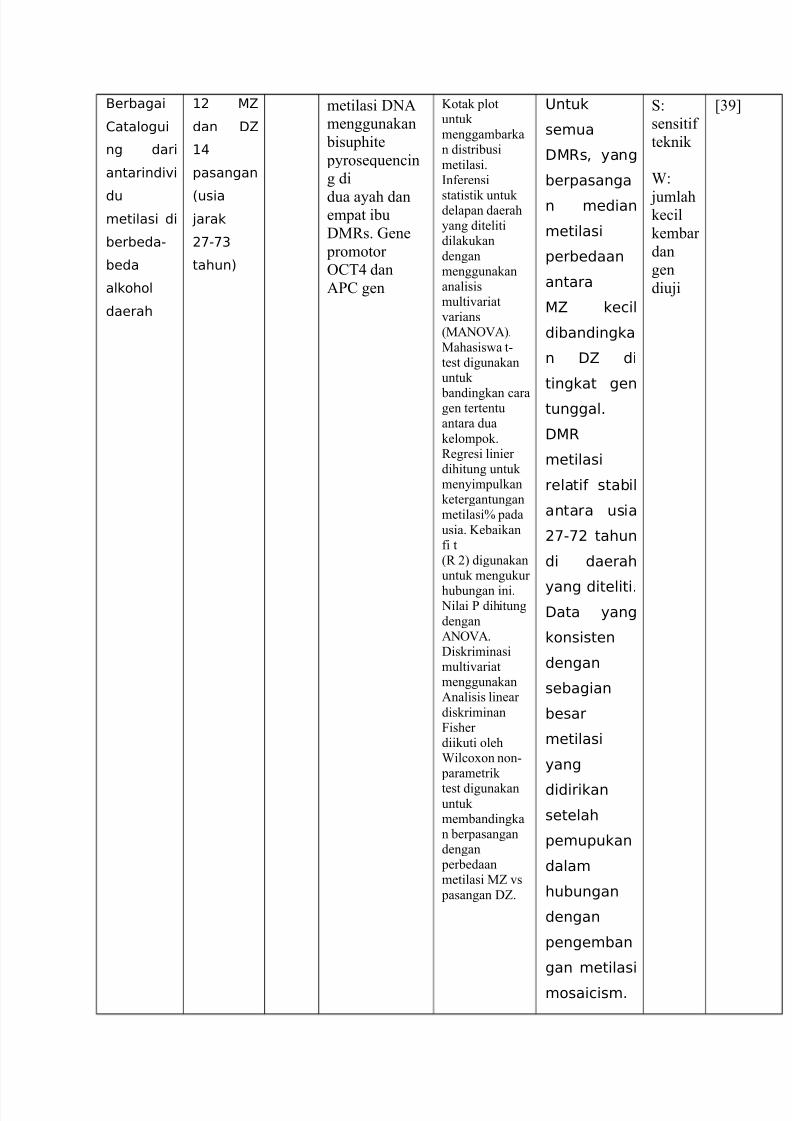
Berbagai
1atalogui
ng dari
antarindi"i
du
metilasi di
berbeda-
beda
alkohol
daerah
+9 5
dan :5
+(
pasangan
%usia
4arak
9'-'&
tahun)
metilasi D01
menggunakan
bisuphite
pyroseGuen#in
g di
dua ayah danempat ibu
DMBs. Aene
promotor
6@< dan
1P@ gen
Kotak plotuntukmenggambarkan distribusi
metilasi.?nferensi
statistik untuk delapan daerahyang ditelitidilakukan
denganmenggunakananalisismulti'ariat'arians%M10671&.
Mahasis"a t(test digunakanuntuk
bandingkan #aragen tertentuantara dua
kelompok.Begresi linier dihitung untukmenyimpulkanketergantunganmetilasiH pada
usia. Kebaikanfi t%B )& digunakanuntuk mengukur
hubungan ini. 0ilai P dihitung
dengan10671.Diskriminasimulti'ariatmenggunakan1nalisis linear
diskriminanCisher diikuti olehIil#oon non( parametrik
test digunakanuntukmembandingkan berpasangandengan perbedaanmetilasi MZ 's
pasangan DZ.
;ntuk
semua
:Rs< yang
berpasanga
n median
metilasi
perbedaan
antara
5 kecil
dibandingka
n :5 di
tingkat gen
tunggal6
:R
metilasi
relati stabil
antara usia
9'-'9 tahun
di daerah
yang diteliti6:ata yang
konsisten
dengan
sebagian
besar
metilasi
yang
didirikan
setelah
pemupukan
dalam
hubungan
dengan
pengemban
gan metilasi
mosaicism6
sensitif
teknik
I
jumlahke#il
kembar
dan
gen
diuji
[2!
7/23/2019 Translate EPM
http://slidepdf.com/reader/full/translate-epm 7/20
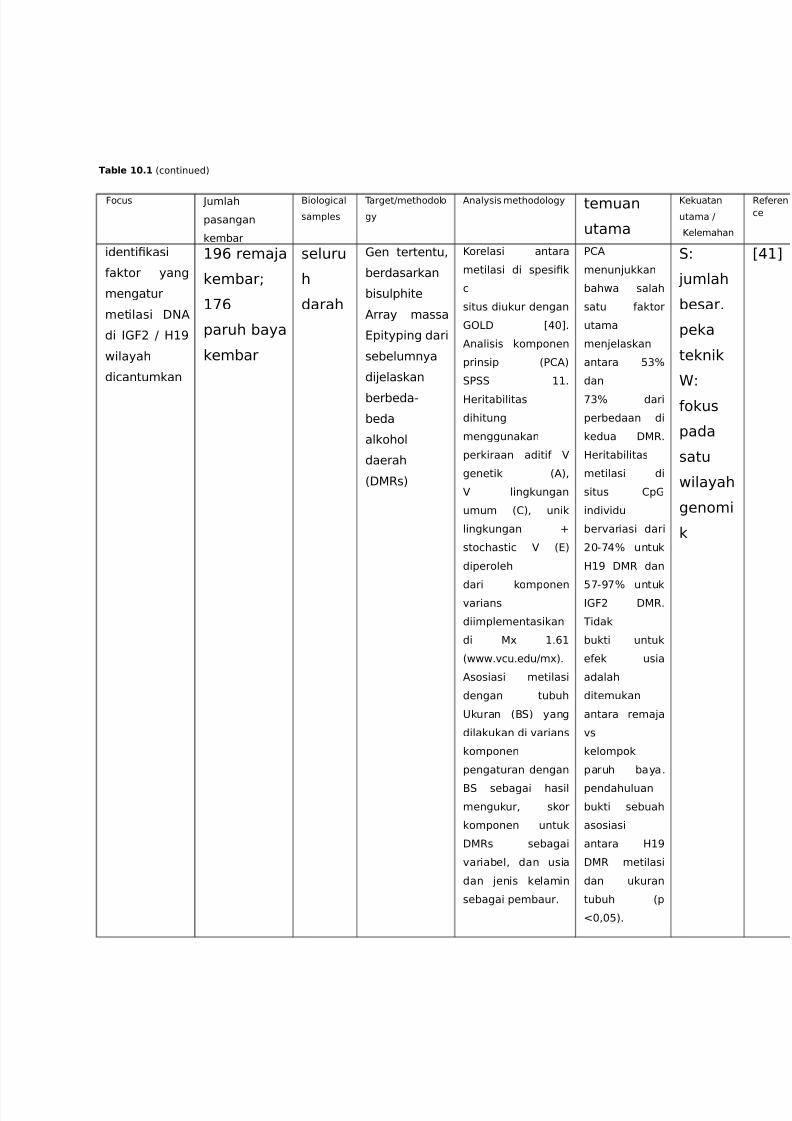
Table 10.1 %continued)
Focus Jumlah
pasangan
kembar
Biological
samples
Target/methodolo
gy
Analysis methodology temuan
utama
Kekuatan
utama /
Kelemahan
Reeren
ce
identi!kasi
aktor yang
mengatur
metilasi :NA
di ,F9 / 2+=
wilayah
dicantumkan
+=> rema4a
kembar?
+'>
paruh baya
kembar
seluru
h
darah
en tertentu<
berdasarkan
bisulphite
Array massa
@pityping dari
sebelumnya
di4elaskan
berbeda-
beda
alkohol
daerah
%:Rs)
Korelasi antara
metilasi di spesi!k
c
situs diukur dengan
: C($D6
Analisis komponen
prinsip %E1A)
.E.. ++6
2eritabilitas
dihitung
menggunakan
perkiraan aditi
genetik %A)<
lingkungan
umum %1)< unik
lingkungan G
stochastic %@)
diperoleh
dari komponen"arians
diimplementasikan
di 3 +6>+
%www6"cu6edu/m3)6
Asosiasi metilasi
dengan tubuh
;kuran %B.) yang
dilakukan di "arians
komponen
pengaturan dengan
B. sebagai hasil
mengukur< skor
komponen untuk
:Rs sebagai
"ariabel< dan usia
dan 4enis kelamin
sebagai pembaur6
E1A
menun4ukkan
bahwa salah
satu aktor
utama
men4elaskan
antara 0&H
dan
'&H dari
perbedaan di
kedua :R6
2eritabilitas
metilasi di
situs 1p
indi"idu
ber"ariasi dari
9$-'(H untuk
2+= :R dan
0'-='H untuk,F9 :R6
Tidak
bukti untuk
eek usia
adalah
ditemukan
antara rema4a
"s
kelompok
paruh baya6
pendahuluan
bukti sebuah
asosiasi
antara 2+=
:R metilasi
dan ukuran
tubuh %p
I$<$0)6
.8
4umlah
besar6
peka
teknik
8
okus
pada
satu
wilayah
genomi
k
C(+D
7/23/2019 Translate EPM
http://slidepdf.com/reader/full/translate-epm 8/20
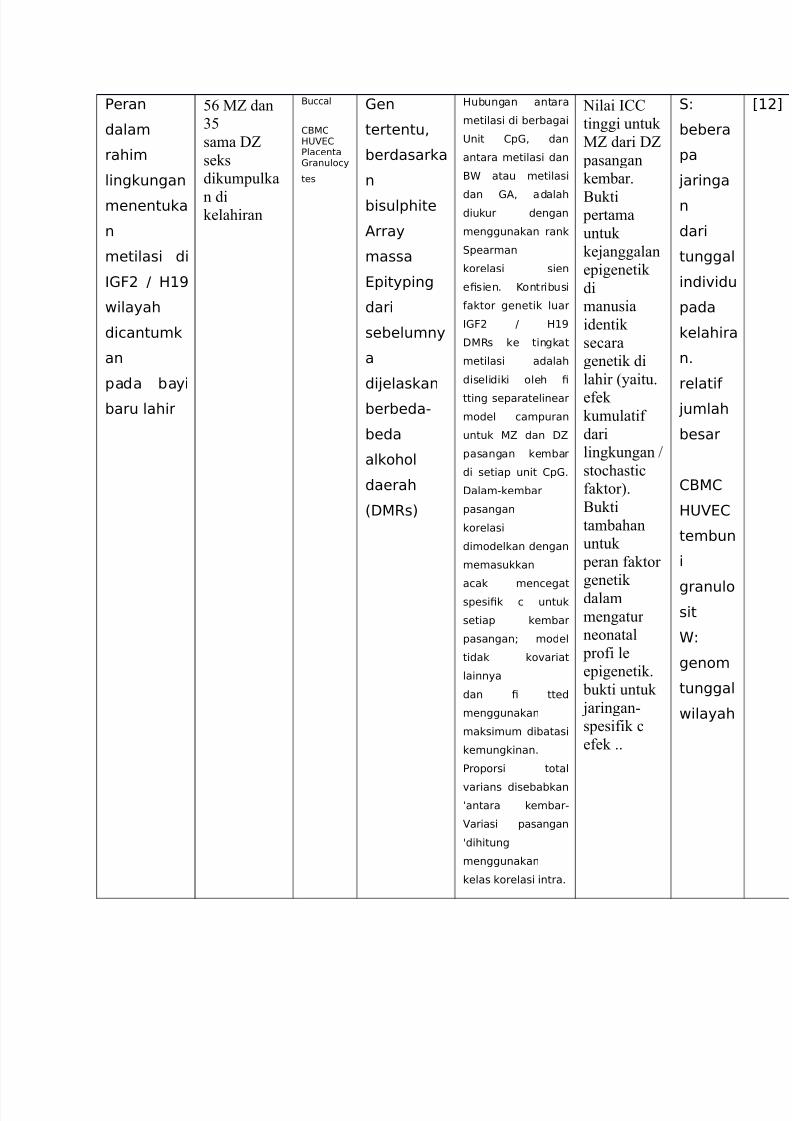
Eeran
dalam
rahim
lingkungan
menentuka
n
metilasi di
,F9 / 2+=
wilayah
dicantumk
an
pada bayi
baru lahir
3= MZ dan
23
sama DZ
seks
dikumpulka
n dikelahiran
Buccal
1B12;@1Elacentaranulocy
tes
en
tertentu<
berdasarka
n
bisulphite
Array
massa
@pityping
dari
sebelumny
a
di4elaskan
berbeda-
beda
alkohol
daerah
%:Rs)
2ubungan antara
metilasi di berbagai
;nit 1p< dan
antara metilasi dan
B atau metilasi
dan A< adalahdiukur dengan
menggunakan rank
.pearman
korelasi sien
e!sien6 Kontribusi
aktor genetik luar
,F9 / 2+=
:Rs ke tingkat
metilasi adalah
diselidiki oleh !
tting separatelinear
model campuran
untuk 5 dan :5
pasangan kembar
di setiap unit 1p6
:alam-kembar
pasangan
korelasi
dimodelkan dengan
memasukkan
acak mencegat
spesi!k c untuk
setiap kembar
pasangan? model
tidak ko"ariat
lainnya
dan ! tted
menggunakan
maksimum dibatasi
kemungkinan6
Eroporsi total
"arians disebabkan
7antara kembar-
ariasi pasangan
7dihitung
menggunakan
kelas korelasi intra6
0ilai ?@@
tinggi untuk
MZ dari DZ
pasangan
kembar.
5ukti pertama
untuk
kejanggalan
epigenetik
di
manusia
identik
se#ara
genetik di
lahir %yaitu.
efekkumulatif
dari
lingkungan 9
sto#hasti#
faktor&.
5ukti
tambahan
untuk
peran faktor
genetik
dalam
mengatur
neonatal
profi le
epigenetik.
bukti untuk
jaringan(
spesifik #
efek ..
.8
bebera
pa
4aringa
n
dari
tunggal
indi"idu
pada
kelahira
n6
relati
4umlah
besar
1B1
2;@1
tembun
i
granulosit
8
genom
tunggal
wilayah
C+9D
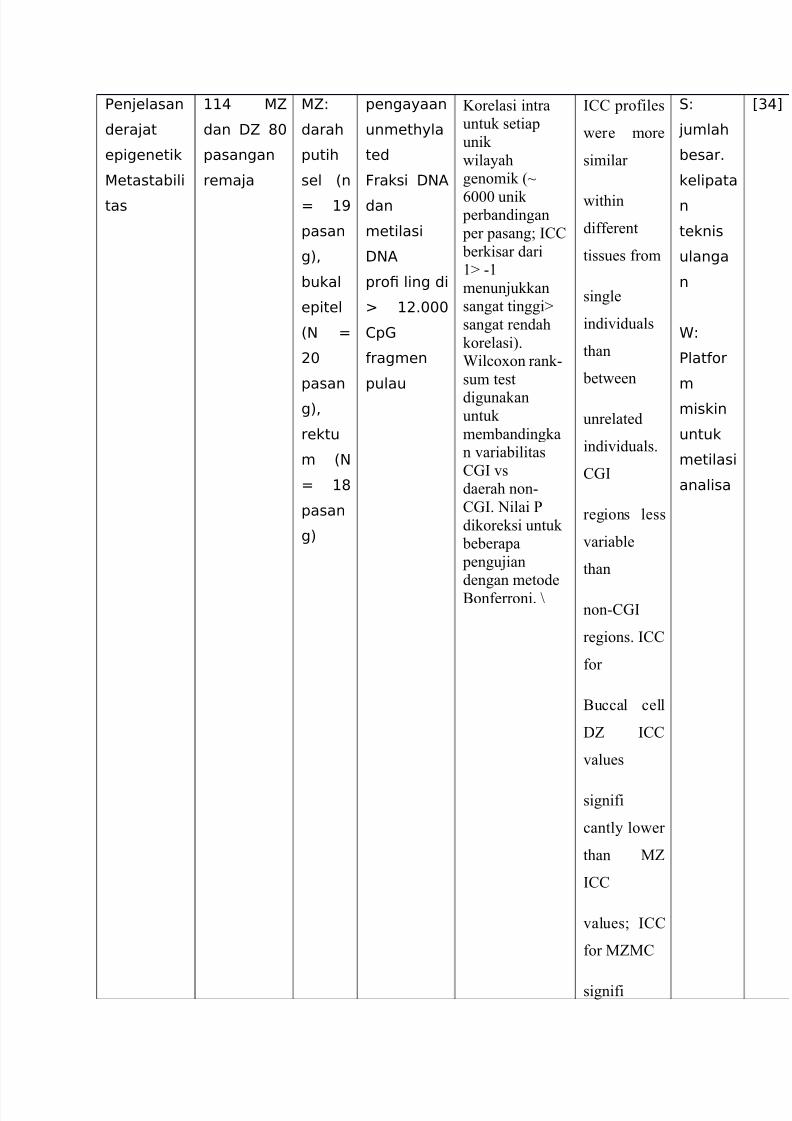
7/23/2019 Translate EPM
http://slidepdf.com/reader/full/translate-epm 9/20
Een4elasan
dera4at
epigenetik
etastabili
tas
++( 5
dan :5 #$
pasangan
rema4a
58
darah
putih
sel %n
* +=
pasan
g)<
bukal
epitel
%N *
9$
pasan
g)<
rektu
m %N
* +#
pasan
g)
pengayaan
unmethyla
ted
Fraksi :NA
dan
metilasi
:NA
pro! ling di
+96$$$
1p
ragmen
pulau
Korelasi intra
untuk setiap
unik
"ilayah
genomik %L
=--- unik perbandingan
per pasang* ?@@
berkisar dari
(
menunjukkan
sangat tinggi
sangat rendah
korelasi&.
Iil#oon rank(
sum test
digunakanuntuk
membandingka
n 'ariabilitas
@A? 's
daerah non(
@A?. 0ilai P
dikoreksi untuk
beberapa
pengujian
dengan metode
5onferroni. N
?@@ profiles
"ere more
similar
"ithin
different
tissues from
single
indi'iduals
than
bet"een
unrelated
indi'iduals.
@A?
regions less
'ariable
than
non(@A?
regions. ?@@
for
5u##al #ell
DZ ?@@
'alues
signifi
#antly lo"er
than MZ
?@@
'alues* ?@@
for MZM@
signifi
.8
4umlah
besar6
kelipata
n
teknis
ulanga
n
8
Elator
m
miskin
untuk
metilasi
analisa
C&(D
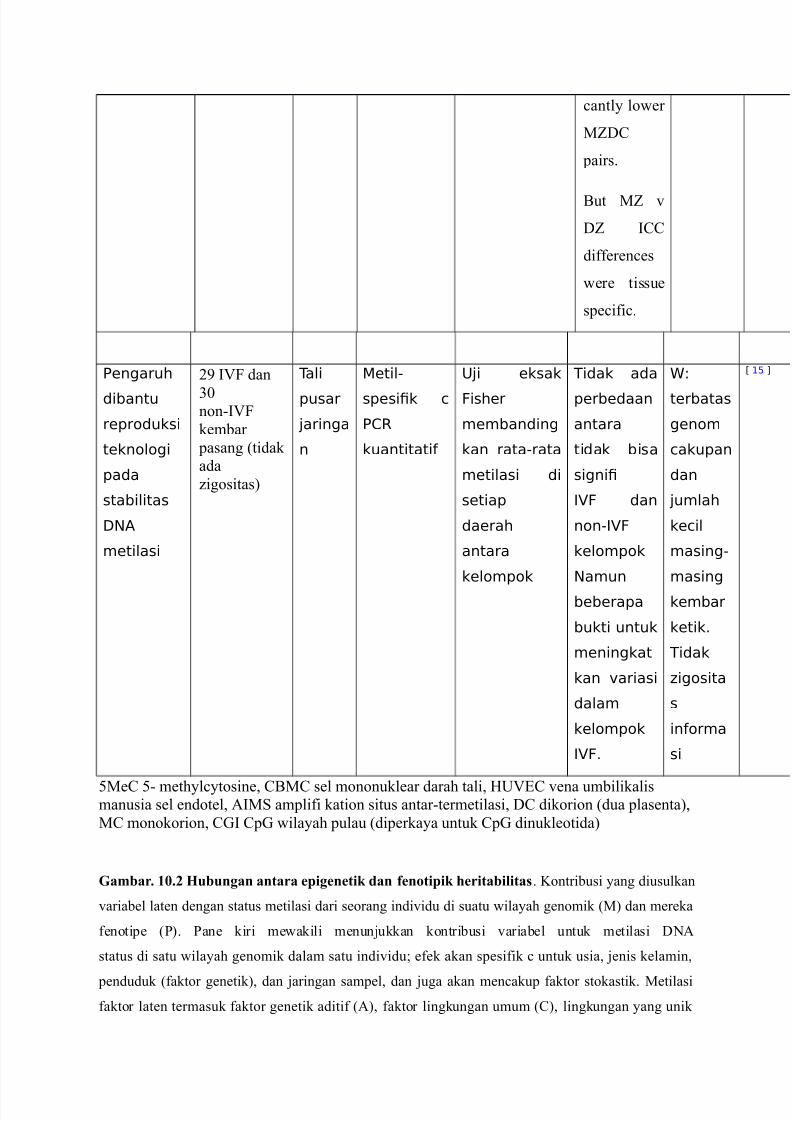
7/23/2019 Translate EPM
http://slidepdf.com/reader/full/translate-epm 10/20
#antly lo"er
MZD@
pairs.
5ut MZ '
DZ ?@@
differen#es
"ere tissue
spe#ifi#.
Eengaruh
dibantu
reproduksi
teknologi
pada
stabilitas
:NA
metilasi
) ?7C dan
2-non(?7C
kembar
pasang %tidak
ada
Eigositas&
Tali
pusar
4aringa
n
etil-
spesi!k c
E1R
kuantitati
;4i eksak
Fisher
membanding
kan rata-rata
metilasi di
setiap
daerah
antara
kelompok
Tidak ada
perbedaan
antara
tidak bisa
signi!
,F dan
non-,F
kelompok
Namun
beberapa
bukti untuk
meningkat
kan "ariasi
dalam
kelompok
,F6
8
terbatas
genom
cakupan
dan
4umlah
kecil
masing-
masing
kembar
ketik6
Tidak
Ligosita
s
inorma
si
C +0 D
3Me@ 3( methyl#ytosine, @5M@ sel mononuklear darah tali, $47/@ 'ena umbilikalismanusia sel endotel, 1?M amplifi kation situs antar(termetilasi, D@ dikorion %dua plasenta&,
M@ monokorion, @A? @pA "ilayah pulau %diperkaya untuk @pA dinukleotida&
Gambar. 10.2 ubungan antara epigenetik dan fenotipik /eritabilita". Kontribusi yang diusulkan
'ariabel laten dengan status metilasi dari seorang indi'idu di suatu "ilayah genomik %M& dan mereka
fenotipe %P&. Pane kiri me"akili menunjukkan kontribusi 'ariabel untuk metilasi D01
status di satu "ilayah genomik dalam satu indi'idu* efek akan spesifik # untuk usia, jenis kelamin,
penduduk %faktor genetik&, dan jaringan sampel, dan juga akan men#akup faktor stokastik. Metilasi
faktor laten termasuk faktor genetik aditif %1&, faktor lingkungan umum %@&, lingkungan yang unik

7/23/2019 Translate EPM
http://slidepdf.com/reader/full/translate-epm 11/20
%/&, dan faktor epigenetik di"ariskan dan stabil yang tidak tergantung urutan D01
%/p&. Pane kanan merupakan model jalan di kembar, yang menggambarkan kontribusi D01
Metilasi dan faktor lainnya untuk fenotip %P& di kembar saya dengan korelasi memperkirakan di MZ
%Oet t& dan DZ %Bigh t& kembar 'ariabel laten termasuk efek genetik aditif %1i&, lingkungan umum
%@i&, metilasi D01 %Mi&, dan lingkungan yang unik %/i&. Perkiraan korelasi yang
diperoleh dari genetik [28! dan studi epigenetik sebelumnya [2<! pada kembar. Dalam saudara,
korelasi di M akan lebih rendah dari yang diamati pada DZ kembar karena perbedaan usia dan terkait
peningkatan tingkat perubahan stokastik kumulatif. %Aambar diadaptasi dari [-!&
10.. Genome-Skala epigenetik $n(e"tiga"i di Twin"
Mengingat relatif mudah se#ara bersamaan pemetaan profil metilasi D01 di
daerah besar genom relatif terhadap tatanan yang lebih tinggi tanda epigenetik, tidak
mengherankan bah"a studi kembar terbatas dilakukan di ruang ini telah berfokus
pada metilasi D01 %abel -.)&.
10...1 e"olu"i Menenga/ 1-2 Tempat per Gene di 1-103 dari Gen
dalam Genome4
Penelitian terbaru telah mulai untuk mengungkapkan sejauh mana sebenarnya metilasi
D01 genom perubahan MZ kembar yang berpotensi bisa menjelaskan kejanggalan fenotip.
Mungkin aplikasi gabungan paling sukses dari model kejanggalan kembar dan epigenetik
analisis telah diselidiki asosiasi perubahan metilasi D01 dengan gangguan kekebalan terkait,
lupus eritematosus sistemik %O/&, rheumatoid arthritis %B1& dan dermatomiositis %DM& [2)!.
Pendekatan berjenjang untuk analisis yang terlibat
Table 10.2 @3amples o epigenetic analysis in discordant twin pairs
Focus Jumlah
pasangan
kembar
Biological
samples
Target/methodology Analysis
methodologytemuan utama Reerence
7/23/2019 Translate EPM
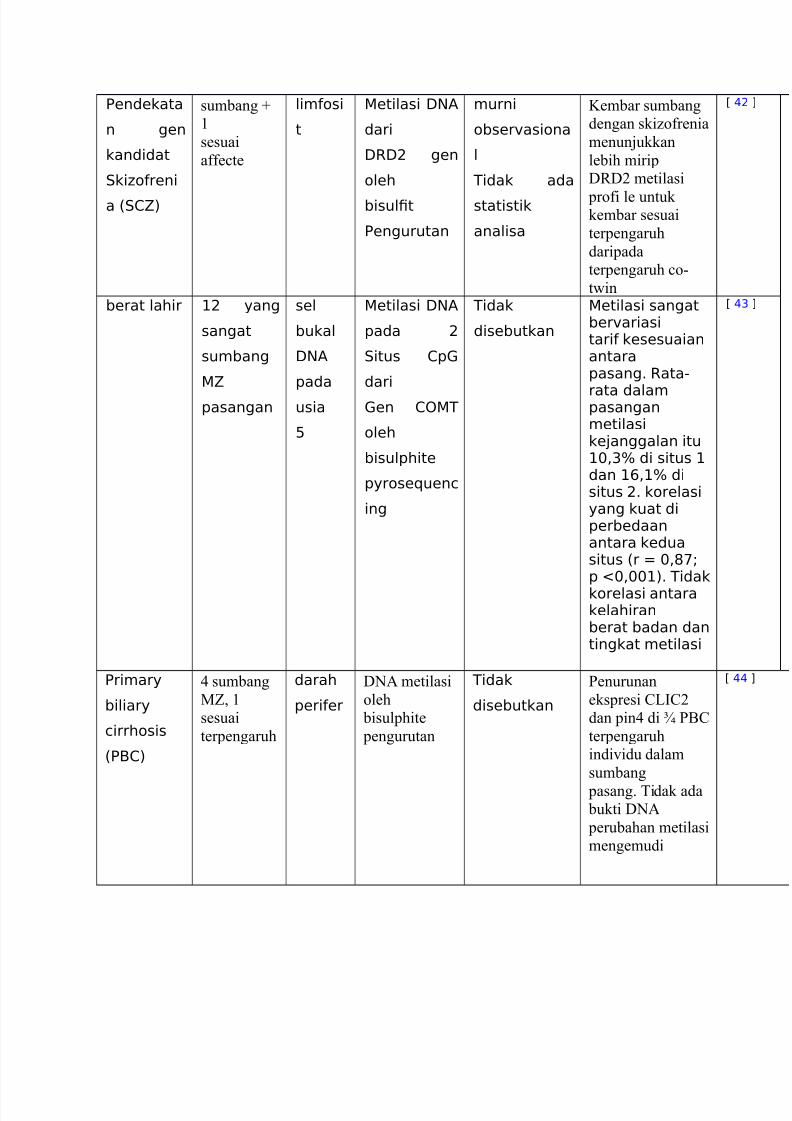
http://slidepdf.com/reader/full/translate-epm 12/20
Eendekata
n gen
kandidat
.kiLoreni
a %.15)
sumbang
sesuai
affe#te
limosi
t
etilasi :NA
dari
:R:9 gen
oleh
bisul!t
Eengurutan
murni
obser"asiona
l
Tidak ada
statistik
analisa
Kembar sumbang
dengan skiEofrenia
menunjukkan
lebih mirip
DBD) metilasi
profi le untuk kembar sesuai
terpengaruh
daripada
terpengaruh #o(
t"in
C (9 D
berat lahir +9 yang
sangat
sumbang
5
pasangan
sel
bukal
:NA
pada
usia
0
etilasi :NA
pada 9
.itus 1p
dari
en 1T
oleh
bisulphite
pyroseMuenc
ing
Tidak
disebutkan
etilasi sangatber"ariasitari kesesuaianantarapasang6 Rata-
rata dalampasanganmetilasike4anggalan itu+$<&H di situs +dan +><+H disitus 96 korelasiyang kuat diperbedaanantara keduasitus %r * $<#'?p I$<$$+)6 Tidak
korelasi antarakelahiranberat badan dantingkat metilasi
C (& D
Erimary
biliary
cirrhosis
%EB1)
< sumbang
MZ,
sesuai
terpengaruh
darah
perier
D01 metilasi
oleh
bisulphite
pengurutan
Tidak
disebutkan
Penurunan
ekspresi @O?@)
dan pin< di Q P5@
terpengaruh
indi'idu dalam
sumbang
pasang. idak ada bukti D01
perubahan metilasi
mengemudi
C (( D

7/23/2019 Translate EPM
http://slidepdf.com/reader/full/translate-epm 13/20
Beckwith
iedeman
n
.yndrome
%B.)
+$
sumbang
5
0 Kendali
5
!brobl
as kulit
dan /
atau
sekelili
ng
darah
:NA metilasi
oleh
Blotting
selatan
urni
obser"asional
ada statistik
analisa
5I terpengaruh
indi'idu
menunjukkan
kehilangan
pen#etakan di
K7DMB dengan bersifat bialel
ekspresi
K@0R6
C (0 D
Eerak-
Russell
.yndrom
e
%.R.)
+ sumbang
5
pasangan
darah
perier
leukosit
Dikombinasika
n bisulfi te
analisis restriksi
%@65B1& dan
bisulfi te
seGuen#ing
untuk dua $
DMBs
urni
obser"asion
al ada
statistik
analisa
Kehilangan $
(DMB di sekitar
setengah dari sel(
sel di terpengaruh
kembar hanya
dengan terkait
penurunan
ekspresi ?AC)
C (> D
ekor
:uplikasi
Anomali
%1:A)
sumbang
5
pasangan
=
terpengaru
h 5
darah
perier
mononukl
ear
sel
etilasi :NA
dari
-A3in + oleh
bisul!t
Eengurutan
binomialumum
linear
di#ampur
Model %lme<
paket&
ebih tinggi
metilasi di
A3in-+
promotor di
terpengaruh "s
terpengaruh
co-twin
%p I$<$$$+)6
lebih tinggi
metilasi di
kedua kembar
dibandingkan
kontrol %p *
$<$9)6
Korelasi ,11
$<'> untuk
semua +$
C (' D
7/23/2019 Translate EPM
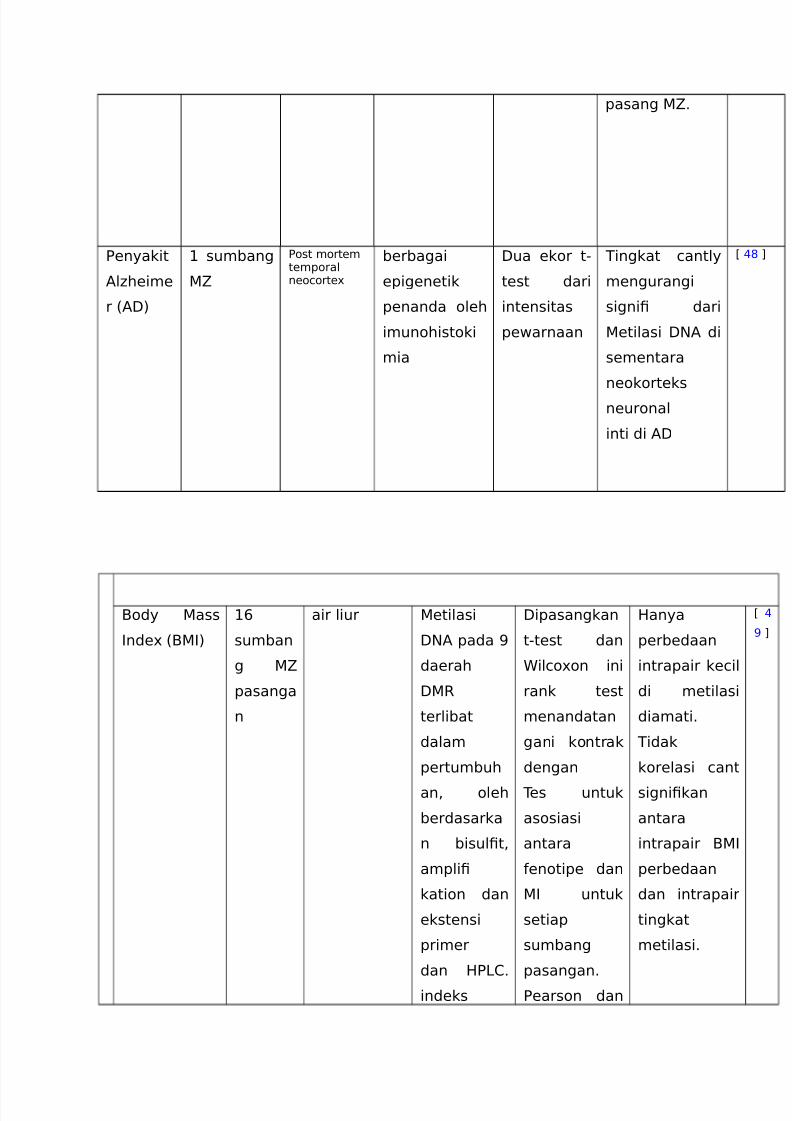
http://slidepdf.com/reader/full/translate-epm 14/20
pasang 56
Eenyakit
AlLheime
r %A:)
+ sumbang
5
Eost mortemtemporalneocorte3
berbagai
epigenetik
penanda oleh
imunohistoki
mia
:ua ekor t-
test dari
intensitas
pewarnaan
Tingkat cantly
mengurangi
signi! dari
etilasi :NA di
sementara
neokorteks
neuronal
inti di A:
C (# D
Body ass
,nde3 %B,)
+>
sumban
g 5
pasanga
n
air liur etilasi
:NA pada =
daerah
:R
terlibat
dalam
pertumbuh
an< oleh
berdasarka
n bisul!t<
ampli!
kation dan
ekstensi
primer
dan 2E16
indeks
:ipasangkan
t-test dan
ilco3on ini
rank test
menandatan
gani kontrak
dengan
Tes untuk
asosiasi
antara
enotipe dan
, untuk
setiap
sumbang
pasangan6
Eearson dan
2anya
perbedaan
intrapair kecil
di metilasi
diamati6
Tidak
korelasi cant
signi!kan
antara
intrapair B,
perbedaan
dan intrapair
tingkat
metilasi6
C (
= D
7/23/2019 Translate EPM
http://slidepdf.com/reader/full/translate-epm 15/20
metilasi
dihitung
sebagai
h %1) / h %1)
G h %T)
di mana h
* puncak
tinggi pada
2E1
.pearman
korelasi
koe!sien sien
dihitung
untuk
mengu4i
untuk
asosiasi
dari intrapair
B,
dan ,
perbedaan
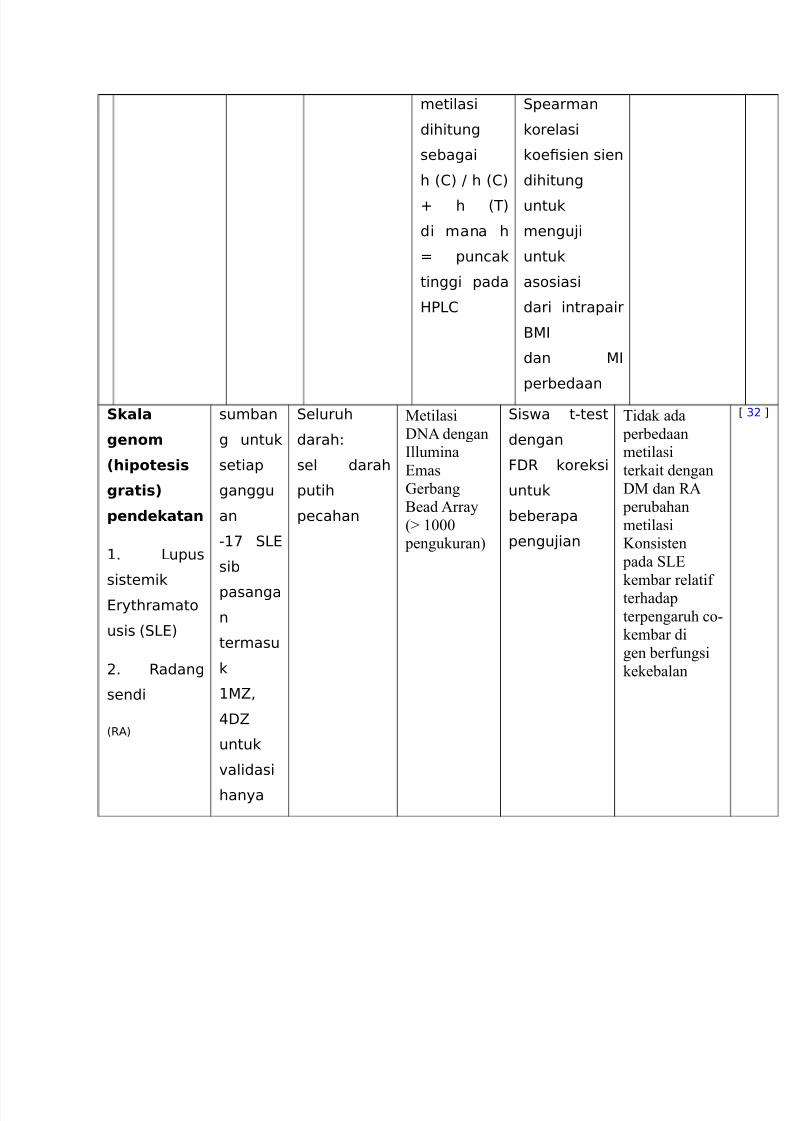
Skala
genom
(hipotesis
gratis)
pendekatan
+6 upus
sistemik
@rythramato
usis %.@)
96 Radang
sendi
%RA)
sumban
g untuk
setiap
ganggu
an
-+' .@
sib
pasanga
n
termasu
k
+5<
(:5
untuk
"alidasi
hanya
.eluruh
darah8
sel darah
putih
pecahan
Metilasi
D01 dengan
?llumina
/mas
Aerbang
5ead 1rray
% ---
pengukuran&
.iswa t-test
dengan
F:R koreksi
untuk
beberapa
pengu4ian
idak ada
perbedaan
metilasi
terkait dengan
DM dan B1
perubahan
metilasi
Konsisten
pada O/
kembar relatif
terhadap
terpengaruh #o(
kembar di
gen berfungsi
kekebalan
C &9 D

7/23/2019 Translate EPM
http://slidepdf.com/reader/full/translate-epm 16/20
&6
:ermatomio
sitis %:)
"alidasi oleh
bisul!t
Eengurutan
pyroseMuenc
ing global
0e1
pengukuran
Eenurunanglobal dalam0-methylcytosine
pada .@ danhypomethylationdari 9# . dan+# gen.r:NA
Table 10.2 %continued)
Caktor ranskripsi terlibat dalam pembentukan A151ergi# interneuron [3)!.
2 S 4B non protein #oding 2 S "ilayah diterjemahkan dari B01 transkrip
DMB daerah berbeda(beda alkohol
Korelasi intra ?@@
DBD) reseptor dopaminergik D)
@6M #ate#hol(6(methyltransferase
BB5 mengurangi representasi bisulphite seGuen#ing
MBD1 metilasi sensitif analisis perbedaan representasional A01P [3-!
PP?/O Peptidylprolyl isomerise /(seperti spermine M synthase
skala genom metilasi D01 profiling dari lebih dari 3-- situs @pA di lima pasang
pasang MZ sumbang, dalam kombinasi dengan gen 'alidasi spesifik di lanjut sumbang
kembar dan subyek kontrol yang #o#ok, dan pengukuran global 3(Methyl#ytosine
tingkat. ebuah t(test digunakan untuk mengidentifikasi probe array yang termetilasi
berbeda(beda antara indi'idu yang terkena dan kembar yang sehat masing(masing. P(nilai
yang dihasilkan adalah dikoreksi untuk beberapa pengujian menggunakan Calse ingkat
Penemuan metodologi [22!. eperti kebanyakan studi metilasi saat di alam ini, perubahan
metilasi berarti se"enang("enang antara kelompok yang terkena dampak dan non(terkena
digunakan sebagai #ut(off %dalam hal ini -H&
untuk menghasilkan daftar kandidat gen untuk 'alidasi berikutnya.
Pendekatan ini mengungkapkan < gen, diperkaya untuk fungsi kekebalan tubuh,
menunjukkan Perubahan konsisten dalam metilasi D01 @ally spesifik pada O/. idak ada
metilasi konsisten perubahan yang ditemukan di B1 atau DM. 7alidasi berikutnya dalam
kelompok kontrol dan selanjutnya menetapkan pasangan kembar sumbang %lagi dengan

7/23/2019 Translate EPM
http://slidepdf.com/reader/full/translate-epm 17/20
koreksi untuk beberapa pengujian&
kerahasiaan rmed hasil array. Menariknya, pemeriksaan kadar 3Me@ global yang
identifi ed kerugian umum metilasi D01 dalam O/ mempengaruhi indi'idu,
dan ini ditemukan %setidaknya sebagian& karena # penurunan spesifik di metilasi
di )8 berulang(ulang dan 8 gen B01 ribosom.
10...2 e"olu"i Tinggi 1-20 Situ" di 10-03 dari Semua Gen4
1nalisis yang paling menarik dari faktor yang mengatur keseluruhan profi le metilasi
adalah diperoleh melalui analisis tiga koleksi DZ dan MZ pasang [2<!. ?ni
terdiri dari sel(sel darah putih %leukosit& dari pasangan kembar MZ yang dikorion
%masing(masing kembar memiliki plasenta mereka sendiri& dan )- pasangan kembar DZ
#o#ok untuk usia, jenis kelamin
dan hitung darah, ditambah sel epitel bukal dari - monokorion MZ dan DZ )-
usia dan jenis kelamin(#o#ok. elain itu leukosit dan sel bukal diperoleh dari
masing(masing - dikorion pasangan MZ dan DZ - dari yang lain kohort kembar
independen dan biopsi usus dikumpulkan dari 8 pasangan MZ dari populasi kembar ketiga.
Korelasi intra(kelas untuk MZ dan DZ pasangan dihitung untuk pasangan perbandingan
bijaksana dari lebih dari =.--- titik data indi'idual sesuai dengan tingkat metilasi di
daerah genom indi'idu. ?ni #ompellingly menunjukkan tingkat yang lebih tinggi
kesamaan antara kembar MZ sebagai kelompok, relatif terhadap kelompok DZ %p + ,) T -
()<&. 0amun, dalam studi yang dilakukan se#ara paralel dengan analisis manusia, mulai luas
epigenetik Perbedaan juga ditemukan di strain tikus hampir identik, memimpin penulis
untuk berspekulasi bah"a diferensial Eigotik epigenetik profi le di DZ dibandingkan MZ
kembar adalah pendorong utama dari perbedaan ?@@ ( 'ariabilitas genetik antara tidak
indi'idu yang berbeda [2<!. 0amun, ini tidak dapat langsung diuji karena
ketidakmampuan untuk langsung mengukur epigenetik profi le pada Eigot bersel tunggal.
10... e"olu"i 5ltra-ig/ Setiap Situ" dan "etiap Gene4
Masa depan kedua analisis genom dan epigenomi# kembar terletak pada penggunaan
throughput tinggi nukleotida urutan. 5aranEini dkk. [23! men#ari genetik, ekspresi dan
metilasi D01 perbedaan sel purifi ed @D< di ke#il
jumlah pasangan kembar MZ sumbang untuk multiple s#lerosis %M&. 1ntara 3- dan =8
juta seGuen#ing berbeda memba#a B01 utusan dihitung untuk setiap pasangan.
e#ara garis besar, ini merupakan ungkapan :output: dari sampel asli.
emakin memba#a hadir untuk gen # spesifik sebagai proporsi dari total jumlah memba#a,
yang lebih tinggi menyatakan itu adalah dalam populasi sel mulai. 1nehnya, sebuah

7/23/2019 Translate EPM
http://slidepdf.com/reader/full/translate-epm 18/20
diagnosis M hanya menyumbang ,<H dari total 'arians* 3,2H adalah disebabkan
untuk antara perbedaan pasangan, dan )=,2H karena 'ariasi sehari(hari dalam satu
indi'idu. Data ini saja menyoroti sifat yang sangat dinamis ekspresi gen
yang menimbulkan tantangan yang #ukup besar ketika men#oba untuk mengidentifikasi
penyakit(spesifik gen #
perubahan ekspresi. idak mengherankan, ada perbedaan ekspresi gen yang kuat bisa
dianggap berasal fenotip M sendiri. Meskipun beberapa bukti untuk ketidakseimbangan alel
%di mana salah satu indi'idu dua salinan gen dinyatakan lebih tinggi daripada
lain& dalam pasangan kembar sumbang, tidak ada perubahan seperti itu umum untuk semua
tiga M
pasangan kembar sumbang diperiksa. Pemeriksaan 3-(-------, berkualitas tinggi
berbunyi berkurang representasi seGuen#ing bisulfit %BB5& dari sel @D<
2 tiga kembar sumbang sama menunjukkan sedikit bukti metilasi D01
perubahan yang bisa @ally spesifik terkait dengan fenotip M. edangkan beberapa
ratus perbedaan yang jelas %di spesifik # situs @pA& yang jelas antara laki(laki
dan perempuan, atau antara jaringan kanker primer dan tidak berhubungan, besarnya
perbedaan epigenetik antara MZ kembar sumbang untuk M setidaknya urutan
besarnya lebih rendah dari perbedaan indi'idu non(terkait, dan L 2 lipat
lebih rendah dari antara jaringan primer dan ganas. ayangnya tidak ada
'alidasi independen yang diamati dalam perbedaan pasangan menggunakan alternatif
metodologi, sehingga ketahanan dari beberapa perubahan metilasi D01 yang diamati
dalam pasangan MZ masih belum jelas [23!.
10.+ &eringatan dan &er/atian
. Mungkin ada efek epigenetik spesifik # kembar yang membatasi utilitas mereka sebagai
model
untuk mengungkapkan kontributor penyebab penyakit yang lebih luas yang berlaku di lebih
luas
populasi tunggal. ebagai #ontoh, proses yang mengarah ke MZ kembar tetap
akan ditentukan tetapi se#ara tradisional telah dianggap hasil dari random
proses. 0amun, data yang lebih baru menunjukkan bah"a faktor epigenetik
sendiri mungkin terlibat dalam pemisahan embrio yang mengarah ke MZ kembar
[2=!. Jika spesifik # untuk MZ kembar, ini akan memiliki efek menga#aukan setiap MZ
perbandingan 's DZ bertujuan untuk mengidentifikasi faktor di"ariskan. $al ini dapat

7/23/2019 Translate EPM
http://slidepdf.com/reader/full/translate-epm 19/20
mengakibatkan
diferensial epigenomes dua baru dibuat embrio MZ se#ara independen dari
onset kemudian kejanggalan fenotipik.
). 5eberapa penelitian telah berusaha untuk memperkirakan kekuatan statistik yangdiperlukan untuk memalsukan
hipotesis epigenetik. $ubungan antara jumlah ulangan teknis
%langkah(langkah independen dari biospe#imen yang sama, yang bertujuan untuk
mengidentifikasi
kontribusi teknis :kebisingan:& dan kekuatan untuk mendeteksi biologis yang bermaknaEerbedaan metilasi :NA di pasangan kembar telah elegan diperiksa oleh
Kaminsky et al6 C&'D6 eperti yang diperkirakan jumlah yang lebih besar dari berbagai
teknik replikasi
$asil di tingkat keseluruhan lebih ke#il dari 'arian teknis di array data indi'idu
poin. Dengan demikian, sedikitnya dua ulangan teknis yang diberikan 8-H kekuatan untuk
mendeteksi
,) kali lipat perubahan pada L -H dari probe array dalam hanya sembilan pasangan
kembar. ?ni meningkat
dari L =-H dari probe dengan hanya mereplikasi teknis tunggal. Perhitungan daya lebih lanjut
meramalkan bah"a dengan dua ulangan teknis dan ), < dan = pasangan kembar,
ada peluang 8-H untuk mendeteksi benar metilasi D01 perbedaan dari ,3,,) dan ,= kali lipat, di 3H dari titik data diuji, antara kelompok [2!. atu peringatan
perhitungan ini adalah potensi 'ariasi dalam metilasi D01 dari "aktu ke "aktu [8, !.
?ni akan memiliki efek meningkatkan 'arians populasi di tingkat metilasi
pada usia(sensitif lokus. 4ntuk menangani hal ini akan diinginkan untuk menggunakan usia
#o#ok kembar pasangan mana mungkin untuk kation identifi biomarker epigenetik
@ally spesifik terkait dengan kejanggalan fenotipik. Meskipun potensi masalah ini,
upaya terbatas pada perhitungan daya sejauh dilakukan adalah untuk mendorong
penerapan analisis genom skala metilasi studi berbasis kembar besar.
ulit untuk per#aya bah"a lebih dari - tahun yang lalu genom manusia belum
ditandai sepenuhnya, B01 mikro dan B01 non(#oding lainnya dianggap
sebagian besar artefak oleh beberapa, dan epigenetik dianggap penting :terbatas:
biologis. epuluh tahun dari sekarang ada kemungkinan bah"a karakterisasi penuh dari
kedua
data genom dan epigenomi# akan berada dalam jangkauan kebanyakan peneliti* dalamkasus data epigenomi#, ini akan menjadi bergantung pada penyediaan segala ma#am

7/23/2019 Translate EPM
http://slidepdf.com/reader/full/translate-epm 20/20
jaringan dari donor yang bersedia. Dalam banyak kasus %seperti jaringan saraf di
neuropsikiatri
gangguan&, ini akan perlu dilakukan post mortem. angat mungkin bah"a tingkat
kompleksitas
dan 'ariasi mengungkapkan dalam diri indi'idu sel 9 jaringan 9 organ dari setiap indi'idu,
dan antara indi'idu akan sangat besar, sangat menghambat kation identifi dari
biologis perbedaan yang berarti dalam profi le epigenetik. Meskipun demikian, melekat
kapasitas untuk mengendalikan 'ariasi genetik dalam pasangan kembar MZ %dan pada tingkat
lebih rendah
faktor lingkungan& harus membuktikan sangat berharga dalam membantu untuk mengungkap
pikiran membingungkan kompleksitas gen lingkungan interaksi epigenetik #enderung
berkontribusi terhadap kesehatan manusia dan penyakit.
Top Related