






Languages
Pages
Legal


MARIANA FEDERICA SÁNCHEZ DE BOCK
EFEITOS DA EXPOSIÇÃO DE TILÁPIAS (Oreochromis niloticus) A SAXITOXINAS
PRODUZIDAS POR Cylindrospermopsis raciborskii (CYANOBACTERIA)
RECIFE
2014

ii
MARIANA FEDERICA SÁNCHEZ DE BOCK
EFEITOS DA EXPOSIÇÃO DE TILÁPIAS (Oreochromis niloticus) A SAXITOXINAS
PRODUZIDAS POR Cylindrospermopsis raciborskii (CYANOBACTERIA)
ORIENTADOR: PROF. DR. RENATO JOSÉ REIS MOLICA
CO-ORIENTADOR: PROF. DR. ÁLVARO JOSÉ DE ALMEIDA BICUDO
RECIFE
2014
Dissertação de Mestrado apresentada ao Programa
de Pós-Graduação em Ecologia - PPGE, da
Universidade Federal Rural de Pernambuco, como
requisito final para a obtenção do título de mestre
em Ecologia.

Ficha Catalográfica
S211e Sánchez De Bock, Mariana Federica Efeitos da exposição de tilápias (Oreochromis niloticus) a saxitoxinas produzidas por Cylindrospermopsis raciborskii (CYANOBACTERIA) / Mariana Federica Sánchez De Bock. – Recife, 2014. 68 f.: il. Orientador (a): Renato José Reis Molica. Dissertação (Programa de Pós-Graduação em Ecologia) – Universidade Federal Rural de Pernambuco, Departamento de Biologia, Recife, 2014. Inclui anexo(s) e referências. 1. Eutrofização 2. Floração 3. Cianotoxinas 4. Piscicultura 5. Desempenho de peixes I. Molica, Renato José Reis, orientador II. Título CDD 574.5

iii
EFEITOS DA EXPOSIÇÃO DE TILÁPIAS (Oreochromis niloticus) A SAXITOXINAS
PRODUZIDAS POR Cylindrospermopsis raciborskii (CYANOBACTERIA)
MARIANA FEDERICA SÁNCHEZ DE BOCK
Dissertação apresentada ao Programa de Pós-Graduação em Ecologia (PPGE), da
Universidade Federal Rural de Pernambuco, como parte dos requisitos para obtenção do título
de Mestre em Ecologia. Dissertação aprovada pela banca examinadora:
Orientador:
______________________________
Prof. Dr. Renato José Reis Molica
Titular / UFRPE
Examinadores:
______________________________
Profª. Drª. Sandra Maria Feliciano de Oliveira e Azevedo
Titular / UFRJ
______________________________
Prof. Dr. Ronaldo Olivera Cavalli
Titular / UFRPE
_________________________________
Profª. Drª. Ariadne do Nascimento Moura
Titular / UFRPE
________________________________
Profª. Drª. Ana Carla Asfora El-Deir
Suplente / UFRPE
Data de aprovação: / / 2014
RECIFE, 2014

iv
“A BUSCA DE NOVOS CAMINHOS NOS LEVA DE VOLTA A NÓS MESMOS E ÀS DIMENSÕES
ESCONDIDAS DE UMA CONSCIÊNCIA MAIS ELEVADA” KEN O´DONNELL.

v
AGRADECIMENTOS
Agradeço a todos e cada um dos momentos e das pessoas que fizeram parte dessa
caminhada. Agradeço imensamente a minha família: Alicia, Juan, Euge e Elbita, que são
minha base e que desde longe e virtualmente foram um grande apoio. A Rodrigo, pela
motivação, força e guia, e por ser a razão de me aventurar por esses novos caminhos. A Beta e
Elias por todo o apoio desde sempre.
A meu orientador Prof. Dr. Renato Molica, por sua confiança em mim desde o início,
pelo apoio através do conhecimento e da experiência pessoal, pelo seu jeito mediante o qual
descobri potencialidades desconhecidas em mim. Pelas viagens a Recife junto com Carol.
Ao meu co-orientador Prof. Dr. Álvaro José de Almeida Bicudo, pela orientação no
trabalho com peixes, os conselhos e suporte para as análises dos dados. Ao Prof. Dr. Kleber
Régis Santoro pela sua paciência e dedicação para descobrirmos informações valiosíssimas
nos dados, mediante as análises estatísticas.
A toda a equipe do CENLAG da Unidade Acadêmica de Garanhuns: Gustavo e
Gláucia por seus importantíssimos ensinamentos; a Rômullo pelo compromisso, carinho, bom
humor e apoio incondicional. A Hugo e Samantha pelos conselhos, companhia e apoio. A
Camila, por me acolher em Garanhuns. A Phillipe, Timóteo, Saulo, Bruno, Vânia e Erivaldo,
pela ajuda imensurável. A Erick, Sheylla, Keila, Patricia, Wando e Marquinho, com quem
compartilhei muitos momentos felizes, e fizeram minha estadia em Garanhuns mais divertida.
A Alana e Carol, que foram uma benção no final do caminho.
A minhas amigas de turma Fernanda, Vanessa, Flávia, Gabriela e Veruska, com quem
a experiência foi mais divertida e compartilhada! Aos professores das disciplinas, que me
aportaram tanto conhecimento e paixão pela ecologia: Professores Geraldo, Raquel, Renato,
Paula, Ariadne, Paulo, Betânia, Elcida e Ana Carla.
A minhas amigas da vida Rocío, Cony, Irene, Vir e Caro, que desde longe me
acompanham sempre. A Basílio, Brena e Cleide, pelo abrigo no momento que tanto precisava,
pela companhia e pelos grandes ensinamentos.
A UFRPE pelo apoio para a participação num congresso, ao Ministério de Relações
Exteriores (MRE) e ao CNPq pela bolsa de mestrado concedida e a FACEPE pelo
financiamento do projeto.

vi
SUMÁRIO
LISTA DE FIGURAS .............................................................................................................. vii
LISTA DE TABELAS ............................................................................................................ viii
RESUMO .................................................................................................................................. ix
ABSTRACT ............................................................................................................................... x
INTRODUÇÃO GERAL .......................................................................................................... 11
REVISÃO BIBLIOGRÁFICA ................................................................................................. 12
REFERÊNCIAS BIBLIOGRÁFICAS ..................................................................................... 18
MANUSCRITO ........................................................................................................................ 23
Resumo ................................................................................................................................... 24
1. Introdução.......................................................................................................................... 25
2. Material e métodos ............................................................................................................ 28
2.1 Experimento ................................................................................................................... 28
2.1.1 Cianobactérias ........................................................................................................... 29
2.1.2 Peixes ......................................................................................................................... 30
2.2 Análises químicas de composição corporal .................................................................. 31
2.3 Análises de saxitoxinas .................................................................................................. 32
2.4 Análises estatísticas ....................................................................................................... 33
3. Resultados e Discussão ..................................................................................................... 34
4. Conclusão .......................................................................................................................... 48
5. Referências bibliográficas ................................................................................................. 50
ANEXO .................................................................................................................................... 56

vii
LISTA DE FIGURAS
MANUSCRITO
Figura 1 – Variações na concentração de saxitoxinas por volume (STX, μg STXequiv.L-1
),
concentração total de saxitoxinas disponível por biomassa de peixe (μg STXequiv. L-1
.g-1
) e
densidade celular de Cylindrospermopsis raciborskii (células.L-1
) no tratamento
Cianobactérias durante o período experimental.
Figura 2 – Sobrevivência acumulada (%) de juvenis de tilápia do Nilo (Oreochromis
niloticus) expostos aos diferentes meios de cultivo.
Figura 3 – Estimativa da influência da mortalidade sobre o peso médio de juvenis de tilápia
do Nilo (Oreochromis niloticus) cultivados nos diferentes meios de cultivo.
Figura 4 – Estimativa da influência da mortalidade sobre o índice de eficiência alimentar de
juvenis de tilápia do Nilo (Oreochromis niloticus) cultivados nos diferentes meios de cultivo.
Figura 5 – Variação no período experimental da concentração de exposição total de
saxitoxinas por biomassa de peixe (μg STXequiv.L-1
g-1
), peso médio diário por peixe (g) e
número de mortes de juvenis de tilápia do Nilo (Oreochromis niloticus) no tratamento
Cianobactérias.
Figura 6 – Estimativa da influência do peso (peso médio estimado por peixe por dia, PM, g)
sobre a mortalidade de juvenis de tilápia do Nilo (Oreochromis niloticus) cultivados no meio
de cultivo com Cylindrospermopsis raciborskii (MA, número de peixes mortos por aquário).
Figura 7 – Estimativa da influência da concentração de exposição total de saxitoxinas por
biomassa de peixe (STXT, μg STX.L-1
.g-1
) sobre a mortalidade de juvenis de tilápia do Nilo
(Oreochromis niloticus) cultivados no meio de cultivo com C. raciborskii (MA, número de
peixes mortos por aquário)..

viii
LISTA DE TABELAS
MANUSCRITO
Tabela 1 – Variáveis de qualidade de água (média ± desvio padrão) durante o período
experimental.
Tabela 2. Registros de mortalidade (nº de peixes) de juvenis de tilápia do Nilo (Oreochromis
niloticus) em cada tratamento durante o período experimental.
Tabela 3. Variáveis de desempenho (média ± desvio padrão) dos juvenis de tilápia do Nilo
(Oreochromis niloticus) expostos aos diferentes meios de cultivo.
Tabela 4. Composição química corporal e taxa de retenção proteica aparente (TRP) (valores
médios ± desvio padrão) de juvenis de tilápia do Nilo (Oreochromis niloticus).

ix
Sánchez De Bock, Mariana Federica; Ma; Universidade Federal Rural de Pernambuco;
Fevereiro de 2014; Efeitos da exposição de tilápias (Oreochromis niloticus) a saxitoxinas
produzidas por Cylindrospermopsis raciborskii (Cyanobacteria); Renato José Reis Molica
(Orientador); Álvaro José de Almeida Bicudo (Co-orientador); Kleber Régis Santoro
(Colaborador científico).
RESUMO
Florações de cianobactérias têm aumentado sua frequência de ocorrência e duração no mundo
inteiro, devido ao aumento do estado trófico dos ecossistemas aquáticos continentais,
inclusive em reservatórios e viveiros utilizados na aquicultura. Várias espécies de
cianobactérias são potenciais produtoras de metabólitos secundários tóxicos (cianotoxinas),
que podem afetar os organismos aquáticos expostos a elas. A influência da exposição crônica
de uma cepa de Cylindrospermopsis raciborskii produtora de saxitoxinas no desempenho de
juvenis de tilápia do Nilo (Oreochromis niloticus) foi avaliada mediante um experimento de
60 dias com três condições de cultivo diferentes: (i) C. raciborskii produtora de saxitoxinas,
em meio de cultura ASM-1; (ii) meio de cultura ASM-1 (controle), e (iii) água desclorada
(controle). A sobrevivência, o peso médio, ganho de peso, índice de eficiência alimentar,
entre outros, foram analisados. Peixes expostos a C. raciborskii apresentaram uma maior
mortalidade e menor crescimento em comparação aos peixes dos tratamentos controle.
Entretanto, o peso dos peixes teve influência na toxicidade, indicando uma resposta dose-
dependente da mortalidade. Os resultados demonstram efeitos prejudiciais das saxitoxinas em
juvenis de tilápia do Nilo, e uma vulnerabilidade em função do tamanho dos peixes e da
concentração de saxitoxinas presentes no meio de cultivo. Reforça-se, a partir da análise dos
resultados, a importância de manter a qualidade da água nos ecossistemas aquáticos. Em
estações de piscicultura isto é particularmente importante, a fim de reduzir riscos de florações
de cianobactérias produtoras de saxitoxinas, evitando, portanto, perdas econômicas aos
produtores.
Palavras-chave: eutrofização, floração, cianotoxinas, piscicultura, desempenho de peixes.

x
Sánchez De Bock, Mariana Federica; M.Sc.; Federal Rural University of Pernambuco;
February 2014; Effects of exposure of tilapia (Oreochromis niloticus) to saxitoxins
(Oreochromis niloticus) produced by Cylindrospermopsis raciborskii (Cyanobacteria);
Renato José Reis Molica (Supervisor); Álvaro José De Almeida Bicudo (Co-supervisor),
Kleber Régis Santoro (Scientific Advisor).
ABSTRACT
Cyanobacterial blooms have increased their frequency of occurrence and duration worldwide,
due to increasing trophic state of freshwater ecosystems, including reservoirs and ponds used
for aquaculture. Several species of cyanobacteria are potential producers of toxic secondary
metabolites (cyanotoxins), which can affect aquatic organisms. The influence of chronic
exposure to saxitoxins-producing strain of Cylindrospermopsis raciborskii on the
performance of juvenile Nile tilapia (O. niloticus) was evaluated by an 60-day experiment
consisting in three different culture conditions: (i) saxitoxins-producing strain C. raciborskii
in ASM-1 culture medium; (ii) culture medium ASM-1 (control), and (iii) dechlorinated water
(control). Survivorship, average weight, weight gain, feed efficiency ratio, among others,
were analyzed. Fish exposed to C. raciborskii showed a higher mortality and lower growth
compared to fish from control treatments. However, fish weight influenced toxicity,
suggesting that mortality was dose-dependent. Results demonstrate harmful effects of
saxitoxins in juvenile Nile tilapia, and a that their vulnerability depends on fish size and
saxitoxins concentration in culture medium. From these results, the importance of maintaining
the water quality in aquatic ecosystems is reinforced. In fish farming, this is particularly
important to reduce risks of saxitoxins-producing cyanobacteria blooms, thus avoiding
economic losses for producers.
Keywords: eutrophization, bloom, cyanotoxins, fish farming, fish performance.

11
INTRODUÇÃO GERAL
Em reservatórios de regiões tropicais, como o Nordeste brasileiro, a ocorrência de
florações de cianobactérias é constante (MOLICA et al., 2005; MOURA et al., 2010), pois
nestas regiões predominam corpos d’ água com longos tempos de retenção e crescente estado
trófico (MEREL et al., 2013). Em adição, as mudanças climáticas, particularmente o
aquecimento global, o aumento da estratificação, a salinização e a intensificação de tormentas
e secas interacionam com o aumento de nutrientes no ambiente, modulando também a
frequência, intensidade, distribuição e duração das florações (PAERL et al., 2011).
Mundialmente, a frequência das florações de cianobactérias tem aumentado em razão
do enriquecimento de nutrientes nos ambientes aquáticos por atividades humanas (CHORUS;
BARTRAM, 1999). A piscicultura é uma atividade em expansão, em razão do seu potencial
de melhorar a qualidade de vida de produtores e oferecer fontes de proteína de pescado, em
vista da incapacidade de atender a demanda apenas com pescado proveniente do extrativismo
(pesca). A tilápia do Nilo (Oreochromis niloticus) (Linnaeus, 1758) é uma das espécies de
peixe de água doce mais cultivada no mundo (FAO, 2012), e é a principal espécie de peixe de
água doce cultivada no Brasil (BRASIL, 2013) - apesar de ser uma espécie exótica e
fortemente invasora - em razão do seu desempenho produtivo. Essa atividade produtiva,
entretanto, pode contribuir para a eutrofização dos ecossistemas aquáticos onde opera
(CYRINO et al., 2010), sendo, em muitos casos, reservatórios de usos múltiplos.
Algumas espécies de cianobactérias podem produzir potentes toxinas (cianotoxinas)
(VAN APELDOORN et al., 2007). Florações de cianobactérias tóxicas podem causar efeitos
nocivos em organismos aquáticos (CARMICHAEL,1992; CODD et al., 2005). Mortalidades
massivas de peixes em ecossistemas aquáticos continentais naturais foram relacionadas a
florações de cianobactérias tóxicas (ERIKSSON et al., 1986; RODGER et al., 1994;
AZEVEDO; CARMOUSE, 1994; CHELLAPPA et al., 2008). Em cativeiro, relatos similares
foram registrados em cultivos de perca (Perca sp) Linnaeus, 1758, bagre do canal (Ictalurus
punctatus) (Rafinesque, 1818), lúcio (Esox sp.) Linnaeus, 1758 e particularmente em carpas
(Cyprinus carpio) Linnaeus, 1758 (SEVRIN-REYSSAC; PLETIKOSIC, 1990), causando
sérios prejuízos econômicos.
A mortalidade de peixes relacionada à ocorrência de florações de cianobactérias pode
ser causada pela liberação de cianotoxinas em altas concentrações. Pode também ser induzida
pelas condições ambientais decorrentes da decomposição destas florações, entre elas: alta

12
temperatura e baixo nível de água, alto pH, altas concentrações de amônia, baixo oxigênio
dissolvido ou pela combinação de todos os fatores (JEWEL et al., 2003; KANGUR et al.,
2005).
Muitos estudos, que relatam efeitos nocivos de cianotoxinas em peixes, utilizaram vias
não naturais de exposição daqueles às cianotoxinas, como injeção intraperitoneal (LI et al.,
2005; SILVA DE ASSIS et al., 2013) e ingestão forçada (ZIMBA et al., 2001, SOARES et
al., 2004). Entretanto, há pouca informação da influência das cianotoxinas em peixes pela
exposição direta, ou seja, do contato dos peixes com células de cianobactérias na água, em
densidades similares às que ocorrem no ambiente durante as florações.
A cianobactéria Cylindrospermopsis raciborskii (Wolosz.) Seena. e Subbar., em razão
de sua grande distribuição em diferentes ecossistemas aquáticos brasileiros e dominância em
florações (BOUVY et al., 2000; SANT'ANNA; AZEVEDO, 2000), destaca-se entre as
cianobactérias produtoras das cianotoxinas saxitoxinas no Brasil (LAGOS et al., 1999;
MOLICA et al., 2002; POMATI et al., 2003).
Em função da importância de aprofundar o entendimento dos efeitos das cianotoxinas
em peixes, este estudo avaliou o efeito da exposição direta de juvenis de tilápia do Nilo
(Oreochromis niloticus) a uma cepa tóxica da cianobactéria Cylindrospermopsis raciborskii.
REVISÃO BIBLIOGRÁFICA
As cianobactérias são micro-organismos procariotos fotossintetizantes que evoluíram e
adaptaram-se a praticamente todos os ecossistemas aquáticos do planeta. São mais
encontradas na zona eufótica do plâncton de ambientes marinhos, salobros e de água doce,
desempenhando um papel ecológico importante na produtividade primária, fixação de
nitrogênio e reciclagem de matéria orgânica (HOWARTH et al., 1988).
Esses micro-organismos possuem mecanismos para tolerar a incidência de raios
ultravioletas, concentrações elevadas de metais pesados, baixas concentrações de oxigênio,
baixas e altas temperaturas, o que contribui para sua ampla ocorrência (WHITTON; POTTS,
2000). Algumas espécies podem formar acinetos, células diferenciadas que funcionam como
esporos de resistência e sobrevivem a diversas situações desfavoráveis. Adicionalmente, a
pigmentação de cianobactérias as protege de processos de fotoinibição causados por altas

13
intensidades de luz e, ao mesmo tempo, também melhora a captação de energia luminosa em
uma região mais larga do espectro visível em comparação com outras espécies de fitoplâncton
(MUR et al., 1999). Essas várias características fisiológicas, vantajosas em relação a outros
componentes do fitoplâncton, permitem explicar porque muitas espécies de cianobactérias
tornam-se dominantes e formadoras de florações em diferentes ecossistemas aquáticos em
todo o mundo (OLIVER; GANF, 2000).
As florações de cianobactérias são o resultado do crescimento exagerado destes
organismos, causando impacto negativo na qualidade da água, tornando os ambientes
impróprios para a pesca e atividades recreativas (CARMICHAEL; FALCONER, 1993). Não
existe uma definição ou quantificação internacional a partir da qual se considera uma
floração. Entretanto, este fenômeno é geralmente considerado como uma significativa
produção de biomassa em um curto período de tempo, associado a uma diminuição da
diversidade do fitoplâncton. Muitas vezes as florações são mono-específicas e podem formar
densas camadas de células na superfície da água, sendo visíveis a olho nu (MEREL et al.,
2013).
Muitas espécies de cianobactérias têm sido reportadas como produtoras de metabólitos
secundários - cianotoxinas - tóxicos para diferentes organismos, tais como microalgas,
zooplâncton, plantas aquáticas e terrestres, insetos terrestres, peixes, aves e mamíferos,
incluído o ser humano (CODD et al., 1995; CODD, 2000; BRIAND et al., 2003; DE
FIGUEIREIDO et al., 2004).
A produção de cianotoxinas ocorre em uns 40 de aproximadamente 150 gêneros de
cianobactérias conhecidos no mundo (VAN APELDOORN et al., 2007). Segundo Molica e
Azevedo (2009), apesar de muitos estudos terem sido realizados, a função ecológica desses
metabolitos secundários para as cianobactérias ainda não são claros. Entretanto, são bem
conhecidos os efeitos das toxinas, sendo classificadas em duas classes principais, de acordo
com seu mecanismo de toxicidade em animais: hepatotoxinas e neurotoxinas. As
hepatotoxinas são identificadas como peptídeos cíclicos contendo cinco (nodularinas) ou sete
aminoácidos (microcistinas), ou alcalóides (cilindrospermopsina) (CHORUS; BARTRAM,
1999). A nodularina é produzida por uma única espécie, encontrada em ambientes marinhos e
salobros, Nodularia spumigena Mertens, 1888 Ex Bornet e Flahaul, enquanto que os mais de
80 análogos de microcistinas identificados (FEURSTEIN et al., 2009) podem ser produzidos
por vários gêneros planctônicos e bentônicos de água doce (FERRÃO-FILHO, 2009). A
cilindrospermopsina é produzida também por vários gêneros de água doce, a saber:

14
Cylindrospermopsis, Aphanizomenon, Umezakia, Raphidiopsis e Anabaena (SIVONEN;
JONES, 1999; MOLICA; AZEVEDO, 2009).
A toxicidade aguda das microcistinas, que envolve disfunções hepáticas agudas com
hemorragia e morte de mamíferos em poucas horas, pode ser explicada pelo mecanismo de
inibição das proteínas fosfatases 1 e 2A (MACKINTOSH et al., 1990). Isto leva a um excesso
de fosforilação de proteínas e a alterações no citoesqueleto, perda da forma dos hepatócitos e
subsequente destruição destas causando hemorragia ou insuficiência hepática (VAN
APELDOORN et al., 2007). As microcistinas também causam estresse oxidativo, podendo
desencadear processos apoptóticos nas células (PRIETO et al., 2009). Os efeitos causados
pelas microcistinas estão mais relacionados à exposição crônica, através do consumo de águas
contaminadas e alimentos (produtos agrícolas, peixes, camarões e moluscos), a exposição
dérmica e inalação (CAMPOS; VASCONCELOS, 2010). O fígado é o órgão mais afetado em
animais, mas a exposição à toxina é susceptível de afetar órgãos como rins e cólon (NOBRE
et al., 1999; BOTHA et al., 2004). As nodularinas são também potentes inibidores de
proteínas fosfatases 1 e 2A (MACKINTOSH et al., 1990), com um mecanismo de ação
similar ao das microcistinas. Em peixes, observou-se a indução de apoptose de linfócitos,
através do caminho de apoptose mitocondrial, destruindo sua resposta imune (ZHANG et al.,
2012). As microcistinas e nodularinas podem causar efeitos de longo prazo em exposições
crônicas, como a promoção de tumores (UENO et al., 1996; HUMPAGE; FALCONER,
2003).
A cilindrospermopsina é um alcalóide guanidínico sulfatado cíclico, que inibe a
glutationa do citocromo P450 e a síntese de proteínas irreversivelmente, causando lesões e
necrose das células no fígado, rins, timo, trato intestinal, músculos, coração e sistema vascular
de vertebrados (TERÃO et al., 1994; FROSCIO et al., 2001).
As neurotoxinas foram classificadas como alcalóides (anatoxina-a), carbamatos
(saxitoxinas) ou organofosforados (anatoxina-a(s)). O mecanismo de ação de cada
neurotoxina é diferente, mas todas atuam sobre o processo de propagação do impulso nervoso.
A anatoxina-a e seus análogos são alcalóides de baixo peso molecular que atuam como
bloqueadores neuromusculares pós-sinápticos, imitando o efeito da acetilcolina ao se ligarem
aos seus receptores na placa motora, causando a despolarização da membrana e impedindo a
transmissão neuromuscular (CARMICHAEL, 1992). A anatoxina-a(s) é um organofosforado
que se liga irreversivelmente à acetilcolinesterase, inibindo-a e impedindo que esta degrade a
acetilcolina (HENRIKSEN et al., 1997). Isto gera um excesso de acetilcolina na junção

15
neuromuscular (sinapses), provocando paralisia dos músculos necessários à respiração e
ocasionando a parada dos batimentos cardíacos.
Em relação às saxitoxinas, são compostos tricíclicos com 57 conhecidos análogos
(WIESE et al., 2010), podendo ser não-sulfatados (saxitoxinas), sulfatados (goniautoxinas) o
duplamente sulfatados (C-toxinas) (VAN APELDOORN et al., 2007). Impedem a propagação
do impulso nervoso pela ligação e bloqueio dos canais de sódio, potássio e cálcio nos axônios
dos neurônios, o que leva a paralisia muscular e consequente morte por parada respiratória
(WIEGAND; PFLUGMACHER, 2005). Entre os sintomas mais comuns de intoxicação em
humanos por saxitoxinas estão: paralisia e dormência progressiva da face e do pescoço,
fraqueza e perda de coordenação motora, baixa progressiva da respiração e, em altas doses,
paralisia e morte (KAO, 1993). Estas toxinas hidrossolúveis podem persistir mais de 90 dias
em água doce (MEREL et al., 2013).
As saxitoxinas são produzidas naturalmente por diversas espécies de dinoflagelados
marinos (LEFEBVRE et al., 2004), e por vários gêneros de cianobactérias em ambientes
aquáticos dulcícolas, como Anabaena, Aphanizomenon, Cylindrospermopsis, Lyngbya e
Planktothrix (WIEGAND; PFLUGMACHER, 2005). Em particular, Cylindrospermopsis
raciborskii vem sendo uma das espécies que desperta maior interesse, pela crescente expansão
de sua distribuição geográfica mundial e potencial de toxicidade (PADISÁK, 1997;
FERRÃO-FILHO, 2009). Nas Américas Central e do Sul já foi registrada em diversos países,
incluindo o Brasil (PADISÁK, 1997; SOUZA et al., 1998; BOUVY et al., 2000; PANOSSO
et al., 2007).
A crescente distribuição geográfica de C. raciborskii em ecossistemas aquáticos do
mundo inteiro pode ser atribuída, entre outros fatores, a sua capacidade de flutuabilidade
(aerótopos), alta tolerância a baixas intensidades luminosas, alta afinidade de captura de
amônio, capacidade de fixar nitrogênio atmosférico e a capacidade de absorver fósforo
mesmo em condições de baixa disponibilidade ambiental (PADISÁK, 1997).
A estacionalidade dos períodos chuvosos e secos no Nordeste brasileiro provoca uma
variação sazonal nas variáveis físico-químicas dos ecossistemas aquáticos, favorecendo as
cianobactérias, entre elas C. raciborskii (SEVRIN-REYSSAC; PLETIKOSIC, 1990;
PADISÁK, 1997). Porém, a eutrofização dos ecossistemas aquáticos é o principal fator que
contribui para que as cianobactérias dominem a comunidade fitoplanctônica e formem
florações de forma cada vez mais frequente (CHORUS; BARTRAM, 1999).

16
Uma das atividades humanas que tem impacto significativo no ambiente aquático é a
aquicultura (CYRINO et al., 2010). Essa atividade foi incentivada na região Nordeste do
Brasil pelo Departamento Nacional de Obras Contra as Secas (DNOCS), através de
introdução de juvenis de tilápia no município de Arcoverde – PE em 1933 (FAO, 1988). Em
1971, foi iniciado um programa oficial de produção de juvenis de tilápia para povoamento dos
reservatórios públicos da região Nordeste pelo DNOCS, como uma atividade econômica
alternativa para populações do semi-árido que estão sujeitas a secas frequentes. A
tilapiacultura firmou-se como atividade empresarial a partir da década de 1980
(FIGUEIREDO JÚNIOR; VALENTE JÚNIOR, 2008).
A tilápia do Nilo Oreochromis niloticus foi escolhida para a introdução no programa
de povoamento do DNOCS por ser uma das espécies mais estudadas e cultivadas no mundo
inteiro. Em razão do seu desempenho produtivo, carne de alta qualidade e com poucas
espinhas, rusticidade e capacidade de tolerar altas densidades de cultivo (SIGNOR et al.,
2010), a tilápia corresponde atualmente à espécie da aquicultura continental mais cultivada
no país, atingindo uma produção de 253.800 toneladas em 2011 (BRASIL, 2013).
O Ministério da Pesca e Aquicultura (MPA) planeja incrementar a produção nacional
de pescado mediante a criação de parques aquícolas continentais entre os milhões de hectares
de lâmina d’água em reservatórios de usinas hidrelétricas (ANEEL) e propriedades
particulares no interior do país (BRASIL, 2012). O. niloticus é uma das principais espécies-
alvo desta política do MPA, devido ao seu alto desempenho produtivo. Entretanto, a
piscicultura precisa ser bem gerenciada e regulada para oferecer uma real oportunidade de
atividade econômica sustentável. Borges et al. (2010) observaram uma alteração na
concentração de nutrientes e na comunidade fito-zooplanctônica de um reservatório no sul do
Brasil em que foram instaladas gaiolas para produção de tilápia do Nilo. As alterações dos
parâmetros da qualidade d´água em estações de cultivo podem ser de maior ou menor impacto
para o ambiente e para própria produção, dependendo do manejo empregado, como por
exemplo, a qualidade da ração fornecida. A otimização do crescimento dos peixes só pode ser
alcançada através do manejo concomitante da qualidade de água, nutrição e alimentação
(CYRINO et al., 2010).
A alimentação excessiva ou o uso de rações não balanceadas reduzem a absorção de
nutrientes pelos peixes, o que pode resultar em excesso de matéria orgânica nos sistemas de
produção. Em condições tropicais, esta matéria orgânica é mineralizada rapidamente
(CYRINO et al., 2010). Nutrientes inorgânicos regenerados neste processo estariam

17
prontamente disponíveis para o florescimento do fitoplâncton, reduzindo a transparência da
água e alterando a sua qualidade (CYRINO et al., 2010). Em ecossistemas aquáticos com
presença de cepas tóxicas de cianobactérias, o risco de intoxicação dos peixes pode acarretar
inclusive maiores perdas econômicas.
Peixes planctófagos têm se mostrado resistentes aos efeitos tóxicos de microcistinas
(XIE et al., 2004) e capazes de acumular toxinas nos seus tecidos. Juvenis de tilápia do Nilo
(Oreochromis niloticus) submetidos a uma dose de 1.000 µg.kg-1
da microcistina-LR,
concentração 20 vezes maior que a DL50 (Dose Letal 50%) para camundongos, apresentaram
apenas efeitos subletais (MOREIRA, 2012). As microcistinas têm sido o principal alvo de
análise em relação aos seus efeitos deletérios em razão da sua maior ocorrência em todo o
mundo (FERRÃO-FILHO, 2009). Muito menos estudos são dedicados às outras toxinas
(FERRÃO-FILHO, 2009; MEREL et al., 2013).
Resultados obtidos em relação aos efeitos das saxitoxinas em embriões, larvas e
juvenis de peixes (ROBINEAU et al., 1991; LEFEBVRE et al., 2004; LEFEBVRE et al.
2005; DA SILVA et al., 2011) demonstraram efeitos prejudiciais similares aos causados pelas
microcistinas, como malformações no desenvolvimento, atraso no tempo de eclosão, dano
genotóxico, perda de coordenação motora, estresse oxidativo e mortalidade. Entretanto, a
maioria desses estudos expuseram os peixes a saxitoxinas purificadas em concentrações
agudas no meio de cultivo, ou via injeção intraperitoneal, lisando previamente as células
concentradas a partir de culturas ou de florações naturais. Embora estas técnicas sejam úteis
no estudo de efeitos patológicos, elas não representam as condições naturais (FERRÃO-
FILHO, 2009).
A rota de exposição tem sido considerada de grande importância na determinação dos
efeitos, uma vez que a exposição a toxinas dissolvidas na água causa efeitos muito menores,
ou, até mesmo, nenhuma mortalidade, quando comparadas às mesmas doses aplicadas
oralmente (FERRÃO-FILHO, 2009). Em condições naturais as toxinas entram nos peixes
principalmente através da ingestão de células e absorção no trato gastrointestinal, e em menor
proporção a absorção pode ocorrer através das brânquias ou epiderme (TENCALLA et al.,
1994). Portanto, espécies de peixes herbívoros como tilápia e carpa são altamente susceptíveis
de serem afetadas pelas cianotoxinas (FERRÃO-FILHO, 2009).
Em razão do exposto acima, o presente trabalho teve por objetivo analisar os
potenciais efeitos de uma cepa da cianobactéria Cylindrospermopsis raciborskii, produtora de

18
saxitoxinas, na sobrevivência e no crescimento de juvenis de tilápia do Nilo (Oreochromis
niloticus). A hipótese de trabalho consistiu a priori que a exposição direta e crônica a uma
cepa de cianobactéria produtora de saxitoxinas influenciaria negativamente a sobrevivência e
o crescimento de juvenis de tilápia.
REFERÊNCIAS BIBLIOGRÁFICAS
AZEVEDO, S.M.F.O.; CARMOUSE, J.P. Une mortalité de poissons dans une lagune
tropicale (Brésil) durant une périod de dominance de Cyanophyceae. Revue d’
Hydrobiologie Tropicale, v.27, p.265-272, 1994.
BORGES, P.A.F. et al. Effects of fish farming on plankton structure in a Brazilian tropical
reservoir. Hydrobiologia, v. 649, p. 279-291, 2010.
BOTHA, N. et al. The effect of intraperitoneally administered microcystin-LR on the
gastrointestinal tract of Balb/c mice. Toxicon, v.43, p.251–254, 2004.
BOUVY, M. et al. Occurrence of Cylindrospermopsis (Cyanobacteria) in 39 Brazilian
tropical reservoirs during the 1998 drought. Aquatic Microbial Ecology, v.23, p. 13-27,
2000.
BRASIL. Ministério da Pesca e a Aquicultura (MPA). Boletim Estatístico da Pesca e
Aquicultura 2010. Brasília: DEMOC, CGMIP, CGCOP, SEPOA, DEFO, DRPA, IBGE
(COAGRO, COMEQ), 2012. 129 p.
BRASIL. Ministério da Pesca e a Aquicultura (MPA). Boletim Estatístico da Pesca e
Aquicultura 2011. Brasília: DEMOC, CGMIP, 2013. 60 p.
BRIAND, J.F. et al. Health hazards for terrestrial vertebrates from toxic cyanobacteria in
surface water ecosystems. Veterinary Research, v. 34, p. 361-377, 2003.
CAMPOS, A.; VASCONCELOS, V. Molecular mechanisms of microcystin toxicity in animal
cells. International Journal of Molecular Sciences, v.11, 268-287, 2010.
CARMICHAEL, W. W. Cyanobacteria secondary metabolites - the cyanotoxins. Journal of
Applied Bacteriology, v.72, p.445-459, 1992.
CARMICHAEL, W. W.; FALCONER, I. R. Disease related to freshwater blue-green algal
toxins, and control measures. In: FALCONER, I. R. Algal toxins in seafood and drinking
water. San Diego, CA: Academic Press, 1993, p. 87-209.
CHELLAPPA, N.T. et al. Harmful phytoplankton blooms and fish mortality in a
eutrophicated reservoir of Northeast Brazil. Brazilian Archives of Biology and Technology,
v. 51, no. 4, p. 833-841, 2008.
CHORUS, I.; BARTRAM, J. Toxic Cyanobacteria in water. A guide to their public health
consequences, monitoring and management. London; New York: E and Fn Spon, 1999. 416 p.

19
CODD, G.A. Cyanobacterial toxins, the perception of water quality, and the priorisation of
eutrophication control. Ecological Engineering, v.16, p.51–60, 2000.
CODD, G.A. et al. Harmful cyanobacteria: from mass mortalities to management measures.
In: HUISMAN, J. et al. Harmful Cyanobacteria. Dordrecht: Springer, 2005, p.1-24.
CODD, G.A. et al. Toxins from cyanobacteria (blue–green algae). IN: WIESSNER, W.;
SCHNEPF, E., STARR, R.C. Algae, environment and human affairs. Bristol, England:
Biopress Ltd., 1995, p. 1–17.
CYRINO, J.E.P. et al. A piscicultura e o ambiente – o uso de alimentos ambientalmente
corretos em piscicultura. Revista Brasileira de Zootecnia, v.39, p. 68-87, 2010.
DA SILVA, C.A. et al. First report about saxitoxins in freshwater fish Hoplias malabaricus
through trophic exposure. Toxicon, v.57, p.141–147, 2011.
DE FIGUEIREDO, D.R. et al. Microcystin-producing blooms— A serious global public
health issue. Ecotoxicology and Environmental Safety, v.59, p.151–163, 2004.
ERIKSSON, J.E. et al. Can Cyanobacterial Toxins Accumulate in Aquatic Food Chains? In:
4th International Symposium of Microbial Ecology, 1986, Ljubljana. Proceeding of the 4th
International Symposium of Microbial Ecolology. 1986. p. 655–658.
FAO. Organização das Nações Unidas para Alimentação e Agricultura. PROJETO
GCP/RLA/075/ITA Apoio as atividades regionais de aquicultura para América Latina e o
Caribe. Manual sobre manejo de reservatórios para a produção de peixes. Roma, 1988.
Disponível em: <http://www.fao.org/docrep/field/003/AB486P/ AB486P00.htm#TOC>.
Acesso em: 10 sept. 2012.
FAO. Organização das Nações Unidas para Alimentação e Agricultura. El estado mundial de
la pesca y la acuicultura 2012. Departamento de Pesca y Acuicultura de la FAO, Roma:
ISSN 1020-5500, 2012.
FERRÃO-FILHO, A.S. Bioacumulação de cianotoxinas e seus efeitos em organismos
aquáticos. Oecologia brasiliensis, v.13, n. 2, p.272-312, 2009.
FEURSTEIN, D.; HOLST, K.; FISCHER, A.; DIETRICH, D.R. Oatp-associated uptake
and toxicity of microcystins in primary murine whole brain cells. Toxicology and Applied
Pharmacoly, v.234, p.247–255, 2009.
FIGUEIREDO JÚNIOR, C.A.; VALENTE JÚNIOR, A.S. XLVI Congresso da Sociedade
Brasileira de Economia, Administração e Sociologia Rural. 2008. Cultivo de tilápias no
Brasil: Origens e cenário atual. Disponível em:<http://www.sober.org.br/
palestra/9/178.pdf>. Acesso em: 5 sept. 2012.
FROSCIO, S.M. et al. Cell-free protein synthesis inhibition assay for the cyanobacterial toxin
cylindrospermopsin. Environmental Toxicology, v.16, p.408-412, 2001.
HENRIKSEN, P. et al. Detection of an anatoxin-a(s)-like anticholinesterase in natural blooms
and cultures of cyanobacteria/blue-green algae from Danish lakes and in the stomach contents
of poisoned birds. Toxicon, v.35, p901-913, 1997.

20
HOWARTH, R.W. et al. Nitrogen-fixation in freshwater, estuarine and marine ecosystems.
Rates and importance. Limnology and Oceanography, v.33, p.669–87, 1988.
HUMPAGE, A.R.; FALCONER, I.R. Oral toxicity of the cyanobacterial toxin
cylindrospermopsin in male swiss albino mice: Determination of no observed adverse
effect level for a drinking water guideline value. Environmental Toxicology, v.18, p. 94-
103, 2003.
JEWEL, M.A.et al. Fish mortality due to cyanobacterial bloom in an aquaculture pond in
Bangladesh. Pakistan Journal of Biological Sciences, v.6, p. 1046–1050, 2003.
KANGUR, K. et al. Fish kill in Lake Peipsi in summer 2002 as a synergistic effect of
a cyanobacterial bloom, high temperature, and low water level. Proceedings of the
Estonian Academy of Sciences: Biology, Ecology, v.54, n.1, p.67–80, 2005.
KAO, C.Y. Paralytic Shellfish Poisoning. In: FALCONER, I.R. Algal toxins in seafood and
drinking water. London: Academic Press, 1993. p. 75-86.
LAGOS, N. et al. The first evidence of paralytic shellfish toxins in the freshwater
cyanobacterium Cylindrospermopsis raciborskii, isolated from Brazil. Toxicon, v.37,
p.1359–1373, 1999.
LEFEBVRE, K.A. et al. Dissolved saxitoxin causes transient inhibition of sensorimotor
function in larval Pacific herring (Clupea harengus pallasi). Marine Biology, v.147, p.1393–
1402, 2005.
LEFEBVRE, K.A. et al. Morphological abnormalities and sensorimotor deficits in larval fish
exposed to dissolved saxitoxin. Aquatic Toxicology, v.66, p. 159–170, 2004.
LI, L. et al. In vivo studies on toxin accumulation in liver and ultrastructural changes of
hepatocytes of the phytoplanktivorous bighead carp i.p.-injected with extracted microcystins.
Toxicon, v. 46, p. 533–545, 2005.
MACKINTOSH, C. et al. Cyanobacterial microcystin-LR is a potent and specific inhibitor of
protein phosphatases 1 and 2A from both mammals and higher plants. FEBS Letters, v. 264,
n.2, p.189-192, 1990.
MEREL, S. et al. State of knowledge and concerns on cyanobacterial blooms and
cyanotoxins. Environment International, v.59, p. 303–327, 2013.
MOLICA, R.; AZEVEDO, S. Ecofisiologia de cianobactérias produtoras de cianotoxinas.
Oecologia Brasiliensis, v. 13, n. 2, p. 229-246, 2009.
MOLICA, R.J.R. et al. Occurrence of saxitoxins and an anatoxin-a(s)-like anticholinesterase
in a Brazilian drinking water supply. Harmful Algae, v.4, p.743–753, 2005.
MOLICA, R.J.R. et al. Toxins in the freshwater cyanobacterium Cylindrospermopsis
raciborskii (Cyanophyceae) isolated from Tabocas reservoir in Caruaru, Brazil, including
demonstration of a new saxitoxin analogue. Phycologia, v.41, p.606-611, 2002.
MOREIRA, D.A. Mecanismos moleculares envolvidos na toxicidade da microscistina-LA
em tilápia (Oreochromis niloticus). Universidade Federal do Rio de Janeiro, Instituto de
Biofísica Carlos Chagas Filho, 2012. 53p.

21
MOURA, A. et al. Reservatórios do nordeste do Brasil: biodiversidade, ecologia, manejo.
Bauru: NUPPEA, 2010. 576p.
MUR, L.R. et al. Cyanobacteria in the Environment. In: CHORUS, I.; BARTRAM, J. Toxic
Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring,
and Management. 1ed. Londres e New York: E&FN Spon, 1999. Cap. 2, p. 15-40.
NOBRE, A.C.L.; JORGE, M.C.M.; MENEZES, D.B.; FONTELES, M.C.; MONTEIRO,
H.S.A. Effects of microcystin-LR in isolated perfused rat kidney. Brazilian Journal Of
Medical and Biological Research, v.32, p.985–988, 1999.
OLIVER, R.L.; GANF, G.G. Freshwater blooms. In: WHITTON, B.A.; POTTS, M. The
Ecology of Cyanobacteria. Dortrecht, The Netherlands: Kluwer Academic Publishers, 2000.
p. 149–194.
PADISÁK, J. Cylindrospermopsis raciborskii (Woloszynnska) Seenayya et Subba Raju, an
expanding, highly adaptive cyanobacterium: worldwide distribution and review of its ecology.
Archiv für Hydrobiology, Stuttgart, v.107, n. 4, p. 563-593, 1997.
PAERL, H.W.; HALL, N.S.; CALANDRINO, E.S. Controlling harmful cyanobacterial
blooms in a world experiencing anthropogenicand climatic-induced change. Science of the
Total Environment, v.409, p.1739–1745, 2011.
PANOSSO, R. et al. Cianobactérias e cianotoxinas em reservatórios do estado do Rio Grande
do Norte e o potencial controle das florações pela tilápia do Nilo (Oreochromis niloticus).
Oecologia Brasiliensis, vol. 11, no. 3, p. 433-449, 2007.
POMATI, F. et al. Enhancement of intracellular saxitoxin accumulation by lidocaine
hydrocloride in the cyanobacterium Cylindrospermopsis raciborskii T3 (Nostocales).
Journal of Phycology, v.39, p.535-542, 2003.
PRIETO, A.I. et al. Time-dependent protective efficacy of Trolox (vitamin E analog)
against microcystin-induced toxicity in tilapia (Oreochromis niloticus). Environmental
Toxicology, v. 24, p.563–579, 2009.
ROBINEAU, B. et al. Comparison of the response of five larval fish species to the toxic
dinoflagellate Alexandrium excavatum (Braarud) Balech. Journal of experimental marine
biology and ecology, v.152, n.2, p. 225 -242, 1991.
RODGER, H.D. et al. Cyanobacterial (blue-green algal) bloom associated pathology in brown
trout (Salmo trutta L.) in Loch Leven, Scotland. Journal of Fish Diseases, v.17, p. 177–181,
1994.
SANT’ANNA, C.L.; AZEVEDO, M.T.P. Contribution to the knowledge of potentially toxic
cyanobacteria from Brazil. Nova Hedwigia, v.71, p. 359-385, 2000.
SEVRIN-REYSSAC, J.; PLETIKOSIC, M. Cyanobacteria in fish ponds. Aquaculture,
Amsterdam, v. 88, p. l-20, 1990.
SIGNOR, A.A. et al. Desempenho de juvenis de tilápia-do-nilo alimentados com rações
contendo complexo enzimático. Revista Brasileira de Zootecnia, v.39, n.5, p.977-983, 2010.

22
SILVA DE ASSIS, H.C. et al. Hematologic and hepatic responses of the freshwater fish
Hoplias malabaricus after saxitoxin exposure. Toxicon, v. 66, p.25–30, 2013.
SIVONEN, K.; JONES, G. Cyanobacterial Toxins. In: CHORUS, I.; BARTRAM, J. Toxic
Cyanobacteria in Water. London: E & FN Spon, 1999. Cap. 3, p.41-111.
SOARES, R.M., MAGALHÃES, V.F., AZEVEDO, S.M.F.O. Accumulation and depuration
of microcystins (cyanobacteria hepatotoxins) in Tilapia rendalli (Cichlidae) under laboratory
conditions. Aquatic Toxicology, v. 70, p. 1-10, 2004.
SOUZA, R.C.R.; CARVALHO, M.C.; TRUZZI, A.C. Cylindrospermopsis raciborskii
(Wolosz.) Seenaya and Subba Raju (Cyanophyceae) dominance and a contribution to the
knowledge of Rio Pequeno Arm, Billings reservoir Brazil. Environmental Toxicology and
Water Quality, v. 13, p. 73–81, 1998.
TENCALLA, F.G. et al. Toxicity of Microcystis aeruginosa peptide toxin to yearling rainbow
trout (Oncorhyncus myskiss). Aquatic Toxicology, v.30, p.215–224, 1994.
TERÃO, K. et al. Electron microscopic studies on experimental poisoning in mice induced by
cylindrospermopsin isolated from blue-green alga Umezakia natans. Toxicon, v.32, n.7,
p.833–843, Jul. 1994.
UENO, Y. et al. Detection of microcystins, a blue-green algal hepatotoxin, in drinking water
sampled in Haimen and Fusui, endemic areas of primary liver cancer in China, by highly
sensitive immunoassay. Carcinogenesis, v.17, p.1317-1321, 1996.
VAN APELDOORN, M.E. et al. Toxins of cyanobacteria. Molecular Nutrition & Food
Research, v. 51, p. 7-60, 2007.
WHITTON, B.A., POTTS, M. The ecology of cyanobacteria, their diversity in time and
space. Dordrecht, London, Boston: Kluwer academic Publishers, 2000. 669p.
WIEGAND, C.; PFLUGMACHER, S. Ecotoxicological effects of selected cyanobacterial
secondary metabolites: a short review. Toxicology and Applied Pharmacology, v.203, p.
201-218, 2005.
WIESE, M., et al. Neurotoxic alkaloids: saxitoxin and its analogs. Marine Drugs 8, 2185–
2211, 2010.
XIE, L.; XIE, P.; OZAWA, K. Dynamics of microcystins-LR and -RR in the
hytoplanktivorous silver carp in a sub-chronic toxicity experiment. Environmental
Pollution, v.127, n.3, p.431-9. 2004.
ZEGURA, B.; VOLCIC, M.; LAH, T.T.; FILIPIC, M. Different sensitivities of
human colon adenocarcinoma (CaCo-2), astrocytoma (IPDDC-A2) and lymphoblastoid
(NCNC) cell lines to microcystin-LR induced reactive oxygen species and DNA damage.
Toxicon, v.52, p.518–525, 2008.
ZHANG, H. et al. Apoptotic Responses of Carassius auratus lymphocytes to nodularin
exposure in vitro. Fish and Shellfish Immunology, V.33, P.1229-1237, 2012.
ZIMBA, P.V. et al. Confirmation of catfish, Ictalurus punctatus (Rafinesque) mortality from
Microcystis toxins. Journal of Fish Diseases, v.24, p. 41–47, 2001.

23
MANUSCRITO
Efeitos da exposição de tilápias (Oreochromis niloticus) a saxitoxinas
produzidas por Cylindrospermopsis raciborskii (Cyanobacteria)
O trabalho será submetido à revista Harmful Algae.

24
Efeitos da exposição de tilápias (Oreochromis niloticus) a
saxitoxinas produzidas por Cylindrospermopsis raciborskii
(Cyanobacteria)
M. F. Sánchez De Bocka,b, G.S.O. Moraesb, R.G.S. Almeidab, A. J. A. Bicudob , K.R.
Sántoro,b & R. J. R. Molicaa,b*
a Programa de Pós-Graduação em Ecologia, Departamento de Biologia,
Universidade Federal Rural de Pernambuco, R. Dom Manoel de Medeiros, s/n, Dois
Irmãos - CEP: 52171-900. Recife, PE, Brasil.
b Unidade Acadêmica de Garanhuns, Universidade Federal Rural de Pernambuco,
Av. Bom Pastor s/n. CEP 55292-270. Garanhuns, PE, Brasil.
Resumo
Florações de cianobactérias têm aumentado sua frequência de ocorrência e
duração no mundo inteiro, devido ao aumento do estado trófico dos ecossistemas
aquáticos continentais, inclusive em reservatórios e viveiros utilizados na
aquicultura. Várias espécies de cianobactérias são potenciais produtoras de
metabólitos secundários tóxicos (cianotoxinas), que podem afetar os organismos
aquáticos expostos a elas. A influência da exposição crônica de uma cepa de
Cylindrospermopsis raciborskii produtora de saxitoxinas no desempenho de juvenis
de tilápia do Nilo (Oreochromis niloticus) foi avaliada mediante um experimento de
60 dias com três condições de cultivo diferentes: (i) C. raciborskii produtora de
saxitoxinas, em meio de cultura ASM-1; (ii) meio de cultura ASM-1 (controle), e (iii)
água desclorada (controle). A sobrevivência, o peso médio, ganho de peso, índice
de eficiência alimentar, entre outros, foram analisados. Peixes expostos a C.
raciborskii apresentaram uma maior mortalidade e menor crescimento em

25
comparação aos peixes dos tratamentos controle. Entretanto, o peso dos peixes
teve influência na toxicidade, indicando uma resposta dose-dependente da
mortalidade. Os resultados demonstram efeitos prejudiciais das saxitoxinas em
juvenis de tilápia do Nilo e uma vulnerabilidade em função do tamanho e da
toxicidade presente nas florações. Reforça-se, a partir da análise dos resultados, a
importância de manter a qualidade da água nos ecossistemas aquáticos. Em
estações de piscicultura isto é particularmente importante, a fim de reduzir riscos de
florações de cianobactérias produtoras de saxitoxinas, evitando, portanto, perdas
econômicas aos produtores.
* Autor para correspondência: Tel./Fax: +55 87 3764 5526
Endereços de e-mail: [email protected].
Palavras-chave: eutrofização, floração, cianotoxinas, piscicultura, desempenho de
peixes.
1. Introdução
As florações de cianobactérias têm aumentado sua frequência e intensidade
em todo o mundo devido ao aumento do processo de eutrofização (Chorus e
Bartram, 1999), particularmente em ecossistemas aquáticos com longos tempos de
retenção de água e com características que favorecem o aumento do estado trófico,
como reservatórios e viveiros utilizados na aquicultura (Sevrin-Reyssac e Pletikosic,
1990; Merel et al., 2013).
O uso humano dos ecossistemas aquáticos para diversos fins, entre eles
utilização de água para consumo, recreação e produção animal, torna as florações

26
um tema de grande importância e preocupação para a saúde pública (Briand et al.,
2003; Codd et al., 2005), pelo fato de que várias espécies de cianobactérias são
potenciais produtoras de metabólitos secundários tóxicos (cianotoxinas).
No Brasil, florações tóxicas de cianobactérias têm sido associadas a áreas de
produção aquícola, como por exemplo, de ostras e mexilhões em ambientes
marinhos costeiros (Tavares et al., 2009) e de tilápias em tanques-rede em
reservatórios (Borges et al., 2010), representando um problema potencial de saúde
humana e possíveis perdas econômicas.
Em reservatórios do nordeste semiárido brasileiro, diversos estudos (Costa et
al., 2006; Bittencourt-Oliveira et al., 2011, Moura et al., 2011; Bittencourt-Oliveira et
al., 2014) reportaram florações multi-espécie de cianobactérias filamentosas dos
gêneros Planktothrix e Cylindrospermopsis, potencialmente tóxicas, com possível
alternância de predominância, segundo as condições climáticas. Dentre as espécies
reportadas nesses trabalhos, Cylindrospermopsis raciborskii (Wolosz.) Seena. e
Subbar. é uma das espécies que desperta maior interesse, pela crescente expansão
de sua distribuição geográfica e potencial de toxicidade (Padisák, 1997; Ferrão-
Filho, 2009). Sua dominância é comumente observada em ambientes eutróficos com
alta turbidez (Figueredo e Giani, 2001; Berger et al., 2006); condições típicas de
áreas de piscicultura em tanques-rede (Borges et al., 2010; Moura et al., 2012).
Em vários países, as cepas tóxicas de C. raciborskii têm sido reportadas
como produtoras da hepatotoxina cilindrospermopsina (Ohtani et al., 1992; Saker e
Griffiths, 2000). No Brasil, todas as cepas tóxicas de C. raciborskii isoladas foram
produtoras de saxitoxinas (Lagos et al., 1999; Molica et al., 2002; Pomati et al.,
2003). Hoff-Risseti et al. (2013) identificaram a presença de genes para a síntese de
cilindrospermopsina e saxitoxinas numa mesma cepa brasileira de C.raciborskii, e

27
sugeriram que evolutivamente houve uma mudança no tipo de cianotoxina produzida
pelas cepas australianas e asiáticas para novos ambientes colonizados na América
do Sul.
As saxitoxinas são alcalóides neurotóxicos que impedem a propagação do
impulso nervoso pela ligação e bloqueio dos canais de sódio nos axônios dos
neurônios, o que leva a paralisia muscular e consequente morte de animais
vertebrados, por parada respiratória (Wiegand e Pflugmacher, 2005). Elas podem
causar malformações no desenvolvimento em peixes, atraso no tempo de eclosão,
dano genotóxico, processos apoptóticos e mortalidade em embriões, larvas e juvenis
(Oberemm et al., 1999; Lefebvre et al., 2004), assim como perda de coordenação
motora, estresse oxidativo e mortalidade em juvenis e adultos (Robineau et al.,
1991; Lefebvre et al., 2005; Ferrão-Filho, 2009; da Silva et al., 2011). Entretanto, a
maioria desses estudos têm demonstrado os efeitos deletérios das saxitoxinas
mediante a sua adição, na forma purificada, ao meio de cultivo, ou através de
lisados de células por vias não naturais, como injeção intraperitoneal. Por sua vez,
Oberemm et al. (1999) e Best et al. (2002) observaram que efeitos deletérios foram
mais intensos com a utilização de extratos aquosos brutos de cianobactérias do que
com toxinas purificadas, sugerindo que substâncias não conhecidas dos extratos
podem incrementar a absorção das toxinas ou causar interações sinergéticas,
incrementando a toxicidade. Portanto, a forma de exposição dos peixes às
cianotoxinas pode levar a diferentes conclusões.
Naturalmente, peixes filtradores, como as tilápias e carpas, estão mais
expostos às cianotoxinas, não só pela entrada de toxinas dissolvidas na água que
passam através das brânquias durante a respiração, como pela ingestão de células
de cianobactérias durante a alimentação (Malbrouck e Kestemont, 2006). Dentre as

28
espécies de peixes fito-planctófagas comercialmente importantes, a tilápia do Nilo
(Oreochromis niloticus) (Linnaeus, 1758) é uma das principais espécies da
aquicultura mundial, e ultimamente, a principal espécie da aquicultura continental no
Brasil (MPA, 2013). Portanto, o objetivo deste trabalho foi avaliar os efeitos da
exposição crônica de uma cepa de C. raciborskii produtora de saxitoxinas na
sobrevivência e índices de crescimento de juvenis de tilápia do Nilo (O. niloticus).
2. Material e métodos
2.1 Experimento
O experimento visou reproduzir as densidades celulares encontradas
normalmente em florações naturais de cianobactérias a fim de analisar o efeito da
exposição crônica a uma cepa produtora de saxitoxinas na sobrevivência e no
crescimento de juvenis de tilápia do Nilo. O desenho experimental incluiu três
tratamentos: (i) cultivo de cianobactérias em meio de cultura ASM-1 -
Cianobactérias, (ii) meio de cultura ASM-1 (controle negativo - Meio) e, (iii) água
(controle positivo - Água), contando cada tratamento com 4 repetições, e totalizando
12 unidades experimentais. O tratamento controle Meio foi incluído para testar se os
sais presentes no meio de cultura ASM-1, onde foram cultivadas as cianobactérias,
poderiam afetar os valores das variáveis resposta analisadas.
O experimento foi realizado em sistema estático com aquários de 40 L e
duração de 60 dias, que é o tempo necessário para que a espécie no estágio juvenil
(1 a 10 gramas) aumente seu peso em 10 vezes, conforme preconizado pelo NRC
(2011). A cada cinco dias, 10 ou 20 L de meio de cultivo, correspondente a cada
tratamento, era renovado em todos os aquários, dependendo das condições físico-
químicas do meio de cultivo, monitoradas através da medição diária da temperatura

29
e pH (pHmetro modelo PA 200, Marconi) e concentração de oxigênio dissolvido
(oxímetro modelo 550A, YSI). O meio de cultura ASM-1 para os tratamentos
Cianobactérias e Meio foi preparado com água de osmose reversa (Marconi
MA1400/570). O meio de cultivo do tratamento Água consistiu em água da torneira
desclorada mediante aeração em tanque reservatório.
Todos os aquários eram dotados de aeração contínua e controle de
temperatura com termostatos (25 ± 2 ºC). Os aquários foram expostos a intensidade
luminosa de 50-80 µmol.fótons m-².s-¹ (Biospherical Instruments QSL-100) em
fotoperíodo de 12h..
2.1.1 Cianobactérias
Para o tratamento Cianobactérias, utilizou-se a cepa ITEP-A1 de C.
raciborskii. Para obtenção de biomassa de C. raciborskii foram realizados cultivos
em frascos de borosilicato de 20 L com meio ASM-1 (Gorham et al., 1964), pH inicial
8.0, 25 ± 2 ºC, intensidade luminosa de 50-80 µmol.fótons m-².s-¹ (Biospherical
Instruments QSL-100), a partir de lâmpadas fluorescentes, e fotoperíodo de 12h
controlado com temporizador.
Diariamente foram coletadas amostras dos aquários de tratamento
Cianobactérias para contagem de filamentos de C. raciborskii, realizada em câmara
de Fuchs-Rosenthal. O tamanho médio dos filamentos foi estimado a partir da
medição de 100 filamentos de amostras aleatórias (da Costa et al., 2013) em
microscópio óptico (Zeiss, AxioScope A1, software AxioVision 4.7.2). O número
médio de células por filamento foi estimado dividindo o comprimento médio dos
filamentos (43 ± 22 μm) pelo comprimento médio de uma célula de C. raciborskii,
considerado 7,5 μm (Padisák, 1997). O número total de células por amostra foi

30
calculado multiplicando-se o número de filamentos contados pelo número médio de
células por filamento.
2.1.2 Peixes
Juvenis machos de tilápia do Nilo (O. niloticus) foram cedidos pela Estação
de Piscicultura do Itiúba da Companhia de Desenvolvimento dos Vales do São
Francisco e do Parnaíba (CODEVASF). Os peixes foram aclimatados às condições
ambientais por dez dias em caixas d'água circulares de fibra (500 L) e alimentados
com ração comercial farelada (40% proteína bruta, marca Duranch) duas vezes ao
dia (8:00 e 16:00 h).
Quinze juvenis de tilápia pesados (1,16 ± 0,3 g) foram distribuídos
aleatoriamente em cada aquário. A densidade de estocagem foi estabelecida após
experimentos pilotos, em função do volume de meio de cultivo e biomassa final
estimada para 60 dias de experimento. A primeira semana após a transferência foi
considerada como período de adaptação às condições experimentais. Na ocorrência
de mortalidade neste período, os peixes mortos foram substituídos por outros de
mesmo peso e tamanho.
Os peixes foram alimentados diariamente na proporção de 5% da biomassa,
com ração comercial (40% proteína bruta) em três refeições diárias (08:00, 12:00 e
17:00 h), sendo utilizada ração farelada nos primeiros 30 dias de cultivo, e a partir do
31º dia até o final do experimento, ração extrusada com granulometria de 2mm. A
quantidade de ração fornecida foi ajustada a cada 10 dias, após pesagem dos lotes
e a cada registro de mortalidade de peixes (Sakomura e Rostagno, 2007). Todas as
pesagens foram precedidas de jejum por 24h e os peixes submetidos a anestesia
com benzocaína (50 mg L-1).

31
As variáveis analisadas foram: Sobrevivência (%) (número diário de peixes no
lote/ número inicial de peixes), Peso médio final (g) (peso final do lote/ número final
de peixes), Ganho de peso médio (g) (peso final – peso inicial), Índice de eficiência
alimentar (g) (ganho de peso médio / consumo de alimento) e Taxa de eficiência
proteica (ganho de peso médio (g) / ingestão de proteína).
2.2 Análises químicas de composição corporal
No início do experimento, 50 peixes da população inicial foram sacrificados
por overdose de benzocaína (500 mg L-1), moídos e homogeneizados para
constituírem uma amostra composta e mantidos a -18oC para posterior determinação
da composição corporal inicial (extrato etéreo, matéria mineral, proteína bruta e
umidade). Ao final do período experimental, quatro peixes de cada aquário foram
submetidos ao mesmo protocolo para determinação da composição corporal.
Todas as amostras foram secas em estufa de ventilação forçada a 50°C por
48h. A matéria seca foi determinada mediante secagem das amostras em estufa a
105ºC por 24 horas. A proteína bruta foi determinada pelo método Micro Kjeldahl
(AOAC, 1990), o extrato etéreo medido com extrator automático (marca Ankom,
mod. XT 15) e a matéria mineral foi calculada a partir da incineração das amostras
por 2 horas a 550 °C em forno mufla. Todas as amostras foram analisadas em
duplicata.
A partir dos dados de composição corporal foi estimada a taxa de retenção
proteica aparente (TRP = ((peso final × proteína corporal final) - (peso inicial ×
proteína corporal inicial))/ ingestão de proteína)× 100).

32
2.3 Análises de saxitoxinas
As amostras de 1L de meio de cultivo do tratamento Cianobactérias,
coletadas a cada troca de meio de cultivo, foram congeladas e liofilizadas para
análise da presença de saxitoxinas por cromatografia líquida de alta eficiência
(HPLC-High Performance Liquid Cromatography). A extração de saxitoxinas foi feita
com 5 mL ácido acético 0,05 M adicionado a uma massa de 100 mg do material
liofilizado (Oshima, 1995). A solução foi agitada por 1 hora e centrifugada a 21.380 g
por 10 minutos, separando o sobrenadante. Este procedimento foi repetido mais
duas vezes com o precipitado, porém com um tempo de extração de 30 minutos, e
ao final todos os sobrenadantes constituíram a amostra que foi utilizada na análise
cromatográfica.
Para a análise cromatográfica, utilizou-se um equipamento Shimadzu, modelo
Proeminance, a qual consistiu em um sistema on line de derivatização pós-coluna
em tubo de teflon (10 m de comprimento e 0,5 mm de diâmetro) e monitoramento em
detector de fluorescência (RF-10AXL) (330 nm emissão; 390 nm excitação),
conforme descrito por Oshima (1995). A fase móvel para detecção dos análogos de
saxitoxinas não-sulfatadas (saxitoxina, neo-saxitoxina e dc-saxitoxina) consistiu em
uma solução de 2mM de 1-heptanosulfonato em 30mM de tampão fosfato de amônia
(30mM), pH 7,1 e 3% acetonitrila. O fluxo da fase móvel foi de 0,8 mL.min-1 e foi
utilizada uma coluna C8 (Shimadzu LC 250 x 4 mm, 5 μm). A oxidação pós-coluna
foi feita com solução oxidante (7,0 mM de ácido periódico em 10,0 mM de tampão
fosfato de potássio pH 9,0) e para interromper a reação utilizou-se ácido acético
0,5M, ambas com fluxo de 04 mL.min-1. A identificação dos análogos de saxitoxinas
foi realizada por comparação dos tempos de retenção dos picos das amostras com
os picos dos padrões. Como garantia, a análise cromatográfica foi repetida, porém

33
as soluções oxidante e ácido acético 0,5 M foram substituídas por água ultra-pura..
Os padrões de saxitoxinas utilizados nas análises foram neosaxitoxina (neo-STX),
saxitoxina (STX), dc-saxitoxina (dc-STX), os quais foram adquiridos da NRC Halifax,
Canadá.
Após a quantificação dos análogos, as suas concentrações foram
transformadas em equivalentes de saxitoxina (STXequiv), considerando a toxicidade
relativa de cada análogo em relação à toxicidade do análogo saxitoxina, que é o
mais tóxico (Oshima, 1995).
A concentração de saxitoxinas foi expressa por volume (μg STXequiv.L-1), por
peso seco de amostra (μg STXequiv.g-1), ou concentração de exposição total por
biomassa de peixe ao longo do período experimental (μg STXequiv.L-1.g-1). Para
esta última, o peso médio diário por peixe foi estimado mediante análise de
regressão, com ajuste ao modelo exponencial (R2 =0,99; p < 0,01) para todo o
período experimental.
2.4 Análises estatísticas
Os resultados obtidos foram submetidos a análise de variância (ANOVA) de
um fator e, quando significativa, as médias foram comparadas pelo teste de Tukey a
5% de probabilidade (p < 0,05).
Para estimar o efeito da mortalidade sobre o peso médio e a eficiência
alimentar dos peixes, foram gerados modelos através de análise de regressão linear
simples. As inclinações das retas foram comparadas pelo teste t de Student.
Incluíram-se nestas análises as remoções de dois peixes por aquário a cada 20 dias,
para cálculo de índices víscero-somático e hepato-somático (resultados não
apresentados), dessa forma a mortalidade representou a densidade de estocagem.

34
A influência do peso médio dos peixes e da concentração total de saxitoxinas
ao longo do período experimental, expressa em proporção do peso vivo corpóreo
médio estimado dos peixes, na mortalidade foi estimada por análise de regressão,
com ajuste ao modelo quadrático. Especificamente, para estas análises, foram
consideradas as mortes registradas no período de adaptação, a fim de incluir os
mínimos valores de peso. Os valores de peso e concentração de saxitoxinas
associados à máxima mortalidade foram calculados pela derivada das equações
quadráticas. Para todas as análises estatísticas utilizou-se o software SAS 9.0.
O presente trabalho foi aprovado pela Comissão de Ética e Uso de Animais
da UFRPE (Processo Nº 23082.012185/2010 – CEUA E07).
3. Resultados e Discussão
Os parâmetros físico-químicos dos meios de cultivo variaram, porém
mantiveram-se dentro das faixas de tolerância da tilápia do Nilo (O. niloticus) durante
todo o experimento. O pH foi similar (p>0,05) em todos os tratamentos (Tabela 1). A
temperatura no tratamento Água foi maior (p<0,05) que no tratamento Meio, embora
ambos tenham sido similares (p>0,05) ao tratamento Cianobactérias. O oxigênio
dissolvido foi menor (p<0,05) no tratamento Cianobactérias em relação aos outros
tratamentos, que por sua vez não diferiram entre si (p>0,05).
As mortalidades de peixes após florações tóxicas de cianobactérias foram
associadas por muitos autores às condições ambientais decorrentes da sua rápida
decomposição: temperatura mais elevada da água, pH alcalino, altas concentrações
de amônio, baixa concentração de oxigênio dissolvido ou pela combinação de todos
esses fatores (Sevrin-Reyssac e Pletikosic, 1990; Jewel et al., 2003; Kangur et al.,
2005). Embora estatisticamente diferentes, não se pode atribuir a uma diferença de

35
0,1ºC das médias de temperatura, um melhor desempenho dos peixes, porque os
valores de temperatura ficaram sempre na faixa ótima - 25,0 a 30,0 ºC (Hussain,
2004) - para o crescimento dos juvenis de tilápia. As concentrações de oxigênio
dissolvido (OD) mantiveram-se entre os valores de tolerância de O. niloticus às
condições ambientais para o cultivo, que segundo Hussain (2004) abrange de 2,0 a
8,0 mg L-1. Entretanto, os valores mínimos de OD foram observados no tratamento
Cianobactérias, após a senescência das células e provável decomposição da
matéria-orgânica. De acordo com Baldisserotto (2002), o crescimento dos peixes
não é afetado enquanto as concentrações de oxigênio na água permanecerem
acima do valor crítico para a espécie. Os peixes alocados no tratamento com C.
raciborskii podem ter experimentado, temporariamente, uma diminuição do consumo
do alimento em razão da menor concentração de oxigênio. Todavia, após a
renovação do meio de cultivo, um ganho de peso compensatório pode ter ocorrido.
O crescimento compensatório é definido por Dobson e Holmes (1984) como uma
fase de crescimento acelerado, maior que o normal, resultado da adequada
realimentação dos peixes, após um período de privação alimentar ou após uma
situação de estresse.
Tabela 1
Variáveis de qualidade de água (média ± desvio padrão) durante o período experimental. Entre
parêntese se informa o intervalo de confiança.
PARÂMETRO TRATAMENTO
CIANOBACTÉRIAS ÁGUA MEIO DE CULTURA
Oxigênio dissolvido
(mg L-1
)
3,73 ± 1,55 b
(2,18 – 5,28)
4,69 ± 1,08 a
(3,61- 5,77)
4,44 ± 1,06 a
(3,38 – 5,55)
Temperatura
(ºC)
26,32 ± 0,63 ab
(25,69 - 26,95)
26,42 ± 0,51 a
(25,91- 26,93)
26,30 ± 0,46 b
(25,84 - 26,76)
pH 6,90 ± 0,38 a
(6,52 - 7,28)
6,90 ± 0,32 a
(6,58 - 7,22)
6,89 ± 0,23 a
(6,66 - 7,12)
Letras diferentes na mesma linha indicam diferença significativa (p < 0,05) pelo Teste de Tukey.
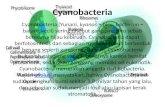
36
No tratamento de exposição à cianobactéria C. raciborskii, a densidade de
células por volume de cultivo nos aquários apresentou variabilidade durante o
período experimental, pela senescência das cianobactérias e consequente reposição
com cultivos novos, sendo que a densidade diminuía gradativamente nos dias
subsequentes à troca (Fig. 1). Os três análogos (STX, neo-STX e dc-STX) foram
encontrados em todas as amostras, sendo, em todos os casos, neo-saxitoxina o
mais abundante. A concentração das saxitoxinas por volume e a concentração de
exposição total por biomassa de peixe ao longo do período experimental também
variaram ao longo do tempo (Fig. 1).
Fig. 1. Variações na concentração de saxitoxinas por volume (STX, μg STXequiv.L-1
), concentração
total de saxitoxinas disponível por biomassa de peixe (μg STXequiv. L-1
.g-1
) e densidade celular de
Cylindrospermopsis raciborskii (células.L-1
) no tratamento Cianobactérias durante o período
experimental.
Tal variação é comumente observada em florações naturais, como registrado
por Borges et al. (2010), Moura et al. (2012) e Bittencourt-Oliveira et al. (2014) na
região nordeste do Brasil. A densidade celular de C. raciborskii encontrou-se a maior

37
parte do tempo entre 1x107 e 2x108 células.L-1, sendo 9x107 células.L-1 a densidade
média.
Na figura 1 pode se observar uma diminuição na densidade de C. raciborskii a
partir do trigésimo dia de experimento, sendo a densidade média nos primeiros 30
dias de 1x108 células.L-1, e nos últimos 30 dias, 6x107 células.L-1. Essa diminuição
de C. raciborskii provavelmente foi sido causada por um maior consumo das
cianobactérias pelos peixes em ativa fase de crescimento. Turker et al. (2003) e
Figueredo e Giani (2005) observaram uma diminuição da densidade de
cianobactérias filamentosas de grande porte, como C. raciborskii, na presença de
tilápia do Nilo em experimentos de biomanipulação in situ em reservatórios, atribuída
pelos autores ao consumo das cianobactérias pelos peixes. Não entanto, a
densidade média de cianobactérias durante todo o experimento foi similar às
densidaded de cianobactérias observadas por Borges et al. (2010), em tanques de
piscicultura em reservatórios, que variaram entre 1,5x107 e 1,8x108 células.L-1.
Adicionalmente, a densidade média esteve bem acima do nível de alerta para águas
destinadas ao consumo humano, que é de 2x104 células.L-1 (Brasil, 2011).
A concentração média de saxitoxinas por volume de cultivo durante os 60 dias
de período experimental foi de 9,19 ± 6,72 µg STXequiv. L-1, com mínimo e máximo
de 2,32 e 26,21 µg STXequiv. L-1, respectivamente (Fig. 1). Essas concentrações
estão em torno e acima do limite máximo aceitável de saxitoxinas em água para
consumo humano no Brasil (3 µg STXequiv. L-1) (Brasil, 2011). As concentrações de
cianotoxinas variam grandemente em eventos de florações (Chorus e Bartram,
1999). Poucos trabalhos no Brasil reportam concentrações de saxitoxinas
produzidas durante florações naturais. Molica et al. (2005) reportaram uma
concentração de 99 ng STXequiv.L-1, e Costa et al. (2006) uma concentração de
38
3,14 µg STXequiv.L-1, ambos estudos realizados durante florações com dominância
de C. raciborskii.
Carmichael et al. (1997) registraram concentrações de 5 a 60 µg STXequiv.g-1
(equivalentes de saxitoxinas por grama de peso seco de sestón) numa floração nos
Estados Unidos, enquanto que em florações de cianobactérias na Austrália as
concentrações de saxitoxinas variaram entre 50 e 3400 µg STXequiv.g-1 (Chorus e
Bartram, 1999). A concentração média de saxitoxinas por grama de peso seco de
sestón à que foram expostos os juvenis de tilápia do Nilo no presente trabalho foi de
35 µg STXequiv.g-1, variando entre 9,92 e 97,57 µg STXequiv.g-1.
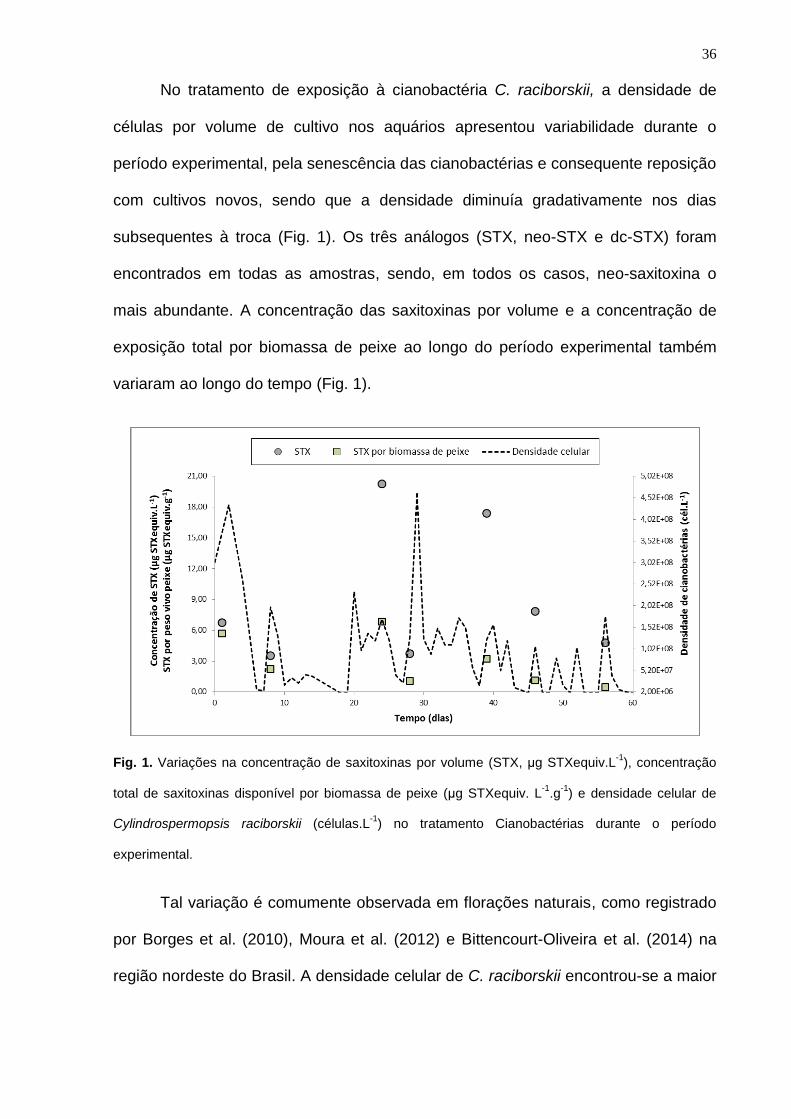
Uma menor sobrevivência dos peixes foi observada no tratamento
Cianobactérias (65 ± 19,9%, p= 0,0043) em relação aos tratamentos controle Meio
(98,33 ± 3,33%) e Água (98,33 ± 3,33%), e não houve nenhuma diferença (p>0,05)
na sobrevivência entre os tratamentos controle (Fig.2, Tabela 2, Tabela 3).
Fig. 2. Sobrevivência acumulada (%) de juvenis de tilápia do Nilo (Oreochromis niloticus) expostos
aos diferentes meios de cultivo. Letras diferentes nas linhas indicam diferença significativa (p < 0,05)
pelo Teste de Tukey.
(a)
(b)
39
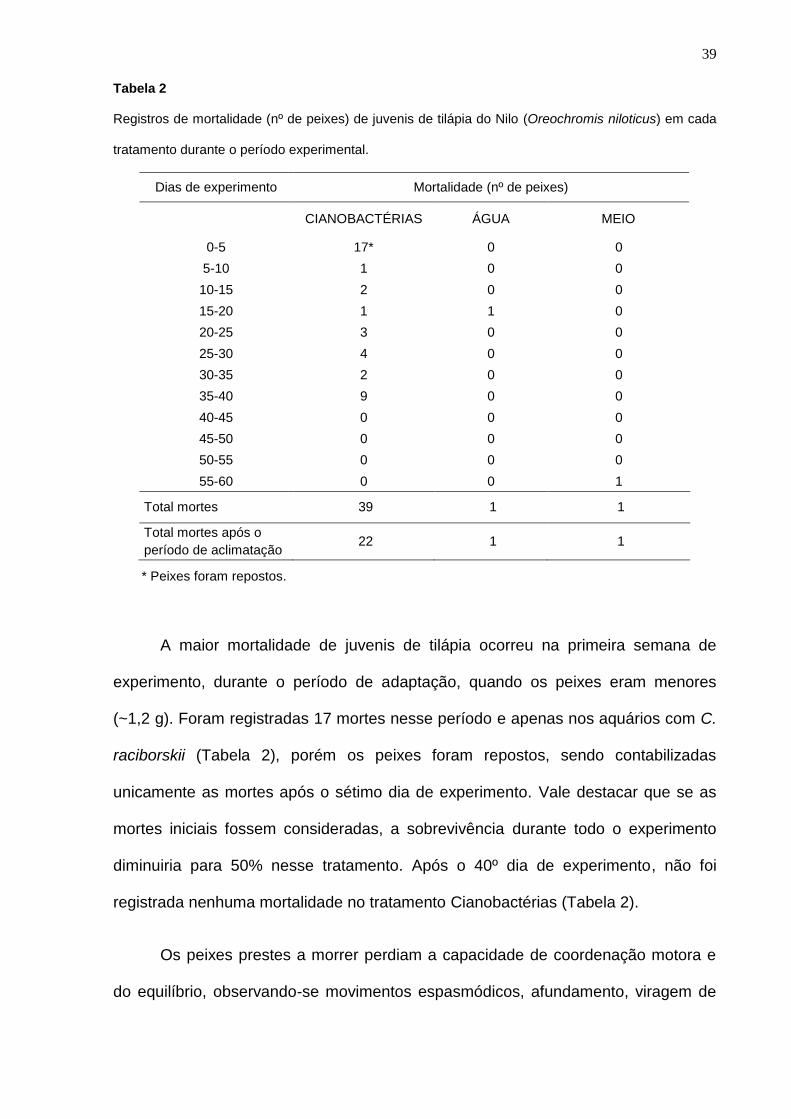
Tabela 2
Registros de mortalidade (nº de peixes) de juvenis de tilápia do Nilo (Oreochromis niloticus) em cada
tratamento durante o período experimental.
Dias de experimento Mortalidade (nº de peixes)
CIANOBACTÉRIAS ÁGUA MEIO
0-5 17* 0 0
5-10 1 0 0
10-15 2 0 0
15-20 1 1 0
20-25 3 0 0
25-30 4 0 0
30-35 2 0 0
35-40 9 0 0
40-45 0 0 0
45-50 0 0 0
50-55 0 0 0
55-60 0 0 1
Total mortes 39 1 1
Total mortes após o
período de aclimatação 22 1 1
* Peixes foram repostos.
A maior mortalidade de juvenis de tilápia ocorreu na primeira semana de
experimento, durante o período de adaptação, quando os peixes eram menores
(~1,2 g). Foram registradas 17 mortes nesse período e apenas nos aquários com C.
raciborskii (Tabela 2), porém os peixes foram repostos, sendo contabilizadas
unicamente as mortes após o sétimo dia de experimento. Vale destacar que se as
mortes iniciais fossem consideradas, a sobrevivência durante todo o experimento
diminuiria para 50% nesse tratamento. Após o 40º dia de experimento, não foi
registrada nenhuma mortalidade no tratamento Cianobactérias (Tabela 2).
Os peixes prestes a morrer perdiam a capacidade de coordenação motora e
do equilíbrio, observando-se movimentos espasmódicos, afundamento, viragem de

40
lado, aumento dos movimentos operculares até paralisia total e morte. O
afundamento e viragem lateral apontam para uma falha nas funções da bexiga
natatória dos juvenis de tilápia do Nilo, como observado por Lefebvre et al. (2004).
Bouvy et al. (1999) também relataram sintomas similares em tilápias que receberam
injeções intraperitoniais de extrato bruto de uma floração de C. raciborskii com
presença de neurotoxinas. Os sintomas observados também concordam com os
descritos por Lefebvre et al. (2004) em larvas de peixe-zebra Danio rerio (Hamilton,
1822) expostas a saxitoxinas dissolvidas no meio de cultivo, cujas concentrações de
exposição (229 ± 7 e 481 ± 40 μg STXequiv.L−1) foram muito superiores às do
presente trabalho (valor médio de 9,19 ± 6,72 µg STXequiv.L−1).
As variáveis de desempenho peso médio final, ganho de peso médio, índice
de eficiência alimentar e taxa de eficiência proteica não apresentaram diferenças
entre os três tratamentos (Tabela 3). Entretanto, a análise de variância avaliou
resultados médios em diferentes densidades de estocagens, pois durante todo o
período experimental foi observada apenas 1 morte em cada tratamento controle
Água e Meio (Tabela 2), sendo suas densidades de estocagem, considerando o
número de indivíduos por volume, praticamente constantes. Já no tratamento
Cianobactérias, a mortalidade aumentou no decorrer do tempo (Fig. 2, Tabela 2),
diminuindo a densidade de estocagem nos aquários deste tratamento.

41
Tabela 3
Variáveis de desempenho (média ± desvio padrão) dos juvenis de tilápia do Nilo (Oreochromis
niloticus) expostos aos diferentes meios de cultivo.
PARÂMETROS TRATAMENTO
CIANOBACTÉRIAS ÁGUA MEIO
Sobrevivência (%) 65,00 ± 19,91 b 98, 32 ± 3,33 a 98.32 ± 3,33 a
Peso médio final (g) 12,70 ± 1,01 a 11,57 ± 0,94 a 11,10 ± 0,88 a
Ganho de peso médio (g) 11,55 ± 1,00 a 10,40 ± 0,93 a 9,94 ± 0,89 a
Índice de Eficiência Alimentar 1,07 ± 0,09 a 1,00 ± 0,04 a 0,99 ± 0,07 a
Taxa de eficiência proteica 2,95 ± 0,24 a 2,74 ± 0,10 a 2,67 ± 0,21 a
Letras diferentes na mesma linha indicam diferença significativa (p < 0,05) pelo Teste de Tukey.
Ao estimar-se a influência da mortalidade nos índices de crescimento, (peso
médio e índice de eficiência alimentar) dos juvenis de tilápia do Nilo cultivados (Figs.
3 e 4), observou-se uma relação positiva entre a mortalidade e o crescimento dos
peixes.
Fig. 3. Estimativa da influência da mortalidade sobre o peso médio de juvenis de tilápia do Nilo
(Oreochromis niloticus) cultivados nos diferentes meios de cultivo. Em parêntesis o grau de
significância do ajuste ao modelo de regressão linear: (**) = p < 0,01.

42
Na regressão entre o número de mortes e o peso médio dos peixes (Fig. 3), a
inclinação das retas dos tratamentos controle Água e Meio foi maior que a inclinação
da reta do tratamento Cianobactérias (1,78, 1,70 e 0,76, teste t de Student,
p<0,0001), o que demonstra um menor peso médio dos peixes neste tratamento. Da
mesma maneira, na regressão entre o número de mortes e o índice de eficiência
alimentar (Fig. 4), a inclinação da reta do tratamento Cianobactérias foi menor que a
do tratamento Água (0,0701 e 0,0954, respectivamente, teste t de Student, p= 0,0109). O tratamento Meio não apresentou ajuste ao modelo de regressão linear
(p= 0,1273), portanto não foi considerado na análise de comparação das inclinações
das retas.
Fig. 4. Estimativa da influência da mortalidade sobre o índice de eficiência alimentar de juvenis de
tilápia do Nilo (Oreochromis niloticus) cultivados nos diferentes meios de cultivo. Em parêntesis o grau
de significância do ajuste ao modelo de regressão linear: (**) = p < 0,01, (*) = p < 0,05, (NS) = p >
0,05.
Portanto, apesar das variáveis de desempenho entre tratamentos não terem
apresentado diferenças nos valores médios (Tabela 3), as análises de regressão
mostradas nas figuras 3 e 4, ao normalizarem a densidade de estocagem dos

43
peixes, mostraram que o peso médio e o índice de eficiência alimentar seriam
menores no tratamento Cianobactérias. Isto indica que os juvenis de tilápia do
tratamento Cianobactérias atingiriam peso médio inferior, com uma diferença de
mais de 100% ao final do período experimental, e também teriam uma menor
eficiência alimentar, caso a densidade de estocagem dos peixes cultivados fosse
similar àquela dos tratamentos controle. Portanto, conclui-se que a maior
mortalidade no tratamento Cianobactérias durante o período experimental propiciou
um efeito compensatório nos índices de desempenho dos peixes expostos a esse
tratamento, em função de um menor gasto energético dispensado em brigas para
defesa do território e maior tempo dedicado à alimentação, como indicado também
por Baldisserotto (2002).
Ao mesmo tempo, em função dos resultados normalizados em relação às
densidades de estocagem, o crescimento dos peixes expostos ao tratamento
Cianobactérias e às saxitoxinas pode ter sido prejudicado por um aumento no gasto
metabólico em razão da biotransformação daquelas toxinas. O principal mecanismo
envolvido na biotransformação das saxitoxinas é o da enzima de detoxificação
glutationa-s-transferase (Wiegand e Pflugmacher, 2005), através da catalisação da
conjugação da glutationa com as saxitoxinas, transformando-as em produtos menos
tóxicos e mais solúveis em água, facilitando a sua excreção (Ferrão-Filho, 2009). A
tolerância dos juvenis de tilápia sobreviventes se deve, provavelmente, a um
processo de adaptação através do aumento da atividade (Silva de Assis et al., 2013)
e/ou da expressão gênica de enzimas de detoxificação (Gustafsson et al., 2005).
Esse mecanismo pode ter permitido a sobrevivência dos peixes, com o aumento do
gasto energético, afetando negativamente o crescimento somático dos peixes, como
observado por Wiegand et al. (1999) no crescimento de larvas de peixe-zebra,
expostos a microcistinas. Os autores observaram uma maior atividade das enzimas
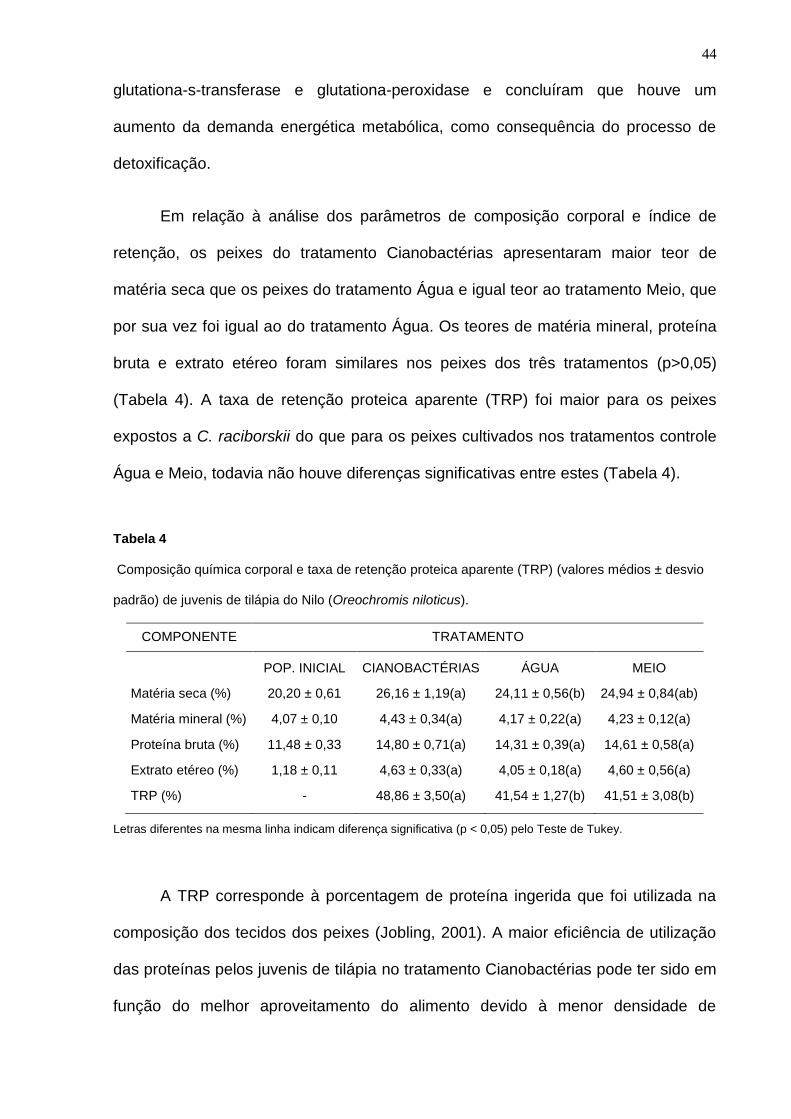
44
glutationa-s-transferase e glutationa-peroxidase e concluíram que houve um
aumento da demanda energética metabólica, como consequência do processo de
detoxificação.
Em relação à análise dos parâmetros de composição corporal e índice de
retenção, os peixes do tratamento Cianobactérias apresentaram maior teor de
matéria seca que os peixes do tratamento Água e igual teor ao tratamento Meio, que
por sua vez foi igual ao do tratamento Água. Os teores de matéria mineral, proteína
bruta e extrato etéreo foram similares nos peixes dos três tratamentos (p>0,05)
(Tabela 4). A taxa de retenção proteica aparente (TRP) foi maior para os peixes
expostos a C. raciborskii do que para os peixes cultivados nos tratamentos controle
Água e Meio, todavia não houve diferenças significativas entre estes (Tabela 4).
Tabela 4
Composição química corporal e taxa de retenção proteica aparente (TRP) (valores médios ± desvio
padrão) de juvenis de tilápia do Nilo (Oreochromis niloticus).
COMPONENTE TRATAMENTO
POP. INICIAL CIANOBACTÉRIAS ÁGUA MEIO
Matéria seca (%) 20,20 ± 0,61 26,16 ± 1,19(a) 24,11 ± 0,56(b) 24,94 ± 0,84(ab)
Matéria mineral (%) 4,07 ± 0,10 4,43 ± 0,34(a) 4,17 ± 0,22(a) 4,23 ± 0,12(a)
Proteína bruta (%) 11,48 ± 0,33 14,80 ± 0,71(a) 14,31 ± 0,39(a) 14,61 ± 0,58(a)
Extrato etéreo (%) 1,18 ± 0,11 4,63 ± 0,33(a) 4,05 ± 0,18(a) 4,60 ± 0,56(a)
TRP (%) - 48,86 ± 3,50(a) 41,54 ± 1,27(b) 41,51 ± 3,08(b)
Letras diferentes na mesma linha indicam diferença significativa (p < 0,05) pelo Teste de Tukey.
A TRP corresponde à porcentagem de proteína ingerida que foi utilizada na
composição dos tecidos dos peixes (Jobling, 2001). A maior eficiência de utilização
das proteínas pelos juvenis de tilápia no tratamento Cianobactérias pode ter sido em
função do melhor aproveitamento do alimento devido à menor densidade de

45
estocagem, como descrito por Baldisserotto (2002), ou à utilização das células de C.
raciborskii no meio de cultivo, como fonte alimentaria adicional, para constituição da
biomassa corporal.
O. niloticus é uma espécie onívora, filtrando eficientemente cianobactérias de
grande porte como a espécie filamentosa C. raciborskii através dos rastros
branquiais (Turker et al., 2003) e possui uma elevada capacidade de digestão de
cianobactérias devido ao baixo pH (=1.25) no seu estômago, como indicado por
Lazzaro (1987). Não entanto, considerando a alta concentração de proteínas na
ração oferecida aos juvenis de tilápia do Nilo (40%) no experimento, provavelmente
as cianobactérias podem ter fornecido nutrientes adicionais aos juvenis de tilápia do
Nilo, favorecendo uma maior retenção proteica aos peixes expostos a C. raciborskii.
Por exemplo, a vitamina B6, que atua como coenzima na desaminação de
aminoácidos e é importante para peixes jovens em fase ativa de crescimento (Lovell,
1998). Aquela vitamina é unicamente aportada no ambiente natural por bactérias e
cianobactérias e apesar de presente nas rações comerciais, por ser hidrossolúvel,
lixívia rapidamente da ração, ficando menos acessível para os peixes cultivados.
Finalmente, em razão das mortes de peixes terem sido observadas apenas
até o 40o dia de experimento (peso médio em torno de 6 g), mesmo os peixes tendo
sido expostos até o dia 60º a concentrações de saxitoxinas que anteriormente
causaram mortalidade (Fig. 5), analisou-se a influência do peso dos peixes e da
concentração de exposição total de saxitoxinas por biomassa de peixe sobre a
mortalidade dos juvenis de tilápia do Nilo (Figs. 6 e 7).
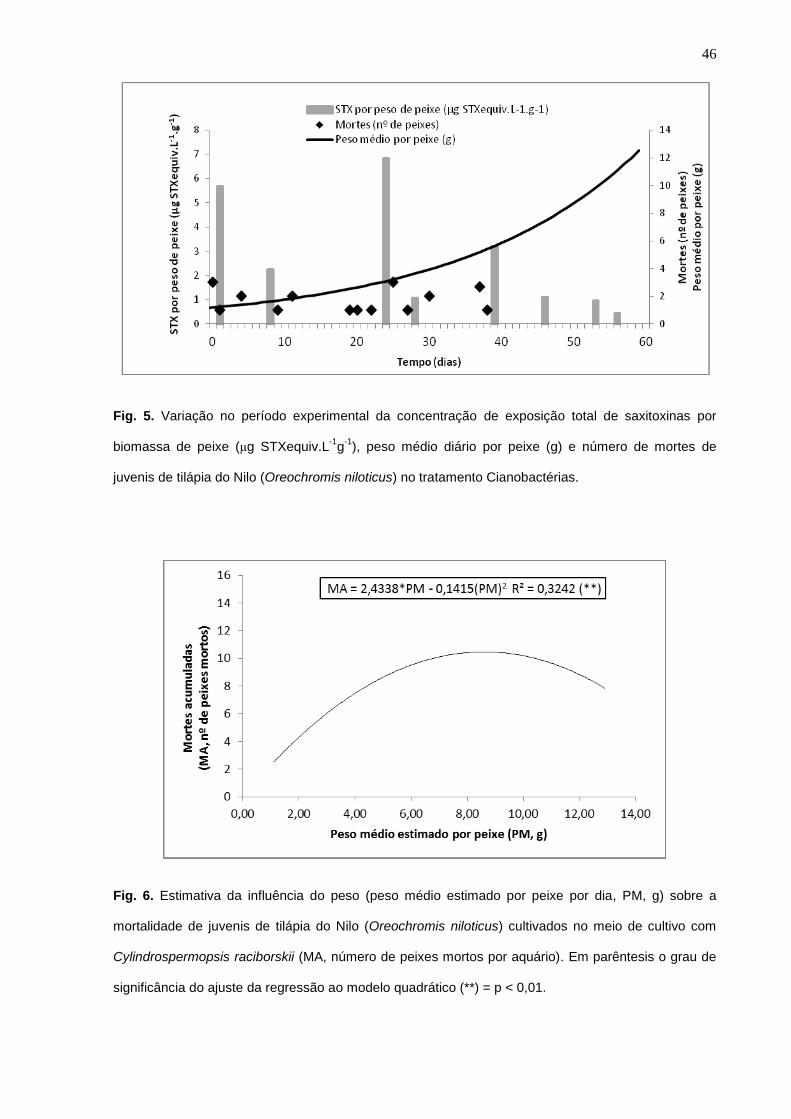
46
Fig. 5. Variação no período experimental da concentração de exposição total de saxitoxinas por
biomassa de peixe (μg STXequiv.L-1
g-1
), peso médio diário por peixe (g) e número de mortes de
juvenis de tilápia do Nilo (Oreochromis niloticus) no tratamento Cianobactérias.
Fig. 6. Estimativa da influência do peso (peso médio estimado por peixe por dia, PM, g) sobre a
mortalidade de juvenis de tilápia do Nilo (Oreochromis niloticus) cultivados no meio de cultivo com
Cylindrospermopsis raciborskii (MA, número de peixes mortos por aquário). Em parêntesis o grau de
significância do ajuste da regressão ao modelo quadrático (**) = p < 0,01.

47
Fig. 7. Estimativa da influência da concentração de exposição total de saxitoxinas por biomassa de
peixe (STXT, μg STX.L-1
.g-1
) sobre a mortalidade de juvenis de tilápia do Nilo (Oreochromis niloticus)
cultivados no meio de cultivo com C. raciborskii (MA, número de peixes mortos por aquário). Em
parêntesis o grau de significância do ajuste da regressão ao modelo quadrático (**) = p < 0,01.
Ambas as regressões, através das quais se avaliou a influência do peso dos
peixes e da concentração de exposição total de saxitoxinas por biomassa de peixe
sobre a mortalidade dos juvenis de tilápia do Nilo, ajustaram-se ao modelo
quadrático. Isto implica que há um valor de peso médio (8,60 g) ao qual se atribui
uma mortalidade máxima e, a partir deste, a mortalidade decai (Fig. 6). Da mesma
maneira, a concentração de exposição total de saxitoxinas por biomassa de peixe de
11,05 μg STXequiv.L-1.g-1 está associada a uma mortalidade máxima, ou seja, a
partir daquela concentração, há cada vez menos efeitos dessas toxinas na
mortalidade dos peixes.
Estes resultados indicam que o efeito das saxitoxinas é dose-dependente e
como a taxa de crescimento dos peixes é muito alta na fase juvenil de

48
desenvolvimento (Baldisserotto, 2002), a relação entre a concentração de
saxitoxinas e o peso e diminui rapidamente. Dessa forma, com o rápido aumento de
biomassa, os peixes poderiam atingir uma maior eficiência na detoxificação das
saxitoxinas. Isso pode ser complementar a um processo de adaptação dos peixes,
que envolveria o aumento da atividade e ou expressão gênica de enzimas de
detoxificação.
Segundo Bakke e Horsberg (2010) a meia-vida de saxitoxinas no salmão do
atlântico (Salmo salar) é de 102,6 minutos, o que corrobora com as baixas
concentrações de saxitoxinas observadas em tecidos de tilápia adultas (440 e 10 ng
STXequiv.g-1 de tecido no fígado e em músculos, respectivamente) cultivadas em
tanques-rede num reservatório da região sudeste do Brasil, durante um evento de
floração da cianobactéria Anabaena spiroides Klebahn com presença de saxitoxinas
(Galvão et al. 2009). Entretanto, Landsberg et al. (2006) e Abbott et al. (2009)
observaram acúmulo de saxitoxinas em concentrações mais elevadas (2 e 10 μg
STXequiv.g-1 de fígado e músculos, respectivamente) em três espécies marinhas de
baiacu, Sphoeroides nephelus (Goode e Bean, 1882), S. testudineus (Linnaeus,
1758) e S. spengleri (Bloch, 1785). Isto demonstra não só uma capacidade
diferencial de detoxificação em relação ao peso, mas também possivelmente a
existência de diversos mecanismos de detoxificação em diferentes espécies de
peixes.
4. Conclusão
Não sendo possível predizer a toxicidade das florações (Merel et al., 2013) e
considerando que a exposição dos peixes às saxitoxinas afetou não só a
sobrevivência, como também o crescimento de juvenis da tilápia do Nilo, conclui-se

49
que florações de espécies de cianobactérias produtoras daquelas toxinas podem
causar sérios prejuízos ao produtor, particularmente nos primeiros estágios de
desenvolvimento.
Considerando-se que a tilápia do Nilo (O. niloticus) é altamente tolerante a
condições ambientais rigorosas (Signor et al., 2010), ecologicamente, os efeitos
nocivos das saxitoxinas poderão ser bem mais severos em populações de espécies
nativas menos rústicas.
Em vista do aumento da frequência de florações tóxicas de cianobactérias e
de densidades muito superiores às citadas previamente, segundo um recente
trabalho de Bittencourt-Oliveira et al. (2014), sugere-se aprofundar os estudos dos
efeitos de cianotoxinas no desempenho de espécies de peixes nativos, adotando-se,
o modelo de exposição utilizado no presente trabalho.

50
5. Referências bibliográficas
Abbott, J.P., Flewelling, L.J., Landsberg, J.H., 2009. Saxitoxin monitoring in three species of Florida
puffer fish. Harmful Algae. 8, 343–348.
Association of Official Analytical Chemists (AOAC). 1990. Official Methods of Analysis. Arlington, VA.
Bakke, M.J.; Horsberg, T.E., 2010. Kinetic properties of saxitoxin in Atlantic salmon (Salmo salar) and
Atlantic cod (Gadus morhua). Comparative Biochemistry and Physiology. 152, 444–450.
Baldisserotto, B., 2002. Fisiologia de peixes aplicada à piscicultura. Ed. UFSM, Santa Maria, Brasil.
Berger, C., Ba, N., Gugger, M., Bouvy, M., Rusconi, F., Couté, A., Troussellier, M., Bernard, C., 2006.
Seasonal dynamics and toxicity of Cylindrospermopsis raciborskii in Lake Guiers (Senegal, West
Africa). FEMS Microbiol. Ecol. 57(3), 355-366.
Best, J.H., Pflugmacher, S., Wiegand ,C., Eddy, F.B., Metcalf, J.S., Codd, G.A., 2002. Effects of
enteric bacterial and cyanobacterial lipopolysaccharides, and of MC-LR, on glutathione S-transferase
activities in zebra fish (Danio rerio). Aquat. Toxicol. 60, 223–231.
Bittencourt-Oliveira, M.C., Piccin-Santos, V., Kujbida, P., Moura, A.N.. 2011. Cylindrospermopsin in
water supply reservoirs in Brazil determined by immunochemical and molecular methods. J. Water
Resour. Prot. 3, 349-355.
Bittencourt-Oliveira, M.C., Piccin-Santos, V., Moura, A.N., Aragão-Tavares, N.K.C., Cordeiro-Araújo,
M.K., 2014. Cyanobacteria, microcystins and cylindrospermopsin in public drinking supply reservoirs
of Brazil. An. Acad. Bras. Cienc. 86(1), 297-309.
Borges, P.A.F. et al., 2010. Effects of fish farming on plankton structure in a Brazilian tropical
reservoir. Hydrobiologia. 649, 279-291.
Bouvy, M., Molica, R., De Oliveira, S., Marinho, M., Beker, B., 1999. Dynamics of a toxic
cyanobacterial bloom (Cylindrospermopsis raciborskii) in a shallow reservoir in the semi-arid region
ofnortheast Brazil. Aquat. Microb. Ecol. 20, 285-297.

51
Brasil, 2011. Ministério da Saúde. Portaria Nº 2.914, 12 de dezembro de 2011. Diário Oficial [da]
República Federativa do Brasil, Poder Executivo, Brasília, DF.
Briand, J.F., Jacquet, S., Bernard, C., Humbert, J.F., 2003. Health hazards for terrestrial vertebrates
from toxic cyanobacteria in surface water ecosystems. Veterinary Research. 34, 361-377.
Carmichael, W.W., Evans, W.R., Yin, Q.Q., Bell, P., Mocauklowski, E., 1997. Evidence for paralytic
shellfish poisons in the freshwater cyanobacterium Lyngbya wollei (Farlow ex Gomont) comb. nov.
Appl. Environ. Microbiol., 63, 3104-3110.
Chorus, I., Bartram, J., 1999. Toxic Cyanobacteria in water. A guide to their public health
consequences, monitoring and management. E and Fn Spon, London; New York.
Codd, G.A., Lindsay, J., Young, F.M., Morrison, L.F., Metcalf, J.S., 2005. Harmful cyanobacteria: from
mass mortalities to management measures. In: Huisman, J., Hans, M.C.P.; Visser, P.M. (Eds),
Harmful Cyanobacteria, Springer, pp.1-24.
Costa, I.A.S., Azevedo, S.M.F.O., Senna, P.A.C., Bernardo, R.R., Costa, S. M., Chellappa, N.T., 2006.
Occurrence of toxin-producing cyanobacteria blooms in a brazilian semiarid reservoir. Braz. J. Biol.
66(1B), 211-219.
da Silva, C.A., Oba, E.T., Ramsdorf, W.A., Magalhães, V.F., Cestari, M.M., Ribeiro, C.A.O., Assis,
H.C.S., 2011. First report about saxitoxins in freshwater fish Hoplias malabaricus through trophic
exposure. Toxicon. 57, 141–147.
Dobson, S.H., Holmes, R.M., 1984. Compensatory growth in rainbow trout Salmo gairdneri. Journal of
Fish Biology. 25, 649-656.
Ferrão-Filho, A.S., 2009. Bioacumulação de cianotoxinas e seus efeitos em organismos aquáticos.
Oecologia brasiliensis. 13(2), 272-312.
Figueredo, C.C., A. Giani, 2001. Seasonal variation in the diversity and species richness of
phytoplankton in a tropical eutrophic reservoir. Hydrobiologia. 445, 165-174.

52
Figueredo, C.C., Giani, A., 2005. Ecological interactions between Nile tilapia (Oreochromis niloticus,
L.) and the phytoplanktonic community of the Furnas Reservoir (Brazil). Freshwater Biology. 50,
1391–1403.
Galvão, J.A., Oetterer, M., Bittencourt-Oliveira, M.C., Gouvêa-Barros, S., Hiller, S., Erler, K. Luckas,
B., Pinto, E. Kujbida, P., 2009. Saxitoxins accumulation by freshwater tilapia (Oreochromis niloticus)
for human consumption. Toxicon. 54, 891-894.
Gorham, P.R., McLachlav, J.R., Hammer, V.T., Kim, W.K., 1964. Isolation and culture of toxic strains
of Anabaena flos-aquae (Lyngb.) de Bréb. Verhandlungen der Internationalen Vereinigung für
Theoretische und Angewandte Limnologie. 15, 796-804.
Gustafsson, S., Rengefors, K., Hansson, L.A., 2005. Increased consumer fitness following transfer of
toxin tolerance to offspring via maternal effects. Ecology. 86, 2561–2567.
Hoff-Risseti, C, Dörr, F.A., Schaker, P.D.C., Pinto, E., Werner, V.R,, Fiore, M.F., 2013.
Cylindrospermopsin and saxitoxin synthetase genes in Cylindrospermopsis raciborskii strains from
Brazilian freshwater. PLoS ONE. 8 (8). doi:10.1371/journal.pone.0074238.
Hussain, M.G., 2004. Farming of tilapia: Breeding plans, mass seed production and aquaculture
techniques. 1st.ed. Momin Offset Press, Dhaka, Bangladesh.
Jewel, M.A., Affan, M.A., Khan, S., 2003. Fish mortality due to cyanobacterial bloom in an aquaculture
pond in Bangladesh. Pakistan Journal of Biological Sciences. 6, 1046–1050.
Jobling, M., 2001. Feed composition and analysis. In: Houlihan, D., Boujard, T., Jobling, M. (Eds).
Food intake in fish. Blackwell Science, Malden, MA, USA, pp. 1-24.
Kangur, K., Kangur, A. Kangur, P., Laugaste, R., 2005. Fish kill in Lake Peipsi in summer 2002
as a synergistic effect of a cyanobacterial bloom, high temperature, and low water level. Proc.
Estonian Acad. Sci. Biol. Ecol. 54(1), 67–80.
Lagos, N., Onodera, H., Zagatto, P.A., Andrinolo, D., Azevedo,S.M.F.O, Oshima, Y., 1999. The first
evidence of paralytic shellfish toxins in the freshwater cyanobacterium Cylindrospermopsis
raciborskii, isolated from Brazil. Toxicon, 37, 1359–1373.

53
Landsberg, J.H., Hall, S., Johannessen, J.N., White, K.D., Conrad, S.M., Abbott, J.P., Flewelling,
L.J., Richardson, R.W., Dickey, R.W., Jester, E.L.E., Etheridge, S.M., Deeds, J.R., Van Dolah, F.M.,
Leighfield, T.A., Zou, Y., Beaudry, C.G., Benner, R.A., Rogers, P.L., Scott, P.S., Kawabata, K., Wolny,
J.L., Steidinger, K.A., 2006. Saxitoxin puffer fish poisoning in the United States, with the first report of
Pyrodinium bahamense as the putative toxin source. Environ. Health Perspect. 114, 1502–1507.
Lazzaro, X.,1987. A review of planktivorous fishes: their evolution, feeding behaviours, selectivities,
and impacts. Hydrobiologia, 146, 97–167.
Lefebvre, K.A., Elder, N.E., Hershberger, P.K., Trainer, V.L., Stehr, C.M., Scholz, N.L., 2005.
Dissolved saxitoxin causes transient inhibition of sensorimotor function in larval Pacific herring (Clupea
harengus pallasi). Mar. Biol. 147, 1393–1402.
Lefebvre, K.A. Trainer, V.L., Scholz, N.L., 2004. Morphological abnormalities and sensorimotor deficits
in larval fish exposed to dissolved saxitoxin. Aquatic Toxicology. 66, 159–170.
Lovell, T., 1998. Nutrition and Feeding of Fish. Springer.
Malbrouck, C., Kestemont, P., 2006. Effects of microcystins on fish. Environmental Toxicology and
Chemistry. 25(1), 72–86.
Merel, S. Walker, D., Chicana, R., Snyder, S., Baurès, E., Thomas, O., 2013. State of knowledge and
concerns on cyanobacterial blooms and cyanotoxins. Environment International. 59, 303–327.
Ministério da Pesca e a Aquicultura – Brasil (MPA), 2013. Boletim Estatístico da Pesca e Aquicultura
2011. Brasília: DEMOC, CGMIP, 2013. 60 p.
Molica, R., Onodera, H., García, C., Rivas, M., Andrinolo, D., Nascimento, S., Meguro, H., Oshima, Y.,
Azevedo, S., Lagos, N., 2002. Toxins in the freshwater cyanobacterium Cylindrospermopsis raciborskii
(Cyanophyceae) isolated from Tabocas reservoir in Caruaru, Brazil, including demonstration of a new
saxitoxin analogue. Phycologia. 41, 606–611.
Molica, R.J.R., Oliveira, E.J.A., Carvalho, P.V.V.C., Costa, A.N.S.F., Cunha, M.C.C., Melo, G.L.,
Azevedo, S. M.F.O., 2005. Occurrence of saxitoxins and an anatoxin-a(s)-like anticholinesterase in a
Brazilian drinking water supply. Harmful Algae. 4, 743–753.

54
Moura, A.N., Dantas, E.W., Oliveira, H.S.B., Bittencourt-Oliveira, M.C., 2011. Vertical and temporal
dynamics of cyanobacteria in the Carpina potable water reservoir in Northeastern Brazil. Braz. J. Biol.
71, 451-459.
Moura, A.N., Nascimento, E.C., Dantas, E.W., 2012. Temporal and spatial dynamics of phytoplankton
near farm fish in eutrophic reservoir in Pernambuco, Brazil. Rev. Biol. Trop. 60 (2), 581-597.
Oberemm, A., Becker, J., Codd, G., Steinberg, C., 1999. Effects of cyanobacterial toxins and aqueous
crude extracts on the development of fish and amphibians. Environ. Toxicol. 14, 77–88.
Ohtani, I., Moore, R.E., Runnegar, M.T.C., 1992. Cylindrospermopsin: a potent hepatotoxin from the
blue-green alga Cylindrospermopsis raciborskii. J. Am. Chem. Soc. 114, 7942– 7944.
Oshima, Y., 1995. Postcolumn derivatization liquid chromatographic method for paralytic shellfish
toxins. Journal of AOAC International, [Gaithersburg]. 78(2), 528- 532.
Padisák, J., 1997. Cylindrospermopsis raciborskii (Woloszynnska) Seenayya et Subba Raju, an
expanding, highly adaptive cyanobacterium: worldwide distribution and review of its ecology. Archiv für
Hydrobiology. 107(4), 563-593.
Pomati, F., Neilan, B.A., Suzuki, T., Manarolla, G., Rossetti, C., 2003. Enhancement of intracellular
saxitoxin accumulation by lidocaine hydrochloride in the cyanobacterium Cylindrospermopsis
raciborskii T3 (Nostocales). J. Phycol. 39, 535– 542.
Robineau, B. Fortier, L., Gagné, J. A., Cembella, A.D., 1991. Comparison of the response of five larval
fish species to the toxic dinoflagellate Alexandrium excavatum (Braarud) Balech. Journal of
experimental marine biology and ecology. 152(2), 225-242.
Saker, M.L., Griffiths, D.J., 2000. The effect of temperature on growth and cylindrospermopsin content
of seven isolates of the cyanobacterium Cylindrospermopsis raciborksii (Woloszynska) Seenayya and
Subba Raju from water bodies in northern Australia. Phycologia 39 (4) 349–354.
Sakomura, N.K., Rostagno, H.S., 2007. Métodos de pesquisa em nutrição de monogástricos. Funep,
Jaboticabal.
Sevrin-Reyssac, J., Pletikosic, M., 1990. Cyanobacteria in fish ponds. Aquaculture. 88, l-20.

55
Signor, A.A., Boscolo, W.R., Bittencourt, F., Feiden, A., Gonçalves, G.S., Freitas, J.M.A., 2010.
Desempenho de juvenis de tilápia-do-nilo alimentados com rações contendo complexo enzimático.
Revista Brasileira de Zootecnia. 39(5), 977-983.
Silva de Assis, H.C., da Silva, C.A., Oba, E.T., Pamplona, J.H., Mela, M., Doria, H.B., Guiloski, I.C.,
Ramsdorf, W., Cestari, M.M., 2013. Hematologic and hepatic responses of the freshwater fish Hoplias
malabaricus after saxitoxin exposure. Toxicon. 66, 25–30.
Tavares, J.F.; Proença, L.A.O.; Odebrecht, C., 2009. Assessing the harmful microalgae occurrence
and temporal variation in a coastal aquaculture area, southern Brazil. Atlântica, Rio Grande. 31(2),
129-144.
Turker H., Eversole A.G. & Brune D.E., 2003. Filtration of green algae and cyanobacteria by Nile
Tilapia, Oreochromis niloticus, in the partitioned aquaculture system. Aquaculture, 215, 93–101.
Wiegand, C., Pflugmacher, S., 2005. Ecotoxicological effects of selected cyanobacterial secondary
metabolites a short review. Toxicology and Applied Pharmacology. 203, 201– 218.
Wiegand, C., Pflugmacher, S., Oberemm, A., Meems, N., Beattie, K.A., Steinberg, C.E.W., Codd,
G.A., 1999. Uptake and effects of microcystin-LR on detoxication enzymes of early life stages of the
zebra fish (Danio rerio). Environmental Toxicology. 14(1), 89–95.

56
ANEXO
NORMAS DA REVISTA HARMFUL ALGAE A QUAL SERÁ SUBMETIDO O ARTIGO.
AUDIENCE
. Marine Biologists, Aquaculturalists, Fisheries Biologists, Oceanographers, Ecologists,
Environmental Scientists, Chemists, Food Scientists, and Toxicologists.
IMPACT FACTOR
.2011: 3.083 © Thomson Reuters Journal Citation Reports 2012
GUIDE FOR AUTHORS
.
INTRODUCTION
Types of paper
Original research articles (Regular Articles) Book Reviews Original research articles should report
the results of original research. The material should not have been previously published elsewhere,
except in a preliminary form. Book Reviews will be included in the journal on a range of relevant
books which are not more than 2 years old.
BEFORE YOU BEGIN
Ethics in publishing
For information on Ethics in publishing and Ethical guidelines for journal publication see
http://www.elsevier.com/publishingethics and http://www.elsevier.com/ethicalguidelines.
Ethics in Publishing
The work described in your article must have been carried out in accordance with The Code of Ethics
of the World Medical Association(Declaration of Helsinki) for animal experiments
http://europa.eu.int/scadplus/leg/en/s23000.htm; Uniform Requirements for manuscripts submitted to
Biomedical journals http://www.nejm.org/general/text/requirements/1.htm. This must be stated at an
appropriate point in the article.
Conflict of interest
All authors are requested to disclose any actual or potential conflict of interest including any financial,
personal or other relationships with other people or organizations within three years of beginning the

57
submitted work that could inappropriately influence, or be perceived to influence, their work. See also
http://www.elsevier.com/conflictsofinterest.;
Submission declaration
Submission of an article implies that the work described has not been published previously (except in
the form of an abstract or as part of a published lecture or academic thesis or as an electronic preprint,
see http://www.elsevier.com/postingpolicy), that it is not under consideration for publication
elsewhere, that its publication is approved by all authors and tacitly or explicitly by the responsible
authorities where the work was carried out, and that, if accepted, it will not be published elsewhere
including electronically in the same form, in English or in any other language, without the written
consent of the copyright-holder.
Contributors
Each author is required to declare his or her individual contribution to the article: all authors must
have materially participated in the research and/or article preparation, so roles for all authors should be
described. The statement that all authors have approved the final article should be true and included in
the disclosure.
Changes to authorship
This policy concerns the addition, deletion, or rearrangement of author names in the authorship of
accepted manuscripts: Before the accepted manuscript is published in an online issue: Requests to add
or remove an author, or to rearrange the author names, must be sent to the Journal Manager from the
corresponding author of the accepted manuscript and must include: (a) the reason the name should be
added or removed, or the author names rearranged and (b) written confirmation (e-mail, fax, letter)
from all authors that they agree with the addition, removal or rearrangement. In the case of addition or
removal of authors, this includes confirmation from the author being added or removed. Requests that
are not sent by the corresponding author will be forwarded by the Journal Manager to the
corresponding author, who must follow the procedure as described above. Note that: (1) Journal
Managers will inform the Journal Editors of any such requests and (2) publication of the accepted
manuscript in an online issue is suspended until authorship has been agreed. After the accepted
manuscript is published in an online issue: Any requests to add, delete, or rearrange author names in
an article published in an online issue will follow the same policies as noted above and result in a
corrigendum.
Copyright
Upon acceptance of an article, authors will be asked to complete a 'Journal Publishing Agreement' (for
more information on this and copyright see http://www.elsevier.com/copyright). Acceptance of the

58
agreement will ensure the widest possible dissemination of information. An e-mail will be sent to the
corresponding author confirming receipt of the manuscript together with a 'Journal Publishing
Agreement' form or a link to the online version of this agreement. Subscribers may reproduce tables of
contents or prepare lists of articles including abstracts for internal circulation within their institutions.
Permission of the Publisher is required for resale or distribution outside the institution and for all other
derivative works, including compilations and translations (please consult
http://www.elsevier.com/permissions). If excerpts from other copyrighted works are included, the
author(s) must obtain written permission from the copyright owners and credit the source(s) in the
article. Elsevier has preprinted forms for use by authors in these cases: please consult
http://www.elsevier.com/permissions.
Retained author rights
As an author you (or your employer or institution) retain certain rights; for details you are referred to:
http://www.elsevier.com/authorsrights.
Role of the funding source
You are requested to identify who provided financial support for the conduct of the research and/or
preparation of the article and to briefly describe the role of the sponsor(s), if any, in study design; in
the collection, analysis and interpretation of data; in the writing of the report; and in the decision to
submit the article for publication. If the funding source(s) had no such involvement then this should be
stated. Please see http://www.elsevier.com/funding.
Funding body agreements and policies
Elsevier has established agreements and developed policies to allow authors whose articles appear in
journals published by Elsevier, to comply with potential manuscript archiving requirements as
specified as conditions of their grant awards. To learn more about existing agreements and policies
please visit http://www.elsevier.com/fundingbodies.
Open access
This journal offers you the option of making your article freely available to all via the ScienceDirect
platform. To prevent any conflict of interest, you can only make this choice after receiving notification
that your article has been accepted for publication. The fee of $3,000 excludes taxes and other
potential author fees such as color charges. In some cases, institutions and funding bodies have entered
into agreement with Elsevier to meet these fees on behalf of their authors. Details of these agreements
are available at http://www.elsevier.com/fundingbodies. Authors of accepted articles, who wish to take
advantage of this option, should complete and submit the order form (available at
http://www.elsevier.com/locate/openaccessform.pdf). Whatever access option you choose, you retain

59
many rights as an author, including the right to post a revised personal version of your article on your
own website. More information can be found here: http://www.elsevier.com/authorsrights .
Language Services
Manuscripts should be written in English. Authors who are unsure of correct English usage should
have their manuscript checked by someone proficient in the language. Manuscripts in which the
English is difficult to understand may be returned to the author for revision before scientific review.
Authors who require information about language editing and copyediting services pre- and
postsubmission please visit http://www.elsevier.com/languagepolishing or our customer support site at
http://epsupport.elsevier.com for more information. Please note Elsevier neither endorses nor takes
responsibility for any products, goods or services offered by outside vendors through our services or in
any advertising. For more information please refer to our Terms & Conditions:
http://www.elsevier.com/termsandconditions.
Submission
Submission to this journal proceeds totally online and you will be guided stepwise through the
creation and uploading of your files. The system automatically converts source files to a single PDF
file of the article, which is used in the peer-review process. Please note that even though manuscript
source files are converted to PDF files at submission for the review process, these source files are
needed for further processing after acceptance. All correspondence, including notification of the
Editor's decision and requests for revision, takes place by e-mail removing the need for a paper trail.
Please submit your article via http://ees.elsevier.com/haralg/
Referees
Please submit, with the manuscript, the names, addresses and e-mail addresses of three potential
referees. Note that the editor retains the sole right to decide whether or not the suggested reviewers are
used.
Page charges
Harmful Algae has no page charges.
PREPARATION
Use of wordprocessing software
It is important that the file be saved in the native format of the wordprocessor used. The text should be
in single-column format. Keep the layout of the text as simple as possible. Most formatting codes will
be removed and replaced on processing the article. In particular, do not use the wordprocessor's
options to justify text or to hyphenate words. However, do use bold face, italics, subscripts,

60
superscripts etc. When preparing tables, if you are using a table grid, use only one grid for each
individual table and not a grid for each row. If no grid is used, use tabs, not spaces, to align columns.
The electronic text should be prepared in a way very similar to that of conventional manuscripts (see
also the Guide to Publishing with Elsevier: http://www.elsevier.com/guidepublication). Note that
source files of figures, tables and text graphics will be required whether or not you embed your figures
in the text. See also the section on Electronic artwork. To avoid unnecessary errors you are strongly
advised to use the 'spell-check' and 'grammar-check' functions of your wordprocessor.
LaTeX
If the LaTeX file is suitable, proofs will be produced without rekeying the text. The article should
preferably be written using Elsevier's document class 'elsarticle', or alternatively any of the other
recognized classes and formats supported in Elsevier's electronic submissions system, for further
information see http://www.elsevier.com/wps/find/authorsview.authors/latex-ees-supported. The
Elsevier 'elsarticle' LaTeX style file package (including detailed instructions for LaTeX preparation)
can be obtained from the Quickguide: http://www.elsevier.com/latex. It consists of the file:
elsarticle.cls, complete user documentation for the class file, bibliographic style files in various styles,
and template files for a quick start.
Article Structure
Manuscripts should be typewritten with numbered lines, with wide margins and double spacing
throughout, i.e. also for abstracts, footnotes and references. Every page of the manuscript, including
the title page, references, tables, etc., should be numbered in the upper righthand corner.
However, in the text no reference should be made to page numbers; if necessary, one may refer to
sections. Avoid excessive usage of italics to emphasize part of the text. Subdivision - numbered
sections Divide your article into clearly defined and numbered sections. Subsections should be
numbered 1.1 (then 1.1.1, 1.1.2, ...), 1.2, etc. (the abstract is not included in section numbering). Use
this numbering also for internal cross-referencing: do not just refer to 'the text'. Any subsection may be
given a brief heading. Each heading should appear on its own separate line.
Introduction
State the objectives of the work and provide an adequate background, avoiding a detailed literature
survey or a summary of the results.
Material and methods
Provide sufficient detail to allow the work to be reproduced. Methods already published should be
indicated by a reference: only relevant modifications should be described.

61
Theory/calculation
A Theory section should extend, not repeat, the background to the article already dealt with in the
Introduction and lay the foundation for further work. In contrast, a Calculation section represents a
practical development from a theoretical basis.
Results
Results should be clear and concise.
Discussion
This should explore the significance of the results of the work, not repeat them. A combined Results
and Discussion section is often appropriate. Avoid extensive citations and discussion of published
literature.
Conclusions
The main conclusions of the study may be presented in a short Conclusions section, which may stand
alone or form a subsection of a Discussion or Results and Discussion section.
Essential title page information
• Title. Concise and informative. Titles are often used in information-retrieval systems. Avoid
abbreviations and formulae where possible.
• Author names and affiliations. Where the family name may be ambiguous (e.g., a double name),
please indicate this clearly. Present the authors' affiliation addresses (where the actual work was done)
below the names. Indicate all affiliations with a lower-case superscript letter immediately after the
author's name and in front of the appropriate address. Provide the full postal address of each
affiliation, including the country name and, if available, the e-mail address of each author.
• Corresponding author. Clearly indicate who will handle correspondence at all stages of refereeing
and publication, also post-publication. Ensure that telephone and fax numbers (with country and
area code) are provided in addition to the e-mail address and the complete postal address.
Contact details must be kept up to date by the corresponding author.
• Present/permanent address. If an author has moved since the work described in the article was done,
or was visiting at the time, a 'Present address' (or 'Permanent address') may be indicated as a footnote
to that author's name. The address at which the author actually did the work must be retained as the
main, affiliation address. Superscript Arabic numerals are used for such footnotes.

62
Graphical abstract
A Graphical abstract is optional and should summarize the contents of the article in a concise, pictorial
form designed to capture the attention of a wide readership online. Authors must provide images that
clearly represent the work described in the article. Graphical abstracts should be submitted as a
separate file in the online submission system. Image size: Please provide an image with a minimum of
531 × 1328 pixels (h × w) or proportionally more. The image should be readable at a size of 5 × 13 cm
using a regular screen resolution of 96 dpi. Preferred file types: TIFF, EPS, PDF or MS Office files.
See http://www.elsevier.com/graphicalabstracts for examples. Authors can make use of Elsevier's
Illustration and Enhancement service to ensure the best presentation of their images also in accordance
with all technical requirements: Illustration Service.
Highlights
Highlights are mandatory for this journal. They consist of a short collection of bullet points that
convey the core findings of the article and should be submitted in a separate file in the online
submission system. Please use 'Highlights' in the file name and include 3 to 5 bullet points (maximum
85 characters, including spaces, per bullet point). See http://www.elsevier.com/highlights for
examples.
Keywords
Immediately after the abstract, provide a maximum of 6 keywords, using American spelling and
avoiding general and plural terms and multiple concepts (avoid, for example, 'and', 'of'). Be sparing
with abbreviations: only abbreviations firmly established in the field may be eligible. These keywords
will be used for indexing purposes.
Abbreviations
Define abbreviations that are not standard in this field in a footnote to be placed on the first page of the
article. Such abbreviations that are unavoidable in the abstract must be defined at their first mention
there, as well as in the footnote. Ensure consistency of abbreviations throughout the article.
Acknowledgements
Collate acknowledgements in a separate section at the end of the article before the references and do
not, therefore, include them on the title page, as a footnote to the title or otherwise. List here those
individuals who provided help during the research (e.g., providing language help, writing assistance or
proof reading the article, etc.).

63
Nomenclature and Units
1. Authors and editors are, by general agreement, obliged to accept the rules governing biological
nomenclature, as laid down in the International Code of Botanical Nomenclature, the International
Code of Nomenclature of Bacteria, and the International Code of Zoological Nomenclature. 2. All
biota (crops, plants, insects, birds, mammals, etc.) should be identified by their scientific names when
the English term is used, with the exception of common domestic animals. 3. The first mention of the
scientific names of the species used in the work - in title or text – should be accompanied by the
taxonomic authority unless they can all be referred to a general work in which the authorities are
given. Scientific names of species referred to in other studies need no authority. Generic names should
only be abbreviated when immediately preceded in the text by the mention of the same species or
another of the same genus. 4. All biocides and other organic compounds must be identified by their
Geneva names when first used in text. Active ingredients of all formulations should be likewise
identified. 5. For chemical nomenclature, the conventions of the International Union of Pure and
Applied Chemistry and the official recommendations of the IUPAC-IUB Combined Commission on
Biochemical Nomenclature should be followed. 6. When referring to salinity, please do not use units
i.e. no PSU or 0/00. Please use "a salinity of X, or salinity" instead of adding units.
Database linking
Elsevier encourages authors to connect articles with external databases, giving their readers oneclick
access to relevant databases that help to build a better understanding of the described research. Please
refer to relevant database identifiers using the following format in your article: Database: xxxx (e.g.,
TAIR: AT1G01020; CCDC: 734053; PDB: 1XFN). See http://www.elsevier.com/databaselinking for
more information and a full list of supported databases.
Artwork
Electronic Artwork
•Make sure you use uniform lettering and sizing of your original artwork.
•Save text in illustrations as 'graphics' or enclose the font.
•Only use the following font in your illustrations: Arial.
•Number the illustrations according to their sequence in the text.
•When labelling composite figures, please label as A,B,C, etc. in Arial font, positioned on the upper
left corner, on the panel whenever possible. Please do not include any periods, parentheses, etc.
•Use a logical naming convention for your artwork files.
•Provide captions to illustrations separately.

64
•Produce images near to the desired size of the printed version.
•Submit each figure as a separate file.
•Extra frames and boxes around figures should be eliminated. Please include only X and Y (and Z if
applicable) axes. Background lines on figures should only be included when absolutely necessary.
•Legend material and explanations of symbols, etc. should be on the panel, not hanging off to the side
of the figure. No frame is necessary. If this material does not fit on the panel, it should be included in
the actual figure legend.
•Submitting figures as they are printed from Excel or other spread sheets is not acceptable formatting
for publication. A detailed guide on electronic artwork is available on our website:
http://www.elsevier.com/artworkinstructions
You are urged to visit this site; some excerpts from the detailed information are given here.
Formats
Regardless of the application used, when your electronic artwork is finalised, please 'save as' or
convert the images to one of the following formats (note the resolution requirements for line drawings,
halftones, and line/halftone combinations given below):
EPS: Vector drawings. Embed the font or save the text as 'graphics'.
TIFF: Color or grayscale photographs (halftones): always use a minimum of 300 dpi.
TIFF: Bitmapped line drawings: use a minimum of 1000 dpi.
TIFF: Combinations bitmapped line/half-tone (color or grayscale): a minimum of 500 dpi is required.
If your electronic artwork is created in a Microsoft Office application (Word, PowerPoint, Excel) then
please supply 'as is'.Please do not:
•Supply files that are optimised for screen use (e.g., GIF, BMP, PICT, WPG); the resolution is too
low;
•Supply files that are too low in resolution;
• Submit graphics that are disproportionately large for the content.
Color Artwork
Please make sure that artwork files are in an acceptable format (TIFF, EPS or MS Office files) and
with the correct resolution. If, together with your accepted article, you submit usable color figures then
Elsevier will ensure, at no additional charge, that these figures will appear in color on the Web (e.g.,
ScienceDirect and other sites) regardless of whether or not these illustrations are reproduced in color
in the printed version. For color reproduction in print, you will receive information regarding the
costs from Elsevier after receipt of your accepted article. Please indicate your preference for color:
in print or on the Web only. For further information on the preparation of electronic artwork, please
see http://www.elsevier.com/artworkinstructions. Please note: Because of technical complications

65
which can arise by converting color figures to 'gray scale' (for the printed version should you not opt
for color in print) please submit in addition highresolution black and white versions of all the color
illustrations. Simply printing color as black and white is not acceptable.
Figure captions
Ensure that each illustration has a caption. Supply captions separately, not attached to the figure. A
caption should comprise a brief title (not on the figure itself) and a description of the illustration. Keep
text in the illustrations themselves to a minimum but explain all symbols and abbreviations used.
Tables
1. Authors should take notice of the limitations set by the size and lay-out of the journal. Large tables
should be avoided. Reversing columns and rows will often reduce the dimensions of a table.
2. If many data are to be presented, an attempt should be made to divide them over two or more tables.
3.Tables should be numbered according to their sequence in the text. The text should include
references to all tables.
4. Each table should be typewritten on a separate page of the manuscript. Tables should never be
included in the text.
5. Each table should have a brief and self-explanatory title.
6. Column headings should be brief, but sufficiently explanatory. Standard abbreviations of units of
measurements should be added between parentheses.
7. Vertical lines should not be used to separate columns. Leave some extra space between the columns
instead.
8. Any explanation essential to the understanding of the table should be given as a footnote at the
bottom of the table.
9. When preparing tables, if you are using a table grid, use only one grid for each individual table and
not a grid for each row. If no grid is used, use tabs, not spaces, to align columns.
References
1. All publications cited in the text should be presented in a list of references following the text of the
manuscript. The manuscript should be carefully checked to ensure that the spelling of author's names
and dates are exactly the same in the text as in the reference list.
2. In the text refer to the author's name (without initial) and year of publication, followed - if necessary
- by a short reference to appropriate pages. Examples: "Since Peterson (1993) has shown that. . ."
"This is in agreement with results obtained later (Kramer, 1993, pp. 12-16)".

66
3. When reference is made to a work by two authors, both names should be given using "and". If
reference is made in the text to a publication written by more than two authors, the name of the first
author should be used followed by "et al.". This indication, however, should never be used in the list
of references. In this list names of first author and co-authors should be mentioned.
4. References cited together in the text should be arranged chronologically. The list of references
should be arranged alphabetically on author's names, and chronologically per author. If an author's
name in the list is also mentioned with co-authors the following order should be used: publications of
the single author, arranged according to publication dates - publications of the same author with one
co-author - publications of the author with more than one co-author. Publications by the same
author(s) in the same year should be listed as 1993a, 1993b, etc. For Volume (Vol.) Bulletin (Bull.),
and No., Arabic numerals should be used (not underlined); the full number of pages should be given in
the form of pp. 123-128.
5. Use the following system for arranging your references:
a. For periodicals
Jones, H.D., Richards, O.G., Southern, T.A., 1992. Gill dimensions, water pumping and body size in
the mussel Mytilus edulis I. J. Exp. Mar. Biol. Ecol. 155(2), 213-237.
b. For books Clark, R.B., 1992. Marine pollution, 3rd ed. Clarendon Press, Oxford.
c. For multi-author books
Hawkins, A.J.S., Baynes, B.L., 1992. Physiological processes, and the regulation of production. In:
Gosling, E. (Ed.), The mussel Mytilus: ecology, physiology, genetics and culture. Elsevier Publishers
B.V., Amsterdam, pp. 171-222.
6. The name of the journal should be abbreviated according to the International List of Periodical Title
Word Abbreviations, published by the International Serials Data Systems; Paris, France.
7. In the case of publications in any language other than English, the original title is to be retained.
However, the titles of publications in non-Latin alphabets should be transliterated, and a notation such
as "(in Russian)" or "(in Greek, with English abstract)" should be added.
8. Work accepted for publication but not yet published should be referred to as "in press".
9. References concerning unpublished data and "personal communications" should not be cited in the
reference list but may be mentioned in the text.
Reference management software
This journal has standard templates available in key reference management packages EndNote
(http://www.endnote.com/support/enstyles.asp) and Reference Manager
(http://refman.com/support/rmstyles.asp). Using plug-ins to wordprocessing packages, authors only
need to select the appropriate journal template when preparing their article and the list of references
and citations to these will be formatted according to the journal style which is described below.

67
Video data
Elsevier accepts video material and animation sequences to support and enhance your scientific
research. Authors who have video or animation files that they wish to submit with their article are
strongly encouraged to include these within the body of the article. This can be done in the same way
as a figure or table by referring to the video or animation content and noting in the body text where it
should be placed. All submitted files should be properly labeled so that they directly relate to the video
file's content. In order to ensure that your video or animation material is directly usable, please provide
the files in one of our recommended file formats with a preferred maximum size of 50 MB. Video and
animation files supplied will be published online in the electronic version of your article in Elsevier
Web products, including ScienceDirect: http://www.sciencedirect.com. Please supply 'stills' with your
files: you can choose any frame from the video or animation or make a separate image. These will be
used instead of standard icons and will personalize the link to your video data. For more detailed
instructions please visit our video instruction pages at http://www.elsevier.com/artworkinstructions.
Note: since video and animation cannot be embedded in the print version of the journal, please provide
text for both the electronic and the print version for the portions of the article that refer to this content.
Supplementary data
Elsevier accepts electronic supplementary material to support and enhance your scientific research.
Supplementary files offer the author additional possibilities to publish supporting applications,
highresolution images, background datasets, sound clips and more. Supplementary files supplied will
be published online alongside the electronic version of your article in Elsevier Web products,
including ScienceDirect: http://www.sciencedirect.com. In order to ensure that your submitted
material is directly usable, please provide the data in one of our recommended file formats. Authors
should submit the material in electronic format together with the article and supply a concise and
descriptive caption for each file. For more detailed instructions please visit our artwork instruction
pages at http://www.elsevier.com/artworkinstructions.
Data at PANGAEA
Electronic archiving of supplementary data enables readers to replicate, verify and build upon the
conclusions published in your paper. We recommend that data should be deposited in the data library
PANGAEA (http://www.pangaea.de). Data are quality controlled and archived by an editor in
standard machine-readable formats and are available via Open Access. After processing, the author
receives an identifier (DOI) linking to the supplements for checking. As your data sets will be citable
you might want to refer to them in your article. In any case, data supplements and the article will be
automatically linked as in the following example: doi:10.1016/0016-7037(95)00105-9. Please use
PANGAEA's web interface to submit your data (http://www.pangaea.de/submit/).

68
Submission checklist
The following list will be useful during the final checking of an article prior to sending it to the journal
for review. Please consult this Guide for Authors for further details of any item.
Ensure that the following items are present:
One author has been designated as the corresponding author with contact details:
• E-mail address
• Full postal address
• Telephone and fax numbers
All necessary files have been uploaded, and contain:
• Keywords
• All figure captions
• All tables (including title, description, footnotes)
Further considerations
• Manuscript has been 'spell-checked' and 'grammar-checked'
• References are in the correct format for this journal
• All references mentioned in the Reference list are cited in the text, and vice versa
• Permission has been obtained for use of copyrighted material from other sources (including the Web)
• Color figures are clearly marked as being intended for color reproduction on the Web (free of
charge) and in print, or to be reproduced in color on the Web (free of charge) and in black-and-white
in print
• If only color on the Web is required, black-and-white versions of the figures are also supplied for
printing purposes
For any further information please visit our customer support site at http://support.elsevier.com.
Top Related