






Languages
Pages
Legal


Meniscus Tissue Engineering:
histologische analyse van In
Vivo biopten
Selma ATMACA
Verhandeling ingediend tot
het verkrijgen van de graad van
Master in de Biomedische Wetenschappen
Promotor: Prof. Dr. Peter VERDONK
Co-promotor: Dr. Heidi DECLERCQ
Begeleider: Dr. Aad DHOLLANDER
Vakgroep: Fysische Geneeskunde en
Orthopedie
Academiejaar 2012-2013


Meniscus Tissue Engineering:
histologische analyse van In
Vivo biopten
Selma ATMACA
Verhandeling ingediend tot
het verkrijgen van de graad van
Master in de Biomedische Wetenschappen
Promotor: Prof. Dr. Peter VERDONK
Co-promotor: Dr. Heidi DECLERCQ
Begeleider: Dr. Aad DHOLLANDER
Vakgroep: Fysische Geneeskunde en
Orthopedie
Academiejaar 2012-2013

Toelating tot bruikleen
“De auteur en de promotor geven de toelating deze masterproef voor consultatie
beschikbaar te stellen en delen ervan te kopiëren voor persoonlijk gebruik. Elk ander
gebruik valt onder de beperkingen van het auteursrecht, in het bijzonder met
betrekking tot de verplichting uitdrukkelijk de bron te vermelden bij het aanhalen van
resultaten uit deze masterproef.”
Datum: 17/05/2013
Selma Atmaca Dr. Heidi Declercq Prof. Dr. Peter Verdonk

Voorwoord
Voor het neerschijven van mijn voorwoord heb ik niet lang moeten nadenken over de mensen
die ik wilde bedanken. De hulp en de ondersteuning die ik kreeg van verschillende mensen
zorgde dat ik mijn einddoel kon behalen voor mijn masterproef.
Ik realiseerde mij dat mijn masterproef het einde van mijn studentenleven betekent. Er staat
mij een nieuwe uitdaging te wachten op het werkveld. Ik besefte dat dit de laatste beproeving
zou zijn van spanning en stress voor het behalen de eindstreep, hopelijk.
In de eerste plaats wil ik mijn promotor Prof. Dr. Peter Verdonk en mijn co-promotor Dr.
Heidi Declercq bedanken om mijn thesis keer op keer na te lezen. Mijn begeleider Dr. Aad
Dhollander wil ook bedanken voor zijn hulp.
Vervolgens wil ik Prof. Dr. Ria Cornelissen bedanken voor het ter beschikking stellen van
haar dienst.
Voor het uitlenen van het patiëntmateriaal wil ik Prof. Dr. Louis Libbrecht en de dienst
Pathologische Anatomie bedanken. Mijn dank gaat ook naar de medewerkers van de dienst
Fysische Geneeskunde en Orthopedie.
Dokter Arne Burssens wil ik ook bedanken, specifiek voor zijn hulp bij het opzoeken en
selecteren van de patiënten.
Laborante Leen Pieters en haar collega’s dank ik voor het gezellig verloop van de saaie
wachttijden bij de immuunkleuringen.
De medewerkers van de dienst Histologie waarmee ik rechtstreeks en onrechtstreeks in
contact was, wil ik ook bedanken. Mijn labopartners en medestudenten, Laure en Hanna, wil
ik ook bedanken voor de toffe momenten in het teaching lab.
Tot slot wil ik mijn ouders, broer, zussen, neven, nichten en vriendinnen bedanken voor de
emotionele ondersteuning doorheen mijn eindverhandeling.
Selma Atmaca

Lijst met afkortingen
BS: blokkingsserum
BSA: bovine serum albumin
CD-RAP: cartilage-derived retinoic acid-sensitive protein
COMP: cartilage oligomeric membrane protein
CS: chondroitinesulfaat
DMSO: dimethylsulfoxide
ECM: extracellulaire matrix
FCS: fetal calf serum
HE: haematoxyline-eosine
IGD: interglobulair domein
IHC: immunohistochemie
KS: keratansulfaat
LSAB: labeled-streptavidin-biotin
MIA: melanoma inhibitory activity
MSCs: mesenchymale stamcellen
NGS: normal goat serum
NRS: normal rabbit serum
NSS: normal swine serum
OA: osteoartritis
PBS: phosphate buffered saline
PCL: polycaprolacton
PSACH: pseudoachondroplasie
TB: tris-buffer
TGFβ: transforming growth factor β
TSP 5: thrombospondine 5
VB: verdunningsbuffer
α-SMA: α-smooth muscle actin

Inhoudsopgave
Samenvatting ................................................................................................................................. 1
1. Inleiding ................................................................................................................................. 2
1.1 Anatomie en functie van de meniscus...............................................................................2
1.2 Histologische opbouw van de meniscus ...........................................................................3
1.3 Extracellulaire matrix ......................................................................................................4
1.4 Meniscusscheuren en meniscectomie ...............................................................................5
1.5 Complicaties geassocieerd met meniscectomie .................................................................6
1.6 Strategieën om blijvende schade bij meniscectomie te herstellen ......................................6
1.6.1 Viabele allogreffes .................................................................................................. 7
1.6.2 Gevroren allogreffe ................................................................................................. 7
1.6.3 Meniscus tissue engineering .................................................................................... 8
1.7 Stamcellen .......................................................................................................................9
1.7.1 Stamcelmerkers ....................................................................................................... 9
1.7.2 Merker voor celproliferatie .....................................................................................11
1.7.3 Merkers in (nieuw) gevormd weefsel......................................................................11
1.7.2.1 Merkers van chondrocyten ................................................................................11
1.8 Doel van de masterproef ................................................................................................ 13
2. Materialen en methoden ........................................................................................................14
2.1 Snijden van de weefsels ................................................................................................. 14
2.2 Histologische analyses ................................................................................................... 14
2.3.1 Deparaffineren .......................................................................................................14
2.3.2 Haematoxyline-eosine kleuring ..............................................................................14
2.3.3 Antigen retrieval ....................................................................................................15
2.3.4 Immunohistochemische kleuring ............................................................................15
2.3.4.1 Geselecteerde merkers ......................................................................................15
2.3.5 Positieve controle voor de verschillende merkers ....................................................18
2.3.6 Picrosirius rood ......................................................................................................19
2.3.7 Dehydrateren .........................................................................................................19

2.3.8 Afdekken met Mounting medium (Richard Allan, Thermo scientific) .....................20
2.3.9 Evaluatie van de preparaten ....................................................................................20
2.3.10 Bepalen van de celdensiteit en het tellen van het aantal vasculaire structuren ..........20
2.3.11 Scores ....................................................................................................................20
2.3.12 Statistiek ................................................................................................................21
2.3 Celcultuur ...................................................................................................................... 21
2.4.1 MG63 cultuurmedium ............................................................................................21
2.4.2 Cellen ontdooien ....................................................................................................22
2.4.3 Cellen tellen ...........................................................................................................22
2.4.4 Cellen splitsen - trypsiniseren .................................................................................22
2.4.5 Cellen invriezen .....................................................................................................22
2.4.6 Celpellet fixeren en inbedden in paraffine ..............................................................23
3. Resultaten .............................................................................................................................24
3.1 Immuunkleuringen op positieve controleweefsels .......................................................... 24
3.1.1 Anti-CD34 immuunkleuring ...................................................................................24
3.1.2 Anti-SMA immuunkleuring ...................................................................................25
3.1.3 Anti-aggrecan immuunkleuring ..............................................................................26
3.1.4 Anti-Ki67 immuunkleuring ....................................................................................26
3.1.5 Anti-Stro1 immuunkleuring ...................................................................................27
3.1.6 Anti-S100 immuunkleuring ....................................................................................28
3.1.7 Collageen type II ....................................................................................................28
3.2 Algemene morfologie van de biopten ............................................................................. 29
3.3 Specifieke kleuringen op de biopten ............................................................................... 31
3.3.1 Orteq scaffold ........................................................................................................31
3.3.2 Deepfrozen allogreffe .............................................................................................34
3.3.3 Viabele allogreffe...................................................................................................36
3.4 Vergelijking van de 3 populaties .................................................................................... 38
4. Bespreking ............................................................................................................................41
5. Algemeen besluit ...................................................................................................................46

6. Referenties ............................................................................................................................48
Bijlagen ............................................................................................................................................. 1
Bijlage I: Deparaffineren ................................................................................................................ 1
Bijlage II: Haematoxyline-eosine kleuring...................................................................................... 2
Bijlage III: Voorbehandeling met citraatbuffer (microgolf) ............................................................. 3
Bijlage IV: Voorbehandeling met citraatbuffer (steamer, Braun) .................................................... 3
Bijlage V: Voorbehandeling met citraatbuffer (combinatie microgolf en steamer) .......................... 4
Bijlage VI: Voorbehandeling met citraatbuffer (warmwaterbad) ..................................................... 4
Bijlage VII: Protocol anti-collageen type II immuunkleuring .......................................................... 5
Bijlage VIII: pepsine (Sigma) 1mg/ml Tris pH2 ............................................................................. 6
Bijlage IX: Protocol anti-aggrecan (AbD Serotec, MCA 4722) (protocol Prof. A. Campos,
Universiteit van Granada) .............................................................................................................. 6
Bijlage X: Chondroitinase ABC (Sigma, C3667) 0,2U/ml in oplossing A ....................................... 7
Bijlage XI: Protocol anti-SMA immuunkleuring (monoclonal antibody, Dako, HHF35) ................. 8
Bijlage XII: Protocol anti-S100 immuunkleuring (polyclonal antibody, Dako, Z0311) .................... 9
Bijlage XIII: Protocol anti-Ki67 immuunkleuring ..........................................................................10
Bijlage XIV: Protocol anti-Stro1 immuunkleuring .........................................................................11
Bijlage XV: Protocol anti-CD34 immuunkleuring .........................................................................12
Bijlage XVI: protocol picrosirius rood (protocol Tissue engineering group, Universiteit van
Granada, Prof. A. Campos) ...........................................................................................................13
Bijlage XVII: Dehydrateren ..........................................................................................................14
Bijlage XVIII: Stijgende alcoholreeks ...........................................................................................14
Bijlage XIX: Tabel 3: Voorstelling van de toegekende scores voor de verschillende kleuringen,
weergegeven per populatie. ...........................................................................................................15

1
Samenvatting
Inleiding: Meniscusscheuren komen met een incidentie van 61 op 100 000 voor in de wereld.
Een scheur, ter hoogte van de witte zone van de meniscus, leidt tot het gedeeltelijk of volledig
verliezen van de meniscus. Om het ontstaan van een degeneratieve aandoening aan het
kniegewricht te voorkomen, worden er verschillende strategieën toegepast. Een van de meest
toegepaste strategieën is het implanteren van deepfrozen of viabele allogreffes. Hoewel deze
strategie goede resultaten geeft, wordt het minder toegepast. De ontwikkeling van tissue
engineering leidde tot het ontwikkelen van een synthetisch meniscus implant (Orteq). Deze
scaffold bezit 80% porositeit en laat een goede weefselingroei toe. Het doel van deze
masterproef is om aan de hand van histologische analyse van in vivo biopten de graad van
regeneratie tussen de 3 populaties te bepalen.
Materiaal en methoden: De geselecteerde antilichamen voor de immunohistochemische
analyse waren: anti-aggrecan, anti-smooth muscle actin (SMA), anti-CD34 en anti-collageen
type II. Voor de algemene morfologie werd de haematoxyline-eosine (HE) kleuring
gehanteerd. De verschillende types collageen werden geëvalueerd met de picrosirius red
kleuring.
Resultaten: Er zijn duidelijk waarneembare morfologische verschillen tussen de drie
populaties op de HE gekleurde coupes. De gemiddelde celdensiteit is verschillend tussen de 3
populaties. De diepe zone wordt bij de populatie met een viabel transplantaat en een Orteq
implantaat beter gekoloniseerd met cellen dan een deepfrozen transplantaat. Uitgerekte
structuren, mogelijks progenitorcellen, zijn CD34+. De populatie met een deepfrozen
transplantaat heeft de zwakste intensiteit in CD34 kleuring, in vergelijking met de twee
andere populaties (onderling geen verschil in intensiteit tussen viabele allogreffe en Orteq).
De biopten met een Orteq implantaat en een viabel allogreffe transplantaat hebben een hoger
aantal bloedvaten dan de biopten met een deepfrozen allogreffe transplantaat. Met de SMA
kleuring konden geen myofibroblastachtige celtypes worden aangetoond. De overwegend
rode kleuring op de picrosirius red kleuring wijst op een hoger collageen type I gehalte.
Besluit: De biopten van de patiënten met een Orteq implantaat en een viabel allogreffe transplantaat,
geven allebei betere resultaten dan de biopten van deepfrozen allogreffes.

2
1. Inleiding
De laatste jaren wordt er veel onderzoek verricht over meniscus tissue engineering. Hierbij
wordt er getracht om te voorkomen dat er blijvende schade zou ontstaan na het gedeeltelijk of
totaal verliezen van de meniscus. Het gedeeltelijk of totaal verliezen van de meniscus leidt
immers op termijn tot het ontstaan van degeneratieve aandoeningen van het kniegewricht
zoals osteoartritis [1, 2, 3].
In de praktijk wordt al een synthetisch construct, Orteq, geïmplanteerd in patiënten [2, 4]. Dit
zou een alternatief kunnen vormen voor de viabele of deepfrozen transplantaten, die eerder
werden toegepast bij patiënten met een meniscaal verlies [4]. Deze drie populaties zullen aan
de hand van histologie onderzocht worden op de graad van regeneratie.
1.1 Anatomie en functie van de meniscus
De menisci zijn halve maan vormige structuren (Figuur 1), die lateraal en mediaal gesitueerd
zijn ter hoogte van het kniegewricht [1, 5, 6]. Deze fibreuze kraakbeenachtige structuren staan
in voor het pijnloos functioneren van het kniegewricht. De menisci zijn voornamelijk
opgebouwd uit water (70 %) en andere componenten zijnde: collageen types I, II, III, V en VI
en proteoglycanen [5].
De primaire functie van de menisci is het efficiënt verdelen van het gewicht en de krachten
die inwerken op de knie [1, 2, 5, 6, 7]. De spreiding van het synoviaal vocht staat in voor de
smering tussen condylus femoris en het tibiaplateau. De avasculaire zones van de meniscus
worden voorzien van voedingstoffen door de diffusie van het synoviaal vocht [5].
Figuur 1: Links: anterieure kijk op het kniegewricht. De meniscus situeert zich tussen de tibia en de femur.
Rechts: superieure kijk op het kniegewricht [8].

3
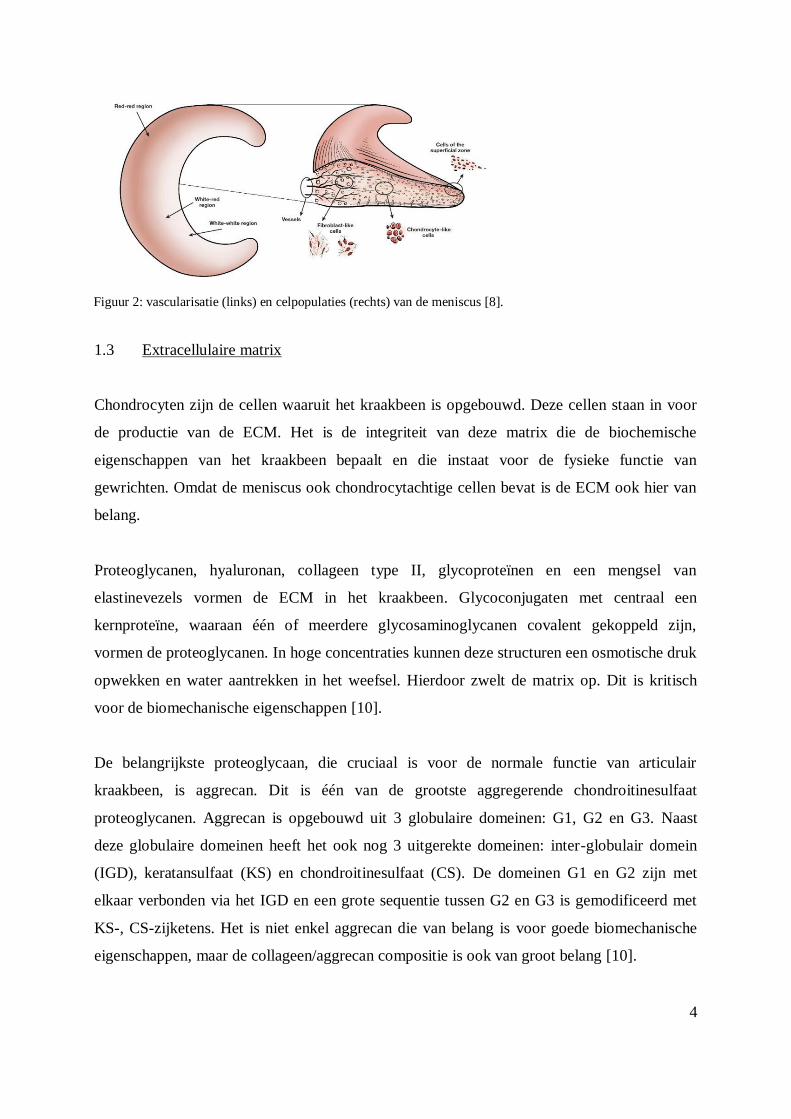
1.2 Histologische opbouw van de meniscus
Histologisch zijn de menisci opgebouwd uit een binnenste avasculaire of witte zone en een
buitenste vasculaire of rode zone (Figuur 2). De cellen van de meniscus, de doorbloeding en
het helingsproces variëren naargelang de zones van de meniscus [1, 2]. Histologie toont ook
aan dat er drie celpopulaties kunnen onderscheiden worden: fibrochondrocyten,
fibroblastachtige cellen en superficiële zone cellen [2]. Verder is er een sterke associatie
tussen de doorbloeding en het helingproces; letsels die ontstaan in de vasculaire zone bezitten
een spontaan regeneratievermogen. Maar de letsels ter hoogte van de avasculaire zone
(binnenste zone) herstellen moeilijk of niet. Zo is er ook weinig geweten welke cellen
verantwoordelijk zijn voor het herstel in de avasculaire zone [2]. In de doorbloede perifere
zone wordt er op de plaats van de schade een fibrineklonter gevormd. Deze fibrineklonter
dient als het ware als een tijdelijk substraat (scaffold) voor het herstelproces. Het zendt
chemotactische en mitogene stimuli uit om de cellen, die deel uitmaken van het herstelproces,
aan te trekken naar de plaats met schade [5].
De ronde tot ovale matrix-geassocieerde cellen, die het humaan meniscusweefsel vormen,
kunnen vergeleken worden met chondrocyten en fibroblastachtige cellen [6, 9]. Deze matrix-
geassocieerde cellen produceren meer collageen type I in tegenstelling tot de articulaire
chondrocyten, die meer collageen type II produceren. Zo is de gemiddelde collageen type
I/type II verhouding in de meniscus 9/1 [6]. Deze verhouding kan wel verschillend zijn
naargelang de zone van de meniscus; de witte zone bestaat bijvoorbeeld voor gemiddeld 67%
uit collageen type I [6]. De verschillende types collageen (type I, II, III, V en VI) en aggrecan,
het meest voorkomende proteoglycaan, vormen de extracellulaire matrix (ECM) van de
meniscus. De ECM van binnenste en middelste zone bevat hoofdzakelijk aggrecan [6, 7]. Het
is deze aggrecan die onder andere instaat voor de visco-elasticiteit en weefselhydratatie [6].
Naast aggrecan komen ook nog andere proteoglycanen (fibromoduline, biglycaan en
decorine) voor in de meniscus [6, 9].
4
1.3 Extracellulaire matrix
Chondrocyten zijn de cellen waaruit het kraakbeen is opgebouwd. Deze cellen staan in voor
de productie van de ECM. Het is de integriteit van deze matrix die de biochemische
eigenschappen van het kraakbeen bepaalt en die instaat voor de fysieke functie van
gewrichten. Omdat de meniscus ook chondrocytachtige cellen bevat is de ECM ook hier van
belang.
Proteoglycanen, hyaluronan, collageen type II, glycoproteïnen en een mengsel van
elastinevezels vormen de ECM in het kraakbeen. Glycoconjugaten met centraal een
kernproteïne, waaraan één of meerdere glycosaminoglycanen covalent gekoppeld zijn,
vormen de proteoglycanen. In hoge concentraties kunnen deze structuren een osmotische druk
opwekken en water aantrekken in het weefsel. Hierdoor zwelt de matrix op. Dit is kritisch
voor de biomechanische eigenschappen [10].
De belangrijkste proteoglycaan, die cruciaal is voor de normale functie van articulair
kraakbeen, is aggrecan. Dit is één van de grootste aggregerende chondroitinesulfaat
proteoglycanen. Aggrecan is opgebouwd uit 3 globulaire domeinen: G1, G2 en G3. Naast
deze globulaire domeinen heeft het ook nog 3 uitgerekte domeinen: inter-globulair domein
(IGD), keratansulfaat (KS) en chondroitinesulfaat (CS). De domeinen G1 en G2 zijn met
elkaar verbonden via het IGD en een grote sequentie tussen G2 en G3 is gemodificeerd met
KS-, CS-zijketens. Het is niet enkel aggrecan die van belang is voor goede biomechanische
eigenschappen, maar de collageen/aggrecan compositie is ook van groot belang [10].
Figuur 2: vascularisatie (links) en celpopulaties (rechts) van de meniscus [8].

5
De grootste onoplosbare, fibreuze proteïne in de ECM en het bindweefsel is collageen. Er zijn
minstens 16 verschillende types van collageen gekend, maar 80 – 90 % van de
collageenvezels, die voorkomen in het lichaam, zijn collageen type I, II of III. De
basisstructuur van collageen wordt gevormd door een tripel helicale structuur, opgebouwd uit
3 α-helices. Daarnaast bevat collageen voornamelijk de aminozuren glycine, proline en
hydroxyproline.
Collageen type I wordt initieel geproduceerd als procollageen in cellen. Dit eiwit bestaat uit
twee α1(I)-procollageen strengen en één α2(I)-procollageen streng. In de huid komt dit type
collageen in grote hoeveelheid voor [11].
Het verschil in de organisatie van de vezels staat in voor het onderscheid tussen verschillende
types van collageen. De fibrillen van collageen type II zijn korter in vergelijking met de
fibrillen van collageen type I. Het is collageen type II dat voornamelijk voorkomt in het
kraakbeen. Deze rigide macromoleculen zijn willekeurig gerangschikt in de visceuze
proteoglycaanmatrix. Hierdoor krijgt de matrix een uitstekende sterkte en samendrukbaarheid.
Deze eigenschap staat in voor de absorptie van shocks in gewrichten [11]. Deze sterkte wordt
nog verbeterd door het collageen/aggrecan netwerk, dat uiterst stijf en resistent is tegen
deformaties [10, 11].
1.4 Meniscusscheuren en meniscectomie
Met een incidentie van gemiddeld 61 op 100 000 per jaar komen meniscusscheuren heel
frequent voor. Sportblessures, die ontstaan als gevolg van het draaien met een gebogen knie
tijdens het voetballen, rugbyen of skiën, zijn de belangrijkste oorzaken van meniscusscheuren
bij jonge patiënten. Maar de meniscus kan degenereren en zijn elasticiteit verliezen bij oudere
patiënten. Hierdoor kunnen de scheuren spontaan ontstaan [12].
Een scheur in de avasculaire zone laat geen spontaan herstel toe [2, 12]. In de jaren 1960 werd
er gedacht dat de enige behandeling van een meniscusscheur een totale meniscectomie was,
zonder enige nadelige consequenties op het kniegewricht [3]. Voor de totale meniscectomie
werd het kniegewricht volledig geopend met een grote incisie [12]. Het duurde enkele
decennia vooraleer er ontdekt werd dat het totaal verwijderen van de meniscus leidt tot

6
ernstige degeneratie van het kniegewricht. Sindsdien werden er verschillende technieken
bestudeerd om de meniscus te sparen en het herstel van de schade te induceren. Als gevolg
van de toenemende populariteit van arthroscopie, werd partiële meniscectomie één van de
meest uitgevoerde chirurgische technieken. Met deze techniek wordt enkel het beschadigd
deel van de meniscus verwijderd [3].
1.5 Complicaties geassocieerd met meniscectomie
Partiële of subtotale meniscectomie uitgevoerd om letsels in de avasculaire zone te
behandelen kunnen op lange termijn aanleiding geven tot degeneratieve veranderingen in het
kniegewricht. Ten gevolge van een hoger stressniveau op het gewrichtskraakbeen kan
osteoartritis (OA) ontstaan [1, 2]. Osteoartritis is de meest gekende vorm van artritis en wordt
gekenmerkt door een gradueel verlies van het kraakbeen [13]. Deze aandoening komt in hoge
prevalentie voor en heeft nadelige effecten op de gezondheid van de patiënt. Hierdoor kan de
patiënt belemmerd worden om de dagelijkse fysieke activiteiten uit te voeren en leiden tot een
daling in de productiviteit van de patiënt. Deze daling in de productiviteit heeft dan op zijn
beurt een impact op de economie [13]. In een studie van Efe et al. werd aangetoond dat het
risico op het ontwikkelen van OA bij patiënten met een mediale meniscectomie significant
hoger ligt [4]. Dus om de stijging van de prevalentie van OA af te remmen moeten er
strategieën onderzocht worden om meniscale schade zo adequaat mogelijk te herstellen.
1.6 Strategieën om blijvende schade bij meniscectomie te herstellen
Een aantal strategieën werden vooropgesteld om patiënten, met blijvende symptomen na het
verlies van de meniscus, te behandelen. Allogene meniscustransplantatie wordt ruim
toegepast in Europa en Noord-Amerika, met goed gerapporteerde klinische resultaten [4]. De
meniscale allogreffes kunnen viabel of diepgevroren zijn [4]. Het transplanteren van
allogreffes houdt verschillende risico’s in. Enerzijds vormen overdracht van infecties en
afstoting een probleem en anderzijds vormt de weefselbeschikbaarheid ook een probleem
[14]. Wanneer er gebruik wordt gemaakt van donorweefsel is het bewerken en het bewaren
van het materiaal van uiterst belang [9]. Aangezien de risico’s geassocieerd met transplantatie
niet weggewerkt kunnen worden, worden transplantaties in sommige centra minder toegepast
[4].

7
De complicaties die bij de verschillende behandelingsvormen (subtotale, totale meniscectomie
en allogreffe transplantatie) de kop opsteken, hebben geleid tot het ontwikkelen van nieuwe
technieken zoals weefselengineering (tissue engineering) [5].
1.6.1 Viabele allogreffes
Viabele, meniscale greffes worden verwijderd van de donor onder steriele omstandigheden.
Tot de implantatie worden deze viabele allogreffes bewaard in een cultuurmedium. Dit
donormateriaal kan maximum 2 weken bewaard worden. Alvorens de allogreffes over te
brengen naar de acceptor moeten die gescreend worden op de aanwezigheid van microbiële
ziekten. Het kan weken tot maanden duren vooraleer de resultaten van de microbiële testen
verkregen worden, dit is langer dan de bewaartijd van de meniscus. Om dit probleem op te
lossen kan er gebruik gemaakt worden van weefselbanken waar het materiaal bevroren wordt
[12].
1.6.2 Gevroren allogreffe
Menisci van verschillende afmetingen kunnen ook bewaard worden in deze weefselbanken.
Dit biedt het voordeel dat er gezocht kan worden naar de best passende meniscus voor de
patiënt Om het materiaal in te vriezen zijn er 2 bewaringstechnieken: cryopreservatie en
versvriezen. Bij de cryopreservatie wordt het materiaal gecontroleerd ingevroren tot -196°C.
Het materiaal wordt in een bad met cryoprotectans van glycerol of dimethylsulfoxide
bewaard. Deze cryoprotectans beschermt de cellen tegen celdood en de vorming van
waterkristallen De cellen die zich het dichtst bij de cryopotectans bevinden blijven viabel. Dit
is afhankelijk van de toegankelijkheid van de cellen voor de cryoprotectans. Het is dit perifere
deel van de meniscus dat het snelst repopuleert na de implantatie in de acceptor [12].
Bij versvriezen (deepfrozen) wordt het donormateriaal diepgevroren bij -80°C, alvorens het
invriezen wordt het materiaal gewassen en verpakt. Met deze methode worden de cellen niet
gespaard. Verschillende methoden zoals chemische decontaminatie of sterilisatie met γ-
stralen worden toegepast alvorens in te vriezen [12].

8
1.6.3 Meniscus tissue engineering
De collaboratie van cel- en moleculaire biologie, cel- en weefselcultuurtechnieken en
materiaalwetenschappen hebben aanleiding gegeven tot de ontwikkeling van een
veelbelovend domein binnen de biomedische wereld en biotechnologie; namelijk tissue
engineering. Binnen dit multidisciplinair onderzoeksdomein ligt de klemtoon op het
ontwikkelen van reconstructieve weefsels. Het zoeken naar alternatieven voor de behandeling
van beschadigde menisci, bracht Arnoczky et al. in 1988 tot tissue engineering binnen de
orthopedie [14]. Tissue engineering omvat het gebruik van scaffolds om defecten op te vullen
en zo het gastheerweefsel te helpen met de regeneratie [4, 14]. De gebruikte scaffolds zijn
artificiële structuren waarop cellen uitgezaaid kunnen worden met al dan niet de
aanwezigheid van groeifactoren. De artificiële structuren kunnen vervolgens geïmplanteerd
worden om de weefselschade te herstellen en zo de driedimensionale weefselvorming
ondersteunen [5].
Het is wenselijk dat de scaffolds bioresorbeerbaar en biodegradeerbaar zijn binnen een
aannemelijke tijdspanne (Figuur 3), zodanig dat de weefselingroei gegarandeerd kan worden
[5]. Een biodegradeerbare, synthetische, acellulaire scaffold, samengesteld met alifatisch
polyurethaan is de ActifitTM
meniscus implant (Orteq) [1, 4]. Deze scaffold bezit 80%
porositeit en laat een goede weefselingroei toe [4].
Figuur 3: Ideale degradatiekinetiek van een resorbeerbare scaffold gerelateerd aan weefselherstel [9].

9
1.7 Stamcellen
Mesenchymale stamcellen (MSCs) zijn multipotente cellen, welke de capaciteit hebben om te
differentiëren tot verschillende mesodermale weefsels, inclusief osteo- en chondrocyten [15].
Het beenmerg is de grootste bron van MSCs, dat gebruikt wordt in weefselengineering.
Wanneer er in vivo een schade optreedt ter hoogte van het gewrichtskraakbeen, die tot in het
subchondraal bot penetreert, wordt er een herstelproces in gang gezet. Tijdens dit
herstelproces migreren cellen vanuit het beenmerg naar de plaats met schade. Op die plaats
met schade ondergaan de cellen chondrogene differentiatie. Maar wanneer de schade
oppervlakkig is en niet tot het subchondraal bot reikt, wordt het herstelproces niet
geactiveerd. Vooral het laatste heeft geleid tot het toenemen van de interesse van
beenmergcomponenten om kraakbeenherstel te ondersteunen. Hoewel er bij een diepe schade,
die zich uitstrekt tot in het subchondraal bot, een herstelproces in gang wordt gezet, wordt er
eerder een fibreus kraakbeen gevormd en geen echte articulair kraakbeen [16].
Naast het beenmerg kunnen MSCs ook geïsoleerd worden uit andere mesenchymale weefsels
zoals vet-, spierweefsel, het synoviaal membraan en het synoviale vocht [17, 18]. Afhankelijk
van de cultuurcondities en cultuurmedium kunnen MSCs in vitro differentiëren in
verschillende weefsels. Zo kunnen MSCs bijvoorbeeld gestuurd worden in de richting van
(fibro)chondrogene differentiatie, mits de toevoeging van dexamethason en TGF-β in het
cultuurmedium [16]. Sinds de ontdekking van de MSCs, begin de jaren 1970, is er een intense
zoektocht naar een betrouwbare merker om MSCs te identificeren en te isoleren [19]. Omdat
er niet één specifieke merker voor MSCs bestaat, worden deze cellen gekarakteriseerd
volgens hun fibroblastachtige morfologie en door de aan- of afwezigheid van verschillende
oppervlaktemerkers [18].
1.7.1 Stamcelmerkers
Ter hoogte van vroeg lymfohematopoietische stam- en progenitorcellen, embryonale
fibroblasten en sommige cellen in foetaal en adult zenuwweefsel komt CD34 tot expressie [6].
CD34 is een stamcelmerker, die vooral ter hoogte van de superficiële zone van de meniscus is
gesitueerd [1, 6]. In de vasculaire en superficiële zone van beschadigde menisci of menisci
van OA-patiënten komen CD34+/CD31
- fibroblastachtige cellen voor [1, 6, 9]. Deze cellen

10
kunnen een mogelijke pool vormen van progenitorcellen verantwoordelijk voor de
vasculogenese, weefselhomeostase en –herstel [1, 6].
Onder pathologische omstandigheden kunnen deze CD34+/CD31
- cellen differentiëren in
myofibroblasten met de vorming van contractiele celfenotypes [1, 9]. Deze contractiele
celfenotypes, die afwezig zijn in de gezonde menisci, worden myofibroblastachtige cellen
genoemd. Deze myofibroblastachtige cellen brengen α-smooth muscle actin (α-SMA) tot
expressie. Declercq et al. bevestigde dat de cellen van de superficiële zone in gezonde
menisci SMA- zijn. De CD34
+ mesenchymale stamcel voorlopers in de superficiële zone
zouden met de juiste stimuli kunnen differentiëren tot MSCs die SMA tot expressie kunnen
brengen [1, 6]. Deze CD34+ en later SMA
+ cellen essentieel zijn voor het intrinsiek
herstelproces van de humane meniscus.
Eén van de inspanningen, die werden geleverd om een geschikte merker te vinden om MSCs
te identificeren en te isoleren, was het gebruikt van CD34+ beenmergcellen. Deze CD34
+
beenmergcellen werden gebruikt als immunogeen en leidden tot de productie van een
monoclonaal antilichaam Stro1 [19]. Initieel werd Stro1 gerapporteerd om stromale
precursoren in het beenmerg te identificeren en er werd aangetoond dat Stro1+ cellen een
multipotentiële capaciteit bezitten om te differentiëren in adipocyten, osteoblasten en
chondrocyten. Hierdoor steeg de interesse in Stro1 als een geschikte merker voor MSCs [20].
Hoewel er nog weinig gekend is over de identiteit van het antigen, dat herkend wordt door
Stro1, wordt het in heel wat studies gebruikt [19].
CD105 of endoglin moduleert de cellulaire respons op TGFβ1 en wordt tot expressie gebracht
op verschillende celtypes: endotheliale cellen, gewrichtschondrocyten en mesenchymale
stamcellen van het beenmerg [6]. CD73 is ook een van de multipele merkers voor MSCs. Dit
eiwit is een goede kandidaat om de chondrogene capaciteit van MSCs te bestuderen [18].
Rolf et al. onderzocht de aanwezigheid van gekende stamcelmerkers CD34, Stro1, CD271 en
CD105. Immunohistochemische kleuring toonde aan dat de cellen in de perivasculaire zone
en het vasculair endotheel Stro1+ zijn. De cellen in de perivasculaire zone zijn ook positief
voor CD271 [21]. Stro1 is veruit de best gekende MSC-merker, die sterk geassocieerd is met
het beenmerg [9]. Verder is CD271 een sleutelmerker van MSCs in het beenmerg. De

11
populatie van MSCs die CD271 tot expressie brengen hebben een hoge capaciteit om te
differentiëren tot adipogene en osteogene cellen [22].
1.7.2 Merker voor celproliferatie
Een eiwit dat sterk geassocieerd wordt met prolifererende cellen is het Ki67 eiwit. Het Ki67
antigen komt tot expressie in alle gezonde en tumorale prolifererende cellen. Dit antigen werd
ontdekt na een gedetailleerde analyse van de volledige celcyclus. Uit deze analyse bleek dat
het Ki67 antigen aanwezig is in de kernen van de cellen in de G1, S en G2 fasen van de
celdeling. Alvorens te kunnen spreken van een efficiënte merker voor celproliferatie, dient er
nagegaan worden of het voldoet aan 2 essentiële criteria. Die criteria houden in dat (a) het
antigen continu aanwezig moet zijn tijdens de celcyclus en (b) dat het antigen snel moet
verdwijnen in cellen die overgaan in een niet-prolifererend stadium. Na het aantonen vanhet
tweede criterium, werd er algemeen aanvaard dat het Ki67 antilichaam een goede merker is
om de fractie van prolifererende cellen te identificeren [23].
1.7.3 Merkers in (nieuw) gevormd weefsel
Bepaalde merkers komen voor in associatie met nieuw gevormde weefsels. Voorbeelden
hiervan zijn de merkers S100, ‘cartilage-derived retinoic acid-sensitive protein’ (CD-RAP) en
‘cartilage oligomeric membrane protein’ (COMP) min of meer specifiek voor (nieuw
gevormde) chondrocyten.
De ECM wordt gekarakteriseerd door de volgende merkers: collageen type I, collageen type
II en aggrecan.
1.7.2.1 Merkers van chondrocyten
Er zijn 21 gekende leden van de S100-familie, welke tot expressie komen ter hoogte van
verschillende weefsels en celtypes. S100 proteïnen zijn calciumbindende eiwitten met een
laag moleculair gewicht en komen voor in vertebraten. Deze eiwitten hebben belangrijke
intra- en extracellulaire functies. Door Stefansson et al. werden de S100 proteïnen voor het
eerst immunohistochemisch geïdentificeerd in humane adulte en foetale chondrocyten. Deze
proteïnen werden zowel in het cytoplasma als in de kernen aangetroffen. Deze merker wordt

12
sterk geassocieerd met de expressie van chondrocyten. In een klein gedeelte van de
verschillende lagen van de gezonde femurkop worden er S100+ cellen aangetroffen. De nieuw
gevormde chondrocyten in de buurt van kraakbeenschade vertonen er een hogere intensiteit
van S100 proteïnen. Deze toename in de intensiteit suggereert dat S100 proteïnen mogelijks
een rol spelen bij het herstel van kraakbeen en dat S100+ cellen vergelijkbaar zijn met
chondrogene progenitorcellen [24].
Het cDNA dat codeert voor een specifiek, eiwit dat mogelijks een belangrijke rol speelt bij
het herstel van kraakbeen, werd geïsoleerd uit twee onafhankelijke bronnen: melanoma
cellijnen en chondrocyten. Het eiwit dat geïsoleerd werd uit melanoma cellijnen wordt
‘Melanoma Inhibitory Activity’ (MIA) genoemd en het eiwit geïsoleerd uit chondrocyten en
kraakbeenachtige weefsels wordt ‘Cartilage-derived retinoic acid-sensitive protein’ (CD-
RAP) genoemd. In normale omstandigheden wordt CD-RAP/MIA tot expressie gebracht door
chondrocyten en in pathologische omstandigheden wordt het tot expressie gebracht door
melanomen. Het CD-RAP/MIA DNA codeert bij de mens een 131 aminozuren lange eiwit
[25]. De expressie van CD-RAP is gelimiteerd tot het kraakbeen in volwassenen en tijdens de
chondrogenese wordt het ook sterk tot expressie gebracht. Dietz et al. isoleerden CD-
RAP/MIA initieel als een molecule die samen met collageen type II tot co-expressie werd
gebracht [26]. Uit de kraakbeenmatrix wordt dit eiwit vrijgesteld als een respons op
kraakbeenschade. Deze kraakbeenschade kan ontstaan als gevolg van rheumatoïde artritis of
een mechanische overbelasting. Voor de geordende fibrillaire structuur van kraakbeen is CD-
RAP/MIA essentieel, want in afwezigheid van dit eiwit wordt er een sterke deformatie gezien
in de morfologie van de chondrocytaire microvilli. Maar het is niet enkel CD-RAP/MIA die
volstaat om de chondrogene differentiatie van humane MSCs te induceren. De differentiatie
wordt tevens in de chondrogene richting gestuurd door transforming growth factor (TGF)-β3
[27].
Cartilage oligomeric matrix protein/thrombospondine 5 (COMP/TSP5) is een pentameer
ECM-proteïne. Dit eiwit is het vijfde lid van de thrombospondine familie. COMP/TSP5 wordt
overvloedig tot expressie gebracht in de ECM van chondrocyten. Daarnaast wordt het ook
aangetroffen in bot, pezen, ligamenten, synovium en bloedvaten. Uit immunohistochemische
analyses van het kraakbeen is gebleken dat er een spatiaal en temporeel distributiepatroon

13
voor dit eiwit bestaat. In foetaal kraakbeen komt het meer voor in de pericellulaire matrix,
terwijl het meer in de interterritoriale matrix in adult kraakbeen voorkomt [28].
De eiwitten die tot de thrombospondine familie behoren, hebben een typische calcium-
bindende capaciteit. De sterk geconserveerde type 3 herhalingssequentie staat in voor deze
calciumbinding [29]. Chen et al. toonde aan dat de structuur van COMP/TSP5 sterk
afhankelijk is van deze calciumbindingscapaciteit [28]. Ter hoogte van dit type 3 calcium
bindende sequentie kunnen er mutaties optreden, die leiden tot het foutief opvouwen van het
eiwit en het ontstaan van pseudoachondroplasie (PSACH) [28, 29]. Die mutatie is een deletie
van 1 aminozuur (D469del) en wordt aangetroffen bij ongeveer 30% van de PSACH
patiënten. Immunohistochemische data en COMP/TSP5 mutaties in PSACH suggereren dat
dit eiwit belangrijk is in kraakbeen. Dankzij de pentamere structuur van dit eiwit vormt het
tevens een interactie met collageen type I, collageen type IX en aggrecan [28].
1.8 Doel van de masterproef
Momenteel is het behandelen van meniscusscheuren de meest algemene, chirurgische ingreep
toegepast door orthopedische chirurgen. Gemiddeld ondergaan 1 miljoen patiënten in de
Verenigde Staten en 400 000 patiënten in Europa een dergelijke chirurgische ingreep [2].
Meniscectomie en OA worden meestal aan elkaar gelinkt. Frequent hebben de OA-patiënten
een meniscus met een abnormale configuratie. Ook is de kans op het krijgen van OA hoger na
een partiële meniscectomie [1, 7]. Daarom is het belangrijk om de complicaties die op termijn
na een subtotale meniscectomie zouden kunnen optreden te minimaliseren. Het doel van deze
masterproef is om op basis van een histologische analyse tussen de verschillende populaties
(viabele en diepgevroren meniscustransplantaten en acellulaire Orteq scaffolds) het
weefselherstel te achterhalen. Naast de algemene histologie zullen de biopten van de patiënten
geëvalueerd worden op de aan- of afwezigheid van specifieke merkers. Vertrekkende van de
histologische analyses kan de aanwezigheid van de verschillende merkers in de
meniscusimplantaten in rekening gebracht worden met de graad van regeneratie van de
meniscus. Dit zal leiden tot het verwerven van inzicht voor orthopedische chirurgen om de
meest geschikte scaffold te implanteren bij patiënten, zodanig dat de meniscus zo volledig
mogelijk regenereert.

14
2. Materialen en methoden
In deze masterproef zullen biopten van 57 patiënten gebruikt worden voor
immunohistochemische analyse (IHC). Deze patiënten werden geclassificeerd in drie
verschillende categorieën: 20 patiënten met een Orteq scaffold implantaat (12 vrouwen en 8
mannen), 19 patiënten met een deepfrozen implantaat (9 vrouwen en 10 mannen) en 18
patiënten met een viabel implantaat (7 vrouwen en 11 mannen). Deze biopten waren gefixeerd
met formaline en ingebed in paraffine. De blokjes werden bewaard in het archief van de
dienst pathologische anatomie (-1 blok A, Universitair ziekenhuis, De Pintelaan 185, Gent,
diensthoofd: Prof. Dr. M. Praet).
2.1 Snijden van de weefsels
De coupes van de biopten en de MG63 celpellet worden gesneden met een rotatiemicrotoom
op een dikte van 5 µm met een microtoommes (Leica Microsystems).
Op de draagglazen (KP-3056, Klinipath) worden enkele druppels gedistilleerd water gebracht,
vervolgens worden de coupes erop gelegd. De coupes spreiden op een hete plaat (Gerhardt)
en worden vervolgens voor minimum één nacht bewaard in een droogoven (Memmert) bij
37°C.
2.2 Histologische analyses
2.3.1 Deparaffineren
Vooraleer de coupes gekleurd worden, worden deze gedeparaffineerd (bijlage I). Dit gebeurt
volgens een programma in het automatisch kleurtoestel MICROM HMS 740.
2.3.2 Haematoxyline-eosine kleuring
Alvorens een immuunkleuring uit te voeren, dienen alle weefsels/cellen een haematoxyline-
eosine (HE) kleuring te ondergaan. Dit wordt gedaan om de algemene histologie te bekijken.
De kleuring wordt uitgevoerd met het automatisch kleurtoestel MICROM HMS 740.

15
De bakjes voor eosine en haematoxyline worden opgevuld met respectievelijk eosine en
gefilterde haematoxyline. In bijlage II staat het protocol voor de HE-kleuring beschreven.
2.3.3 Antigen retrieval
Tijdens deze masterproef werd er voor de Ki67, CD34 en S100 immunohistochemische
kleuring een antigen retrieval uitgetest. Om te voorkomen dat de coupes zouden loskomen
van het draagglas werden er vier methoden uitgetest. Voor de eerste methode werd
citraatbuffer (pH 6.0) voorverwarmd in de microgolf (bijlage III) en voor de tweede methode
werd citraatbuffer voorverwarmd in de steamer (bijlage IV). Een derde methode was de
combinatie van de eerste twee methoden (bijlage V).
De coupes kwamen in alle drie de gevallen gemakkelijk los van het draagglaasje. Daarom
werd een vierde methode uitgetest (bijlage VI). Deze laatste methode werd gebruikt voor alle
immuunkleuringen waar een antigen retrieval vereist is.
2.3.4 Immunohistochemische kleuring
2.3.4.1 Geselecteerde merkers
Tabel 1: Overzicht van de geselecteerde merkers voor de IHC kleuringen.
Merker 1ste
Antilichaam
(AL1)
2de
Antilichaam
(AL2)
Peroxidase Verdunningen
α-SMA
(alpha smooth
muscle actin)
Mouse anti-
SMA
DAKO HHF35
Biotin-Rabbit
anti-mouse
DAKO P0397
Streptavidin-
HRP
DAKO P0397
AL1: 1/400
AL2: 1/200
Str-HRP: 1/200
S100
(chondrocyt)
Rabbit anti-S100
DAKO Z0311
Biotin-Swine
anti-rabbit
DAKO E0431
Streptavidin-
HRP
Dako P0397
AL1: 1/400-1/2400
AL2: 1/200
Str-HRP: 1/200
Aggrecan
(ECM)
Mouse anti-
aggrecan
AbD Serotec
MCA4722
Anti-mouse
IgG-peroxidase
Vector MP-7402
gebonden aan
AL2
AL1: 1/250
AL2: ready to use

16
Stro1
Mouse anti-
Stro1
R&D Systems
MAB 1038
Biotin-Rabbit
anti-mouse
DAKO E0413
Streptavidin-
HRP
DAKO P0397
AL1: 1/50-1/100
AL2: 1/200
Str-HRP: 1/200
Collagen
type II
(ECM)
Mouse anti-
collagen type II
Acris Antibodies
AM00618PU-N
Biotin-Rabbit
anti-mouse
DAKO E0413
Streptavidin-
HRP
DAKO P0397
AL1: 1/100
AL2: 1/200
Str-HRP: 1/200
Ki67
(proliferatie)
Mouse anti-Ki67
DAKO M7240
Biotin-Rabbit
anti-mouse
DAKO E0413
Streptavidin-
HRP
DAKO P0397
AL1: 1/50
AL2: 1/200
Str-HRP: 1/200
CD34 Mouse anti-
CD34
(QBEnd10)
DAKO M7165
Biotin-Rabbit
anti-mouse
DAKO E0413
Streptavidin-
HRP
DAKO P0397
AL1: 1/100
AL2: 1/200
Str-HRP: 1/200
De gesneden coupes worden immunohistochemisch gekleurd volgens de labeled-Streptavidin-
Biotin (LSAB) methode.
Benodigdheden
Tabel 2: De samenstelling van het blokkingsserum en de verdunningsbuffer voor de IHC
kleuringen.
Blokkingsserum (BS) hoeveelheid Verdunningsbuffer (VB) hoeveelheid
PBS (Phosphate buffered saline) 10 ml PBS 9 ml
5% NRS (normal rabbit serum) 0,5 ml
(DAKO X0902)
5% NGS (normal goat serum) 0,5 ml
(DAKO X0907)
5% NSS (normal swine serum) 0,5 ml
(DAKO X0901)
Blokkingsserum 1 ml

17
(afhankelijk van AL2)
0,2% Tween 20 200 µl
1% BSA* (bovine serum albumin) 0,1g
* 1% BSA wordt als laatste toegevoegd aan het BS, omdat dit tijdens het vortexen schuimt.
Principe
Protocol
Nadat de preparaten gedeparaffineerd zijn, worden de coupes afgegrensd met toluolstift en
kan de immuunkleuring gestart worden.
Het protocol voor de verschillende merkers zijn, afhankelijk van de vereiste voorbehandeling
lichtjes verschillend van elkaar. De collageen type II kleuring (bijlage VII) vereist
respectievelijk een voorbehandeling met pepsine (bijlage VIII). Na deze voorbehandeling
worden de coupes gespoeld met PBS (5 x 2 min). Voor de aggrecan-kleuring (bijlage IX)
dienen de preparaten eerst gedurende 15 minuten gespoeld te worden met PBS (0,1M) bij
37°C. Daarna moeten de coupes gedurende 1h bij 37°C voorbehandeld worden met
Chondroitinase ABC (bijlage X).
Figuur 4: het principe van de labeled-Streptavidin-Biotin (LSAB) methode [30].

18
Hierna zijn de gevolgde protocols voor de SMA- (bijlage XI), S100 (bijlage XII) collageen
type II (bijlage VII), Ki67 (bijlage XIII)-, Stro1- (bijlage XIV) en CD34-kleuring (bijlage
XV) ongeveer hetzelfde, behalve voor aggrecan (bijlage IX).
De coupes worden gedurende 10 minuten voorbehandeld met 3% H2O2 (verdund in PBS).
Gedurende 30 minuten wordt het BS (tabel 2) toegevoegd en vervolgens afgenomen met
cleanex. Het eerste antilichaam wordt verdund in VB (tabel 1) en kan toegevoegd worden
gedurende een incubatietijd van 1h en respectievelijk 2h, voor Stro1 en respectievelijk voor
SMA, collageen type II, CD34 en Ki67. Vervolgens wordt alles 2 keer 5 minuten gespoeld
met PBS. Het secundair antilichaam, verdund in VB (tabel 1), wordt toegevoegd en de coupes
worden gedurende 30 minuten geïncubeerd met dit antilichaam. Alles nogmaals 2 keer
spoelen met PBS. Vervolgens streptavidin-peroxidase, verdund in VB (tabel 1) toevoegen
voor een incubatietijd van 30 minuten. Alle coupes 2 keer 5 minuten spoelen met PBS en nog
een laatste keer 5 minuten spoelen met Trisbuffer (TB). Tijdens de laatste 5 minuten wordt
het chromogeen substraat (5 ml TB; 5 µl H2O2 (0,03%) 0,03 g DAB (0,06%)) bereid. Het
chromogeen substraat wordt toegevoegd voor een incubatietijd van 10 minuten. De preparaten
moeten tijdens deze stap wel afgeschermd worden van licht. Na deze stap worden de
preparaten gedurende 10 minuten gespoeld met stromend leidingwater.
Tot slot wordt alles tegengekleurd met haematoxyline volgens Mayer (1 seconde) en
gedurende 1 minuut afgespoeld met stromend water. De preparaten kunnen hierna
gedehydrateerd en gemonteerd worden.
2.3.5 Positieve controle voor de verschillende merkers
Voor iedere merker is het van belang om een positieve controle mee te nemen in de
immunohistochemische kleuring.
Deze positieve controle kan een stukje weefsel zijn of cellen, die gebruikt worden om de
kleuring voor een specifieke merker op punt te zetten. Voor collageen type I wordt de humane
huid gebruikt als positief controleweefsel, voor collageen type II en aggrecan kan de humane
epiglottis gebruikt worden als positieve controle.

19
Om de Stro1 kleuring op punt te zetten worden MG-63 celpellets (humane osteosarcoma
cellijn) gebruikt.
Om de S100-kleuring op punt te zetten werd gebruik gemaakt van coupes van
varkensembryo’s en de humane epiglottis. De laatste zal verder ook gebruikt worden als
positieve controle bij het uitvoeren van de kleuring op patiëntmateriaal.
De humane buikhuid wordt gebruikt om de CD34-kleuring op punt te zetten en als positief
controleweefsel voor de CD34- en SMA-kleuringen.
2.3.6 Picrosirius rood
Om de verschillende collageenvezels te onderscheiden werd de picrosirius methode gebruikt
(bijlage XVI). Deze kleuring werd uitgevoerd in het laboratorium van Prof. A. Campos
(Universiteit van Granada). De foto’s van deze kleuring werden genomen met een Nikon
Eclipse 90i microscoop met een gepolariseerd en fluorescent systeem. De gebruikte digitale
camera was een Nikon DXM 1200c camera in hetzelfde laboratorium van Prof. A. Campos.
Voor deze kleuring wordt Sirius red F3B gemengd in een oplossing met picrinezuur.
Sirius red F3B is een uitgerekte molecule met zes sulfongroepen. Deze sulfongroepen van de
kleurstof interageren bij een lage pH met de aminogroepen van lysine en hydroxylysine en de
guanidinegroepen van arginine van het basisch eiwit, collageen [31, 32]. Met de
gepolariseerde lichtmicroscoop kan het dubbelbrekingseffect van de collageenvezels en hun
organisatie en distributie geëvalueerd worden [33]. De verschillende types collageen vertonen
een kleurverschil, en –intensiteit in de Picrosirius-polarisatie methode [31]. Zo toonde
Junqueira et al. aan dat de rode kleur correspondeert met collageen type I, de gele kleur met
collageen type II en de groene kleur met collageen type III op de gepolariseerde
lichtmicroscopische opnames van de picrosirius red gekleurde coupes [34].
2.3.7 Dehydrateren
Na de tegenkleuring worden de preparaten gedehydrateerd met het automatisch kleurtoestel
MICROM HMS 740 (bijlage XVII).

20
2.3.8 Afdekken met Mounting medium (Richard Allan, Thermo scientific)
De coupes, die immunohistochemisch gekleurd zijn, worden gemonteerd met Mounting
medium en afgedekt met een dekglaasje (24x50mm, Knittel glass). Zo kunnen de preparaten
bewaard worden voor verdere microscopische analyse.
2.3.9 Evaluatie van de preparaten
De gekleurde coupes worden eerst bekeken met de lichtmicroscoop en vervolgens worden er
foto’s genomen van de gewenste zones met de virtuele microscoop (Olympus, U-CMAD3; U-
TV1-X2).
2.3.10 Bepalen van de celdensiteit en het tellen van het aantal vasculaire structuren
De celdensiteit ter hoogte van de superficiële zone en de diepe zone wordt, per
patiëntenpopulatie, bepaald met foto’s (vergroting: 12,5x genomen met de lichtmicroscoop,
Jenaval) van HE-gekleurde coupes. De superficiële zone wordt van de diepe zone
onderscheiden op basis van de verandering van de weefselstructuur. Er wordt één foto
genomen van de HE-gekleurde coupes van elke patiënt. Op deze foto’s worden met het
programma Photoshop CS6 per coupe at random 6 vierkante zones met een zijde van 100 µm
geselecteerd. Het aantal cellen wordt in iedere zone geteld en vervolgens wordt het
gemiddelde berekend. De gemiddelde celdensiteit werd uitgedrukt in aantal cellen per mm².
Hetzelfde principe wordt toegepast om het aantal vasculaire structuren te tellen op de CD34-
gekleurde coupes.
2.3.11 Scores
Om de celdensiteiten van de superficiële zone en de diepere zone van de biopten, afkomstig
van patiënten met een Orteq implantaat, deepfrozen en viabele allogreffes te vergelijken,
werden verschillende scores gebruikt. Het scoresysteem voor de celdensiteit van de
superficiële zone: score 0 = 0; score 1 = 0 – 500; score 2 = 500 – 1000; score 3 = 1000 –
1500; score 4 = 1500 – 2000 en score 5 = > 2000 cellen/mm². Voor de diepere zone worden

21
de volgende scores toegekend: score 0 = 0; score 1 = 0 – 100; score 2 = 100 – 200; score 3 =
200 – 300; score 4 = 300 – 400 en score 5 = > 400 cellen/mm².
Op de CD34 gekleurde coupes werden het aantal vasculaire structuren geteld. Voor het aantal
vasculaire structuren worden er scores toegekend naargelang het aantal: score 0 = 0; score 1 =
0 – 100: score 2 = 100 – 200; score 3 = 200 – 300; score 4 = 300 – 400 en score 5 = > 400
bloedvaten per mm².
De CD34+ cellen en de aggrecan kleuring werden geëvalueerd op basis van de intensiteit van
de immuunkleuring en er werd gewerkt met het volgend scoresysteem: score 0 = geen
kleuring; score 1 = zwakke kleuring; score 2 = middelmatige kleuring en score 3 = sterke
kleuring.
Tot slot werd voor de analyse van de picrosirius red kleuring (op de gepolariseerde
lichtmicroscopische opnames) het volgende scoresysteem gebruikt: score 0 = zwart; score 1 =
overwegend geel/groen; score 2 = overwegend oranje/rood en score 3 = 50/50 geel/rood.
2.3.12 Statistiek
De statistische verwerking van de resultaten werd met SPSS Statistics 19 uitgevoerd. Met de
Mann Whitney U-test wordt statistische significantie tussen de populaties onderling
geëvalueerd; het significantieniveau wordt ingesteld op p < 0.05.
2.3 Celcultuur
Om de Stro1 en de Ki67 kleuringen op punt te zetten wordt er voor deze masterproef gewerkt
met de humane MG63 osteosarcoma cellijn.
2.4.1 MG63 cultuurmedium
De MG63 osteosarcoma cellen worden in cultuur gebracht in het MG63 medium. Dit medium
bevat DMEM glutamax (Life TechnologiesTM
), 10% FCS en 1% Na-pyruvaat.

22
2.4.2 Cellen ontdooien
Een maatbeker wordt gevuld met steriel water en in de microgolf opgewarmd (37°C).
Vervolgens worden de vials met cellen uit de stikstofvaten gehaald om die langzaam te
ontdooien in het warm waterbad op 37°C. Nadat de cellen ontdooid zijn, worden ze
overgebracht in een proefbuis met voldoende cultuurmedium.
2.4.3 Cellen tellen
De ontdooide cellen worden overgebracht in een proefbuis, waar 10 ml voorverwarmd
cultuurmedium (37°C) wordt toegevoegd. Na het centrifugeren (5 min, 1000 rpm) van de
proefbuis wordt het medium uitgegoten en worden de cellen geresuspendeerd in 3 ml
cultuurmedium. Vervolgens worden de cellen 1/5 verdund in Trypaanblauw (Invitrogen®)
(20 µl celsuspensie, 80 µl Trypaanblauw) om het aantal cellen te tellen in een
Bürkertelkamer.
2.4.4 Cellen splitsen - trypsiniseren
Om de cellen te splitsen wordt het medium verwijderd uit de T75 cultuurfles. Vervolgens
wordt er ongeveer 3 ml PBS toegevoegd om het resterend medium te spoelen. Na het
verwijderen van PBS, wordt 3 ml trypsine toegevoegd. De cultuurfles wordt dan gedurende 3
minuten in de incubator (37°C, 5% CO2) geplaatst. Er wordt gecontroleerd of de cellen
losgekomen zijn en nadien wordt 1 ml FCS toegevoegd. Deze suspensie wordt overgebracht
in een proefbuis om het te centrifugeren (5 min, 1000 rpm). Na het centrifugeren wordt de
supernatans uitgegoten en worden de cellen geresuspendeerd met 5 ml cultuurmedium. Het
celaantal wordt geteld met behulp van een Bürkertelkamer (zie §2.2.3) en het gewenst aantal
cellen wordt verdeeld over verschillende cultuurflessen.
2.4.5 Cellen invriezen
De cellen, die gecentrifugeerd werden, worden geresuspendeerd in het vriesmedium (FCS +
10% DMSO). Vervolgens worden de cellen verdeeld over verschillende vials. Op iedere vial
wordt het type cel, het aantal en de datum genoteerd. Gedurende één uur worden de vials op

23
een vat geplaatst. Dat vat is voor de helft gevuld met vloeibare stikstof en voor de andere met
stikstofdamp. Het verblijf in dit vat laat een geleidelijke daling van de temperatuur toe. Hierna
worden de vials overgebracht in de stikstofvaten.
2.4.6 Celpellet fixeren en inbedden in paraffine
De getrypsiniseerde cellen worden overgebracht in een proefbuis. Na het centrifugeren (5
min, 1500 rpm) wordt het supernatans uitgegoten. Aan de celpellet wordt 1 ml 4% neutraal
gebufferde formol (pH 6.9) toegevoegd. Dit wordt vervolgens bewaard in de koude kamer
(4°C).
Om de celpellet in te bedden in paraffine wordt het behandeld met een stijgende alcoholreeks
(bijlage XVIII). Na de laatste stap (tolueen, 30 min) wordt de celpellet in een paraffine-
cassette gebracht. Na overnacht in de incubator (50°C) kan de paraffine-cassette in de
koelkast (2 h bij 4°C) geplaatst worden. Vervolgens kan het plastiek rond de paraffine
verwijderd worden en op een houten blokje geplaatst worden.

24
3. Resultaten
3.1 Immuunkleuringen op positieve controleweefsels
Voor immunohistochemische kleuringen is het van belang om positieve controleweefsels of
cellen mee te nemen. Deze controleweefsels/cellen dienen om na te gaan of de kleuring
specifiek is. Ook worden deze controleweefsels/cellen gebruikt om immunohistochemische
kleuringen op punt te zetten.
Van de vier methoden met de citraatbuffer voorbehandeling bleven de meniscale coupes het
best op het draagglas met de methode in het warmwaterbad (Bijlage VI). Deze methode werd
dan ook gebruikt voor alle immunohistochemische kleuringen waar een citraatbuffer
voorbehandeling is vereist.
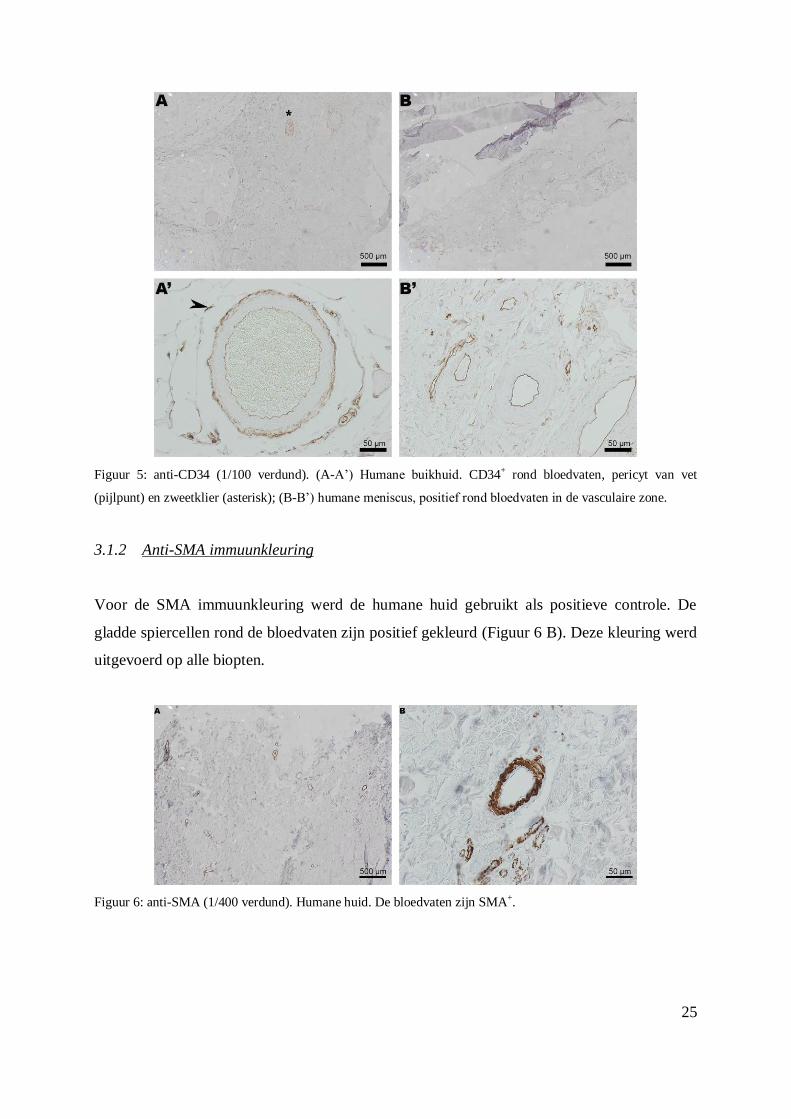
3.1.1 Anti-CD34 immuunkleuring
Om deze kleuring op punt te zetten werd de humane buikhuid gebruikt als positief
controleweefsel. Eveneens werden er coupes van een gezonde meniscus van humane
oorsprong gebruikt. Deze menisci werden bekomen na amputatie van het onderbeen van
diabetespatiënten (Tissue engineering group, Universiteit van Granada, Prof. A. Campos).
Een verdunning van 1/100 werd geselecteerd om de biopten te kleuren. De bloedvaten op de
humane buikhuid, zweetklieren en de pericyten ter hoogte van het vet kleurden positief
(Figuur 5 A – 5 A’). De bloedvaten in de vasculaire zone van de humane meniscus zijn ook
positief voor deze kleuring (Figuur 5 B – 5 B’). Deze kleuring werd uitgevoerd op alle
biopten.
25
Figuur 5: anti-CD34 (1/100 verdund). (A-A’) Humane buikhuid. CD34+ rond bloedvaten, pericyt van vet
(pijlpunt) en zweetklier (asterisk); (B-B’) humane meniscus, positief rond bloedvaten in de vasculaire zone.
3.1.2 Anti-SMA immuunkleuring
Voor de SMA immuunkleuring werd de humane huid gebruikt als positieve controle. De
gladde spiercellen rond de bloedvaten zijn positief gekleurd (Figuur 6 B). Deze kleuring werd
uitgevoerd op alle biopten.
Figuur 6: anti-SMA (1/400 verdund). Humane huid. De bloedvaten zijn SMA+.

26
3.1.3 Anti-aggrecan immuunkleuring
De aggrecan kleuring werd op punt gezet mits gebruik van de humane epiglottis als positief
controleweefsel. De extracellulaire matrix en de zone rond de chondrocyten zijn positief voor
aggrecan (Figuur 7 B).
Figuur 7: anti-aggrecan (1/250 verdund). Humane epiglottis. Positief rond de chondrocyten en in de
extracellulaire matrix.
3.1.4 Anti-Ki67 immuunkleuring
De Ki67 merker is een typische proliferatiemerker. Daarom werden MG63 osteosarcoma
celpellets gekleurd tegen Ki67. Omdat de MG63 in cultuur werden gehouden, waren deze
cellen in een proliferatiefase. De resultaten met de MG63 celpellet waren positief voor de
Ki67 merker (Figuur 8 B). Deze immuunkleuring vereist een voorbehandeling met
citraatbuffer (pH 6.0), die hitte gemedieerd is (bijlage V). Er werd beslist om deze kleuring
niet uit te voeren op de biopten. Want in de meeste gevallen was er een lange tijd verstreken
tussen de datum van implantatie van allogreffe/scaffold en de datum van biopsie. Door deze
lange periode zijn er weinig/geen cellen in de proliferatiefase.

27
Figuur 8: anti-Ki67 (1/50 verdund). MG63 osteosarcoma celpellet.
3.1.5 Anti-Stro1 immuunkleuring
Om de oppervlaktemerker anti-Stro1 immuunkleuring op punt te zetten werd de MG63
celpellet gebruikt. Een verdunning van 1/50 van dit antilichaam gaf de resultaten
weergegeven in figuur 9 A – A’. Omwille van de oriëntatie van de cellen in de celpellet zijn
sommige kernen blauw gekleurd. De randen van deze blauwgekleurde kernen zijn lichtbruin
gekleurd, omdat deze cellen als het ware doormidden werden gesneden met de
rotatiemicrotoom. Cellen die anders georiënteerd waren dan deze cellen vertonen een bruine
kleur aan de volledige oppervlakte, dit wijst op de aanwezigheid van de oppervlaktemerker op
deze cellen.
Deze kleuring werd niet uitgevoerd op de biopten, want het moet nog op punt gezet worden
op humane meniscus biopten.
Figuur 9: anti-Stro1 immuunkleuring op de MG63 celpellet. (A-A’) 1/50 verdund.

28
3.1.6 Anti-S100 immuunkleuring
Om deze immuunkleuring op punt te zetten werden de humane epiglottis en het
varkensembryo (Carnegie stage 20) gebruikt als positieve controleweefsels. De kernen van de
chondrocyten en het cytoplasma zijn positief gekleurd met een verdunning van 1/800 van het
primair antilichaam S100 (Figuur 10 A’, B’). Bij de meniscus zijn de kernen en het
cytoplasma van de fibrochondrocyten positief gekleurd (Figuur 10 C’). Het bindweefsel rond
de epiglottis is ook S100+.
Figuur 10: anti-S100 (1/800 verdund). (A-A’) Humane epiglottis. De kernen van de chondrocyten en het
cytoplasma is positief; (B-B’) varkensembryo, Carnegie stage 20, de wervelkolom die gevormd wordt, is positief
in de kernen en het cytoplasma van chondrocyten; (C-C’) humane meniscus, de fibrochondrocyten zijn positief
in de kernen en het cytoplasma. Cr: chondrocyten, BW: bindweefsel. Pijlpunten: (A’, B’) chondrocyten; (C’)
fibrochondrocyten.
3.1.7 Collageen type II
Voor deze kleuring werd de humane epiglottis, die opgebouwd is uit elastisch kraakbeen,
gebruikt als positief controleweefsel. De verdunning, die werd gebruikt is 1/100. Hoewel de
epiglottis opgebouwd is uit elastisch kraakbeen, is slechts de periferie weinig intens positief
gekleurd met dit antilichaam (Figuur 11 A).

29
Figuur 11: anti-collageen type II (1/100 verdund). Humane epiglottis. Slechts positief ter hoogte van één perifere
zone.
3.2 Algemene morfologie van de biopten
Van iedere patiënt werd één coupe gekleurd met HE om de algemene morfologie en de
celdensiteit te bepalen. Om een algemeen beeld te geven van de resultaten van de HE kleuring
werd at random één patiënt per populatie geselecteerd (Figuur 12 A - F). Op de meeste coupes
zijn er 2 verschillende celtypes waar te nemen. Het eerste celtype zijn de fibrochondrocyten,
die gemakkelijk te onderscheiden zijn op de meeste coupes (Figuur 12 A - C). Daarnaast zijn
er de fibroblastachtige cellen, die gelokaliseerd zijn ter hoogte van de buitenste vasculaire
zone, die soms moeilijker te onderscheiden zijn (Figuur 12 D - F). Deze twee celtypes hebben
duidelijk een verschil in morfologie (Figuur 12 A’ - E’). De fibrochondrocyten hebben eerder
een ronde morfologie met een duidelijke kern met daarrond lacunes (Figuur 12 A’ – C’). De
fibroblastachtige cellen hebben een meer uitgerekte morfologie (Figuur 12 D’ – F’). Ter
hoogte van de superficiële zone komen de fibroblastachtige cellen voor (Figuur 12 D’ – F’).
De morfologie van de cellen wijzigt in de richting van de diepe zone, daar hebben de cellen
een fibrochondrocytaire morfologie (Figuur 12 A’ – C’).

30
Figuur 12: HE-kleuring van de verschillende populaties. x = zone waarop werd ingezoomd. Biopten afkomstig
van patiënten met: (A - A’, D - D’) Orteq scaffold; (B - B’, E - E’) deepfrozen allogreffe; (C - C’, F - F’) viabele
allogreffe. *Fc: fibrochondrocyten. Fb: fibroblastachtige cellen.
31
3.3 Specifieke kleuringen op de biopten
Op alle coupes werden de volgende immuunkleuringen uitgevoerd: SMA, aggrecan, CD34 en
picrosirius red (Figuur 14 - 18). De foto’s van de verschillende immuunkleuringen werden
genomen met de virtuele microscoop (Olympus).
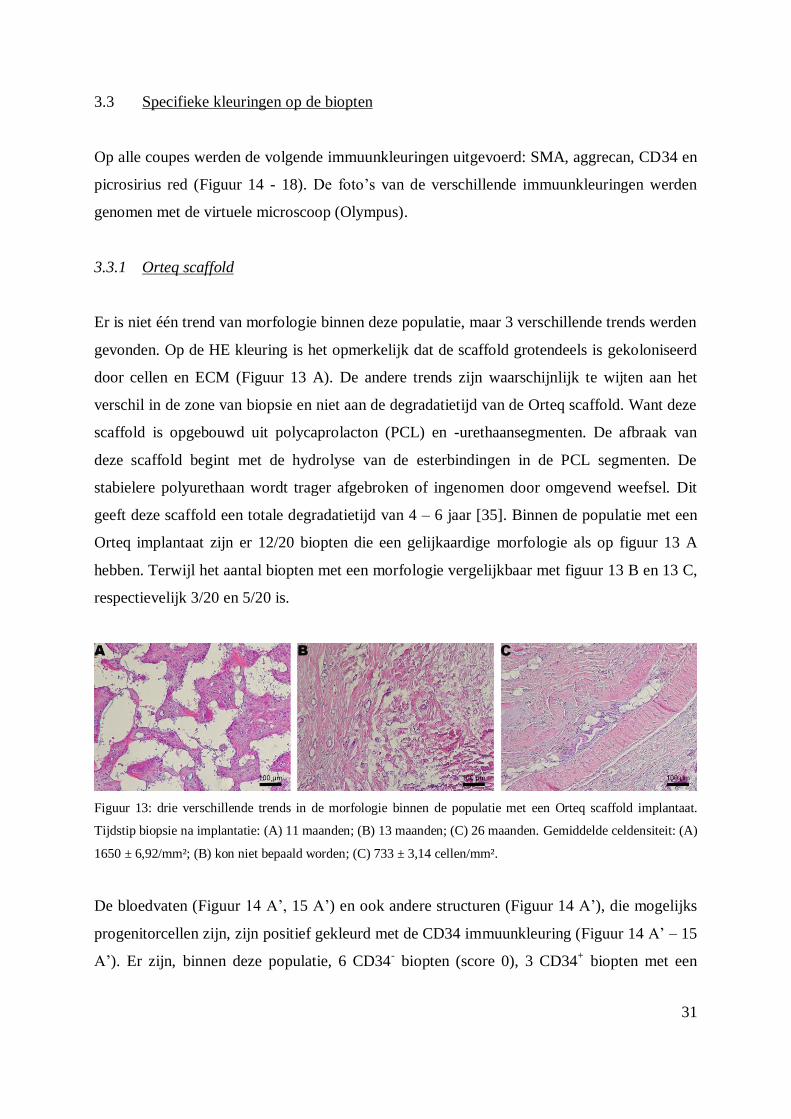
3.3.1 Orteq scaffold
Er is niet één trend van morfologie binnen deze populatie, maar 3 verschillende trends werden
gevonden. Op de HE kleuring is het opmerkelijk dat de scaffold grotendeels is gekoloniseerd
door cellen en ECM (Figuur 13 A). De andere trends zijn waarschijnlijk te wijten aan het
verschil in de zone van biopsie en niet aan de degradatietijd van de Orteq scaffold. Want deze
scaffold is opgebouwd uit polycaprolacton (PCL) en -urethaansegmenten. De afbraak van
deze scaffold begint met de hydrolyse van de esterbindingen in de PCL segmenten. De
stabielere polyurethaan wordt trager afgebroken of ingenomen door omgevend weefsel. Dit
geeft deze scaffold een totale degradatietijd van 4 – 6 jaar [35]. Binnen de populatie met een
Orteq implantaat zijn er 12/20 biopten die een gelijkaardige morfologie als op figuur 13 A
hebben. Terwijl het aantal biopten met een morfologie vergelijkbaar met figuur 13 B en 13 C,
respectievelijk 3/20 en 5/20 is.
Figuur 13: drie verschillende trends in de morfologie binnen de populatie met een Orteq scaffold implantaat.
Tijdstip biopsie na implantatie: (A) 11 maanden; (B) 13 maanden; (C) 26 maanden. Gemiddelde celdensiteit: (A)
1650 ± 6,92/mm²; (B) kon niet bepaald worden; (C) 733 ± 3,14 cellen/mm².
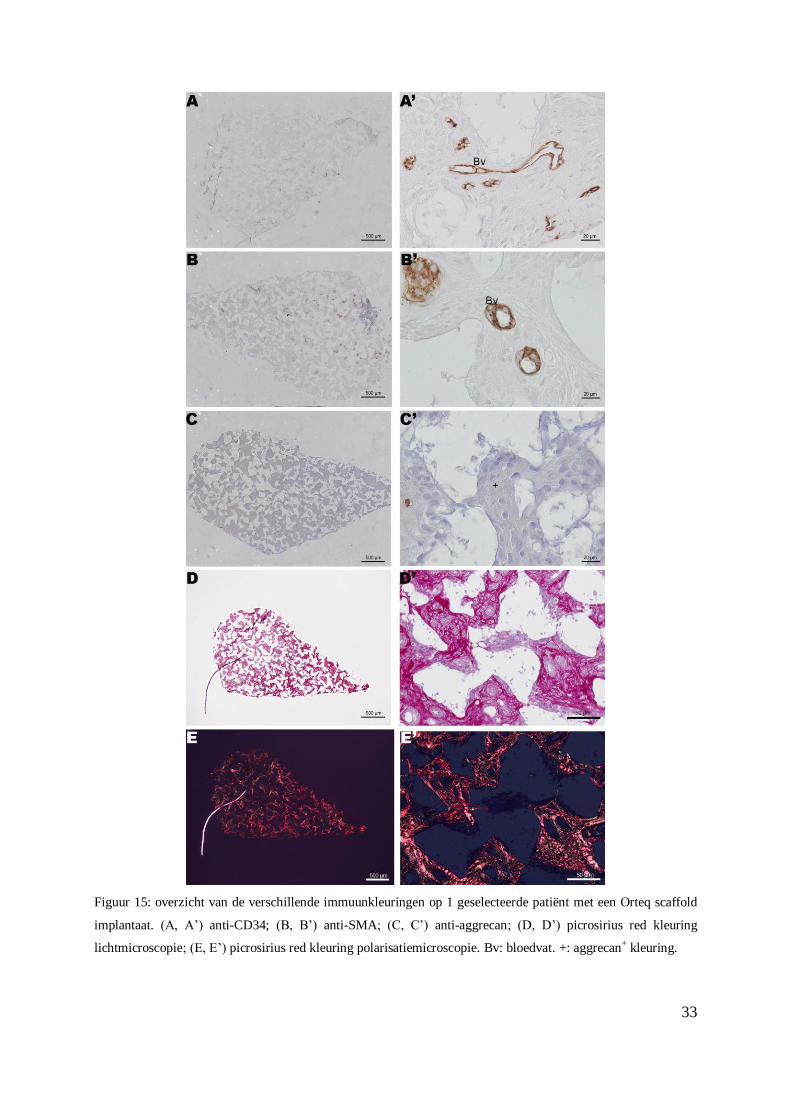
De bloedvaten (Figuur 14 A’, 15 A’) en ook andere structuren (Figuur 14 A’), die mogelijks
progenitorcellen zijn, zijn positief gekleurd met de CD34 immuunkleuring (Figuur 14 A’ – 15
A’). Er zijn, binnen deze populatie, 6 CD34- biopten (score 0), 3 CD34
+ biopten met een

32
zwakke intensiteit, 5 CD34+ biopten met een gemiddelde intensiteit en 6 CD34
+ biopten met
een sterke intensiteit (Tabel 3, Bijlage XIX).
De resultaten met de SMA kleuring tonen aan dat enkel de gladde spiercellen rond de
bloedvaten (Figuur 15 B’) positief kleurden met dit antilichaam.
De ECM van het biopt op figuur 15 C – C’ vertoont een middelmatige intensiteit voor de
aggrecan kleuring.
Op de lichtmicroscopische en polarisatie microscopische opnames van de picrosirius red
kleuring is er een opmerkelijke rode kleur overheersend. Dit toont aan dat het type collageen
dat overheersend is in de meniscus, collageen type I is (Figuur 15 D’ - E’).
Figuur 14: coupe van een biopt van een patiënt met een Orteq implantaat. De endotheliale aflijning en
voorlopercellen zijn CD34+. Bv: bloedvat. *Pr: mogelijke progenitorcel.
33
Figuur 15: overzicht van de verschillende immuunkleuringen op 1 geselecteerde patiënt met een Orteq scaffold
implantaat. (A, A’) anti-CD34; (B, B’) anti-SMA; (C, C’) anti-aggrecan; (D, D’) picrosirius red kleuring
lichtmicroscopie; (E, E’) picrosirius red kleuring polarisatiemicroscopie. Bv: bloedvat. +: aggrecan+ kleuring.

34
3.3.2 Deepfrozen allogreffe
De resultaten van de HE kleuring wijzen erop dat de biopten binnen deze populatie ingedeeld
kunnen worden in 2 verschillende categorieën. Zo zijn er twee trends met verschillen in de
morfologie (Figuur 16 A - B). Binnen de populatie met een deepfrozen allogreffe
transplantaat zijn er 8/19 biopten die een gelijkaardige morfologie als op figuur 16 A hebben.
Terwijl de rest van de biopten (11/19) een morfologische gelijkenis met figuur 16 B vertonen.
Figuur 16: twee verschillende trends in de morfologie binnen de populatie met een deepfrozen allogreffe
transplantaat. Tijdstip biopsie na implantatie: (A) kon niet achterhaald worden; (B) 11 maanden. Gemiddelde
celdensiteit: (A) 750 ± 3,02 cellen/mm²; (B) 1317 ± 3,82 cellen/mm².
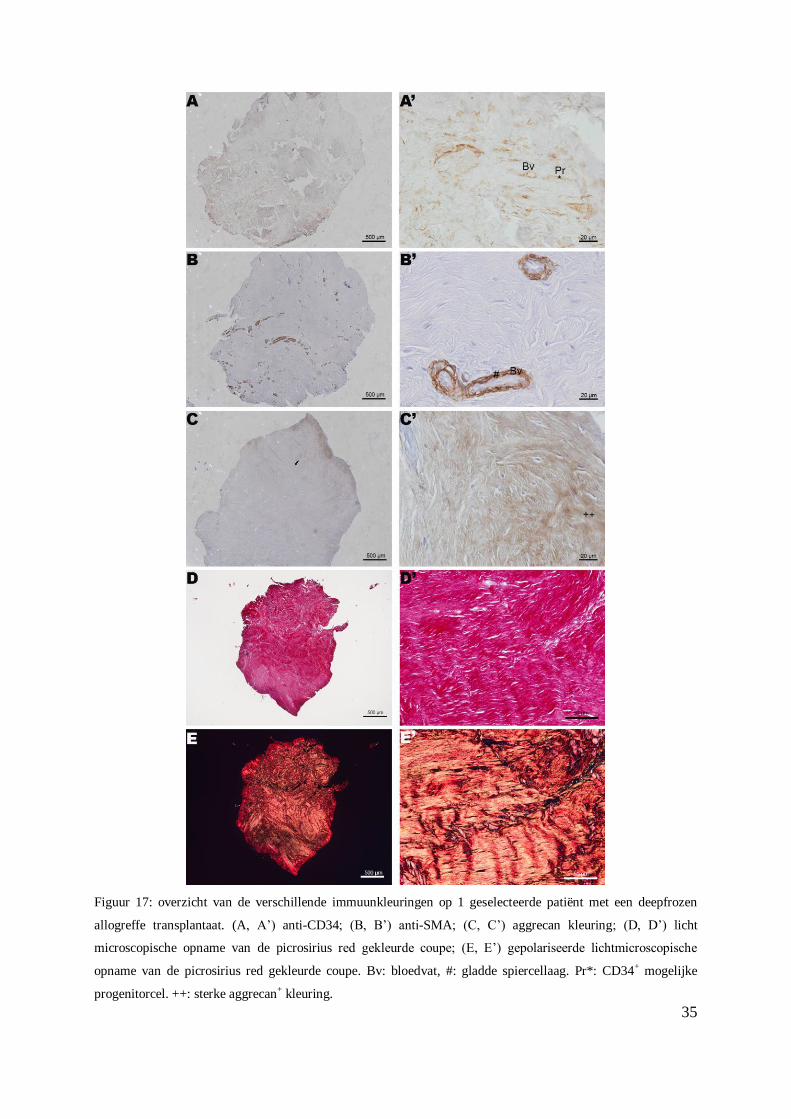
De resultaten van de CD34 immuunkleuring tonen aan dat de bloedvaten (Figuur 17 A - A’)
en ook mogelijkse voorlopercellen (Figuur 17 A’) positief zijn binnen deze populatie. Er zijn,
binnen deze populatie, 12 biopten die CD34- zijn (score 0), 1 CD34
+ biopt met een zwakke
intensiteit, 0 CD34+ biopten met een gemiddelde intensiteit en 4 CD34
+ biopten met een
sterke intensiteit (Tabel 3, Bijlage XIX). Verder waren er 2 biopten die geen score toegekend
kregen, omdat deze verloren waren gegaan tijdens de voorbehandeling van de
immuunkleuring.
Enkel de gladde spiercellen rond de bloedvaten kleurden positief met de anti-SMA
immuunkleuring (Figuur 17 B - B’).
Een zeer sterke intensiteit voor de aggrecan kleuring is waar te nemen op figuur 17 C’.
De intens rode kleuring op de lichtmicroscopische en polarisatie microscopische opnames van
de picrosirius red kleuring wijst op een hoger gehalte collageen type I in de meniscus (Figuur
17 D’ - E’).
35
Figuur 17: overzicht van de verschillende immuunkleuringen op 1 geselecteerde patiënt met een deepfrozen
allogreffe transplantaat. (A, A’) anti-CD34; (B, B’) anti-SMA; (C, C’) aggrecan kleuring; (D, D’) licht
microscopische opname van de picrosirius red gekleurde coupe; (E, E’) gepolariseerde lichtmicroscopische
opname van de picrosirius red gekleurde coupe. Bv: bloedvat, #: gladde spiercellaag. Pr*: CD34+ mogelijke
progenitorcel. ++: sterke aggrecan+ kleuring.

36
3.3.3 Viabele allogreffe
De biopten binnen deze populatie kunnen ingedeeld worden in 2 verschillende categorieën op
basis van de HE kleuring (Figuur 18 A - B). Er zijn 8/18 biopten die morfologisch een
gelijkenis vertonen met figuur 18 A. De rest van de biopten (10/18 van de populatie met een
viabel allogreffe transplantaat een gelijkaardige morfologie als op figuur 18 B hebben.
Figuur 18: twee verschillende trends in de morfologie binnen de populatie met een viabele allogreffe
transplantaat. Tijdstip biopsie na implantatie: (A) 32 maanden; (B) 4 maanden. Gemiddelde celdensiteit: (A) 567
± 4,13 cellen/mm²; (B) 900 ± 3,69 cellen/mm².
Mogelijkse voorlopercellen en vasculaire structuren (Figuur 19 A’) zijn positief gekleurd met
de anti-CD34 immuunkleuring (Figuur 19 A - A’). Er zijn, binnen deze populatie, 4 CD34-
biopten (score 0), 3 CD34+ biopten met een zwakke intensiteit, 4 CD34
+ biopten met een
gemiddelde intensiteit en 5 CD34+ biopten met een sterke intensiteit (Tabel 3, Bijlage XIX).
Aan 1 biopt kon geen score worden toegekend, omdat het verloren was gegaan tijdens de
voorbehandeling van de immuunkleuring.
De anti-SMA immuunkleuring leverde enkel positieve resultaten rond de bloedvaten (Figuur
18 B’). De gladde spiercellaag rond de bloedvaten was SMA+.
Het geselecteerd biopt op vertoont een zwakke intensiteit voor de aggrecan kleuring (Figuur
19 C – C’).
De opnames met de lichtmicroscoop en de polarisatiemicroscoop van de coupes gekleurd met
picrosirius red geven een overwegend rode kleur, met een hoge intensiteit. Deze rode kleur
wijst op collageen type I (Figuur 19 D’ - E’).

37
Figuur 19: overzicht van de verschillende immuunkleuringen op 1 geselecteerde patiënt met een viabele
allogreffe transplantaat. (A, A’) anti-CD34; (B, B’) anti-SMA; (C, C’) anti-aggrecan; (D, D’)
lichtmicroscopische opname van picrosirius red; (E, E’) polarisatie microscopische opname van picrosirius red.
Pr*: mogelijke progenitorcel. Bv: bloedvat. +/-: zwakke aggrecan kleuring.

38
3.4 Vergelijking van de 3 populaties
Een overzicht van het procentueel aantal positieven per immuunkleuring en populatie wordt
gegeven in figuur 20. Voor de anti-SMA immuunkleuring stellen de positieve resultaten in de
grafiek, de resultaten voor waarop de gladde spiercellen rond de vasculaire structuren positief
waren. Op geen enkel biopt konden myofibroblastachtige cellen aangetoond worden met de
SMA immuunkleuring.
Figuur 20: procentueel overzicht van het aantal positieven per immuunkleuring per populatie. SMA (α-smooth
muscle actin): enkel positief voor de gladde spiercellen rond de bloedvaten. Wanneer vasculaire en/of cellulaire
structuren (mogelijke progenitorcellen) positief gekleurd waren met de CD34 immuunkleuring, werden deze
meegeteld als CD34+ en opgenomen in de grafiek.
Een overzicht van alle geselecteerde biopten met de daarbij horende resultaten voor sommige
kleuringen wordt weergegeven in tabel 3 (Bijlage XIX). De drie populaties werden onderling
onderzocht op statistisch significante verschillen voor de celdensiteit in de superficiële zone,
de celdensiteit in de diepe zone, het aantal vasculaire structuren (inclusief de capillairen),
intensiteit van de anti-CD34 immuunkleuring, intensiteit van de anti-aggrecan
immuunkleuring en het kleurverschil van de picrosirius red kleuring op de gepolariseerde
lichtmicroscopische opnames (Tabel 3, Bijlage XIX). Het aantal CD34+
vasculaire structuren
werd geteld op de anti-CD34 gekleurde coupes. Er werd geen onderscheid gemaakt op basis
van de diameter van de verschillende vaten.

39
Morfologisch is er op de HE gekleurde coupes zowel een verschil tussen de populaties als
binnen de populaties (Figuur 12). Echter, dit verschil is beperkt tot wat er visueel wordt
waargenomen op de coupes, want er is geen statistisch significant verschil tussen de
populaties betreffende de gemiddelde celdensiteit. Zo is de gemiddelde celdensiteit voor een
biopt van een patiënt met een Orteq implantaat 896 ± 5,77 cellen/mm², voor die met een
deepfrozen transplantaat is deze 850 ± 6,18 cellen/mm² en de gemiddelde celdensiteit voor
een biopt van een viabel transplantaat is 658 ± 5,31 cellen/mm² (resultaten niet weergegeven
in tabel).
Wanneer de celdensiteit in de superficiële zone en de diepe zone afzonderlijk wordt
geanalyseerd, worden er significant verschillende resultaten bekomen. In de diepere zone is
de gemiddelde celdensiteit voor de biopten van deepfrozen allogreffes 177 ± 2,1 cellen/mm²
en voor de biopten van viabele allogreffes 426 ± 3,71 cellen/mm². Het verschil in gemiddelde
celdensiteit in de diepe zone is statistisch significant verschillend (p < 0,01) tussen deze 2
populaties. De diepere zone van de viabele allogreffes is meer gekoloniseerd met cellen dan
de deepfrozen allogreffes. In de superficiële zone is er geen statistisch significant verschil
tussen deze twee populaties, wat betreft de celdensiteit (Tabel 3, Bijlage XIX). De
gemiddelde celdensiteit van de superficiële zone van een biopt met een deepfrozen
transplantaat is 1200 ± 8,32 cellen/mm² en de gemiddelde celdensiteit van een biopt met een
viabel transplantaat is 1083 ± 6,99 cellen/mm².
Er is een statistisch significant verschil (p < 0,01) in de gemiddelde celdensiteit in de diepe
zone tussen de populaties met een Orteq implantaat en viabele allogreffe transplantaat. Het
Orteq implantaat is meer gekoloniseerd met cellen in de diepe zone dan de viabele allogreffes.
De gemiddelde celdensiteit van de diepe zone voor een biopt met een Orteq implantaat is 896
± 5,77 cellen/mm² en voor de biopten met viabele allogreffes is dit 426 ± 3,71 cellen/mm².
Maar er is geen significant verschil tussen deze twee populaties, wat betreft de celdensiteit
van de superficiële zone. Wanneer het Orteq implantaat vergeleken wordt met de deepfrozen
allogreffes, is er een statistisch significant verschil in de gemiddelde celdensiteit in zowel de
superficiële (p < 0,05) als de diepe zone (p < 0,001). De superficiële zone van deepfrozen
allogreffes (gemiddelde celdensiteit: 1200 ± 8,32 cellen/mm²) worden beter gekoloniseerd
met cellen dan Orteq implantaten (gemiddelde celdensiteit: 896 ± 5,77 cellen/mm²). Terwijl
de diepe zone van Orteq implantaten (gemiddelde celdensiteit: 896 ± 5,77 cellen/mm²) beter
gekoloniseerd worden met cellen dan de deepfrozen allogreffes (177 ± 2,1 cellen/mm²). Er is
wel een verschil in celtype in de superficiële zone van de biopten met Orteq implantaat. Hier

40
zijn er vooral fibrochondrocyten in de superficiële zone, terwijl deze zone in biopten van
deepfrozen en viabele allogreffes gekoloniseerd is met fibroblastachtige cellen.
Wanneer de 3 populaties, Orteq scaffold, deepfrozen allogreffe en viabele allogreffe, met
elkaar vergeleken worden aan de hand van de uitgevoerde kleuringen is er een statistisch
significant verschil in de intensiteit van de CD34+ progenitorcellen tussen de groepen met een
deepfrozen (17 A - A’) en een viabele allogreffe transplantaat (19 A - A’) (Tabel 3, Bijlage
XIX). De intensiteit van de CD34+ progenitorcellen is sterker in de populatie van viabele
allogreffes dan in de populatie met deepfrozen allogreffes. Er is geen significant verschil (p >
0,05) in de intensiteit van de CD34+ biopten van het Orteq implantaat en de intensiteit van de
deepfrozen en viabele allogreffes (Tabel 3, Bijlage XIX). Wanneer de p-waarde op 0,06 wordt
ingesteld, dan is er wel een statistisch significant verschil in de intensiteit van de CD34
kleuring tussen de biopten met het Orteq implantaat en deepfrozen allogreffe. De intensiteit
van de CD34+ progenitorcellen van de biopten met een Orteq implantaat is sterker dan de
intensiteit van de biopten met een deepfrozen allogreffe.
In tegenstelling tot wat er gezien wordt op de geselecteerde coupes (Figuur 17 A’, 19 A’) is
de kleuring van de CD34+
progenitorcellen binnen de groep met de deepfrozen allogreffe
zwakker dan binnen de groep met een viabele allogreffe. Ook de endotheliale aflijning van de
vasculaire structuren is CD34+. Hier is er ook een statistisch significant verschil in het aantal
bloedvaten tussen de populaties met een deepfrozen (Figuur 17 A - A’) en een viabele
allogreffe transplantaat (19 A - A’) én tussen de populaties met een Orteq implantaat (14 A -
A’) en een deepfrozen allogreffe transplantaat (17 A - A’) (Tabel 3, Bijlage XIX). De biopten
van de deepfrozen-groep hebben het minst gemiddeld aantal bloedvaten (74 ± 1,50
bloedvaten/mm²) in vergelijking met de groep met een viabel allogreffe implantaat (241 ±
2,17 bloedvaten/mm²) en de groep met een Orteq implantaat (158 ± 1,65 bloedvaten/mm²).
Er waren geen SMA+ myofibroblastachtige cellen in de drie vergeleken populaties; enkel de
gladde spiercellaag rond de bloedvaten waren SMA+ (Figuur 15 B’, 17 B’, 19 B’).
De grafische voorstelling in figuur 20 geeft de indruk dat de resultaten van de anti-aggrecan
immuunkleuring tussen de populaties sterk verschillend zijn. Na statistische analyse is echter
gebleken dat er geen statistisch significant verschil is in de intensiteit van de aggrecan
kleuring tussen de verschillen populaties (Tabel 3, Bijlage XIX). Het verkregen resultaat en

41
de intensiteit zijn dus niet verschillend tussen de groepen Orteq, deepfrozen en viabel (Figuur
15 C’, 17 C’, 19 C’).
Het achterhalen van de verhoudingen van de verschillende types collageen, op basis van de
picrosirius red kleuring, is onmogelijk zonder de nodige computersoftware om de
verschillende kleurschakeringen en –intensiteiten te onderscheiden.
Maar de analyse van de biopten, enkel op basis van de visueel waarneembare kleuren op de
opnames met de polarisatiemicroscoop, toont dat er geen statistisch significant verschil is op
te merken in het kleurverschil tussen de verschillende populaties (Tabel 3, Bijlage XIX). De
visueel waarneembare kleuren geel, oranje/rood en groen, stemmen overeen met
respectievelijk collageen type II, collageen type I en collageen type III [31].
Bij alle drie de biopten is de rode kleur duidelijk overheersend en de intensiteit van de rode
kleur varieert naargelang de zone (Figuur 15 F, 17 F, 19 F). Aan de perifere zone van de
coupes is er duidelijk een intensere, rode kleur dan centraal op de coupes (Figuur 15 E, 17 E,
19 E). Op de 20x vergroting is er variatie tussen de verschillende populaties, maar deze
variatie is vooral ontstaan als gevolg van de zone waarop werd ingezoomd (Figuur 15 F’, 17
F’, 19 F’).
4. Bespreking
Er wordt een duidelijk verschil in de morfologie tussen de verschillende populaties
waargenomen met de haematoxyline eosine kleuring (Figuur 12). De twee verschillende
celtypes, die kunnen onderscheiden worden, zijn: fibrochondrocyten en fibroblastachtige
cellen (Figuur 12 A - F). De fibroblastachtige cellen komen voor ter hoogte van superficiële
zone, terwijl de fibrochondrocyten ter hoogte van de diepe zone voorkomen. Op enkele
coupes konden de fibroblastachtige cellen onderscheiden worden (Figuur 12 DEF). Maar de
superficiële zone van de biopten met een Orteq implantaat vertonen een morfologisch verschil
in de superficiële zone; deze zone is voornamelijk gekoloniseerd met fibrochondrocyten
(Figuur 12 A).
De celdensiteit in het centraal gedeelte van biopten met een deepfrozen allogreffe is opvallend
lager (Tabel 3, Bijlage XIX). Dit resultaat stemt overeen met de resultaten van een eerdere
studie van Arnoczky et al [36]. Onze resultaten tonen dat de gemiddelde celdensiteit van de
biopten van patiënten met een deepfrozen transplantaat significant lager is dan de gemiddelde

42
celdensiteit van de biopten van patiënten met een viabel transplantaat. Dit komt overeen met
de resultaten van recente histologische analyses. Die histologische analyses toonden aan dat
de biopten, die werden genomen van patiënten met een deepfrozen allogreffe, weinig
gerepopuleerd werden door gastheercellen. De repopulatie was beperkt tot de superficiële
zone en de kern van het transplantaat bleef acellulair [9]. De diepe zone van de biopten met
een Orteq implantaat is meer gekoloniseerd met cellen dan de biopten met een deepfrozen
allogreffe. De deepfrozen allogreffes laten weinig/geen ingroei toe van cellen in de richting
van de diepe zone. Dit is waarschijnlijk te wijten aan de procedure, die wordt doorlopen
tijdens de transplantatie, waarbij cellen afsterven [12].
Er werd eerder verwacht dat de morfologie binnen iedere populatie vergelijkbaar zou zijn,
tegen deze verwachting in waren er morfologische verschillen binnen de populaties en tussen
de 3 populaties. De implantaten worden gerepopuleerd met verschillende celtypes na de
implantatie. Rodeo et al. postuleerde dat allogreffes gerepopuleerd worden met cellen
afkomstig van het synoviaal membraan van de gastheer [36]. Waarschijnlijk zijn de cellen die
geïncorporeerd zijn in de implantaten ook afkomstig van de synoviale membraan.
Declercq et al. en Verdonk et al. deden al onderzoek op de expressie van CD34 in de
meniscale cellen [1, 6].
Verdonk et al. toonde dat er drie verschillende celtypes positief zijn voor CD34. Vooreerst
zijn de fibroblastachtige cellen ter hoogte van de buitenste vasculaire zone van de meniscus
CD34+. Ten tweede zijn er CD34
+ cellen ter hoogte van de superficiële zone van de meniscus,
deze cellen hebben een ronde tot ovale morfologie. Tot slot is de endotheliale aflijning van de
vasculaire structuren positief voor CD34 [6]. Op het positief controleweefsel, de humane
buikhuid, zijn de bloedvaten positief gekleurd met CD34. Ook tonen de resultaten van de
CD34 immuunkleuring op de verschillende populaties aan dat de vasculaire structuren
positief zijn ter hoogte van de endotheliale aflijning. Overeenstemmend met de resultaten van
Verdonk et al. waren er ter hoogte van de buitenste vasculaire zone ook structuren met een
uitgerekte morfologie, die CD34+
waren [6]. Deze CD34+ cellen komen voor in de drie
populaties. Zoals eerder aangetoond door Declercq et al. waren in onze studie ook geen
CD34+ cellen in de binnenste zone van de meniscus [1]. In de groep met een viabel implantaat
hebben de CD34 gekleurde progenitorcellen een hogere intensiteit dan de groep met een
deepfrozen implantaat. Rijk et al. toonde aan dat de ingroei van cellen en bloedvaten start
vanaf de periferie naar het centraal gedeelte toe [37]. Nieuwe capillairen revasculariseren het

43
implantaat slechts gradueel en incompleet [36]. Resultaten vergelijkbaar met onze resultaten,
in verband met de repopulatie van het deepfrozen implantaat werd bekomen in een andere
studie [36].
De hogere intensiteit van de CD34 kleuring binnen de groep met een viabel implantaat zou
het gevolg kunnen zijn dat er meer progenitorcellen gespaard worden tijdens de procedure
vóór de implantatie. Deze allogreffes moeten binnen de 2 weken geïmplanteerd worden in de
acceptor [12]. Door de minder lange tijd, die wordt verstreken tussen de explantatie en
implantatie zouden er minder cellen sterven. Het probleem met deze techniek is de lange
tijdsduur van de microbiële testen [12]. Wanneer gekeken wordt naar de bewaringstechniek
van de deepfrozen allogreffes, zijn er op het moment van implantatie al heel veel cellen dood.
Het donormateriaal wordt hier onmiddellijk bij een temperatuur van -80°C gebracht [12].
Deze bewaringstechniek zou een verklaring kunnen vormen voor de minder goede resultaten,
die we bekomen hebben binnen de groep met een deepfrozen implantaat (Tabel 3, Bijlage
XIX).
In tegenstelling tot de articulaire chondrocyten produceren de matrix-geassocieerde cellen in
de meniscus meer collageen type I dan collageen type II. De verhouding collageen type I/type
II is in de meniscus gemiddeld 9/1 [6]. Deze verhouding toont aan dat er een heel lage
hoeveelheid collageen type II aanwezig is in de meniscus. Aan de hand van de resultaten van
de anti-collageen type II kleuring op de epiglottis (Figuur 11); waar de kleuring enkel positief
is aan de periferie, kunnen de negatieve resultaten verklaard worden. Er werd echter verwacht
dat de epiglottis min of meer overal positief zou zijn met een gelijke intensiteit. De epiglottis
zou normaal gezien positief moeten kleuren voor collageen type II met een sterke intensiteit
in zowel de territoriale matrix als de interterritoriale matrix [38]. De resultaten die verkregen
werden met het muis anti-humaan collageen type II (Acris Antibodies) antilichaam waren
negatief voor alle biopten. De slechte resultaten die werden verkregen, zowel op het positief
controleweefsel als de biopten, zou kunnen wijzen dat de voorbehandeling voor dit
antilichaam niet de gepaste is. Deze kleuring zou verder op punt gezet moeten worden.
De analyse van de biopten voor de verschillende types collageen in de meniscus zullen verder
besproken worden op basis van de picrosirius red kleuring.
Met de anti-SMA immuunkleuring werd er gehoopt om de contractiele fenotypes, die
ontstaan tijdens het herstel van weefsels, aan te tonen [1]. De resultaten van deze

44
immuunkleuring toonde aan dat slechts de gladde spiercellen rond de bloedvaten positief
kleurden. In geen enkel biopt werd een contractiel fenotypische celtype waargenomen met
deze kleuring. Dit is waarschijnlijk te wijten aan het feit dat er een te lange periode is
verstreken tussen de datum van operatie en de datum van biopsie. De periode varieert tussen
10 weken en 21 jaar. Declercq et al. toonde aan dat er geen SMA+ cellen aanwezig zijn in de
gezonde menisci en menisci afkomstig van patiënten met OA. Biopten afkomstig van
patiënten met een gescheurde meniscus waren wel positief voor SMA. Het aantal SMA+
cellen nam toe in aantal in functie van de tijd die verstreken is sinds de schade [1]. In de
studie van Declercq et al. varieerde de tijd tussen het ontstaan van schade en biopsie tussen
0,5 weken tot 27 weken. In ons onderzoek was deze tijd meer gespreid, gaande van 10 weken
tot 21 jaar. Dit zou een verklaring kunnen zijn voor de negatieve resultaten die we hebben
verkregen met deze immuunkleuring. De myofibroblastachtige cellen komen mogelijks enkel
tot expressie gedurende een bepaalde periode van het helingsproces.
De belangrijkste proteoglycaan in de meniscus is aggrecan, het komt in hogere hoeveelheden
voor in de binnenste en de middelste zone van de meniscus dan in de perifere zone [6]. Onze
resultaten voor deze kleuring op de verschillende biopten zijn uiteenlopend (Tabel 3, Bijlage
XIX). De perifere zone is minder intens positief voor aggrecan (Figuur 14 C, 16 C, 18 C) dan
de binnenste zone. Sun et al. toonde aan dat de aggrecan kleuring meer intens is aan de
diepere zone van OA menisci dan de oppervlakkige zone. Dit wijst op een actieve aggrecan
synthese en turnover in de meniscus. In dezelfde studie werd eveneens aangetoond dat het
aggrecan gehalte in OA menisci hoger is dan in gezonde menisci [39]. Aangezien wij in ons
onderzoek geen significant verschil hebben kunnen aantonen in de intensiteit van aggrecan
tussen de 3 populaties (Tabel 3, Bijlage XIX), zou dit kunnen wijzen op een minder actieve
aggrecan synthese in deze menisci. Veronderstellend dat de patiënten van de 3 populaties,
waarvan biopsie werd genomen, geen OA patiënten zijn, zouden onze resultaten erop kunnen
wijzen dat in de drie populaties de menisci een goed herstel tonen.
Informatie betreffende de verschillende types collageen in de meniscus werd gewonnen met
de picrosirius red kleuring. De collageen type II immuunkleuring leverde geen positieve
resultaten op en de evaluatie van de verschillende types collageen in de biopten berust enkel
op de picrosirius red kleuring. Vroegere studies tonen aan dat er met de combinatie van
picrosirius red en gepolariseerde lichtmicroscopie wel kwalitatieve informatie gewonnen kan

45
worden, maar dat kwantitatieve informatie eerder gelimiteerd is [34]. Ondanks de specificiteit
die verkregen kan worden met deze combinatie, werd deze kleuring tot in de jaren 1980 niet
veel gebruikt [34]. Verder is de kleur van de verschillende types collageen, op de
gepolariseerde lichtmicroscopische opnames van de picrosirius red gekleurde coupes,
afhankelijk van de dikte van de collageenvezels. Door de verschillen in dikte worden er
verschillende kleuren gezien. Zo verandert de kleur met de toenemende dikte van vezels van
geel naar oranje/rood [34]. De dunnere vezels vertonen een lichtere, moeilijk te onderscheiden
kleur. Dit brengt met zich het nadeel mee dat het collageen gehalte onderschat kan worden
[31, 34]. Junqueira et al. beweert dat de rode kleur correspondeert met collageen type I, de
gele kleur met collageen type II en de groene kleur met collageen type III op de
gepolariseerde lichtmicroscopische opnames van de picrosirius red gekleurde coupes [31].
Echter niet iedere onderzoeker is ervan overtuigd dat deze interpretatie correct is. Zo zegt
Rich et al. dat het type collageen niet kan geïdentificeerd worden aan de hand van de
waargenomen kleur op de gepolariseerde lichtmicroscopische opnames van de picrosirius red
gekleurde coupes. Hoewel de collageen type III vezels dunner zijn dan de type I vezels, zou
het kunnen zijn dat de groene kleur een immature collageen type I vezel is [34].
Vermits er nogal wat tegenstrijdigheden zijn wat betreft de interpretatie van de gepolariseerde
lichtmicroscopische opnames van de picrosirius red gekleurde coupes kunnen de populaties
ook niet met deze kleuring vergeleken worden op de collageen type I/type II verhouding.
Wanneer de resultaten, gebaseerd op de bevindingen van Junqueira et al., vergeleken zullen
worden, dan is er duidelijk een overheersing van collageen type I (Figuur 14 F’, 16 F’, 18 F’).
Junqueira et al. schrijft de rode kleur op de opnames van de gepolariseerde lichtmicroscoop
toe aan collageen type I [34]. In mindere mate wordt er op de 2x vergroting geel en groen
waargenomen. De gele kleur slaat op de aanwezigheid van collageen type II. Verder zou de
groene kleur in de meniscus de immature collageen type I voorstellen [34]. De collageen type
I/type II verhouding in de meniscus is verschillend naargelang de zone in de meniscus [6].
Aangezien biopsie meestal wordt genomen van de witte zone van de meniscus, zou een
overheersende gele kleur een normaal resultaat kunnen zijn (Tabel 3, Bijlage XIX).

46
5. Algemeen besluit
De histologische analyse van in vivo biopten, afkomstig van patiënten met een Orteq scaffold
implantaat, deepfrozen of viabel transplantaat gaf uiteenlopende resultaten. Zo werd er geen
verschil gevonden, tussen de drie populaties, in verband met de intensiteit van de aggrecan
kleuring. Ook konden de verschillende types collageen niet met 100% zekerheid van elkaar
onderscheiden worden. De overwegend rode kleuring op de gepolariseerde
lichtmicroscopische opnames van de picrosirius red gekleurde coupes, toont dat collageen
type I het meest voorkomend type collageen is in de meniscus. Aangezien de gemiddelde
verhouding van collageen type I/type II in de meniscus 9/1 is, zijn deze resultaten
aanvaardbaar [6].
Hoewel er binnen iedere populatie CD34+ cellen, die mogelijks progenitorcellen zijn, konden
worden aangetoond, waren er geen SMA+ myofibroblastachtige cellen binnen de
geselecteerde steekproeven. Deze CD34+ progenitorcellen zijn gelokaliseerd ter hoogte van de
buitenste, vasculaire zone van de meniscus. Het waren niet enkel deze progenitorcellen die
CD34+ waren, maar ook de endotheliale aflijning van vasculaire structuren was CD34
+. Het
gemiddeld aantal bloedvaten op deze CD34+ coupes werd geteld. Uit deze telling bleek dat
biopten van viabele transplantaten een significant hoger aantal bloedvaten hadden dan de
biopten van deepfrozen transplantaten. De biopten van het Orteq implantaat hadden een
significant hoger aantal bloedvaten dan de biopten van deepfrozen transplantaten. De
resultaten van de CD34 kleuring tonen eveneens aan dat de biopten van de populatie met een
deepfrozen allogreffe het minst aantal progenitorcellen bevat.
De resultaten van de HE kleuring en de anti-CD34 immuunkleuring geven aan dat er
verschillende celtypes gevormd worden in de menisci met verschillende implantaten. De
diepe zone van de deepfrozen allogreffe wordt minder gekoloniseerd met cellen dan de diepe
zone van viabele allogreffe en Orteq implantaat. Er is verder een ingroei van bloedvaten
mogelijk in de implantaten, ter hoogte van de vasculaire zone.
Deze resultaten geven aan dat de biopten met een Orteq implantaat en een viabel allogreffe
het best uit de studie komen, wat betreft de graad van regeneratie. De biopten met een
deepfrozen allogreffe worden minder gekoloniseerd met cellen in de diepe zone en er komen
ook minder vasculaire structuren voor op deze biopten dan op de twee andere populaties. De

47
vergelijking van de biopten met een Orteq implantaat en viabele allogreffe toont dat de diepe
zone van de Orteq implantaten beter gekoloniseerd worden met cellen dan de diepe zone van
de viabele allogreffes. Dit is waarschijnlijk te danken aan de hoge porositeit (80 %) van de
Orteq scaffold.
Hoewel de biopten van de Orteq scaffold en de viabele allogreffe, beide betere resultaten
leveren dan de biopten van deepfrozen allogreffe, is de Orteq scaffold waarschijnlijk de beste
uit de drie populaties. Want met de Orteq scaffold moet er geen extra inzet geleverd worden
om de verschillende nadelen, die verbonden zijn aan de transplantatie van viabele allogreffes,
zoals de weefselbeschikbaarheid, het risico op de overdracht van infecties, HLA-mismatching
en het zoeken naar een transplantaat met de gepaste grootte, te proberen wegwerken. Het
voordeel van het gebruik van een Orteq scaffold is dat het op maat gesneden kan worden en
kan ingebracht worden in de patiënt, zonder lange procedures die moeten uitgevoerd worden
alvorens de implantatie. Ook worden er na afbraak van de Orteq scaffold geen toxische
stoffen vrijgesteld, die schadelijk zouden kunnen zijn voor de patiënt. Naast deze opgesomde
voordelen, toont ons onderzoek aan dat de Orteq scaffold een goede weefselingroei en
kolonisatie met cellen toelaat.
Het gebruik van een Orteq scaffold betekent tijdswinst zowel voor de patiënt en als voor de
chirurg. De patiënt moet niet langer wachten op een geschikte meniscus en de chirurg kan de
scaffold onmiddellijk, in de operatiekamer, op maat snijden én implanteren.

48
6. Referenties
1. Declercq HA, Forsyth RG, Verbruggen A, Verdonk R, Cornelissen MJ, Verdonk PCM (2012). CD34 and
SMA expression of superficial zone cells in the normal and pathological human meniscus. Journal of
orthopaedic research 30: 800-808.
2. Verdonk R, Verdonk PC, Huysse W, Forsyth RG, Heinrichs EL (2011). Tissue ingrowth after implantation of a novel, biodegradable polyurethane scaffold for treatment of partial meniscal lesions. The American
journal of sports medicine 39: 774-782.
3. Buma P, Ramrattan NN, van Tienen TG, Veth RP (2004). Tissue engineering of the meniscus. Biomaterials
25: 1523-1532.
4. Efe T, Getgood A, Schofer MD, Fuchs-Winkelmann S, Mann D, Paletta JR (2012). The safety and short-
term efficacy of a novel polyurethane meniscal scaffold for the treatment of segmental medial meniscus
deficiency. Knee surgery, sports traumatology, arthroscopy: official journal of the ESSKA 20: 1822-1830.
5. Kawamura S, Lotito K, Rodeo SA (2003). Biomechanics and healing response of the meniscus. Operative Techniques in Sports Medicine 11: 68-76.
6. Verdonk PC, Forsyth RG, Wang J, ALmqvist KF, Verdonk R, Veys EM, Verbruggen G (2005).
Characterisation of human knee meniscus cell phenotype. Osteoarthritis and Cartilage 13: 548-560.
7. Vanderploeg EJ, Wilson CG, Imler SM, Ling CH, Levenston ME (2012). Regional variations in the
distribution and colocalization of extracellular matrix proteins in the juvenile bovine meniscus. Journal of
anatomy 221: 174-186.
8. Makris EA, Hadidi P, Athanasiou KA (2011). The knee meniscus: structure-function, pathophysiology,
current repair techniques, and prospects for regeneration. Biomaterials 32: 7411-7431.
9. Van der Bracht H, Verdonk R, Verbruggen G, Elewaut D, Verdonk P (2007). Topics in tissue engineering 3
In: Cell-based meniscus tissue engineering.
10. Kiani C, Chen L, Wu YJ, YEE AJ, Yng BB (2002). Structure and function of aggrecan. Cell research 12:
19-32.
11. Lodish H, Berk A, Zipursky SL (2000). Section 22.3, Collagen: the fibrous proteins of the matrix. In:
Molecular Cell Biology. 4th edition.
12. McDermott ID (2010). What tissue bankers should know about the use of allograft meniscus in
orthopaedics. Cell and tissue banking 11: 75-85.
13. Wang SY, Olson-Kellogg B, Shamliyan TA, Choi JY, Ramakrishnan R, Kane RL (2012). Physical therapy
interventions for knee pain secondary to osteoarthritis: a systemic review. Annals of internal medicine 157:
632-644.
14. Ibarra C, Koski JA, Warren RF (2000). Tissue engineering meniscus: cells and matrix. The Orthopedic
clinics of North America 31: 411-508.
15. Koc ON, Lazarus HM (2001). Mesenchymal stem cells: heading into the clinic. Bone Marrow
Transplantation 27: 235-239.
16. Yoo JU, Barthel TS, Nishimura K, Solchaga L, Caplan AI, Goldberg VM, Johnstone B (1998). The
chondrogenic potential of human bone-marrow-derived mesenchymal progenitor cells. Journal of Bone and
Joint Surgery 80-A: 1745-1757.

49
17. Cornelissen M, Beele H (2012). Cursus: weefselengineering. IN: Culture of human embryonic stem cells
for tissue engineering.
18. Arufe MC, De la Fuente A, Diaz S, Duentes I, De Toro FJ (2010). Chondrogenic potential of
subpopulations of cells expressing mesenchymal stem cell markers derived from human synovial
membranes. Journal of cellular biochemistry 111: 834-845.
19. Ning HX, Lin GT, Lue T, Lin CS (2011). Mesenchymal stem cell marker Stro-1 is a 75kd endothelial
antigen. Biochemical and biophysical research communications 413: 353-357.
20. Otsuki S, Grogan SP, Miyaki S, Kinoshita M, Asahara H, Lotz MK (2010). Tissue neogenesis and Stro-1 expression in immature and mature articular cartilage. Journal of Orthopaedic research 28: 96-102.
21. Rolf HJ, Kierdorf U, Kierdorf H, Schulz J, Seymour N, Schliephake H, Napp J, Niebert S, Woelfel H,
Wiese KG (2008). Localization and characterization of STRO-1(+) cells in the deer pedicle and
regenerating antler. Plos one 3: e2064.
22. Hermida-Gomez T, Fuentes-Boquete I, Gimeno-Longas MJ, Muinos-Lopez E, Diaz-Prado S, de Toro FJ,
Blanco FJ (2011). Bone marrow cells immunomagnetically selected for CD271+antigen promote in vitro
the repair of articular cartilage defects. Tissue engineering 17: 1169-1179.
23. Scholzen T, Gerdes J (2000). The Ki-67 protein: from the known and the unknown. Journal of cellular physiology 182: 311-322.
24. Yammani RR (2012). S100 proteins in cartilage: role in arthritis. Biochimica et biophysica acta 1822: 600-
606.
25. Sakano S, Shu Y, Sandell LJ (2000). Cartilage-derived retinoic acid-sensitive protein and type II collagen
expression during fracture healing are potential targets for sox9 regulation. Journal of bone and mineral
research 15: 609.
26. Bosserhoff AK, Kondo S, Moser M, Dietz UH, Copeland NG, Gilbert DJ, Jenkins NA, Buettner R, Sandell
LJ (1997). Mouse CD-RAP/MIA gene: Structure, chromosomal localization, and expression in cartilage
and chondrosarcoma. Developmental Dynamics 208: 516-525.
27. Schmid R, Schiffner S, OPolka A, Grassel S, Schubert T, Moser M, Bosserhoff AK (2010). Enhanced
cartilage regeneration in MIA/CD-RAP deficient mice. Cell death and disease 1: 1-10.
28. Chen FH, Herndon ME, Patel N, Hecht JT, Tuan RS, Lawler J (2007). Interaction of cartilage oligomeric
matrix protein/thrombospondin 5 with aggrecan. The journal of biological chemistry 282: 24591-24598.
29. Hecht JT, Deere M, Putnam E, Cole W, Vertel B, Chen H, Lawler J (1998). Characterization of cartilage
oligomeric matrix protein (COMP) in human normal and pseudoachondroplasia musculoskeletal tissues.
Matrix Biology 17: 269-278.
30. http://microscopy.bitesizebio.com/articles/troubleshooting-immunohistochemistry/
31. Montes GS, Junqueira LC (1991). The use of the Picrosirius-polarization method for the study of the
biopathology of collagen. Memorias do Instituto Oswaldo Cruz 86: 1-11.
32. Junqueira LC, Bignolas G, Brentani RR (1979). Picrosirius staining plus polarization microscopy, a
specific method for collagen detection in tissue sections. The Histochemical journal 11: 447-55.
33. Carriel VS, Aneiros-Fernandez J, Arias-Santiago S, Garzon IJ, Alaminos M, Campos A (2011). A novel
histochemical method for a simultaneous staining of melanin and collagen fibers. The journal of
histochemistry and cytochemistry : official journal of the Histochemistry Society 59: 270-7.

50
34. Rich L, Whittaker P (2005). Collagen and picrosirius red staining: a polarized light assessment of fibrillar
hue and spatial distribution. Journal of morphological sciences 22: 97-104.
35. http://orteq.com/wp-content/uploads/2011/01/LR-25-5-400174.02-UK-Degradation.pdf
36. Lubowitz JH, Verdonk PC, Reid JB, 3rd, Verdonk R (2007). Meniscus allograft transplantation: a current
concepts review. Knee surgery, sports traumatology, arthroscopy: official journal of the ESSKA 15: 476-
92.
37. Rijk PC (2004). Meniscal allograft transplantation—part II: alternative treatments, effects on articular
cartilage, and future directions. Arthroscopy: the journal of arthroscopic & related surgery: official publication of the Arthroscopy Association of North America and the international Arthroscopy
Association 20: 851-9.
38. Wachsmuth L, Soder S, Fan Z, Finger F, Aigner T (2006). Immunolocalization of matrix proteins in
different human cartilage subtypes. Histology and histopathology 21: 477-485.
39. Sun Y, Mauerhan DR, Kneisl JS, James Norton H, Zinchenko N, Ingram J, Hanley Jr EN, Gruber HE
(2012). Histological examination of collagen and proteoglycan changes in osteoarthritic menisci. The open
rheumatology journal 6: 24-32.
Bijlagen
Bijlage I: Deparaffineren
Bad 1 tolueen 5'
Bad 2 tolueen 5'
Bad 3 tolueen 5'
Bad 4 isopropanol 2'
Bad 5 isopropanol 2'
Bad 6 alcohol 96% 2'
Bad 7 alcohol 96% 2'
Bad 29 leidingwater 2'
Bad 19 aqua dest. 1'
Bad 40 aqua dest. 1'

Bijlage II: Haematoxyline-eosine kleuring
Bad 1 tolueen 5’
Bad 2 tolueen 5'
Bad 3 tolueen 5'
Bad 4 isopropanol 2'
Bad 5 isopropanol 2'
Bad 6 alcohol 96% 2'
Bad 7 alcohol 96 % 2'
Bad 29 leidingwater 2'
Bad 19 aqua dest. 1'
Bad 15 haematoxyline 15”
Bad 29 leidingwater 2’
Bad 16 clarifier I 1'
Bad 29 leidingwater 1'
Bad 17 bluing reagent 1'
Bad 29 leidingwater 1'
Bad 19 aqua dest. 1'
Bad 18 eosine+phloxine 30”
Bad 29 leidingwater 2’
Bad 20 alcohol 96% 2'
Bad 21 alcohol 96% 2'
Bad 22 isopropanol 2'
Bad 8 isopropanol 2'
Bad 9 tolueen 2'
Bad 23 tolueen 2'
Bad 38 tolueen 1'

Bijlage III: Voorbehandeling met citraatbuffer (microgolf)
Citraatbuffer pH 6,0:
- 0,2 g citroenzuur in 800 ml AD
- Op pH brengen met 1M NaOH
- Aanlengen tot 1000 ml
Citraatbuffer opwarmen in een grote beker in de microgolf met deksel erop, totdat het kookt
(± 5’)
Preparaten in glazen preparatenhouder 2 x 5’ koken
20’ – 30’ afkoelen
Spoelen met PBS (+ coupes afgrenzen)
Bijlage IV: Voorbehandeling met citraatbuffer (steamer, Braun)
Citraatbuffer pH 6,0:
- 0,2 g citroenzuur in 800 ml AD
- Op pH brengen met 1M NaOH
- Aanlengen tot 1000 ml
Plastieken coplin jar vullen met citraatbuffer en 30’ opwarmen in de steamer (± 95°C)
Preparaten in coplin jar plaatsen 20’ – 30’ in de steamer
20’ – 30’ afkoelen
Spoelen met PBS (+ coupes afgrenzen)

Bijlage V: Voorbehandeling met citraatbuffer (combinatie microgolf en steamer)
Citraatbuffer pH 6,0:
- 0,2 g citroenzuur in 800 ml AD
- Op pH brengen met 1M NaOH
- Aanlengen tot 1000 ml
Citraatbuffer opwarmen in een grote beker in de microgolf met deksel erop, totdat het kookt
(± 5’)
De hete citraatbuffer voorzichtig overgieten in de plastieken coplin jar.
Preparaten in coplin jar plaatsen 20’ – 30’ in de steamer (± 95°C)
20’ – 30’ afkoelen
Spoelen met PBS (+ coupes afgrenzen)
Bijlage VI: Voorbehandeling met citraatbuffer (warmwaterbad)
Citraatbuffer pH 6,0:
- 0,1 g citroenzuur in 400 ml AD
- Op pH brengen met 5M en 1M NaOH
- Aanlengen tot 500 ml
Citraatbuffer opwarmen in het warmwaterbad in een glazen preparatenhouder met deksel erop
(20’ tot 87,5°C – 90°C)
Preparaten in de glazen preparatenhouder plaatsen en 20’ incuberen
20’ – 30’ afkoelen
Spoelen met PBS (+ coupes afgrenzen)

Bijlage VII: Protocol anti-collageen type II immuunkleuring
1e AL Anti-collageen II (Acris AMOO 616 PUN): 1/100
2e AL gebiotinyleerd rabbit anti-mouse 1/200
streptavidin-peroxidase
DAB
Methode
1) Deparaffineren + afgrenzen coupes met toluolstift
2) Immuunkleuring
Stap Tijd
Voorbehandeling met pepsine 1mg/ml Tris pH=2 10’ op
37°C
5 x wassen in PBS 2’
Voorbehandeling met 3%H2O2 in PBS 10’
Blokkingsserum (met mormal serum) *
Afnemen met cleanex
30’
AL1
2 x wassen met PBS
2h
2 x 5’
AL2 1/200
2 x wassen met PBS
30’
2 x 5’
Streptavidin-peroxidase 1/200
2 x wassen met PBS
wassen met Trisbuffer
30’
2 x 5’
5’
Chromogeen-substraat**
wassen met stromend water
10’
10’
* Blokkingsserum: 5% NRS 0,5 ml
1% BSA 0,1 g
PBS 10 ml
0,2% Tween 20 200 µl
** Chromogeen-substraat:
0.06% DAB (DAB is sterk kankerverwekkend!!) 0.003g DAB
0.03% H2O2 5µl H2O2
Trisbuffer 5ml Tris
Het DAB-poeder afwegen in zilverpapier (zodoende is er geen statische elektriciteit)
3) Tegenkleuring
haematoxyline volgens Mayer (1/4 verdund) 5”
wassen met leidingwater 5’

Bijlage VIII: pepsine (Sigma) 1mg/ml Tris pH2
Tris HCl pH 7,6 100 ml op pH2 brengen door het druppelsgewijs toevoegen van HCl (1
M).
2 mg pepsine afwegen op aluminiumpapier en oplossen in 2 ml Tris pH2.
Bijlage IX: Protocol anti-aggrecan (AbD Serotec, MCA 4722) (protocol Prof. A.
Campos, Universiteit van Granada)
1e AL muis anti-aggrecan (AbD Serotec, MCA4722) 1/250
2e AL anti-muis IgG (Vector, cat n° MP-7402)
1) Deparaffineren en afgrenzen van de coupes met stift
2) Immuunkleuring
Stap Tijd
2 x wassen met PBS 0,1 M 15’ op
37°C
Voorbehandeling met chondroitinase ABC 0,2U/ml (Sigma, ref: C3667
zie bijlage X)
1h op 37°C
3 x wassen met PBS 0,1 M 5’ op Tr
Voorbehandeling met 3% H2O2 in PBS 10’
3 x wassen met PBS 0,1 M 5’
CAS BLOCK1 (Invitrogen, cat n°: 008120) 15’
AL1 (1/250 verdunnen in PBS 0,1M + 0,3% Tween20) 1h
3 x wassen met PBS 0,1 M 5’
AL2 (Vector kit) 30’
3 x wassen met PBS 0,1 M 5’
DAB Impact (Vector kit, cat n°: SK-4105)* 2’ tot 5’
Wassen met stromend water 10’
3) Tegenkleuring
Haematoxyline volgens Mayer (onverdund) 1”
Wassen met leidingwater 1’
4) Dehydrateren met automatisch kleurtoestel MICROM (DehydraUG)
5) Monteren (Mounting medium)
*Onder de microscoop kijken wanneer de kleuromslag begint (2’ – 5’). Wanneer er na 5’ niets
te zien is, is de kleuring mislukt. De geselecteerde incubatietijd voor DAB Impact is 1’ 45”. 1 CAS BLOCK is een universeel blocking agens om de niet-specifieke achtergrondkleuring te
reduceren bij immuunkleuringen.

Bijlage X: Chondroitinase ABC (Sigma, C3667) 0,2U/ml in oplossing A
Oplossing A:
- 80 ml AD
- 0,6 g Tris-base (Sigma) op pH8 brengen door het druppelsgewijs
- 1,62 g Na-acetaat (Merck) toevoegen van HCl (1 M).
0,02 g BSA toevoegen en alles goed laten oplossen!
40 µl Chondroitinase ABC (5U/ml) + 960 µl oplossing A Chondroitinase ABC 0,2U/ml

Bijlage XI: Protocol anti-SMA immuunkleuring (monoclonal antibody, Dako, HHF35)
1e AL mouse anti-human smooth muscle actine: 1/400
2e AL gebiotinyleerd rabbit anti-mouse: 1/200
streptavidin-peroxidase
DAB
Methode
1 Deparaffineren + afgrenzen met toluolstift
2 Immuunkleuring
Stap Tijd
Voorbehandeling met 3% H2O2 in PBS 10’
Blokkingsserum*
Afnemen met een cleanex
30’
AL1
2 x wassen met PBS
2h
2 x 5’
AL2 (1/200)
2 x wassen met PBS
30’
2 x 5’
Streptavidin-peroxidase (1/200)
2 x wassen met PBS
Wassen met Trisbuffer
30’
2 x 5’
5’
Chromogeen-substraat**
Wassen met stromend water
10’
10’
* Blokkingsserum: 5% NRS 0,5 ml
1% BSA 0,1 g
PBS 10 ml
0,2% Tween20 200 µl
** Chromogeen-substraat:
0.06% DAB (DAB is kankerverwekkend!!!)
0.03% H2O2
Trisbuffer
Het DAB-poeder afwegen in zilverpapier (zodoende is er geen statische elektriciteit)
3 Tegenkleuring
Haematoxyline volgens Mayer (onverdund) 1”
Wassen met leidingwater 5’
4 Ontwateren + monteren met Mounting media

Bijlage XII: Protocol anti-S100 immuunkleuring (polyclonal antibody, Dako, Z0311)
1e AL rabbit anti-human S100: 1/1000
2e AL gebiotinyleerd swine anti-rabbit: 1/200
streptavidin-peroxidase
DAB
Methode
1 Deparaffineren
2 Antigen Retrieval (volgens bijlage VI)
3 Afgrenzen met toluolstift
4 Immuunkleuring
Stap Tijd
Voorbehandeling met 3% H2O2 in PBS 10’
Blokkingsserum*
Afnemen met een cleanex
30’
AL1
2 x wassen met PBS
2h
2 x 5’
AL2 (1/200)
2 x wassen met PBS
30’
2 x 5’
Streptavidin-peroxidase (1/200)
2 x wassen met PBS
Wassen met Trisbuffer
30’
2 x 5’
5’
Chromogeen-substraat**
Wassen met stromend water
10’
10’
* Blokkingsserum: 5% NSS 0,5 ml
1% BSA 0,1 g
PBS 10 ml
0,2% Tween20 200 µl
** Chromogeen-substraat:
0.06% DAB (DAB is kankerverwekkend!!!)
0.03% H2O2
Trisbuffer
Het DAB-poeder afwegen in zilverpapier (zodoende is er geen statische elektriciteit)
5 Tegenkleuring
Haematoxyline volgens Mayer (onverdund) 1”
Wassen met leidingwater 5’
6 Ontwateren + monteren met Mounting media

Bijlage XIII: Protocol anti-Ki67 immuunkleuring
1e AL Anti-Ki67 (Dako, M7240, clone MIB-1): 1/50
2e AL gebiotinyleerd rabbit anti-mouse 1/200
Streptavidin-peroxidase
DAB
Methode
1 Deparaffineren
2 Antigen retrieval (volgens bijlage VI)
3 Afgrenzen coupes met toluolstift 4 Immuunkleuring
Stap Tijd
Voorbehandeling met 3%H2O2 in PBS 10’
Blokkingsserum (met NRS)*
Afnemen met cleanex
30’
AL1 1/50
2 x wassen met PBS
2h
2 x 5’
AL2 1/200
2 x wassen met PBS
30’
2 x 5’
Streptavidin-peroxidase 1/200
2 x wassen met PBS wassen met Trisbuffer
30’
2 x 5’ 5’
Chromogeen-substraat**
wassen met stromend water
10’
10’
* Blokkingsserum: 5% NRS 0,5 ml
1% BSA 0,1 g
PBS 10 ml
0,2% Tween 20 200 µl
** Chromogeen-substraat:
0.06% DAB (DAB is sterk kankerverwekkend!!) 0.003g DAB
0.03% H2O2 5µl H2O2
Trisbuffer 5ml Tris
Het DAB-poeder afwegen in zilverpapier (zodoende is er geen statische elektriciteit)
5 Tegenkleuring
haematoxyline volgens Mayer (onverdund) 1”
wassen met leidingwater 1’ 6 Dehydrateren + monteren met Mounting media

Bijlage XIV: Protocol anti-Stro1 immuunkleuring
1e AL muis anti-Stro1 (R&D Systems, MAB1038) 1/50, 1/100
2e AL gebiotinyleerd RAM 1/200
1 Deparaffineren en afgrenzen van de coupes met stift
2 Immuunkleuring
Stap Tijd
Voorbehandeling met 3% H2O2 verdund in PBS 10’
BS
Afnemen met cleanex
30’ op Tr
AL1 (verdund in VB) 1h op Tr
2 x wassen met PBS 0,1 M 2 x 5’
AL2 (verdund in PBS) 30’
2 x wassen met PBS 0,1 M 2 x 5’
Streptavidin-peroxidase (HRP) (1/200 verdund in VB) 30’
2 x wassen met PBS 0,1 M 2 x 5’
Wassen met Trisbuffer 5’
Chromogeen substraat* (DAB) 10’
Wassen met stromend water 10’
BS: PBS 9,8 ml
0,2% Tween 20 200 µl
5% NRS 500 µl
BSA 0,1 g
VB: 9 ml PBS + 1 ml BS
*Chromogeen substraat 5 ml Trisbuffer
5 µl 0,03% H2O2
0,003g 0,06% DAB (DAB is sterk kankerverwekkend!)
(DAB afwegen in zilverpapier, zodoende is er geen statische elektriciteit)
3 Tegenkleuring:
Haematoxyline volgens Mayer (onverdund) 1”
Wassen met stromend water 1’
4 Dehydrateren met automatisch kleurtoestel MICROM (DehydraUG)
5 Monteren (Mounting media)

Bijlage XV: Protocol anti-CD34 immuunkleuring
1e AL Anti-CD34 (Dako, M7165, clone QBEnd10): 1/100
2e AL gebiotinyleerd rabbit anti-mouse 1/200
Streptavidin-peroxidase
DAB
Methode
1 Deparaffineren
2 Antigen retrieval (volgens bijlage VI) 3 Afgrenzen coupes met toluolstift
4 Immuunkleuring
Stap Tijd
Voorbehandeling met 3%H2O2 in PBS 10’
Blokkingsserum (met NRS)*
Afnemen met cleanex
30’
AL1 1/100
2 x wassen met PBS
2h
2 x 5’
AL2 1/200
2 x wassen met PBS
30’
2 x 5’
Streptavidin-peroxidase 1/200
2 x wassen met PBS
wassen met Trisbuffer
30’
2 x 5’
5’
Chromogeen-substraat**
wassen met stromend water
10’
10’
* Blokkingsserum: 5% NRS 0,5 ml
1% BSA 0,1 g
PBS 10 ml
0,2% Tween 20 200 µl
** Chromogeen-substraat:
0.06% DAB (DAB is sterk kankerverwekkend!!) 0.003g DAB
0.03% H2O2 5µl H2O2
Trisbuffer 5ml Tris
Het DAB-poeder afwegen in zilverpapier (zodoende is er geen statische elektriciteit)
5 Tegenkleuring
haematoxyline volgens Mayer (1/4 verdund) 5”
wassen met leidingwater 1’
6 Dehydrateren + monteren met Mounting media

Bijlage XVI: protocol picrosirius rood (protocol Tissue engineering group, Universiteit
van Granada, Prof. A. Campos)
Oplossingen:
Sirius red F3B (Sigma-Aldrich, ref: 365548) 0,2 g
Picric acid (Sigma-Aldrich, ref: 80450) verzadigde oplossing 100 ml
Harris haematoxyline
Protocol:
1. Deparaffineren van de coupes in xyleen en hydrateren in een dalende alcoholreeks (100 –
70°), eindigend in AD.
2. Coupes kleuren met 0,2 % sirius red F3B in een verzadigde oplossing van picric acid (30
min bij kamertemperatuur).
3. 2 x wassen met AD.
4. Tegenkleuren met Harris haematoxyline (3 min).
5. Wassen met stromend leidingwater (3 – 5 min).
6. Dehydrateren in een stijgende reeks van ethanol.
7. Reinigen met xyleen (2 x) en monteren met een hydrofobe medium.

Bijlage XVII: Dehydrateren
Bad 29 leidingwater 4'
Bad 20 alcohol 96% 2'
Bad 21 alcohol 96% 2'
Bad 22 isopropanol 2'
Bad 8 isopropanol 2'
Bad 9 tolueen 2'
Bad 23 tolueen 2'
Bad 38 tolueen 1'
Bijlage XVIII: Stijgende alcoholreeks
alcohol 30% 1h
alcohol 50% 1h
alcohol 70% 1h
Disolol 99% + toluidineblauw 1h
Isopropanol 100% 1h
Isopropanol + tolueen 1h
Tolueen EXACT 30 min

Bijlage XIX: Tabel 3: Voorstelling van de toegekende scores voor de verschillende
kleuringen, weergegeven per populatie.
Tijdstip Celdensiteit (HE kleuring)
aantal vasculaire structuren*** (CD34 kleuring)
CD34+ cellen****
Aggrecan* Picrosirius red* (polarisatie microscopie)
Ort 1 2 3 4 5 6 7 8 9
10 11 12 13 14 15 16 17 18 19 20
29 wk 10 wk
1 j, 19d 1 j, 1 m 50 wk
n/a 1 j, 1 m
1 j 11 m 11 m
1 j 1 j
10 m 2 j, 4 m
9 m 1 j 4 m
n/a 2 j
2 j, 2 m 4 m
DZ* SZ** 0 0 3 1 4 4 3 4 2 1 2 4 3 0 0 2 0 3 3 0
0 0 2 1 2 3 2 2 2 0 3 3 1 0 1 3 0 3 3 0
3 2 1 0 0 2 0 2 2 2 1 1 1 1 1 3 3 1 1 2
0 3 1 2 2 3 3 2 2 2 2 1 1 3 0 2 2 3 2 2
5 5
n/a 5 5 5
n/a 5 5 5 5 5 3 5 0 5 5 5 5 5
2 3
n/a 1 3 2
n/a 2 4 2 5 2 1 3 0 3 2 2 2 2
Df 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39
Viabel 40
1 j, 1 m
11 m n/a 14 j
11 m 1 j, 5 m
n/a 11 m n/a
1 j, 2 m 11 m 5 m n/a 9 m 8 m
1 j, 6 m n/a 5 m 5 m
2 j, 10
1 2 1 2 0 4 2 1 5 2 5 2 3 0 2 1
n/a 0 1
3
2 2 4 5 0 3 4 2 4 2 4 5 3 5 5 4
n/a 0 5
2
0
n/a 2 0 0
n/a 5 1 0 0 0 1 0 0 0 3 3 0 0
0
0
n/a 3 0 0
n/a 3 0 0 0 0 1 0 0 0 3 3 0 0
0
1 2 2 1 0 0 0 3 2 1 1 2 2 0 1 0 3 2 2
1
2 2 2 1 2 2 2 1 2 3 1 2 2 2 3 2 2 2 2
2
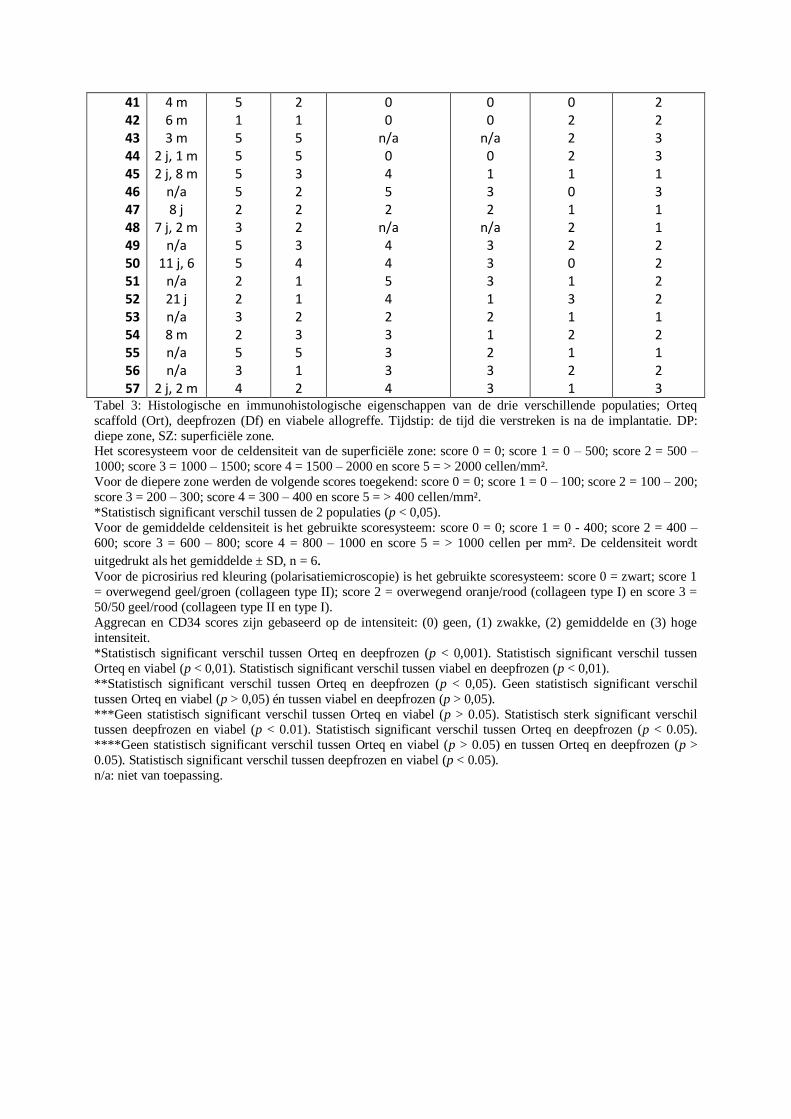
41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57
4 m 6 m 3 m
2 j, 1 m 2 j, 8 m
n/a 8 j
7 j, 2 m n/a
11 j, 6 n/a 21 j n/a 8 m n/a n/a
2 j, 2 m
5 1 5 5 5 5 2 3 5 5 2 2 3 2 5 3 4
2 1 5 5 3 2 2 2 3 4 1 1 2 3 5 1 2
0 0
n/a 0 4 5 2
n/a 4 4 5 4 2 3 3 3 4
0 0
n/a 0 1 3 2
n/a 3 3 3 1 2 1 2 3 3
0 2 2 2 1 0 1 2 2 0 1 3 1 2 1 2 1
2 2 3 3 1 3 1 1 2 2 2 2 1 2 1 2 3
Tabel 3: Histologische en immunohistologische eigenschappen van de drie verschillende populaties; Orteq
scaffold (Ort), deepfrozen (Df) en viabele allogreffe. Tijdstip: de tijd die verstreken is na de implantatie. DP:
diepe zone, SZ: superficiële zone. Het scoresysteem voor de celdensiteit van de superficiële zone: score 0 = 0; score 1 = 0 – 500; score 2 = 500 –
1000; score 3 = 1000 – 1500; score 4 = 1500 – 2000 en score 5 = > 2000 cellen/mm².
Voor de diepere zone werden de volgende scores toegekend: score 0 = 0; score 1 = 0 – 100; score 2 = 100 – 200;
score 3 = 200 – 300; score 4 = 300 – 400 en score 5 = > 400 cellen/mm².
*Statistisch significant verschil tussen de 2 populaties (p < 0,05).
Voor de gemiddelde celdensiteit is het gebruikte scoresysteem: score 0 = 0; score 1 = 0 - 400; score 2 = 400 –
600; score 3 = 600 – 800; score 4 = 800 – 1000 en score 5 = > 1000 cellen per mm². De celdensiteit wordt
uitgedrukt als het gemiddelde ± SD, n = 6. Voor de picrosirius red kleuring (polarisatiemicroscopie) is het gebruikte scoresysteem: score 0 = zwart; score 1
= overwegend geel/groen (collageen type II); score 2 = overwegend oranje/rood (collageen type I) en score 3 =
50/50 geel/rood (collageen type II en type I).
Aggrecan en CD34 scores zijn gebaseerd op de intensiteit: (0) geen, (1) zwakke, (2) gemiddelde en (3) hoge
intensiteit. *Statistisch significant verschil tussen Orteq en deepfrozen (p < 0,001). Statistisch significant verschil tussen
Orteq en viabel (p < 0,01). Statistisch significant verschil tussen viabel en deepfrozen (p < 0,01).
**Statistisch significant verschil tussen Orteq en deepfrozen (p < 0,05). Geen statistisch significant verschil
tussen Orteq en viabel (p > 0,05) én tussen viabel en deepfrozen (p > 0,05).
***Geen statistisch significant verschil tussen Orteq en viabel (p > 0.05). Statistisch sterk significant verschil tussen deepfrozen en viabel (p < 0.01). Statistisch significant verschil tussen Orteq en deepfrozen (p < 0.05).
****Geen statistisch significant verschil tussen Orteq en viabel (p > 0.05) en tussen Orteq en deepfrozen (p >
0.05). Statistisch significant verschil tussen deepfrozen en viabel (p < 0.05).
n/a: niet van toepassing.
Top Related