







Languages
Pages
Legal


Genomika hospodářských zvířat
1
1.
2

Mapování QTL pomocí
vazbové nerovnováhy
Linkage Disequilibrium (LD)
3

Mapování QTL pomocí vazbové nerovnováhy
• založeno na existenci LD mezi QTL a
markerem v náhodně se pářící populaci.
• Z efektu testovaného markeru na fenotypový
znak usuzujeme na přítomnost a efekt QTL.
4

Faktory ovlivňující vznik a degradaci LD
v populaci
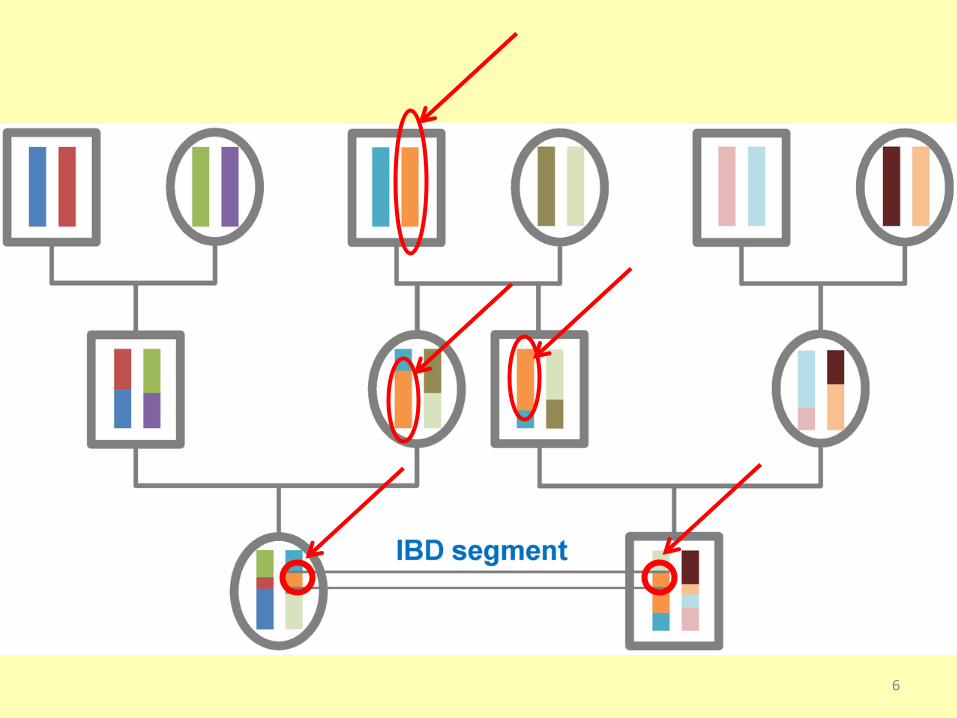
IBD, Identity By Descends
Úsek chromozómu se shodnými alelami (haplotyp),
jaké měl předek, u kterého vznikla příčinná mutace
ovlivňující variabilitu užitkového znaku.
Je potomkům předáván bez rekombinace.
5
6

Úplná LD
Není LD
Mapová
vzdálenost
7
LD je kontinuálně erodována
rekombinací

Faktory ovlivňující velikost a rozsah LD v populaci
DŮLEŽITÉ
Náhodný posun (random drift)
U populací s relativně malým počtem jedinců ovlivňuje frekvence
alel a holotypů. Čím je menší Ne populace tím větší je efekt driftu
(viz výše).
8

Faktory ovlivňující velikost a rozsah LD v populaci
DŮLEŽITÉ
Selekce
Vliv selekce na LD závisí na směru, intenzitě a délce trvání
selekce. Selekce redukuje genetickou variabilitu v následující
generaci a produkuje negativní LD mezi lokusy. Když je selekce
aplikovaná na určitý lokus, sousední lokusy, které jsou s tímto
lokusem v LD budou mít zvýšenou LD. Tzv. hitchhiking efekt.
9

Faktory ovlivňující velikost a rozsah LD v populaci
DŮLEŽITÉ
Křížení
Tvoří nové falešné LD mezi lokusy, které dříve nebyly v LD
v původních populacích a mění rozsah LD pro lokusy které byly v LD
v původních populacích.
Tyto falešné LD se rychle ztrácejí v následujících generacích.
10

Hlavní faktory ovlivňující LD u hospodářských zvířat
DŮLEŽITÉ
V populacích hospodářských zvířat je pro LD
rozhodující
velikost populace.
Efektivní velikost
populace je u většiny hospodářských zvířat malá, a proto je
rozsah LD
velký!!!11

Efektivní velikost populace Ne
je počet jedinců, který zapříčiní stejný přírůstek inbreedingu, jako
kdyby se rozmnožovali jako ideální populace.
Jinak: máme velkou populaci, která není ideální, např. je v ní větší
počet samců. V ní je přírůstek inbreedingu stejný, jako v menší
populaci, která je ideální.
Ne N
1
Ne =
2 F
4 Nm Nf
Ne =
Nm + Nf
stejný počet
samců
a samic
nestejný
počet samců
a samic
Tato metoda je založena na analýze poměru pohlaví,
existuje více metod. 12

Run of Homozygosity ROH
DŮLEŽITÉ
Inbreeding koeficient F je pravděpodobnost, že na náhodně
vybraném lokusu v populaci je pár alel „identic by descent“, IBD.
F se obvykle počítá z rodokmenu, pravděpodobnost že pár alel
je IBD je odhadována statisticky.
S použitím high-density SNP analýzy může být míra inbreedingu
pro populaci určena, i když není rodokmen k dispozici.
Skutečná hodnota inbreedingu pro jedince může být vypočtena z
celogenomové sekvence.
Dva postupy:
1/ testování markerů identických by state IBS
2/ runs of homozygosity ROH13

Run of Homozygosity ROH
ROH jsou úseky DNA s nepřerušenými úseky lokusů, které jsou
homozygotní u jedince, ale polymorfní v populaci.
Samozřejmě, to nemusí znamenat, že každý takový úsek zdědil
jedinec od společného předka bez rekombinace.
Nicméně, i když ROH může vzniknout z různých důvodů,
předpokládá se, že hlavním důvodem je inbreeding.
Pro jedince platí, že
koeficient inbreedingu je podíl genomu, který
je „identical by descent“, IBD, což lze určit z
analýzy „run of homozygosity“, ROH.
14


Inbrední deprese
Studie z poslední doby ukazují následující.
Nebyl zjištěn efekt druhu, ale populace (plemene). Tj. deprese
závisí více na frekvencích alel segregujících uvnitř plemen, než na
fyziologických specifikách druhů.
U hosp. zvířat jakákoliv selektovaná vlastnost může být postižena
inbrední depresí.
16

Inbrední deprese
Drůbež
Produkční systémy jsou založeny téměř výhradně na křížení.
Inbreedingová deprese může být z ekonomického hlediska pominuta.
Mléčný skot
Vět. čistokrevná plemena. Proto deprese musí být pod kontrolou.
U ostatních druhů je situace intermediární.
17

Inbrední deprese
U produkce mléka se odhaduje pokles 0,37% na 1% inbreedingu.
U holštýna je roční přírůstek inbreedingu ca 0,12%, tj. za generaci
0,6% při generačním intervalu 5 let.
Potom, je-li roční genetický zisk u holštýna ca 100 kg mléka na
laktaci, tj. 1% současného fenotypového průměru 10 tis. kg. Inbrední
deprese za rok potom odpovídá 0,044% fenotypového průměru, to je
4,4% genetického zisku.
To sice není mnoho pro jednu vlastnost, ale počítáme-li s depresí i u
ostatních vlastností, je ekonomický význam nepochybný.18

2.
19

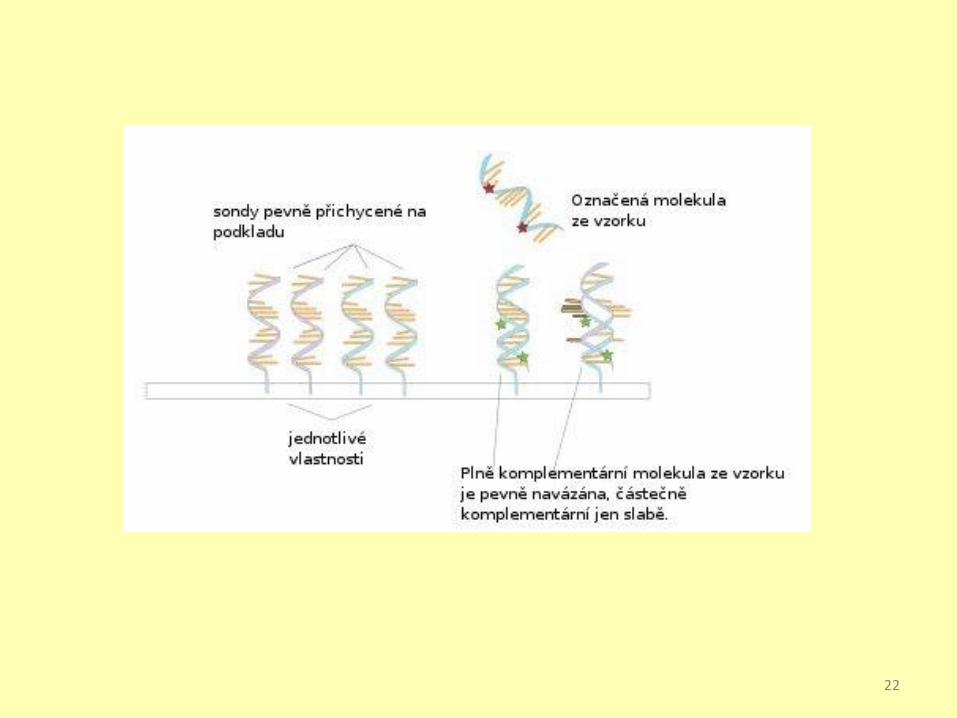
DNA chips, DNA microarrays
20

21
Získání DNA
Kontrola kvality
Celogenomová
amplifikace
Fragmentace
Hybridisace
Scan
Softwarová analýza
Data
22
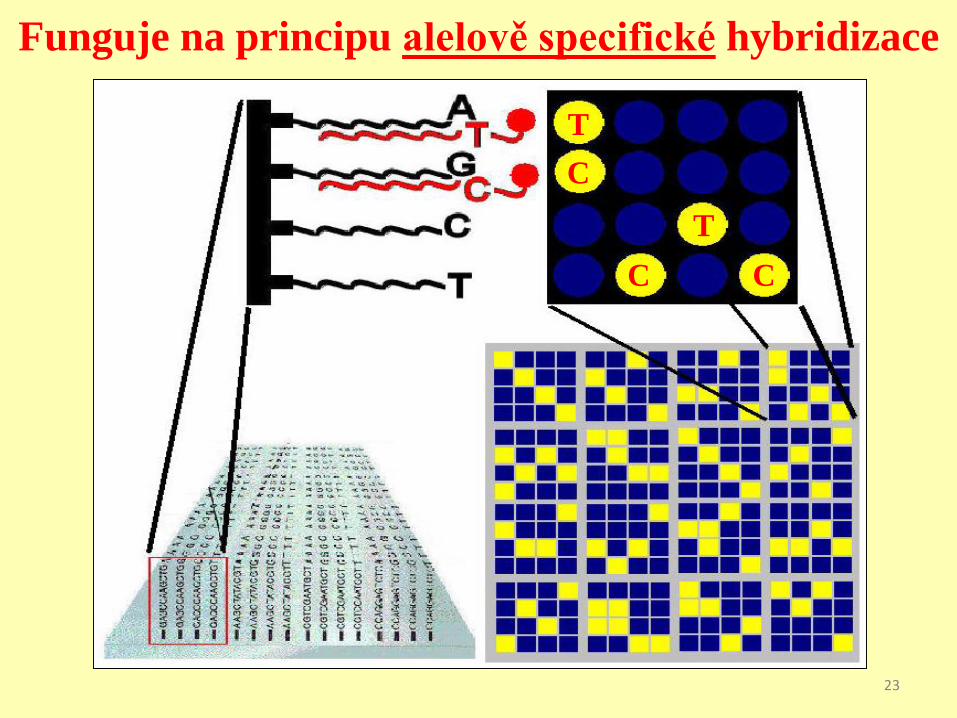
Funguje na principu alelově specifické hybridizace
23
T
C
C C
T

Řada aplikací. Nás zajímá zvl.
identifikace alel SNP.
Následné využití při genotypizaci, forenzní analýze, odhadu
predispozice k chorobám, identifikaci mutací v zárodečných liniích,
somatických mutací v nádorech.
Pro naše účely je důležitá genotypizace alel SNP a následná
analýza vazbové nerovnováhy.24

Často se používají čipy fy Illumina (říjen 2016):
Bovinní SNP čipy:
BovineLD Genotyping BeadChip: 7 931 SNPs
BovineSNP50 v3 DNA Analysis BeadChip: 53 714 SNPs
BovineHD DNA Analysis Kit: 777 962 SNPs
Sekvenační technologie:
TruSeq Bovine Parentage Sequencing: panel 200 ISAGem
doporučených SNPs pro parentitu plus 66 SNP asociovaných s
recesivními dědičnými chorobami
25
Prase: PorcineSNP60 DNA Analysis Kit v2: 64 232 SNP

3.
26

Genomová selekce
27
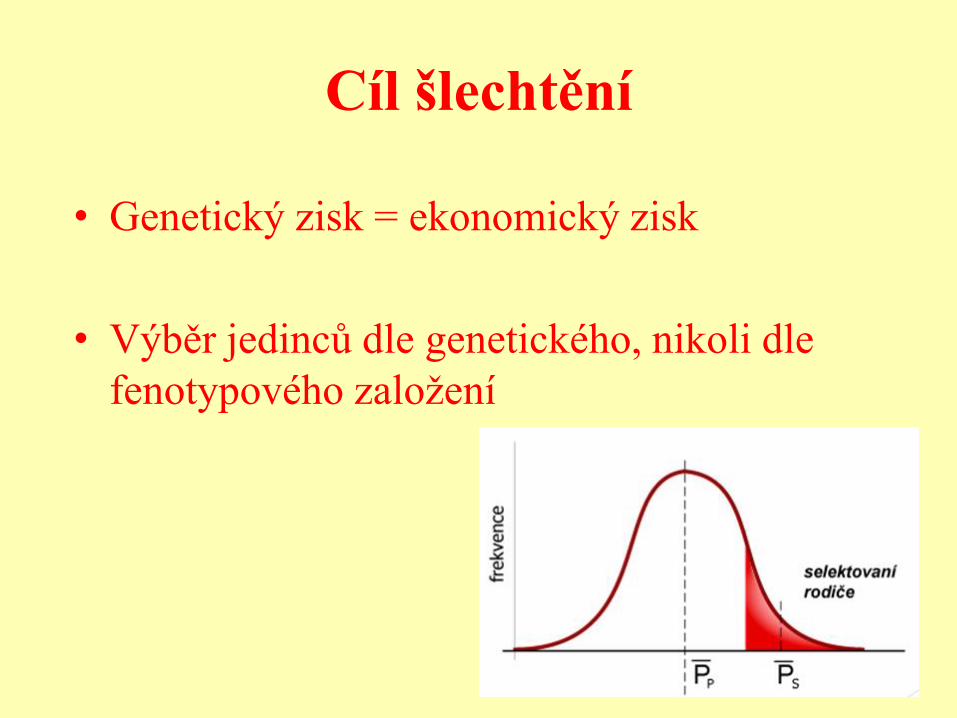
Cíl šlechtění
• Genetický zisk = ekonomický zisk
• Výběr jedinců dle genetického, nikoli dle
fenotypového založení
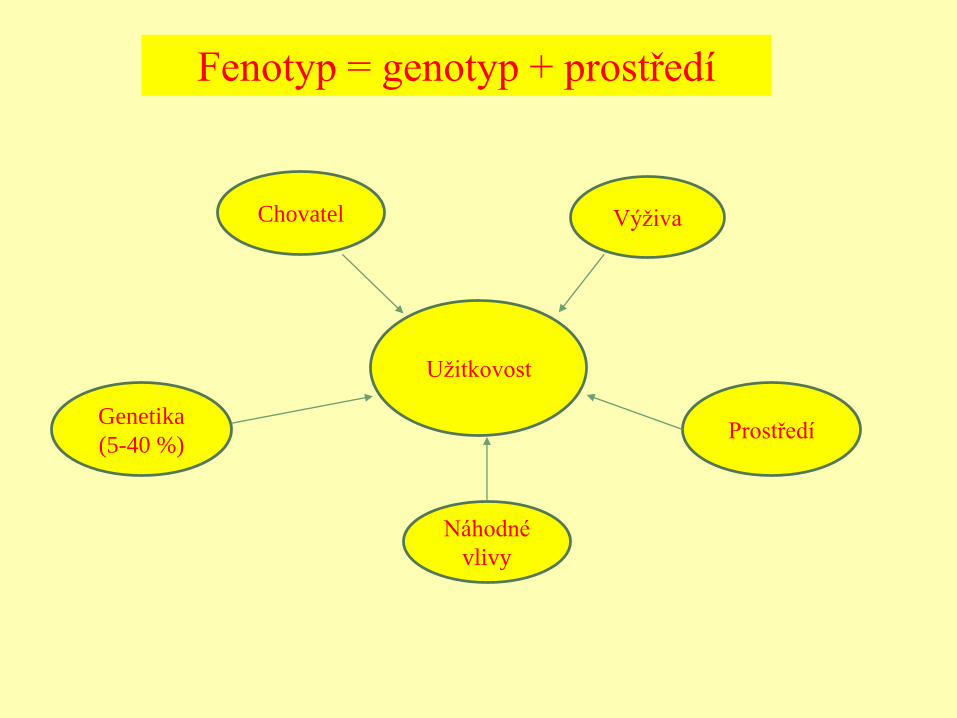
Užitkovost
Chovatel Výživa
Genetika
(5-40 %)Prostředí
Náhodné
vlivy
Fenotyp = genotyp + prostředí

Plemenná hodnota
• Odhad genetického založení jedince pro
odchylku v užitkové vlastnosti od průměru
vrstevníků
• Aditivní účinek genů
• Zohlednění příbuznosti všech jedinců
vstupujících do modelu
• RPH – PH převedeny na průměr
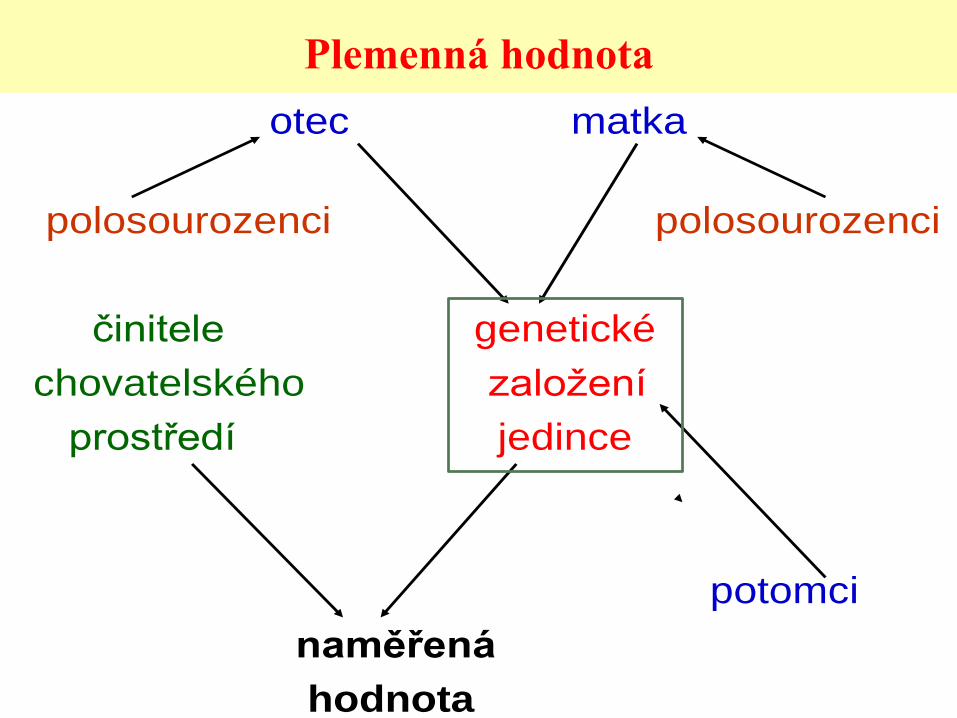
otec matka
polosourozenci polosourozenci
činitele genetické
chovatelského založení
prostředí jedince
potomci
naměřená
hodnota
Plemenná hodnota

Genomová selekce
Pracuje na základě rozdělení celého genomu do
chromozómových úseků definovaných sousedními
markery a následného sledování efektů těchto
chromozómových úseků.
32

Genomová selekce
Využívá předpokladu, že efekty chromozómových úseků budou
shodné v celé populaci, protože markery jsou v LD s QTL mezi
markery (mají stejnou vazbovou fázi).
GS předpokládá, že všechny markery mohou být ve vazbě ke genu
ovlivňujícím vlastnost.
Proto hustota markerů musí být dostatečná, aby zajistila, že
všechny QTL jsou v dostatečném LD s markery nebo haplotypy
markerů.
To umožňují SNP.
33

Praktické použití genomové selekce
1. Odhad efektů chromozómových segmentů v referenční
populaci. Zvířata referenční populace tedy musí mít
stanoveny genotypy markerů a mít záznamy o užitkovosti.
2. Předpověď GPH pro zvířata mimo referenční populaci, pro
mladé býčky, jalovice, kanečky atd. Tito kandidáti mají
k dispozici pouze genotypy markerů, ale nemají k dispozici
údaje o vlastní užitkovosti nebo užitkovosti potomstva.
Formální kvantitativní genetika počítá s tím, že potomek
zdědí polovinu vlastností od každého z rodičů. Ve skutečnosti
to tak docela neplatí. 34

Praktické použití genomové selekce
Všechny SNP efekty jsou simultánně odhadovány v referenční
populaci, která je genotypována a fenotypována s použitím
statistického modelu (předpokládáme 50 000 SNP):
yi = m + Xi1 x b1 + X2i x b2 + … + X50000i x b50000 + ei
yi je fenotyp zvířete i; m průměr; Xi1 genotyp zvířete i pro marker
1; ei residuální efekt
35

Praktické použití genomové selekce
Metoda používá BLUP k odhadu SNP efektů, nazývá se někdy
SNP-BLUP.
36
Genomická plemenná hodnota selekčního kandidáta „j“ je
předpovídána jako:
GEBVj = X1j x 1 + X2j x 2 + … + X50000j x 50000
1 je odhad efektu SNP1, X1j genotyp zvířete j pro SNP 1

Ale:
Vlastní výpočet plemenné hodnoty se
provádí např. metodou:
• BLUP-AM
• RR BLUP
• G BLUP
• ssG BLUP
• Blending ss G BLUP 37
Při praktickém použití genomové
selekce se používají tyto metody

V tradiční BLUP je PH odhadována s použitím
fenotypů a rodokmenových příbuzností.
V GBLUP je GPH odhadována s použitím
fenotypů a genomových příbuzností, které jsou
založeny na celogenomových hustých
markerových datech.
38

Genomová příbuznost mezi zvířaty 1 a 2 je
počítána jako korelace mezi jejich SNP genotypy
Xj1 a Xj2 přes všechny SNP j.
GBLUP je tedy velmi podobná tradiční BLUP, ale
rodokmenové příbuznosti jsou
nahrazeny genomovými.
Mohou být použity tradiční metody výpočtu a
software. 39

Rodokmenová příbuznost sourozenců je 0,5
tedy by měli sdílet 50% společných alel. Ve
skutečnosti mohou sdílet 40% nebo 60%
společných alel. Tato odchylka může být
detekována genotypováním markerů.
GBLUP je přesnější, protože genomová
příbuznost je přesnější, než rodokmenová.40

Plemenná hodnota BLUP – AMBest Linear Unbiased Prediction – Animal Model
nejlepší lineární nevychýlená předpověď – model zvířete
Genetické založení jedince pro konkrétní vlastnost
Y = X.b + Z.a + e
Y vektor naměřených užitkovostí (v. pozorování)
X, Z matice plánu pokusu pro systematické chovatelské
prostředí (X) a pro náhodné efekty včetně plemenných
hodnot (Z)
b vektor odhadovaných fixních efektů
a vektor odhadovaných náhodných efektů
e vektor náhodných vlivů prostředí (v. reziduí)

Plemenná hodnota
A je matice příbuznosti
b je vektor odhadovaných fixních efektů,
a vektor předpovídaných náhodných efektů,
plemenná hodnotaa je poměr residuálního a aditivně genetického rozptylu,
a = s2e / s2
a

Co vstupuje do odhadu
Před genomikou
• Původ, tj. příbuzenské vztahy
• Kontrola užitkovosti
Po zavedení genomiky
• Původ, tj. příbuzenské + genomické vztahy
• Kontrola užitkovosti

Vícekroková metoda
• Nejrozšířenější
• V praxi od 2010
• Předpověď PH pouze pro genotypizovaná zvířata
Nevýhody:
• U malých populací
• Neschopnost porovnat genotypované a negenotypované
jedince

Vícekroková metoda
• 1. krok – pomocí statistických analýz přiřazeny
užitkové hodnoty jednotlivých SNP (jakou
užitkovost má jaká mutace v genotypu jedince)
• 2. krok - ,,sečteny“ jednotlivé účinky genů –
sečteny účinky jednotlivých SNP a sestavena
genomická PH (GPH) jedince

Jednokroková metoda
• Předpověď PH pro celou populaci
• Neodhaduje účinky jednotlivých SNP
• Upřesnění příbuzenských vztahů
• Pro genotypované a negenotypované jedince, tedy
porovnání pořadí• Místo A-1 je H-1 = (G + A)
• Přesnější, jednodušší
• Mírné zvýšení spolehlivosti i pro negenotypovanéjedince, tedy
zpřesnění odhadu PH

Jednokroková metoda
ssGBLUP
užitkovost
Y = Xb + Zu + e
H
G+A

Při ssGBLUP
se upravuje matice příbuznosti H, a to
spojením genomové matice příbuznosti G s
rodokmenovou maticí příbuznosti A
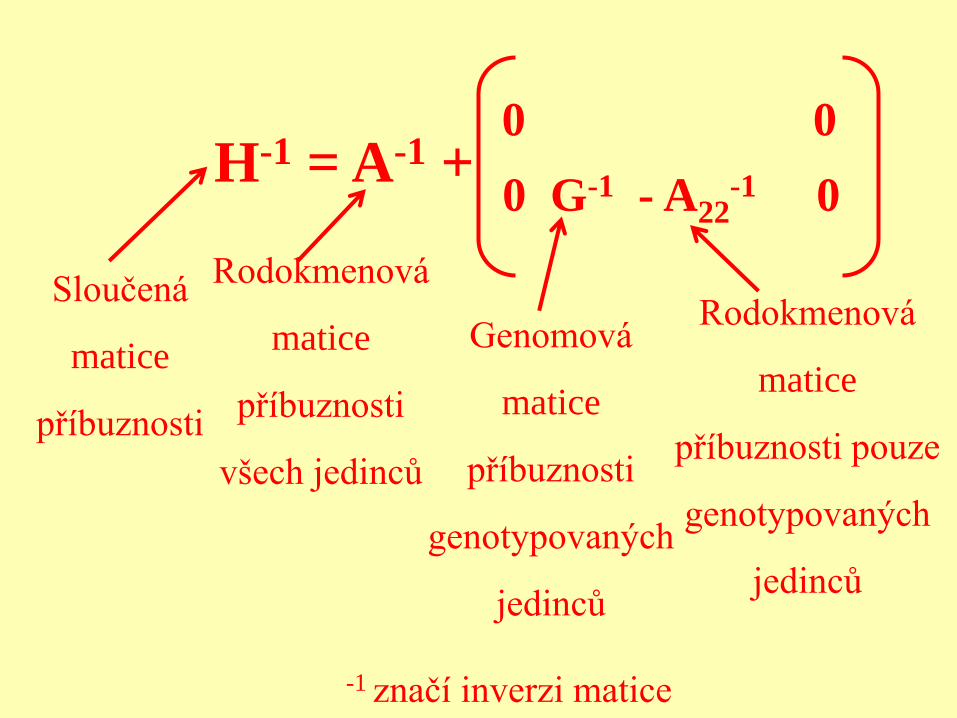
H-1 = A-1 + 0 0
0 G-1 - A22-1 0
Sloučená
matice
příbuznosti
Rodokmenová
matice
příbuznosti
všech jedinců
Genomová
matice
příbuznosti
genotypovaných
jedinců
Rodokmenová
matice
příbuznosti pouze
genotypovaných
jedinců
-1 značí inverzi matice

ssGBLUP
H je sloučená matice příbuznosti, spojuje rodokmenovou (A) a genomickou (G) příbuznost.
1

Imputace
Na základě známých genotypů SNP zvířete je určen
jeho haplotyp, protože stejný haplotyp byl pozorován u
jiných zvířat.
Tedy: chybějící genotyp může být přečten z genotypu
jiných zvířat se stejným haplotypem.
Software: Beagle, Fimpute, Alphaimpute.
51

Budoucnost
ss odhad užívající nelineární
statistickou metodu a data ze
sekvenace!
52

Faktory ovlivňující přesnost GS
Přesnost odhadu GPH závisí na několika parametrech.
1) Rozsah a velikost LD mezi markery a QTL.
2) Počet zvířat s fenotypy a genotypy v referenční populaci,
ze které jsou efekty SNP odhadovány.
3) Příbuznost (genetické propojení) mezi referenční populací
(která je genotypovaná a fenotypovaná) a populací s kandidátními
zvířaty (která je genotypovaná, ale není fenotypovaná).
4) Dědivost (heritabilita - h2) sledovaných znaků.
5) Distribuce QTL efektů.
První tři faktory lze ovlivnit, druhé dva ne!
53

První komerční čip v r. 2008 – Illumina BovineSNP50
BeadChip.
Dostupné pro všechny důležité druhy hosp. zvířat, ovce, prase,
kůň, kur, koza, pstruh, losos, atd.
Počet SNP v první generaci čipů 50 – 60 tis.
V souč. (2015) je jen pro skot nabízeno 11 komerčních čipů od
3 největších firem, Illumina, Neogen-GeneSeek, Affymetrix.
Dále se nabízí tzv. custom čipy.54

4.
55

Celogenomové asociační studie
GWAS
Genome-Wide Association Study
56

Graf znázorňuje několik silně asociovaných podezřelých lokusů.
Každý bod představuje SNP, osa X je genomická
lokalizace, osa Y úroveň asociace.
Příklad ukazuje asociaci pro mikrocirkulaci, body nahoře indikují genetické
varianty, častěji nalézané u jedinců konstrikcemi v malých cévách.57
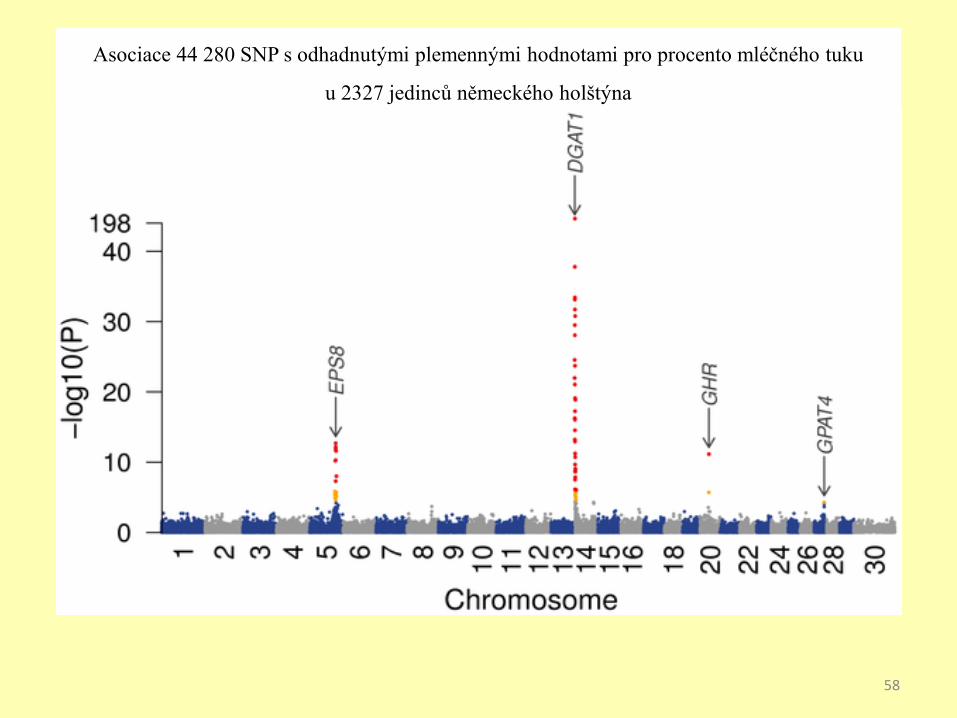
Asociace 44 280 SNP s odhadnutými plemennými hodnotami pro procento mléčného tuku
u 2327 jedinců německého holštýna
58

5.
59

Ještě pár poznámek
60

Customer chips
61

62

Softwarové nástroje pro
zpracování dat z analýzy SNP
PLINK
Je považován za určitý standard pro management dat.
Atd…
63

Do 10 let (?) se bude u hospodářských zvířat
provádět sekvenování kompletního genomu.
64

65
6.

Současný stav genomové selekce
66

Genomová selekce teoreticky
popsána 2001.
67

V ČRU holštýnského skotu probíhá ve spolupráci Svazu
chovatelů holštýnského skotu, ČMSCH a.s., Plemdat s.r.o.
a VÚŽV.
Zahájen byl výpočet GPH u mladých býčků a jalovic,
jednokroková metoda.
Proběhla validace GPH pro znaky produkce v
Interbullu.
Rutinní výpočet GPH pro znaky produkce a somatické
buňky realizuje Plemdat s.r.o.
Genotypováno bylo 450 starších krav. Cílem je
zpřesnění národního odhadu GPH. 68

V ČR
U holštýnského skotu jsou výsledky odhadu GPH
pravidelně publikovány na stránkách Svazu chovatelů
holštýnského skotu.
69
70
71

PO
R
LIN
-REG
JMÉN
O B
ÝKA
RO
K N
AR
OZE
NÍ
OR
G
DĚD
. VA
DY
OTE
C J
MÉN
O
OM
JM
ÉNO
gSIH
DC
ER P
RO
D
STÁ
D P
RO
D
R -
PR
OD
DI-
MLK
MLÉ
KO
KG
TUK
KG
TUK
%
BÍL
K K
G
BÍL
K %
1 NEO-437 SUNNYDAY 2013 401 NUMERO UNO ALTAIOTA 150,8 0 0 82 141 305 67 0,74 25 0,26
2 NEO-513 SILVER 2013 170 *TV*TL*TY MOGUL SNOWMAN 150,2 0 0 77 147 2072 78 0,08 51 -0,02
3 NXB-120 RASTY 2012 201 *TV*TL SUDAN O MAN 145,2 0 0 79 150 2093 72 0,01 56 0,03
4 NEO-572 ABBADO 2012 121 *TV*TL*TY NUMERO UNO PLANET 145,0 0 0 85 126 388 48 0,45 18 0,12
DC
ER S
B
STÁ
D S
B
SB
DC
ER E
XT
STÁ
D E
XT
R -
EXT
DI-
KO
N
DI-
VEM
RÁ
MEC
ŠÍŘ
KA
HR
UD
.
HL.
TĚL
A
HR
AN
ATO
ST
SKL.
ZA
DĚ
ŠÍŘ
. ZÁ
DĚ
PZK
-ZE
ZAD
U
PZK
-ZB
OK
U
ÚH
EL P
AZ
PŘ
. UP
N. V
EM.
RO
ZM. P
Ř. S
TR.
DÉL
KA
STR
.
HLO
UB
KA
VEM
.
VÝŠ
KA
ZA
D. U
P.
ZÁV
. VA
Z
RO
ZM. Z
. STR
.
ŠÍŘ
KA
ZA
D. U
P.
KV
AL.
KO
STI
CH
OD
IVO
ST
KO
ND
ICE
ML.
SÍL
A
0 0 100 0 0 61 112 120 108 107 108 102 81 105 100 97 101 109 119 101 109 109 123 119 108 109 113 110 117
0 0 104 0 0 55 118 117 121 113 109 104 98 98 129 98 98 108 108 97 110 127 104 106 133 116 111 97 129
0 0 106 0 0 55 109 103 106 104 104 103 111 106 112 87 94 100 103 88 105 102 90 99 105 101 113 111 104
0 0 109 0 0 56 112 123 106 97 102 109 80 105 108 96 95 119 121 97 119 109 110 120 107 119 118 101 110
72

STA
VB
A T
ĚLA
KO
NČ
ETIN
Y
VEM
ENO
TYP
CEL
KEM
INS-
JAL
PLO
DN
OST
-JA
L
INS-
KR
PLO
DN
OST
-K
R
PLO
DN
OST
-P
L
DC
ER D
LV
VYŘ
R -
DLV
DLO
UH
OV
ĚKO
ST
ID-B
YKA
117 115 118 123 0 112 0 106 110 0 0 0 135 CZEM000700787053
116 123 122 131 0 102 0 99 100 0 0 0 106 USAM000072156794
110 112 104 108 4 101 0 106 105 0 0 0 92 CZEM000796586061
121 117 121 125 0 115 0 113 117 0 0 0 147 ITAM017991251571
73

„V éře genomiky je fenotyp
králem.“
U všech druhů mají klíčový význam
fenotypová data vysoké kvality.
74

U skotu odhalilo sekvenování 1000 býků více
než 30 mil. SNP.
75

Zavedena úspěšně u velkých populací, tj.
holštýn
simentál
jersey
brown swiss.76

Mléčný skot
GS umožňuje omezit resp. ukončit tzv.
„star system“.
Nadměrné použití několika vynikajících
plemeníků, kteří jsou využíváni jako otcové
býků. To vede k vyššímu inbreedingu.
Pozor! V praxi bylo dosud zavedení GS
spojeno s vyšším inbreedingem!77

Mléčný skot
Implementace znamenala vyšší genetický
zisk, např. v Kanadě se zdvojnásobil.
Zároveň se ale zvýšil inbríding. Úkol do
budoucna – zvýšit zisk a omezit inbríding.
Zkrácení odhadu PH u býků z více než 6 let
na 3 roky.
Stačí menší počet inseminačních stanic.78

Masný skot
GS nebyla dosud uplatněna v takové míře,
jako u mléčného:
nižší přesnost
ekonomické výhody nejsou tak vysoké
jako u mléčného skotu
79

Masný skot
Přesnost GPH nižší, než u mléčného skotu:
Referenční populace má nižší kvalitu.
Méně zvířat v ref. populaci pro jedno
plemeno, ta nejsou testována podle už.
potomstva.
Ref. populace a populace v níž se
selektuje jsou méně geneticky propojené
než u mléč. plemen.80

Masný skot
Holštýn je převažující mléčné plemeno.
Jeho Ne je méně než 100.
Masných plemen je mnoho.
Každé má Ne větší,
než holštýn!!!
Každé má své specifické vlastnosti. 81

Prasata
82

Nejdůležitějším krokem v selekci je výběr
kanců v nukleových chovech.
Výsledky testování kanců přichází
většinou před výběrem elitních kanců,
takže zlepšení výsledků v důsledku
redukce generačního intervalu je
limitováno, i když s pomocí GS lze
dosáhnout asi 25% redukce generačního
intervalu.83

Proto je GS orientována zvl. na
znaky, kde je získání dat invazivní
(jatečná hodnota);
maternální znaky u kanců;
užitkovost kříženců, která nemůže být
hodnocena u čistokrevných zvířat.
84

V komerčním šlechtění prasat je v souč. době
nejčastěji používanou aplikací GS
jednokrokový odhad, založený na
imputovaných genotypech.
85

Cena kance a býkaU skotu je býk testován ve vyšším věku (dcery
dokončí 1. laktaci), náklady jsou vysoké. Býk
musí být chován, musí se shromáždit velký
počet dávek, test potomstva je zahájen po
dlouhé době.
Náklad na genotypizaci i sekvenaci býka je
relativně nízký ve srovnání s náklady na
konvenční testování.86

Cena kance a býkaU prasat jsou náklady na odchov nižší,
selekčními kandidáty jsou všichni testovaní
čistokrevní kanečci s rodokmenem.
Proto jsou náklady na genotypizaci kance
relativně významnější.
87

Další rozvoj u prasat
U prasat hraje významnou roli u mnoha
znaků heteróze. Je-li GPH je založena na
datech o užitkovosti kříženců, je užitečné
zařadit do modelu ne-aditivní genetické
efekty, které způsobují heterózi.
To může zvýšit přesnost GPH a snížit její
vychýlení. 88

Drůbež
89

Tradiční šlechtitelské programy
již mají
krátký generační interval.
Vícečetné překrývající se generace za rok se
selekcí každých 6 týdnů u brojlerů,
nepřekrývající se roční generace u nosných.90

Velmi vysoký počet selekčních kandidátů a
vysoká intenzita selekce.
Desetitisíce sel. kand. na linii a generaci, jen 1-
3% samců selektováno pro šlechtění.
Proto
genotypizace musí být levná!
91

U drůbeže je genetický pokrok z čistých linií
přenášen křížením pyramidově do velkého
počtu brojlerů nebo nosnic.
Jedna linie vnáší jen 25%!!!
92

U drůbeže není spolehlivá metoda
kryokonzervace, selektovaný samec má jen
místní význam.
93

Optimální je
sledování haplotypů zpětně k původním
liniím
a následná
selekce čistokrevných linií podle přítomnosti
výhodných haplotypů.
94

Hlavní přínos
GS u drůbeže je zvýšení přesnosti odhadu
PH ve věku puberty u znaků, manifestovaných
pohlavně specificky.
95

Koně
96

Hlavní výzvou
je limitovaná dostupnost kvalitních
fenotypových dat ve srov. s mléčným
skotem.
GS má enormní potenciál.97

Koně mají značně různorodé využití.
Menší podíl profesionálů mezi chovateli
než u ostatních druhů hospodářských
zvířat (eufemisticky řečeno)!!!
Vysoká cena koní peněžní a citová.
Proto je management populace a design
šlechtění stále problémem. 98

Generační interval
U koní delší, než byl u mléčného skotu před
GS. Přesnost odhadu PH nižší.
Kůň je ca 9-10 let starý!!! když je známa jeho
konvenční PH. Pokud se vůbec zjišťuje!!!99

U sportovních koní
je značná výměna genetického materiálu
mezi plemeny a zeměmi.
GS poskytuje možnost přístupu k plem.
hodnotám pro zahraniční koně.
Podm. je gen. propojení mezi populacemi.
Odhady pro více znaků, jednokroková
metoda. 100

Budoucnost (pro všechny druhy)
Často je překážkou cena genotypizace. S
jejím poklesem bude použití GS stoupat.
To pomůže sestavení větších a
aktuálnějších refer. populací.
101

Budoucnost
Editace genomu
Cílená změna sekvence DNA v buňce.
Uměle syntetizované sekvence jsou
integrovány do přesně definované pozice.
CRISPR/Cas9 !!
Praktické aplikace jsou na dosah. 102

Budoucnost
Editace genomu
je slibná u majorgenů, ale podstatně méně u
komplexně podmíněných vlastností. Sem
ale patří většina ekonomicky důležitých
vlastností.
Praktický význam editace proto bude
omezený. 103

Srovnání s rostlinami
Zcela rozdílné.
Genetické zlepšení mnoha rostlinných
druhů je založeno na klonech a jejich
testování v různých podmínkách.
Cílem je klon s jasnými přednostmi v
resistenci.
104

Srovnání s rostlinami
Velkou výhodou práce s klony je, že
genomické hodnocení může být velmi
důkladné, protože
příslušný fenotyp může být využíván
velmi dlouho.
105
Top Related