







Languages
Pages
Legal


BAB 1
PENDAHULUAN
Leptin merupakan hormon terutama dihasilkan sel adiposit, terdiri dari
164 asam amino dengan berat molekul 16 kD (Singla et al,.2010; Kershaw &
Flier, 2004). Leptin pertama kali ditemukan oleh Freidman dan kawan kawan
yang mengklon gen ob pada tikus. Leptin berperan dalam pengaturan
keseimbangan energi (Zang et al., 1994; Masuzaki et al., 1995; Leibel, 2008).
Leptin mempunyai efek menghambat nafsu makan, dengan memberikan
respons rasa kenyang. Kadar leptin sebanding dengan jumlah jaringan adiposa
dalam tubuh. Leptin telah dikenal sebagai hormon yang berperan dalam mengatur
berat badan, pada penelitian akhir akhir ini diketahui bahwa leptin secara primer
berperan dalam pengaturan homeostasis glukosa yang tidak tergantung pada
aksinya terhadap asupan makanan, penggunaan energi atau berat badan. Leptin
diekspresikan pada banyak jaringan di perifer melalui reseptornya. Reseptor
leptin terdapat pada pankreas, hepar, otot rangka dan jaringan adiposa, aksi leptin
secara langsung maupun tidak langsung berperan dalam mengontrol homeostasis
glukosa (Denroche et al., 2012).
Berbagai penelitian membuktikan bahwa leptin dan insulin mempunyai
peranan penting di susunan saraf pusat (SSP) dan di perifer dalam mengontrol
homeostasis energi. Resistensi leptin diketahui berhubungan dengan resistensi
insulin melalui aksinya di sentral maupun di jaringan perifer. Resistensi leptin
akan menyebabkan obesitas dengan hiperfagia yang juga akan meningkatkan
kadar insulin (Singla et al., 2010).
1

Tinjauan pustaka ini akan membahas mengenai leptin, peran leptin dalam
homeostasis energi, insulin, resistensi insulin, peran leptin pada resitensi insulin
dan pemeriksaan laboratorium leptin dan insulin.
2

BAB 2TINJAUAN PUSTAKA
2.1 LEPTIN DAN RESISTENSI LEPTIN
2.1.1 Leptin
2.1.1.1 Sejarah dan Definisi
Leptin pertama kali ditemukan pada tahun 1994, oleh Friedman dan
kawan kawan yang berhasil mengklon gen ob pada tikus. Produk yang dihasilkan
gen ob ini disebut leptin. Leptin berasal dari kata leptos, bahasa Yunani yang
berarti kurus (Zang et al., 1994; Patani et al., 2012).
Leptin adalah produk protein dari gen ob yang dihasilkan terutama oleh
sel adiposit, memberikan pengaruh terhadap asupan makanan, pengeluaran energi,
berat badan dan fungsi neuroendokrin melalui targetnya pada sel saraf di
hipotalamus (Smith et al, 2000; Patani et al., 2012).
2.1.1.2 Struktur Leptin
Gambar 2.1 Molekul Leptin (Kline et al. 1997)
Leptin terdiri dari rantai heliks yang dihubungkan dengan loop. Loop AB,
BC, dan CD yang mempunyai tambahan heliks E. Secara umum panjang heliks
3
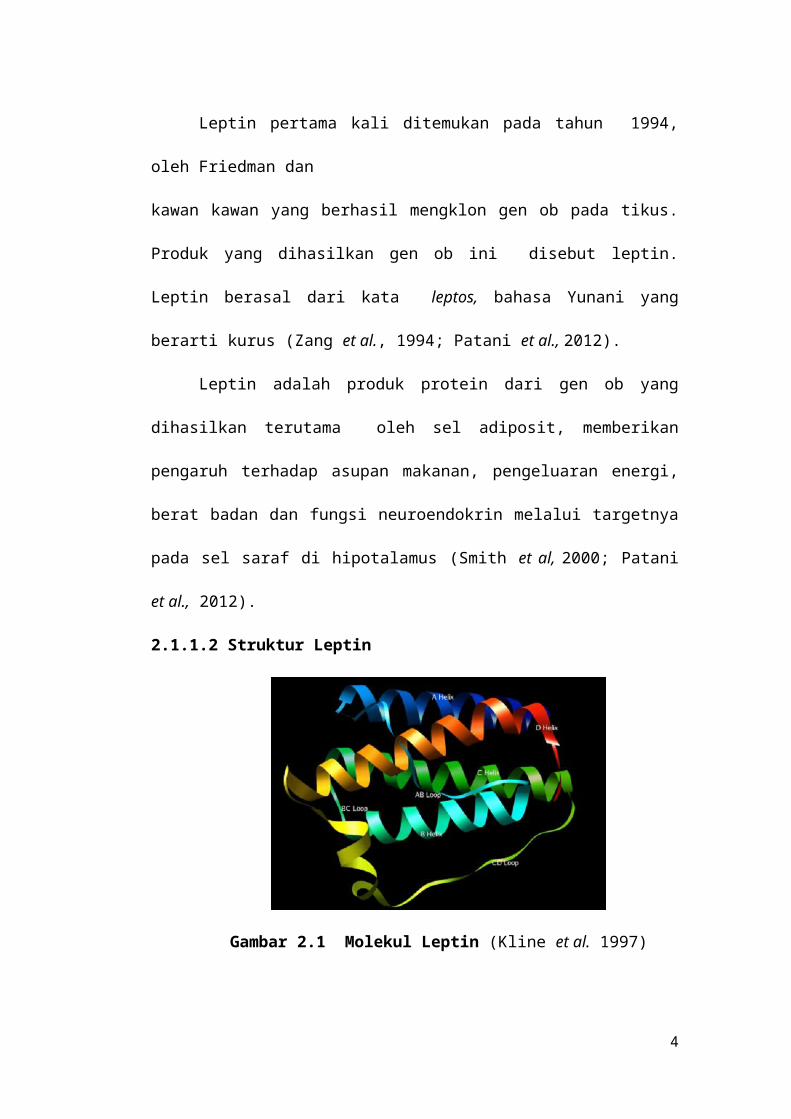
dan ikatan disulfida menunjukkan kemiripan dengan kelompok sitokin heliks
pendek (Kline et al. 1997; Zhang et al. 1997).).
Leptin manusia terdiri dari 164 asam amino dengan ikatan disulfida
tunggal, terletak di C-terminal akhir antara residu sistein 96 dan 146 (Patani et
al.,2012). Gen ob yang mengode leptin berlokasi pada kromosom 7 lengan
panjang pada posisi 31.3 (Genetic Home Reference, 2014).
2.1.1.3 Fisiologi Leptin
Kadar leptin dalam darah berfluktuasi mengikuti irama sirkadian, dengan
kadar terendah pada pagi hingga tengah hari dan kadar tertinggi setelah tengah
malam hingga dini hari. Karakteristik fluktuasi sekresi leptin serupa pada
individu kurus dan gemuk sedangkan volume leptin yang disekresikan lebih
besar pada orang gemuk (Schoeller et al,1997; Minocci, 2000; Radic et al.,2003).
Kadar leptin di sirkulasi secara langsung sebanding dengan jumlah lemak
tubuh dan berfluktuasi sesuai dengan asupan kalori. Kadar leptin basal lebih
tinggi pada perempuan dibandingkan pada laki-laki dan berkorelasi dengan
distribusi lemak tubuh serta indeks IMT (Radic et al.,2003; Leibel, 2008).
Produksi leptin lebih tinggi di lemak subkutan dibandingkan lemak viseral.
Puasa yang lama secara substansial mengurangi kadar leptin, sedangkan pada
keadaan kenyang jumlah leptin akan meningkat (Radic et al., 2003; Patani et
al.,2012). Peningkatan kadar leptin juga mengaktifkan hormon tiroid, hormon
pertumbuhan, axis gonad dan menekan aksis hipofisis-adrenal (Mantzoro, 1999).
Leptin, di sirkulasi dalam bentuk bebas dan terikat protein. Sebagian
besar leptin bersirkulasi dalam bentuk makromolekul terikat. Leptin dalam bentuk
terikat mempunyai waktu paruh yang lebih lama dari pada leptin bebas. Leptin
4

dibersihkan dari sirkulasi dan dikeluarkan melalui ginjal (Sharma et al., 1997;
Mantzoro, 1999; Fruhbeck, 2006). Penelitian pada individu kurus dengan massa
jaringan adiposa yang relatif kecil, mayoritas leptin dalam bentuk terikat,
sedangkan pada orang gemuk terjadi peningkatan leptin bebas (Mantzoro, 1999;
Fruhbeck, 2006).
2.1.1.4 Peran Leptin dalam Metabolisme Energi
Leptin dapat bekerja di susunan saraf pusat dan di perifer dalam
mengontrol homeostasis glukosa. Kerja leptin di susunan saraf pusat adalah di
hipotalamus sedangkan diperifer leptin bekerja pada jaringan otot, hepar, pankreas
dan jaringan adiposit (Cedia et al., 2002)
Kerja leptin di susunan saraf pusat terutama pada area nukleus arkuatus,
hipotalamus lateral dan nukleus paraventrikular yang merupakan area utama
pengatur homeostasis energi. Leptin di susunan saraf pusat menstimulasi produksi
neuropeptida anoreksigenik antara lain proopiomelanocortin (POMC)/cocaine
and amphetamine related peptide (CART) dan mengurangi neuropeptida
oreksigenik antara lain neuropeptide Y (NPY)/agouti related protein (AgRP)
yang berlokasi di area tersebut (Sahu, 2004).
Kerja leptin di jaringan perifer melalui reseptor leptin yang diekspresikan
dijaringan tersebut, termasuk pankreas sebagai organ endokrin dan jaringan yang
sensitif terhadap insulin. Kerja leptin secara langsung di pankreas adalah
menghambat sekresi insulin dari sel beta dan glukagon dari sel alfa, sedangkan di
sel adiposa leptin menghambat sinyal dan kerja insulin. Penelitian secara invivo
mendapatkan efek antagonis leptin terhadap sensitifitas insulin di hepar. Efek
5

langsung leptin di otot rangka adalah menurunkan asupan glukosa dan
menstimulasi insulin dalam metabolisme glukosa (gambar 2.2) (Denroche et al.,
2012).
Gambar 2.2 Kerja Leptin Secara Langsung pada Jaringan yang Berperan dalam Homeostasis Glukosa (Denroche et al., 2012) AMPK= adenosine monophosphate-activated protein kinase; BAT= brown adipose tissue; cAMP = cyclic adenosine monophosphat; WAT = white adipose tissue
Peran leptin dalam mengatur keseimbangan energi dipengaruhi insulin
(Nelson & Cox, 2004). Leptin dilepaskan dari jaringan adiposit dan insulin
dilepaskan dari sel pankreas. Dua hormon ini beraksi pada sel anoreksigenik
untuk memicu pelepasan melanocytes stimulatting hormone (MSH), hal ini
mengaktifkan sinyal neuronal untuk mengurangi nafsu makan dan meningkatkan
metabolisme bahan bakar. Leptin dan insulin juga beraksi pada sel oreksigenik
6

untuk menghambat pelepasan NPY dan mengurangi sinyal nafsu makan yang
akan dikirim ke jaringan ( Nelson & Cox, 2004)
Beberapa penelitan membuktikan bahwa leptin secara invfitro
mengurangi fosforilasi glucose transporter 2 (GLUT2), transpor glukosa dan
kadar adenosine triphosphate (ATP) intraselular. Leptin mengaktivasi saluran
ATP-sensitive kalium (K+ATP) dan hiperpolarisasi sel beta sehingga menurunkan
kadar kalsium intraselular. Leptin juga memperlihatkan efek menekan cyclic
adenosin monophosphate (cAMP) yang akan mengaktivasi phosphodiesterase-3B
(PDE3B). Leptin juga menghambat protein kinase C (PKC), menghambat
asetilkolin dan glucagon-like peptide 1 (GLP-1). Leptin secara langsung menekan
insulin dengan menghambat glukosa dan penutupan saluran KATP, dan dengan
menghambat cAMP. Protein kinase C memerantarai sekresi insulin di sel beta
pankreas (Denroche et al., 2012)
2.1.2 Resistensi Leptin
Resistensi leptin merupakan keadaan berkurangnya atau tidak adanya
respons hormon dalam memberikan efek menghambat nafsu makan dan
mengontrol berat badan (Vaselli, 2012). Hal ini akan menyebabkan asupan energi
yang berlebihan sehingga keseimbangan energi terganggu. Resistensi leptin
ditemukan pada individu obesitas (Ramadhinara et al., 2008; Farooqi &
O’Rahilly, 2009; Myers, et al., 2012). Individu dengan obesitas mempunyai
kadar leptin yang lebih tinggi dibandingkan dengan individu kurus (Manzoros et
al., (2011).
7

Terjadinya resistensi leptin disebabkan kegagalan leptin dalam menembus
sawar darah otak akibat adanya gangguan transpor leptin di susunan saraf pusat,
berkurangnya reseptor leptin pada sel target, defek pada reseptor (akibat proses
mutasi) sehingga leptin tidak dapat berikatan dengan reseptornya, dan adanya
polimorfisme reseptor yang akan mengubah ikatan dan transmisi sinyal pada
resptor leptin ( Pijl, 2007; Ramadhinara et al., 2008; Moon et al., 2013).
Faktor yang berperan dalam mekanisme terjadinya resistensi leptin antara
lain (Martin et al., 2008). :
- Mutasi genetik, adanya mutasi pada gen leptin maupun reseptor leptin
dapat menyebabkan sinyal yang ditimbulkan leptin tidak efektif
sehingga terjadi hiperleptinemia dan resistensi leptin. Penelitian pada
tikus diabetes dan tikus zucker yang mengalami disfungsi reseptor leptin
didapatkan adanya hiperleptinemia dan resistensi leptin. Kasus mutasi gen
ini jarang terjadi pada manusia tetapi faktor genetik ini diwariskan dan
berperan dalam terjadinya resistensi leptin
- self-regulation, seperti pada jalur sinyal biologis lain, leptin juga
mempunyai efek pengaturan tersendiri, adanya gangguan dalam sistem
pengaturan ini mengakibatkan penurunan ekspresi reseptor terhadap sinyal
leptin.
- Adanya hambatan pada akses leptin di sawar darah otak. Berdasarkan
penelitian didapatkan zat yang dapat menghalangi leptin dalam melintasi
sawar darah otak, antara lain kadar trigliserida yang tinggi dan (Banks et
al., 2004) dan peningkatan kadar C reactive protein (CRP) (Chen et
al,2006).
8

- Efek molekular yang akan menghambat leptin pada tingkat selular dan di
sirkulasi. Adanya induksi suppressor-of cytokine-signaling-3 (SOCS3)
intraselular yang berlebihan akan menghambat sinyal janus kinase/signal
transducer and activator of transcription (JAK/STAT) leptin.
Kelebihan asupan makanan akan mengakibatkan kelebihan energi,
peningkatan jaringan adiposa dan kadar leptin meningkat. Resistensi leptin di
sentral akan meningkatkan neuropeptida oreksigenik seperti NPY di
hipotalamus. Peningkatan neuropeptida ini meningkatkan rangsang parasimpatis
dan simpatis sehingga terjadi peningkatan produksi glukosa hepatik, peningkatan
produksi insulin, pengurangan produksi insulin-independent glucose dan
resistensi insulin di otot, serta pengurangan lipolisis di jaringan adiposa
(Ramadhinara et al., 2008; Farooqi & O’Rahilly, 2009; Myers, et al., 2012).
Fungsi leptin dalam mengatur keseimbangan energi (asupan dan
pengeluaran kalori) dimediasi terutama melalui jalur sinyal JAK/STAT. Ikatan
leptin dan reseptornya menghasilkan autofosforilasi JAK2, yang akan
memerantarai fosforilasi dan aktivasi STAT3 (Fruhbeck, 2006). Kaskade sinyal
yang diperantarai leptin ini akan dihambat oleh SOCS 3, yang merupakan
umpan balik negatif. Sinyal independen JAK/STAT menginduksi fosforilasi
phosphatidylinositol-3-kinase (PI3K), yang berperan dalam memerantarai kerja
insulin hipotalamus dan homeostasis glukosa. (Ramadhinara et al., 2008; Marino
et al., 2011).
9

Gambar 2.3 Mechanisme SOC3 dalam Menekan Efek Fisiologis Leptin (Ramadanira et al., 2008)
Suppressor-of cytokine-signaling-3 merupakan inhibitor utama sinyal
leptin yang diinduksi oleh jalur JAK2/STAT3. Kerja SOCS3 bertujuan untuk
menghambat fosforilasi dan aktivasi JAK2. Leptin secara khusus menginduksi
ekspresi messenger ribonucleic acid (mRNA) SOCS-3 di hipotalamus yang
mengekspresikan kadar long form reseptor leptin. Ekspresi SOCS-3 akan
menghambat reseptor leptin dalam transduksi sinyal. Aktivitas SOCS-3 yang
berlebihan pada leptin, merupakan mekanisme yang potensial untuk resistensi
leptin yang dapat menimbulkan obesitas (Bjorbæk et al., 1998).
10

2.2 INSULIN DAN RESISTENSI INSULIN
2.2.1. Insulin
Insulin merupakan hormon peptida yang disekresikan oleh sel beta
pankreas. Insulin berfungsi menjaga homeostasis glukosa darah dengan cara
menurunkan kadar glukosa darah. Insulin bekerja dengan memerantarai uptake
glukosa seluler, regulasi metabolisme karbohidrat, lemak, dan protein (Mayes &
Bender , 2003).
Struktur insulin terdiri dari rantai A dan rantai B, kedua rantai ini
dihubungkan dengan jembatan sulfida yang menghubungkan struktur heliks
terminal N-C dari rantai A dengan struktur heliks sentral dari rantai B. Insulin
mengandung 51 asam amino yang terbagi dalam rantai A terdiri dari 21 asam
amino dan rantai B terdiri dari 30 asam amino (Greenstein, 1994; Dadson &
Steiner, 1998).
2.2.1.1 Sintesis dan Pelepasan Insulin
Insulin disintesis oleh sel beta pankreas sebagai preproinsulin di
retikulum endoplasma. Preproinsulin akan diubah menjadi proinsulin dengan
bantuan enzim peptidase lalu disimpan dalam vesikel sekretori dan dikirim ke
badan golgi. Proinsulin akan bergabung dengan zink dan kalsium sehingga
menjadi bentuk heksamer yang tidak larut air di badan golgi. Enzim peptidase
juga akan merubah proinsulin menjadi insulin dan C-peptide dan akan dikeluarkan
melalui membran sel (Dadson & Steiner, 1998).
Peningkatan kadar glukosa setelah makan menginduksi sekresi insulin
pada tahap awal. Glukosa akan dibawa oleh GLUT 2 dan masuk ke dalam sel
11

beta pankreas. Glukosa di dalam sel beta pankreas akan difosforilasi menjadi
glukosa-6-fosfat (G6P) dengan bantuan enzim glukokinase dan ATP. Penutupan
saluran K+-ATP ini mengakibatkan depolarisasi membran plasma dan aktivasi
saluran kalsium yang menyebabkan peningkatan konsentrasi kalsium intraselular
yang memicu sekresi insulin oleh sel beta pankreas (Soria et al., 2004).
Pelepasan insulin juga diinduksi oleh aktivasi fosfolipase dan protein
kinase C serta rangsangan dari aktivitas adenilat siklase dan protein kinase-A sel
beta. Mekanisme induksi sekresi insulin ini akan mengaktifkan hormon, seperti
vasoactive intestinal peptide (VIP), pituitary adenylate cyclase-activating
polypeptide (PACAP), GLP-1, dan glucose-dependent insulinotropic polypeptide
(GIP), yang berperan dalam “fase kedua” sekresi insulin, yaitu pelepasan insulin
yang baru disintesis maupun insulin yang tersimpan dalam granula sekretorik
(Bratanova et al., 2002).
2.2.1.2 Reseptor Insulin
Reseptor insulin berada di membran sel, terdiri dari subunit glikoprotein 2
alfa dan 2 beta yang dihubungkan ikatan disulfida. Insulin berikatan dengan
subunit alfa di ekstraselular kemudian mengalami perubahan bentuk dengan
mengikat ATP pada subunit beta di intraselular. Ikatan ATP akan memicu
fosforilasi subunit beta melalui aktivitas enzim tirosin kinase. Fosforilasi tirosin
pada substrat intraselular/insuline receptor substrate (IRS), yang akan
mengaktivasi sinyal molekul lain dan berperan dalam kerja insulin di intraselular
(Kido et al., 2001).
Insuline receptor substrate merupakan substrat protein yang difosforilasi
oleh reseptor insulin. Insuline receptor substrate terdiri dari empat jenis IRS 1,
12

IRS 2, IRS 3 dan IRS 4. Insuline receptor substrate 1 merupakan IRS yang
terbanyak ditemukan di otot rangka, IRS2 ditemukan di hepar yang berfungsi
dalam aktivitas insulin dan pertumbuhan dari sel beta pankreas, IRS 3 ditemukan
pada jaringan adiposa, sel beta pankreas, dan hepar, sedangkan IRS 4 ditemukan
di timus, otak dan ginjal. Insuline receptor substrate yang telah terfosforilasi akan
mengikat src-homology-2 domain protein (SH2) spesifik, yang akan
memengaruhi beberapa enzim seperti PI3K dan phosphotyrosine phosphatase
(SHPTP2 ) (Withers & White, 2000; Kido et al.,2001).
Insuline receptor substrate 1 merupakan IRS yang mengalami fosforilasi
dan berhubungan langsung dengan PI3K segera setelah adanya stimulasi insulin
(Myers et al.,, 1992). Phosphatidylinositol-3-kinase akan mengakibatkan
translokasi protein GLUT, sintesis glikogen lipid dan protein, anti-lipolisis, serta
mengatur glukoneogenesis di hepar. Phosphatidylinositol-3-kinase bekerja
melalui serin dan threonin kinase seperti PKB, PKC dan phosphatidylinositol
dependent protein kinases 1 & 2 (PIPD 1&2) (Kasuga, 2006).
2.2.1.3 Mekanisme Kerja Insulin
Insulin berinteraksi dengan sel target melalui reseptor tirosin kinase yang
berfungsi memicu respons intrasel untuk memengaruhi jalur metabolisme.
Respons awal yang terjadi adalah translokasi GLUT dari aparatus golgi ke
membran plasma yang akan membawa glukosa masuk kedalam sel (Michael et
al.,2007).
Glucose transport unit 4 adalah transporter glukosa utama yang terdapat
pada sel otot dan sel lemak. Efek intraselular GLUT-4 dimulai dengan pengikatan
13

insulin pada bagian ekstraseluler dari reseptor insulin transmembran. Ikatan ini
mengaktifkan fosforilasi tirosin kinase pada bagian intraseluler dari reseptor,
aktivasi selanjutnya melaui PI3K yang diperlukan untuk stimulasi transpor
glukosa oleh insulin yang akan merangsang translokasi GLUT-4 ke membran
plasma (Sheperd et al, 1999).
Translokasi GLUT-4 di intraselular ke membran plasma dirangsang oleh
ekspresi bentuk aktif PKB atau isoform atipikal PKC. Translokasi GLUT-4 akan
mengangkut glukosa dari ekstrasel ke intrasel (Sheperd et al, 1999; Shulman,
2000)
2.2.2 Resistensi Insulin
Resistensi insulin berarti berkurangnya kemampuan insulin dalam
memberi efek biologik yang normal pada kadar gula darah tertentu, pada
keadaan ini dibutuhkan kadar insulin yang lebih banyak untuk mencapai kadar
glukosa darah yang normal (ADA, 2004). Resistensi insulin merupakan suatu
kondisi yang berhubungan dengan kegagalan organ target yang secara normal
merespons aktivitas hormon insulin (Savage, 2005).
2.2.21 Mekanisme Terjadinya Resistensi Insulin
Mekanisme terjadinya resistensi insulin dipengaruhi oleh berbagai faktor
antara lain faktor inflamasi dan obesitas. Peningkatan produksi berbagai sitokin
proinflamasi akibat akumulasi jaringan lemak pada obesitas menyebabkan
keadaan inflamasi subakut pada obesitas. Ikatan molekul sitokin proinflamasi
dengan reseptor spesifiknya akan mengaktifkan jalur janus kinase dan inhibitor
κB kinase (IκβK). Selanjutnya keadaan ini akan mengaktifkan faktor trankripsi
14

nuclear factor κβ (NFκβ). Translokasi NFκβ ke dalam nucleus akan menginduksi
transkripsi berbagai macam mediator inflamasi yang mengarah pada keadaan
resistensi insulin (Hotamisligil, 2000).
Jalur janus kinase dan IKKβ/NFκβ juga dapat diaktivasi oleh ikatan dari
pattern recognition receptor (PRR) yang merupakan reseptor pada permukaan
membran dengan substansi dari luar sel. Pattern recognition receptor pada
membran sel ini antara lain adalah toll-like receptor (TLR) dan receptor for
advanced glycation end products (RAGE). Receptor for advanced glycation end
products akan berikatan dengan advanced glycation end products (AGE)
endogen yang merupakan subtansi nonenzimatik dan merupakan produk dari
metabolisme glukosa dan protein dengan laju turnover yang lambat (Shoelson,
et al., 2006).
Resistensi insulin juga dapat diinduksi oleh faktor yang berasal dari dalam
sel. Adanya stres intraselular seperti reactive oxygen species (ROS) atau reactive
nitrogen species (RNS), stres pada retikulum endoplasmikum, ceramide, dan
beragam isoform dari PKC. Beberapa faktor intrasel ini akan mengaktifkan jalur
janus kinase dan IKKβ/NFκβ yang akan menginduksi resistensi insulin pada sel
target (Shoelson, et al., 2006).
Faktor lain yang berperan dalam terjadinya resistensi insulin adalah
obesitas. Obesitas dapat menimbulkan resistensi insulin melalui peningkatan
produksi asam lemak bebas. Asam lemak bebas yang terakumulasi di jaringan
akan menginduksi resistensi insulin terutama pada hati dan otot. Mekanisme
induksi resistensi insulin oleh asam lemak ini terjadi akibat kompetisi asam lemak
dan glukosa dalam berikatan dengan reseptor insulin. Oksidasi asam lemak akan
15

menyebabkan peningkatan asetil koA pada mitokondria dan inaktivasi enzim
piruvat dehidrogenase. Mekanisme ini akan menginduksi peningkatan kadar sitrat
intraselular yang akan menghambat akumulasi fosfofruktokinase dan glukosa
phosphat yang menyebabkan akumulasi glukosa interselular dan mengurangi
uptake glukosa dari ekstrasel (Rothman et al., 1995; Misra, 2007)).
Berdasarkan penelitian didapatkan bahwa akumulasi asam lemak dan
metabolitnya di dalam sel akan menyebabkan aktivasi jalur serin/threonin kinase.
Aktivasi jalur ini menyebabkan fosforilasi pada gugus serin dari kompleks IRS,
sehingga fosforilasi dari gugus tironin seperti pada mekanisme kerja insulin yang
normal akan terhambat. Hambatan pada fosforilasi gugus tironin kompleks IRS
ini menyebabkan jalur PI3K tidak teraktivasi sehingga glukosa tetap berada di
ekstrasel (Savage et al., 2005; Shulman, 2000).
Resistensi insulin menyebabkan penggunaan glukosa yang dimediasi oleh
insulin di jaringan perifer menjadi berkurang dan akan menyebabkan kegagalan
fosforilasi kompleks IRS, penurunan translokasi GLUT4 dan penurunan oksidasi
glukosa sehingga glukosa tidak dapat masuk ke dalam sel dan terjadi
hiperglikemia (Williams et al., 2002; Lee, 2006).
Sel beta pankreas pada tahap awal akan melakukan kompensasi untuk
merespons keadaan hiperglikemia dengan memproduksi insulin dalam jumlah
yang lebih besar sehingga terjadi hiperinsulinemia (Williams et al., 2002; Lee,
2006). Kegagalan sel beta dalam merespons kadar glukosa darah yang tinggi, akan
menyebabkan abnormalitas jalur transduksi sinyal insulin pada sel beta dan terjadi
resistensi insulin di sel beta pankreas. Hal ini akan mengaktivasi jalur caspase dan
peningkatan kadar ceramide yang menginduksi apoptosis sel beta, terjadi
16

kerusakan sel beta pankreas yang akan diikuti oleh berkurangnya massa sel beta.
Pengurangan massa sel beta pankreas ini akan menyebabkan sintesis insulin
berkurang dan menyebabkandiabetes melitus tipe 2 (Ten & MacLaren, 2004).
Resistensi insulin juga menyebabkan peningkatan lipolisis dan produksi
asam lemak bebas akibat berkurangnya inhibisi enzim lipase pada sel lemak.
Produksi asam lemak bebas ini akan ditransportasikan ke hati dan akan
menginduksi peningkatan produksi very low density lipoproteni (VLDL),
apolipoprotein B (apoB) dan sekresi trigliserida (McNeal & Wilson, 2008).
2.3 PERAN LEPTIN PADA RESISTENSI INSULIN
Berbagai penelitian yang dilakukan menunjukkan bahwa leptin dan insulin
mempunyai peran penting di SSP dalam mengontrol homeostasis energi. Insulin
diskresikan dari endokrin pankreas dan berefek pada cadangan nutrisi di perifer.
Insulin juga memberikan sinyal afferen ke SSP yang menyebabkan efek inhibitor
jangka lama pada asupan energi. Reseptor leptin dan reseptor insulin
diekspresikan oleh sel otak yang memengaruhi asupan energi dan mengatur
neuropeptida secara langsung ke otak dan mengurangi asupan makanan.
Resistensi leptin menyebabkan obesitas, dengan hiperfagia yang persisten
sehingga kadar insulin juga akan meningkat (Singla et al.,2010).
Penelitian lain mendapatkan bahwa insulin memegang peranan secara
kronis dalam mengatur ekspresi gen leptin dan produksinya di white adipose
tissue (WAT). Keadaan hiperinsulinemia meningkatkan kadar leptin plasma dan
ekspresinya di WAT. Hal ini terbukti pada insulinoma yang menunjukkan
peningkatan leptin plasma, mRNA leptin serta kadar leptin plasma dan akan
kembali normal setelah insulinoma diatasi. Insulin nampaknya berpengaruh
17

langsung dengan kadar leptin di adiposit dengan meningkatkan sekresi leptin dan
ekspresi gen yang berhubungan dengan metabolisme dan transport glukosa
(Margetic, et al.,2002).
Insulin dan leptin merupakan sinyal adiposit yang beraksi di nukleus
arkuatus hipotalamus untuk menghambat ekspresi neuron NPY/AgRP yang
memacu anabolik (meningkatkan asupan makanan, menurunkan penggunaan
energi) dan menstimulasi ekspresi neuron POMC/CART yang memicu efek
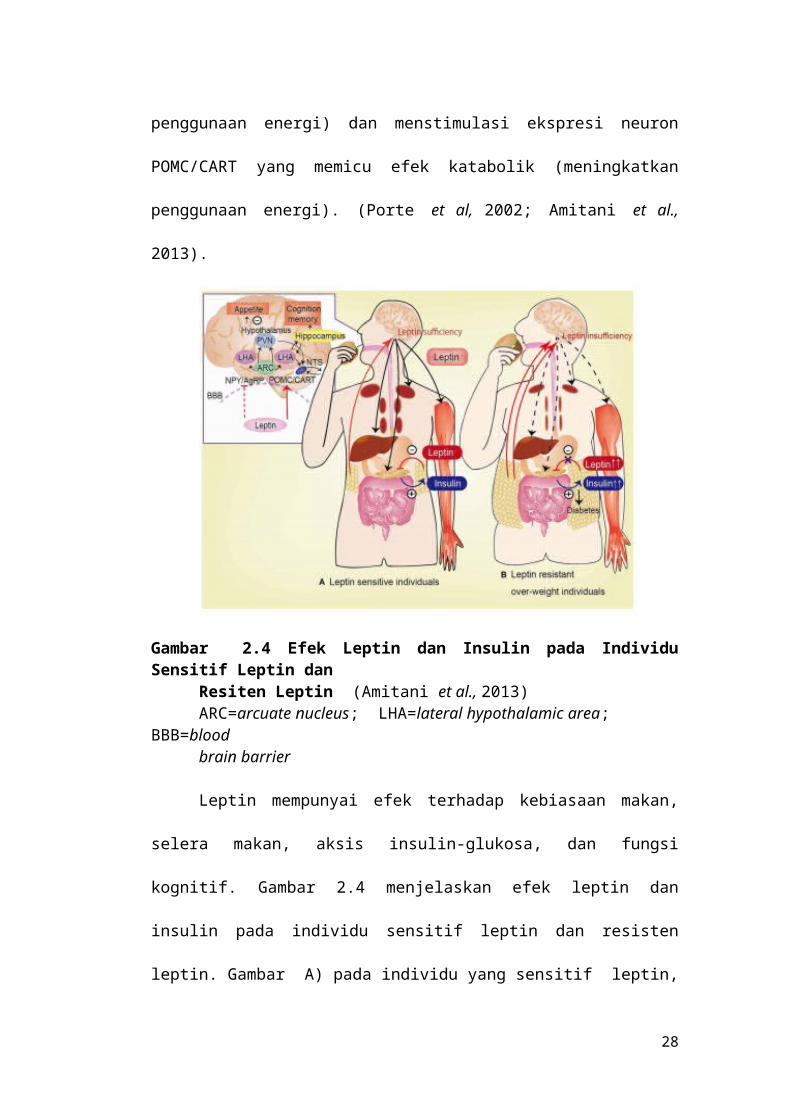
katabolik (meningkatkan penggunaan energi). (Porte et al, 2002; Amitani et al.,
2013).
Gambar 2.4 Efek Leptin dan Insulin pada Individu Sensitif Leptin dan Resiten Leptin (Amitani et al., 2013)ARC=arcuate nucleus; LHA=lateral hypothalamic area; BBB=blood brain barrier
Leptin mempunyai efek terhadap kebiasaan makan, selera makan, aksis
insulin-glukosa, dan fungsi kognitif. Gambar 2.4 menjelaskan efek leptin dan
18

insulin pada individu sensitif leptin dan resisten leptin. Gambar A) pada individu
yang sensitif leptin, leptin akan menghambat biosintesis insulin dan sekresi
insulin oleh sel beta pankreas. Sebaliknya insulin menstimulasi sekresi leptin dari
jaringan adiposa. Leptin menstimulasi glukoneogenesis di hepar dan sensitivitas
insulin di hepar melalui cabang nervus vagus di hepar. Leptin juga meningkatkan
uptake glukosa di otot, jantung dan lemak coklat melalui sistem nervus
simpatis.Gambar (B) pada keadan resistensi leptin (obesitas) permeabilitas sawar
darah otak akan menurun terhadap leptin karena diet yang tinggi lemak walaupun
kadar leptin di dalam serum meningkat. Kegagalan transport leptin melintasi
sawar darah otak merupakan salah satu penyebab resistensi leptin. Insufisiensi
sinyal leptin di hipotalamus (diinduksi oleh hiperleptinemia pada obesitas) akan
menyebabkan hiperglikemia dan hiperinsulinemia, yang akan mengarah ke
diabetes melitus. (Amitani et al., 2013).
Peran leptin dan insulin dalam terjadinya resistensi insulin dan DM tipe 2
akibat adanya disregulasi aksis adipoinsular (gambar 2.5). Gambar (A) pada
keadaan normal, sekresi leptin dari jaringan adiposa akan membatasi seksresi
insulin dari sel beta pankreas untuk menyesuaikan homeostasis glukosa dan
menyimpannya menjadi cadangan lemak. gambar (B) pada keadaan leptin resisten
(obesitas) pengurangan sinyal leptin pada sel beta pankreas akan menyebabkan
hipersekresi kronik pada insulin, sehinggga terjadi hiperinsulinemia. Peningkatan
kadar insulin ini akan memicu resistensi insulin dan peningkatan biosintesis leptin
serta sekresinya dari jaringan adiposa yang akan menurunkan sensitivitas sinyal
leptin di pankreas dan meningkatkan resistensi leptin. Hipersekresi insulin kronik
19

oleh sel beta pankreas yang disebabkan penurunan inhibisi oleh leptin yang pada
akhirnya akan menyebabkan kerusakan sel beta pancreas dan terjadi diabetes
melitus tipe 2 pada pasien obesitas ( Seufert, 2004).
Gambar 2.5 Disregulasi Aksis Adipoinsular dan Terjadinya Diabetes Tipe 2. ( Seufert, 2004).
Hubungan antara sinyal intraselular insulin dan leptin di duga melalui
enzim PI3K. Leptin berikatan dengan reseptornya, menyebabkan penggabungan
dengan enzim tirosin kinase, JAK-2 dengan domain di sitoplasma pada reseptor
leptin. Insulin berikatan dengan subunit alfa di ekstraselular yang akan
mengaktivasi tirosin kinase pada subunit beta di intraselular. Reseptor tirosin
kinase insulin akan mengatalisis protein IRS-2 yang berhubungan dengan PI3K,
Aktivasi enzim PI3K yang menurunkan transduksi sinyal pada jalur neuronal dan
mengakibatkan hambatan pada asupan makanan. Diduga bahwa aksi JAK-2
melalui mekanisme yang mirip dengan IRS-dependent dalam mengaktivasi
PI3K. Baik resptor leptin maupun reseptor insulin berinteraksi dengan sejumlah
20

jalur transduksi sinyal lain dapat memengaruhi asupan makanan dan berat badan
tetapi mekanismenya belum dapat dijelaskan (gambar 2.6) (Porte et al, 2002)
Gambar 2.6 Hipotesis Kaitan Antara Sinyal Intraselular Insulin dan Leptin Melalui Enzim Phosphotidylinositol 3-kinase (Porte et al, 2002)
Penelitian pada hewan menunjukkan bahwa leptin menghambat ekspresi
gen insulin dan sekresi insulin (gambar 2.7) Di sel beta pankreas leptin
menginduksi aktivasi STAT3 dan STAT5 yang akan bermigrasi ke nukleus dan
menginduksi ekspresi SOC3. SOC3 merupakan inhibitor jalur JAK/STAT, di sel
beta ini akan menghambat ekspresi proinsulin (Marroqui et al.,2012).
Gambar 2.7 Efek Leptin di Sel Beta Pankreas (Marroqui et al.,2012)
21

Hubungan antara sinyal leptin dan jalur yang diinduksi insulin dijelaskan
pada gambar 2.8. Jalur PI3K/PDE3B)/cAMP. Aktivitas PI3K merupakan target
utama yang diatur oleh suatu ligan dengan spektrum yang luas, yang memerlukan
insulin sebagai suatu faktor khusus yang penting. Pada kenyataannya kebanyakan
aksi yang tergantung insulin memerlukan aktivasi PI3K, sehingga hal ini
merupakan kaitan antara jalur sinyal insulin dan leptin. Produk PI3K secara
khusus menstimulasi protein kinase seperti Akt/PKB dan PKC (Fruhbeck, 2006).
Gambar 2.8 Hubungan Antara Sinyal Leptin Dan Jalur Yang Diinduksi Insulin. Aktivasi Leptin (L) receptor (OB-Rb) melalui beberapa komponen dari kaskade sinyal insulin yang merekrut insulin receptor substrate (IRS), phosphatidylinositol4,5-bisphosphate (PIP2), Phosphatidylinositol3,4 trisphosphate (PIP3) (Fruhbeck, 2006).
Leptin dilaporkan berkaitan dengan insulin melalui beberapa
komponen.Ikatan insulin pada reseptornya merekrut IRS yang difosforilasi oleh
aktivitas intrinsik reseptor. Fosforilasi IRS meningkatkan afinitasnya dalam
mengikat sinyal molekul lain, sehingga akan memengaruhi tahap selanjutnya.
Protein IRS menyebabkan aktivasi PI3K melalui gabungannya dengan regulator
subunitnya p85, dan meningkatkan aktivitas katalisisnya. Stimulasi PI3K
mengaktivasi PIP3 yang dipengaruhi oleh serin/threonin kinase seperti PDK1
22

yang dapat mengaktivasi Akt, selanjutnya serin/threonin kinase berperan dalam
downregulasi sinyal. Di susunan saraf pusat PI3K-dependent mengaktivasi
PDE3B dan reduksi cAMP. Leptin juga menstimulasi IRS2-dimediasi oleh
aktivitas hipotalamik PI3K, yang menghambat PI3K pada leptin dan menginduksi
hiperpolarisasi neuron NPY/AgRP, sehingga akan menghambat efek
anoreksigenik dari leptin (Fruhbeck, 2006).
23

BAB 3PEMERIKSAAN LABORATORIUM
3.1 Pemeriksaan Leptin
Metode yang dapat dipakai untuk mengukur kadar leptin total serum
antara lain dengan metode Enzyme Linked Immuno Assay (ELISA)
sedangkan untuk mengukur kadar leptin bebas dan leptin terikat digunakan
metode High Performance Liquid Chromatography (HPLC).
3.1.1 Praanalitik Pemeriksaan
Pengambilan sampel pada pagi hari atau awal sore dianggap cukup,
untuk pemeriksaan tunggal dengan siklus makan yang normal. Sampel
berupa serum, plasma dengan antikoagulan EDTA atau heparin.
Pengukuran kadar leptin masih dapat dilakukan pada sampel dengan kadar
hemoglobin serum/plasma sampai 1,0 mg/ml, kadar bilirubin sampai 170
µmol/l dan kadar trigliserida sampai 5,0 mmol/l.
Sampel serum atau plasma dapat disimpan pada suhu -70°C
selama beberapa tahun, suhu -20°C selama beberapa bulan, pencairan dan
pembekuan sampel secara berulang tidak dianjurkan.
3.1.2. Pemeriksaan Leptin dengan Metode Enzyme Linked Immuno Assay
3.1.2.1. Prinsip Pemeriksaan (Bio Vendor, 2014)
Terjadi ikatan anti gen dalam sampel dengan antibodi (polyconal
anti human leptin antibody) yang terdapat pada dinding well, dengan
penambahan konjugat dan substrat akan terjadi perubahan warna, sehingga
dapat dibaca oleh alat ELISA reader.
24

3.1.2.2 Alat dan Bahan
- Mikrotiter yang dilapisi antibodi
- Larutan konjugat
- Larutan standar
- Quality control high
- Quality control low
- Larutan bufer pengencer
- Larutan pencuci
- Larutan substrat
- Stop solution
- Deionized (distilled) water
- Tabung reaksi
- Gelas ukur
- Pipet ukur volume 10 µl -1000 µl dengan disposable tips
- Multichannel pipette ukuran 100 µl dengan disposable tips
- Kertas tisu
- Vortex mixer
- Orbital microplate shaker
- ELISA reader
Sebelum digunakan sampel dan reagen diletakkan pada suhu kamar.
3.1.2.3 Cara Kerja
- Sebanyak 100 µl larutan standar yang diencerkan, quality control,
blanko (larutan bufer yang diencerkan) dan sampel dimasukkan ke
dalam well
25

- Plate diinkubasi pada suhu ruangan(25°C ) selama 60 menit, shake pada
300 rpm diatas orbital microplate shaker.
- Well dicuci tiga kali dengan larutan pencuci (tiap well sebanyak 0,35 ml)
- Sebanyak 100 µl larutan konjugat ditambahkan pada tiap well
- Plate diinkubasi pada suhu ruangan (25°C) selama 60 menit, shake pada
300 rpm diatas orbital microplate shaker.
- Well dicuci tiga kali 3 dengan larutan pencuci ( tiap well 0,35 ml)
- Sebanyak 100 µl larutan substrat ditambahkan kedalam well, dan plate
dilhindarkan dari paparan cahaya secara langsung dengan ditutup
memakai aluminium foil
- Diinkubasi selama 10 menit pada suhu ruangan
- Ditambahkan 100 µl stop solution.
- Absorbansi dibaca dengan ELISA reader pada panjang gelombang 450
nm, dalam waktu tidak lebih dari lima menit. Kadar leptin dibaca sesuai
dengan kurva standar.
Gambar 3.1 Kurva Standar untuk Human Leptin ELISA(BioVendor 2014)
26

Nilai referensi Laki laki 0,7 - 5,3 ng/mL Perempuan 3,3 – 18,3 ng/mL
3.1.3 Metode High Performance Liquid Chromatography (HPLC)
Metode ini untuk menentukan fraksi leptin bebas dan leptin yang terikat.
3.1.3.1 Prinsip pemeriksaan:
Prinsip kerja HPLC adalah pemisahan komponen analit berdasarkan sifat
kepolaran masing-masing zat dalam sampel, dengan bantuan pompa, fase
gerak cair dialirkan melalui kolom detektor. Sampel dimasukkan ke dalam
aliran fase gerak dengan cara penyuntikan, di dalam kolom akan terjadi
pemisahan tiap komponen campuran.
3.1.3.2 Alat dan Bahan
- Lyophilized leptin
- Pelarut: HCl 1,5 mM, NaOH 7,5 mM
- Larutan elusi : 70 % acetonitile dalam 0,1 % three fluroacetic
acid/TFA
3.1.3.3 Cara Kerja
Standar murni diperoleh dengan melarutkan satu vial yang terdiri dari
1 mg lyophilized leptin ke dalam 0,5 ml HCl ( 1,5 mM) lalu dinetralisir
dengan 0,3 ml NaOH (7,5 mM) dan dipakai untuk HPLC column kemudian
dielusi dengan B (70 % acetonitile dalam 0,1 % three fluroacetic acid/TFA)
dari 30% sampai 70% selama 40 menit menggunakan chromatography pump.
Sampel diletakkan di C18 reverse-phase catridge column, dicuci dengan 2
ml larutan TFA 0,1% (larutan A). Analisis HPLC pada nucleosil C18 gel
27
column (ukuran celah 10 nm ukuran partiket 5 µm dengan larutan A dan
larutan B (70% acetonitrile dalam 0,1% TFA). Leptin dielusi selama 40 menit
menggunakan chromatography pump. Elusi peptida ini dianalisis
menggunakan sinar ultraviolet pada panjang gelombang 220 nm.(Ardekani et
al., 2008)
3.2 Pemeriksaan Insulin
Metode yang digunakan adalah metode Electro chemiluminescence
immunoassay (ECLIA)
3.2.1 Preanalitik Pemeriksaan
Sampel untuk pemeriksaan tunggal sebaiknya dikumpulkan setelah pasien
berpuasa dan tidak minum obat hipoglikemik minimal delapan jam sebelum
pengambilan spesimen.
Sampel yang digunakan adalah serum atau plasma dengan antikoagulan
lithium heparin, K3 EDTA dan natrium sitrat. Sampel hemolisis tidak boleh
digunakan, hasil pemeriksaan menjadi lebih rendah karena eritrosit akan
melepas insulin peptidase.
Stabilitas sampel
3.2.2 Prinsip Tes Metode Electro Chemiluminescence Immunoassay
(ECLIA) (Roche, 2014)
Terjadi ikatan antara insulin dengan antibodi monoklonal insulin-
spesifik terbiotinilasi, dan antibodi monoklonal spesifik insulin-berlabel
dengan kompleks ruthenium dan membentuk kompleks sandwich,
dengan penambahan streptavidin yang dilapisi mikropartikel, kompleks ini
akan terikat pada fase padat lalu dihisap ke dalam pengukur sel, secara
28

magnetis ditangkap oleh permukaan elektroda. Zat terikat ini kemudian
dipindahkan dengan procells. Adanya prubahan tegangan elektroda
kemudian menginduksi chemiluminescent emisi yang diukur dengan
photomultiplier
3.2.3 Alat dan Bahan :
- Streptavadin yang dilapisi mikropartikel
- Reagen1 : antibodi monoklonal spesifik insulin biotinylated
- Reagen 2: antibodi monoklonal spesifik insulin yang dilabel rhutenium
- Larutan bufer
3.2.4 Cara Kerja
- inkubasi petama : 20 µl sampel ditambahkan antibodi monoklonal insulin
spesifik terbiotinilasi, dan antibodi monoklonal spesifik insulin-berlabel
dengan kompleks ruthenium.
- inkubasi kedua: setelah penambahan streptavidin-dilapisi mikropartikel,
kompleks menjadi terikat pada fase padat melalui interaksi antara biotin
dan streptavadin.
- campuran reaksi dihisap ke dalam pengukur yang secara magnetis
ditangkap oleh permukaan elektroda lalu dipindahkan dengan ProCells.
Tegangan yang ditimbulkan elektroda kemudian menginduksi emisi
chemiluminescent yang diukur dengan photomultiplier hasil
ditentukan melalui kurva kalibrasi instrumen khusus yang dihasilkan oleh
2 titik kalibrasi dan kurve standar yang terdapat pada barcode reagen.
Nilai normal: 6 µIU/mL –29 µIU/mL (43 pmol/L SI units –208 pmol/L SI
units).
29

BAB 4RINGKASAN
Leptin merupakan hormon berperan dalam pengaturan keseimbangan
energi. Leptin memberikan pengaruh terhadap asupan makanan, pengeluaran
energi, berat badan dan fungsi neuroendokrin melalui targetnya pada sel saraf di
hipotalamus Kadar leptin sebanding dengan jumlah jaringan adiposa dalam tubuh.
Leptin diekspresikan pada banyak jaringan di perifer melalui reseptornya.
Reseptor leptin terdapat pada pankreas, hepar, otot rangka dan jaringan adiposa,
aksi leptin secara langsung maupun tidak langsung berperan dalam mengontrol
homeostasis glukosa.
Resistensi leptin merupakan keadaan berkurangnya atau tidak adanya
respons hormon ini dalam memberikan efek menghambat nafsu makan dan
mengontrol berat badan, resistensi leptin ditemukan pada kebanyakan individu
obesitas. Terjadinya resistensi leptin kemungkinan disebabkan kegagalan leptin
dalam menembus sawar darah otak akibat adanya gangguan transpor leptin di
susunan saraf pusat, berkurangnya reseptor leptin pada sel target, defek pada
reseptor (akibat proses mutasi) dan adanya polimorfisme reseptor.
Leptin dan insulin mempunyai peranan penting di SSP dan perifer dalam
mengontrol homeostasis energi. Resistensi leptin diketahui berhubungan dengan
resistensi insulin melalui aksinya di sentral maupun di jaringan perifer. Insulin
dan leptin merupakan sinyal adiposit yang beraksi di nukleus arkuatus
hipotalamus untuk menghambat ekspresi neuron NPY/AGRP dan menstimulasi
ekspresi POMC/CART. Pada keadan resistensi leptin terjadi kegagalan transport
leptin melintasi sawar darah otak sehingga terjadi insufisiensi sinyal leptin di
30

hipotalamus menyebabkan hiperglikemia dan hiperinsulinemia, yang akan
mengarah ke reistensi insulin atau bahkan diabetes melitus
Hubungan antara sinyal intraselular insulin dan leptin di duga melalui
enzim PI3K. Leptin berikatan dengan reseptornya (ObRb), menyebabkan
penggabungan dengan enzim tyrosine kinase enzyme JAK-2. Reseptor tirosin
kinase insulin akan mengatalisis protein IRS-2 yang berhubungan dengan PI3K,
menghasilkan aktivasi enzim PI3K yang menurunkan transduksi sinyal pada
jalur neuronal dan mengakibatkan hambatan pada asupan makanan. Diduga
bahwa aksi JAK-2 melalui suatu yang mirip dengan mekanisme IRS-dependent
untuk mengaktivasi PI3K. Stimulasi PI3K mengaktivasi PDK1 yang akan
mengaktivasi Akt, selanjutnya melaui serine/threonine kinase merupakan langkah
penting dalam downregulasi sinyal. Di susunan saraf pusat PI3K-dependent
mengaktivasi PDE3B dan reduksi cAMP. Leptin juga menstimulasi IRS2-
dimediasi oleh aktivitas hipotalamik PI3K, yang menghambat PI3K pada leptin
dan menginduksi hiperpolarisasi neuron NPY/AgRP.
Pemeriksaan laboratorium leptin dapat dilakukan dengan metode ELISA
dan HPLC, sedangkan pemeriksaan laboratorium insulin dengan metode ECLIA.
31