



![0& · 2 &!!!!m))nopq!)"*+$#"(%,&-./0&j&6%,&e3(a($%&4#,(%,&3,&26*4*(2,2&&&&&;h.0?.c& 3 -./0&j&6%,&e3(a($%&4#,(%,&3,&26*4*(2,2&!]"&0ws&e3(a($%&3,&)"*+$#"(%,b&#,&4#62 ...](https://static.fdocument.pub/doc/165x107/6020c2c04c7b540c9043abf1/0-2-mnopq-0j6e3a4326422h0c.jpg)



Languages
Pages
Legal


!
!!!!!!!!
!!!!!!!!!!!!!!!!!
!"#!$!%!&"'()'*+',)-*!.+%!&"'(/'0!#'-+,'/"'()"(,!1),)2-)34526$(4!
!"#$%&'(%'"%)*%")*%'+,-',./,'0121"34&5'+"'!67!68'0%9:!
!
!
!
!
!
!
!
! !
Dendrimer Team
Chardon Florian, Gomes Aurélie, Hoareau Coralie, Planas Delphine
Master 2 Expression Génique
et Protéines Recombinantes


!
Remerciements
En premier lieu, nous tenons à remercier Cédric-Olivier (CO) Turrin pour le temps
qu’il nous a consacré et pour avoir continué à nous encourager même pendant nos
« grands » moments de faiblesses.
Nous remercions également Fabrice Dumas pour avoir écouté attentivement notre
histoire de dendrimère-inhibiteur de la réplication virale, et pour nous avoir éclairé dans ce
vaste domaine qu’est le SIDA.
Merci à Nathalie Doncescu et Honoré Mazarguil de nous avoir fait partager leur savoir
en chimie de synthèse.
Nous remercions l’ensemble de l’équipe pédagogique EGPR pour avoir répondu à nos
questions.
Enfin un grand merci à notre « Papa », Rémy Poupot, pour nous avoir initié au monde
des dendrimères mais aussi d’avoir toujours été disponible entre deux longueurs de piscine.
Dendrimer Team
Florian, Aurélie, Coralie et Delphine


Résumé
Le virus de l’immunodéficience humaine a été mis en évidence au début des années
1980. Ce rétrovirus est responsable du syndrome de l’immunodéficience acquise (SIDA) chez
l’Homme. Malgré les avancées de la science sur la biologie et le mode d’infection de ce virus
environ 3 millions de personnes sont infectées chaque année. De nouvelles cibles
thérapeutiques sont activement recherchées, notamment pour augmenter l’efficacité de
l’HAART (highly active antiretroviral therapy).
Les récepteurs aux chémokines jouent un rôle important dans la régulation de
l’immunité des muqueuses parmi eux le récepteur CCR6. Il a été mis en évidence par
différents travaux que l’activation de ce récepteur par hBD2 (humaine !-defensin2) induisait
une expression d’APOBEC3G (apolipoptotein B mRNA-editingcatalytic polypeptide-like
3G), mécanisme naturel de défense antivirale.
Dans le cadre de notre étude, la construction d’un dendrimère portant à sa surface le
peptide hBD2 sera réalisé. Les propriétés de multivalence des dendrimères permettraient
d’augmenter l’avidité de liaison de hBD2 pour CCR6 et ainsi augmenter considérablement
l’expression d’APOBEC3G, ce qui inhiberait la réplication virale.
Des résultats concluants pourraient mener à une étude clinique chez l’Homme.


Abréviations
!M : micro mol
" : facteur de multivalence
ADN : acide désoxyribonucléique
AMP : adénosine monophosphate
APOBEC3G: apolipoprotein B mRNA-editingcatalytic polypeptide-like 3G
ARN : acide ribonucléique
AZT : azidothymidine
CD : cellule dendritique
DAB : di-amino-butane
DHFR : Dihydrofolate réductase
DICT50 : dose de virus qui, injectée dans une culture cellulaire, peut détruire 50 % des cellules
DMSO : diméthylsulfoxyde
DO : densité optique
DOSY : Diffusion Ordered SpectroscopY
EDA : éthylènediamine
Elisa : enzyme-linked immunosorbent assay
FA : acide folique
FBP : Protéine de liaison au folate
fmoc : Fluorenylmethyloxycarbonyl chloride
FR : récepteur au folate
GALT : gut-associated lymphoïd tissue
HAART : highly active antiretroviral therapy
hBD2 : beta-defensine2 humaine
HPLC : chromatographie liquide à haute performance
HSQC : Heteronuclear single quantum coherence
IgG : immunoglobuline G
IPBS : institut de pharmacologie et de biologie structurale
KD : constante de dissociation
Ki : constante d’inhibition
koff : vitesse de dissociation
kon : vitesse d’association
LTR : Long terminal Repeat


MBHA : 4-Methylbenzhydrylamine Hydrochloride
MTT : 3-(4,5-diméthylthiazol-2-yl)-2,5-diphényl tétrazolium
MTX : méthotrexate
NADP : nicotinamide adénine dinucléotide phosphate
NADPH : nicotinamide adénine dinucléotide phosphate oxydase
NHS : N-Hydroxysuccinimide
nm : nanomètre
PAMAM: polyamidoamine
PBMC : Peripheral Blood MononuclearCell
PE : phycoérythrine
PEG: polyéthylène glycol
PPI: poly (propylèneimine)
qPCR : réaction en chaîne de polymérisation quantitative
RFR : reduced folate carrier
RMN : résonnance magnétique nucléaire
RU : unité de réponse
SDS-PAGE : sodium dodecyl sulfate-polyacrylamide gel electrophoresis
SIDA : syndrome de l’immunodéficience acquise
TFA : acide trifluoro acétique
UPLC : Chromatographie Liquide Ultra Performante
VIH : virus de l’immunodéficience humaine
XTT: 2,3-bis-(2-methoxy-4-nitro-5-sulfophenyl)-2H-tetrazolium-5-carboxanilide
!


!
TABLE DES MATIERES
I. INTRODUCTION AU PROJET DE RECHERCHE 1
I.1. Le VIH 1
I.2. Une nouvelle voie d’action : la voie CCR6 2
I.3. La multivalence 3
I.4. Les dendrimères 3
II. LA MULTIVALENCE DES DENDRIMERES PERMET UNE AUGMENTATION
DE LA REPONSE BIOLOGIQUE 5
II.1. Utilisation d’un dendrimères multivalent pour cibler les récepteurs à
l’acide folique dans le cadre de la lutte contre le cancer (TP. Thomas et al.,
2012) 5
II.2. Résultats 5
II.2.1. Synthèse de plusieurs dendrimères 5
II.2.2. Détermination des constantes d’affinités 6
II.2.3. Internalisation du G5-5T3-MTX10 6
II.2.4. Test à la DHFR (dihydrofolate réductase) 7
II.2.5. Cytotoxicité des conjugués G5-MTXn 7
II.2.6. Cytotoxicité du conjugué G5-MTX10 : 8
II.3. Conclusion 8
III. PROJET DE RECHERCHE 9
III.1.Synthese chimique 9
III.1.1.Choix des dendrimères 9
III.1.1.1.Les dendrimères poly (amido amine) ou PAMAM 9


!
III.1.1.2.Les dendrimères Poly (propylène imine) ou PPI 9
III.1.1.3.Les dendrimères poly mélamines 10
III.1.2. Cytotoxicité intrinsèque aux dendrimères 10
III.1.3. Synthèse chimique de hBD2 10
III.1.4. Greffage du PEG sur le dendrimère 11!
III.1.5. Greffage de hBD2 sur les dendrimères-PEG24 12
III.1.6. Dendrimères fluorescents 12
III.2. Caractérisation des macromolécules par RMN 13!
III.3. Criblage des dendrimères 13
III.3.1. Lignées cellulaires. 13
III.3.2. Cytotoxicité des dendrimères-protéique 14
III.3.3. Tests de compétitions 14
III.3.4. Tests d’activité 15
III.3.5. Infection des cellules par le VIH 16
III.3.6. Expression d’APOBEC3G et taux de Vif 16
III.3.7. Test d’inhibition de la réplication virale 16
III.3.8. Taux d’encapsidation d’APOBEC3G 17
IV. DISCUSSION ET PERSPECTIVES 18
V. REFERENCES 19
VI. ANNEXES 21


! ! "!
I. INTRODUCTION AU PROJET DE RECHERCHE
I.1. Le VIH
Le virus de l’immunodéficience humaine (VIH) a été mis en évidence au début des
années 1980. Ce rétrovirus est responsable du syndrome de l’immunodéficience acquise
(SIDA) chez l’Homme, qui se traduit par un affaiblissement du système immunitaire et donc
une vulnérabilité aux maladies opportunistes (tuberculose, zona, gale, toxoplasmose …)
(K.Pluta et MM. Kacprzak, 2009).
L’infection par le VIH est initiée par l’interaction entre une glycoprotéine de
l’enveloppe du virus (gp120) et la glycoprotéine CD4 de la cellule cible. Cela entraine une
cascade d’interactions moléculaires entre gp120 et les co-récepteurs CCR5 et/ou CXCR4
aboutissant à une fusion de l’enveloppe virale à la membrane plasmique. La capside virale
contenant l’ARN-viral, la reverse transcriptase, l’intégrase et d’autres protéines virales est
libérée dans la cellule hôte. L’ARN-viral subit alors une transcription inverse simultanément à
sa décapsidation et à sa migration vers le noyau le long des microtubules. L’ADN-viral est
transporté dans le noyau via les complexes de pores nucléaires et est intégré à l’ADN de la
cellule hôte. L’ARN et les protéines virales peuvent alors être synthétisés en utilisant la
machinerie de la cellule hôte. Ils sont ensuite transportés jusqu’à la membrane plasmique d’où
bourgeonne de nouveaux virions (K.Pluta et MM. Kacprzak, 2009,NIAIDS).
Certaines cellules immunitaires telles que les lymphocytes T CD4+, les macrophages,
les cellules dendritiques (CD) et les cellules microgliales présentent à leur surface la
glycoprotéine CD4 en combinaison avec les récepteurs aux chémokines CCR5 et/ou CXCR4
faisant de ces cellules des cibles potentielles du VIH (V.A. Evans et al., 2012; MM.
Goodenow et RG. Collman 2006 ; K.Pluta et MM. Kacprzak, 2009). Les personnes infectées
possèdent donc un pool de cellules immunitaires considérablement diminué expliquant
l’affaiblissement de leur système immunitaire. Durant la phase précoce d’infection, la quantité
de lymphocytes T CD4+est fortement diminuée, ce qui entraine de graves conséquences dans
le GALT (gut-associated lymphoïd tissue), site majeur de lutte contre les infections de la
muqueuse intestinale.
Malgré les avancéesde la science sur la biologie et le mode d’infection de ce virus,
environ 3 millions de personnes sont infectées chaque année. En 2011, 34 millions de
personnes sont connues porteuses du virus (Organisation Mondiale de la Santé :
!
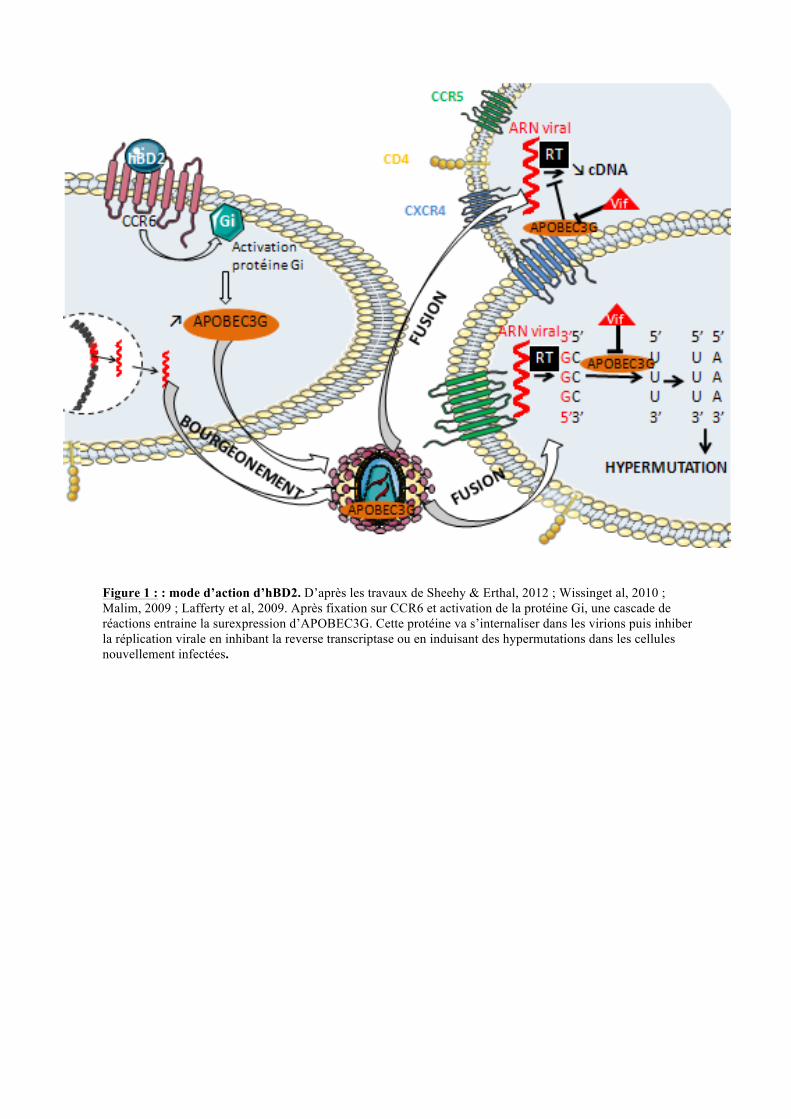
Figure 1 : : mode d’action d’hBD2. D’après les travaux de Sheehy & Erthal, 2012 ; Wissinget al, 2010 ; Malim, 2009 ; Lafferty et al, 2009. Après fixation sur CCR6 et activation de la protéine Gi, une cascade de réactions entraine la surexpression d’APOBEC3G. Cette protéine va s’internaliser dans les virions puis inhiber la réplication virale en inhibant la reverse transcriptase ou en induisant des hypermutations dans les cellules nouvellement infectées.

! ! #!
http://www.who.int/fr/). De nouvelles thérapies sont attendues car à ce jour il n’existe pas de
traitement curatif et des mécanismes de résistance aux médicaments actuels sont observés.
C’est pour cela que de nouvelles cibles thérapeutiques sont activement recherchées,
notamment pour augmenter l’efficacité de l’HAART (highly active antiretroviral therapy),
aussi appelée trithérapie (J.A Esté et T.Cihlar, 2009)
I.2. Une nouvelle voie d’action : la voie CCR6
Les récepteurs aux chémokines jouent un rôle important pour la mobilisation des
cellules immunitaires dans l’organisme. Des études ont mis en évidence l’importance de ces
récepteurs transmembranaires couplés aux protéines G, dans la régulation de l’immunité des
muqueuses ainsi que leur capacité à recruter les diverses cellules de l’immunité vers les sites
de l’inflammation (MK. Lafferty et al., 2010).
Parmi eux, le récepteur CCR6 est notamment exprimé par les cellules de l’épithélium
intestinal, les cellules dendritiques et les lymphocytes T CD4+. De plus, il a été montré que les
cellules CD4+CCR6+, largement retrouvées dans le GALT (site majeur d’infection du VIH),
sont hautement permissives à l’infection du VIH en comparaison aux cellules CD4+CCR6-. La
prédisposition à l’infection par le VIH des cellules CD4+CCR6+ font de celles-ci une cible
intéressante pour la conception de nouvelles stratégies thérapeutiques (A. Gosselin et al.,
2010).
Les récepteurs CCR6 possèdent deux ligands : CCL20 et hBD2 (beta-defensine2
humaine). Le peptide hBD2 (4.3kDa) est sécrété par les cellules épithéliales de nombreux
organes et son expression est induite lors de l’infection par le VIH (A. Weinberg et al ;,
2006). De plus, il a été démontré que hBD2 possède une activité inhibitrice anti-VIH sans
présenter d’effets négatifs sur le métabolisme cellulaire même à très haute concentration (L.
Sun et al. 2005). Les travaux de Mark K. Lafferty, ont mis en évidence une voie impliquant
hBD2 et CCR6, permettant de diminuer la réplication du VIH (figure 1). En effet, après
interaction de hBD2 avec le récepteur CCR6, les protéines Gi activées vont induire une
cascade de réactions, qui va provoquer une augmentation de l’expression d’APOBEC3G
(apolipoprotein B mRNA-editingcatalytic polypeptide-like 3G). La protéine APOBEC3G,
possédant la fonction de cytosine désaminase, est alors empaquetée dans les virions (MK.
Lafferty et al., 2010 ; MH. Malim., 2009). Lorsque ces derniers infectent de nouvelles
cellules, les produits de la réverse transcription portent alors des mutations dues à
APOBEC3G. Les cystéines transformées en uraciles par la réaction de désamination induisent


! ! $!
des hypermutations de l’ADN viral qui provoquent alors sa dégradation. Il a aussi été montré
qu’ APOBEC3G inhibe la reverse transcriptase mais le mécanisme n'est pas connu (EN.
Newman et al., 2005). Cependant, il a été montré que les virus peuvent vaincre les
mécanismes de défense intracellulaire (KN. Bishop et al., 2012)). En effet, l’ADN viral code
pour des protéines dites accessoires/régulatrices telles que la protéine Vif. Cette protéine
interagit avec APOBEC3G formant un complexe qui provoque l’ubiquitinylation et la
dégradation d’APOBEC3G par le protéasome.Cependant, les travaux d’Andrew Mehle
suggèrent que la surexpression d’APOBEC3G provoque la polyubiquitinylation et la
dégradation par le protéasome de Vif. Ainsi, Vif et APOBEC3G possèdent une relation
interdépendante qui promeut leur ubiquitinylation mutuelle et ainsi leur dégradation. Ces
résulats suggèrent donc qu’APOBEC3G pourrait contrer l’effet de Vif ( A. Mehle et al.,
2003).
I.3. La multivalence
Le concept de multivalence repose sur l’augmentation d’une interaction entre un
récepteur et son ligand par le biais d’une macro molécule portant plusieurs ligands à sa
surface, multipliant ainsi les interfaces avec le récepteur. Si l’un des ligands attaché à la
macro molécule interagit avec son récepteur, alors les ligands sont rapprochés des récepteurs
voisins. Il y a ainsi une augmentation des probabilités de rencontres ligand-récepteur. Cela
augmente l’affinité du ligand pour le récepteur, on parle alors d’avidité ou d’affinité
fonctionnelle.
Notre étude se portera donc sur la construction d’un dendrimère portant à sa
surface le peptide hBD2. Les propriétés de multivalence des dendrimères permettraient
d’augmenter l’avidité de liaison de hBD2 pour CCR6 et ainsi augmenter
considérablement l’expression d’APOBEC3G, ce qui contrerait les effets de la protéine
virale Vif et permettrait d’inhiber la réplication virale.
I.4. Les dendrimères
Les dendrimères sont des macromolécules hyper ramifiées de structure parfaitement
définie, leurs caractéristiques physico chimiques sont donc contrôlables. Ils sont caractérisés
par six paramètres structuraux: la nature du cœur, les branches, les points de divergence, la
génération, les groupements terminaux et les éventuelles molécules greffées en surface. La
synthèse de dendrimères est simple, elle se fait le plus souvent de manière dite « divergente »,

!
!
!
!
!
!
!
!
!
!
!
!
Figure 2 : Schématisation de la synthèse divergente d’un dendrimère. r1 : greffage des branches sur le cœur. r2: ajout de points de divergence. Etapes répétées jusqu’à l’obtention de la génération voulue.

! ! %!
du cœur vers la périphérie (figure 2). Elle consiste en la répétition d'une séquence de
réactions. La 1ère réaction consiste en un greffage sur le cœur de branches, puis une 2ème
réaction permet l’ajout de points de divergence. Ces deux étapes sont ensuite répétées jusqu’à
l’obtention de la génération voulue. Enfin, les groupements terminaux sont ajoutés aux
branches formant la surface multivalente qui peut contenir un grand nombre de fonctions
capables d’interagir avec l’environnement externe.
Les dendrimères sont intéressants du fait de leur faible dispersité, de leur morphologie
spécifique souvent globulaire (R. Esfand et al.,2001), de leur multivalence, de leur densité en
groupements fonctionnels, et de leur poids moléculaire contrôlable. La première synthèse de
dendrimère a été réalisée par Vögtle et al. à la fin des années 1970. En 1993, Mülhaupt et al.
et Meijer et al. ont décrit pour la première fois la préparation des dendrimères poly (propylène
imine) (PPI) à cœur diaminobutane. Le groupe de Tomalia a synthétisé des macromolécules
polyamidoamines (PAMAM) branchées en forme d’étoile. Il est le premier à employer le
terme « dendrimère ». Plus récemment, E. Simanek et al. en 2001, ont synthétisé un nouveau
type de dendrimère basé sur les mélamines (polymélamine) présentant des sites amines à leur
surface.
Dans le cadre de l’étude présenté, les dendrimères seront utilisés comme support pour
étudier l’effet de multivalence afin d'augmenter la réponse biologique. De plus, en maîtrisant
hautement la structure, les paramètres physico chimiques pourront être adaptés, tels que leur
cytotoxicité, leur solubilité, leur taille et donc leur valence. De plus, il sera aisé d’ajouter des
molécules fluorescentes à la surface des dendrimères pour des études d’interaction avec les
récepteurs.
Dans cette étude les dendrimères PAMAM, PPI et poly mélamine de générations 4 et
5 vont être utilisés. Ils présenteront des groupements amines à leur surface simplifiant la
liaison de la protéine au dendrimère. La protéine hBD2 sera liée aux dendrimères par le biais
d'une molécule de PEG (poly-éthylène glycol) servant d’espaceur flexible facilitant ainsi
l'interaction entre hBD2 et son récepteur.
Il sera nécessaire de cribler ces dendrimères afin de déterminer le meilleur candidat
selon la biocompatibilité, la toxicité, la solubilité et l’efficacité d'interaction avec CCR6.
L’utilisation de dendrimère-PEG-HBD2 a pour but de cibler les cellules exprimant le
récepteur CCR6 à leur surface afin d’augmenter l’avidité de liaison. Cette interaction

!
Figure 3 : mode d’action du méthotrexate. Le MTX peut être internaliser dans la cellules via les récepteurs à l’acide folique (FR) ou les transporteurs RFC (reduced folate carrier), afin d’inhiber la DHFR (enzyme clé de la voie métabolique de la biosynthèse des nucléotides), aboutissant à la mort cellulaire.
! !

! ! &!
permettra d'augmenter l’expression d’APOBEC3G dans les nouveaux virions pour induire des
hypermutations sur l'ADN viral et inhiber sa réplication dans les cellules infectées. Ce
processus permettrait d’avoir un effet sur l’ensemble des cellules y compris les cellules
inaccessibles par la trithérapie (cellules présentes dans le cerveau, cellules en latence…). En
effet, l’ajout d’hypermutations sur l’ADN viral par APOBEC3G couplé aux drogues de la
trithérapie (anti-intégrase, anti-protéase, inhibiteur de la réverse transcriptase) permettrait
d’augmenter l’espérance de vie des patients.
II. LA MULTIVALENCE DES DENDRIMERES PERMET UNE AUGMENTATION
DE LA REPONSE BIOLOGIQUE
II.1. Utilisation d’un dendrimère multivalent pour cibler les récepteurs à
l’acide folique dans le cadre de la lutte contre le cancer (TP. Thomas et al.,
2012)
Le méthotrexate (MTX) est une molécule cytotoxique chémothérapeutique utilisée dans le
traitement de cancers. Son rôle est d’inhiber la DHFR (Dihydrofolate réductase), enzyme
impliquée dans le métabolisme de biosynthèse des nucléotides (figure 3). Il a été
précédemment montré que le couplage de nanoparticules aux molécules chémothérapeutiques
présente plus d’avantages que l’utilisation de la molécule seule. Les travaux précédents de JR.
Baker ont démontrés l’efficacité de dendrimères couplés à l’acide folique (FA), permettant le
ciblage des cellules exprimant le récepteur au folate, et à du MTX par une liaison ester.
Cependant la liaison de deux molécules différentes aux dendrimères entrainait des problèmes
d’homogénéité de synthèse. Ainsi, cette étude permet de démontrer le maintient de l’activité
biologique du MTX lié covalamment aux dendrimères par l’intermédiaire d’un espaceur
cyclooctyne.
II.2. Résultats
II.2.1. Synthèse de plusieurs dendrimères
Pour cette étude, plusieurs dendrimères ont été synthétisés à partir du dendrimère poly
(amido-amine) de génération 5 (G5-PAMAM). Le nombre de molécules de MTX fixées à la
surface du dendrimère par l’intermédiaire d’un espaceur cyclooctyne a un effet sur la
multivalence et la toxicité de celui-ci. Les G5-MTXn (n=5, 10, 17) et l’analogue fluorescent,
le G5-5T3-MTX10 ont donc été synthétisés. Le nombre de molécules de MTX fixées à la
surface des dendrimères ainsi que leur pureté a été vérifié respectivement par RMN du proton
et par UPLC (Chromatographie Liquide Ultra Performante).
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!! !
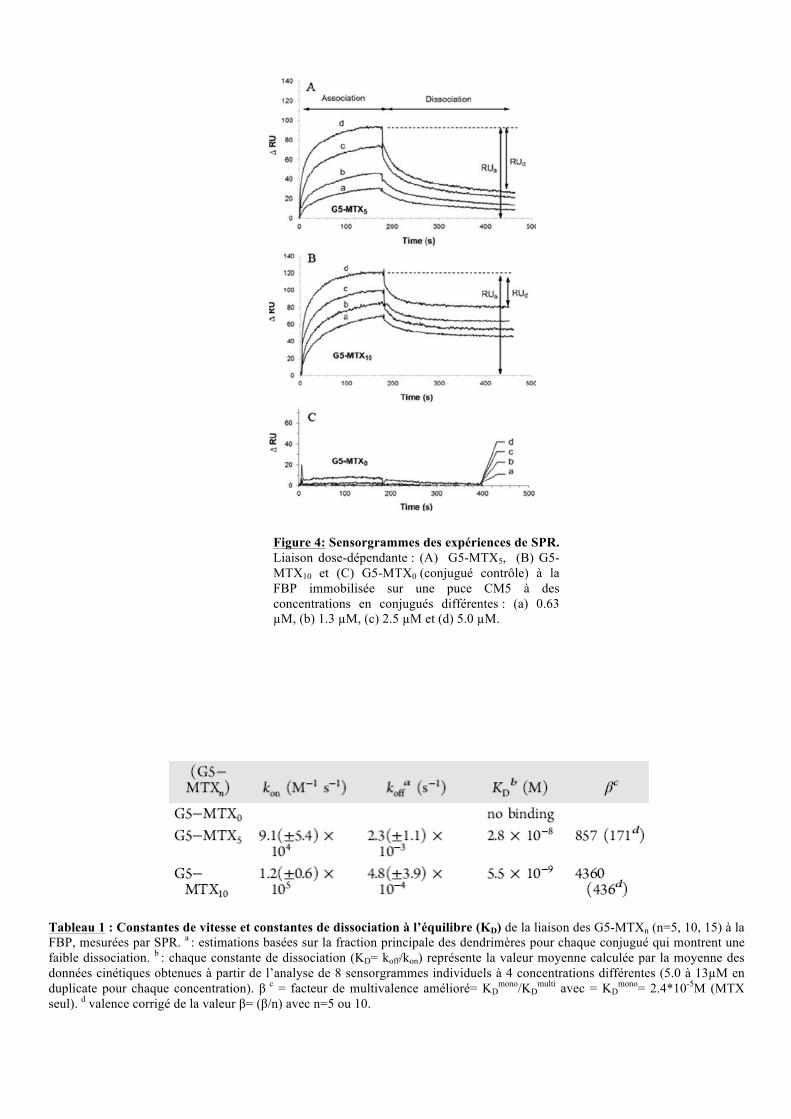
Tableau 1 : Constantes de vitesse et constantes de dissociation à l’équilibre (KD) de la liaison des G5-MTXn (n=5, 10, 15) à la FBP, mesurées par SPR. a : estimations basées sur la fraction principale des dendrimères pour chaque conjugué qui montrent une faible dissociation. b : chaque constante de dissociation (KD= koff/kon) représente la valeur moyenne calculée par la moyenne des données cinétiques obtenues à partir de l’analyse de 8 sensorgrammes individuels à 4 concentrations différentes (5.0 à 13!M en duplicate pour chaque concentration). " c = facteur de multivalence amélioré= KD
mono/KDmulti avec = KD
mono= 2.4*10-5M (MTX seul). d valence corrigé de la valeur "= ("/n) avec n=5 ou 10.
!
Figure 4: Sensorgrammes des expériences de SPR. Liaison dose-dépendante : (A) G5-MTX5, (B) G5-MTX10 et (C) G5-MTX0 (conjugué contrôle) à la FBP immobilisée sur une puce CM5 à des concentrations en conjugués différentes : (a) 0.63 !M, (b) 1.3 !M, (c) 2.5 !M et (d) 5.0 !M.
!

! ! '!
II.2.2. Détermination des constantes d’affinités
Afin de déterminer l’affinité des différents dendrimères pour les récepteurs folates, des
expériences de spectroscopie par résonance plasmonique de surface (SPR) ont été réalisées.
Ces expériences permettront de suivre la cinétique de liaison des dendrimères à la FBP
(Folate Binding Protein de lait bovin).
Les expériences réalisées avec le conjugué G5-MTX0 (contrôle négatif) ne montrent
aucune liaison significative avec la FBP (figure 4C). Au contraire, les conjugués G5-MTX5
(figure 4A) et le G5-MTX10 (figure 4B) montrent une cinétique de liaison dépendante de la
concentration. De plus, à temps et à concentration égal le G5-MTX10 présente une meilleure
association soit une moins bonne dissociation que le G5-MTX5. Ceci est illustré par le
pourcentage de désorption (RU dissociation/RU association), 62% pour le G5-MTX5 contre 39% pour
le G5-MTX10, lors d’une injection de 5!M de conjugué. Ces résultats sont confirmés par la
détermination des constantes cinétiques (kon, koff) obtenues par régression non-linéaire
(tableau 1). En effet, le kon du G5-MTX5 est plus faible que celui du G5-MTX10, et
inversement pour le koff. Cela démontre bien une association plus rapide et une dissociation
plus lente du G5-MTX10 pour la FBP.
Le KD (koff/kon), reflet de l’affinité de liaison, est la concentration de conjugués
nécessaire à la saturation de 50% des sites de liaisons à la FBP. Le tableau 1 montre que le
G5-MTX10 a un KD 5 fois inférieur au G5-MTX5, il est donc 5 fois plus affin. De plus, le
facteur " (KDmono/KD
multi) montre que les conjugués G5-MTXn ont une affinité supérieure au
MTX libre. La valeur !/n indique qu'une interaction multivalente entre le MTX et la FBP a
une plus grande affinité fonctionnelle (avidité) qu’une liaison monovalente.
Ces résultats mettent en évidence l’avidité des dendrimères couplés au MTX pour les
récepteurs folates. Leur internalisation dans les cellules surexprimant le récepteur au folate et
l’effet inhibiteur de la DHFR (dihydrofolate réductase) ont ensuite été testés.
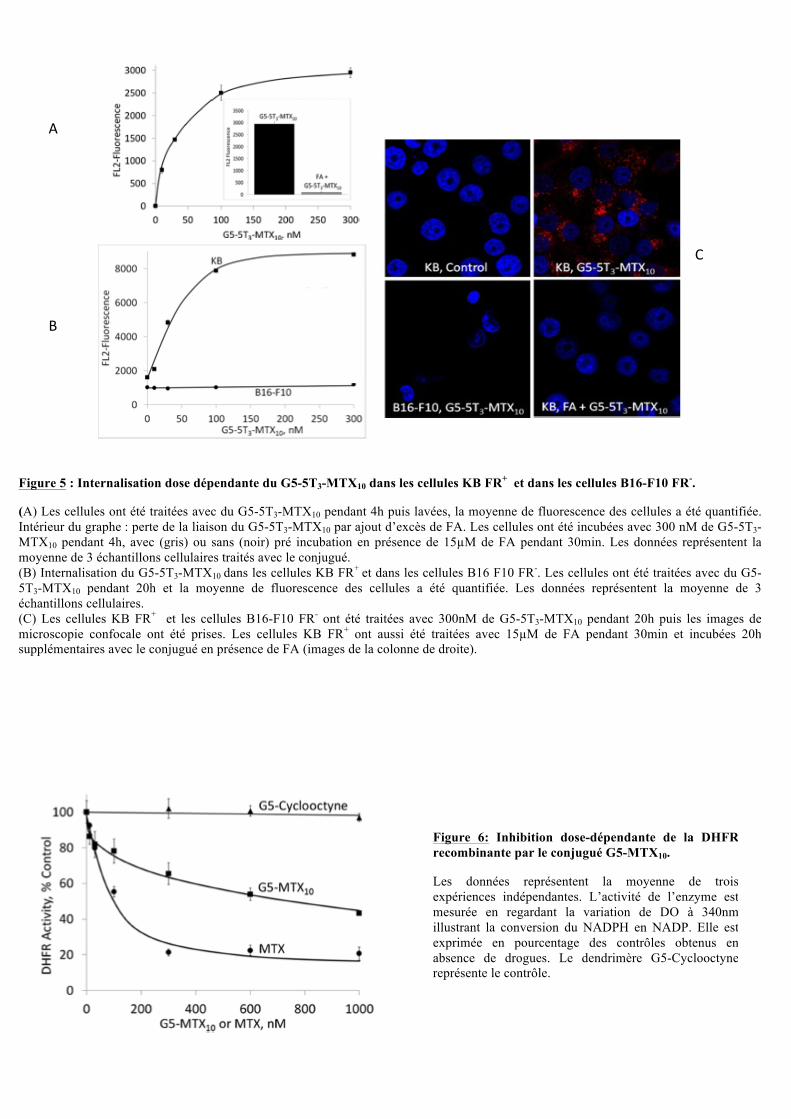
II.2.3. Internalisation du G5-5T3-MTX10
Dans le but de caractériser l’internalisation du conjugué fluorescent G5-5T3-MTX10 dans
les cellules après interaction avec les récepteurs folate (FR), des expériences de FACS ont été
réalisées (Figure 5). L’hyperbole de la fluorescence en fonction de la concentration en G5-
5T3-MTX10 dans les cellules KB suggère que l’internalisation se fait via les FR. Ceci est
démontré par l’absence d’internalisation du G5-5T3-MTX10 lors d’expériences similaires en
!
!
"!
!
!
!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!! #!
!
$!
!
!
!
!
!
!
!
!
!
!
!
!
Figure 5 : Internalisation dose dépendante du G5-5T3-MTX10 dans les cellules KB FR+ et dans les cellules B16-F10 FR-.
(A) Les cellules ont été traitées avec du G5-5T3-MTX10 pendant 4h puis lavées, la moyenne de fluorescence des cellules a été quantifiée. Intérieur du graphe : perte de la liaison du G5-5T3-MTX10 par ajout d’excès de FA. Les cellules ont été incubées avec 300 nM de G5-5T3-MTX10 pendant 4h, avec (gris) ou sans (noir) pré incubation en présence de 15!M de FA pendant 30min. Les données représentent la moyenne de 3 échantillons cellulaires traités avec le conjugué. (B) Internalisation du G5-5T3-MTX10 dans les cellules KB FR+ et dans les cellules B16 F10 FR-. Les cellules ont été traitées avec du G5-5T3-MTX10 pendant 20h et la moyenne de fluorescence des cellules a été quantifiée. Les données représentent la moyenne de 3 échantillons cellulaires. (C) Les cellules KB FR+ et les cellules B16-F10 FR- ont été traitées avec 300nM de G5-5T3-MTX10 pendant 20h puis les images de microscopie confocale ont été prises. Les cellules KB FR+ ont aussi été traitées avec 15!M de FA pendant 30min et incubées 20h supplémentaires avec le conjugué en présence de FA (images de la colonne de droite).
!
Figure 6: Inhibition dose-dépendante de la DHFR recombinante par le conjugué G5-MTX10.
Les données représentent la moyenne de trois expériences indépendantes. L’activité de l’enzyme est mesurée en regardant la variation de DO à 340nm illustrant la conversion du NADPH en NADP. Elle est exprimée en pourcentage des contrôles obtenus en absence de drogues. Le dendrimère G5-Cyclooctyne représente le contrôle.

! ! (!
présence d’un excès d’acide folique, ligand naturel au FR, jouant le rôle de compétiteur
(Figure 5A). De plus, la spécificité de liaison du G5-5T3-MTX10 au FR est vérifié par
l’absence d’internalisation du conjugué dans des cellules B16-F10 FR- (Figure 5B).
L’internalisation des conjugués via une liaison spécifique aux FR est confirmée par des
analyses de microscopie confocale. Ces analyses démontrent également la présence des
conjugués dans le compartiment cytosolique (figure 5C).
II.2.4. Test à la DHFR (dihydrofolate réductase)
Pour mesurer la capacité du conjugué G5-MTX10 à inhiber l’activité de la DHFR, la
cinétique de conversion du NADPH en NADP a été suivi par spectrophotométrie à 340nm
pendant 5 minutes (Figure 6). De façon similaire au MTX libre, le conjugué G5-MTX10 inhibe
l’activité de la DHFR de façon dose-dépendant. Le contrôle négatif (G5-cyclooctyne) montre
que cette inhibition est spécifique à la présence du MTX libre ou conjugué. Cependant,
l’efficacité du G5-MTX10 est moins importante que celle du MTX libre, suggérant que le
conjugué a une moins bonne affinité pour la DHFR.
Après avoir démontré que les conjugués étaient capables de se lier aux FR, de
s’internaliser et d’inhiber l’activité de la DHFR (in vitro), des études de cytotoxicité sur les
cellules KB ont été réalisées.
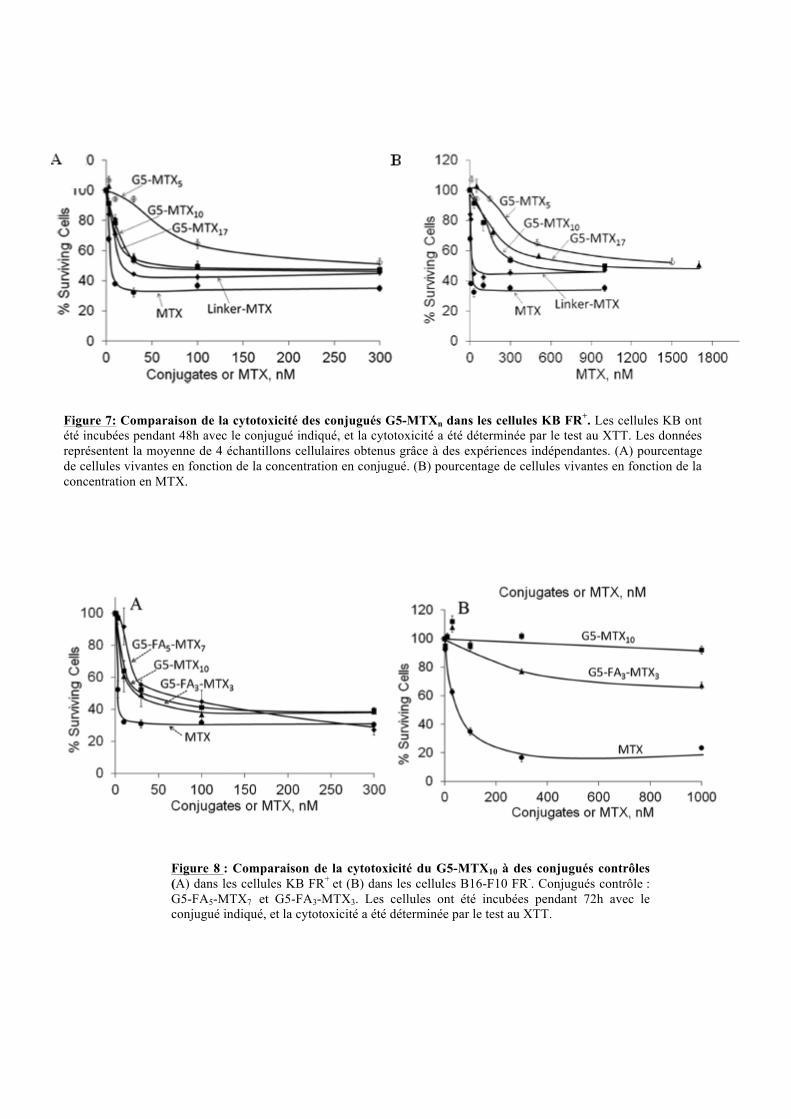
II.2.5. Cytotoxicité des conjugués G5-MTXn
L’utilisation du MTX comme drogue chémotherapeutique est due à sa cytotoxicité; le
maintient de cette fonction lorsqu’il est conjugué au dendrimère a été vérifié par des tests au
XTT, qui colore les cellules KB viables (Figure 7). Ces études démontrent que les conjugués
et le MTX libre induisent une cytotoxicité dose dépendante, que se soit en fonction de la
concentration en conjugués ou en fonction de la concentration en MTX (Figure 7A et 7B). Le
conjugué G5-MTX5 présente une plus faible cytotoxicité que les deux autres conjugués ayant
un plus grand nombre de MTX (10,17). Ceci est probablement le reflet d’une plus grande
avidité des formes G5-MTX10,17 pour le FR. Cependant, à même concentration en MTX, le
conjugué G5-MTX17 montre une moins bonne cytotoxicité que le G5-MTX10. Les auteurs
expliquent ce résultat par la diminution de la disponibilité du conjugué G5-MTX17 due à sa
plus faible hydro-solubilité. Le MTX libre reste tout de même le plus cytotoxique. De plus,
l’ajout d’un linker à ce dernier diminue sa cytotoxicité, laissant penser que le MTX libre est
internalisé via une autre voie non saturante.
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
! !
Figure 7: Comparaison de la cytotoxicité des conjugués G5-MTXn dans les cellules KB FR+. Les cellules KB ont été incubées pendant 48h avec le conjugué indiqué, et la cytotoxicité a été déterminée par le test au XTT. Les données représentent la moyenne de 4 échantillons cellulaires obtenus grâce à des expériences indépendantes. (A) pourcentage de cellules vivantes en fonction de la concentration en conjugué. (B) pourcentage de cellules vivantes en fonction de la concentration en MTX.
Figure 8 : Comparaison de la cytotoxicité du G5-MTX10 à des conjugués contrôles (A) dans les cellules KB FR+ et (B) dans les cellules B16-F10 FR-. Conjugués contrôle : G5-FA5-MTX7 et G5-FA3-MTX3. Les cellules ont été incubées pendant 72h avec le conjugué indiqué, et la cytotoxicité a été déterminée par le test au XTT.

! ! )!
II.2.6. Cytotoxicité du conjugué G5-MTX10 :
Le composé G5-MTX10 s’étant révélé être le meilleur candidat, des études de spécificité et
d’efficacité ont été réalisés en le comparant à des conjugués contrôles : G5-FA5-MTX7 et G5-
FA3-MTX3 (H. Zong et al.,2012). Dans les cellules KB, le conjugué G5-MTX10 montre une
cytotoxicité similaire aux conjugués contrôle, mais tout de même inférieure au MTX libre.
Dans les cellules B16-F10 FR-, le MTX libre et le G5-FA3-MTX3 présente une cytotoxicité
pour ces cellules contrairement au conjugué G5-MTX10 (figure 8). Le conjugué G5-FA3-
MTX7 est lié au FA et au MTX par des liaisons ester qui sont hydrolysées par des estérases
présentent dans le sérum. Cette hydrolyse permet la libération du MTX qui pourra alors
diffuser passivement via les RFC (reduced folate carrier) (figure 3). Il en résulte une forte
diminution de la spécificité du conjugué. Le G5-MTX10 possédant des liaisons amide n’est
pas sujet à ce problème, il est donc le meilleur candidat. Ce conjugué est aussi efficace que les
conjugués contrôles avec un gain de spécificité pour les cellules surexprimant les récepteurs
FR.
II.3. Conclusion
Cet article ainsi que d’autres travaux de J.R. Baker mettent en évidence les points forts des
dendrimères. Parmi eux, l’effet de multivalence, la biocompatibilité, la solubilité et l’avidité
de liaison dont la constante de dissociation peut être 4000 fois plus faible que celle du ligand
seul. Cet article a aussi mis en évidence le fait que la chimie des dendrimères est adaptative et
que le problème de spécificité due à une liaison chimique labile peut être contourné.
Cette publication justifie donc l’utilisation de dendrimères pour activer une voie de
signalisation permettant d’inhiber la réplication du VIH. Cette voie faisant intervenir une
interaction entre une protéine et son récepteur, la protéine sera greffée à la surface d'un
dendrimère afin d’augmenter l’avidité de liaison au récepteur et donc accentué l’activation de
la voie de signalisation.

!!
!
!
!
!!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
! !
Figure 9 : Structure des dendrimères. (A) PAMAM G2 : en noir, le cœur d’éthyle diamine (EDA) ; en rouge, les branches de méthyl acrylate ; en bleu, les points de divergence d’EDA ; et en marron, les groupements terminaux d’EDA. (B) PPI G2 : en noir, le cœur de diamino butane (DAB) ; en bleu, les branches divergentes de diamino propane ; et en marron les groupements terminaux amine primaire. (C) poly-mélamines G3 : en noir, le cœur de trichlorotriazide ; en bleu, les branches de piperazine ; en rouge, les points de divergence de trichlorotriazide ; et en marron, les groupements terminaux de diamino propane. !
"!#!

! ! *!
III. PROJET DE RECHERCHE
III.1.Synthese chimique
III.1.1.Choix des dendrimères
Parmi les dendrimères existant, nous nous sommes plus particulièrement intéressés à
certain d’entre eux satisfaisant des critères tels que la biocompatibilité, la disponibilité
commerciale et les groupements de surface. De plus, leur architecture particulière leur confère
des propriétés physico-chimique telles que la solubilité, la flexibilité et l’accessibilité des
groupements de surface que les polymères classiques ne possèdent pas.
III.1.1.1.Les dendrimères poly (amido amine) ou PAMAM
Les PAMAM (figure 9A) sont les premiers dendrimères à avoir été commercialisés, ils
sont aujourd’hui les dendrimères les plus répandus et les plus utilisés, notamment dans des
applications biologiques (A.K. Patri et al., 2002). Ils sont synthétisés de manière divergente
(du cœur vers la périphérie) et de telle façon que d’une génération à une autre, le poids
moléculaire ainsi que le nombre de groupement de surface du dendrimère double
approximativement.
Les dendrimères PAMAM sont composés d’un cœur central d’éthylène diamine (EDA),
de branches composées de méthyl acrylate qui vont venir se greffer sur les amines, et de
points de divergence d’EDA (figure 9A). Les groupements de surface peuvent être variables.
Dans le cadre du projet, les dendrimères PAMAM porteront à leur surface des
groupements terminaux NH2 (64 pour la génération 4 et 128 pour la génération 5) qui
permettent une solubilité dans les solvants polaires protéiques et qui s’accordent au mode de
liaison utilisé pour le greffage de la protéine au dendrimère (paragraphe 1.4) (J. Peterson et
al., 2001).
III.1.1.2.Les dendrimères Poly (propylène imine) ou PPI
Les dendrimères PPI sont aussi connus sous le nom de DAB (diaminobutane). Ces
dendrimères biocompatibles ont déjà été utilisés dans des applications biologiques,
notamment pour des drogues anti-cancer (F. Wang et al., 2012). Dans le cadre du projet le
cœur du dendrimère est une molécule de 1, 4 diaminobutane où sont greffées des branches
divergentes de 1,3 diaminopropane (figure 9B). Ces dendrimères possèdent également des


! ! "+!
NH2 à leur surface (32 pour un G4 et 64 pour un G5). (E.M.M. de Brabander-van den Berg et
E.W Meijer, 1993).
III.1.1.3.Les dendrimères poly mélamines
Les dendrimères basés sur les mélamines sont eux aussi commercialisés, biocompatibles
et solubles. Leur synthèse peut se faire de manière divergente comme convergente. Le cœur
du dendrimère est un cycle trichlorotriazide (C3N3Cl3), les branches sont composées de cycle
piperazine et de cycles trichlorotriazide (figure 9C) (E.J. Acosta et al., 2004). Le cœur et les
branches de ce dendrimère sont des cycles et peuvent jouer un rôle sur la rigidité de la
structure dendritique, et donc modifier leur comportement en milieu physiologique.
Similairement aux dendrimères PAMAM et PPI les dendrimères poly mélamines
possèdent des amines comme groupement de surface (64 pour la génération 4 et 96 pour la
génération 5).
III.1.2. Cytotoxicité intrinsèque aux dendrimères
Les dendrimères sélectionnés possèdent des fonctions NH2 à leur surface, qui vont se
protoner en milieu physiologique et former ainsi une macromolécule poly cationique toxique
pour l’organisme (F. Wang, 2012). Pour palier à cela, approximativement 50% des amines de
surface des dendrimères seront acétylées pour empêcher leur protonation (D.Astruc et al.,
2010). Par exemple, pour un G4 PAMAM présentant 64 amines à sa surface, le dendrimère
sera mis en présence de chlorure d’acétyle dans un ratio de 1 : 32.
Les amines libres restantes seront utilisées pour fixer la protéine ou la molécule
fluorescente sur le dendrimère.
III.1.3. Synthèse chimique de hBD2
La protéine hBD2 sera synthétisée chimiquement sur support solide. Le peptide hBD2 se
compose de 41 acides-aminés et de trois ponts disulfure dont la séquence est la suivante :
1
,-./!01223456!!"#$%/7!#+"#8!9:-/!;<=4>?34!35!9/!@14?34/7!"***A!
!
!
!
!
!
!
!
!
!
!
!
!
!
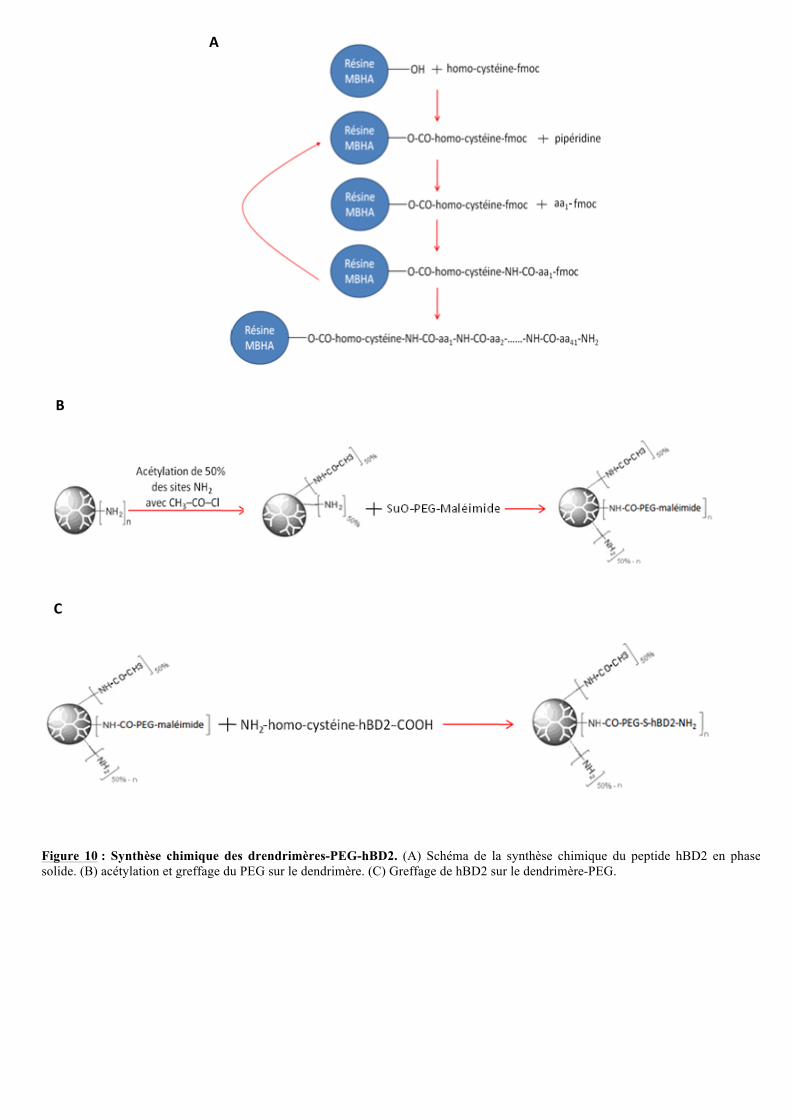
Figure 10 : Synthèse chimique des drendrimères-PEG-hBD2. (A) Schéma de la synthèse chimique du peptide hBD2 en phase solide. (B) acétylation et greffage du PEG sur le dendrimère. (C) Greffage de hBD2 sur le dendrimère-PEG.
"!
#!
$!

! ! ""!
Cette synthèse sera réalisée par l’équipe de Nathalie DONCESCU (Plateau technique et
synthèse peptidique) à l’IPBS (figure 10A). Pour permettre le greffage de hBD2 sur le
dendrimère, une homocystéine sera ajoutée en C terminal.
La protéine sera synthétisée de l’extrémité C terminale vers l’extrémité N terminale.
L’homocystéine sera fixée à la résine MBHA par une liaison ester. Sa fonction NH2 sera
protégée par un groupement fmoc (Fluorenylmethyloxycarbonyl chloride) pour éviter des
réactions indésirables. Une étape de déprotection à la pipéridine sera nécessaire avant
d’ajouter le second acide aminé. Les étapes de greffage et déprotection seront réitérées
jusqu’à l’obtention de la protéine entière. Les chaînes latérales de l’ensemble des acides-
aminés seront protégées, pour être sûr que ce soit l’amine terminale qui réagisse. La protéine
est ensuite décrochée du support et les groupements latéraux seront déprotégés au TFA (acide
tri fluoro acétique). La renaturation de la protéine sera effectuée sur colonne HPLC (high
performance liquid chromatography) en présence de DMSO (diméthylsulfoxyde, oxydant
doux), permettant la formation de trois ponts disulfure nécessaire à la bonne conformation de
la protéine. Des étapes de purification (HPLC: chromatographie liquide à haute performance)
et de caractérisation (Spectrométrie de masse) seront réalisées par l’équipe de Nathalie
DONCESCU pour vérifier l’intégrité de la protéine.
III.1.4. Greffage du PEG sur le dendrimère!
Le polyéthylène glycol (PEG) est un polyéther composé d’unités répétées d’oxyde
d’éthylène -(CH2-CH2-O)n-. Il est non chargé, hydrophile, soluble dans l’eau et dans la
plupart des solvants organiques.
Depuis plusieurs années, l’ajout de PEG à des protéines, peptides ou molécules
thérapeutique est en plein essor, notamment en pharmacologie. En effet, la PEGylation des
molécules biologiques améliore la solubilité, augmente la stabilité et le temps de rétention
dans le sang et diminue l’immunogénicité de ces molécules.
Le PEG a été également greffé à des dendrimères dans le but de diminuer leur cytotoxicité
(R. Qi et al., 2009) mais aussi comme espaceur entre le dendrimére et la partie variable d’un
anticorps (N. Nanaware-Kharade et al., 2012). De ce fait, le PEG sera utilisé dans le projet
afin d’éloigner hBD2 de l’encombrement stérique à la surface du dendrimère qui pourrait
empêcher la protéine d'interagir avec CCR6.
2


! ! "#!
D’après les résultats de J. Röhrl et al. (2012), la fusion d’une immunoglobuline G (IgG) à
l’extrémité C-terminale de hBD2 ne perturbe ni sa liaison au récepteur CCR6 ni son
activation. Ainsi, la protéine hBD2 sera lié au PEG via son extrémité C-terminale.
Le PEG utilisé pour ce projet sera composé de ving-quatre unités d’oxyde d’éthyléne
(PEG24) (N. Nanaware-Kharade et al., 2012). Pour permettre les réactions de liaison du PEG24
au dendrimère puis à la protéine sans qu’il y ait de réactions inter moléculaires indésirables
(entre deux molécules de PEG24 ou entre une molécule de PEG24 et deux dendrimères), les
molécules de PEG24 porteront une extrémité maléimide, qui réagira avec l’homocystéine de
hBD2, et une extrémité acide carboxylique qui permettra la liaison à la surface des
dendrimères (Bioconjugates Techniques, 2eme Edition, G.T. Hermanson).
Pour greffer le PEG24 sur le dendrimère, l’extrémité acide carboxylique sera activée
par de l’anhydride succinique. En mettant les PEG24 activés en présence de dendrimères, les
groupements amines primaires de surface vont réagir avec les extrémités succinimidyl ester
du PEG24 (figure 10B).
Le nombre de protéines liées à la surface du dendrimère pouvant influencer la réponse
biologique, plusieurs synthèses seront réalisées en faisant varier le nombre de protéines liées
au dendrimère. Pour chacun d’entre eux, 5, 10 ou 15 équivalents de PEG24 activés seront mis
en présence d’un équivalent de dendrimère. Ainsi des dendrimères liés à un nombre moyen de
molécules de PEG24 égal à 5, 10 ou 15 seront obtenus.
III.1.5. Greffage de hBD2 sur les dendrimères-PEG24
Dans un deuxième temps, les protéines synthétisées chimiquement seront greffées aux
conjugués dendrimères-PEG24 via une liaison thio éther stable (figure 10C), grâce à la
maléimide du PEG qui va réagir avec la fonction thiol de l’homocystéine en C-terminal de
hBD2. Il en résultera un nombre moyen de protéines liées via le PEG24 au dendrimère égal à
5, 10 ou 15.
III.1.6. Dendrimères fluorescents
Afin de pouvoir suivre les dendrimères au cours des tests de compétition, il est nécessaire
de marquer les dendrimères. L’utilisation de fluorophores présente des avantages, notamment
la simplicité de synthèse, et de manipulation. Pour marquer des dendrimères présentant des
amines primaires à leur surface, il est possible d’utiliser de la 5-carboxytetramethyl-
rhodamine, succimidyl ester (NHS rhodamine). Cette molécule dérivée de la rhodamine a une


! ! "$!
longueur d’onde d’excitation de 546nm et un maximum d’émission à 579nm. La fonction
succinimidyl ester est hautement réactive, elle pourra ainsi réagir avec les amines primaires
présentes à la surface des dendrimères. Cinq équivalents de NHS rhodamine seront mis en
présence d’un équivalent de dendrimère, comme il a été décrit par I.J.Majoros et al. en 2006.
L'ensemble des dendrimères qui seront synthétisés pour le projet sont présentés en
annexe 1.
III.2. Caractérisation des macromolécules par RMN!
Les produits dendrimères-PEG24-hBD2 seront purifiés par dialyse et leur pureté sera
analysée par HPLC.
Afin de caractériser la structure dendritique, des expériences de résonnance
magnétique nucléaire (RMN) du proton et du carbone sont requises. Le ratio PEG/dendrimère
et PEG/protéine seront déterminés par comparaison des intégrales propres à chaque composé.
Un nombre statistique de protéines conjuguées aux dendrimères sera alors déduit.
Des expériences de 1H-13C HSQC (heteronuclear single quantum correlation) aideront
à attribuer les pics des expériences RMN 1D.
Des expériences de RMN DOSY (Diffusion Ordered SpectroscopY) permettront de
déterminer les tailles des molécules ou agrégats présents dans le mélange. Les spectres des
différents composants du mélange pourront être séparés en fonction de la valeur de leurs
coefficients de diffusion apparents. D’après l’équation de Stokes-Einstein, la diffusion est
proportionnelle au rayon hydrodynamique de la molécule considérée comme sphérique.
Connaissant le coefficient de diffusion, la taille de la molécule peut donc être estimée.'
III.3. Criblage des dendrimères
Après avoir synthétisé et caractérisé les dendrimères protéiques, une étape de criblage
sera nécessaire afin d’évaluer leur cytotoxicité et leur capacité à activer la voie APOBEC3G.
L’ensemble des tests détaillés ci-dessous permettront de sélectionner le(s) meilleur(s)
candidat(s) pour inhiber la réplication virale
III.3.1. Lignées cellulaires.
Les PBMC (Peripheral Blood Mononuclear Cell) sont des cellules mononucléaires
circulant dans le sang (monocytes et lymphocytes). Une partie de ces cellules possèdant à leur


! ! "%!
surface les récepteurs nécessaires à l’infection (CD4, CCR5, CXCR4) elles peuvent être
naturellement infectées par le VIH. Ces cellules seront isolées du sang de patients en utilisant
le kit Histopaque (Sigma-Aldrich). Les cellules T CD4+ seront séparées des PBMC par
utilisation du kit CD4+ T Cell Isolation Kit II (MiltenyiBiotec). Les cellules T CD4+CCR6+
seront isolées des T CD4+CCR6- grâce à un anticorps anti-CCR6-PE (phycoérythrine) (BD
biosciences) et des microbilles anti-PE (MiltenyiBiotec).
III.3.2. Cytotoxicité des dendrimères-protéique
Pour évaluer la cytotoxicité des dendrimères sur les cellules T CD4+CCR6+! et T
CD4+CCR6- utilisées, le test au MTT sera réalisé (In vitro toxicology assay Kit MTT based).
Ce test consiste à mesurer l’activité métabolique des cellules. En effet, les cellules
métaboliquement actives seront capables, grâce à l’activité de leur déshydrogénase
mitochondriale, de réduire le MTT (3-(4,5-diméthylthiazol-2-yl)-2,5-diphényl tétrazolium) de
couleur jaune, en formazan, de couleur bleu violet qui précipite sous forme de cristaux dans
les cellules. Les cristaux de formazan seront solubilisés dans l’isopropanol et la quantité de
formazan produite sera mesurée par densité optique à 540 nm.
Le pourcentage de toxicité est calculé par rapport aux cellules non traitées par les
dendrimères protéiques.
% cytotoxicité = 100 - !" !"# !"#!!"##$#"%!!"#$!!!"!!"#$!!"#!!"#!$%&!!"#!!"#$!!"#$%!" !"# !"#!!"##$#"%!!"!!!"#$!!!" ! !""
III.3.3. Tests de compétitions.
Afin de déterminer l’avidité de liaison entre les dendrimères-PEG24-hBD2n (n=5,
10,15) et les récepteurs CCR6, des expériences de compétition seront réalisées sur les cellules
T CD4+CCR6+. La cytométrie de flux permettra de mesurer l’intensité de fluorescence
relative à la quantité de dendrimères-PEG24-hBD2n fluorescent liés de façon spécifique et non
spécifique aux récepteurs CCR6 (liaison totale).
Pour réaliser ces expériences, une concentration croissante en dendrimères-PEG24-
hBD2n ou hBD2 seule sera mise en présence d’un excès de dendrimères-PEG24-hBD2n
fluorescent. Les constantes d’inhibition (KI) de hBD2 seule et des dendrimères-PEG24-hBD2n,
correspond à la concentration nécessaire pour déplacer 50% de la liaison des dendrimères-
PEG24-hBD2n fluorescents sur CCR6, seront mesurées. Les KI des dendrimères-PEG24-hBD2n


! ! "&!
sont attendus plus faible que le KI de hBD2 seul, reflétant ainsi l’avidité de liaison due à la
multivalence des dendrimères.
Les KI des dendrimères-PEG24-hBD2n étant la concentration nécessaire à déplacer 50%
de leur homologue fluorescent liés à CCR6, leur KI sera équivalent à leur constante d’affinité
(KD)
Un contrôle négatif sera réalisé sur les cellules T CD4+CCR6-.
Les concentrations en dendrimères utilisées pour les tests suivants seront déterminées
à partir des KD des dendrimères-PEG24-hBD2n.
III.3.4. Tests d’activité
Dans le but de vérifier que le dendrimères-PEG24-hBD2n est capable d’activer les
protéines Gi associées aux récepteurs CCR6 qui vont d'inhiber l'AMP cyclase, des
expériences de dosage de l'AMP cyclique seront réalisées grâce au cAMP enzyme
immunoassay kit direct de Sigma. Ces tests seront réalisés sur les cellules T CD4+CCR6+.
L'échantillon prétraité avec les dendrimères-PEG24-hBD2n ou hBD2 seule à différentes
concentrations sera traité à l'acide chloridrique afin de stopper l'hydrolyse de l'ATP en AMPc
par l'AMP cyclase. Les cellules seront ensuite lysées et mises en présence d'une concentration
connue en AMPc liée à la phosphatase alcaline (AMPc du kit) ainsi qu'un anticorps poly
clonal anti-AMPc. Cet anticorps va reconnaître sans distinction les molécules d'AMPc-
phosphatase alcaline et d'AMPc endogène, s'ensuit une compétition entre les deux molécules
d'AMPc pour l'anticorps. L'échantillon sera ensuite déposé sur une plaque où les anticorps
secondaires dirigés contre l'anticorps primaire seront fixés. Un lavage permettra d'éliminer
l'AMPc non lié aux anticorps primaires. Le pNpp (incolore), substrat de la phosphatase
alcaline sera ajouté puis le pNp, produit de la réaction de couleur jaune sera dosé.
L'intensité de la D.O à 405 nm sera inversement proportionnelle à la quantité d'AMPc
endogène présent initialement dans l'échantillon.
Des courbes de pourcentage d’inhibition en fonction de la concentration en
dendrimères-PEG24-hBD2n ou hBD2 seul pourront être alors obtenues. Le conjugué le plus
efficace présentera le taux d’inhibition le plus élevé. Un contrôle négatif sera aussi réalisé sur
les cellules T CD4+CCR6-.


! ! "'!
III.3.5. Infection des cellules par le VIH
Les souches des virus VIHIIIB (tropisme X4) et VIHBa-L (tropisme R5) seront achetées
à la société advanced biotechnology.
Les cellules T CD4+ CCR6+ et T CD4+ CCR6- seront infectées pendant 2 heures avec
100 DICT50 (dose de virus qui permet d'infecter 50 % des cellules) de VIHIIIB ou VIHBa-L.
Après infection, les cellules seront lavées avec du tampon phosphate salin (PBS) et du milieu
complet contenant le traitement approprié sera ajouté (voir paragraphe 3.6 et 3.7) (L. Sun et
al, 2005).
Le kit p24 Elisa (PerkinElmer Life and Analytical Sciences) permettra de suivre
l’infection des cellules, p24 étant une protéine de la nucléocapside virale.
III.3.6. Expression d’APOBEC3G et taux de Vif
L’activation des protéines Gi associées aux récepteurs CCR6 entraine une cascade de
réactions qui induit l’augmentation de l’expression de la protéine APOBEC3G. Les travaux
expérimentaux de A. Mehle et al ont montré que l’augmentation d’APOBEC3G entrainait
l’ubiquitinylation des protéines virales Vif ainsi que leur dégradation. De ce fait, dans une
même expérience, un Western Blot sera réalisé avec un anticorps anti APOBEC3G de lapin
(sigma) et un anticorps anti-Vif de souris (abcam).
Les expériences seront réalisées sur un lysat de cellules T CD4+ CCR6+ et T CD4+
CCR6- infectées par VIHIIIB ou VIHBa-L (voir paragraphe 3.5), traitées ou non à différentes
concentrations (voir paragraphe 3.3) de dendrimères-PEG24-hBD2n ou de la protéine hBD2
seule. Après lyse cellulaire, les protéines totales seront déposées sur le gel SDS-PAGE
(sodium dodecyl sulfate-polyacrylamide gel electrophoresis) en quantité équivalente
(méthode de Bradford). La révélation se fera grâce à des anticorps secondaires dirigés contre
l’anticorps primaire de lapin pour visualiser APOBEC3G et contre l’anticorps primaire de
souris pour visualiser Vif.
Une normalisation sera faite avec un anticorps anti-"-actine (sigma).
III.3.7. Test d’inhibition de la réplication virale
Les travaux de M.K.Lafferty et al. (2010) ont démontré que l’augmentation
d’APOBEC3G via l’activation du récepteur CCR6, induit une inhibition de la réplication
virale dans les cellules infectées par le VIH. Par les mêmes expériences, l’inhibition de la


! ! "(!
réplication virale par les dendrimères-PEG24-hBD2n en comparaison à hBD2 seule sera
quantifiée par qPCR (quantitative polymerisation chain reaction).
Les cellules T CD4+ CCR6+ et T CD4+ CCR6- infectées par VIHIIIB ou VIHBaL, (voir
paragraphe 3.5) seront traitées ou non par les dendrimères-PEG24-hBD2n ou hBD2 seule à
différentes concentrations (voir paragraphe 3.3). Après 4, 8 et 24h de traitement, l’ADN total
sera extrait par précipitation à l'éthanol (DNeasy Blood & Tissue Kit Qiagen) et les produits
de la reverse transcription seront quantifiés en utilisant l'iQSYBR Green supermix (Bio-Rad).
Un témoin sera réalisé en prétraitant les cellules à l’azidothymidine (AZT : médicament
antirétroviral bloquant l’infection des cellules). Les amorces utilisées lors de cette qPCR
correspondent aux régions LTR/RU5 (amorce sens 5#CTCTCTGGTTAGACCAGATCTG3#et
amorce anti-sens 5#ACTGCTAGAGATTTTCCACACTG3#). La normalisation se fera avec le
gène de l'albumine (amorce sens 5'TGTTGCATGAGAAAACGCCA3' et amorce anti-sens
5'GTCGCCTGTTCACCAAGGAT3') (SD. Mahajan et al., 2001 ; L. Sun et a.l, 2005 ; MK.
Lafferty et al., 2009).
Le pourcentage d’inhibition de la réplication virale pour chaque essai sera calculé par
rapport à la quantité d’ADN viral obtenue avec les cellules infectées non traitées.
III.3.8. Taux d’encapsidation d’APOBEC3G.
Afin de vérifier que l’augmentation de l’expression d’APOBEC3G induit une
augmentation de son encapsidation dans les nouveaux virions, le taux de protéines
APOBEC3G présentes dans les particules virales sera analysé par Western Blot.
Le surnageant des cultures de cellules T CD4+ CCR6+ et T CD4+ CCR6- infectées par
VIHIIIB ou VIHBa-L (voir paragraphe 3.5), prétraitées ou non avec différentes concentrations
(voir paragraphe 3.3) de dendrimères-PEG24-hBD2n ou hBD2 seule, est récupéré. Les
particules virales seront séparées des débris cellulaires par centrifugation et par filtration
(taille des pores 0,2 !m) (Milipore). Les particules virales seront ensuite concentrées par
centrifugation et le culot obtenu sera resuspendu dans un tampon de lyse. Le Western Blot
sera réalisé avec un anticorps anti APOBEC3G de lapin (sigma) sur le lysat comme décrit
dans le paragraphe 3.6.
Le taux d’encapsidation d’APOBEC3G sera comparé au taux d’APOBEC3G obtenu
avec les cellules infectées non traitées.


! ! ")!
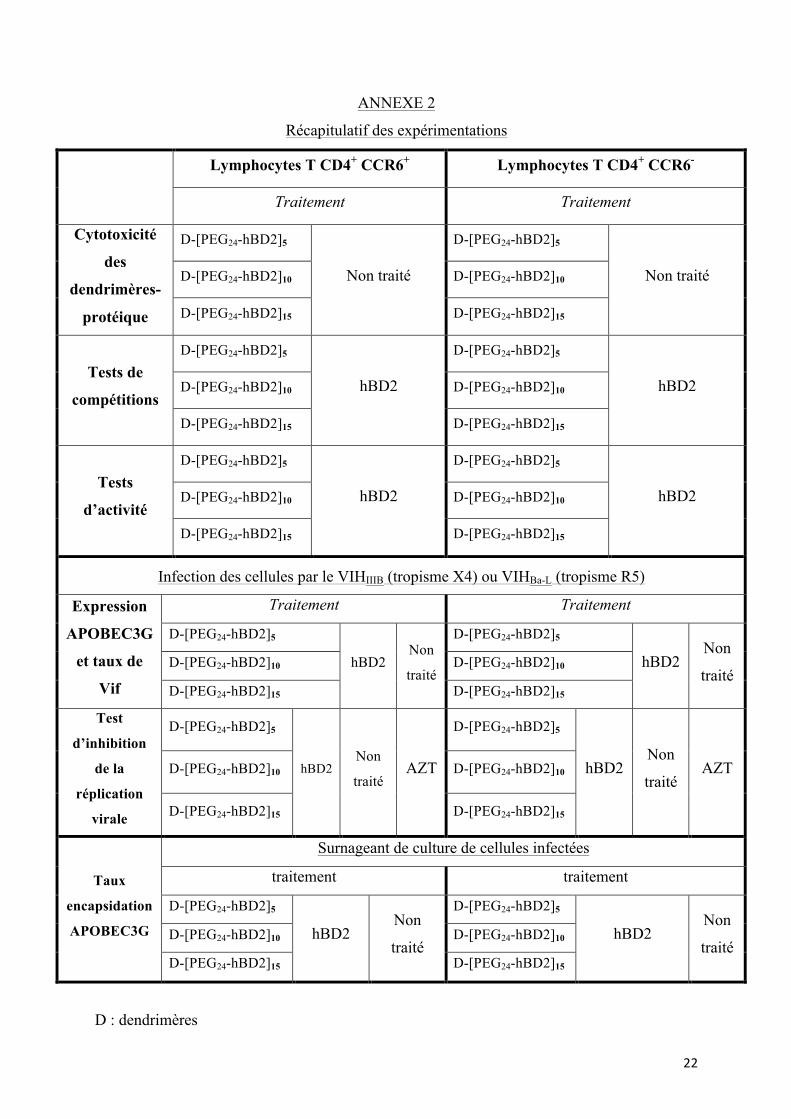
L'ensemble des expérimentations qui seront mises en œuvre pour le projet sont présentées
en annexe 2.
IV. DISCUSSION ET PERSPECTIVES
L’ensemble des expérimentations développées ci-dessus ont pour but de sélectionner
le(s) meilleur(s) candidat(s) pour l’inhibition de la réplication par VIHIIIB ou VIHBa-L. Ce
criblage permettra de retenir le(s) conjugué(s) activant au mieux la voie hBD2-CCR6-
APOBEC3G tout en présentant une faible cytotoxicité. Il est néanmoins possible que la
conception de certains conjugués pose problème à l’avancé de l’étude et demande une
révision de leur structure. La nécessité de réévaluer la structure des conjugués est notamment
illustré par les travaux de E.J. Baker sur le méthotrexate.
Afin de compléter l’étude, il est envisageable d’étudier l’effet préventif des
dendrimère-PEG24-hBD2 sur l’expression d’APOBEC3G, le taux de Vif, et la réplication
virale. Pour cela les cellules T CD4+CCR6+ pourront être prétraitées avec les dendrimères-
PEG24-hBD2 avant d’être infectées par le VIH.
Si l’étude révèle un ou plusieurs conjugués efficaces, des tests in vivo pourront être
envisagés, notamment chez le singe, afin de valider les résultats obtenus in vitro. De plus, les
expériences in vivo permettront d’étudier l’élimination des conjugués et de déceler leurs
éventuels effets indésirables sur l’organisme. Des résultats concluants pourraient mener à une
étude clinique chez l’Homme.
Le succès de cette étude serait un élément positif qui participerait à l’application des
dendrimères protéiques à d’autres contextes thérapeutiques.


! ! "*!
V. REFERENCES
Acosta EJ, Sergio O. Gonzalez, Eric E. Simanek (2004) Synthesis, characterization, and application of melamine-based dendrimers supported on silica gel. Journal of Polymer Science Part A: Polymer Chemistry 43: 168–177
Astruc D, Boisselier E, Ornelas C (2010) Dendrimers designed for functions: from physical, photophysical, and supramolecular properties to applications in sensing, catalysis, molecular electronics, photonics, and nanomedicine. Chem Rev.110: 1857-959
Bioconjugates Technique, 2ème Edition, GT. Hermanson
Bishop KN, Verma M, Kim EY, Wolinsky SM, Malim MH (2012) APOBEC3G inhibits elongation of HIV-1 reverse transcripts. PLoS Pathog 4: 1000231
Ellen M.M. de Brabander-van den Berg, E. W Meijer (1993) Poly(propylene imine) dendrimers: Large-Scale synthesis by hetereogeneously catalyzed hydrogenations. Angewandte Chemie, International Edition in English 32: 1308–1311
Esté JA, Cihlar T (2010) Current status and challenges of antiretroviral research and therapy. Antiviral Research 85: 25-33
Evans VA, Khoury G, Saleh S, Cameron PU, Lewin SR (2012) HIV persistence: chemokines and their signalling pathways. Cytokine Growth Factor Rev. 23: 151-7
Goodenow MM, Collman RG (2006) HIV-1 coreceptor preference is distinct from target cell tropism: a dual-parameter nomenclature to define viral phenotypes. J Leukoc Biol. 80: 965-72
Gosselin A, Monteiro P, Chomont N, Diaz-Griffero F, Said EA, Fonseca S, Wacleche V, El-Far M, Boulassel MR, Routy JP, Sekaly RP, Ancuta P (2010) Peripheral blood CCR4+CCR6+ and CXCR3+CCR6+CD4+ T cells are highly permissive to HIV-1 infection. The Journal of immunology 184: 1604-16
Lafferty MK, Sun L, DeMasi L, Lu W, Garzino-Demo A (2010) CCR6 ligands inhibit HIV by inducing APOBEC3G. Blood 115:1564-71
Majoros IJ, Myc A, Thomas T, Mehta CB, Baker JR Jr. (2006) PAMAM dendrimer-based multifunctional conjugate for cancer therapy: synthesis, characterization, and functionality. Biomacromolecules 7: 572-9
Malim MH (2009) APOBEC proteins and intrinsic resistance to HIV-1 infection. Philosophical Transactions of the Royal Society B: Biological Sciences 364: 675-87
Mehle A, Strack B, Ancuta P, Zhang C, McPike M, Gabuzda D (2003) Vif overcomes the innate antiviral activity of APOBEC3G by promoting its degradation in the ubiquitin-proteasome pathway. The Journal of Biological Chemistry 279: 7792-8
Nanaware-Kharade N, Gonzalez GA 3rd, Lay JO Jr, Hendrickson HP, Peterson EC (2012) Therapeutic anti-methamphetamine antibody fragment-nanoparticle conjugates: synthesis and in vitro characterization. Bioconjugate Chemistry 23: 1864-72


! ! #+!
Newman EN, Holmes RK, Craig HM, Klein KC, Lingappa JR, Malim MH, Sheehy AM (2005) Antiviral function of APOBEC3G can be dissociated from cytidine deaminase activity. Curr Biol. 15: 166-70.
Patri AK, Majoros IJ, Baker JR (2002) Dendritic polymer macromolecular carriers for drug delivery. Current Opinion in Chemical Biology 6: 466-471
Peterson J, Arkadi Ebber, Veiko Allikmaa, Margus Lopp (2001) Synthesis and CZE analysis of PAMAM dendrimers with an ethylenediamine core. Proceedings of the Estonian Academy of Sciences, Chemistry 50:156-166
Pluta K, Kacprzak MM (2009) Use of HIV as a gene transfer vector. Acta Biochim Pol 56: 531-95
Qi R, Gao Y, Tang Y, He RR, Liu TL, He Y, Sun S, Li BY, Li YB, Liu G (2009) PEG-conjugated PAMAM dendrimers mediate efficient intramuscular gene expression. The APPS Journal 11: 395-405
Röhrl J, Yang D, Oppenheim JJ, Hehlgans T(2012) Specific binding and chemotactic activity of mBD4 and its functional orthologue hBD2 to CCR6-expressing cells. J Biol Chem. 285: 7028-34
Schröder JM, Harder J (1999) Human beta-defensin-2. Int J Biochem Cell Biol. 3: 645-51
Sun L, Finnegan CM, Kish-Catalone T, Blumenthal R, Garzino-Demo P, La Terra Maggiore GM, Berrone S, Kleinman C, Wu Z, Abdelwahab S, Lu W, Garzino-Demo A (2005) Human beta-defensins suppress human immunodeficiency virus infection: potential role in mucosal protection. Journal of Virology 79: 14318-29
Thomas TP, Huang B, Choi SK, Silpe JE, Kotlyar A, Desai AM, Zong H, Gam J, Joice M, Baker JR Jr. (2012) Polyvalent dendrimer-methotrexate as a folate receptor-targeted cancer therapeutic. Molecular Pharmaceutics 9: 2669-2676
Wang F, Cai X, Su Y, Hu J, Wu Q, Zhang H, Xiao J, Cheng Y (2012) Reducing cytotoxicity while improving anti-cancer drug loading capacity of polypropylenimine dendrimers by surface acetylation. Acta Biomaterialia 8: 4304-4313
Weinberg A, Quiñones-Mateu ME, Lederman MM (2006) Role of human beta-defensins in HIV infection. Advances in Dental Research 19: 42-8
Zong H, Thomas TP, Lee KH, Desai AM, Li MH, Kotlyar A, Zhang Y, Leroueil PR, Gam JJ, Banaszak Holl MM, Baker JR Jr (2012) Bifunctional PAMAM dendrimer conjugates of folic acid and methotrexate with defined ratio. Biomacromolecules 13: 982-91


! ! #"!
VI. ANNEXES
ANNEXE1
Récapitulatif des dendrimères synthétisés
Dendrimères poly amido amine ou PAMAM
• Génération 4
G4-PAMAM-[PEG24-hBD2] n
G4-PAMAM-[PEG24-hBD2] n-[Rhodamine]5
• Génération 5
G5-PAMAM-[PEG24-hBD2] n
G5-PAMAM-[PEG24-hBD2] n-[Rhodamine]5
Dendrimères Poly propylène imine ou PPI
• Génération 4
G4-PPI-[PEG24-hBD2] n
G4-PPI-[PEG24-hBD2] n-[Rhodamine] 5
• Génération 5
G5-PPI-[PEG24-hBD2] n
G5-PPI-[PEG24-hBD2] n-[Rhodamine]5
Les dendrimères poly mélamines
• Génération 4
G4-mélamine-[PEG24-hBD2] n
G4-mélamine-[PEG24-hBD2] n-[Rhodamine] 5
• Génération 5
G5-mélamine-[PEG24-hBD2] n
G5-mélamine-[PEG24-hBD2] n-[Rhodamine] 5

! ! ##!
ANNEXE 2
Récapitulatif des expérimentations
Lymphocytes T CD4+ CCR6+ Lymphocytes T CD4+ CCR6-
Traitement Traitement
Cytotoxicité
des
dendrimères-
protéique
D-[PEG24-hBD2]5
Non traité
D-[PEG24-hBD2]5
Non traité D-[PEG24-hBD2]10 D-[PEG24-hBD2]10
D-[PEG24-hBD2]15 D-[PEG24-hBD2]15
Tests de
compétitions
D-[PEG24-hBD2]5
hBD2
D-[PEG24-hBD2]5
hBD2 D-[PEG24-hBD2]10 D-[PEG24-hBD2]10
D-[PEG24-hBD2]15 D-[PEG24-hBD2]15
Tests
d’activité
D-[PEG24-hBD2]5
hBD2
D-[PEG24-hBD2]5
hBD2 D-[PEG24-hBD2]10 D-[PEG24-hBD2]10
D-[PEG24-hBD2]15 D-[PEG24-hBD2]15
Infection des cellules par le VIHIIIB (tropisme X4) ou VIHBa-L (tropisme R5)!
Expression
APOBEC3G
et taux de
Vif
Traitement Traitement
D-[PEG24-hBD2]5
hBD2 Non
traité
D-[PEG24-hBD2]5
hBD2 Non
traité D-[PEG24-hBD2]10 D-[PEG24-hBD2]10
D-[PEG24-hBD2]15 D-[PEG24-hBD2]15
Test
d’inhibition
de la
réplication
virale
D-[PEG24-hBD2]5
hBD2 Non
traité AZT
D-[PEG24-hBD2]5
hBD2 Non
traité AZT D-[PEG24-hBD2]10 D-[PEG24-hBD2]10
D-[PEG24-hBD2]15 D-[PEG24-hBD2]15
Taux
encapsidation
APOBEC3G
Surnageant de culture de cellules infectées
traitement traitement
D-[PEG24-hBD2]5
hBD2 Non
traité
D-[PEG24-hBD2]5
hBD2 Non
traité D-[PEG24-hBD2]10 D-[PEG24-hBD2]10
D-[PEG24-hBD2]15 D-[PEG24-hBD2]15
D : dendrimères

Top Related