
TRƢỜNG ĐẠI H C KHOA H C T NHIÊN Cao Lệ Quyên NGHIÊN CỨ … · OsDREB1A/2A mã hóa...
27
ĐẠI HỌC QUỐC GIA HÀ NỘI TRƢỜNG ĐẠI HỌC KHOA HỌC TỰ NHIÊN ----------------------- Cao Lệ Quyên NGHIÊN CỨU PHÂN LẬP VÀ CHUYỂN GEN LIÊN QUAN ĐẾN TÍNH CHỊU HẠN VÀO GIỐNG LÚA Ở VIỆT NAM Chuyên ngành: Di truyền học Mã số: 62 42 01 21 DỰ THẢO TÓM TẮT LUẬN ÁN TIẾN SĨ SINH HỌC Hà Nội - 2016
Transcript of TRƢỜNG ĐẠI H C KHOA H C T NHIÊN Cao Lệ Quyên NGHIÊN CỨ … · OsDREB1A/2A mã hóa...

ĐẠI HỌC QUỐC GIA HÀ NỘI
TRƢỜNG ĐẠI HỌC KHOA HỌC TỰ NHIÊN
-----------------------
Cao Lệ Quyên
NGHIÊN CỨU PHÂN LẬP VÀ CHUYỂN GEN LIÊN QUAN
ĐẾN TÍNH CHỊU HẠN VÀO GIỐNG LÚA Ở VIỆT NAM
Chuyên ngành: Di truyền học
Mã số: 62 42 01 21
DỰ THẢO TÓM TẮT LUẬN ÁN TIẾN SĨ SINH HỌC
Hà Nội - 2016

Công trình đƣợc hoàn thành tại:
Trƣờng Đại học Khoa học Tự nhiên, Đại học Quốc gia Hà Nội.
Ngƣời hƣớng dẫn khoa học:
1. PGS.TS. Phạm Xuân Hội
2. PGS.TS. Đinh Đoàn Long
Phản biện 1:
Phản biện 2:
Phản biện 3:
Luận án đƣợc bảo vệ tại Hội đồng chấm luận án cấp Đại học
Quốc gia họp tại:
Trƣờng Đại học Khoa học Tự nhiên – Đại học Quốc gia Hà Nội
vào hồi…giờ 00, ngày…tháng…năm…
Có thể tìm hiểu luận án tại:
- Thƣ viện Quốc gia Việt Nam
- Trung tâm thông tin - Thƣ viện, Đại học Quốc gia Hà Nội

Đặt vấn đề
Gần đây, do ảnh hƣởng của hiện tƣợng biến đổi khí hậu toàn
cầu, hạn hán xảy ra ngày càng thƣờng xuyên hơn, với mức độ ngày
càng trầm trọng, gây tác động lớn tới tổng sản lƣợng lƣơng thực quốc
gia, ảnh hƣởng tới công tác xuất khẩu lúa gạo, đe doạ trực tiếp tới an
ninh lƣơng thực trong khu vực và trên thế giới.Trong điều kiện khí
hậu toàn cầu biến đổi mạnh mẽ, việc tạo ra các giống cây trồng có
năng suất cao, đồng thời chống/chịu đƣợc với các điều kiện bất lợi
của môi trƣờng trở thành một trong những nhiệm vụ then chốt của
khoa học nông nghiệp. Tuy nhiên, cho đến này tất cả những giống
cây trồng chống chịu điều kiện môi trƣờng bất lợi đƣợc sử dụng
trong sản xuất đều đƣợc lai tạo dựa trên các phƣơng pháp chọn giống
truyền thống, hầu nhƣ chƣa có giống cây trồng chịu hạn nào đƣợc tạo
ra b ng phƣơng pháp công nghệ sinh học. Sang thế k I, với sự
phát triển b ng nổ của sinh học phân tử và công nghệ tế bào, hai định
hƣớng chọn giống chịu hạn đang đƣợc các nhà khoa học đặc biệt
quan tâm là chọn giống phân tử và chọn giống chuyển gen.
Tính trạng chịu hạn là tính trạng đa gen (với sự tham gia của
hàng trăm gen khác nhau), vì vậy định hƣớng chọn giống phân tử
đang gặp khó khăn lớn trong việc quy tụ các tính trạng, gen quan
trọng liên quan đến chịu hạn. Gần đây, dựa trên những thành tựu
nghiên cứu về chức năng hệ gen thực vật, c ng với sự phát triển các
kỹ thuật microaray, proteomic..., các nhà khoa học đã phát hiện và
chứng minh vai trò quan trọng của nhóm gen điều khiển trong việc
tăng cƣờng tính chịu hạn ở thực vật. Nhóm gen điều khiển mặc d
không tham gia trực tiếp vào quá trình đáp ứng với điều kiện hạn của
thực vật nhƣng sự biểu hiện của chúng lại có vai trò điều hòa biểu
hiện của rất nhiều gen chức năng khác tham gia vào quá trình đáp

ứng hạn, dẫn tới làm tăng cƣờng khả năng chịu hạn ở thực vật. Phát
hiện này đã mở ra một hƣớng nghiên cứu rất mới cho lĩnh vực chọn
giống chuyển gen ở thực vật, đó là ch cần chuyển một hay một vài
gen điều khiển thay vì vài trăm gen chức năng vào cây để tăng cƣờng
tính chống chịu của cây trồng. Chính vì lí do này mà các nghiên cứu
phân lập, đặc tính hoá các gen điều khiển liên quan đến tính chịu hạn
đang trở thành định hƣớng nghiên cứu đầy tiềm năng trong việc chọn
tạo giống chịu hạn.
Ở Việt Nam, trong khoảng 10 năm trở lại đây, các nghiên
cứu về phân lập gen chịu hạn và tạo giống cây trồng chống chịu hạn
b ng công nghệ chuyển gen thực vật đã bắt đầu đƣợc một số phòng
thí nghiệm quan tâm. Tuy nhiên, hầu hết các chƣơng trình, dự án
nghiên cứu về gen chống chịu stress môi trƣờng nói chung và chống
chịu hạn nói riêng đều sử dụng các nguồn gen từ nƣớc ngoài hoặc
các gen đã đƣợc công bố bởi các nhóm nghiên cứu khác trên thế giới.
Cho đến nay, chúng ta vẫn chƣa có một đề tài nghiên cứu cơ bản
hoàn ch nh nào về phân lập gen liên quan đến đáp ứng chống chịu
hạn ở thực vật. Từ đó, chúng tôi đề xuất và tiến hành thực hiện đề tài:
“Nghiên cứu phân lập và chuyển gen liên quan đến tính chịu hạn
vào giống lúa ở Việt Nam” với mục đích để tạo nguồn vật liệu, số
liệu quan trọng trong việc tạo ra các giống cây trồng chuyển gen có
khả năng chống chịu điều kiện hạn.
Mục tiêu nghiên cứu
i) Phân lập đƣợc gen mã hóa nhân tố phiên mã s R 1 /2
liên quan tính chịu hạn trên giống lúa Việt Nam.
ii) Tối ƣu hoá đƣợc quy trình biến nạp gen thông qua vi khuẩn
Agrobacterium tumefaciens trên một số giống lúa Việt Nam.

iii) Tạo ra đƣợc một số dòng lúa chuyển gen mã hóa nhân tố
phiên mã s R 1 /2 có kiểu hình tƣơng đồng cây đối chứng
(cây không chuyển gen) trong điều kiện bình thƣờng và có khả năng
chống/chịu (tiếp tục sinh trƣởng, cho thu hạt) khi bị xử l hạn trong
điều kiện phòng thí nghiệm.
Đối tƣợng v nội ung nghiên cứu
Đối tượng nghiên cứu của luận án là các gen
OsDREB1A/2A mã hóa nhân tố điều hòa phiên mã liên quan tới tính
chịu hạn của giống lúa Việt Nam và vai trò, biểu hiện của các gen mã
hóa nhân tố phiên mã này trong các dòng chuyển gen dƣới sự điều
khiển của các promoter liên tục hoặc cảm ứng.
ác n i ung nghiên cứu chính
Nội dung 1: Phân lập gen hai gen OsDREB1A và OsDREB2A
liên quan đến tính chịu hạn từ thƣ viện c N của giống lúa Việt
Nam; và thiết kế các cấu trúc chuyển gen thực vật (binary vector) cho
gen OsDREB1A/OsDREB2A dƣới sự điều khiển của promoter liên
tục (Ubiquitine)/cảm ứng (Lip9).
Nội dung 2: Tối ƣu hóa quá trình phát sinh callus và tái sinh
cây từ callus của một số giống lúa Việt Nam; và hoàn thiện kỹ thuật
biến nạp gen thông qua vi khuẩn Agrobacterium tumefaciens trên
một số giống lúa Việt Nam có tiềm năng phát sinh callus và khả năng
tái sinh cao.
Nội dung 3: iến nạp các cấu trúc biểu hiện nhân tố phiên
mã vào giống lúa Việt Nam và sàng lọc các dòng cây chuyển gen
(kiểu hình- so sánh với cây đối chứng, kiểu gen- xác định sự tồn tại
ổn định và số lƣợng cấu trúc biến nạp trong cây chuyển gen).

Nội dung 4: Đánh giá biểu hiện (kiểu hình và kiểu gen) của
các dòng chuyển gen (T3, một cấu trúc biến nạp) trong điều kiện xử
l hạn tại phòng thí nghiệm.
Địa điểm nghiên cứu
Luận án đƣợc thực hiện tại 2 địa điểm chính: ộ môn ệnh
học Phân tử, Viện i truyền Nông nghiệp (Viện hoa học Nông
nghiệp Việt Nam) là địa ch uy tín, dẫn đầu trong cả nƣớc về công tác
phân lập, nghiên cứu chức năng của các gen mã hóa nhân tố phiên
mã; và Phòng Thí nghiệm tƣơng tác cây trồng-vi sinh vật thuộc
Trung tâm nghiên cứu vì sự phát triển (IR ) tại ontpellier là phòng
thí nghiệm có trang bị hiện đại, có các chuyên gia uy tín trong nghiên
cứu chức năng, biểu hiện của gen/bộ gen thực vật (đặc biệt là lúa).
Đóng góp mới của luận án
Luận án là công trình đầu tiên ở Việt Nam thực hiện một
cách có hệ thống về nghiên cứu cơ bản theo định hƣớng ứng dụng
trên hai gen mã hóa nhân tố phiên mã OsDREB1A và OsDREB2A
liên quan tính chịu hạn. Trong nghiên cứu này, trình tự mã hóa
OsDREB1A/OsDREB2A dƣới sự điều khiển bởi promoter hoạt động
liên tục (Ubiquitin) hoặc promoter cảm ứng stress (Lip9) đã đƣợc
nghiên cứu biểu hiện và ảnh hƣởng đến cây lúachuyển gen trong cả
điều kiện thông thƣờng và điều kiện bất lợi. Luậnánđã chọn đƣợc 04
dòng lúa Chành trụi chuyển gen (OsDREB1A) kiểu hình bình thƣờng
và có khả năng tăng cƣờng chịu hạn (đã duy trì đến T3).
Ngoài ra, trong nghiên cứu này, lần đầu tiên ở Việt Nam
chúng tôi tiến hành khảo sát chi tiết khả năng phát sinh callus và khả
năng tái sinh cây hoàn ch nh từ callus của nhiều giống lúa Việt Nam
(giống phổ biến, giống chịu hạn, giống địa phƣơng...). ết quả
nghiên cứu đã ch ra một số giống lúa có tiềm năng để tiếp tục tối ƣu

quy trình chuyển gen cho từng giống lúa sau này. Đây là tiền đề cho
các nghiên cứu tiếp theo nh m tạo ra các giống lúa công nghệ sinh
học b ng công nghệ ch nh sửa hệ gen không qua lai tạo có các đặc
điểm nông sinh học vƣợt trội mà vẫn giữ các phẩm chất của giống
gốc ban đầu.
Luận án lần đầu tiên ở Việt Nam đã thành công trong việc
ứng dụng phản ứng PCR định lƣợng trong việc ƣớc tính số lƣợng bản
sao của cấu trúc chuyển gen và xác định các dòng chuyển gen đồng
hợp ngay từ thế hệ T1. Với kết quả từ nghiên cứu này, chúng tôi tin
r ng đây có thể là phƣơng pháp ph hợp cho điều kiện phòng thí
nghiệm còn nhiều hạn chế trong nƣớc khi tiến hành nghiên cứu, sàng
lọc các dòng cây chuyển gen.
Ứng ụng thực tiễn của luận án
ết quả nghiên cứu của luận án đã tạo ra đƣợc các dòng T2
(đồng hợp, một bản sao gen OsDREB1A dƣới sự điều khiển của
promoter cảm ứng Lip9) có kiểu hình tƣơng đồng cây đối chứng
không chuyển gen và đặc biệt có khả năng phục hồi và kết hạt sau 3
tuần ngừng tƣới nƣớc. Đây là kết quả quan trọng, chứng tỏ tiềm năng
ứng dụng của OsDREB1A và điều hòa biểu hiện của gen mã hóa
nhân tố phiên mã dƣới sự điều khiển của promoter cảm ứng trong
công tác tạo giống cây chuyển gen chịu hạn. Trên cơ sở kết quả của
luận án, các vector biểu hiện gen OsDREB1A và OsDREB2A do luận
án tạo ra đang đƣợc nghiên cứu chuyển vào các giống cây trồng quan
trọng nhƣ ngô, đậu tƣơng, bông… để chọn tạo các giống cây chuyển
gen chịu hạn nh m đáp ứng nhu cầu cấp thiết của sản xuất trong điều
kiện thay đổi khí hậu toàn cầu. Các dòng chuyển gen đƣợc duy trì
theo dõi trong nhà lƣới để làm nguyên liệu cho công tác lai tạo giống

khi điều kiện thực tế cho phép (chấp nhận cây lúa chuyển gen và vật
liệu di truyền do sự kiện chuyển gen tạo ra).
1. Bố cục của luận án
Luận án gồm 137 trang, bao gồm: Phần mở đầu (06 trang); Tổng
quan tài liệu (32 trang); Nguyên liệu và phƣơng pháp nghiên cứu (18
trang); Kết quả nghiên cứu và thảo luận (36 trang); Kết luận (02
trang); Kiến nghị (01 trang); Các công trình khoa học của tác giả liên
quan đến luận án (01 trang); Tài liệu tham khảo (12 trang) với 135 tài
liệu gồm 2 thứ tiếng: tiếng Việt (12 tài liệu) và tiếng Anh (123 tài
liệu); Phụ lục (14 trang). Luận án có 18 bảng, 24 hình.

Chƣơng 1. TỔNG QUAN TÀI LIỆU
1.1. ẢNH HƢỞNG CỦA HẠN HÁN ĐẾN NÔNG NGHIỆP &
THỰC VẬT
Hạn đối với thực vật là khái niệm đƣợc d ng để ch trạng thái thiếu nƣớc
do môi trƣờng gây nên trong suốt quá trình sống hay trong từng giai đoạn
sống, làm ảnh hƣởng đến quá trình sinh trƣởng và phát triển của thực vật.
ôi trƣờng khô hạn gây ra hàng loạt những tác động tiêu cực tới thực vật ở
tất cả các cấp độ, từ hình thái tới phân tử và ở tất cả các giai đoạn phát triển.
Để đối phó với điều kiện hạn hán, thực vật sẽ khởi động cơ chế phòng vệ
chống lại sự thiếu hụt nƣớc, sau đó sẽ là một loạt các cơ chế ở các cấp độ
khác nhau. Đáp ứng chống chịu stress hạn của thực vật đƣợc thực hiện
thông qua một chuỗi các quá trình rất phức tạp với sự tham gia của hàng
loạt các yếu tố và có thể chia thành 3 giai đoạn: nhận tín hiệu – dẫn truyền
tín hiệu – đáp ứng chống chịu khởi đầu chuỗi truyền tín hiệu.
1.2. PHẢN ỨNG CỦA THỰC VẬT TR NG ĐIỀU KIỆN HẠN
Thực vật thích nghi với môi trƣờng sống nhờ khả năng điều hòa
nhanh biểu hiện của các gen chức năng. Quá trình điều hòa biểu hiện
gen đóng một vai trò quan trọng trong đáp ứng của thực vật với các
điều kiện stress phi sinh học, trong đó có hạn hán. Các nghiên cứu
gần đây đã chứng minh stress hạn tác động tới tất cả các bƣớc trong
quá trình điều hòa hoạt động gen của thực vật, bao gồm: điều hòa quá
trình phiên mã, điều hòa sau phiên mã, điều hòa quá trình dịch mã,
điều hòa sau dịch mã, điều hòa hoạt động gen thông qua các phân tử
RNA không mã hóa và yếu tố di truyền ngoại sinh (ví dụ nhƣ quá
trình methyl hóa DNA hay quá trình biến đổi histone).

1.3. VAI TRÒ CỦA NHÂN TỐ PHIÊN Ã TR NG ĐÁP ỨNG
STRESS HẠN
Trong hệ gen thực vật có khoảng 7% gen mã hóa nhân tố
phiên mã (TF), trong số đó có rất nhiều gen liên quan đến đáp ứng
chống chịu stress. Các TF tham gia trong mạng lƣới điều hòa hoạt
động gen đáp ứng stress hạn đã biết cho tới nay đƣợc xác định
thuộc hầu hết các nhóm TF lớn của thực vật, trong đó đƣợc nghiên
cứu nhiều nhất là các nhóm AP2/ERF, NAC, WRKY, MYB/MYC,
bZIP hay ZF.
1.4. TÌNH HÌNH NGHIÊN CỨU TẠO GIỐNxG LÚA CHUYỂN
GEN CHỊU HẠN TRÊN THẾ GIỚI VÀ Ở VIỆT NAM
Việc phát triển các giống lúa chịu hạn và giảm lƣợng nƣớc tiêu
thụ trong sản xuất lúa gạo có nghĩa vô c ng to lớn để tăng sản
lƣợng và đảm bảo an ninh lƣơng thực. Một hƣớng nghiên cứu phổ
biến và rất đƣợc quan tâm hiện nay là tăng cƣờng biểu hiện các gen
đáp ứng hoặc liên quan đến đáp ứng chống chịu hạn trong cây lúa
chuyển gen. o cơ chế chống chịu hạn phức tạp của thực vật, việc
sử dụng các gen điều hòa (mã hóa các protein tham gia vào mạng
lƣới điều hòa đáp ứng stress) trong nghiên cứu chuyển gen tạo
giống lúa chịu hạn thƣờng mang lại hiệu quả cao hơn so với các gen
chức năng (mã hóa protein/enzyme trực tiếp tham gia vào đáp úng
chống chịu stress). Tuy nhiên, cho đến nay mới ch có một vài
nghiên cứu chứng minh khả năng chịu hạn của các dòng lúa chuyển
gen trong điều kiện thử nghiệm trên đồng ruộng. Ở Việt Nam, cho
đến nay chƣa có một nghiên cứu hoàn ch nh nào về các gen liên
quan đến đáp ứng chống chịu stress ở thực vật, đặc biệt là nhóm
gen mã hoá nhân tố phiên mã liên quan tới tăng cƣờng đáp ứng
chống chịu hạn ở lúa.

Chƣơng 2. VẬT LIỆU & PHƢƠNG PHÁP NGHIÊN CỨU
2.1. VẬT LIỆU
2.1.1. Đối tƣợng nghiên cứu
Thƣ viện c N sơ cấp của lúa Oryza sativaL.xử lý hạn và mặn
đƣợc cung cấp bởi Bộ môn Bệnh học phân tử, Viện Di truyền Nông
nghiệp. 47 giống lúa đƣợc cung cấp bởi Trung tâm Tài nguyên Di
truyền.
2.1.2. Chủng vi sinh vật
Chủng vi khuẩn Escherichia coli H5α đƣợc mua từ hãng
Fermentas (Mỹ); chủng vi khuẩn E. coli XLOR, XL1-Blue MRF và
phage “trợ giúp” đƣợc mua từ Công ty Agilent Technologies (Mỹ);
chủng vi khuẩn E. coli Rosetta (DE3) khả biến đƣợc mua từ Công ty
Merck Millipore (Mỹ); chủng vi khuẩn Agrobacterium tumefaciens
LBA4404 khả biến.
2.1.3. Vector và oligonucleotide
Các cặp mồi sử dụng làm mồi và các mẫu oligonucleotide đầu dò
đƣợc đặt mua từ hãng Invitrogen (Mỹ) và Sigma (Mỹ).
2.2. PHƢƠNG PHÁP NGHIÊN CỨU
- Xử lý mẫu thực vật theo phƣơng pháp của Qin (2007), Dubouzet (2003)
và Tran (2004).
- Tách chiết, định lƣợng DNA/RNA.
- Nhân dòng trình tự mã hóaOsDREB1A/OsDREB2Avào vectorpUC19.
- Thiết kế vector biểu hiện OsDREB1A/OsDEB2A.

Hình 2.2 | Giản đồ quy trình thiết kế vector chuyển gen
- Đánh giá khả năng tạo callus và tái sinh chồi từ callus của một số giống
lúaở Việt Nam
- Khảo sát một số quy trình chuyển gen cho lúa indica
- Tạo cây chuyển gen biểu hiện OsDREB1A/OsDREB2A.
- Sàng lọc kiểu gen, kiểu hình của các cây chuyển gen
- Đánh giá khả năng chịu hạn của cây chuyển gen ở giai đoạn sinh
trƣởng, ra hạt

Chƣơng 3. KẾT QUẢ & THẢO LUẬN
3.1. PHÂN LẬP THIẾT KẾ VECTOR CHUYỂN GEN
OSDREB1A/OSDREB2A
3.1.1. Phân lập trình tự mã hóa gen OsDREB1A/OsDREB2A
Trình tự mã hóa gen OsDREB1A đã đƣợc phân lập b ng phản
ứng PCR với cặp mồi gen OsDREB1A-Fw/Rv từ DNA hệ gen giống
Cƣờm Dạng và thƣ viện cDNA xử lý hạn của giống lúa Mộc tuyền.
Trình tự mã hóa s R 2 đã đƣợc phân lập b ng phản ứng PCR
với cặp mồi gen OsDREB2A-Fw/Rv từ khuôn cDNA xử lý hạn của
giống lúa Cƣờm Dạng. Sản phẩm khuếch đại trình tự mã hóa của hai
gen đã đƣợc nhân dòng vào vector pUC19. Vector tái tổ hợp đã đƣợc
biến nạp vào vi khuẩn E. coli chủng H5α, sàng lọc b ng phƣơng
pháp PCR trực tiếp khuẩn các lạc màu trắng(hình 3.5) và tiếp đó
đƣợc giải trình tự theo phƣơng pháp Sanger với cặp mồi vector .
Hình 3.5 | Kết quả PCR kiểm tra trực tiếp khuẩn lạc mang
vector pUC19 tái tổ hợp Ghi chú: ên trái- i h ợc biến n p vector tái tổ hợ
i giế g - 5: sản ph h h n l c biến n p sản
ph m ghép n i với o n chèn là sản ph m khuế h i trình tự mã hóa OsDREB1A t DNA
tổng s gi g ú ờm D ng; (ii) giếng 6-9: sản ph h h n l c biến n p sản ph m ghép n i với o n chèn là sản ph m khuế h i trình tự mã hóa OsDREB1A t
cDNA xử lý h n của gi ng lúa M c Tuyề giế g i chứng âm (không có DNA khuôn).
ên ph i- i h ợc biến n p vector tái tổ hợp pUC19/OsDREB2A: giế g i chứng âm (không có DNA khuôn); 12-15: sản ph h h n
l c biến n p sản ph m ghép n i giữa pUC19 và sản ph m khuế h i trình tự mã hóa
OsDREB2A t cDNA của gi g ú ờm D ng.

3.1.2. Thiết kế vector chuyển gen
Trình tự mã hoá OsDREB1A/OsDREB2A trong vector pUC19 và
vector chuyển gen pBIG - Lip9/Ubi đƣợc cắt và xử lý b ng enzyme
giới hạn BamHI, rồi đƣợc ghép nối b ng enzyme T4 ligase theo quy
trình giản lƣợc trong hình 2.2. Vector pBIG-Lip9/Ubi tái tổ hợp đƣợc
biến nạp vào vi khuẩn E. coli chủng DH5α, sử dụng phƣơng pháp
PCR trực tiếp khuẩn lạc để sàng lọc các thể tái tổ hợp trong đó trình
tự mã hóa n m cùng chiều với promoter điều khiển trong cấu trúc
chuyển gen (hình 3.10).
Hình 3.10 | Điện i sản phẩm PCR khuẩn lạc trên gel agarose 1% i- Sản ph m PCR t các khu n l c mang pBIG-Lip9-OsDREB1A. Giếng M: Thang chu n
DNA 1kb; giếng 1, 2: khu n l c 1 và 2; giế g 3 i chứng âm
B- Sản ph m PCR t các khu n l c mang pBIG-Lip9-OsDREB2A. Giếng M: Thang chu n DNA 1kb; giế g i chứng âm; giếng 2-5: khu n l c 1-4
3.2. NGHIÊN CỨU QUI TRÌNH CHUYỂN G N VÀ GIỐNG
LÚ Ở VIỆT N
3.2.1. Khả hình th nh callus của tập đo n giống lúa Việt Nam
Sử dụng 03 loại môi trƣờng MS (nồng độ 2,4D từ 1,5 mg/l đến
2,5 mg/l) để khảo sát khả năng tạo callus của 47 giống lúa ở Việt
Nam chúng tôi nhận thấy có 26 giống có khả năng tạo callus tốt.
Trong đó, có 05 giống bao gồm giống 20, 26, 34, 44 và 47 có tiềm
năng tạo callus cao nhất. Cụ thể là với giống 20 môi trƣờng có bổ
sung 1,5mg/l 2,4 (môi trƣờng S1) cho tỷ lệ tạo callus cao nhất

(78,49%) còn môi trƣờng có bổ sung 2mg/l 2,4 ( môi trƣờng S2)
là thích hợp nhất cho sự hình thành callus của giống lúa 26, 34, 44 và
47 (tƣơng ứng 84,94, 76,53%, 79,61% và 90,06%).
3.2.2. Khả năng tái sinh của tập đo n giống lúa Việt Nam
26 giống lúa có khả năng tạo callus tốt trong mục 3.2.1 đã đƣợc
nghiên cứu đánh giá khả năng tái sinh chồi từ callus trên 04 loại môi
trƣờng trên nền MS có bổ sung BAP và kinetin. Kết quả khảo sát cho
thấy, có 05 giống bao gồm YUNLU103-1 , giống LUYIN46, LP-
5M, IR80416-B-152-4 và Chành trụi có khả năng tạo callus tốt nhất
đƣợc lựa chọn tiếp tục nghiên cứu đánh giá khả năng tiếp nhận gen
và hoàn thiện quy trình chuyển gen.
3.2.3. Khảo sát khả năng tiếp nhận gen của giống lúa Việt Nam
Bảng 3.5 | hả năng tiếp nhận gen của 5 giống lúa Việt Nam
Qui trình Ch nh trụi
YUNLU10
3-1B LUYIN46 LP-5M
IR80416-B-
152-4
Phòng
Bệnh học
phân tử
Chọn lọc 1 50 ± 2,04 20,33± 1,67 15,53± 1,78 19,39± 1,57 21,53± 1,98
Chọn lọc 2 38 ± 1,53 5,26± 1,01 2,34± 0,67 4,33± 0,51 6,21± 0,62
Tái sinh 27,67± 1,62 0 0 0 0
PCR 17,33± 1,12
Rahman
(2011)
Chọn lọc 1 55,05± 1,94 20,13± 1,94 10,53± 1,94 17,32± 1,94 11,06± 1,94
Chọn lọc 2 15,12± 1,94 7,05± 1,94 1,12± 1,94 3,54± 1,94 4,57± 1,94
Tái sinh 0 0 0 0
PCR
Hiei
(2008)
Chọn lọc 1 40 20,33 15,53 19,39 9,34
Chọn lọc 2 15,24 5,19 4,31 8,03 3,47
Tái sinh 0 0 0 0 0
PCR 0
05 giống có khả năng tạo callus và tái sinh tạo chồi tốt nhất đƣợc

nghiên cứu đánh giá khả năng tiếp nhận gen thông qua vi khuẩn A.
tumefaciens với 03 quy trình khác nhau (Bảng 3.5) bao gồm: 01 quy
trình đã phát triển bởi chính tác giả và 01 quy trình cho lúa japonica
(Hiei, 2008) và 01 quy trình cho lúa india (Rahman, 2011). ết quả
nghiên cứu cho thấy ch có giống Chành Trụi có khả năng tiếp nhận
gen lạ thông qua vi khuẩn A. tumefaciens sử dụng quy trình đã đƣợc
phát triển bởi chính nghiên cứu sinh (Hình 3.15).
Hình 3.15 | Quy trình chuyển gen Ch nh trụi nền môi trƣờng MS Q y ì h ợc thiết lập cho gi ng Chành trụi t i hó về thời gian t g b ớc, nồ g
các chất bổ sung, nhiệ . Chú ý nồ g vi khu . ef ie ã i 600 = 0,03,
sử dụng vi khu n ở nồ g o hơ ,5 h t s nghiên cứ ớ ó ó h gây giảm hiệu quả củ i ờng chọn lọ b ớc ra rễ (7) có th sử dụ g i ờng MS giảm m t nửa
nồ g (MS/2) giúp rễ xuất hiện sớm (rễ mảnh và phân chia nhiề hơ , y hiê ở b ớc tiếp
theo (8) cầ hú ý hi ây v o ù hè hó i o m và nhiệ ). Cây sau khi h ới 03 tuần sẽ ợc thu mẫu lá, tách DNA tổng s sàng lọc bằ g x ịnh
ây d ơ g í h MS: i ờng Murashige and Skoog.
3.3. TẠO GIỐNG LÚA CHÀNH TRỤI CHUYỂN GEN
Các cấu trúc p IG-Lip9:OsDREB1A, pBIG-Ubi:OsDREB1A,
pBIG-Lip9:OsDREB2A và pBIG-Ubi: s R 2 , các vector trống p IG-
Lip9 và pBIG-Ubi đã đƣợc biến nạp vào giống lúa Chành trụi theo quy trình
giản lƣợc trong hình 3, kết quả từ 600 callus ban đầu đã thu đƣợc 99 chồi tái
sinh. ết quả theo dõi cây chuyển gen và sử dụng phƣơng pháp PCR, qRT-

PCR với cả mồi đặc hiệu gen và mồi vector đã xác định đƣợc 44 dòng
chuyển gen một bản copy và trong đó ch có 10 dòng cho hạt ( ảng 3.7).
Bảng 3.1 | Kết quả kiểm tra các cây chuyển gen T0
Cấu trúc gen chuyển Tái
sinh
PCR ƣơng
tính Số bản copy qRT-PCR
1 bản copy > 1 bản copy
Lip9: OsDREB2A 15 12 5 7
Ubi: OsDREB2A 12 10 6 4
Lip9: OsDREB1A 23 19 13 6
Ubi: OsDREB1A 20 15 11 4
pBIG-Lip9 16 10 5 5
pBIG-Ubi 13 9 4 5
Tổng 99 75 44 31
Tiếp tục đánh giá cây chuyển gen T1 b ng kỹ thuật qRT-PCR, nảy mầm
trên Hygromycin đã lựa chọn đƣợc 9 dòng chuyển gen đồng hợp một bản
copy để tiếp tục theo dõi đánh giá khả năng chịu hạn trong các thế hệ tiếp
theo. Tuy nhiên trong số đó có 03 dòng là L4, U2 và U3 không cho hạt T2 nên
ch còn 06 dòng đƣợc tiếp tục nghiên cứu trong các thí nghiệm đánh giá kiểu
hình kế tiếp ( ảng 3.10).
Bảng 3.2 | Kết quả sinh trƣởng các òng chuyển gen đồng hợp
Cấu trúc gen Số cây Sinh trƣởng
bình thƣờng Trổ bông Kết hạt
Thu hạt
T2
Lip9:
OsDREB1A
L1 2 2 1 1 1
L2 2 2 2 1 1
L3 2 2 2 2 2
L4 1 0
L5 2 1 1 1 1
Ubi:
OsDREB1A
U1 3 2 2 2 2
U2 1 0

U3 2 1 1 0
U4 2 2 2 2 2
Tổng 17 12 11 9 9
3.4. ĐÁNH GIÁ IỂU HÌNH C Y CHUYỂN GEN
Các cây chuyển gen T2 đƣợc nghiên cứu đánh giá kiểu hình so
sánh với cây đối chứng trong cả điều kiện hạn và điều kiện hạn. Kết
quả so sánh đánh giá trong điều kiện bình thƣờng có 04 dòng chuyển
gen một bản copy có kiểu hình tƣơng đồng với cây đối chứng không
chuyển gen (Bảng 3.11).
Bảng 3.3 | Một số chỉ tiêu sinh trƣởng của cây chuyển gen và cây
đối chứng
Cây
Chỉ tiêu sinh trƣởng
Số nhánh/
khóm
Chiều cao
cây (cm)
Lƣợng hạt
chắc trên
bông
Thời gian sinh
trƣởng (ngày)
Cây đối chứng 5,02 ± 1,21 110,65 ± 3,51 110 ± 3,5 110 ± 3
L1 4,51 ± 1,09 90,12 ± 1,56 100 ± 1,2 105 ± 5
L2 4,49 ± 0,82 115,68 ± 2,02 100 ± 3,5 116 ± 4
L3 5,12 ± 0,89 105,79 ± 2,50 115 ± 2,5 107 ± 5
L5 5,09± 0,95 108,54 ± 1,59 116 ± 1,8 106 ± 4
U1 4,99 ± 0,32 105,12 ± 2,01 107± 3,5 106 ± 5
U4 5,16 ± 0,98 112,46 ±1,58 109± 2,5 108 ± 3
Khả năng giữ nƣớc là một trong những ch tiêu quan trọng để
đánh giá tính chịu hạn ở thực vật và khả năng này phụ thuộc vào các
đặc tính nhƣ: sự cuộn lá, đóng mở khí khổng, tham gia cơ chế giảm
áp suất trƣơng của lá, giảm lƣợng nƣớc bị mất của cây và hoạt hoá sự
biểu hiện hàng loạt các gen liên quan đến tính chịu hạn. Trong thí
nghiệm của chúng tôi, mặc d các gen điều khiển chịu hạn đã đƣợc
xác định sự tồn tại và có thể đã biểu hiện tốt trong các cây chuyển

gen. Tuy nhiên, nghiên cứu khả năng giữ nƣớc cho kết quả có sự
khác biệt rất ít giữa cây chuyển gen và cây đối chứng không chuyển
gen (Bảng 3.12).
Bảng 3.4 | Khả năng giữ nƣớc của các òng lúa chuyển gen
Độ hụt khối lƣợng tƣơng đối (%) Tỷ lệ sống sót
(%) Sau 1 giờ Sau 4 giờ Sau 7giờ
Cây đối chứng 10,00 ± 0,25 14,55 ± 0,45 30,55 ± 0,36 30,55 ± 0,36
L3 9,67 ± 0,36 14,01 ± 0,56 27,08 ± 0,52 80,08 ± 0,52
L5 8,67 ± 0,32 13,98 ± 0,49 26,57 ± 0,78 90,57 ± 0,78
U1 9,67 ± 0,23 14,01 ± 0, 32 28,45 ± 0,62 78,45 ± 0,62
U4 8,99 ± 0,36 13,95 ± 0,28 27,45 ± 0,54 83,45 ± 0,54
Hình 3.20 | Khả năng giữ nƣớc của các òng lúa chuyển gen
Ngƣợc lại về thí nghiệm đánh giá khả năng phục hồi cho thấy có
sự khác biệt lớn giữa cây chuyển gen và cây đối chứng. Các cây sau
thí nghiệm đánh giá khả năng giữ nƣớc đƣợc trồng trở lại trong điều
kiện bình thƣờng, khả năng phục hồi đƣợc ghi nhận ở ngày thứ 1.
Kết quả cho thấy các cây đối chứng phát triển rất yếu xuất hiện các lá
vàng và xoăn; các cây chuyển gen phát triển khỏe mạnh bình thƣờng
các lá xanh khỏe mạnh. Tỷ lệ sống sót giữa cây đối chứng và các
dòng chuyển gen có sự khác biệt rất lớn trong khi cây đối chứng ch
sống sót có 30,55% còn các dòng chuyển gen có tỷ lệ sống sót rất cao
từ 78,45 – 90,57%. Kết quả này cho thấy các cây chuyển gen
OsDREB1A có khả năng phục hồi rất tốt sau quá trình mất nƣớc
(Hình 3.20).
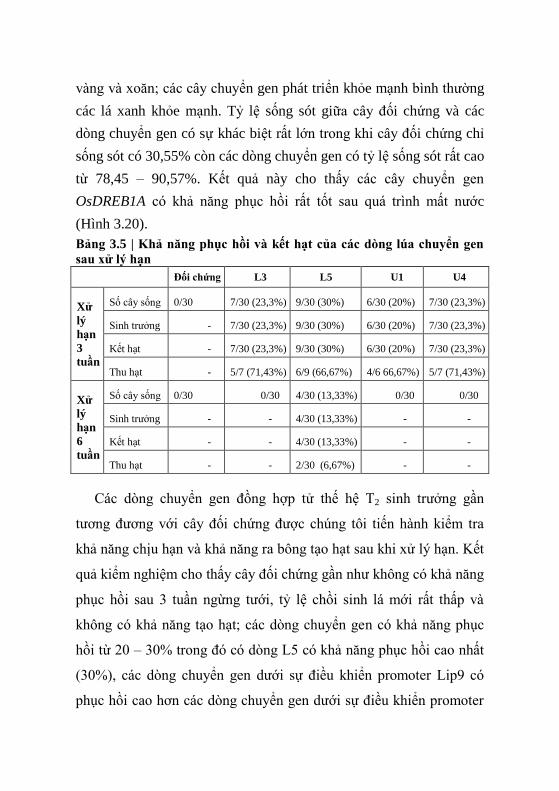
Bảng 3.5 | Khả năng phục hồi v kết hạt của các òng lúa chuyển gen
sau xử lý hạn
Đối chứng L3 L5 U1 U4
Xử
lý
hạn
3
tuần
Số cây sống 0/30 7/30 (23,3%) 9/30 (30%) 6/30 (20%) 7/30 (23,3%)
Sinh trƣởng - 7/30 (23,3%) 9/30 (30%) 6/30 (20%) 7/30 (23,3%)
Kết hạt - 7/30 (23,3%) 9/30 (30%) 6/30 (20%) 7/30 (23,3%)
Thu hạt - 5/7 (71,43%) 6/9 (66,67%) 4/6 66,67%) 5/7 (71,43%)
Xử
lý
hạn
6
tuần
Số cây sống 0/30 0/30 4/30 (13,33%) 0/30 0/30
Sinh trƣởng - - 4/30 (13,33%) - -
Kết hạt - - 4/30 (13,33%) - -
Thu hạt - - 2/30 (6,67%) - -
Các dòng chuyển gen đồng hợp tử thế hệ T2 sinh trƣởng gần
tƣơng đƣơng với cây đối chứng đƣợc chúng tôi tiến hành kiểm tra
khả năng chịu hạn và khả năng ra bông tạo hạt sau khi xử l hạn. ết
quả kiểm nghiệm cho thấy cây đối chứng gần nhƣ không có khả năng
phục hồi sau 3 tuần ngừng tƣới, tỷ lệ chồi sinh lá mới rất thấp và
không có khả năng tạo hạt; các dòng chuyển gen có khả năng phục
hồi từ 20 – 30% trong đó có dòng L5 có khả năng phục hồi cao nhất
(30%), các dòng chuyển gen dƣới sự điều khiển promoter Lip9 có
phục hồi cao hơn các dòng chuyển gen dƣới sự điều khiển promoter

Ubiquitin (Ngƣợc lại về thí nghiệm đánh giá khả năng phục hồi cho
thấy có sự khác biệt lớn giữa cây chuyển gen và cây đối chứng. Các
cây sau thí nghiệm đánh giá khả năng giữ nƣớc đƣợc trồng trở lại
trong điều kiện bình thƣờng, khả năng phục hồi đƣợc ghi nhận ở
ngày thứ 1. ết quả cho thấy các cây đối chứng phát triển rất yếu
xuất hiện các lá vàng và xoăn; các cây chuyển gen phát triển khỏe
mạnh bình thƣờng các lá xanh khỏe mạnh. Tỷ lệ sống sót giữa cây
đối chứng và các dòng chuyển gen có sự khác biệt rất lớn trong khi
cây đối chứng ch sống sót có 30,55% còn các dòng chuyển gen có tỷ
lệ sống sót rất cao từ 78,45 – 90,57%. ết quả này cho thấy các cây
chuyển gen OsDREB1A có khả năng phục hồi rất tốt sau quá trình
mất nƣớc (Hình 3.20).
ảng 3.5).
Sau khi đánh giá khả năng chịu hạn các dòng tiếp tục đƣợc trồng
ở điều kiện bình thƣờng trong nhà lƣới, kết quả cho thấy các dòng
chuyển gen có khả năng phục hồi tốt, sinh trƣởng gần nhƣ bình
thƣờng hiện tƣợng lá bị vàng và bị cháy rất ít, các dòng ra hoa kết
quả bình thƣờng. Tỷ lệ hạt chắc ch đạt từ 66,67% đến 71,43%, điều
này cho thấy hạn có khả năng ảnh hƣởng khá lớn đến năng suất của
cây trồng.

-
Hình 3.21 | Khả năng phục hồi, tạo hạt của các òng T2 chuyển gen khi
xử lý hạn 3 tuần
Ghi chú: 1: Cây đối chứng; 1: Dòng L3; 3: Dòng L5; 4: Dòng U1; 5: Dòng U4
Các cây thế hệ T3 của các dòng chuyển gen U1, U4, L3 và L5
đƣợc gieo trong điều kiện nhà lƣới, tiến hành xử lý hạn ở giai đoạn 3
lá mạ để đánh giá mức độ biểu hiện của các gen quan tâm trong hai
điều kiện xử lý hạn 48h và không xử lý hạn. Kết quả nghiên cứu biểu
hiện OsDREB1A trong cây không chuyển gen, cây chuyển gen trong
điều kiện thƣờng và điều kiện hạn b ng kỹ thuật RT-PCR sử dụng
hai cặp mồi OsDREB1A-RT-Fw/Rv và mồi Actin-Fw/Rv (sử dụng để
nội chuẩn mức độ biểu hiện của OsDREB1A trong tất cả các điều
kiện) đƣợc biểu đồ hóa trong Hình 3.. Kết quả đánh giá cho thấy
OsDREB1A biểu hiện thấp ở cả cây Chành trụi không chuyển gen và
dòng chuyển gen L3 và L5 trong điều kiện không xử lý hạn và ngƣợc
lại với các cây thuộc dòng U1 và U4 gen có mức độ biểu hiện cao
hơn đáng kể (LogFC2 >2). Điều này chứng tỏ với nền cây Chành
Trụi OsDREB1A không có mức biểu hiện cao mặc d đây là giống
lúa nƣơng và dƣới sự điều khiển của promoter Lip9 gen này cũng có
1 2 3 4 5

mức biểu hiện tƣơng đƣơng cây không chuyển gen trong điều kiện
bình thƣờng. Ngƣợc lại, dƣới sự điều khiển của promoter liên tục gen
này có mức độ biểu hiện mạnh (Hình 3.22).
Hình 3.22 | Nghiên cứu mức độ biểu hiện OsDREB1A trong cây chuyển
gen T3

KẾT LUẬN
1. Luận án đã phân lập đƣợc trình tự mã hóa OsDREB1A từ DNA tổng
số giống Cƣờm dạng và cDNA của giống Mộc Tuyền và trình tự mã
hóa OsDREB2A từ cDNA giống Cƣờm dạng. Đã nhân dòng thành
công hai trình tự mã hóa riêng rẽ vào vector pUC19. Kết quả giải trình
tự sau đó cho thấy hai trình tự mã hóa nhân tố phiên mã của hai giống
lúa Việt Nam có trình tự và kích thƣớc (720 bp và 825 bp) tƣơng đồng
với hai gen tƣơng ứng trong ngân hàng gen thế giới là OsDREB1A
(AF300970) và OsDREB2A (AM495895) của giống Nipponbare. Hai
trình tự này sau đó đã đƣợc đƣa thành công một cách riêng rẽ vào hai
hệ vector pBIG/Ubi (promoter Ubiquitine - nguốc gốc từ ngô, là
promoter biểu hiện liên tục) và pBIG/Lip (promoter Lip9 - nguồn gốc
từ lúa, là promoter cảm ứng hạn) để tạo 04 vector biểu hiện (binary
vector): Ubi:OsDREB1A, Ubi:OsDREB2A, Lip9:OsDREB1A và
Lip9:OsDREB2A làm vật liệu phục vụ công tác tạo giống cây trồng
chịu hạn theo định hƣớng chuyển gen.
2. Luận án đã thành công bƣớc đầu trong việc đánh giá khả năng phát sinh
callus (trên 04 nền môi trƣờng MS có bổ sung 2,4D nồng độ từ 1,5 – 2,5
mg/l) của 47 giống lúa nƣơng Việt Nam và khả năng tái sinh chồi từ
callus (trên 04 nền môi trƣờng MS bổ sung BAP và kinetine có nồng độ
khác nhau từ 0,5 – 2 mg/l) của 26 giống lúa nƣơng Việt Nam có khả năng
tạo callus trên 50%. Kết quả cụ thể cho thấy, tập đoàn 47 giống lúa nƣơng
đã khảo sát có khả năng tạo calus không cao, ch có 05 giống lúa có khả
năng tạo callus trên 75% (giống LP-5 trên môi trƣờng MS + 1,5 mg/l
2,4D, giống IR80416-B-152-4, YUNLU103-1B và Nếp hau Để Dỏn
trên môi trƣờng S + 2 mg/l 2,4 , đặc biệt có giống Chành Trụi có khả
năng tạo callus lớn hơn 80% trên cả 3 môi trƣờng MS bổ sung 2,4D). Khả
năng tái sinh chồi từ callus của các giống lúa nƣơng đã khảo sát trong luận

án không cao, khó phát triển quy trình chuyển gen cho các giống lúa. Ch
có 05/26 giống lúa (LUYIN46, IR80416-B-152-4, LHY-4, YUNLU103-
1B và Chành Trụi) có khả năng tái sinh chồi từ callus đạt trên 70% trên
môi trƣờng tái sinh TS2 (MS + 2mg/l BAP + 0,5mg/l kinetine). Luận án
đã thành công trong việc thiết lập quy trình chuyển gen cho giống lúa
Chành Trụi là giống lúa nƣơng địa phƣơng của Việt Nam và đây là kết
quả chuyển gen thành công đầu tiên cho một giống lúa bản địa của Việt
Nam đã đƣợc công bố.
3. Luận án đã thành công trong việc biến nạp 04 vector biểu hiện
Ubi:OsDREB1A, Ubi:OsDREB2A, Lip9:OsDREB1A và
Lip9:OsDREB2A vào giống lúa Chành Trụi. Tỷ lệ chuyển gen trên
giống Chành Trụi dao động từ 9% đến 15%, trung bình là 10%. Luận
án cũng đã lần đầu tiên áp dụng thành công kỹ thuật PCR định lƣợng
(qRT-PCR) ở Việt Nam để thay thế cho: kỹ thuật lai Southern Blotting
trong việc ƣớc tính số lƣợng bản sao của cấu trúc chuyển gen ở các cây
chuyển gen thế hệ T0, T1; t lệ nảy mầm trên môi trƣờng Hygromycin
để xác định cây chuyển gen T1 đồng hợp.
4. Luận án đã thành công trong việc sàng lọc đƣợc 04 dòng Chành Trụi
(Lip9:OsDREB1A) một bản sao, đồng hợp ở thế hệ T2 có kiểu hình (chiều
cao cây, số nhánh, thời gian sinh trƣởng…) tƣơng đồng với cây đối chứng
không chuyển gen trong điều kiện thông thƣờng và 100% có khả năng
phục hồi, kết hạt (tỷ lệ hạt chắc đạt 40-60%) sau 03 tuần ngừng cung cấp
nƣớc. Luận án cũng đã thành công trong việc sử dụng PCR định lƣợng và
bán định lƣợng cho gen chuyển (OsDREB1A) và gen ch thị khả năng
chịu hạn Dehydration-Induced Protein (DIP)/Salt-induced protein (SALT)
trong các cây chuyển gen T2. Kết quả cho thấy cả gen chuyển và gen ch
thị đều ch đƣợc tăng cƣờng biểu hiện trong điều kiện gây hạn nhân tạo so
với cây đối chứng (Fold change-FC: 2,5-3,3).

KIẾN NGHỊ
Dựa trên các kết luận rút ra từ kết quả nghiên cứu trên đây, chúng tôi
đƣa ra một số hƣớng nghiên cứu tiếp theo nhƣ sau:
- Tiếp tục nghiên cứu khả năng duy trì tính ổn định ở các thế hệ
T3, T4… của 04 dòng chuyển gen Chành Trụi đã tạo ra trong nghiên
cứu trong các thế hệ tiếp theo (tính bền vững di truyền, khả năng duy
trì tính chống chịu, năng suất…).
- Mở rộng nghiên cứu chuyển gen OsDREB2A vào các đối
tƣợng cây trồng khác với các promoter điều khiển hoạt động cảm ứng
điều kiện stress để phục vụ công tác chọn tạo giống.

DANH MỤC CÁC CÔNG TRÌNH KHOA HỌC CỦA TÁC GIẢ LIÊN
QUAN ĐẾN LUẬN ÁN
1. Cao Lệ Quyên, Phạm Thị Vân, Phạm uân Hội (2011), “Nghiên
cứu khả năng tạo callus và tái sinh cây từ một số giống lúa nƣơng
Việt Nam nhập nội phục vụ cho công tác chuyển gen”, hí
Si h họ , 33(1), tr. 80-85.
2. Cao Lệ Quyên, Trần Tuấn Tú, Phạm Thị Vân, Phạm uân Hội
(2012), “Thiết kế vector chuyển gen điều khiển OsDREB1A có
tiềm năng tăng cƣờng khả năng chịu điều kiện bất lợi”, hí
g ghệ Si h họ , 10(2), tr. 271-279.
3. Cao Lệ Quyên, Trần Tuấn Tú, Đinh Đoàn Long, Phạm uân
Hội (2016), “Nghiên cứu chuyển gen nhân tố phiên mã
MtOsDREB1A liên quan đến tính chịu hạn vào giống lúa Việt
Nam”. hí N g ghiệ v h i N g h , 8, tr. 13-
19.


















