
The Cooperation Between Two Silencers Creates an Enhancer Element that Controls Both the...
10
Eur. J. Biochem. 239, 23-32 (1996) 0 FEBS 1996 The cooperation between two silencers creates an enhancer element that controls both the lens-preferred and the differentiation stage-specific expression of the rat fl2-crystallin gene Ron P H. DIRKS, Harry J. KRAFT, Siebe T. VAN GENESEN, Erik J. KLOK, Rolph PFUNDT, John G. G. SCHOENMAKERS and Nicolette H. LUBSEN Department of Molecular Biology, University of Nijmegen, The Netherlands (Received 19 December 1995/24 April 1996) - EJB 95 2082/2 The rat /332-crystallin gene is active only during a specific stage of the differentiation of rat lens fibre cells directed by basic fibroblast growth factor. The regulatory elements that determine the transient activity of this gene are located in the -750/-123 region and in the first intron. Singly, these elements act as silencers, together they constitute an enhancer that is active only during the specific differentiation stage. An additional silencer is found between -123 and -77. The proximal promoter region contains a Pax-6 binding site at -65/-51. In vitro, binding to this site could be detected but, according to in vivo footprinting experiments, this site is not occupied in the endogenous gene. Furthermore, co-expression of Pax-6 did not enhance promoter activity. Finally, mutation or deletion of this site did not affect promoter activity: the region -37/+ 10 sufficed for basal promoter activity. The cooperation between the -750/ - 123 region and the first intron of the m2-crystallin gene not only determines the differentiation stage- specific activity of the gene, but also contributes to the highly increased expression in lens cells compared with non-lens cells. Keywords: crystallin; fibroblast growth factor; lens differentiation. The vertebrate lens contains two cell types : epithelial cells, which form an anterior monolayer, and fibre cells, which form the bulk of the lens. The lens grows by differentiation of epithe- lial cells to fibre cells at the equator of the lens. The newly formed fibre cells are thus found at the periphery of the lens, while the older fibre cells occupy the core of the lens. The differ- entiation of the fibre cell is marked by morphological changes, such as cell elongation and loss of cell organelles during termi- nal differentiation, and by the synthesis of the crystallins, the abundant water-soluble structural proteins of the lens. Some of the crystallin genes are used elsewhere as housekeeping or stress genes and the expression of these genes is merely upregulated in the lens (for reviews, see Wistow and Piatigorsky, 1987; Piati- gorsky and Wistow, 1989,1991; De Jong et al., 1989, 1994; Bloemendal and De Jong, 1991). Examples of such genes are the a-crystallin genes and the taxon-specific crystallin genes. The p- and y-crystallin genes have long been thought to be lens-spe- Curresputzdence to N. H. Lubsen, Department of Molecular Biology, University of Nijmegen, Toernooiveld 1, NL-6525 ED Nijmegen, The Netherlands Fax: +31 24 3652938. Note. The first two authors contributed equally to this work. Abbreviations. bFGF, basic fibroblast growth factor; CAT, chloram- phenicol acetyltransferase; CMV, cytomegalovirus; Me2S0,, dimethyl- sulfate; HSV, herpes simplex virus; LM-PCR, ligation-mediated poly- merase chain reaction; RLE cells, rat lens epithelial cells; RNR cells, rat neural retina cells; SRE, serum response element; SRF, serum re- sponse factor; TK, thymidine kinase. Note. The novel nucleotide sequence data published here has been deposited with the EMBL sequence data bank and is available under accession number X83671 cific, but there is accumulating evidence that they may also be expressed at low levels outside the lens. Expression of two J- crystallins has been detected in the chicken retina (Head et al., 1991) and, recently, extralenticular expression of y-crystallin mRNA was detected in early embryonic stages of Xenopus de- velopment (Smolich et al., 1994). The regulation of crystallin gene activity depends upon the state of development as well as on the state of differentiation of the fibre cell. The differentiation and development of the verte- brate lens is thus characterized by a continuously changing pattern of crystallin synthesis (McAvoy, 1978a; Hejtmancik et al., 1985; Van Leen et al., 1987; Aarts et al., 1989; Goring et al., 1992). The resulting complex protein gradient determines the gradient in the index of refraction necessary for the optical properties of the lens. The mode of crystallin gene regulation can be followed in more detail in an in vitro system that has been established in rat. This system is based upon explanted lens epithelial cells, which differentiate synchronously to lens fibre cells in vitro, provided that a high concentration of basic fibroblast growth factor (bFGF) is present (Chamberlain and McAvoy, 1989; McAvoy and Chamberlain, 1989), and has the unique advantage that a homogenous population of fibre cells in intermediate stages of differentiation can be examined. In vitro, a-crystallin mRNA ac- cumulation precedes p-crystallin mRNA synthesis, which in turn precedes y-crystallin mRNA synthesis (Peek et al., 1992 b). Thus, the sequential pattern of crystallin expression in the in vitro system mimics that observed in vivo according to McAvoy (1978b). The crystallin mRNAs reach a steady-state level after two or three days, and, as we have shown that aA- and pB2-
-
Upload
ron-p-h-dirks -
Category
Documents
-
view
213 -
download
1
Transcript of The Cooperation Between Two Silencers Creates an Enhancer Element that Controls Both the...

Eur. J. Biochem. 239, 23-32 (1996) 0 FEBS 1996
The cooperation between two silencers creates an enhancer element that controls both the lens-preferred and the differentiation stage-specific expression of the rat fl2-crystallin gene Ron P H. DIRKS, Harry J. KRAFT, Siebe T. VAN GENESEN, Erik J. KLOK, Rolph PFUNDT, John G. G. SCHOENMAKERS and Nicolette H. LUBSEN Department of Molecular Biology, University of Nijmegen, The Netherlands
(Received 19 December 1995/24 April 1996) - EJB 95 2082/2
The rat /332-crystallin gene is active only during a specific stage of the differentiation of rat lens fibre cells directed by basic fibroblast growth factor. The regulatory elements that determine the transient activity of this gene are located in the -750/-123 region and in the first intron. Singly, these elements act as silencers, together they constitute an enhancer that is active only during the specific differentiation stage. An additional silencer is found between -123 and -77. The proximal promoter region contains a Pax-6 binding site at -65/-51. In vitro, binding to this site could be detected but, according to in vivo footprinting experiments, this site is not occupied in the endogenous gene. Furthermore, co-expression of Pax-6 did not enhance promoter activity. Finally, mutation or deletion of this site did not affect promoter activity: the region -37/+ 10 sufficed for basal promoter activity. The cooperation between the -750/ - 123 region and the first intron of the m2-crystallin gene not only determines the differentiation stage- specific activity of the gene, but also contributes to the highly increased expression in lens cells compared with non-lens cells.
Keywords: crystallin; fibroblast growth factor; lens differentiation.
The vertebrate lens contains two cell types : epithelial cells, which form an anterior monolayer, and fibre cells, which form the bulk of the lens. The lens grows by differentiation of epithe- lial cells to fibre cells at the equator of the lens. The newly formed fibre cells are thus found at the periphery of the lens, while the older fibre cells occupy the core of the lens. The differ- entiation of the fibre cell is marked by morphological changes, such as cell elongation and loss of cell organelles during termi- nal differentiation, and by the synthesis of the crystallins, the abundant water-soluble structural proteins of the lens. Some of the crystallin genes are used elsewhere as housekeeping or stress genes and the expression of these genes is merely upregulated i n the lens (for reviews, see Wistow and Piatigorsky, 1987; Piati- gorsky and Wistow, 1989, 1991; De Jong et al., 1989, 1994; Bloemendal and De Jong, 1991). Examples of such genes are the a-crystallin genes and the taxon-specific crystallin genes. The p- and y-crystallin genes have long been thought to be lens-spe-
Curresputzdence to N. H. Lubsen, Department of Molecular Biology, University of Nijmegen, Toernooiveld 1, NL-6525 ED Nijmegen, The Netherlands
Fax: +31 24 3652938. Note. The first two authors contributed equally to this work. Abbreviations. bFGF, basic fibroblast growth factor; CAT, chloram-
phenicol acetyltransferase; CMV, cytomegalovirus; Me2S0,, dimethyl- sulfate; HSV, herpes simplex virus; LM-PCR, ligation-mediated poly- merase chain reaction; RLE cells, rat lens epithelial cells; RNR cells, rat neural retina cells; SRE, serum response element; SRF, serum re- sponse factor; TK, thymidine kinase.
Note. The novel nucleotide sequence data published here has been deposited with the EMBL sequence data bank and is available under accession number X83671
cific, but there is accumulating evidence that they may also be expressed at low levels outside the lens. Expression of two J- crystallins has been detected in the chicken retina (Head et al., 1991) and, recently, extralenticular expression of y-crystallin mRNA was detected in early embryonic stages of Xenopus de- velopment (Smolich et al., 1994).
The regulation of crystallin gene activity depends upon the state of development as well as on the state of differentiation of the fibre cell. The differentiation and development of the verte- brate lens is thus characterized by a continuously changing pattern of crystallin synthesis (McAvoy, 1978a; Hejtmancik et al., 1985; Van Leen et al., 1987; Aarts et al., 1989; Goring et al., 1992). The resulting complex protein gradient determines the gradient in the index of refraction necessary for the optical properties of the lens.
The mode of crystallin gene regulation can be followed in more detail in an in vitro system that has been established in rat. This system is based upon explanted lens epithelial cells, which differentiate synchronously to lens fibre cells in vitro, provided that a high concentration of basic fibroblast growth factor (bFGF) is present (Chamberlain and McAvoy, 1989; McAvoy and Chamberlain, 1989), and has the unique advantage that a homogenous population of fibre cells in intermediate stages of differentiation can be examined. In vitro, a-crystallin mRNA ac- cumulation precedes p-crystallin mRNA synthesis, which in turn precedes y-crystallin mRNA synthesis (Peek et al., 1992 b). Thus, the sequential pattern of crystallin expression in the in vitro system mimics that observed in vivo according to McAvoy (1978b). The crystallin mRNAs reach a steady-state level after two or three days, and, as we have shown that aA- and pB2-

24 Dirks et al. (EUK J . Biochern. 239)
crystallin mRNAs are stable, the transcriptional activity of these crystallin genes must cease at that time. The activity of the crys- tallin genes is thus restricted to a narrow differentiation window (Peek et al., 1992bj.
To understand the molecular basis of crystallin gene regula- tion, we thus need )to ask not only how the crystallin promoters are switched on at a specific stage of lens fibre cell differentia- tion but also how they are switched off at a later stage. Recently, we identified the regulatory events that mediate the differentia- tion stage-specific (activity of the rat yD-crystallin gene (Dirks et al., 1996). We focus here on the mechanism(s) responsible for the restriction of pB2-crystallin gene activity to a particular differentiation state and have used the in vitvo differentiating system to assay for fibre cell stage-specific recognition of the regulatory elements of the rat m2-crystallin promoter. We show that the differentiation stage-specific enhancer of the m2-crys- tallin gene is the result of a complex interplay between upstream and intronic elements. We also demonstrate that the same tran- scription regulatory elements are active in rat neural retina (RNR) cells and in fibroblasts.
MATERIALS AND METHODS
RNA and DNA sequence determination. RNA was iso- lated using standand methods (Ausubel et al., 1987). To deter- mine the 5’ mRNA sequence and map the transcription initiation site of the m2-crystallin gene, an oligonucleotide complemen- tary to the last non-coding and the first 18 coding nucleotides of the PB2-crystalIin mRNA (pr2-2: 5’-CTG GTG GTC TGA GGC CAT G-3’) (Aarts et al., 1989) was extended on total rat RNA with reverse transcriptase in the presence of dideoxy- nucleotides (Geliebter et al., 1986). On the basis of the RNA sequence results, an oligonucleotide was designed (pr2-1: 5’- GAC CAG ATN GCC AGC CCC NGC GAG-3’), complemen- tary to the last 21 nucleotides of the first exon and the first three nucleotides of the second exon, and used to identify the location of the first non-cod.ing exon on the restriction map of the cosmid clone pRcosp-3 (Aarts et al., 1987; Aarts, 1989). To obtain the genomic sequence, the appropriate restriction fragments were cloned into pBluescript and sequenced by the dideoxy chain- termination method (Sanger et al., 1977). Sequence data were recorded, edited and compared using Intelligenetics Suite soft- ware. .
/?B2-crystallin/CAT gene fusion constructs. For the con- struction of the /B:!-crystallin/CAT gene fusion plasmids a Ball restriction site (primer: 5’-GGG CTG GCC ATC TGG-3’) was created by site-directed mutagenesis (Sambrook et al., 1989) at position +20 in the first non-coding exon of the PB2-crystallin gene. Subsequently, NcoI (-750), SmuI (- 123) and PstI (-77) restriction sites were used to clone the -750/+20, -123/+20 and -77/+20 fragments of the /B2-crystallin gene into the pSCAT vector, in which the promoter-less CAT gene is preceded by the pUC12 polylinker. The -750/+1000 plasmid containing the first intron was constructed in two steps. First, the 250-bp NcoI fragment, containing the 3‘ end of the first intron up to the translation initiation site in the second exon, was isolated and blunt-ended with rriung bean nuclease to remove the translation initiation site, after which it was cloned in pSCAT. Second, the Ncol fragment containing the rest of the first intron, the first exon and approximately 750 bp of upstream sequences, was cloned in front of the 250-bp NcoI fragment. The - 123/+ 1000 and -77/+ 1000 constructs were derived from the -7501+1000 construct by internal deletion of, respectively, the -750/- 123 and the -750/-77 region. The +20/+1000 construct was made by cloning the I-kb Thul-Hind111 fragment of the -750/+1000
construct into pSCAT. The same 1-kb intron fragment was cloned in either orientation in front of the /lB2-crystallin pro- moter fragment of the -750/+20 construct. Site-directed mu- tants of the pB2-crystallin promoter were made by first cloning the -77/+20 fragment into pBluescript I1 KS- (Stratagene). Sin- gle-stranded DNA was isolated and mutageiiesis was performed according to the manual of the Sculptor in vitro mutagenesis system version 3 (Amersham). The oligonucleotide (5’-CAG TGT TCA CAG ATC TTG GCA C-3’) was used for the C-61/ C-60 + A-61/G-60 mutation and the oligonucleotide (5’-CAC CCA TCT TTT CAC TGG CTA C-3‘) for the G-54/G-53 - T-54/T-53 mutation. Mutagenesis was checked by dideoxy se- quence analysis (Sanger et al., 1977) and the mutated -77/+20 fragments were recloned into pSCAT. The -611-60 mutation also created a BglII restriction site, which was used to generate the -59/+20 construct by internal deletion of the -77/-60 re- gion from the -77/+20 construct. Similarly, a unique EcoNI site was used to make the -37/+20 construct by internal dele- tion of the -77/-38 region. /332-crystallin-thymidine-kinase fusion constructs were made by cloning the -1231-77 or the +20/+1000 fragment of the /B2-crystallin gene in either orien- tation in front of the herpes simplex virus (HSV) thymidine ki- nase (TK) promoter in pBLCAT2. pBLCAT2 has been described previously (Luckow and Schiitz, 1987). The -38/+20 pB2- crystallin fragment of pRcosp-3 (Aarts et al., 1987) was ampli- fied in 30 PCR cycles using the sense primer (5’-CGG GAT CCG GGA AGG TAT AAA TAC C-3’) and the antisense primer (5’-CCC AAG CTT CCA GCC CCC GCG-3’). Denaturation was for 90 s at 94”C, annealing for 2 min at 44°C and polymer- ization for 3 min at 72°C. The -38/+10 fragment was derived from the -38/+20 fragment by using the ThuI site at + lo . The -77/+20, -38/+20 and -38/+10 fragments were gel- purified and cloned into the pEUCAT vector (Piaggio and DeSi- mone, 1991). All constructs were confirmed by restriction site mapping and sequence determination.
Cell cultures. Rat lens epithelial cells were isolated and cul- tured as described (Peek et al., 1992a). RNR cells were obtained by trypsinization of the neural retina isolated from newborn rats, followed by culturing under the same conditions as for rat lens epithelial cells. Lens epithelial explants were made from 1-3- days-old rat lenses, essentially as described (McAvoy and Cham- berlain, 1989). Explants were cultured in M199 medium (Gibco Laboratories), supplemented with 0.1 % BSA, either with or without 25 ng/ml bFGF (the optimal concentration for this batch, kindly donated by Scios Inc., Mountain View CA). Ltk- cells were cultured in Dulbecco’s modified Eagle’s medium enriched with 10% fetal calf serum.
Transfections, chloramphenicol acetyltransferase assay and P-galactosidase assay. Supercoiled plasmid DNA was puri- fied by CsCl gradient centrifugation (Sambrook et al., 1989) and quantitated by means of both ethidium bromide staining and spectrophotometric measurement. Cultured cells were transfected at approximately 70 % confluence. Explants (five/ 3.5-cm dish) were traiisfected after culture in the presence or absence of bFGF, as indicated. In each dish, 2 pg pB2-crystallin/ CAT reporter construct and 200 ng cytoinegalovirus (CMV)/P- gal construct (Jain and Magrath, 1991) were transfected using lipofectamine (Gibco BRL) according to the manufacturer’s pro- tocol. In Pax-6 cotransfection experiments, 500 ng pCMV or pCMV-Pax-6 (Epstein et al., 1994b) (kindly provided by Dr Maas) was combined with the /B2-crystallin/CAT and CMVIP- gal constructs. Three days after addition of the DNA, the cells were harvested in 200 pl reporter lysis buffer (25 mM bicine pH 7.8, 0.05% Tween-20, 0.05% Tween-80). To determine the transfection efficiency, 25 pl cell lysate was used to assay for P-galactosidase activity (Jain and Magrath, 1991). After the P-

25 Dirks et al. ( E m J . Biochem. 239)
-150 -100 -200 ttgcccagccgcctgacacactgcgcagccgaatctgaaagctaatgacattattgtgtggagacacaatgtctgtgggcatttgctgacccgggctttggtgtaggcttagccctcatt
ctcaccctgctttctgcagggatacagtgttcacccatct tggcactggctaccctggggaaggtataaataccacctcccaccggcctggcttcACGGCACTCGCGGGGGCCGATC - 5 0 1
TGgt~agaaggctt~tgctgcttgctctgccctt~gctcttctttcattcttgt~ggaaagtgc~gagtggagacttcggcacttggcctcagttttcccttct~ttaattggg~agaag 50 100
2 0 0 2 5 0 150 gggtcgccaaagcctctctgacatgggttaggaagcaatctagcagactggcccagtgcacagccagtgcctggacaaggatactcattaatatattaaccagcttctcagcacctattt
TGgt~agaaggctt~tgctgcttgctctgccctt~gctcttctttcattcttgt~ggaaagtgc~gagtggagacttcggcacttggcctcagttttcccttct~ttaattggg~agaag 50 100
2 0 0 2 5 0 150 gggtcgccaaagcctctctgacatgggttaggaagcaatctagcagactggcccagtgcacagccagtgcctggacaaggatactcattaatatattaaccagcttctcagcacctattt
300 350 ttttaatactagaaaagaagactagaaaggaagatgcatttggcttgtgaaccagaggtgaccccaccttgaccatgtctaggttaccgagccagttttctcatgtctagtgattgcagt
400 4 5 0 5 0 0 tcccttgggccagccttggggaaaacttgtggactaacctcagctttacgtgttcctgagtgctataatttcagtgtcaataaatcccctggaaatgtcctaaccatgtcaccctggttg
tcccttcttgggtgcagggtgtgccttccagggccctttcagatgcacacagttgtatgcagtcttggttcagagatggtgaggcacctctattactttcctgatgtacagagtggtaaa
ctgaggcacaaggagagggtaggacttacgggagaccctttagctgctgggtggcagagcaagtacttgggctggcctaattgagcccaacacctaaaggtagagaatttggtgagctgg
aagtggtctcagggaagccttctgacatgactccatagcgctggagtctagcatcaggggagacaatccatgggagagatccttctgctcagagctgggttggattcatagtgaaggtga
agccagagcgcagatggctgcccccagaacaatctgaaacccagtaaaagaggagagggctgaaggctggaccaggttcatccagaagtcctctggcagcttgaagccaagaccagcaag
g t C C t C a t g C a g t C t t t C t t t C a ~ T ~ C ~ C G A ~ C ~ G A G A G ~ ~ C ~ ~ C ~ ~ G A C ~ C ~ G
550 600
6 5 0 700
750 800 850
900 950
1000 1050 - Fig. 1. Nucleotide sequence of the promoter region, exon 1, intron 1 and part of exon 2 of the rat /3BZ-crystallin gene. The exonic sequences are capitalized, intronic and flanking sequences are shown in lower case. The transcription initiation site is denoted by 1. The TATA box is underlined. The ATG initiation codon in the second exon is double underlined.
galactosidase assay, cellular deacetylases were inactivated by heating for 15 min at 65°C. Aliquots of cell lysate correspond- ing to equal p-galactosidase activities were assayed for CAT ac- tivity according to Gorman et al. (1982) or using the Quan-T- CATTM system (Amersham). Every experiment was performed at least twice with two batches of DNA.
In vivo footprinting. Lens explants (10/3.5-cm dish) were cultured in the presence of bFGE The culture medium was re- placed by medium containing 0.1 % dimethyl sulfate (Me,SO,; Fluka). After a 2-min incubation at 37 "C, the Me,SO,-contain- ing medium was removed and the cells were washed four times with NaClP, (140 mM NaCI, 2.5 mM KC1, 8.1 mM NalHPO,, 1.5 mM KH,PO,, pH 7.5) at 37°C. Me,SO,-treated explants were transferred to 100 pl lysis buffer (300 mM NaCl, 50 mM Tris/HCl pH 8.0, 25 mM EDTA pH 8.0, 200 pg/ml proteinase K, 0.5% sodium dodecyl sulfate) and incubated overnight at 37 "C. The lysate was extracted once with phenol/chloroform and once with n-butanol and subsequently diluted with 10 mM Tris/HCI pH 7.5, 1 mM EDTA (Tris/EDTA) to 500 pi. The ly- sate was applied to a Microcon-30 (Amicon) filter unit and spun for 15 min at room temperature in an Eppendorf centrifuge. The chromosomal DNA was washed with 400 p1 Tris/EDTA and re- suspended in 50 p1 TrNEDTA. Me,SO, methylation of control protein-free chromosomal DNA was performed as described (Maxam and Gilbert, 1980). The Me,SO, modified DNA was precipitated in ethanol and redissolved in 100 pl 10% piperidine (Sigma). After incubation for 30 rnin at 90"C, the DNA was lyophilized in a Speedvac and redissolved in Tris/EDTA. To re- move trace amounts of piperidine the DNA was successively precipitated in ethanol, redissolved in Tris/EDTA, precipitated in isopropanol, redissolved in water, lyophilized in a Speedvac and redissolved in TrislEDTA. The @,-crystallin promoter frag- ments in an amount of DNA, equivalent to the yield from five explants, were amplified by ligation-mediated polymerase chain reaction (LM-PCR) as described below.
Ligation-mediated polymerase chain reaction. LM-PCR was performed essentially as described (Mueller and Wold, 1989; Garrity and Wold, 1992). For visualization of the upper strand of the m2-crystallin promoter the following primers were
used: rp2 - l ( a ) , (5'-CCG CAC TTT CCA ACA AGA ATG A-3'), position +91 to +70; rpB2-2(a), (5'-AAG GGC AGA GCA AGC AGC AGA AGC-3'), position +59 to +36; r p 2 - 3(a), (5'-AAG GGC AGA GCA AGC AGC AGA AGC CTT C- 3'), position +59 to +32. Primers r/B2-l(a), rpB2-2(a) and rj3B2-3(a) were annealed at 60°C, 72°C and 73"C, respectively. Primers that were used for visualization of the lower strand were: rpB2-l(s), (5'-AGC CGA ATC TGA AAG CTA ATG ACA-3'), position -189 to -166; r@2-2(s), (5'-CAC AAT GTC TGT GGG CAT TTG CTG AC-3'), position -151 to -126; rpB2-3(s), (5'-CAC AAT GTC TGT GGG CAT TTG CTG ACC C-3'), position -151 to -124. Primers rpB2-1(s), rpS2-2(s) and rpB2-3(s) were annealed at 60"C, 72°C and 73 "C, respectively. The pB2-crystallin gene fragments were am- plified in 20 PCR cycles and labelled in two additional PCR rounds. Labelled fragments were electrophoresed in a 6% se- quencing gel and exposed to Fuji AX film. The footprinting data were quantified by scanning the autoradiograms with a densi- tometer (Kipp & Zonen, Delft).
Nuclear extract preparation and gel shift assay. Whole cell extracts of explant cells were prepared and binding reactions were performed as described (Ray et al., 1992), except that Hepes pH 6.8 was used instead of Hepes pH 7.9. Nuclear ex- tracts from whole rat lenses were made as described previously (Peek et al., 1992a). Extracts were first incubated for 10 rnin on ice in the presence of 1 pg poly(d1-dC) and 0.4 pg herring sperm DNA to suppress non-specific binding. DNA restriction frag- ments containing j332-crystallin promoter regions were then end-labelled by filling in sticky ends with Klenow DNA poly- merase in the presence of [(x-32P]dATP. Approximately 0.05 - 0.5 ng double-stranded probe (10000-20000 cpm) was mixed with the samples. For competition experiments, the specific competitor was added before the labelled probe to the sample in the molar excess indicated in the figures. Binding was allowed to proceed for 10 min at room temperature, after which the sam- ple was loaded directly on a native 5 % polyacrylamide gel and run in 0.25XTrishoric acid/EDTA (1 XTris/boric acid/EDTA is 0.089 M Tris,,0.089.M boric acid, and 0.0025 M EDTA pH 8.3), at 1.5 V/cm with recirculation of the buffer. Gels were run

26 Dirks et al. (Eur: J . Biochem. 239)
-77 t 2 D
-123 t 2 D
-750 t i 0 0 0
rel. rel. rel. rel. CAT CAT 10-13d tbFGF
act. act. 0-3d -bFGF CAT 4-7d tbFGF CAT 7-10d tbFGF
act. act.
100 100 100 100
35 15 16 20
19
17
4
5
5
143
4
a
Fig. 2. Differentiation stage-specific antisilencer activity of the first intron of the rat /lB2-crystallin gene. Explants at various stages of the differentiation process were transfected with the D2-crystallinlCAT fusion genes shown on the left. Black, white and striped boxes represent restriction fragments used for their construction (see also Materials and Methods). The amount of extract used for the CAT assays was normalized for the activity of the co-transfected CMV-lacZ construct and can be directly compared at anyone time point. They cannot be compared between time points as the transfection efficiencylp-galactosidase activity is variable. The /I-galactosidase activities were (relative to day 0-3 set at 1) : day 4-7 = 5 ; day 7-10 = 8.5; day 10-13 = 4. CAT activities (rel. CAT act.) were determined by densitornetric scanning of the autoradiographic signal of the acetylated chloramphenicol and are indicated for each separate time point, as values relative to the activity of the -77/+20 construct (100%)
for 60 min prior to loading. The synthetic double-stranded Pax-6 binding site of the mouse aA-crystallin promoter (sense strand 5’-CCC TTA ATT CCT CCA TTC TGT GCA GGC ATA T-3’) was according to Cvekl et al. (1995a). Anti-Pax-6 antibody (Carriere et al., 1903) waq kindly provided by Dr S. Saule. The anti-/lB2-crystallin antibody was a rabbit polyclonal antibody raised against recoiinbinant rat PB2-crystallin.
RESULTS Identification of transcription regulatory elements in the rat j?BZ-crystallin gene. To map the regulatory elements involved in transcription of rat p-crystallin genes, we choose the promoter of the late /7B2-crystallin gene, which is primarily expressed postnatally (Aarts et al., 1989). This gene was chosen because it is abundantly expressed during days 5-8 of in vitro differenti- ation (Peek et al., 1992b) and also because it is located only 5 kb downstream of the jK-crystallin gene (Aarts et al., 1987), thus limiting the 5‘ upstream region that needs to be scanned for regulatory elements. Fig. 1 shows the sequence of the promoter region, exon 1, intron 1 and part of exon 2 of the rat pB2-crystal- lin gene. We first tried to map the regulatory elements of the m2-crystallin gene in primary cultures of newborn rat lens epi- thelial (RLE) cells. Although these cells do not express the en- dogenous p- and y-crystallin genes, in a transient reporter gene analysis the rat yD-crystallin promoter is highly active in these cells (Peek et al., 1’992a). However, when the jB2-crystallin pro- moter was tested fix activity in these cells, no activity could be detected. To determine whether the lack of expression of the pB2-crystallin proimoter constructs was due to the absence of essential sequence information in our constructs, we scanned the 5-kb intergenic region between the pB2- and the jB3-crystallin gene, as well as the introns and 6 kb of the 3’ flanking region of the /jB2-crystallin gene for enhancing elements by cloning overlapping fragments of these regions in front of the HSV TK promoter. No enhancing elements were found (data not shown). We then turned to explanted lens epithelial cells as transfection hosts, since accumulation of /332-crystallin mRNA during days 5-8 of in vitro differentiation of these cells implies that at least the endogenous promoter is active (Peek et al., 1992b).
Explanted rat Itms epithelial cells differentiate synchronously to lens fibre cells in vitro i n the presence of bFGF (McAvoy and Chamberlain, 1985)). This experimental system has the unique advantage that lens fibre cells at defined intermediate stages of
differentiation can be used as transfection hosts. Thus the possi- bly changing recognition of crystallin promoters in lens fibre cells during differentiation can be assayed. However, it must be remembered that the activity of the constructs is measured over a three-day period (explants were harvested three days after transfection), in which the differentiation stage of the cells con- tinues to change.
To test the hypothesis that the transient accumulation of m2-crystallin mRNA during fibre cell differentiation is due to the differentiation stage-specific activity of the /B2-crystallin promoter, ,8B2-~rystallin/chloramphenicol acetyltransferase (CAT) fusion genes (four of which are depicted in Fig. 2) were transfected into explants at various times after the initiation of the differentiation process. Of the four constructs initially tested, only the -750/+1000 construct, which contains the entire first intron of the m2-crystallin gene, shows the pattern of activity that corresponds with the transient accumulation of m2-crystal- lin mRNA: it has transcriptional activity only in explants transfected after 7 days of exposure to bFGF, not at earlier or later differentiation stages (Fig. 2). The -750/+20 construct, in which the first intron is deleted, is completely inactive through- out the differentiation process, which clearly indicates that the first intron is essential for the transient promoter activity. Dele- tion of the region between -750 and -123 bp partly restores the activity of the promoter and deletion up to -77 bp increases promoter activity even further, which suggests that both the - 750/- 123 and the - 1231- 77 region contain a negative tran- scription regulatory element (Fig. 2). In contrast with the tran- sient activity of the -750/+1000 construct, the -123/+20 and -771-t 20 constructs are constitutively active throughout the differentiation process. The activity of the -77/+20 construct is weak, only 2-3-fold higher than the background activity of a promoterless CAT construct (data not shown). In these initial experiments four regulatory elements can be discriminated : an activator in the -77/+20 region, two negative regulatory ele- ments located between -750 and -123 and between -123 and -77, respectively, and an element in the first intron. As will be outlined below, the interaction between the -7501-123 element and the first intron abolishes the silencer activity and creates an enhancer at a specific stage of differentiation, a stage that coincides with the activity of the endogenous /B2-crystallin gene.
The -77/+20 region. A putative transacting factor required for activity of the pB2-crystallin promoter is expected to bind to

Dirks et al. (ELK J. Biochem. 239) 27
C BE2 probe w
anti-Pax-6 antibody - - 1 1 1 3pl - - 0 r -61/-60 -541-53 0 0 Pax-6 competitor - - ; ; anti- /3B2-crystallin antibody - - - - ip i 3p1
I 1 - n , . , I
ens extract - t - t - t lens extract - t t t t lens extract - t t t t t
F BE2 WT probe BE2 WT probe
Fig.3. A nuclear lens factor binding to the -77/+20 /B2-crystallin promoter fragment. (A) Binding capacity of wild-type and mutant rat /B2- crystallin promoter fragments in a gel shift assay using newborn rat lens nuclear extracts. 'zP-labelled wild-type (WT) and mutant (-61/-60, -54/-53) -77/+20 /lB2-crystallin promoter fragments were incubated in the presence (+) or absence (-) of newborn rat lens nuclear extract and DNA-protein complex formation was analysed by electrophoresis in a native polyacrylamide gel. F, free probe. (B) Competition between a synthetic Pax-6 binding site and the rat fB2-crystallin promoter for binding to a nuclear lens factor. "P-labelled wild-type -77/+20 m2-crystallin promoter fragment was incubated in the absence (-) or presence (+) of newborn rat lens nuclear extract and in either the absence (-) or presence of 1 - 100-fold molar excess of an oligonucleotide containing the Pax-6 binding site of the mouse aA-crystallin promoter (Cvekl et al., 1995a). DNA- protein complex formation was analysed by electrophoresis in a native polyacrylamide gel. (C) Binding of Pax-6 to the rat /B2-crystallin promoter. 32P-labelled wild-type -77/+20 /B2-crystallin promoter fragment was incubated in the absence (-) or presence (+) of newborn rat lens nuclear extract and in either the absence (-) or presence of 1-3 p1 of anti-Pax-6 antibody or of anti-fB2-crystallin antibody. DNA-protein complex formation was analysed by electrophoresis in a native polyacrylamide gel.
the -77/+20 region. Use of this region as probe in a band shift assay, in which nuclear extracts prepared from whole rat lenses were used as the source of nuclear factors, yields a prominent DNA-protein complex (Fig. 3). The specificity of the interaction of the -77/+20 region with factors in the (whole lens) extracts was tested by competing either with an unlabelled -77/+20 fragment or with an unlabelled -122/-77 fragment (data not shown). Only the -77/+20 fragment competed for binding, the -122/-77 region did not, showing that the interaction is sequence-specific. Complex formation with the - 77/+20 region is found at all stages of the differentiation process including epi- thelial cells, which is in accordance with our observation that the -77/+20 construct is constitutively active (data not shown).
In search for consensus binding sites for known transcription factors in the -77/+20 region, we recognized that the sequence CCCATCTTGG (positions -62 to -53) closely resembles the consensus binding site CC[A/T],GG of the serum response factor (SRF), a direct target of the signal transduction cascade initiated by several polypeptide growth factors (Faisst and Meyer, 1992). To test the possibility that the m2-crystallin pro- moter interacts with an SRF, we disrupted the serum response element-like sequence by site-directed mutagenesis: C-61/C-60 was changed into A-61/G-60, and G-54/G-53 was changed into T-541T-53 (Fig. 4). Mutagenesis of nucleotides -61/-60 com- pletely abolishes the capacity of the -77/+20 fragment to form the prominent DNA-protein complex (Fig. 3 A), indicating that C-61 and/or C-60 specifically interact in vitro with a lens nuclear protein. In contrast, complex formation between the -54/-53 mutant and the nuclear lens factor is not significantly affected (Fig. 3A), making it highly unlikely that an SRF interacts with the -61/-60 element.
Recently, Pax-6 has been shown to act as a regulator of eye development (reviewed by Hanson and Van Heyningen, 1995). When we searched the sequence of the pB2-crystallin gene (Fig. 1 ) for the presence of putative Pax-6 binding sites (Epstein et al., 1994a), the best match was found at a site located be- tween -65 and -51 bp (Fig. 4). Whereas the -61/-60 muta- tion decreases the similarity with the Pax-6 consensus binding
site, the -54/-53 mutation rather increases it, which corres- ponds exactly with the binding activity of the mutant m2-crys- tallin promoter fragments in the gel shift assay (Fig. 3A). When we used the Pax-6 binding site of the mouse nA-crystallin pro- moter (Cvekl et d., 1995a) as a competitor i n the gel shift assay, formation of the DNA-protein complex was partially inhibited (Fig. 3 B). Moreover, the mobility of the same DNA-protein complex was affected by the addition of anti-Pax-6 antibodies, whereas anti-m2-crystallin antibodies had no effect (Fig. 3 C). Thus, the in vitro binding studies suggest that the m2-crystallin promoter contains a binding site for Pax-6 at the -61/-60 re- gion. To test whether Pax-6 can directly activate the m2-crystal- lin promoter, differentiating lens explants were cotransfected with a Pax-6 expression construct and wild-type or mutant -77/ + 20 @2-crystallin/CAT construct. Expression of Pax-6 does not significantly affect the activity of the -77/+20 /B2-crystal- lin promoter fragment (Fig. 5) . More importantly, the activity of the -61/-60 mutant is equally high as the activity of the wild- type promoter fragment (Fig. 5). These results suggest that the lens nuclear protein that interacts with the -61/-60 element in vitro is not necessary for activity of the -77/+20 promoter frag- ment in vivo. To examine whether an interaction at -61/-60 does occur at the endogenous promoter, we performed in vivo Me,SO, footprint analysis of explants at several stages of the differentiation process. No footprint could be detected within the first 77 bp upstream of the transcription start site in explants that had been exposed to bFGF for 7 days (Fig. 6), or at other stages of the differentiation process (data not shown). Only C+11 in the lower strand is almost fully protected, whereas a site of strong hypermethylation is visible at G-204 in the upper strand. Thus, although the -61/-60 element interacts with a nuclear lens factor (probably Pax-6) in vitro, both our reporter gene assays and the Me,SO, footprint analysis indicate that it does not play a role in vivo.
To identify the minimal region required for basal promoter activity, -59/+20 and -37/+20 deletion constructs were made (Fig. 4) and their activities tested in differentiating rat lens ex- plants. Both constructs were as active as the -77/+20 construct
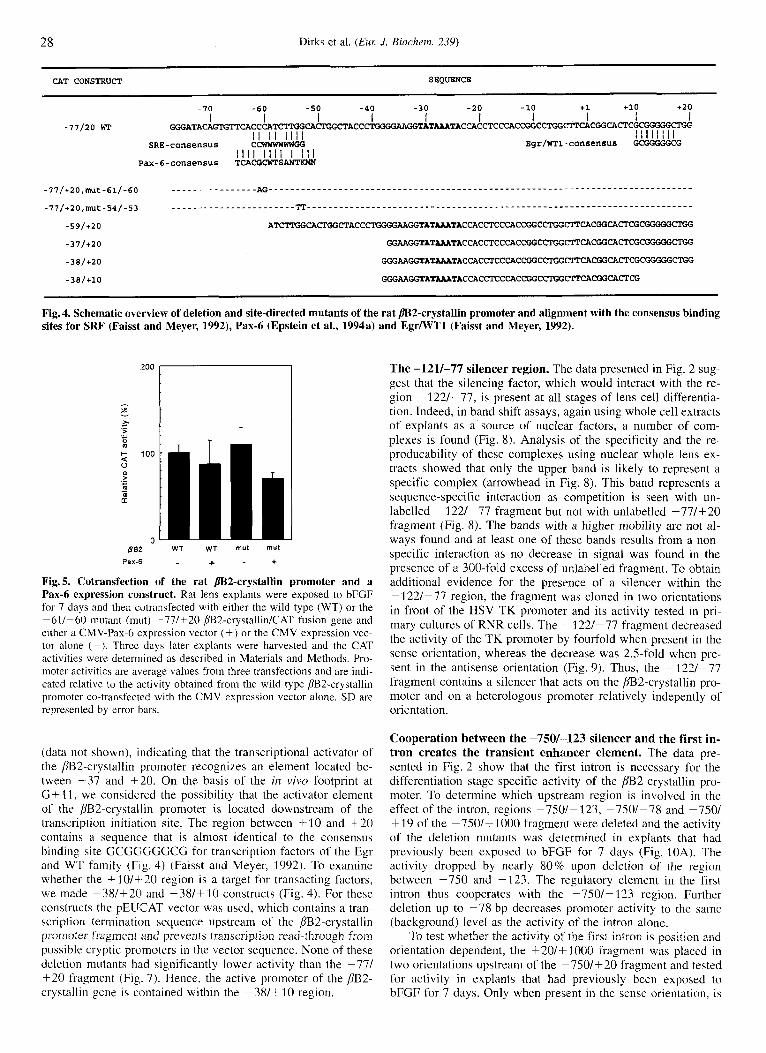
28 Dirks et al. ( E m J. Biochern. 239)
CAT CONSTRUCT SEQUENCE
- I0 -60 -50 -40 -30 -20 -10 +1 +10 +20
GGGRTACAGTGTPCACCCAT~~~~ACCCT I I I I I I I I I I
I I I I I I I I -77120 WT
Bgr/WTl-cOnSensus GGGGGGCG I I I I I I I I
I I I I I I I I I i l l SRE-consensus CCWWWWWWGG
Yax-6-conaen~us TCACGCWTS"
Fig. 4. Schematic overview of deletion and site-directed mutants of the rat m2-crystallin promoter and alignment with the consensus binding sites for SRF (Faisst and Meyer, 1992), Pax-6 (Epstein et al., 1994a) and EgrlWTl (Faisst and Meyer, 1992).
:zoo
- s - ._ w 0 .- - t 100 0 m ._ c m m U -
0 5Bil
P a x 4
T
WT WT mut mut
+ - +
Fig.5. Cotransfection of the rat /BZ-crystallin promoter and a Pax4 expression construct. Rat lens explants were exposed to bFGF for 7 days and then cvtransfected with either the wild-type (WT) or the -61/-60 mutant (mut) -77/+20 /332-~rystallin/CAT fusion gene and either a CMV-Pax-6 expression vector (+) or the CMV expression vec- tor alone (-). Three days later explants were harvested and the CAT activities were determined as described in Materials and Methods. Pro- moter activities are average values from three transfections and are indi- cated relative to the activity obtained from the wild-type @2-crystallin promoter co-transfected with the CMV expression vector alone. SD are represented by error bars.
(data not shown), indicating that the transcriptional activator of the /IB2-crystallin promoter recognizes an element located be- tween -37 and +20. On the basis of the in vivo footprint at G + 11, we considered the possibility that the activator element of the pB2-crystallin promoter is located downstream of the transcription initiation site. The region between +10 and +20 contains a sequence that is almost identical to the consensus binding site GCGC;GGCCC for transcription factors of the Egr and WT family (Fig. 4) (Faisst and Meyer, 1992). To examine whether the +10/+-20 region is a target for transacting factors, we made -38/+20 and -38/+10 constructs (Fig. 4). For these constructs the pEUCAT vector was used, which contains a traii- scription termination sequence upstream of the BB2-crystallin promoter fragment and prevents transcription read-through from possible cryptic promoters in the vector sequence. None of these deletion mutants had significantly lower activity than the -77/ +20 fragment (Fig. 7). Hence, the active promoter of the BB2- crystallin gene is contained within the -38/+10 region.
The -121/-77 silencer region. The data presented in Fig. 2 sug- gest that the silencing factor, which would interact with the re- gion -122/-77, is present at all stages of lens cell differentia- tion. Indeed, in band shift assays, again using whole cell extracts of explants as a source of nuclear factors, a number of com- plexes is found (Fig. 8). Analysis of the specificity and the re- producability of these complexes using nuclear whole lens ex- tracts showed that only the upper band is likely to represent a specific complex (arrowhead in Fig. 8). This band represents a sequence-specific interaction as competition is seen with un- labelled - 122/-77 fragment but not with unlabelled -77/+20 fragment (Fig. 8). The bands with a higher mobility are not al- ways found and at least one of these bands results from a non- specific interaction as no decrease in signal was found in the presence of a 300-fold excess of unlabelled fragment. To obtain additional evidence for the presence of a silencer within the -122/-77 region, the fragment was cloned in two orientations in front of the HSV TK promoter and its activity tested in pri- mary cultures of RNR cells. The -122/-77 fragment decreased the activity of the T K promoter by fourfold when present in the sense orientation, whereas the decrease was 2.5-fold when pre- sent in the antisense orientation (Fig. 9). Thus, the -122/-77 fragment contains a silencer that acts on the /IB%-crystallin pro- moter and on a heterologous promoter relatively indepently of orientation.
Cooperation between the -7501-123 silencer and the first in- tron creates the transient enhancer element. The data pre- sented in Fig. 2 show that the first intron is necessary for the differentiation stage-specific activity of the [1'B2-crystallin pro- moter. To determine which upstream region is involved in the effect of the intron, regions -750/-123, -7.501-78 and - I N / +19 of the -750/+1000 fragment were deleted and the activity of the deletion mutants was determined in explants that had previously been exposed to bFGF for 7 days (Fig. 10A). The activity dropped by nearly 80% upon deletion of the region between -750 and -123. The regulatory element in the first intron thus cooperates with the -750/-I23 region. Further deletion up to -78 bp decreases promoter activity to the same (background) level as the activity of the intron alone.
To test whether the activity of the first intron is position and orientation dependent, the +20/+ 1000 fragment was placed in two orientations upstream of the -750/+20 fragment and tested for activity i n explants that had previously been exposed to bFGF for 7 days. Only when present in the sense orientation, is

Dirks et al. (Eur: .I. Aiochem. 239) 29
lower strand F
T 7
upper strand F
T 7 r-7 tll- t 2-
-12- -20-
-41-
-50-
- Gt11 -204-
-153-
-121-
-105-
-80-
-69-
-60- -53-
-71- -39-
-32-
-10-
- G-204
Fig. 6. Genomic footprint analysis of the rat /BZ-crystallin gene pro- moter. Newborn rat lens explants were exposed to bFGF and then treated with Me2S0, as described in Materials and Methods. The chro- mosomal DNA was nicked at sites of modified G residues by treatment with piperidine. @2-crystallin gene fragments were amplified and la- belled by LM-PCR and analysed on a sequencing gel. Open boxes indi- cate sites of hypomethylation (at least 50% protection) and closed boxes sites of hypermethylation (at least twofold increase). Numbers indicate the positions of the G residues relative to the transcription start site. (T) G ladder obtained from Me,SO,-treated naked chromosomal DNA. F(7), G ladder obtained from explants that had been exposed to bFGF for 7 days.
the first intron able to overcome the silencing effect of the up- stream region, indicating that the anti-repressor activity is rela- tively independent of position but not of orientation (Fig. 1 OB).
As shown in Fig. 10A, the activity of the -77/+1000 frag- ment is threefold lower than the activity of the -77/+20 pro- moter fragment, indicating that, in the absence of the upstream region, the first intron can also act as a silencer of the pS2- crystallin promoter. To test the possibility that the first intron contains a transcriptional silencer directly, we placed the +20/ +lo00 fragment in two orientations in front of the HSV TK promoter and tested its activity in RNR cells. The first intron decreases activity of the TK promoter by more than 30-fold when present in the sense orientation and by 10-fold when in the antisense orientation, thus indicating that the +20/+ 1000 fragment contains a highly potent transcriptional silencer (Fig. 10C).
Activity of the fl2-crystallin promoter in non-lens cells. It has been reported that the /332-crystallin protein is expressed at low levels in the chicken retina (Head et al., 1991). To examine whether the transcription regulatory elements that are active i n differentiating rat lens explants could also play a role in retinal expression, the same four /3Ei2-crystallinlCAT gene constructs as shown in Fig. 2 were tested in RNR cells. Mouse Ltk fibro-
-77/*20 -38/+20 -38/+10 pEUCAT
CAT construct
Fig. 7. Deletion mutant analysis of the rat /B2-crystallin promoter. Newborn rat lens explants were exposed to bFGF for 7 days and then transfected with the indicated jB2-crystallinlCAT constructs (see also Fig. 4) or with the promoterless CAT gene vector (pEUCAT). Three days later explants were harvested and the CAT activities were determined as described in Materials and Methods. Promoter activities are average val- ues froin three transfections and are indicated relative to the activity obtained from the -77/+20 /332-crystallin promoter. SD are represented by error bars.
0 0 0 0 - _ - -122/-77 competitor - - - - - - - - - - % x
0 - 0 - - -77/+20 competitor - - - - - - - - - - - extract explant lens
' 0 4 5 6 7 8 9 1 2 '
*
-122/-77 probe
Fig.8. A nuclear factor binding to the jIBZ-crystallin upstream si- lencing region is present in rat lens explants at all stages of differen- tiation. "P-labelled - 122/-77 jB2-crystallin promoter region was used as a probe in a gel shift assay with whole cell extracts of explants har- vested after various days of exposure to bFGF (as indicated) or with newborn rat lens nuclear extract and either in the absence or presence of 100-300-fold molar excess of unlabelled -122/-77 or -77/+20 rn2-crystallin fragment.
blasts, which do not detectably express /332-crystallin mRNA, were included as a negative control. Essentially the same pattern of reporter gene activities as described for the explants, was found in the RNR cells and, surprisingly, in the mouse Ltk- fibroblasts (Fig. 11). In both cell types the activity of the -77/ +20 promoter fragment is repressed by a negative regulatory element located between -77 and -121 bp, and is partially re- stored by an element in intron 1. However, the activity of the -750/+1000 construct relative to the activity of the -77/+20 construct is lower in RNR and Ltk- cells while it is higher in differentiating lens explants. In conclusion, the regulatory ele- ments identified thus far may not only play a role in determining the transient expression of the m2-ci-ystallin gene in differenti-

30 Dirks et al. (EUK J . Biochem. 23Y)
construct CAT activity (X) A construct CAT activity (X)
100 f 32
25 * 1
iqm 40 5 7 -123 -77
-77 -123
Fig. 9. The /?BZ-crysfallin upstream silencing region downregulates a heterologous promoter. The -123/-77 /{B2-crystallin fragment was placed in either orientation in front of the HSV TK promoter and its activity was tested by transient reporter gene analysis in RNR cells as described in Materials and Methods. CAT activity is expressed relative to the activity obtaine4d from the HSV TK construct (100%).
ating lens cells, but also contribute to the high expression level of the gene in lens cells compared with non-lens cells.
DISCUSSION
The cooperation between the first intron and the -750/ - 123 region of the /B2-crystallin gene plays a key role in the transient activation of the gene during bFGF-mediated differen- tiation of explanted newborn rat lens epithelial cells. Either re- gion has the unusual property that it combines two opposite tran- scription regulatory activities. Separately, the first intron and the -750/-123 region act as a silencer of the /B2-crystallin promoter (Fig. 12A), yet, surprisingly, when the -750/- 123 re- gion and the first intron are combined, an enhancer is created (Fig. 12B). The anti-silencer activity of the intron could be ex- plained by anti-termination of RNA polymerase 11; however, our observation that the intron has the same activity when placed in front of the -750/-t20 region, indicates that it acts at the DNA level. Unexpectedly, the intron activity is dependent on orienta- tion, possibly because the antisense orientation does not favour the interaction with the -750/- 123 region. The lens-specific expression of the chicken d1-crystallin gene also results from a complex interplay between overlapping positive and negative transcription regulatory elements located within the third intron of the gene, which together form a lens-specific enhancer (Kamachi and Konidoh, 1993). Whether the silencer and anti- silencer activities of the intron of the /?B2-crystallin gene co- localize remains to be established.
According to our bandshift assays, a factor that interacts irz vitro with nucleotides -61/-60 is present throughout differenti- ation from epithelial cells onwards. The -61/-60 dinucleotide is part of a sequence that closely resembles the consensus bind- ing site for Pax-6, the regulator of eye development. A synthetic oligonucleotide containing the Pax-6 binding site of the aA-crys- tallin promoter cornpetes with the rat /jB2-crystallin promoter for binding to the nuclear lens factor. In addition, anti-Pax-6 antibodies affect the mobility of the same DNA-protein complex in a gel shift assay. Although these observations all support a role for Pax-6 in regulating /iB2-crystallin promoter activity, several other data do not. First, in vivo Me,SO, footprint analysis did not reveal DNA-protein interactions at nucleotides -61/ -60 of the endogenous [lB2-crystallin promoter at any stage of the differentiation process. This is not likely due to the experi- mental system, as our recently published irz viiw yD-crystallin promoter footprints indicate that Me2S0, footprinting is possible in newborn rat lens explants (Dirks et al., 1996). Second, site- directed mutageneais of nucleotides -61/-60 completely abo- lishes the capacity of the /?B2-crystallin promoter to bind to the Pax-6-like factor i r i vitro, but does not affect its activity in a
l o o * 1 -750 t1000
ElESa-- 22 i 4 -- 1 1 f 1
I-- 13 i 7
-123 t 1000
-77 tl000
tZO t1000
WCUI 32 f 3 -77 t20
l o o * 1 -750 t1000
B
-750 t20 1 9 i 0
1105 1 t20
w 4 t20 +lOOO -750
1 6 i 4 120
m4 +lo00 t20 -750
1 2 + 1
C 100 * 32
t- 3 * 0
m- 10 f 1
t20 t1000
t1000 t20
Fig. 10. Context-dependent silencer or antirepressor activity of the first intron of the rat j?BZ-crystaIlin gene. (A) The presence of the -750/-123 region determines whether the first intron has silencer or antirepressor activity. Rat lens explants were exposed to bFGF for 7 days and then transfected with the indicated ~B2-crystallin deletion con- structs. Three days later explants were harvested and the CAT activities were determined as described in Materials and Methods. Promoter activ- ities are average values from two transfections and are indicated relative to the activity obtained from the -750/+1000 construct. (B) The antire- pressor activity of /IB2-crystallin intron 1 is position-independent but orientation-dependent. ,!B2-crystallin intron 1 was placed in either orien- tation in front of the -750/+20 region and the resulting CAT constructs were transfected into newborn rat lens explants that had been exposed to bFGF for 7 days. Three days later explants were harvested and the CAT activities were determined as described in Materials and Methods. CAT activity is the average value from two transfections and is ex- pressed relative to the activity obtained From the -750/+1000 construct (100%). (C) The first intron of the rat bB2-crystallin gene silences a heterologous promoter. PB2-crystallin intron 1 was placed in either ori- entation in front of the HSV TK promoter and its activity was tested by transient reporter gene analysis in RNR cells as described in Materials and Methods. CAT activity is the average value from two transfections and is expressed relative to the activity obtained from the HSV TK con- struct (100%).
reporter gene assay. Thirdly, activity of the fl2-crystallin pro- moter does not increase upon cotransfection of a Pax-6 expres- sion construct, either in lens explants or i n rat fibroblasts (our unpublished data). Recent in vifro footprint data showed that the -XO/-40 region of the mouse [lB2-crystallin promoter may contain a binding site for Pax-6-5a. a splice variant of Pax-6 (Chambers et al., 1995). Comparison of the [jB2-crystallin pro- moter sequences of rat and mouse (data not shown) indicated that the first 330 nucleotides upstream of the transcription start site are 93 % identical. For a promoter region, this is a strikingly high level of evolutionary conservation. However, the sequence located between -44 and -73 is only 74% identical between both species, in accordance with our observation that this region is not important for activity of the pB2-crystallin promoter in

Dirks et al. (Eur: J. Riochcm. 239) 31
CAT activity (%)
construct RNR Ltk-
100 100
[ h3 CAT ] 18 13 -77 +20
-123 +20
25 1 6 -750 t20
71 67 -750 t 1000
Fig. 11. Extralenticular activity of the rat /3B2-crystallin promoter. The same constructs as depicted in Fig. 2 were transiently transfected into RNR and Ltk- cells. CAT activity is indicated relative to the activity obtained from the -77/+20 construct (100%).
-750 -123 -77+20 t 1000
-750 -123 -77 t20
Fig. 12. Model for the regulation of the differentiation stage-specific activity of the rat /3BZ-crystallin gene during in vitro differentiation of explanted newborn rat lens epithelial cells. (A) The activator of the promoter is constitutively repressed by two silencer elements located upstream of the promoter. (B) The differentiation stage-specific activity of an anti-silencer located within the first intron not only counteracts both upstream silencers, but also creates an enhancer for the /SB2-crystal- lin promoter.
vivo. Our combined gel shift and reporter gene analyses also did not support an activating role for the serum-response-factor-like sequence at -621-53 or the Egr/WTI-like sequence at + l o / +20, but rather indicated that the activator of the P2-crystallin promoter recognizes an element located between -37 and +lo. Comparison of the -37/+ 10 sequence with transcription regula- tory elements that have earlier been found important for activity of the chicken P1-crystallin promoter (Roth et al., 1991) or of other crystallin promoters (reviewed in Piatigorsky and Zelenka, 1992) did not reveal any significant similarities. Although Pax-6 does not activate the rat pB2-crystallin promoter, it does bind to and activate the promoters of the chicken (Cvekl et al., 1994) and mouse (Cvekl et al., 1995a) crA-crystallin genes, the enhancer of the chicken 0'1-crystallin gene (Cvekl et al., 199513) and the promoter of the guinea pig c-crystallin gene (Richardson et al., 1995). We are currently examining the possibility that Pax-6 plays an indirect regulatory role in the formation of the promoter-enhancer complex of the m2-crystallin gene. Re- cently, Sox-2 was identified as a transcriptional activator of the chicken 61 -crystallin enhancer and of the mouse yF-crystallin promoter (Kamachi et al., 1995). Whether Sox-2 is also involved in regulation of P-crystallins remains to be determined.
The observation that the rat m2-crystallin promoter is quite active in at least some non-lens cells, such as fibroblasts or ret- ina cells, is consistent with an earlier report that the chicken PI-crystal l in promoter is active in fibroblasts, albeit much less than in primary lens epithelial cells (Roth et al., 1991). In fact,
the DNA elements that regulate the activity of the rat P 2 - c r y s - tallin promoter have essentially the same properties in differenti- ating explants as in fibroblasts and retina cells. The major differ- ence is that, in the latter two cell types, the first intron only partially relieves the activity of the upstream silencers, whereas in lens cells it is involved in the formation of an enhancer. The enhancer that is created by the cooperation between the -7501 -123 region and the first intron may thus contribute to the highly increased activity of the PB2-crystallin gene in lens cells compared with the activity in non-lens cells. There is accumulat- ing evidence that members of the Ply-crystallin superfamily are also expressed extralenticularly. Activity of the /332-crystallin promoter i n RNR cells is consistent with the previously de- scribed expression of j7l32-crystallin protein in the chicken retina (Head et al., 1991).
We have previously shown that the crystallin genes are only active during a restricted stage of fibre cell differentiation (Peek et al., 1992b). Recently, we identified the regulatory events that determine the differentiation stage-specific expression of the rat yD-crystallin gene (Dirks et al., 1996). The experiments pre- sented here begin to unravel the complex mechanism that under- lies the differentiation stage specificity of the /ll32-crystallin gene. A second variable in the control of crystallin promoter activity is the developmental age of the lens cells. In the future we hope to dissect the developmental control elements in vitro by using explants isolated from older animals (Lovicu and McAvoy, 1992; Peek et al., 1992b).
We thank Bart Aarts for his assistance in constructing /332-crystallin/ CAT reporter plasmids, Dr Henk Aarts for mapping the transcription start site of the pB2-crystallin gene and Drs R. Clayton and P. Russell for communicating their results prior to publication. We are grateful to Dr Maas for providing the Pax-6 expression constructs and to Dr Saule for providing anti-Pax-6 antibody. This work has been carried out under the auspices of the Netherlands Foundation for Chemical Research (SON) and with financial aid from the Netherlands Organization for the Advancement of Pure Research (NWO); it was also supported by a grant (1R01 EY09187) from the National Institutes of Health and a Science Twinning grant (ERBSCI TT920804) from the EC.
REFERENCES Aarts, H. J. M. (1 989) The /?-&crystiiIliu multi-gene family: expression
cind evolution, PhD Thesis, University of Nijmegen. Aarts, H. J . M., Den Dunnen, J. T., Lubsen, N. H. & Schoenmakers, J.
G. G. (1987) Linkage between the /332 and j'B3 crystallin genes in man and rat: a remnant of an ancient /krystallin gene cluster, Gene 59, 127-13.5.
Aarts, H. J. M., Lubsen, N. H. & Schoenmakers, J. G. G. (1989) Crystal- lin gene expression during rat lens development, Eur: J. Rioclzcm.
Ausubel, F. M., Brent, R., Kingston, R. E., Moore, D. D., Seidman, J . G., Smith, J . A. & Struhl, K. (ed.) (1987) C~irrentprotoc,ol.s in molecular biology. Greene Publishing Associates and John Wiley & Sons, New York.
Bloemendal, H. & De Jong, W. W. (1991) Lens proteins and their genes, Pmg. Nucleic Acids Rex Mol. B i d . 41, 259-281.
Carriere, C., Plaza, S., Martin, P., Quatannens, B., Bailly, M., Stehelin, D. & Saule, S. (1993) Characterization of the quail Pax-6 (Pax-QNR) proteins expressed in the neuroretina, Mol. Cell. B i d . 13. 72.57- 1266.
Chamberlain, C. G. & McAvoy, J. W. (1989) lnduction of lens fibre differentiation by acidic and basic fibroblast growth factor (FGF), Growllz Factor.v I , 12.5-134.
Chambers, C., Cvekl, A., Sax, C. M. & Russell. P. (199.5) Sequence, initial functional analysis and protein-DNA binding sites of the mouse /332-crystallin-encoding gene, Gene 166, 287-292.
Cvekl, A,, Kashanchi, F., Sax, C. M., Brady, J. N. & Piatigorsky, J. (1995a) Transcriptional regulation of the mouse nA-crystallin gene:
183, 31 -36.

32 Dirks et al. (Eur: J . Biochem. 239)
activation dependNent on a cyclic AMP-responsive element (DEIl CRE) and a Pax-6,-binding site, Mol. Cell. B id . 15, 653-660.
Cvekl, A,, Sax, C. M., Bresnick, E. H. & Piatigorsky, J. (1994) A com- plex array of positive and negative elements regulates the chicken nA-crystallin gene: involvement of Pax-6, USF, CREB andlor CREM, and AP-1 proteins, Mol. Cell. Biol. 14, 7363-7376.
Cvekl, A,, Sax, C. M., Li, X., McDermott, J. B. & Piatigorsky, J . (1995bj Pax-6 and lens-specific transcription of the chicken n’l -crystallin gene, Proc. Natl Acud. Sci. USA 92, 4681 -4685.
De Jong, W. W., Hendriks, W., Mulders, J. W. M. & Bloemendal, H. (1989) Evolution of eye lens crystallins: the stress connection, Trends Biochem. Sci. 14, 365-368.
De Jong, W. W., Lubien, N. H. & Kraft, H. J. (1994) Molecular evolu- tion of the eye lens, Prvgr: Ret. Eye Res. 13, 391-442.
Dirks, R. P. H., Klok, E. J., Van Genesen, S. T., Schoenmakers, J. G. G. & Lubsen, N. H. (1996) The sequence of regulatory events con- trolling the expression of the yD-crystallin gene during fibroblast growth factor mediated rat lens fiber cell differentiation, Dev. Biol. 173, 14-25.
Epstein, J., Cai, J., Glaser, T., Jepeal, L. & Maas, R. (1994a) Identifica- tion of a Pax paired domain recognition sequence and evidence for DNA-dependent conformational changes, J . Bid . Chern. 269, 8355 - 8361.
Epstein, J . A., Glaser, T., Cai, J., Jepeal, L., Walton, D. S. & Maas, R. L. (1994b) Two independent and interactive DNA-binding subdo- mains of the Pax6 paired domain are regulated by alternative splic- ing, Genes & Deli. 8, 2022-2034.
Faisst, S. & Meyer, !$. (1992) Compilation of vertebrate-encoded tran- scription factors, .Vucleic Acids Res. 20, 3-26.
Garrity, P. A. & Wold, B. J. (1992) Effects of different DNA polymerases in ligation-mediated PCR : enhanced genomic sequencing and in vivo footprinting, Proc. Nut1 Acnd. Sci. USA 89, 1021 - 1025.
Geliebter, J., Zeff, R. A., Melvold, R. W. & Nathenson, S. G. (1986) Mitotic recombination in germ cells generated two major histocom- patibility complex. mutated genes shown to be identical by RNA se- quence analysis: Kh”” and Kh”’”, Proc. Natl Acad. Sci. USA 83,
Goring, D. R., Breitman, M. L. & Tsui, L.-C. (1 992) Temporal regulation of six y-crystallin transcripts during mouse lens development, Exp. Eye Res. 54, 785~r795.
Gorman, C., Moffat, L. & Howard, B. H. (1982) Recombinant genomes which express chloramphenicol acetyltransferase in mammalian cells, Mol. Cell. tliol. 2, 1044-1054.
Hanson, 1. & Van He:yningen, V. (1995) Pax6: more than meets the eye, Trenh Genet. 11, 268-272.
Head, M. W., Peter, A. & Clayton, R. M. (1991) Evidence for the extra- lenticular expression of members of the p-crystallin gene family in the chick and a comparison with Scrystallin during differentiation and traiisdifferentiation, D#erentiution 48, 147- 156.
Hejtmancik, J. F., Beebe, D. C., Ostrer, H. & Piatigorsky, J. (1985) 6- and p-crystallin inRNA levels in the embryonic and posthatched chicken lens : temporal and spatial changes during development, Dei: Biol. 109, 72-81.
Jain, V. K. & Magratl-i, I. T. (1991) Chemiluminiscent assay for the quan- titation of /,’-gala,ctosidase in the femtogram range : application to the quantitation of 0-galactosidase in lncZ-transfected cells, Anal. Bioclienz. 199, 1 19 - 124.
Kamachi, Y. & Kondoh, H. (1993) Overlapping positive and negative regulatory elements determine lens-specific activity of the 61-crystal- lin enhancer, Mol. Cell. Biol. 13, 5206-5215.
Kamachi, Y., Sockanathan, S., Liu, Q., Breitman, M., Lovell-Badge, R. & Kondoh, H. (1995) Involvement of SOX proteins in lens-spe- cific activation of crystallin genes EMBO J. 14, 3510-3519.
3371 -3375.
Lovicu, F. J. & McAvoy, J. W. (1992) The age of rats affects the response of lens epithelial explants to fibroblast growth factor, Invest. Oph- thalmol. Vis. Sci. 33, 2269-2278.
Luckow, B. & Schiitz, G. (1987) CAT constructions with multiple unique restriction sites for the functional analysis of eukaryotic promoters and regulatory elements, Nucleic Acids Res. 15, 5490.
Maxam, A. & Gilbert, W. (1980) Sequencing end-labeled DNA with base specific chemical cleavages, Methods Enzymol. 65, 499 -560.
McAvoy, J . W. (1978a) Cell division, cell elongation and distribution of a-, p- and y-crystallins in the rat lens, .I. Ernbryol. Exp. Morplz. 44,
McAvoy, J. W. (1978b) Cell division, cell elongation, and the co-ordina- tion of crystallin gene expression during lens morphogenesis in the rat, J. Embryo/. Exp. Morph. 45, 271 -281.
McAvoy, J. W. & Chamberlain, C. G. (1989) Fibroblast growth factor (FGF) induces different responses in lens epithelial cells depending on its concentration, Development 107, 221 -228.
Mueller, P. R. & Wold, B. (1989) In vivo footprinting of a muscle spe- cific enhancer by ligation mediated PCR, Science 246, 780-786.
Peek, R., Kraft, H. J., Klok, E. J., Lubsen, N. H. & Schoenmakers, J. G. G. (1992a) Activation and repression sequences determine the lens- specific expression of the rat ;Ocrystallin gene, Nucleic Acids Res.
Peek, R., McAvoy, J. W., Lubsen, N. H. & Schoenmakers, J. G. G. (1992bj Rise and fall of crystallin gene messenger levels during fi- broblast growth factor induced terminal differentiation of lens cells, Deli. Biol. 152, 152-160.
Piaggio, G. & DeSimone, V. (1991) A new expression vector to study weak promoters, Foctts 12, 85-86.
Piatigorsky, J. & Wistow, G. J. (1989) Enzymekrystallins: gene sharing as an evolutionary strategy, Cell 57, 197-199.
Piatigorsky, J. & Wistow, G. (1991) The recruitment of crystallins: new functions precede gene duplication, Science 25, 1078 - 1079.
Piatigorsky, J. & Zelenka, P. S. (1992) Transcription regulation of crys- tallin genes: cis elements, trans-factors, and signal transduction sys- tems in the lens, Adv. Dev. Biochem. I , 211 -2.56.
Ray, D., Bosselut, R., Ghysdael, J., Mattei, M.-G., Tavitian, A. & Mo- reau Gachelin, F. (1992) Characterization of Spi-B, a transcription factor related to the putative oncoprotein Spi-1lPU.1, Mol. Cell. Biol. 12, 4297-4304.
Richardson, J., Cvekl, A. & Wistow. G. (1995) Pax-6 is essential for lens-specific expression of (-crystallin, Proc. Nut/ Acud. Sci. USA
Ruth, H. .I., Das, G. C. & Piatigorsky, J. (1991) Chicken /IBI-crystallin gene expression: presence of conserved functional pol yomavirus en- hancer-like and octamer binding-like promoter elements found in non-lens genes, Mul. Cell. Bid . 11, 1488-1499.
Sambrook, J., Fritsch, E. F. & Maniatis, T. (1989) Molectilur cloning: a laboratory mcnzuul, 2nd edn, Cold Spring Harbor Laboratory Press, Cold Spring Harbor NY.
Sanger, F., Nicklen, S. & Coulson, A. R. (1977) DNA sequencing with chain-termination inhibitors, Proc. Nut1 Acad. Sci. USA 74, 5463 - 5467.
Smolich, B. D., Tarkington, S. K., Saha, M. S. & Grainger, R. M. (1994) Xenopus y-crystallin gene expression: evidence that the y-crystallin gene family is transcribed in lens and nonlens tissues, Mol. Cell. Biol. 14, 1355-1363.
Van Leen, R. W., Van Roozendaal, K. E. P.. Lubsen, N. H. & Schoen- makers, J. Ci. G. (1987) Differential expression of crystallin genes during development of the rat eye lens, Dev. Biol. 120, 457-464.
Wistow, G. & Piatigorsky, J. (1987) Recruitment of enzymes as lens structural proteins, Science 236, 1554- 1556.
149 - 165.
20, 4865-4871.
92, 4676-4680.












![Characterization of an antibody that recognizes peptides ... · in αA-crystallin (Asp 58 and Asp 151) [3], αB-crystallin (Asp 36 and Asp 62) [4], and βB2-crsytallin (Asp 4) [5]](https://static.fdocument.pub/doc/165x107/5ff1e68e89243b57b64135f8/characterization-of-an-antibody-that-recognizes-peptides-in-a-crystallin-asp.jpg)





