TECNOLOGIA DEL DNA RICOMBINANTE · di appaiarsi l’una con l’ altra. ... Cellula vegetale...
75
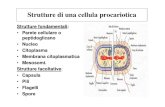
TECNOLOGIA DEL DNA RICOMBINANTE Vettore (plasmidi) DNA sorgente (uno o più frammenti) Taglio (enzimi di restrizione) Unione (DNA ligasi) Ricombinazione Trasformazione Espressione (omologa o eterologa) Clonaggio (amplificazione)
Transcript of TECNOLOGIA DEL DNA RICOMBINANTE · di appaiarsi l’una con l’ altra. ... Cellula vegetale...
TECNOLOGIA DEL DNA RICOMBINANTE
Vettore (plasmidi)
DNA sorgente (uno o più frammenti)
Taglio (enzimi di restrizione)
Unione (DNA ligasi)
Ricombinazione
Trasformazione
Espressione (omologa o eterologa)
Clonaggio (amplificazione)

Alcuni batteri hanno molecole di DNA circolari extracromosomico dette plasmidi che si duplicano in maniera indipendente rispetto al cromosoma batterico e producono proteine sfruttando i meccanismi biosintetici della cellula. E’ possibile usare i plasmidi come vettori per introdurre uno specifico gene che codifica per la proteina desiderata. Per questa operazione è necessario conoscere le sequenze nucleotidiche dei segmenti da clonare, e manipolare il DNA plasmidico tale da renderlo idoneo per l’acquisizione dell’inserto. Il DNA viene quindi estratto, digerito e poi inserito nel vettore plasmidico. Questo nuovo DNA, detto ricombinante perché ottenuto attraverso tecniche di ricombinazione in vitro, può essere utilizzato per trasformare una cellula ospite, batterio o un altro microrganismo capace di riprodursi rapidamente e generare un gran numero di copie di DNA ricombinante.
TECNOLOGIA DEL DNA RICOMBINANTE

DIGESTIONE DEL DNA - ENZIMI DI RESTRIZIONE
Vari tipi di enzimi sono in grado di idrolizzare i legami fosfoestere del DNA (nucleasi)
Endonucleasi : tagliano all’interno del filamento di DNA
Esonuclesi: tagliano a partire dalle estremità 5’ o 3’
ENDONUCLEASI DI RESTRIZIONE
Sono enzimi prodotti da batteri che tagliano le molecole di DNA a livello di sequenze nucleotidiche specifiche dette siti di restrizione o di riconoscimento. Scindono specificamente il legame estere tra l’ossigeno del C3’ dello zucchero di un nucleotide e il gruppo fosfato in C5 ‘ dello zucchero del nucleotide successivo.
Varie classi
Classe I e III necessitano di energia e scindono il DNA in modo imprevedibile;
Classe II sono quelle più utili in quanto non richiedono energia e tagliano in siti specifici riconoscendo delle sequenze di 4-8 bp dette palindromiche poiché risultano identiche se lette su entrambi i filamenti.
Il loro taglio può produrre frammenti con estremità piatte o sfalsate (coesive), a seconda del tipo di enzima utilizzato.

Esempi di tagli effettuati da diversi enzimi di restrizione
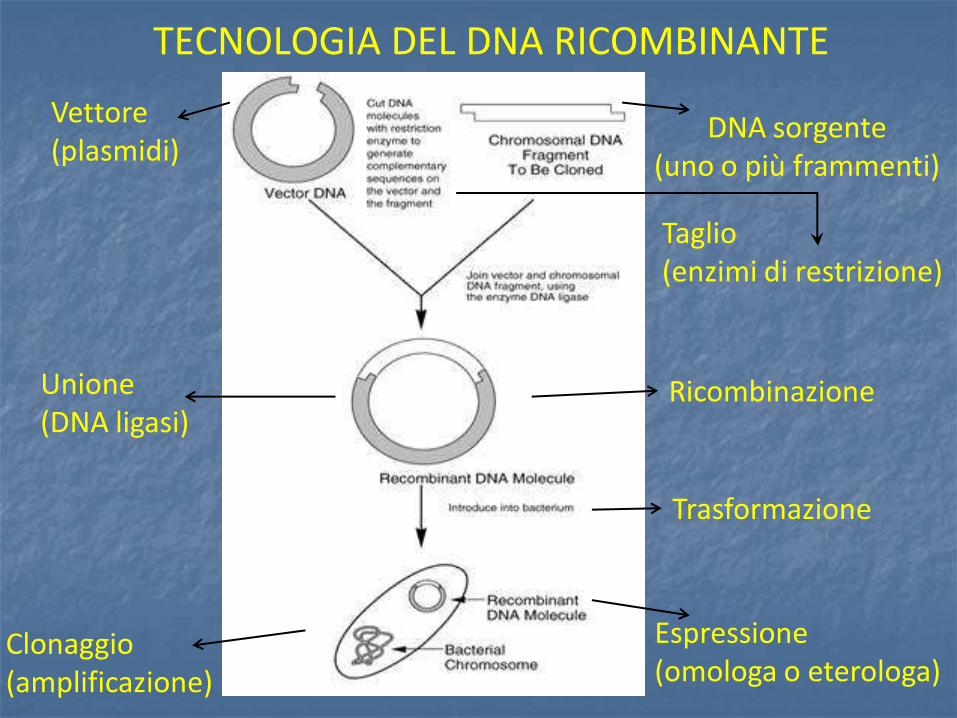
Le estremità coesive prodotte dallo stesso enzima su molecole di DNA diverse sono complementari e quindi in grado di appaiarsi l’una con l’ altra. Le catene associate sono tenute insieme da legami idrogeno tra basi complementari su filamenti opposti ma necessitano di un unione covalente tra il gruppo fosfato 5’ di un’ estremità e il gruppo ossidrilico 3’ della catena adiacente. Tale reazione è catalizzata dall’enzima DNA ligasi.
DNA ligasi

DNA ligasi formazione del legame covalente
tra due molecole di DNA.

Gran parte degli straordinari progressi ottenuti dalla biotecnologia e dalla biologia
molecolare, dipendono dall'acquisizione della capacità di amplificare e propagare
indefinitivamente i geni.
Clonare un gene significa isolarlo da un genoma ed inserirlo in un vettore capace di
replicarsi in un certo ospite (di solito E.coli o lievito).
Esistono diversi tipi di vettori di clonaggio, ciascuno con vantaggi e svantaggi.
La principale considerazione da fare é relativa alle dimensioni dell'inserto di DNA che ogni
vettore può accettare.
PLASMIDI da 0,1 a 10 Kb
FAGI da 8 a 22 kb
COSMIDI da 32 a 45 kb
BAC (CROMOSOMI ARTIFICIALI BATTERICI ) da 75 a 300 kb
YAC (CROMOSOMI ARTIFICIALI DI LIEVITO ) da 100 a 2000 kb
Vettori di clonaggio

I plasmidi sono degli elementi genetici extracromosomali che si replicano autonomamente. Variano da 1 a 200 kb e sono molto diffusi tra i procarioti. Esempi di plasmidi batterici naturali sono i plasmidi ColE1 di E.coli, i plasmidi Sym di rhizobium, I plasmidi Ti o Ri di Agrobacterium.
Plasmide superavvolto (supercoiled)
Plasmide circolare rilassato
Plasmide linearizzato
nick
Nell'ospite batterico i plasmidi si presentano come molecole circolari superavvolte, che, durante le manipolazioni sperimentali, possono rilassarsi o linearizzarsi in seguito a rotture a singolo o a doppio filamento. In un gel di agarosio e bromuro di etidio le tre forme migrano a velocità diverse e possono essere distinte.
I plasmidi possono essere lineari o integrati nel cromosoma batterico ma, nella maggior parte dei casi, sono molecole di DNA circolari (episomi)
PLASMIDI

Dai plasmidi batterici naturali sono derivati i vettori di clonaggio, le cui caratteristiche essenziali sono
• Origine di replicazione
• Marcatore selezionabile
• Siti di restrizione unici
Vettori plasmidici

VETTORI PLASMIDICI
Ori: origine della replicazione
Amp: gene reistenza ampicillina Tet: gene reistenza tetraciclina
Piccoli DNA circolari con capacità di replicazione autonoma (specie specifica) contenenti geni che possono essere utili al batterio (es. resistenza ad antibiotici) Utili se contengono siti unici per enzimi di restrizione

VETTORI PLASMIDICI
Plasmidi artificiali: creati per soddisfare meglio le esigenze di clonaggio
MCS (multiple cloning sequence): breve tratto di DNA contenente diversi siti di restrizione unici e posizionata in un punto specifico del vettore LacZ: gene delle beta-galattosidasi (enzima in grado di convertire X-gal in un composto colorato)
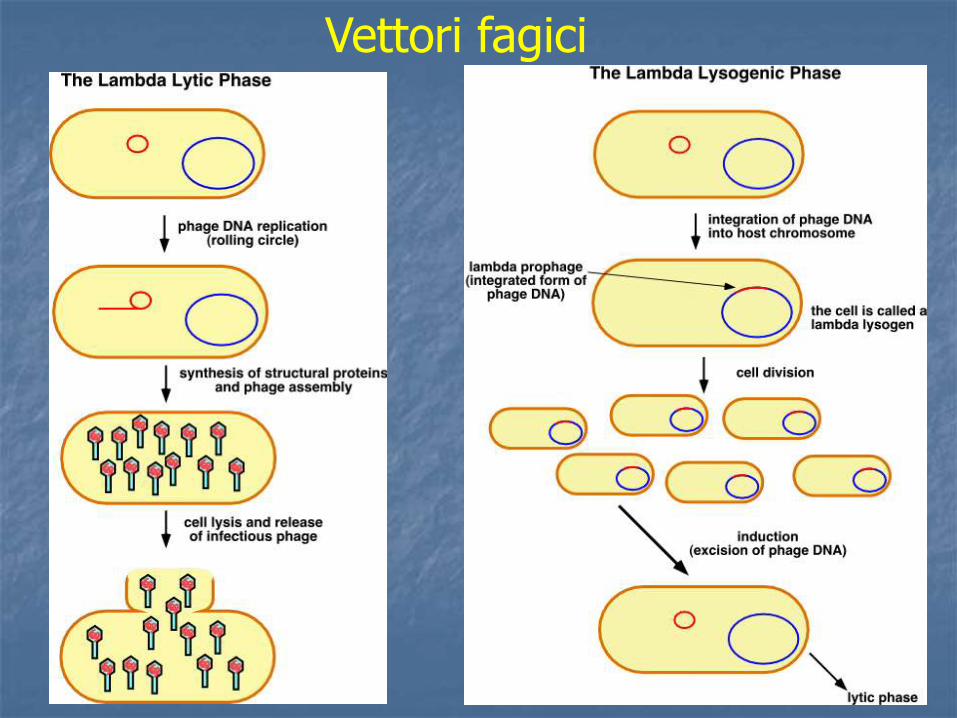
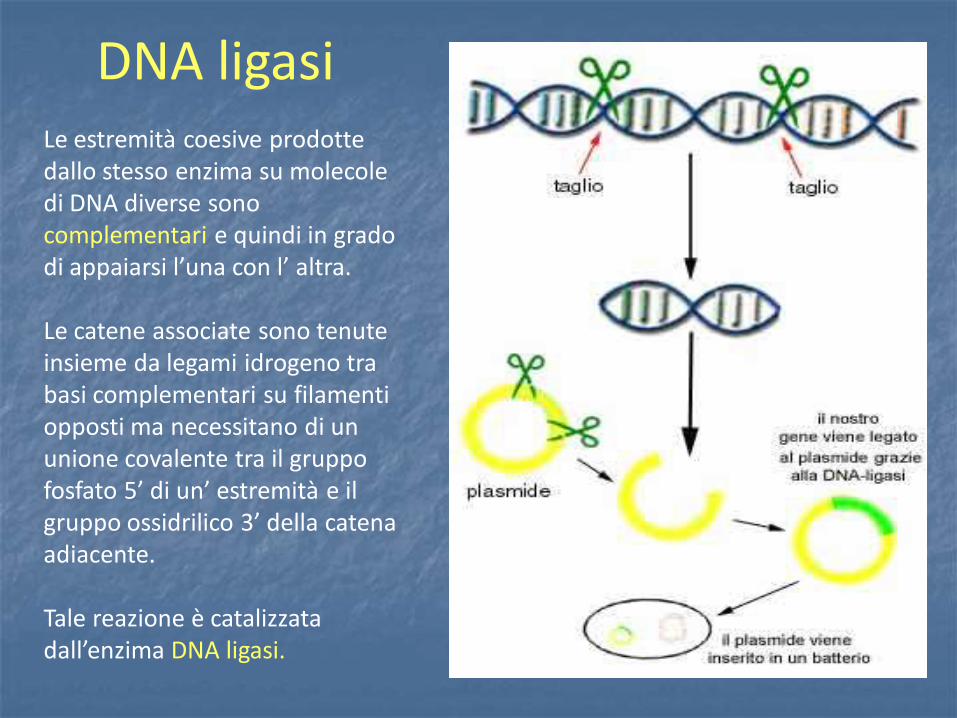
Vettori fagici
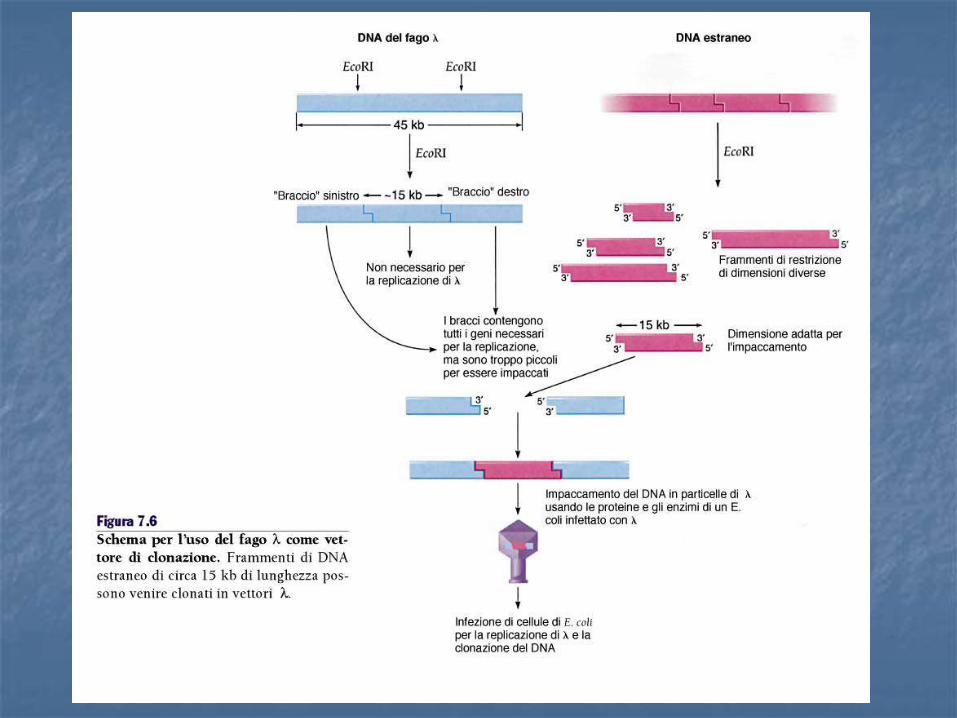

Vettori fagici

Per trasformare la cellula batterica, vista la dimensione del cosmide, si può utilizzare un packaging in vitro analogamente a un fago Il cosmide infatti possiede un sito cos che può essere considerato un buon substrato per una reazione di packaging, purchè abbia dimensione comprese tra 38 e 51 Kb. Un cosmide si clona come un plasmide e si propaga come un fago λ
I COSMIDI I cosmidi permettono di trasportare DNA fino a 45 Kb. Un cosmide è un plasmide, di solito intorno alle 5 Kb, contenente, un sito cos. Contiene una ori, un marcatore di resistenza e siti unici di restrizione (sito di clonaggio) e si può utilizzare come un plasmide.

I COSMIDI
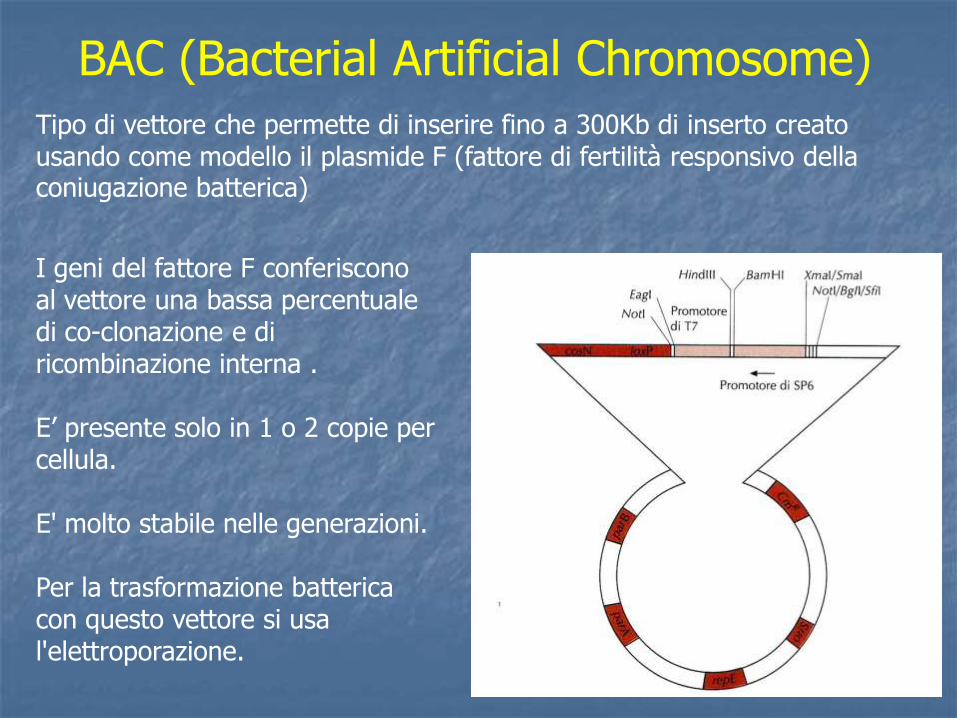
I geni del fattore F conferiscono al vettore una bassa percentuale di co-clonazione e di ricombinazione interna . E’ presente solo in 1 o 2 copie per cellula. E' molto stabile nelle generazioni. Per la trasformazione batterica con questo vettore si usa l'elettroporazione.
BAC (Bacterial Artificial Chromosome)
Tipo di vettore che permette di inserire fino a 300Kb di inserto creato usando come modello il plasmide F (fattore di fertilità responsivo della coniugazione batterica)
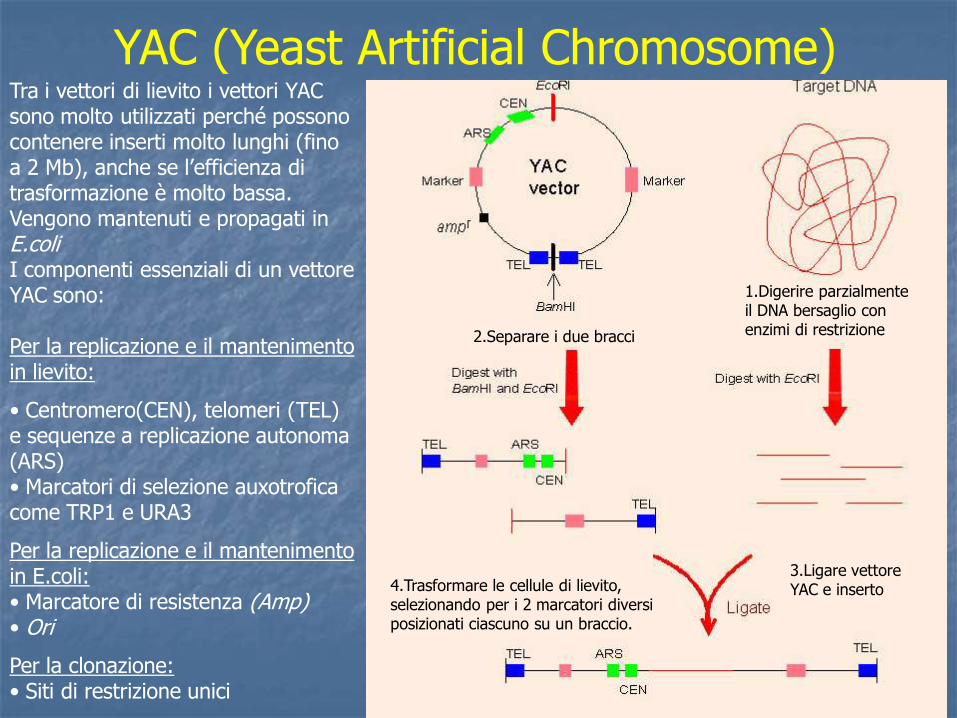
YAC (Yeast Artificial Chromosome) Tra i vettori di lievito i vettori YAC sono molto utilizzati perché possono contenere inserti molto lunghi (fino a 2 Mb), anche se l’efficienza di trasformazione è molto bassa. Vengono mantenuti e propagati in E.coli I componenti essenziali di un vettore YAC sono: Per la replicazione e il mantenimento in lievito:
• Centromero(CEN), telomeri (TEL) e sequenze a replicazione autonoma (ARS) • Marcatori di selezione auxotrofica come TRP1 e URA3
Per la replicazione e il mantenimento in E.coli: • Marcatore di resistenza (Amp) • Ori
Per la clonazione: • Siti di restrizione unici
4.Trasformare le cellule di lievito, selezionando per i 2 marcatori diversi posizionati ciascuno su un braccio.
1.Digerire parzialmente il DNA bersaglio con enzimi di restrizione 2.Separare i due bracci
3.Ligare vettore YAC e inserto
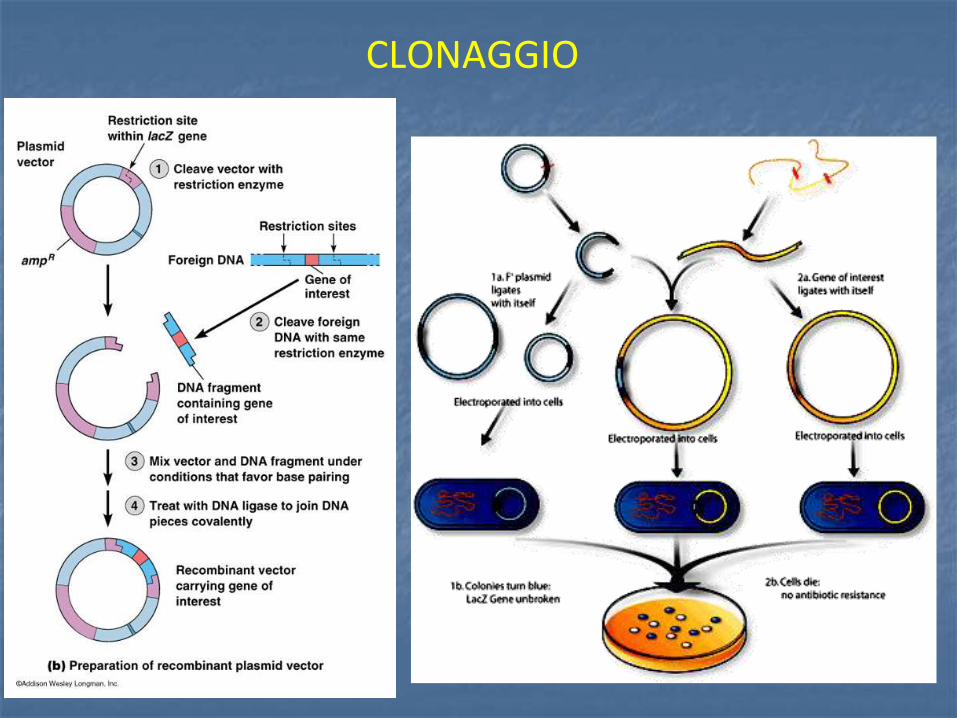
CLONAGGIO

DNA SORGENTE
- Genoma
frammentazione del DNA e inserimento in vettori
frammenti casuali (necessità di analizzare molti cloni)
presenza di sequenze non codificanti
- Trascrittoma (RNA)
necessità di covertire RNA in DNA (cDNA)
non tutti i geni sono espressi nella stessa cellula

Librerie geniche o genoteche
Quando un DNA genomico viene estratto dalle cellule di un organismo, tagliato con uno o più enzimi di enzima di restrizione e la popolazione dei frammenti di DNA ottenuti vengono clonati in un vettore, si ottiene una collezione di cloni, contenente almeno una copia di tutte le sequenze di DNA presenti nel genoma. Tale collezione è definita LIBRERIA GENOMICA (o GENOTECA)
Imparentate alle genoteche genomiche sono le genoteche di DNA complementare (cDNA), collezioni di cloni contenenti molecole di DNA copiate dai mRNA isolati dalle cellule. In tali genoteche è clonato il DNA privo delle regioni introniche e quindi di solo la porzione codificante dei vari geni analizzati.



Sintesi del cDNA


Libreria di cDNA
La libreria di cDNA è cellula specifica

Trasformazione
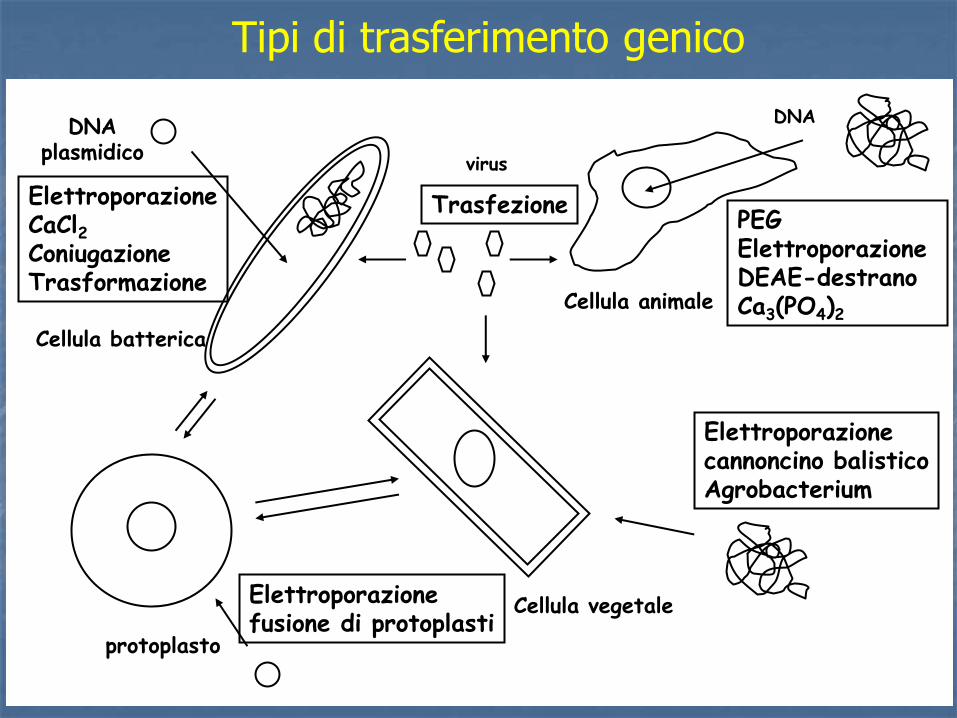
Tipi di trasferimento genico
DNA plasmidico
Elettroporazione CaCl2 Coniugazione Trasformazione
Cellula batterica
Cellula animale
PEG Elettroporazione DEAE-destrano Ca3(PO4)2
virus
Cellula vegetale
Elettroporazione cannoncino balistico Agrobacterium
protoplasto
DNA
Elettroporazione fusione di protoplasti
Trasfezione

Metodi di Trasformazione
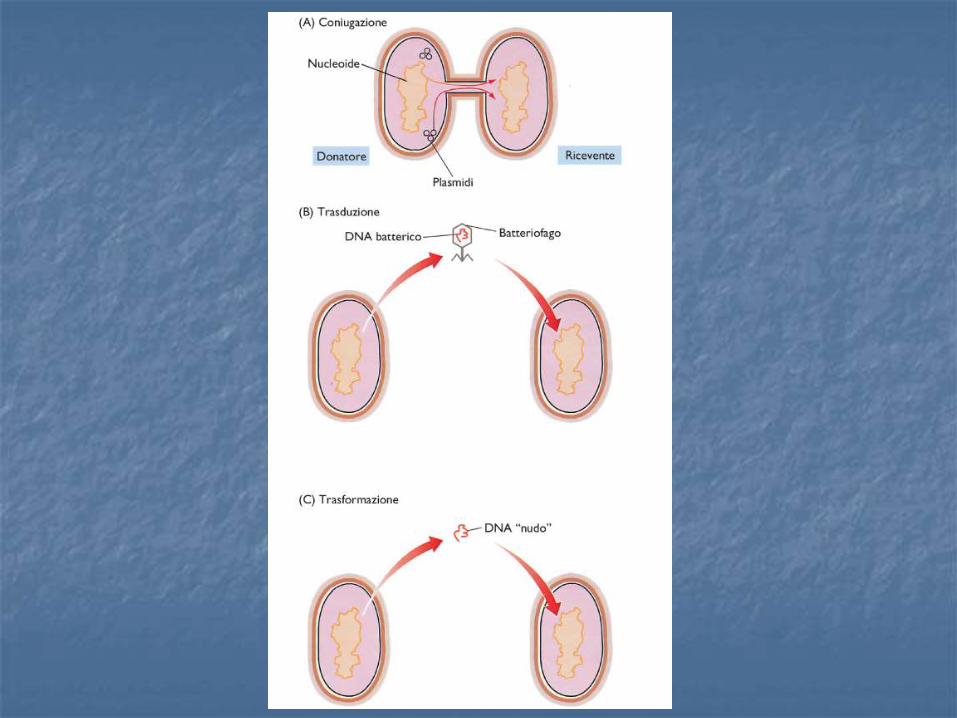
CONIUGAZIONE
Trasferiento mediato da pili che congiungono 2 cellule
CaCl2 - Ca3(PO4)2
I Sali di Calcio creano dei pori nella membrana cellulare attraverso la quale possono transitare le molecole di DNA all’interno della cellula.
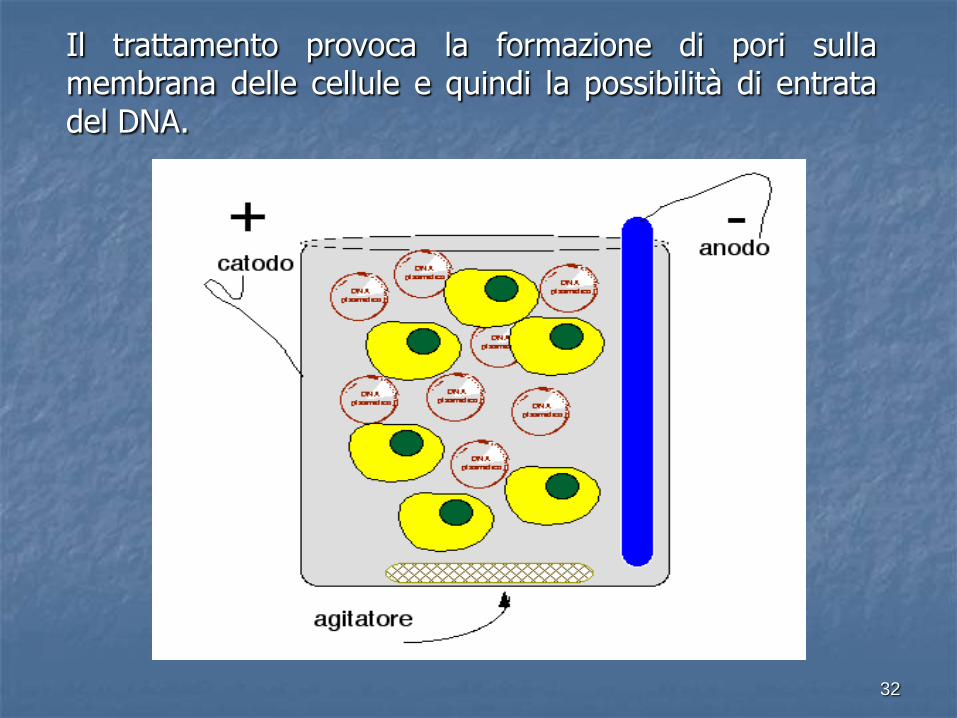
ELETTROPORAZIONE
Metodica che applica un impulso elettrico ad alto voltaggio a una sospensione di cellule in presenza di DNA al fine di far entrare quel DNA nelle cellule.

31
Elettroporazione
L’elettroporazione è una metodica efficiente e praticamente universale.
32
Il trattamento provoca la formazione di pori sulla membrana delle cellule e quindi la possibilità di entrata del DNA.

33
Trasduzione (Vettori virali)
o Elevata efficienza
Si sfruttano le proprietà naturali del virus
o Uso attento
Rischio di creare virus patogeni per ricombinazione o Molti possibili vettori diversi:
Oncoretrovirus, lentivirus, adenovirus, HSV, virus ibridi

34
Ingegnerizzando il DNA contenuto nel capside del fago è possibile ottenere trasformazione della cellula bersaglio. Il sistema è ad elevata efficienza perché non si fa altro che sfruttare, manipolandolo, un processo naturale (la naturale tendenza del virus all’infezione).

SELEZIONE DEI CLONI TRASFORMATI

SELEZIONE DEI CLONI TRASFORMATI

SELEZIONE DEI CLONI TRASFORMATI
Inattivazione inserzionale


SELEZIONE DEI CLONI SPECIFICI
- PCR con primers specifici.
- Ibridazione con sonde a DNA specifiche.
- Tecniche immunologiche
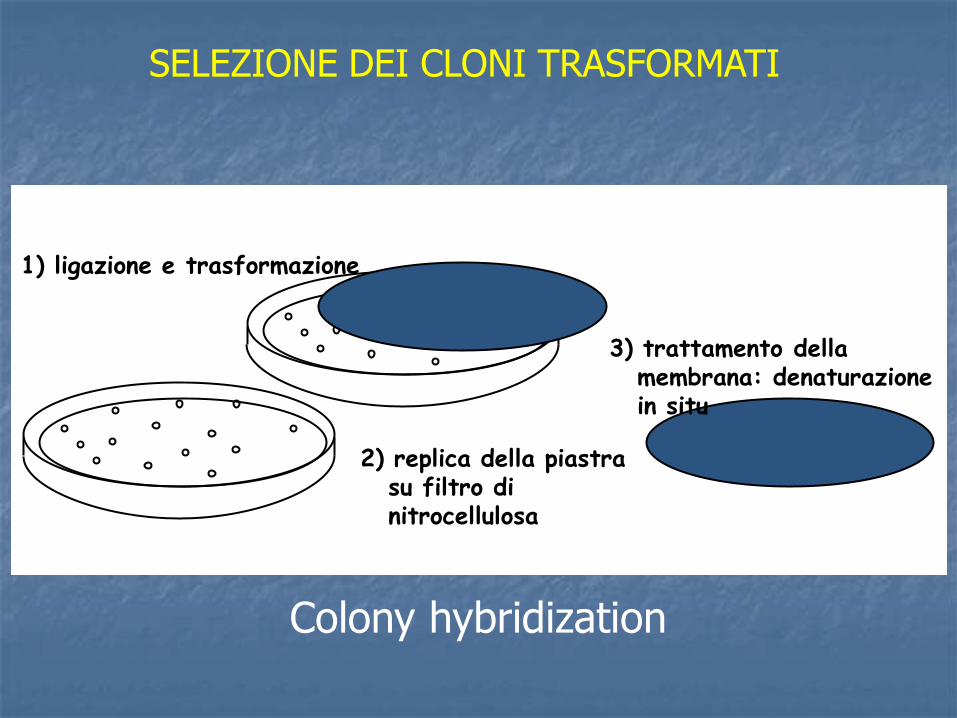
1) ligazione e trasformazione
2) replica della piastra su filtro di nitrocellulosa
3) trattamento della membrana: denaturazione in situ
Colony hybridization
SELEZIONE DEI CLONI TRASFORMATI
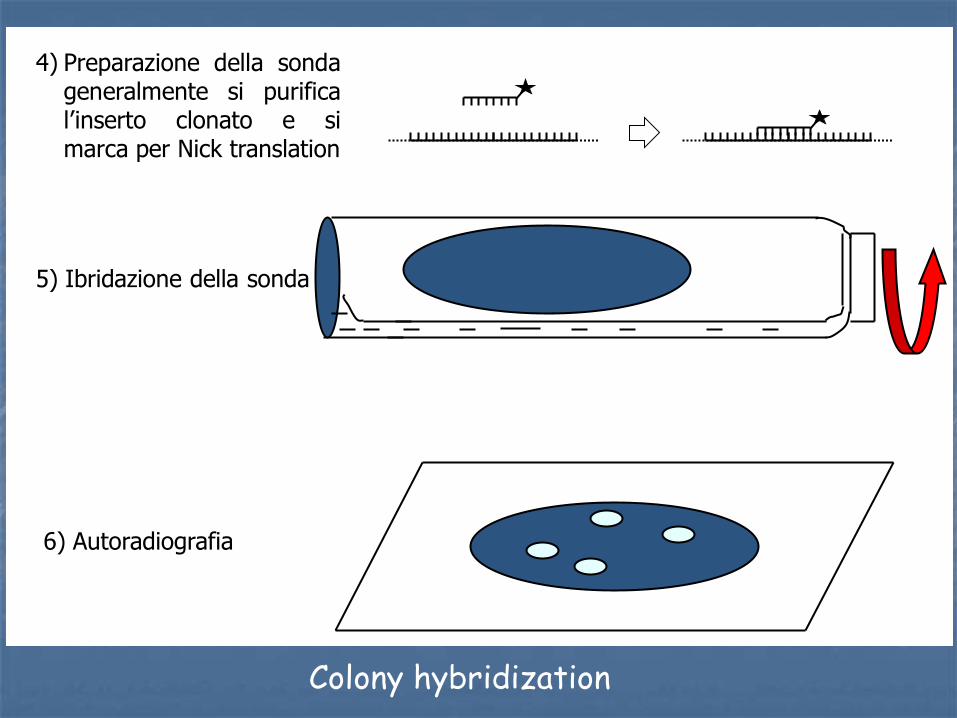
4) Preparazione della sonda generalmente si purifica l’inserto clonato e si marca per Nick translation
5) Ibridazione della sonda
6) Autoradiografia
Colony hybridization
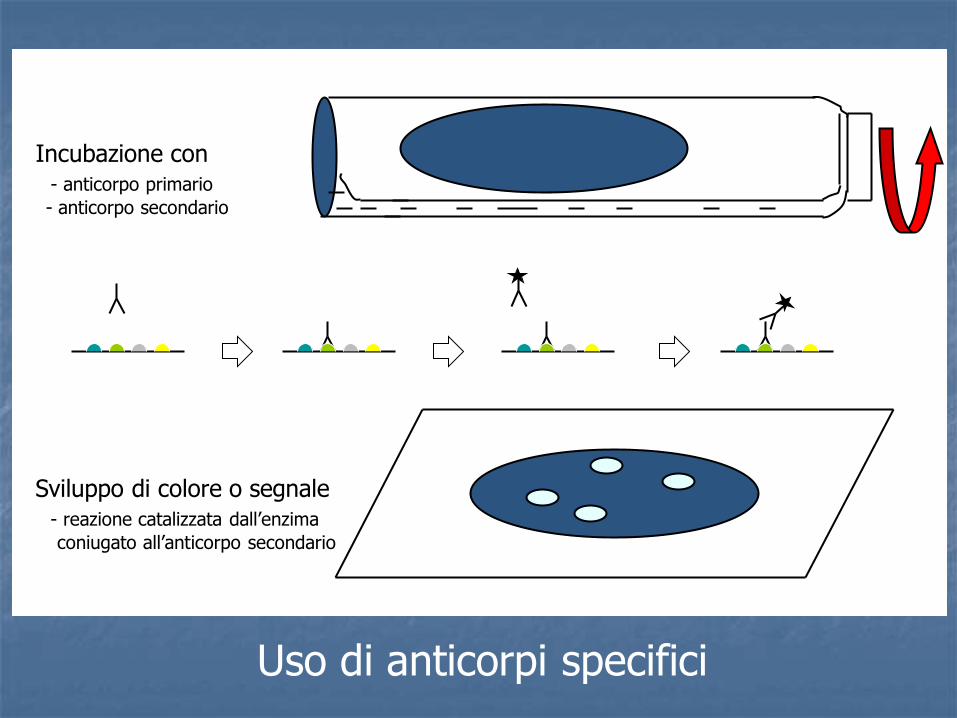
Incubazione con - anticorpo primario
- anticorpo secondario
Sviluppo di colore o segnale - reazione catalizzata dall’enzima
coniugato all’anticorpo secondario
Uso di anticorpi specifici

ESPRESSIONE DI PROTEINE RICOMBINANTI

ESPRESSIONE DI PROTEINE RICOMBINANTI
Clonaggio di un gene o di un cDNA codificante una particolare
proteina
Scelta del tipo di cellula ospite in funzione della proteina da
produrre
Inserimento del gene nella cellula ospite ove si voglia farlo
esprimere. I sistemi di espressione più diffusi sono Escherichia coli,
Bacillus subtilitis, lievito e cellule di mammifero.
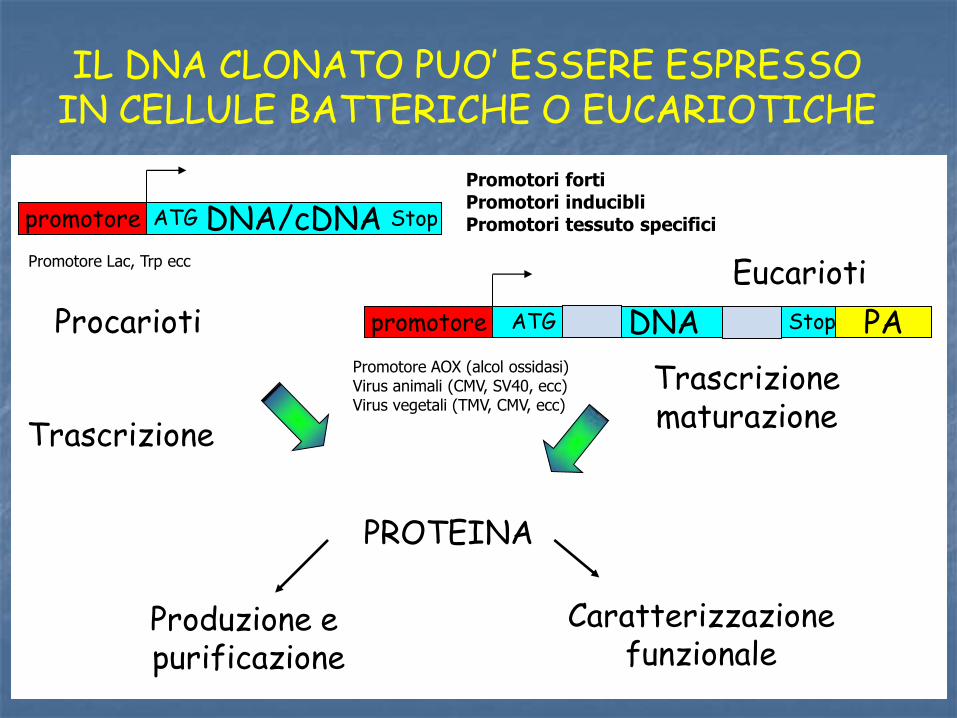
Trascrizione maturazione
IL DNA CLONATO PUO’ ESSERE ESPRESSO IN CELLULE BATTERICHE O EUCARIOTICHE
promotore DNA/cDNA ATG Stop
PROTEINA
Produzione e purificazione
Caratterizzazione funzionale
PA promotore DNA ATG Stop
Trascrizione
Procarioti
Eucarioti Promotore Lac, Trp ecc
Promotore AOX (alcol ossidasi) Virus animali (CMV, SV40, ecc) Virus vegetali (TMV, CMV, ecc)
Promotori forti Promotori inducibli Promotori tessuto specifici

CELLULE BATTERICHE
Vantaggi
Semplicità di manipolazione
Velocità di replicazione
Alta resa di prodotto a basso costo.
Svantaggi
A volte non assumono la conformazione corretta, formano dei corpi
inclusi generando proteine biologicamente inattive
E’necessario un promotore inducibile per attivare la trascrizione
del gene relativo alla proteina desiderata.
Mancano di enzimi che effettuano modificazioni postraduzionali
sulle proteine necessarie affinchè quest’ultime possano funzionare.

LIEVITO
Eucariote unicellulare ma cresce in modo rapido ed economico (batteri). Realizza molte delle modificazioni postraduzionali osservate nelle cellule di mammifero Possiede proteasi attive che degradano le proteine estranee (desiderate) riducendo il recupero quantitativo del prodotto.

MAMMIFERI
Sede di elezione dove produrre una proteina di mammifero. I promotori, i vettori, i protocolli di trasformazione e i sistemi di cellule ospiti sono stati notevolmente migliorati determinando la produzione su ampia scala di proteine quali l’ attivatore del plasminogeno tissutale (tPA). Vengono usate spesso per testare l’ attività di un gene appena clonato o la funzionalità di una proteina modificata.
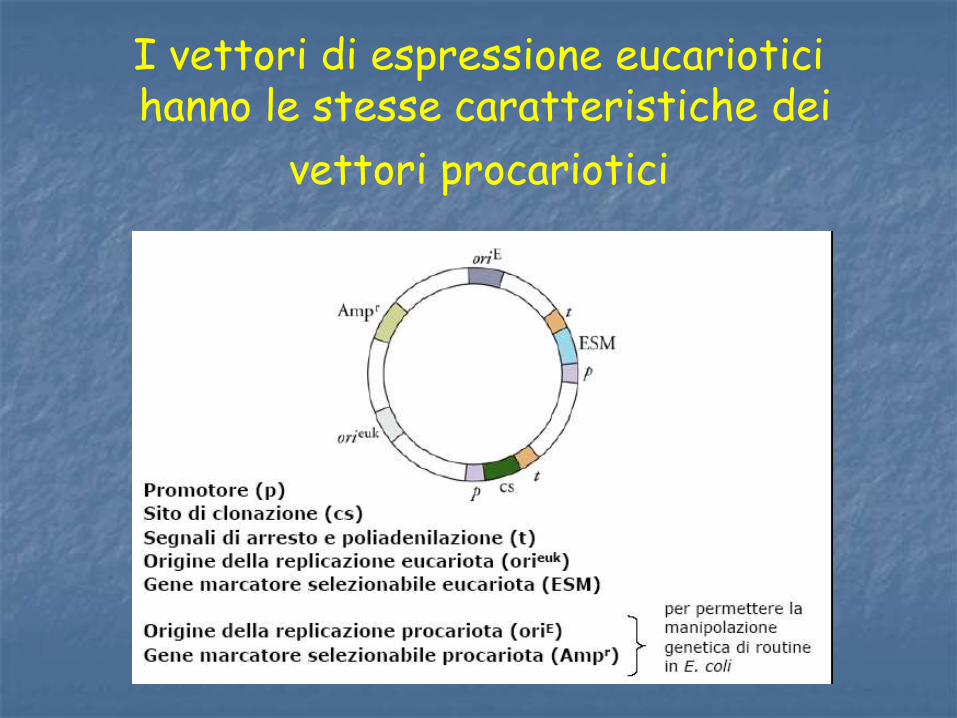
I vettori di espressione eucariotici hanno le stesse caratteristiche dei
vettori procariotici

VETTORI di Saccharomyces cerevisiae
Vettori episomiali o plasmidici Molto diffusi Possibile instabilità su larga scala
Vettori che si integrano Limitazione del numero di copie del gene clonato (uno per cromosoma) e quindi limitazione della resa in proteina geni spesso instabili
Cromosomi artificiali di lievito ( YAC ) Lungo frammento di DNA (100Kb) mantenuto nella cellula come cromosoma separato Altamente stabile Non ancora usato per produrre proteine eterologhe su scala industriale.

Il vettore YAC simula un cromosoma

Espressione in E.coli

INSULINA : PRIMO FARMACO RICOMBINANTE
Ormone di natura proteica che regola il metabolismo degli zuccheri,
iene prodotta dalle cellule β delle isole Langerhans del pancreas
e successivamente secreta nel torrente circolatorio.
L’ impossibilità di produrre insulina determina l’insorgenza del diabete.
Nel tentativo di sopperire al deficit di tale molecola si è iniziato
dapprima ad estrarla dal maiale e poi a sintetizzarla tramite tecnologia
del DNA ricombinante in sistemi batterici.
L’insulina ricavata dal pancreas di maiali, anche se biologicamente attiva
nell’ uomo, possedeva delle sequenze amminoacidiche diverse tali che
alcuni pazienti trattati producevano anticorpi contro-insulina che
determinavano occasionalmente gravi reazioni immunitarie.
.

Nei mammiferi l’ insulina è espressa come un singolo filamento di pre-prormone che viene secreto attraverso la membrana plasmatica; esso contiene degli amminoacidi aggiuntivi non presenti nell’ormone maturo. Durante la secrezione tali amminoacidi vengono eliminati ad opera di proteasi cellulari per rilasciare l’insulina matura che consta di due corte catene polipeptidiche A e B unite da due legami disolfuro.

CATENA A 30 aa CATENA B 21 aa peptide di collegamento C 35 aa
Unite da 2 ponti S-S
ESONE 2 ESONE 1
C A
B
PREPROINSULINA
PROINSULINA (forma S-S)
INSULINA
Peptide segnale
Nel Golgi un enzima rimuove il pepdide
di collegamento

INSULINA RICOMBINANTE L’insulina ricombinante è stata ottenuta inizialmente facendo esprimere separatamente le catene A e B e poi assemblandole nella configurazione della molecola matura. I segmenti che codificano per ciascuna catena sono stati sintetizzati chimicamente e inseriti in un vettore batterico tale che, dopo la traduzione, l’insulina fosse fusa all’ estremità carbossiterminale del gene β-gal di E.coli. I batteri producono elevate quantità di proteina di fusione e, dopo purificazione le catene insuliniche possono essere separate con BrCN il quale taglia tutti i legami successivi ad ogni metionina.
A
B β-galattosidasi
β-galattosidasi
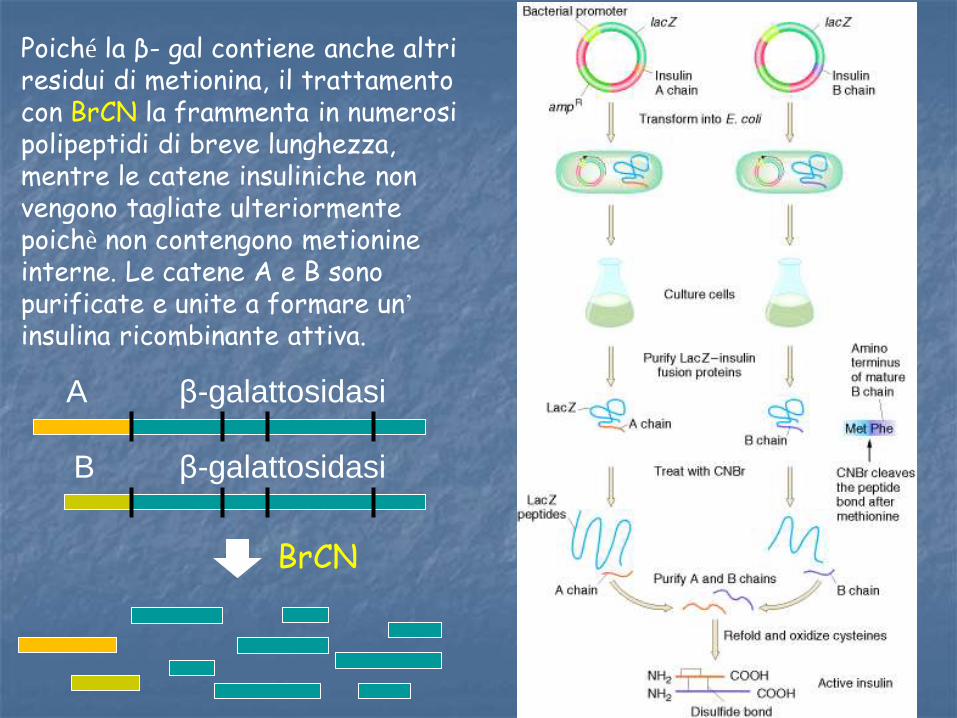
Poiché la β- gal contiene anche altri residui di metionina, il trattamento con BrCN la frammenta in numerosi polipeptidi di breve lunghezza, mentre le catene insuliniche non vengono tagliate ulteriormente poichè non contengono metionine interne. Le catene A e B sono purificate e unite a formare un’ insulina ricombinante attiva.
A
B β-galattosidasi
β-galattosidasi
BrCN

hGH UMANO RICOMBINANTE
E’ una proteina di 191 amminoacidi prodotta dall’ adenoipofisi
regola la crescita e lo sviluppo. Una carenza di hGH non consente,
nei bambini, di raggiungere una statura normale ma iniezioni
regolari di tale ormone ne stimolano lo sviluppo. A differenza di
quanto visto per l’ insulina, gli ormoni della crescita di origine
animale non sono efficaci: funziona solo la proteina di origine umana
Per molti anni la si è estratta dai cadaveri ma l’infezione da virus è
stata un’inevitabile conseguenza di tale trattamento.
La tecnologia ricombinante ha rappresentato una fonte sicura e
affidabile di tale ormone.

METODO A SECREZONE INTRACELLULARE
La manipolazione è stata necessaria poiché il sistema batterico non era in grado di produrre una forma normale della proteina umana matura. La sintesi di hGH si è ottenuta mediante la costruzione di un gene ibrido a partire da un cDNA di hGH naturale (aa 24-191) e da un oligonucleotide sintetico (aa 1-24) che codificavano la porzione aminoacidica terminale della forma matura della proteina. Tale sequenza codificante è stata ligata in un plasmide in posizione adiacente ad un promotore batterico e ad un gene che codifica per una Met d’inizio. Dopo transfezione in E. coli l’ormone inizia ad essere prodotto e ad accumularsi nella cellula batterica e infine può essere recuperato mediante opportune tecniche di purificazione.

METODO A SECREZIONE ESTERNA
E’ possibile produrre nei batteri una proteina senza una Met in più indirizzandola per la secrezione. A tale scopo è stato costruito un pre-ormone unendo il segmento di DNA codificante l’hGH ad una sequenza segnale ricavata da una proteina batterica. L’ ormone della crescita umano viene così prodotto in E. coli insieme alla sequenza segnale che indirizza le proteine verso la secrezione. Le proteine si accumulano nello spazio periplasmatico fra le membrane batteriche interna ed esterna e possono essere rilasciate all’esterno in seguito a rottura ipotonica della membrana. Una proteasi batterica al momento della secrezione rimuove il peptide segnale cosicchè le proteine prodotte non contengono la Met d’inizio.

Espressione in lievito

ESPRESSIONE DIRETTA in S.cerevisiae (accumulo di proteina nel citoplasma dell’ospite)
Cu/Zn - SOD ( superossido dismutasi )
catalasi
SOD, H + perossidasi
O2¯ H2O2 H2O + O2
anione perossido di
superossido idrogeno
La SOD trova un impiego terapeutico come agente antinfiammatorio
contro osteoartite, artrite reumatoide e spondilite anchilosante.

Il vettore della Cu/Zn-SOD umana-lievito
GAPD = gliceraldeide 3-fosfato deidrogenasi
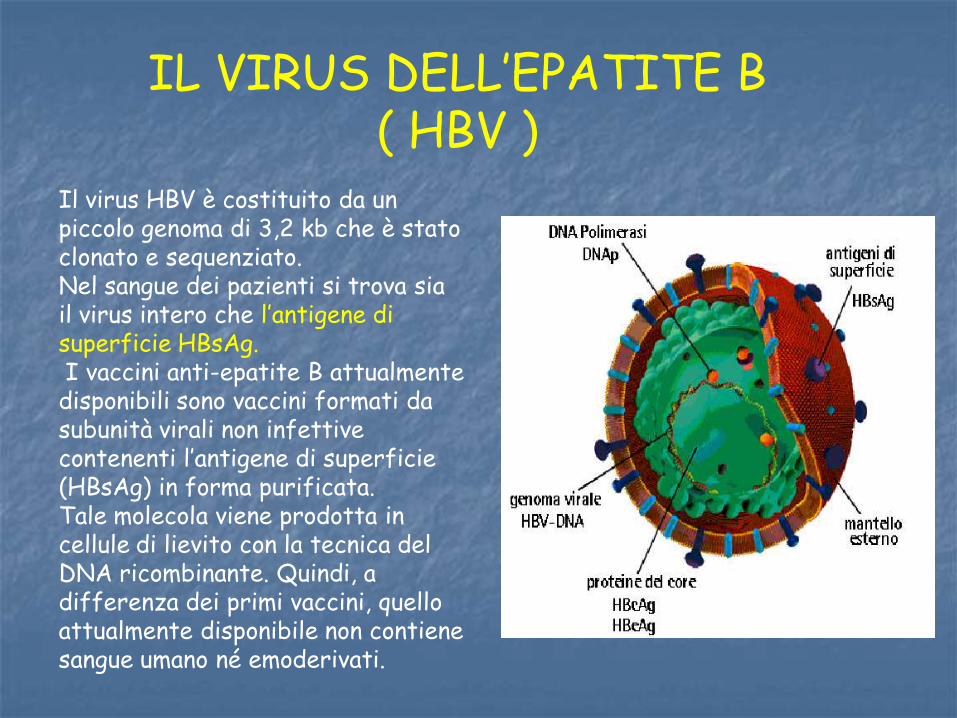
IL VIRUS DELL’EPATITE B ( HBV )
Il virus HBV è costituito da un piccolo genoma di 3,2 kb che è stato clonato e sequenziato. Nel sangue dei pazienti si trova sia il virus intero che l’antigene di superficie HBsAg. I vaccini anti-epatite B attualmente disponibili sono vaccini formati da subunità virali non infettive contenenti l’antigene di superficie (HBsAg) in forma purificata. Tale molecola viene prodotta in cellule di lievito con la tecnica del DNA ricombinante. Quindi, a differenza dei primi vaccini, quello attualmente disponibile non contiene sangue umano né emoderivati.

Preparazione di HBsAg espressione nel lievito
HBV è un virus difficile da propagare in coltura si è ricorso pertanto ad un vettore d’espressione in S. cerevisiae per clonare l’antigene di superficie. La trascrizione è indotta da un promotore forte a valle del quale viene inserito un terminatore di trascrizione. Il vettore contiene, ora, le origini di duplicazione e i marcatori sia per i batteri che per il lievito. Il lievito trasformato con tale plasmide può crescere in incubatori sino a concentrazioni cospicue tali da produrre un accumulo di quote elevate di HBsAg. Dopo opportuna purificazione le particelle antigeniche si aggregano a formare una strutture di circa 20 nm di diametro, le stesse che ritroviamo nei pazienti infettati da HBV.

Pichia pastoris: lievito metanoltrofico

Espressione in cellule di mammifero

Vettori di espressione per cellule di mammifero
Simili ai vettori eucariotici
Studio della funzione e
regolazione dei geni di
mammifero
Produzione di proteine
ricombinanti “autentiche”
utilizzabili in applicazioni
terapeutiche.

Sistemi marcatori selezionabili per vettori di espressione in cellule di mammifero
Neo: gene che codifica per la neomicina transferasi G-418: agente che blocca la traduzione e uccide la cellula neomicina fosfotransferasi
attiva inattiva
G-418 G-418
P

DHRF: gene che codifica la diidrofolato reduttasi Utilizzabile con cellule DHFR- DHFR- inibizione della crescita
MTX (metotrexato) DHFR+ proliferazione
normale
aumento di [ Mtx ] e selezione di cellule contenenti più copie del vettore
Sistemi marcatori selezionabili per vettori di espressione in cellule di mammifero
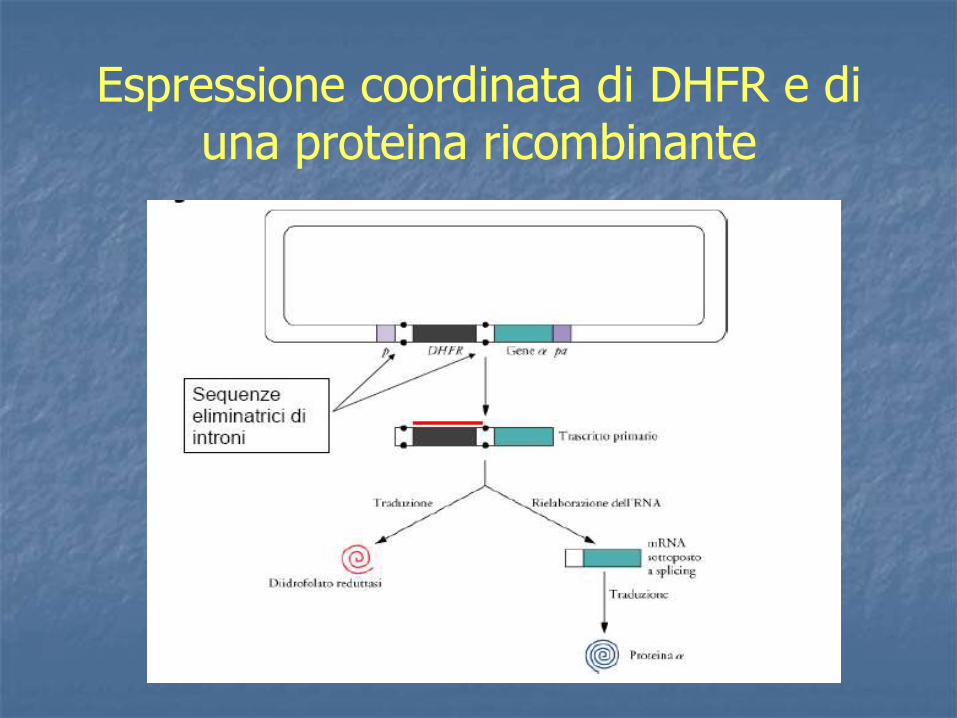
Espressione coordinata di DHFR e di una proteina ricombinante

COLTURE CELLULARI DI MAMMIFERO
Alcune proteine di interesse clinico sono caratterizzate da una complessità strutturale e funzionele tale da non potere essere prodotte in batteri o lieviti.
La moderna biotecnologia ha permesso il ricorso all’uso di colture di cellule di mammifero che pur richiedendo trattamenti dispendiosi consentono di produrre in modo corretto proteine modificate pienamente attive.

E’ il primo farmaco ad essere stato prodotto per mezzo di una coltura di cellule di mammifero. La proteina viene utilizzata come agente trombolitico nell’infarto miocardico, nell’embolia polmonare o in casi di
trombosi venosa. Il tPA è un enzima coinvolto nel processo di fibrinolisi che ha lo scopo di mantenere l’integrità del coagulo solo per il tempo strettamente necessario all’arresto dell’emorragia e di impedire altresì la formazione di trombi intravascolari che possono verificarsi a seguito dell’attivazione spontanea della coagulazione. La fibrinolisi inizia con la conversione del plasminogeno, precursore inattivo, in plasmina per opera degli attivatori del plasminogeno, i più noti fra i quali sono l’urochinasi e varie sostanze
di origine tissutale tra cui il tPA. La plasmina è una potente proteasi capace di scindere la fibrina
responsabile della formazione dei coaguli ematici. La rapida somministrazione di un attivatore del plasminogeno a seguito di un attacco cardiaco scioglie i trombi che, altrimenti, produrrebbero danni irreversibili al muscolo cardiaco.
tPA: attivatore del plasminogeno

Il cDNA del tPA umano è stato clonato e inserito in un vettore di espressione contenente un promotore forte e un terminatore. Con il vettore così costruito sono state transfettate stabilmente cellule di mammifero. Le cellule trasformate secernono il tPA nel mezzo di coltura ma con basse rese. Per aumentare i livelli di produzione si è ricorsi all’ uso di metotrexato che permette di selezionare solo quelle cellule in cui è avvenuta l’ amplificazione del vettore e quindi quella del gene codificante il tPA. Le linee cellulari ad alta capacità di espressione sono poste in coltura in grandi incubatori da cui il tPA ricombinante può essere purificato.

ACTILYSE: nuovo farmaco rtPA per il
trattamento dell’ictus ischemico acuto
Farmaco "coagulo selettivo “ che attiva la cascata fibrinolitica trasformando il plasminogeno in plasmina la quale scioglie la fibrina presente nel trombo. I nuovi dati clinici dimostrano che i pazienti trattati con rt-PA possono vedere aumentate la loro probabilità di evitare qualsiasi forma di invalidità a seguito di un ictus. Il farmaco è risultato efficace in una finestra temporale compresa tra le 3 e le 4,5 ore dalla comparsa dell’ictus.