
Study on the binding of Thioflavin T to β-sheet-rich and non-β-sheet cavities
12
Journal of Structural Biology 158 (2007) 358–369 www.elsevier.com/locate/yjsbi 1047-8477/$ - see front matter © 2006 Elsevier Inc. All rights reserved. doi:10.1016/j.jsb.2006.12.010 Study on the binding of ThioXavin T to -sheet-rich and non--sheet cavities Minna Groenning a,b,¤ , Lars Olsen c , Marco van de Weert a , James M. Flink b , Sven Frokjaer a , Flemming S. Jørgensen c a Department of Pharmaceutics and Analytical Chemistry, The Danish University of Pharmaceutical Sciences, Universitetsparken 2, 2100 Copenhagen, Denmark b Department of Biophysics, Novo Nordisk A/S, Novo Allé, 2880 Bagsvaerd, Denmark c Department of Medicinal Chemistry, The Danish University of Pharmaceutical Sciences, Universitetsparken 2, 2100 Copenhagen, Denmark Received 6 October 2006; received in revised form 18 December 2006; accepted 19 December 2006 Available online 31 December 2006 Abstract Amyloid Wbril formation plays a role in more than 20 diseases including Alzheimer’s disease. In vitro detection of these Wbrils is often performed using ThioXavin T (ThT), though the ThT binding mode is largely unknown. In the present study, spectral properties of ThT in binding environments representing -sheet-rich and non--sheet cavities were examined. Acetylcholinesterase and -cyclodextrin induced a characteristic ThT Xuorescence similar to that with amyloid Wbrils, whereas -cyclodextrin and the -sheet-rich transthyretin did not. The cavities of acetylcholinesterase and -cyclodextrin were of similar diameter and only these cavities could accommodate two ThT ions according to molecular modelling. Binding stoichiometry studies also showed a possible binding of two ThT ions. Thus, the characteristic ThT Xuorescence is induced in cavities with a diameter of 8–9 Å and a length able to accommodate the entire length of the ThT ion. The importance of a cavity diameter capable of binding two ThT ions, among others, indicates that an excimer formation is a plausible mechanism for the characteristic Xuorescence. We propose a similar ThT binding mode in amyloid Wbrils, where cavities of an appropriate size running parallel to the Wbril axis have previously been proposed in several amyloid Wbril models. © 2006 Elsevier Inc. All rights reserved. Keywords: Amyloid Wbril; Binding; Cavity; Dimer; Excimer; Fluorescence; Isothermal titration calorimetric; Molecular modelling; ThioXavin T 1. Introduction Amyloid Wbrils are associated with a number of diseases such as Alzheimer’s disease, Type 2 diabetes, senile amyloi- dosis, and dialysis amyloidosis. In some of these diseases the amyloid Wbril deposits cause the disease; in others it is still not established whether the Wbrils cause the diseases or are just associated symptoms (Pepys, 2006). The ability of proteins to form amyloid Wbrils has been observed for a large range of proteins both in vivo and in vitro and is sug- gested to be a generic feature of all polypeptide sequences (Dobson, 2003). Amyloid Wbrillation also takes place in protein-based drugs such as insulin where amyloid Wbrilla- tion causes problems during production, storage, and deliv- ery (Nielsen et al., 2001; Wang, 2005). The presence of amyloid Wbrils is a therapeutic problem since amyloid Wbril- lated proteins are biologically inactive (Brange et al., 1997; Grillo et al., 2001) and may cause immunological responses in patients (Brange et al., 1997; Onoue et al., 2004). There- fore, understanding the Wbrillation process and especially focusing on how to prevent or inhibit the Wbril formation are important issues in both biotechnology and medicine. A method for detection of amyloid Wbrils is essential to gain increased knowledge about the Wbrillation process. ThioXavin T (ThT) (Fig. 1) is a dye which is widely used to determine the presence of amyloid Wbrils and to examine Wbrillation kinetics in situ. Detection is based on the * Corresponding author. Fax: +45 3530 6030. E-mail address: [email protected] (M. Groenning).
-
Upload
minna-groenning -
Category
Documents
-
view
213 -
download
1
Transcript of Study on the binding of Thioflavin T to β-sheet-rich and non-β-sheet cavities

Journal of Structural Biology 158 (2007) 358–369
www.elsevier.com/locate/yjsbi
Study on the binding of ThioXavin T to �-sheet-rich and non-�-sheet cavities
Minna Groenning a,b,¤, Lars Olsen c, Marco van de Weert a, James M. Flink b, Sven Frokjaer a, Flemming S. Jørgensen c
a Department of Pharmaceutics and Analytical Chemistry, The Danish University of Pharmaceutical Sciences, Universitetsparken 2, 2100 Copenhagen, Denmark
b Department of Biophysics, Novo Nordisk A/S, Novo Allé, 2880 Bagsvaerd, Denmarkc Department of Medicinal Chemistry, The Danish University of Pharmaceutical Sciences, Universitetsparken 2, 2100 Copenhagen, Denmark
Received 6 October 2006; received in revised form 18 December 2006; accepted 19 December 2006Available online 31 December 2006
Abstract
Amyloid Wbril formation plays a role in more than 20 diseases including Alzheimer’s disease. In vitro detection of these Wbrils is oftenperformed using ThioXavin T (ThT), though the ThT binding mode is largely unknown. In the present study, spectral properties of ThTin binding environments representing �-sheet-rich and non-�-sheet cavities were examined. Acetylcholinesterase and �-cyclodextrininduced a characteristic ThT Xuorescence similar to that with amyloid Wbrils, whereas �-cyclodextrin and the �-sheet-rich transthyretindid not. The cavities of acetylcholinesterase and �-cyclodextrin were of similar diameter and only these cavities could accommodate twoThT ions according to molecular modelling. Binding stoichiometry studies also showed a possible binding of two ThT ions. Thus, thecharacteristic ThT Xuorescence is induced in cavities with a diameter of 8–9 Å and a length able to accommodate the entire length of theThT ion. The importance of a cavity diameter capable of binding two ThT ions, among others, indicates that an excimer formation is aplausible mechanism for the characteristic Xuorescence. We propose a similar ThT binding mode in amyloid Wbrils, where cavities of anappropriate size running parallel to the Wbril axis have previously been proposed in several amyloid Wbril models.© 2006 Elsevier Inc. All rights reserved.
Keywords: Amyloid Wbril; Binding; Cavity; Dimer; Excimer; Fluorescence; Isothermal titration calorimetric; Molecular modelling; ThioXavin T
1. Introduction
Amyloid Wbrils are associated with a number of diseasessuch as Alzheimer’s disease, Type 2 diabetes, senile amyloi-dosis, and dialysis amyloidosis. In some of these diseasesthe amyloid Wbril deposits cause the disease; in others it isstill not established whether the Wbrils cause the diseases orare just associated symptoms (Pepys, 2006). The ability ofproteins to form amyloid Wbrils has been observed for alarge range of proteins both in vivo and in vitro and is sug-gested to be a generic feature of all polypeptide sequences(Dobson, 2003). Amyloid Wbrillation also takes place in
* Corresponding author. Fax: +45 3530 6030.E-mail address: [email protected] (M. Groenning).
1047-8477/$ - see front matter © 2006 Elsevier Inc. All rights reserved.doi:10.1016/j.jsb.2006.12.010
protein-based drugs such as insulin where amyloid Wbrilla-tion causes problems during production, storage, and deliv-ery (Nielsen et al., 2001; Wang, 2005). The presence ofamyloid Wbrils is a therapeutic problem since amyloid Wbril-lated proteins are biologically inactive (Brange et al., 1997;Grillo et al., 2001) and may cause immunological responsesin patients (Brange et al., 1997; Onoue et al., 2004). There-fore, understanding the Wbrillation process and especiallyfocusing on how to prevent or inhibit the Wbril formationare important issues in both biotechnology and medicine.
A method for detection of amyloid Wbrils is essential togain increased knowledge about the Wbrillation process.ThioXavin T (ThT) (Fig. 1) is a dye which is widely used todetermine the presence of amyloid Wbrils and to examineWbrillation kinetics in situ. Detection is based on the

M. Groenning et al. / Journal of Structural Biology 158 (2007) 358–369 359
Xuorescence characteristics of ThT; free ThT Xuorescesfaintly with excitation and emission maxima of 350 and440 nm, respectively, whereas upon interaction with amy-loid Wbrils a sizably enhanced ThT Xuorescence is observedwith excitation and emission maxima at about 450 and480 nm, respectively (LeVine, 1993; Naiki et al., 1989). Ingeneral, the characteristic Xuorescence of ThT is consideredspeciWc towards amyloid Wbrils and does not occur on bind-ing to the precursor proteins, or amorphous aggregates ofproteins (LeVine, 1999).
The characteristic Xuorescence of ThT is observed uponbinding of ThT to amyloid Wbrils formed from proteinswith a wide variety of size and primary, secondary, and ter-tiary structure (LeVine, 1995). Based on these observations,the primary structure of the proteins is not considered cru-cial in obtaining a proper binding between ThT and amy-loid Wbrils, although it may account for some of theobserved diVerences in characteristic ThT Xuorescenceintensity. Furthermore, the characteristic ThT Xuorescenceintensity may vary considerably depending on the mor-phology of the amyloid Wbrils, as shown for diVerent typesof glucagon Wbrils (Pedersen et al., 2006). Common struc-tural characteristics of the amyloid Wbrils are likely to beimportant for the binding of ThT. For example, amyloidWbrils are suggested to contain a common secondary struc-ture with a characteristic arrangement of contiguous �-sheet polypeptide chains aligned perpendicular to the longaxis of the Wbril, as revealed by X-ray diVraction studies(Eanes and Glenner, 1968; Serpell et al., 2000). Further-more, electron microscopy and atomic force microscopystudies generally show that Wbrils have a common appear-ance, being typically about 10 nm in diameter, though withhighly variable lengths up to several micrometer, and areoften twisted and unbranched (Cohen and Calkins, 1959;Jimenez et al., 2002; Khurana et al., 2003). A central cavityhas also been proposed, though not in all amyloid struc-tural models. For example, a number of tubular modelshave been proposed (Malinchik et al., 1998; Lazo andDowning, 1998; Perutz et al., 2002). Cross-� sheet modelscontaining a cavity are also proposed, e.g., for transthyretinwhere a �-sheet helix with a central cavity is proposed(Blake and Serpell, 1996) and for SH3 where cryo-electronmicroscopy reveals a double helix of two protoWlamentpairs wound around a hollow core (Jimenez et al., 1999).Notably, the presence of elongated cavities parallel to theWbril axis has been suggested both to be an essential part ofthe protoWlament structure and to be formed between the
Fig. 1. Structure of ThioXavin T with the dimensions 15.2, 6.1, and 4.3 Å(Krebs et al., 2005).
protoWlaments when associating to form protoWbrils andmature Wbrils.
Increased knowledge on the mode of ThT binding isessential for understanding the structural characteristic ofthe amyloid Wbrils being detected by ThT. This is crucial fora more elaborate interpretation of the characteristic ThTsignal, e.g., occurring when studying the Wbrillation kinetics.However, the lack of an atomic resolution structure ofamyloid Wbrils complicates the examination of the bindingmode to amyloid Wbrils. ThT most likely binds to a com-mon structural characteristic of the amyloid Wbril andexamination of the eVect of diVerent environments, like theproposed presence of cavities, on the occurrence of thecharacteristic ThT Xuorescence may be crucial in the searchfor a proper binding mode. Therefore, the aim of the cur-rent work is to further examine the binding mode of ThTwhich induces the characteristic ThT Xuorescence. Theexamination of the binding mode is done by studying theinteraction between ThT and selected compounds repre-senting �-sheet-rich and non-�-sheet cavity structures usingXuorescence spectroscopy, isothermal titration calorimetry(ITC) and molecular modelling.
2. Materials and methods
2.1. Materials
ThT was purchased as the chloride salt from Sigma–Aldrich Chemie GmbH (Schnelldorf, Germany). Beforeuse, ThT was recrystallized three times in demineralizedwater. The purity of ThT was veriWed by proton NMR. Amolar extinction coeYcient of 36,000 M¡1 cm¡1 at 412 nmwas determined for the recrystallized ThT in demineralizedwater. Human insulin with zinc, batch JMOHO2801, wasobtained from Novo Nordisk A/S (Bagsvaerd, Denmark).The zinc content was 0.37%, corresponding to approxi-mately 2 Zn2+ ions per insulin hexamer. Transthyretin(TTR) was purchased from Sigma–Aldrich Chemie GmbH(Schnelldorf, Germany). �-Cyclodextrin (�-CD) and �-cyclodextrin (�-CD) were purchased from Fluka ChemieGmbH (Buchs, Switzerland). Acetylcholinesterase (AChE)from electric eel, type VI-s, lyophilized powder, was pur-chased from Sigma (St. Louis, MO, USA). The AChE con-centration was determined by UV absorbance at 280 nmusing an extinction coeYcient of 1.8 for 1 mg/ml. All otherchemicals were of analytical grade.
2.2. Anisotropy
Anisotropy was measured using a Spex Fluorolog 3-22Xuorescence spectrometer (Jobin Yvon Horiba, France)with a 450 W Xenon lamp. The Xuorescence of free ThTwas used and therefore, the samples were excited at 350 nmusing slits of 5 nm, while the emission anisotropy was deter-mined at 450 nm using slits of 10 nm. All measurementswere carried out at 20 °C, and the anisotropy was measuredusing right angle settings. The integration time was 1 s and

360 M. Groenning et al. / Journal of Structural Biology 158 (2007) 358–369
three measurements were performed on each sample. Theexamination was conducted at a ThT and �-CD concentra-tion of 83 and 417�M, respectively, and at a concentrationof 8.3�M for both ThT and TTR.
2.3. Job plot
The binding stoichiometry of ThT binding to AChE in50 mM Tris-buVer (pH 7.5) was estimated by a continuousvariation analysis (Job plot) (Huang, 1982) using the char-acteristic Xuorescence of bound ThT. Eleven samples con-taining molar fractions between 0–1 and 1–0 (ThT–AChE)were prepared. The total concentration of AChE plus ThTfor any given sample was 20�M. Technical Xuorescencespectra were measured with the Spex Fluorolog 3-22 Xuo-rescence spectrometer. An emission scan from 460 to600 nm using emission slits of 5 nm was excited at 450 nmusing slits of 1 nm. The integration time was 0.1 s and anincrement of 0.5 nm was used. All measurements were car-ried out at 20 °C. Inner Wlter corrections were unnecessarydue to the low absorbance of AChE and ThT at this con-centration and wavelength range. Each sample was cor-rected for the signal of AChE in the absence of ThT and forthe signal of ThT in the absence of AChE.
2.4. Isothermal titration calorimetry
13.6 mM �-CD was titrated into 1.4002 ml of 1 mM ThTusing a VP-ITC MicroCalorimeter from MicroCal (North-ampton, USA). Both solutions were adjusted to pH 6.5 withNaOH and were degassed under vacuum with a Thermo-Vac accessory also from MicroCal. An initial injection of3 �l was followed by 29 injections of 10 �l with a 5-minequilibration period between the injections. The system wasequilibrated at 20 °C and 290 rpm mixing. Afterwards,excess cell solution was removed and the titration was con-tinued twice with 30 injections of 10�l. Three independenttitrations were performed. Titration of �-CD into milli-Qwater (pH 6.5) and milli-Q water (pH 6.5) titrated into ThTonly contributed with constant minor endothermic heatchanges upon dilution. Therefore, the Y-axis was translatedto correct for the heat of dilution. Data analysis of each ofthe three ITC runs was individually performed by ITCData Analysis in MicroCal Origin 7.0. A similar experimentwas performed with 13.7 mM �-CD titrated into 1.4002 mlof 1 mM ThT at 20 °C.
2.5. Preparation of insulin Wbrils
A Fluostar Optima platereader from BMG Labtech-nologies (OVenburg, Germany) was used with 96-well,black, polystyrene, non-sterile plates with optical bottomfrom Nalge Nunc International (Rochester, NY, USA).To avoid dehydration, the plates were covered with Poly-oleWn non-sterile sealing tape also from Nalge NuncInternational. A concentration of 5.2 mM insulin in0.025 mM HCl (pH 1.6) was used and Wbrillated for 120 h
at 37 °C using 1 mm linear shaking (350 rpm) for 300 sduring each cycle of 400 s.
2.6. Fluorescence characteristic of ThT
Technical Xuorescence spectra were monitored using aVarian Cary Eclipse Fluorescence Spectrophotometer (Vic-toria, Australia). The occurrence of the characteristic ThTXuorescence was evaluated by performing emission scansfrom 370 to 600 nm and from 460 to 600 nm when excited at350 and 450 nm, respectively. Following, the exact excita-tion and emission peak maxima were determined for thecharacteristic ThT Xuorescence. The Scan Control is set toSlow and an appropriate PMT Detector Voltage was set,i.e., either medium or high. Excitation and emission slits of5 or 10 nm were used and all measurements were performedat 21 °C. The positive control contained 15 �M insulin inWbril form (prepared as described above) and 20 �M ThT.The Xuorescence characteristics of ThT in �-CD wereexamined at a condition where a Xuorescent 2:2 complexpreviously has been detected, i.e., 21 mM �-CD and 26 �MThT (Retna Raj and Ramaraj, 1997). The ThT Xuorescenceupon binding to �-CD has been examined where one ThTion is proposed to be in complex with two �-CDs (RetnaRaj and Ramaraj, 1999), i.e., 15 mM �-CD with 26 �M ThT.The spectral properties of AChE in the range 0.1 �M–1.1 mM AChE with 26�M ThT in 50 mM Tris-buVer (pH7.5) were examined. The Xuorescence characteristics of ThTupon binding to TTR were examined at diVerent pH valuesand a range of concentrations i.e., the Xuorescence charac-teristics were examined at 0.18 and 1.19 �M TTR in both1 M citrate buVer (pH 4) and in milli-Q water (pH 7.5), andwith ThT concentrations in the range 1.1–26 �M.
2.7. Molecular modelling
A short energy minimization (200 iterations) of thestructure of one ThT and four possible combinations oftwo ThT ions (ThT dimers) (Fig. 2) was performed with theMMFF94s force Weld (Halgreen, 1999) in the Sybyl soft-ware version 6.9 (Tripos Associates Inc., St. Louis, USA).These structures were used for the subsequent dockingstudies. The examination of possible complexes betweenThT and the binding models did not require examination ofa ThT trimer due to the size of the cavities.
Possible orientations of ThT ligands within TTR andAChE were investigated using the docking program GOLD(Generic Optimization of Ligand Docking) version 1.2(Jones et al., 1997). The program allows positioning of thefully Xexible ligand within a partially Xexible protein, i.e.,the polar hydrogens are Xexible, by a genetic docking algo-rithm. Several crystal structures of the two proteins wereavailable from the Protein Data Bank (PDB). The calcula-tions were based on the X-ray structure of TTR (PDB entry1BM7) (Peterson et al., 1998) and AChE (PDB entry1EVE) (Kryger et al., 1999). In addition, a modiWed TTRmodel with a more open entrance to the binding site was

M. Groenning et al. / Journal of Structural Biology 158 (2007) 358–369 361
constructed by adjusting the Lys15 side chains into a posi-tion similar to the position in the TTR structure with PDBentry 1BZD (Schormann et al., 1998) where no ligand wasbound. All water molecules, and if present also unspeciWcbound ligands, were removed from the proteins prior to thedocking studies. The N-terminal and Arg and Lys residueswere protonated whereas the C-terminal and Asp and Gluresidues were deprotonated. An active site for docking wasdeWned as a sphere of 20 Å around a central atom in theactive site cavity of the proteins.
The various ThT combinations were manually placedinto the middle of the cavity of two �-CDs or two �-CDs.The length of one ThT corresponds to the length of theconnected cavity of two cyclodextrins, since the depth ofone cavity is 8 Å (Connors, 1997). Several structures of �-CD and �-CD were found in the Cambridge StructuralDatabase (CSD). For �-CD, a tubular structure of twoaligned �-CDs in a complex, with no side chains attached tothe CDs, was chosen (CSD entry AGAZIR) (Alexanderet al., 2002). For �-CD, a tubular structure of three aligned�-cyclodextrins in a complex, with no side chains attachedto the CDs, was selected (CSD entry SIBJES) (Ding et al.,1991). Both combinations of two �-CDs were investigated,i.e., the combination with the exterior rim with reduceddiameter of each CD facing towards each other, and wherethe other rim consisting of secondary hydroxyl groups faceseach other. The complex with two exterior rims consistingof secondary hydroxyl groups facing each other was exam-ined for �-CDs. The energy minimization of ThT was per-formed with the MMFF94s force Weld. ThT andcyclodextrin hydrogens were allowed to relax.
The energy associated with rotation around the bondbetween the benzothiazole and benzamine rings was cal-culated by freezing the S-C-C-C angle at appropriate val-ues and relaxing all other degrees of freedom usingdensity functional theory (DFT). Geometry optimiza-tions and subsequent single point calculations were per-formed using the 6-31G¤¤ and 6-311++G(2d,2p) basissets, respectively, with the B3LYP method (Becke,1993a,b; Lee et al., 1988), as implemented in the Gauss-ian03 program (Frisch et al., 2004). To estimate the eVectfrom the surrounding solvent on the energies for thediVerent geometries, calculations at the B3LYP/6-31G¤¤
level with the CPCM model (Barone and Cossi, 1998;Cossi et al., 2003) were performed.
3. Results
3.1. Binding of ThT
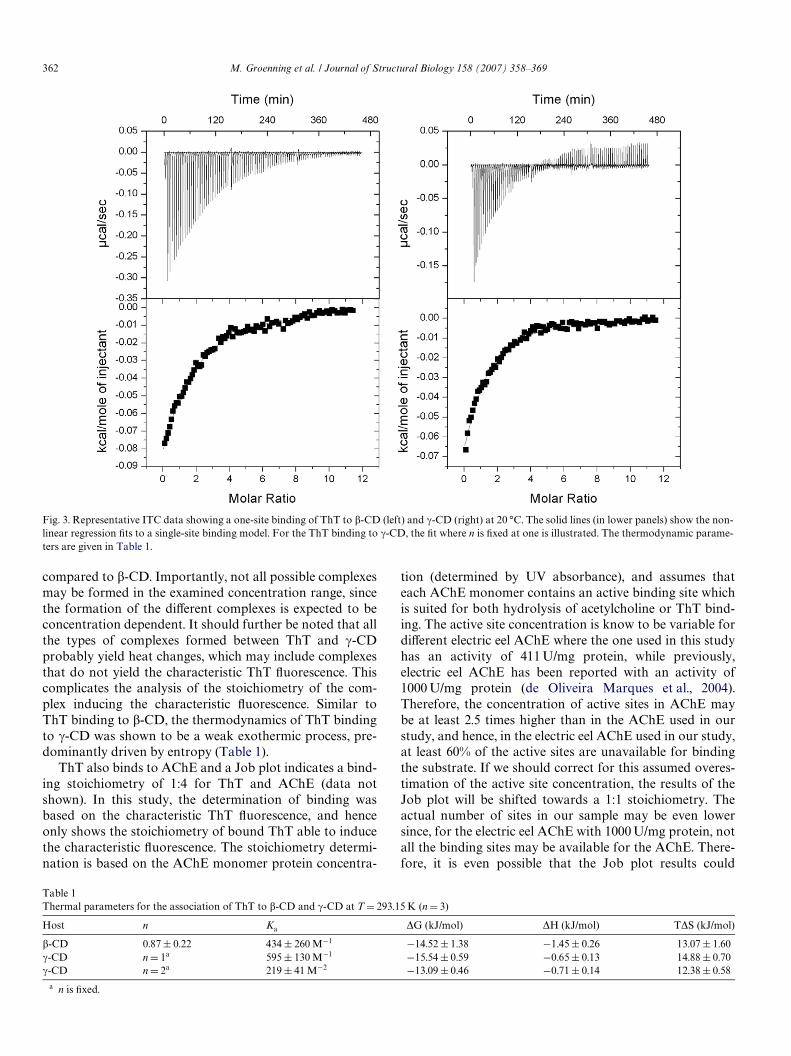
The binding of ThT to TTR and �-CD was studied byanisotropy measurements. The anisotropy of ThTincreased from 0.072§0.001 to 0.096§ 0.001 when ThTwas mixed with �-CD, and from 0.071§0.004 to0.280§0.004 when ThT was mixed with TTR. These signiW-cant increases in the anisotropy showed that ThT has lessrotational motion in the presence of the binding com-pounds compared to free ThT. As expected, the homotetr-amer of TTR with a total molecular mass of 55 kDa(Hornberg et al., 2000) has a larger eVect on the anisotropyof ThT than �-CD which has a much smaller molecularmass. The anisotropy increases indicate that ThT is boundto both �-CD and TTR. Furthermore, ThT binding to �-CD was evaluated by ITC measurements at 20 °C (Fig. 3)where a low aYnity binding of ThT to �-CD was deter-mined with a binding constant, Ka, of about 400 M¡1 usinga single-site binding model. In addition, the number ofbinding sites (n) was determined to be 0.87§0.22 (Table 1).The ITC experiments also showed negative enthalpies (�H)for ThT associating with �-CD in milli-Q water (pH 6.5)(Table 1) which shows that the formation of the inclusioncomplexes is a weak exothermic process. The entropy eVect(T�S) is positive and evidently gives larger contribution tothe negative change of Gibbs energy (�G) than theenthalpy does (Table 1). Therefore, the formation of thisinclusion complex is predominantly driven by entropy.
ThT was also shown to bind to �-CD using ITC (Fig. 3).A low aYnity binding of ThT to �-CD was determined witha Ka similar to the one obtained for �-CD (Table 1). Thelow aYnity binding to the �-CD complicates an exact deter-mination of the number of sites, and it was necessary to Wxn, which is often the case for low aYnity systems (Turnbulland Daranas, 2003; Tellinghuisen, 2005). A single bindingsite model was used and n was Wxed at either 0.5, 1, or 2.The binding of ThT in �-CD is possible to have both nD1and nD2 as judged from the Wts having similar �2-values, incontrast to nD0.5 which haves a higher �2. This indicatesthat at the given conditions, one ThT does not bind withtwo �-CDs. However, ThT and �-CD may bind in a 1:1 (or2:2) complex, or two ThT may bind in one �-CD which isexpected to be possible due to the larger diameter of �-CD
Fig. 2. The four ways two ThioXavin T ions were stacked in the molecular modelling.
362 M. Groenning et al. / Journal of Structural Biology 158 (2007) 358–369
compared to �-CD. Importantly, not all possible complexesmay be formed in the examined concentration range, sincethe formation of the diVerent complexes is expected to beconcentration dependent. It should further be noted that allthe types of complexes formed between ThT and �-CDprobably yield heat changes, which may include complexesthat do not yield the characteristic ThT Xuorescence. Thiscomplicates the analysis of the stoichiometry of the com-plex inducing the characteristic Xuorescence. Similar toThT binding to �-CD, the thermodynamics of ThT bindingto �-CD was shown to be a weak exothermic process, pre-dominantly driven by entropy (Table 1).
ThT also binds to AChE and a Job plot indicates a bind-ing stoichiometry of 1:4 for ThT and AChE (data notshown). In this study, the determination of binding wasbased on the characteristic ThT Xuorescence, and henceonly shows the stoichiometry of bound ThT able to inducethe characteristic Xuorescence. The stoichiometry determi-nation is based on the AChE monomer protein concentra-
tion (determined by UV absorbance), and assumes thateach AChE monomer contains an active binding site whichis suited for both hydrolysis of acetylcholine or ThT bind-ing. The active site concentration is know to be variable fordiVerent electric eel AChE where the one used in this studyhas an activity of 411 U/mg protein, while previously,electric eel AChE has been reported with an activity of1000 U/mg protein (de Oliveira Marques et al., 2004).Therefore, the concentration of active sites in AChE maybe at least 2.5 times higher than in the AChE used in ourstudy, and hence, in the electric eel AChE used in our study,at least 60% of the active sites are unavailable for bindingthe substrate. If we should correct for this assumed overes-timation of the active site concentration, the results of theJob plot will be shifted towards a 1:1 stoichiometry. Theactual number of sites in our sample may be even lowersince, for the electric eel AChE with 1000 U/mg protein, notall the binding sites may be available for the AChE. There-fore, it is even possible that the Job plot results could
Fig. 3. Representative ITC data showing a one-site binding of ThT to �-CD (left) and �-CD (right) at 20 °C. The solid lines (in lower panels) show the non-linear regression Wts to a single-site binding model. For the ThT binding to �-CD, the Wt where n is Wxed at one is illustrated. The thermodynamic parame-ters are given in Table 1.
Table 1Thermal parameters for the association of ThT to �-CD and �-CD at T D 293.15 K (n D 3)
a n is Wxed.
Host n Ka �G (kJ/mol) �H (kJ/mol) T�S (kJ/mol)
�-CD 0.87§ 0.22 434 § 260 M¡1 ¡14.52 § 1.38 ¡1.45§ 0.26 13.07 § 1.60�-CD n D 1a 595 § 130 M¡1 ¡15.54 § 0.59 ¡0.65§ 0.13 14.88 § 0.70�-CD n D 2a 219 § 41 M¡2 ¡13.09 § 0.46 ¡0.71§ 0.14 12.38 § 0.58
M. Groenning et al. / Journal of Structural Biology 158 (2007) 358–369 363
indicate that two ThT ions bind to the binding site in theAChE monomer.
3.2. Fluorescence characteristics of ThT bound in �-sheet-rich and non-�-sheet cavities
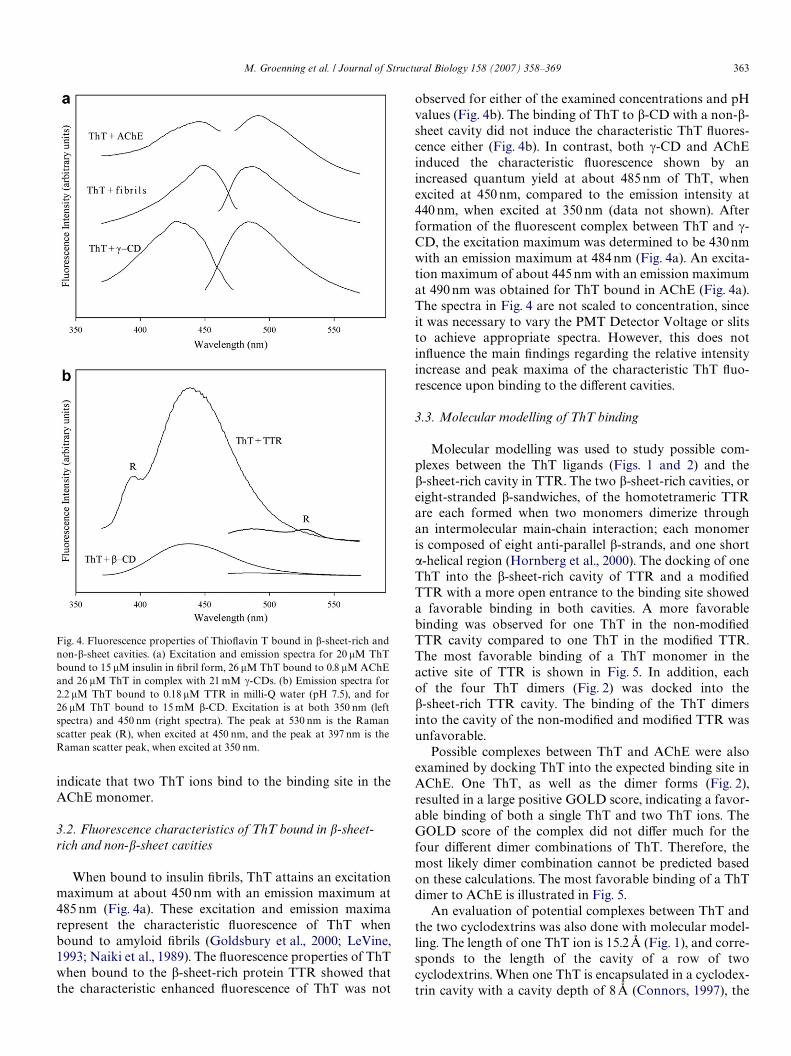
When bound to insulin Wbrils, ThT attains an excitationmaximum at about 450 nm with an emission maximum at485 nm (Fig. 4a). These excitation and emission maximarepresent the characteristic Xuorescence of ThT whenbound to amyloid Wbrils (Goldsbury et al., 2000; LeVine,1993; Naiki et al., 1989). The Xuorescence properties of ThTwhen bound to the �-sheet-rich protein TTR showed thatthe characteristic enhanced Xuorescence of ThT was not
Fig. 4. Fluorescence properties of ThioXavin T bound in �-sheet-rich andnon-�-sheet cavities. (a) Excitation and emission spectra for 20 �M ThTbound to 15 �M insulin in Wbril form, 26 �M ThT bound to 0.8 �M AChEand 26 �M ThT in complex with 21 mM �-CDs. (b) Emission spectra for2.2 �M ThT bound to 0.18 �M TTR in milli-Q water (pH 7.5), and for26 �M ThT bound to 15 mM �-CD. Excitation is at both 350 nm (leftspectra) and 450 nm (right spectra). The peak at 530 nm is the Ramanscatter peak (R), when excited at 450 nm, and the peak at 397 nm is theRaman scatter peak, when excited at 350 nm.
observed for either of the examined concentrations and pHvalues (Fig. 4b). The binding of ThT to �-CD with a non-�-sheet cavity did not induce the characteristic ThT Xuores-cence either (Fig. 4b). In contrast, both �-CD and AChEinduced the characteristic Xuorescence shown by anincreased quantum yield at about 485 nm of ThT, whenexcited at 450 nm, compared to the emission intensity at440 nm, when excited at 350 nm (data not shown). Afterformation of the Xuorescent complex between ThT and �-CD, the excitation maximum was determined to be 430 nmwith an emission maximum at 484 nm (Fig. 4a). An excita-tion maximum of about 445 nm with an emission maximumat 490 nm was obtained for ThT bound in AChE (Fig. 4a).The spectra in Fig. 4 are not scaled to concentration, sinceit was necessary to vary the PMT Detector Voltage or slitsto achieve appropriate spectra. However, this does notinXuence the main Wndings regarding the relative intensityincrease and peak maxima of the characteristic ThT Xuo-rescence upon binding to the diVerent cavities.
3.3. Molecular modelling of ThT binding
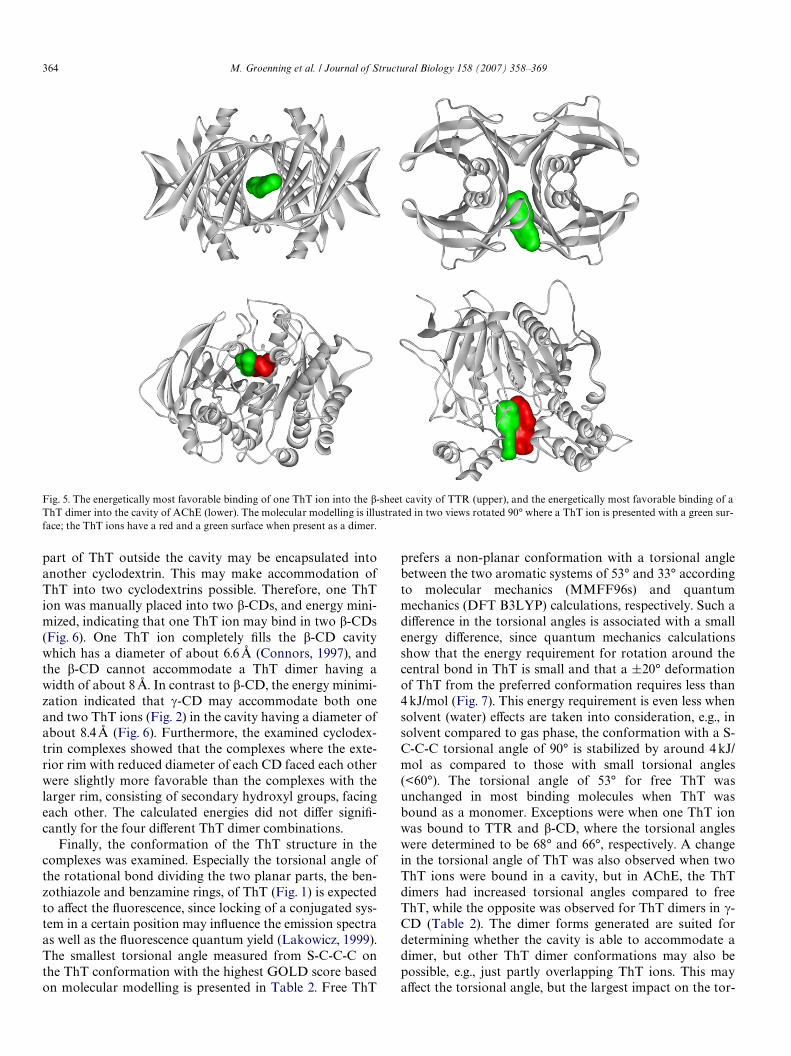
Molecular modelling was used to study possible com-plexes between the ThT ligands (Figs. 1 and 2) and the�-sheet-rich cavity in TTR. The two �-sheet-rich cavities, oreight-stranded �-sandwiches, of the homotetrameric TTRare each formed when two monomers dimerize throughan intermolecular main-chain interaction; each monomeris composed of eight anti-parallel �-strands, and one short�-helical region (Hornberg et al., 2000). The docking of oneThT into the �-sheet-rich cavity of TTR and a modiWedTTR with a more open entrance to the binding site showeda favorable binding in both cavities. A more favorablebinding was observed for one ThT in the non-modiWedTTR cavity compared to one ThT in the modiWed TTR.The most favorable binding of a ThT monomer in theactive site of TTR is shown in Fig. 5. In addition, eachof the four ThT dimers (Fig. 2) was docked into the�-sheet-rich TTR cavity. The binding of the ThT dimersinto the cavity of the non-modiWed and modiWed TTR wasunfavorable.
Possible complexes between ThT and AChE were alsoexamined by docking ThT into the expected binding site inAChE. One ThT, as well as the dimer forms (Fig. 2),resulted in a large positive GOLD score, indicating a favor-able binding of both a single ThT and two ThT ions. TheGOLD score of the complex did not diVer much for thefour diVerent dimer combinations of ThT. Therefore, themost likely dimer combination cannot be predicted basedon these calculations. The most favorable binding of a ThTdimer to AChE is illustrated in Fig. 5.
An evaluation of potential complexes between ThT andthe two cyclodextrins was also done with molecular model-ling. The length of one ThT ion is 15.2 Å (Fig. 1), and corre-sponds to the length of the cavity of a row of twocyclodextrins. When one ThT is encapsulated in a cyclodex-trin cavity with a cavity depth of 8 Å (Connors, 1997), the
364 M. Groenning et al. / Journal of Structural Biology 158 (2007) 358–369
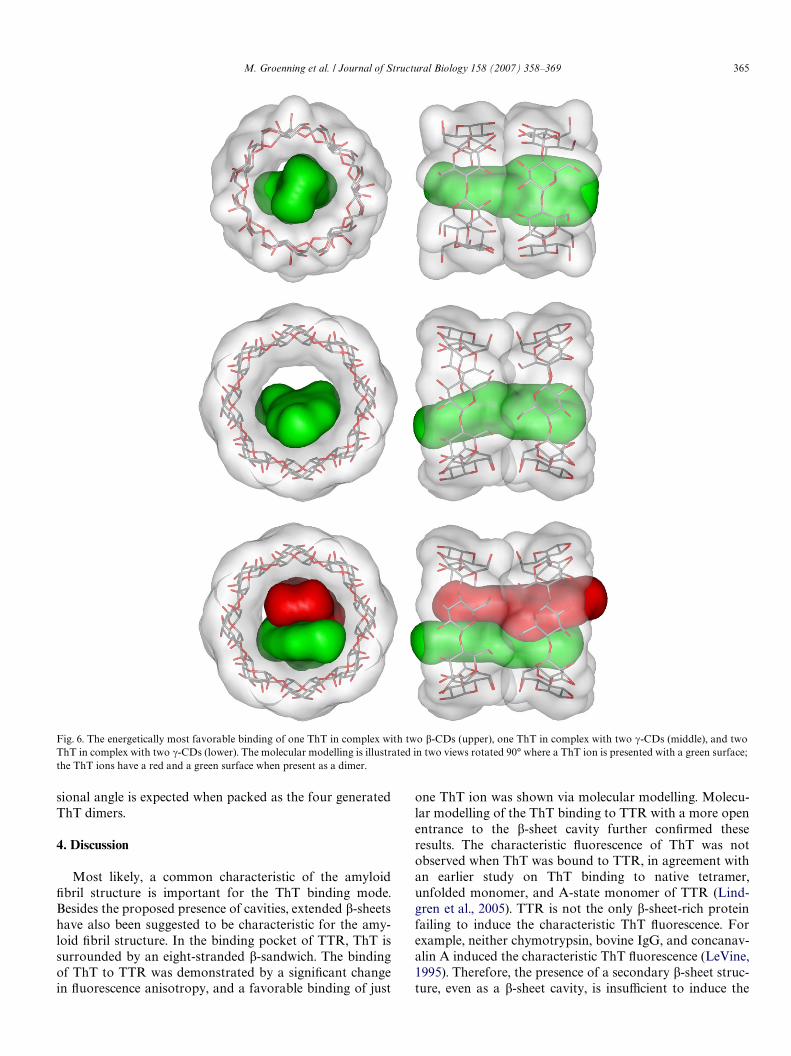
part of ThT outside the cavity may be encapsulated intoanother cyclodextrin. This may make accommodation ofThT into two cyclodextrins possible. Therefore, one ThTion was manually placed into two �-CDs, and energy mini-mized, indicating that one ThT ion may bind in two �-CDs(Fig. 6). One ThT ion completely Wlls the �-CD cavitywhich has a diameter of about 6.6 Å (Connors, 1997), andthe �-CD cannot accommodate a ThT dimer having awidth of about 8 Å. In contrast to �-CD, the energy minimi-zation indicated that �-CD may accommodate both oneand two ThT ions (Fig. 2) in the cavity having a diameter ofabout 8.4 Å (Fig. 6). Furthermore, the examined cyclodex-trin complexes showed that the complexes where the exte-rior rim with reduced diameter of each CD faced each otherwere slightly more favorable than the complexes with thelarger rim, consisting of secondary hydroxyl groups, facingeach other. The calculated energies did not diVer signiW-cantly for the four diVerent ThT dimer combinations.
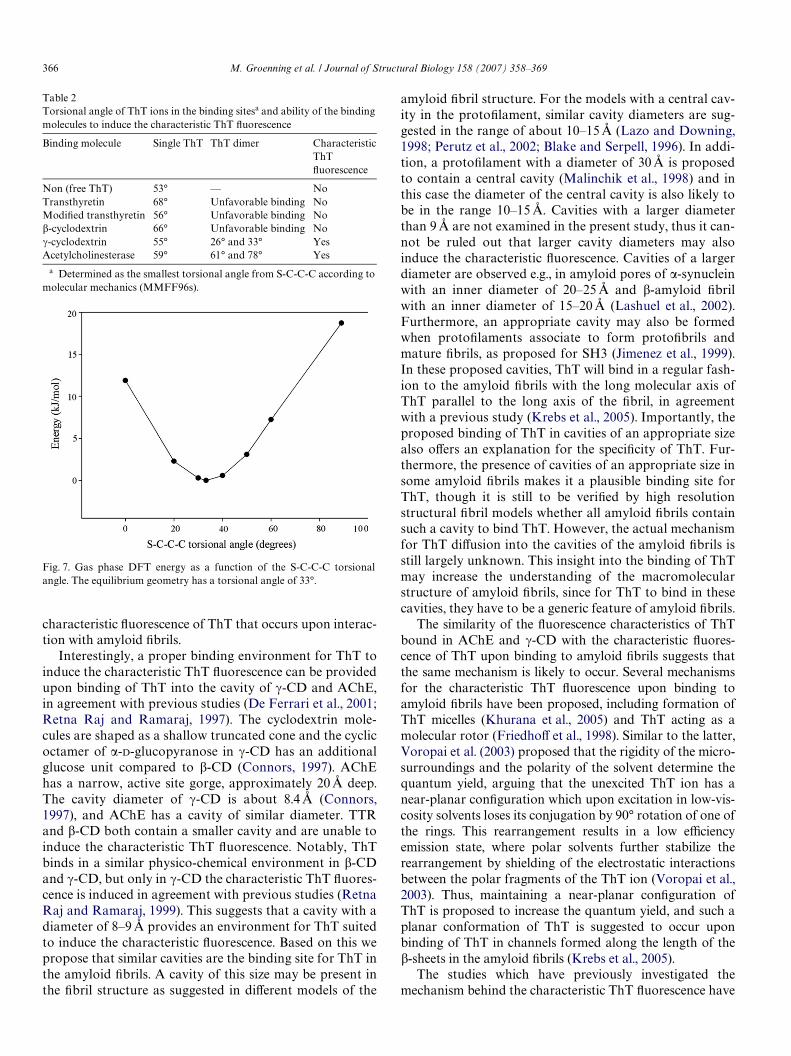
Finally, the conformation of the ThT structure in thecomplexes was examined. Especially the torsional angle ofthe rotational bond dividing the two planar parts, the ben-zothiazole and benzamine rings, of ThT (Fig. 1) is expectedto aVect the Xuorescence, since locking of a conjugated sys-tem in a certain position may inXuence the emission spectraas well as the Xuorescence quantum yield (Lakowicz, 1999).The smallest torsional angle measured from S-C-C-C onthe ThT conformation with the highest GOLD score basedon molecular modelling is presented in Table 2. Free ThT
prefers a non-planar conformation with a torsional anglebetween the two aromatic systems of 53° and 33° accordingto molecular mechanics (MMFF96s) and quantummechanics (DFT B3LYP) calculations, respectively. Such adiVerence in the torsional angles is associated with a smallenergy diVerence, since quantum mechanics calculationsshow that the energy requirement for rotation around thecentral bond in ThT is small and that a §20° deformationof ThT from the preferred conformation requires less than4 kJ/mol (Fig. 7). This energy requirement is even less whensolvent (water) eVects are taken into consideration, e.g., insolvent compared to gas phase, the conformation with a S-C-C-C torsional angle of 90° is stabilized by around 4 kJ/mol as compared to those with small torsional angles(<60°). The torsional angle of 53° for free ThT wasunchanged in most binding molecules when ThT wasbound as a monomer. Exceptions were when one ThT ionwas bound to TTR and �-CD, where the torsional angleswere determined to be 68° and 66°, respectively. A changein the torsional angle of ThT was also observed when twoThT ions were bound in a cavity, but in AChE, the ThTdimers had increased torsional angles compared to freeThT, while the opposite was observed for ThT dimers in �-CD (Table 2). The dimer forms generated are suited fordetermining whether the cavity is able to accommodate adimer, but other ThT dimer conformations may also bepossible, e.g., just partly overlapping ThT ions. This mayaVect the torsional angle, but the largest impact on the tor-
Fig. 5. The energetically most favorable binding of one ThT ion into the �-sheet cavity of TTR (upper), and the energetically most favorable binding of aThT dimer into the cavity of AChE (lower). The molecular modelling is illustrated in two views rotated 90° where a ThT ion is presented with a green sur-face; the ThT ions have a red and a green surface when present as a dimer.
M. Groenning et al. / Journal of Structural Biology 158 (2007) 358–369 365
sional angle is expected when packed as the four generatedThT dimers.
4. Discussion
Most likely, a common characteristic of the amyloidWbril structure is important for the ThT binding mode.Besides the proposed presence of cavities, extended �-sheetshave also been suggested to be characteristic for the amy-loid Wbril structure. In the binding pocket of TTR, ThT issurrounded by an eight-stranded �-sandwich. The bindingof ThT to TTR was demonstrated by a signiWcant changein Xuorescence anisotropy, and a favorable binding of just
one ThT ion was shown via molecular modelling. Molecu-lar modelling of the ThT binding to TTR with a more openentrance to the �-sheet cavity further conWrmed theseresults. The characteristic Xuorescence of ThT was notobserved when ThT was bound to TTR, in agreement withan earlier study on ThT binding to native tetramer,unfolded monomer, and A-state monomer of TTR (Lind-gren et al., 2005). TTR is not the only �-sheet-rich proteinfailing to induce the characteristic ThT Xuorescence. Forexample, neither chymotrypsin, bovine IgG, and concanav-alin A induced the characteristic ThT Xuorescence (LeVine,1995). Therefore, the presence of a secondary �-sheet struc-ture, even as a �-sheet cavity, is insuYcient to induce the
Fig. 6. The energetically most favorable binding of one ThT in complex with two �-CDs (upper), one ThT in complex with two �-CDs (middle), and twoThT in complex with two �-CDs (lower). The molecular modelling is illustrated in two views rotated 90° where a ThT ion is presented with a green surface;the ThT ions have a red and a green surface when present as a dimer.
366 M. Groenning et al. / Journal of Structural Biology 158 (2007) 358–369
characteristic Xuorescence of ThT that occurs upon interac-tion with amyloid Wbrils.
Interestingly, a proper binding environment for ThT toinduce the characteristic ThT Xuorescence can be providedupon binding of ThT into the cavity of �-CD and AChE,in agreement with previous studies (De Ferrari et al., 2001;Retna Raj and Ramaraj, 1997). The cyclodextrin mole-cules are shaped as a shallow truncated cone and the cyclicoctamer of �-D-glucopyranose in �-CD has an additionalglucose unit compared to �-CD (Connors, 1997). AChEhas a narrow, active site gorge, approximately 20 Å deep.The cavity diameter of �-CD is about 8.4 Å (Connors,1997), and AChE has a cavity of similar diameter. TTRand �-CD both contain a smaller cavity and are unable toinduce the characteristic ThT Xuorescence. Notably, ThTbinds in a similar physico-chemical environment in �-CDand �-CD, but only in �-CD the characteristic ThT Xuores-cence is induced in agreement with previous studies (RetnaRaj and Ramaraj, 1999). This suggests that a cavity with adiameter of 8–9 Å provides an environment for ThT suitedto induce the characteristic Xuorescence. Based on this wepropose that similar cavities are the binding site for ThT inthe amyloid Wbrils. A cavity of this size may be present inthe Wbril structure as suggested in diVerent models of the
Table 2Torsional angle of ThT ions in the binding sitesa and ability of the bindingmolecules to induce the characteristic ThT Xuorescence
a Determined as the smallest torsional angle from S-C-C-C according tomolecular mechanics (MMFF96s).
Binding molecule Single ThT ThT dimer CharacteristicThT Xuorescence
Non (free ThT) 53° — NoTransthyretin 68° Unfavorable binding NoModiWed transthyretin 56° Unfavorable binding No�-cyclodextrin 66° Unfavorable binding No�-cyclodextrin 55° 26° and 33° YesAcetylcholinesterase 59° 61° and 78° Yes
Fig. 7. Gas phase DFT energy as a function of the S-C-C-C torsionalangle. The equilibrium geometry has a torsional angle of 33°.
amyloid Wbril structure. For the models with a central cav-ity in the protoWlament, similar cavity diameters are sug-gested in the range of about 10–15 Å (Lazo and Downing,1998; Perutz et al., 2002; Blake and Serpell, 1996). In addi-tion, a protoWlament with a diameter of 30 Å is proposedto contain a central cavity (Malinchik et al., 1998) and inthis case the diameter of the central cavity is also likely tobe in the range 10–15 Å. Cavities with a larger diameterthan 9 Å are not examined in the present study, thus it can-not be ruled out that larger cavity diameters may alsoinduce the characteristic Xuorescence. Cavities of a largerdiameter are observed e.g., in amyloid pores of �-synucleinwith an inner diameter of 20–25 Å and �-amyloid Wbrilwith an inner diameter of 15–20 Å (Lashuel et al., 2002).Furthermore, an appropriate cavity may also be formedwhen protoWlaments associate to form protoWbrils andmature Wbrils, as proposed for SH3 (Jimenez et al., 1999).In these proposed cavities, ThT will bind in a regular fash-ion to the amyloid Wbrils with the long molecular axis ofThT parallel to the long axis of the Wbril, in agreementwith a previous study (Krebs et al., 2005). Importantly, theproposed binding of ThT in cavities of an appropriate sizealso oVers an explanation for the speciWcity of ThT. Fur-thermore, the presence of cavities of an appropriate size insome amyloid Wbrils makes it a plausible binding site forThT, though it is still to be veriWed by high resolutionstructural Wbril models whether all amyloid Wbrils containsuch a cavity to bind ThT. However, the actual mechanismfor ThT diVusion into the cavities of the amyloid Wbrils isstill largely unknown. This insight into the binding of ThTmay increase the understanding of the macromolecularstructure of amyloid Wbrils, since for ThT to bind in thesecavities, they have to be a generic feature of amyloid Wbrils.
The similarity of the Xuorescence characteristics of ThTbound in AChE and �-CD with the characteristic Xuores-cence of ThT upon binding to amyloid Wbrils suggests thatthe same mechanism is likely to occur. Several mechanismsfor the characteristic ThT Xuorescence upon binding toamyloid Wbrils have been proposed, including formation ofThT micelles (Khurana et al., 2005) and ThT acting as amolecular rotor (FriedhoV et al., 1998). Similar to the latter,Voropai et al. (2003) proposed that the rigidity of the micro-surroundings and the polarity of the solvent determine thequantum yield, arguing that the unexcited ThT ion has anear-planar conWguration which upon excitation in low-vis-cosity solvents loses its conjugation by 90° rotation of one ofthe rings. This rearrangement results in a low eYciencyemission state, where polar solvents further stabilize therearrangement by shielding of the electrostatic interactionsbetween the polar fragments of the ThT ion (Voropai et al.,2003). Thus, maintaining a near-planar conWguration ofThT is proposed to increase the quantum yield, and such aplanar conformation of ThT is suggested to occur uponbinding of ThT in channels formed along the length of the�-sheets in the amyloid Wbrils (Krebs et al., 2005).
The studies which have previously investigated themechanism behind the characteristic ThT Xuorescence have

M. Groenning et al. / Journal of Structural Biology 158 (2007) 358–369 367
mainly focused on the eVect of solvent dielectric constantand solvent viscosity (FriedhoV et al., 1998; LeVine, 1999;Voropai et al., 2003). The Xuorescence quantum yield ofThT is observed to be dependent on viscosity when aproper environment, i.e., with a lower dielectric constant,surrounds ThT (FriedhoV et al., 1998). When ThT is boundin a cavity, rotation of the benzothiazole and benzaminerings in ThT is expected to be restricted, like in high viscos-ity surroundings. The examination of the Xuorescence char-acteristics of ThT showed that some cavities succeeded ininducing the characteristic Xuorescence of ThT. However,this was not the case for a tight packing of ThT in �-CDand the �-sheet cavity in TTR. Therefore, restriction of therotations around the bond between the benzothiazole andbenzamine rings by being bound in a rigid environment isprobably not suYcient to induce the characteristic ThTXuorescence. The inXuence of the dielectric constant on thequantum yield has not been fully clariWed, though no rela-tionship has been proposed (FriedhoV et al., 1998; LeVine,1995). However, a diVerence in dielectric constant can aVectthe wavelength of the excitation and emission maxima, e.g.,illustrated by the red shift of the absorption of about 20 nmfor ThT in dichlorobenzene compared to in ethanol(Voropai et al., 2003). Therefore, the minor diVerences inthe excitation and emission maxima for ThT bound toAChE, �-CD and insulin amyloid Wbrils may be attributedto diVerences in dielectric constant of the surroundingenvironment.
The binding stoichiometry of ThT in the cavities induc-ing the characteristic ThT Xuorescence, versus the cavitiesthat do not, may be essential for elucidation of the mecha-nism behind the characteristic Xuorescence. The cavitiesshown to induce the characteristic ThT Xuorescence arelarge enough to accommodate two ThT ions and a favor-able binding of both one and two ions could take place atthe binding site according to molecular modelling. ForAChE, a Job plot also indicated that the complex inducingthe characteristic Xuorescence may be one ThT ion, or pos-sibly two ThT ions, binding at the active site of AChE. For�-CD, ITC also indicated formation of 1:1 (or 2:2) or 2:1ThT:�-CD complexes, and thereby indicates that the �-CDcavity is able to accommodate one or two ThT ions. Inter-estingly, the cavities of both �-CD and AChE, inducing thecharacteristic ThT Xuorescence, are large enough for twoThT ions to bind, and they may do so, in contrast to thesmaller cavities of TTR and �-CD which were unable toyield the characteristic Xuorescence of ThT. A 2:2 complexof ThT and �-CD has previously been proposed to be thecomplex inducing the characteristic Xuorescence (RetnaRaj and Ramaraj, 1999). The actual presence of a 2:2 com-plex is diYcult to determine by ITC. Furthermore, analysisof the stoichiometry of the complex inducing the character-istic Xuorescence cannot be done by, e.g., a Job plot, since aconcentration of �-CD well above the dissociation constantis needed, which is impossible due to the limited solubilityof �-CD. However, both ITC and molecular modelling indi-cate that the 2:2 complex can be formed. Importantly, the
2:2 complex, and for example not the 2:1 ThT:�-CD com-plex, induces the characteristic Xuorescence (Retna Raj andRamaraj, 1999), meaning that the cavity needs to be longenough to cover the entire length of the ThT ions for theXuorescence to occur. According to ITC and molecularmodelling of ThT binding to �-CD, a 1:1 or a 1:2 ThT:�-CD complex can be formed. The presence of a 1:2 complex,where the entire ThT ion is covered with �-CD, has alsopreviously been shown (Retna Raj and Ramaraj, 1999).Therefore, lack of covering the entire ThT ion cannotexplain that �-CD is unable to induce the characteristicThT Xuorescence. This further indicates that the dimer for-mation is crucial.
The possible ThT dimer formation indicates that therepulsion of the positive charge on the two ThT ions doesnot dominate the interaction in the cavities. This may bedue to a diVuse location of the positive charge in the benzo-thiazole region making a dipole–dipole interaction possibleupon association in a cavity. Furthermore, the ITC experi-ments showed that ThT binding to �-CD and �-CD waspredominantly entropically driven, a characteristic of bind-ing primarily driven by hydrophobic eVects. The thermody-namics also suggest that the formation of inclusioncomplexes is a weak exothermic process indicating thatsome favorable hydrogen bonding, van der Waals and/orelectrostatic interactions take place. However, it should benoted that the 2:2 complex of ThT and �-CD which inducesthe characteristic Xuorescence may not be formed duringthe ITC experiment, or is only present in small amounts,and that the thermodynamics thus are dominated byanother complex.
Molecular modelling indicates that the two ThT ions inthe cavity formed of two aligned �-CDs adopt conforma-tions diVerently from the global energy conformation(Table 2), which suggests that the binding imposes strain onThT and thereby restricts the Xexibility of ThT, like in highviscosity surroundings. It is interesting to note that a ThTdimer in complex with �-CD is more planar than free ThT,while the opposite is observed for a ThT dimer in complexwith AChE. Binding of one ThT in the active site cavity ofAChE showed a similar, or slightly less planar, globalenergy conformation as free ThT. A less planar conforma-tion of a single ThT is also observed in TTR and �-CDwithout inducing the characteristic ThT Xuorescence(Table 2). Thus, no direct correlation between the degree ofdeformation and the occurrence of Xuorescence is observedin the present studies. A correlation would be expected iflocking of ThT in a speciWc conformation is the genericmechanism behind the characteristic ThT Xuorescence.Another plausible mechanism for the characteristic ThTXuorescence in �-CDs and AChE is an excimer formationof ThT. Excimer formation is observed for molecules with astructure similar to ThT, e.g., 2-phenylindone with a mono-mer emission centered near 390 nm and a red-shifted exci-mer emission at 420 nm (Lakowicz, 1999). A red shift is alsoobserved for the characteristic ThT Xuorescence. Such ashift most often accompanies an excimer formation due to

368 M. Groenning et al. / Journal of Structural Biology 158 (2007) 358–369
the formation of a more conjugated system. An excimerformation of ThT has also previously been proposed forthe 2:2 complex with �-CD (Retna Raj and Ramaraj, 1997)and for ThT in DNA (Ilanchelian and Ramaraj, 2004).
5. Conclusions
The study on the binding mode of ThT in diVerent cavitystructures showed that the characteristic ThT Xuorescenceis induced in cavities with a diameter of about 8–9 Å and alength able to accommodate the entire length of the ThTion. Smaller cavities, just able to bind one ThT ion, wereunable to provide a proper binding environment to inducethe characteristic Xuorescence. This was also the case forthe �-sheet-rich cavity of TTR. The importance of a cavitydiameter capable of binding two ThT ions, among others,indicates that an excimer formation is a plausible mecha-nism for the occurrence of the characteristic Xuorescence.We propose a similar binding mode of ThT in amyloidWbrils, where cavities of an appropriate size running parallelto the Wbril axis have previously been proposed in severalamyloid Wbril models.
Acknowledgments
The authors thank Henrik Olsen for performing the 1HNMR analysis and the Tripos Associates for supportregarding access to Sybyl. Apotekerfonden af 1991 isacknowledged for funding the Spex Fluorolog 3-22 Xuores-cence spectrometer. The project is supported by the DrugResearch Academy (M.G.) and Carlsberg Foundation(L.O.).
References
Alexander, J.M., Clark, J.L., Brett, T.J., Stezowski, J.J., 2002. Supramolecu-lar chemistry and self-assembly special feature: chiral discrimination incyclodextrin complexes of amino acid derivatives: �-cyclodextrin/N-acetyl-L-phenylalanine and N-acetyl-D-phenylalanine complexes. Proc.Natl. Acad. Sci. USA 99, 5115–5120.
Barone, V., Cossi, M., 1998. Quantum calculation of molecular energiesand energy gradients in solution by a conductor solvent model. J. Phys.Chem. A 102, 1995–2001.
Becke, A.D., 1993a. Density-functional thermochemistry III. The role ofexact exchange. J. Chem. Phys. 98, 5648–5652.
Becke, A.D., 1993b. A new mixture of Hartree-Fock and local density-functional theories. J. Chem. Phys. 98, 1372–1377.
Blake, C., Serpell, L., 1996. Synchrotron X-ray studies suggest that the coreof the transthyretin amyloid Wbril is a continuous �-sheet helix. Struc-ture 4, 989–998.
Brange, J., Andersen, L., Laursen, E.D., Meyn, G., Rasmussen, E., 1997.Toward understanding insulin Wbrillation. J. Pharm. Sci. 86, 517–525.
Cohen, A.S., Calkins, E., 1959. Electron microscopic observations on aWbrous component in amyloid of diverse origins. Nature 183, 1202–1203.
Connors, K.A., 1997. The stability of cyclodextrin complexes in solution.Chem. Rev. 97, 1325–1357.
Cossi, M., Rega, N., Scalmani, G., Barone, V., 2003. Energies, structures,and electronic properties of molecules in solution with the C-PCM sol-vation model. J. Comput. Chem. 24, 669–681.
De Ferrari, G.V., Mallender, W.D., Inestrosa, N.C., 2001. ThioXavin T is aXuorescent probe of the acetylcholinesterase peripheral site that revealsconformational interactions between the peripheral and acylation sites.J. Biol. Chem. 276, 23282–23287.
de Oliveira Marques, P.R.B., Nunes, G.S., dos Santos, T.C.R., Andreescu,S., Marty, J.L., 2004. Comparative investigation between acetylcholin-esterase obtained from commercial sources and genetically modiWedDrosophila melanogaster: Application in amperometric biosensors formethamidophos pesticide detection. Biosens. Bioelectron. 20, 825–832.
Ding, J., Steiner, T., Saenger, W., 1991. �-Cyclodextrin 1-propanol clath-rate hydrate. Acta Crystallogr. B-Struct. Sci. 47, 731–738.
Dobson, C.M., 2003. Protein folding and misfolding. Nature 426, 884–890.Eanes, E.D., Glenner, G.G., 1968. X-ray diVraction studies on amyloid Wla-
ments. J. Histochem. Cytochem. 16, 673–677.FriedhoV, P., Schneider, A., Mandelkow, E.-M., Mandelkow, E., 1998.
Rapid assembly of Alzheimer-like paired helical Wlaments from micro-tubule-associated protein tau monitored by Xuorescence in solution.Biochemistry 37, 10223–10230.
Frisch, M.J., Trucks, G.W., Schlegel, H.B., Scuseria, G.E., Robb, M.A.,Cheeseman, J.R., Montgomery, Jr.J.A., Vreven, T., Kudin, K.N.,Burant, J.C., Millam, J.M., Iyengar, S.S., Tomasi, J., Barone, V., Men-nucci, B., Cossi, M., Scalmani, G., Rega, N., Petersson, G.A., Nakatsuji,H., Hada, M., Ehara, M., Toyota, K., Fukuda, R., Hasegawa, J., Ishida,M., Nakajima, T., Honda, Y., Kitao, O., Nakai, H., Klene, M., Li, X.,Knox, J.E., Hratchian, H.P., Cross, J.B., Bakken, V., Adamo, C., Jara-millo, J., Gomperts, R., Stratmann, R.E., Yazyev, O., Austin, A.J.,Cammi, R., Pomelli, C., Ochterski, J.W., Ayala, P.Y., Morokuma, K.,Voth, G.A., Salvador, P., Dannenberg, J.J., Zakrzewski, V.G., Dap-prich, S., Daniels, A.D., Strain, M.C., Farkas, O., Malick, D.K.,Rabuck, A.D., Raghavachari, K., Foresman, J.B., Ortiz, J., Cui, Q.,Baboul, A.G., CliVord, S., Cioslowski, J., Stefanow, B.B., Liu, G., Lias-henko, A., Piskorz, P., Komaromi, I., Martin, R.L., Fox, D.J., Keith, T.,Al-Laham, M.A., Peng, C.Y., Nanayakkara, A., Challacombe, M., Gill,P.M.W., Johnson, B., Chen, W., Wong, M.W., Gonzalez, C., Pople, J.A.,2004. Gaussian 03, Revision C.02, Gaussion, Inc., Wallingford CT.
Goldsbury, C.S., Wirtz, S., Müller, S.A., Sunderji, S., Wicki, P., Aebi, U.,Frey, P., 2000. Studies on the in vitro assembly of Ab 1-40: implicationsfor the search for Ab Wbril formation inhibitors. J. Struct. Biol. 130,217–231.
Grillo, A.O., Edwards, K.T., Kashi, R.S., Shipley, K.M., Hu, L., Besman,M.J., Middaugh, C.R., 2001. Conformational origin of the aggregationof recombinant human factor VIII. Biochemistry 40, 586–595.
Halgreen, T.A., 1999. MMFF VI. MMFF94s option for energy minimiza-tion studies. J. Comput. Chem. 20, 720–729.
Hornberg, A., Eneqvist, T., Olofsson, A., Lundgren, E., Sauer-Eriksson,A.E., 2000. A comparative analysis of 23 structures of the amyloido-genic protein transthyretin. J. Mol. Biol. 302, 649–669.
Huang, C.Y., 1982. Determination of binding stoichiometry by the contin-uous variation method: the Job plot. Methods Enzymol. 87, 509–525.
Ilanchelian, M., Ramaraj, R., 2004. Emission of thioXavin T and its controlin the presence of DNA. J. Photochem. Photobiol. A 162, 129–137.
Jimenez, J.L., Guijarro, J.I., Orlova, E., Zurdo, J., Dobson, C.M., Sunde,M., Saibil, H.R., 1999. Cryo-electron microscopy structure of an SH3amyloid Wbril and model of the molecular packing. EMBO J. 18, 815–821.
Jimenez, J.L., Nettleton, E.J., Bouchard, M., Robinson, C.V., Dobson,C.M., Saibil, H.R., 2002. The protoWlament structure of insulin amyloidWbrils. Proc. Natl. Acad. Sci. USA 99, 9196–9201.
Jones, G., Willett, P., Glen, R.C., Leach, A.R., Taylor, R., 1997. Develop-ment and evaluation of a generic algoritm for Xexible docking. J. Mol.Biol. 267, 727–748.
Khurana, R., Coleman, C., Ionescu-Zanetti, C., Carter, S.A., Krishna, V.,Grover, R.K., Roy, R., Singh, S., 2005. Mechanism of thioXavin T bind-ing to amyloid Wbrils. J. Struct. Biol. 151, 229–238.
Khurana, R., Ionescu-Zanetti, C., Pope, M., Li, J., Nielson, L., Ramirez-Alvarado, M., Regan, L., Fink, A.L., Carter, S.A., 2003. A generalmodel for amyloid Wbril assembly based on morphological studiesusing atomic force microscopy. Biophys. J. 85, 1135–1144.

M. Groenning et al. / Journal of Structural Biology 158 (2007) 358–369 369
Krebs, M.R.H., Bromley, E.H.C., Donald, A.M., 2005. The binding of thio-Xavin-T to amyloid Wbrils: localisation and implications. J. Struct. Biol.149, 30–37.
Kryger, G., Silman, I., Sussman, J.L., 1999. Structure of acetylcholinester-ase complexed with E2020 (Aricept): implications for the design of newanti-Alzheimer drugs. Struct. Fold. Des. 7, 297–307.
Lakowicz, J.R., 1999. Principles of Fluorescence Spectroscopy, second ed.Kluwer Academic/Plenum Publishers, New York.
Lashuel, H.A., Hartley, D., Petre, B.M., Walz, T., Lansbury, P.T., 2002.Neurodegenerative disease: amyloid pores from pathogenic mutations.Nature 418, 291.
Lazo, N.D., Downing, D.T., 1998. Amyloid Wbrils may be assembled from�-helical protoWbrils. Biochemistry 37, 1731–1735.
Lee, C.T., Yang, W.T., Parr, R.G., 1988. Development of the Colle-Salvetticorrelation-energy formula into a functional of the electron-density.Phys. Rev. B 37, 785–789.
LeVine III, H., 1995. ThioXavin T interaction with amyloid �-sheet struc-tures. Int. J. Exp. Clin. Invest. 2, 1–6.
LeVine III, H., 1999. QuantiWcation of �-sheet amyloid Wbril structureswith thioXavin T. Methods Enzymol. 309, 274–284.
LeVine III, H., 1993. ThioXavine T interaction with synthetic Alzheimer’sdisease �-amyloid peptides: detection of amyloid aggregation in solu-tion. Protein Sci. 2, 404–410.
Lindgren, M., Sörgjerd, K., Hammarström, P., 2005. Detection and charac-terization of aggregates, preWbrillar amyloidogenic oligomers, and pro-toWbrils using Xuorescence spectroscopy. Biophys. J. 88, 4200–4212.
Malinchik, S.B., Inouye, H., Szumowski, K.E., Kirschner, D.A., 1998.Structural analysis of Alzheimer’s �(1-40) amyloid: protoWlamentassembly of tubular Wbrils. Biophys. J. 74, 537–545.
Naiki, H., Higuchi, K., Hosokawa, M., Takeda, T., 1989. Fluorometricdetermination of amyloid Wbrils in vitro using the Xourescent dye, Thi-oXavin T. Anal. Biochem. 177, 244–249.
Nielsen, L., Frokjaer, S., Carpenter, J.F., Brange, J., 2001. Studies of thestructure of insulin Wbrils by fourier transform infrared (FTIR) spec-troscopy and electron microscopy. J. Pharm. Sci. 90, 29–37.
Onoue, S., Ohshima, K., Debari, K., Koh, K., Shioda, S., Iwasa, S.,Kashimoto, K., Yajima, T., 2004. Mishandling of the therapeutic pep-tide glucagon generates cytotoxic amyloidogenic Wbrils. Pharm. Res.21, 1274–1283.
Pedersen, J.S., Dikov, D., Flink, J.L., Hjuler, H.A., Christiansen, G., Otzen,D.E., 2006. The changing face of glucagon Wbrillation: structural poly-morphism and conformational imprinting. J. Mol. Biol. 355, 501–523.
Pepys, M.B., 2006. Amyloidosis. Annu. Rev. Med. 57, 223–241.Perutz, M.F., Finch, J.T., Berriman, J., Lesk, A., 2002. Amyloid Wbers are
water-Wlled nanotubes. Proc. Natl. Acad. Sci. USA 99, 5591–5595.Peterson, S.A., Klabunde, T., Lashuel, H.A., Purkey, H., Sacchettini, J.C.,
Kelly, J.W., 1998. Inhibiting transthyretin conformational changes thatlead to amyloid Wbril formation. Proc. Natl. Acad. Sci. USA 95, 12956–12960.
Retna Raj, C., Ramaraj, R., 1997. �-Cyclodextrin induced intermolecularexcimer formation of thioXavin T. Chem. Phys. Lett. 273, 285–290.
Retna Raj, C., Ramaraj, R., 1999. InXuence of cyclodextrin complexationon the emission of thioXavin T and its oV-on control. J. Photochem.Photobiol. A 122, 39–46.
Schormann, N., Murrell, J.R., Benson, M.D., 1998. Tertiary structures ofamyloidogenic and non-amyloidogenic transthyretin variants: newmodel for amyloid Wbril formation. Amyloid 5, 175–187.
Serpell, L.C., Blake, C.C., Fraser, P.E., 2000. Molecular structure of a Wbril-lar Alzheimer’s A� fragment. Biochemistry 39, 13269–13275.
Tellinghuisen, J., 2005. Optimizing experimental parameters in isothermaltitration calorimetry. J. Phys. Chem. B 109, 20027–20035.
Turnbull, W.B., Daranas, A.H., 2003. On the value of c: can low aYnitysystems be studied by isothermal titration calorimetry? J. Am. Chem.Soc. 125, 14859–14866.
Voropai, E.S., Samtsov, M.P., Kaplevskii, K.N., Maskevich, A.A., Stepuro,V.I., Povarova, O.I., Kuznetsova, I.M., Turoverov, K.K., Fink, A.L.,Uversky, V.N., 2003. Spectral properties of thioXavin T and its com-plexes with amyloid Wbrils. J. Appl. Spectrosc. 70, 868–874.
Wang, W., 2005. Protein aggregation and its inhibition in biopharmaceu-tics. Int. J. Pharm. 289, 1–30.


















