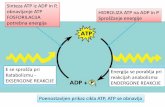
Sinteza ATP iz ADP in P, obnavljanje ATP HIDROLIZA ATP na ...
Upload
paulina-francisCategory
view
218download
2
Structure of ATP and NAD

Purposes/Roles/Functions of Glycolysis
• 1. Produce ATP
• 2. Feed ATP production via PDH, TCA, ET, OP
• 3. Feed production of TCA intermediates
• 4. Feed FA synthesis via PDH
• 5. Feed AA synthesis via TCA intermediates


Glycolysis Net Rxn (adding all rxns)
• ATP + G G6P + ADP• G6P F6P• ATP + F6P F1,6BP + ADP• F1,6BP GAP + DHAP• DHAP GAP• 2Pi + 2NAD+ + 2GAP 2 (1,3BPG) + 2NADH• 2 (1,3BPG) + 2ADP 2 (3PG) + 2ATP• 2 (3PG) 2 (2PG)• 2 (2PG) 2 PEP • 2 PEP + 2 ADP 2pyr + 2ATP
___________________________________________• G + 2Pi + 2 ADP + 2NAD+ 2pyr + 2 ATP + 2NADH

Lactate Dehydrogenase (LDH)
• Purpose of LDH: to covert the NADH produced in GAPDH back to NAD+ for use in GAPDH.
• This is required to keep glycolysis (G’lys) going rapidly during rapid muscle activity.
• The sum: [NAD+] + [NADH] is small. All of it would soon be converted to NADH if it was not recycled.

LDH
• In muscle, NADH is converted to NAD+ via ET: 2NADH + 2H+ + O2 ---> 2H2O + 2NAD+;
• but in active muscle, NADH production exceeds the capacity to deliver O2 to ET, so LDH also
converts NADH to NAD+.
• This removes the limitation that O2 delivery rate
would place on the maximum rate of muscle activity: we can sprint and go into O2 debt rather
than just jog.

LDH• LDH rxn: • pyruvate + NADH + H+ <-----> lactate + NAD+
• In active muscle, the capacity to consume O2 exceeds ability to deliver, so [O2] decreases: anaerobic conditions.
• But O2 delivery by heart, lungs, and Hb is at the maximum possible rate and O2 consumption in muscle is the highest ever: aerobic conditions.
• Anaerobic G’lys produces 2 ATP per G converted to lactate at a high rate only when aerobic G’lys is rapid (and only in addition to aerobic G’lys) . Aerobic glycolysis (working with PDH, TCA, ET, and OP) produces 36 or 38 ATP per G converted to carbon dioxide.

Fates of Pyruvate
• Depending on the organism in question and the conditions under which it is operating, pyruvate may be converted to:
• Lactate
• Acetyl coenzyme A (ACoA)
• Ethanol
• Oxaloacetate
• Alanine

Gluconeogenesis• Gluconeogenesis: in the liver, pyruvate (or
oxaloacetate) is converted to G to increase [G] in the blood:
• 1. in short-term CH2O starvation: muscle protein is hydrolysed to AAs which go to the blood and to the liver where they are converted to oxac G to supply G to the brain when [G] is low in the blood.
• 2. in rapid muscle activity, lactate from muscles is converted back to G to go back to the muscles
• Reactions: 7 of the reactions are the same reactions (same Es) as in glycolysis: they just go faster in reverse during gluconeogenesis (G’neo). (Q becomes greater than K)
• The irreversible reactions of glycolysis (G’lysis) are bypassed (NOT reversed)

Net Reaction for G’neo• To bypass PK (twice/G) consumes 2ATP +
2GTP.
• Reverse of PGK (2(1,3BPG) + 2ADP 2 (3PG) + 2ATP) twice/G consumes 2ATP.
• No ATP is produced in the bypass rxns of G’neo, nor in the reversed rxns of G’lys.
• So the net rxn for G’neo is:
• 2pyr + 4ATP + 2GTP + 2NADH G + 4ADP + 2GDP + 6Pi + 2NAD+

Energy Released Uphill and Down?• Both glycolysis and gluconeogenesis are energy
releasing processes that occur rapidly when their regulated enzymes are active. But the enzymes have no effect on the energy, so how can both directions (G 2pyr; 2pyr G) have negative ΔGs?
• The net reaction for glycolysis has ΔG ~ -100 kJ: G + 2ADP + 2Pi + 2NAD+ 2pyr + 2ATP + 2NADH
• And 2ATP 2ADP + 2Pi has ΔG ~ -100 kJ; so the sum: G + 2NAD+ 2pyr + 2NADH has ΔG ~ -200 kJ
• So 2pyr + 2NADH G + 2NAD+ has ΔG ~ +200 kJ• Add: 4ATP + 2GTP 4ADP + 2GDP + 6Pi ~ -300 kJ• This yields ~ -100 kJ for the net rxn for G”neo

Energy Released Uphill and Down?
• So, how can both directions (G 2pyr; 2pyr G) have negative ΔGs?
• G 2pyr releases about 200 kJ. Of this, 100 is used to make 2ATP and the other 100 is released in G’lys.
• 2pyr G consumes 200 kJ, so 6 ATP equivalents (4ATP + 2GTP) are used to “pay” that 200 kJ and have 100 kJ left to be released.

LDH
• LDH rxn: • pyruvate + NADH + H+ <-----> lactate + NAD+
• Q = [lactate][NAD+] / [pyruvate][NADH][H+]• Active Muscle: [pyruvate], [H+], and [NADH] are
all high from rapid production in G’lys; this pushes the LDH rxn to the right (LeChatelier).
• Also, [NAD+] is low from rapid use in glycolysis so the concentrations of these four substances make the value of Q low.

Liver LDH• Glucagon causes liver enzymes to convert
pyruvate to glucose in gluconeogenesis (G’neo).
• Glucagon is secreted in response to adrenalin or epinephrine, which is secreted during rapid muscle activity.
• Cori cycle: • Lactate is transported in blood from muscle to
liver, where it is converted back to G via G’neo. • Then G goes back out into the blood, and is
taken back into muscle cells, where it is then used or stored for later.

Liver LDH
• Liver: [pyr], [H+], and [NADH] are all low from rapid consumption in hormonally activated G’neo and [NAD+] is high from rapid production in G’neo, so Q is high.
• In gluconeogenesis, GAPDH is reversed (as are all reversible reactions of glycolysis) so NAD+ is produced, NADH is consumed.
• LDH rxn: pyruvate + NADH + H+ <-----> lactate + NAD+
• Q = [lactate][NAD+] / [pyruvate][NADH][H+]

Direction of LDH• Sign of ΔG and direction of LDH rxn depends on Q
vs K:• ΔG0’ = -RT ln K• ΔG = ΔG0’ + RT ln Q = -RT ln K + RT ln Q • = RT ln (Q/K)
• Q < K -ln; so ΔG is - and LDH rxn goes forward in active muscle where Q is low
• Q > K +ln; so ΔG is + and LDH rxn goes in reverse (L R) in liver at the same time as it is going forward in muscle
•• (If Q = K ΔG = 0; rxn is at EQ)

“Irreversible” Reactions of Glycolysis
• We can divide the reactions into two groups, based on their ΔG values: “large negatives” or “near zero”

“Irreversible” Reactions of Glycolysis• Unlike the LDH reaction, the reactions with large
negative values of ΔG never have a large enough value of Q to cause their net direction to be in reverse in a cell. For this reason they are said to be “physiologically irreversible”. (Like all enzymes, these enzymes do catalyze the reverse reaction. It has a nonzero rate, but a rate that is always slower than that of the forward rxn.)
• These reactions are catalyzed by regulated enzymes. • The reactions with values of ΔG that are near zero are
easily shifted one way or the other by changes in the value of Q that occur in cells.
• These “reversible” reactions are catalyzed by enzymes that are not regulated: these reactions are always moving rapidly toward equilibrium.

Regulation of Glycolysis Enzymes• Hexokinase (HXK) is inhibited by glucose-6-phosphate
(G6P)• Metabolic Relationship (MR): G6P is a product of the
HXK reaction (“direct product”)• Metabolic Logic (ML): if [G6P] is high, there is no need
for HXK to produce more.• What does this mean: “[G6P] is high”?
The regulated enzymes have evolved so that their affinity for their effectors causes minimal binding when the [effector] is barely enough to meet needs, but binding to most of the molecules of E when the [effector] is ample to meet needs.
• A similar situation exists for rate and the binding of substrate: the steep part of the rate curve usually extends across the usual range of [substrate] in the cell where that enzyme exists.

Regulation of Glycolysis Enzymes
• PFK is inhibited by ATP.
• MR: ATP is a product of glycolysis (in which PFK acts) and is a product of OP via processes fed by this pathway: PDH + TCA + ET + OP. (“indirect product”)
• ML: When the [ATP] is high there is no need for PFK to feed production of more.

Regulation of Glycolysis Enzymes• PFK is inhibited by citrate.• MR: citrate is an indirect product of PFK. Note the
metabolic relationship diagram (MRD):
• When [citrate] is high, this indicates that the TCA and all processes fed by it are well supplied, especially ET/OP and FA synthesis, and that PFK does not need to produce more rapidly.

Regulation of Glycolysis Enzymes• F1, 6BP is an activator of PK• MR: F1, 6BP is an indirect substrate (it will be
converted to PEP, the substrate) of PK. Note the MRD:
• ML: when [F1, 6BP] is high, PFK is rapid, there is plenty of PEP, there is a need for PK to be active to be in sync w/PFK
• HXK also “follows” PFK rate: When PFK is rapid, F6P is consumed rapidly and G6P is “pulled” to F6P, so [G6P] will not build up; HXK not inhibited
• HXK has a very high affinity for G; at “any” [G] the v 90% Vmax; main effect on rate is [G6P].

Reciprocal RegulationReciprocal Regulationof G’lys and G’neo Enzymes
• AMP activates PFK by overriding the ATP inhibition of PFK; and AMP inhibits FBPase.
• MR for PFK: its activity feeds ATP production in glycolysis AND in OP via PDH + TCA + ET + OP
• ML: if [AMP] is high then ATP is being consumed rapidly and needs to be produced, which it will be by PFK activation
• MR for FBPase: G’neo consumes 6ATP/G• ML: if [AMP] is high then ATP is being consumed
rapidly and needs to be produced; cell can’t afford to consume ATP in G’neo when ATP production is needed, FBPase/G’neo slows in response to AMP


AMP vs ATP Effects• PFK is more responsive to [AMP] than to [ATP]: • Adenylate Kinase (myokinase): 2ADP ↔ ATP + AMP• The [AMP] changes by a much larger factor (though
not by a greater amount) than [ATP].• This is because ATP is at a much higher concentration
and only changes by ~10%. The [AMP] is only about 1/50th (2%) of the [ATP] at rest; so if [ATP] goes from 2 mM to 1.8 mM, the [AMP] may go from about 0.04 mM to .16 - .20 mM, a factor of 4 or 5
• Because the factor change is greater for [AMP], it has more effect on PFK than ATP, overrides.
• (PFK has higher affinity for allosteric binding of AMP than it does for ATP)

Hormonal Regulation of G’lys and G’neo In Liver• Glucagon and insulin exert their effects on
these pathways by regulating the “tandem enzyme” See pages 649-650 in V+V.
• PFK2 and FBPase2 activities are at 2 different active sites on each of the polypeptide chains of “the tandem E”, a homodimer (two identical subunits).
• PFK2 catalyzes: F6P + ATP F2,6BP + ADP• FBPase2 catalyzes: F2,6BP F6P + Pi • F2,6BP activates PFK (overrides PFK inhibition
by ATP)• F2,6BP inhibits FBPase• There is only one purpose (and only one effect)
of F2,6BP: to regulate PFK and FBPase.

Hormonal Regulation of G’lys and G’neo In Liver
• Glucagon is secreted in short-term CH2O starvation in response to low [G] in blood; its purpose is to cause the liver to release G into the blood by activating G’neo and glycogenolysis.
• When the tandem E is phosphorylated in response to glucagon, only its FBPase2 is active (PFK2 is NOT); this causes the [F2,6BP] to decrease.
• This removes the inhibition of FBPase by F2,6BP and G’neo goes rapidly.
• G’lys is slowed by ATP inhibition of PFK.• G’lys has to go slow (NOT consume G when [G] is
low) so as to not oppose G’neo.

Hormonal Regulation of G’lys and G’neo In Liver
• Insulin is secreted after a CH2O meal in response to high [G] in blood; its purpose is to cause the liver to store fuel by G’lys, PDH, CS, and FA synthesis, and by glycogen synthesis.
• When the tandem E is dephosphorylated in response to insulin, only its PFK2 is active (FBPase2 is NOT) ); this causes the [F2,6BP] to increase.
• PFK is activated by the [F2, 6BP] increase, and glycolysis goes rapidly to “feed” FA synthesis (The E of FA synth is also dephos, active).
• FBPase is inhibited by F2,6BP, to keep it from opposing G’lys

Hormonal Regulation of G’lys and G’neo• Liver PK is also phosphorylated in response to
glucagon. This inhibits PK to slow G’lys and keep it from working against G’neo.
• Liver PK is also dephosphorylated in response to insulin. This activates PK to work with PFK to make G’lys go to feed FA synthesis.
• In heart muscle, the tandem E has the opposite response to phosphorylation, which occurs in response to adrenalin, as compared to tandem E in liver:
• The heart muscle tandem E’s PFK2 is active (its FBPase2 is inactive), so that the [F2, 6BP] increases and PFK is activated.
• The purpose here is to activate glycolysis, so that ATP can be rapidly produced (in G’lys and in PDH, TCA, and ET + OP) so that heart muscle can have rapid activity.

HXK is NOT
an ATPase

HXK is NOT an ATPase• HXK can bind ATP and G in either orderin either order, but if ATP
binds first, it’s not in contact with HXK’s catalytic side chains.
• G must bind to HXK and cause a conformational change to place ATP in position for catalysis (whenever ATP binds).
• Otherwise, ATP could bind to HXK (if HXK had only one, active conformation) and the γ phosphate would usually be transferred to water: ATP + H2O ADP + Pi (ATPase activity)
• ATP produced from fuel (food) burning would be wasted; and so would fuel (FOOD).
• Xylose converts HXK to an ATPase by binding to HXK and inducing the conformational change, putting ATP in contact with the catalytic groups. It lacks the #6C group of glucose (-H instead of -CH2OH); is otherwise the same. H2O can bind where the #6C would.


Substrate Cycling• Effect of “substrate cycling” on relative rate
of enzymes:
• Consider rapid PFK with high [AMP], say 90 units PFK, while FBPase is slow, say 1 unit; so net is 89 units of G consumed.
• At rest, with low [AMP], PFK will be much lower, say 10 units, and FBPase may be 9 units, then net is 1 unit G consumed.
• Difference is 89 fold net increase in G consumed, even though rates of G’lys and G’neo actually only differed by 9X


















