
Saggio in vitro sull'effetto di funghi ambientali e ... · 1.1 Classificazione e tassonomia ......
85
UNIVERSITÀ DEGLI STUDI DI PISA Dipartimento di Scienze Veterinarie Tesi di Laurea Magistrale in Medicina Veterinaria Saggio in vitro sull'effetto di funghi ambientali e estratti vegetali nei confronti delle uova di Ascaris suum Candidato: Federico Rossi Relatori: Dott. Roberto Amerigo Papini Prof.ssa Francesca Mancianti Controrelatore: Prof. ssa Stefania Perrucci ANNO ACCADEMICO 2013-14
-
Upload
hoangxuyen -
Category
Documents
-
view
214 -
download
0
Transcript of Saggio in vitro sull'effetto di funghi ambientali e ... · 1.1 Classificazione e tassonomia ......

UNIVERSITÀ DEGLI STUDI DI PISA
Dipartimento di Scienze Veterinarie
Tesi di Laurea Magistrale in Medicina Veterinaria
Saggio in vitro sull'effetto di funghi ambientali e estratti
vegetali nei confronti delle uova di Ascaris suum
Candidato: Federico Rossi Relatori: Dott. Roberto Amerigo Papini
Prof.ssa Francesca Mancianti
Controrelatore: Prof. ssa Stefania Perrucci
ANNO ACCADEMICO 2013-14

INDICE
RIASSUNTO.......................................................................................................................3
INTRODUZIONE...............................................................................................................5
CAPITOLO 1: ASCARIDIOSI DEL SUINO.....................................................................7
1.1 Classificazione e tassonomia.........................................................................................7
1.2 Descrizione morfologica..............................................................................................11
1.2.1 Parassita adulto..................................................................................................
1.2.2 Uova..................................................................................................................
1.2.3 Genoma.............................................................................................................
1.3 Ciclo biologica.............................................................................................................17
1.4 Immunità naturale e acquisita......................................................................................20
1.5 Patogenesi....................................................................................................................22
1.6 Segni clinici e rilievi anatomo-patologici....................................................................22
1.7 Epidemiologia e danni economici...............................................................................25
1.8 Tecniche diagnostiche..................................................................................................31
1.8.1 Diagnosi in allevamento....................................................................................
1.8.2 Diagnosi di laboratorio......................................................................................
1.8.3 Diagnosi in sede di macellazione.......................................................................
1.9 Prevenzione e controllo...............................................................................................34
1.9.1 Prevenzione........................................................................................................
1.9.2 Trattamenti antielmintici....................................................................................
1.9.3 Vaccinazione......................................................................................................
CAPITOLO 2: STRATEGIE ALTERNATIVE DI CONTROLLO DELLE
PARASSITOSI..................................................................................................................43
2.1 Lotta biologica.............................................................................................................43
2.1.1 Principi...............................................................................................................
2.1.2 Storia..................................................................................................................
2.1.3 Funghi nematofagi.............................................................................................
2.2. Fitoterapia...................................................................................................................52
CAPITOLO 3: MATERIALI E METODI.........................................................................55
3.1 Obbiettivo........................................................................................................................
1

3.2 Raccolta dei parassiti.......................................................................................................
3.3 Preparazione dei campioni..............................................................................................
3.4 Prove in vitro con funghi ambientali nei confronti di uova embrionate..........................
3.5 Prove in vitro con oli essenziali.......................................................................................
CAPITOLO 4: RISULTATI...............................................................................................62
4.1 Saggio dei funghi ambientali...........................................................................................
4.2 Saggio degli oli essenziali...............................................................................................
CAPITOLO 5:DISCUSSIONI E CONCLUSIONI...........................................................72
5.1 Saggio dei funghi ambientali...........................................................................................
5.2 Saggio degli oli essenziali...............................................................................................
BIBLIOGRAFIA...............................................................................................................77
2

RIASSUNTO
PAROLE CHIAVE: Ascaris suum, Lotta biologica, Estratti vegetali, Suino, funghi
nematofagi
L'ascaridiosi è una parassitosi a diffusione cosmopolita, responsabile di importanti
perdite economiche e di frequente riscontro nell'allevamento suino.
Il controllo di questa infestazione si basa sull'utilizzo di farmaci antielmintici, anche se
negli ultimi anni sono stati effettuati numerosi studi al fine di valutare alternative
terapeutiche basate sull'utilizzo di sostanze di origine naturale e sul ricorso alla lotta
biologica.
Lo scopo del nostro lavoro è stato quello di saggiare, in vitro, l'attività antielmintica di
estratti vegetali e di funghi ambientali nei confronti delle uova di Ascaris suum.
A questo scopo, è stata valutata l'inibizione dello sviluppo larvale da parte di oli
essenziali (Origanum vulgaris 5%, Mentha spicata 5%, Thymus serpyllum 5%,
Eucalyptus globulus 5%, Origanum majorana 5%, llicium verum 5%, Foeniculum
vulgare 5%) e di estratti vegetali ( Balanites aegyptiaca5%, Acacia auriculiformis 5%).
Gli oli essenziali di Timo, Origano e Eucalipto hanno mostrato i risultati migliori e
potrebbero essere prese in considerazione per ulteriori studi. Inoltre è stata valutata
l'attività ovicida di Beauveria sp.,Acremonium sp., Trichoderma sp., Trichothecium sp.,
Trichoderma viride coltivati in 2%water-agar e agar Sabouraud addizionato di
cicloeximide e cloramfenicolo. Beauveria sp. e Trichothecium sp. hanno mostrato una
potenziale attività nematofaga poiché sono stati capaci di danneggiare le uova in 2%
water-agar.
ABSTRACT
KEYWORDS: Ascaris suum, Biological Control, Plant Extracts, Pig, Nematophagous
Fungus
Ascariasis is a cosmopolitan infection caused by round-worm, which produces important
3

economical losses and frequently observed in pig farms.
The control of the infection is based on anti-helminthic drugs, but recently have been
studied several alternative strategies based on the use of nematophagous fungi and plant
extracs.
The purpose of this study was the in vitro testing of the anti-helmintic activity showed by
various enviromental fungi and plant extracs towards the Ascaris suum eggs.
For this purpose, was examinated the inhibition of larval development via the application
of essential oils (Origanum vulgaris 5%, Mentha spicata 5%, Thymus serpyllum 5%,
Eucalyptus globulus 5%, Origanum majorana 5%, llicium verum 5%, Foeniculum
vulgare 5%) and plants extracts (Balanites aegyptiaca 5%, Acacia auriculiformis 5%). T.
serpyllum, O. vulgaris and E. globulus can be considered for future antihelmintich
studies. Furthermore, the ovicidal activity of Beauveria sp., Acremonium sp.,
Trichoderma sp., Trichothecium sp., Trichoderma viride cultured in 2% water agar and
agar Sabouraud added with cycloheximid and chloramphenicol, was evaluated in vitro.
Beauveria sp. e Trichothecium sp. showed potential nematophagous activity causing
eggs' damaging in 2% water agar.
4

INTRODUZIONE
Fin dalla sua domesticazione, avvenuta circa 11000 anni fa, l'uomo ha allevato il suino
come risorsa di cibo per la sua sopravvivenza. Oggi l'industria suinicola è una delle
maggiori per produzione di carne ma, nonostante lo sviluppo della zootecnia, la presenza
di parassitosi rappresenta un fattore limitante dal punto di vista economico e produttivo.
Tra queste, l'ascaridiosi è una delle più importanti e rappresenta un problema diffuso in
tutto il mondo intersecando aspetti di sanità pubblica rilevanti, soprattutto nelle aree
meno sviluppate.
Il controllo di questo parassita viene effettuato attraverso l'uso di farmaci di sintesi anche
se negli ultimi anni è cresciuto l'interesse verso metodi alternativi di controllo.
Il rischio dell'insorgenza di farmaco-resistenze, la presenza di residui farmacologici nella
carne e la necessità di rispettare i tempi di sospensione, e la crescente espansione degli
allevamenti biologici dove l'uso dei farmaci è limitato, spingono verso la ricerca di
soluzioni naturali dotate di attività antielmintica come la fitoterapia e la lotta biologica.
Lo scopo del nostro studio è stato quello di saggiare in vitro l'efficacia antielmintica di
5

alcuni funghi ambientali e estratti vegetali nei confronti delle uova di Ascaris suum.
6

CAPITOLO 1: ASCARIDIOSI DEL SUINO
1.1. CLASSIFICAZIONE E TASSONOMIA
La classe dei nematodi comprende circa 12000 specie, tra cui specie parassitarie di
frequente riscontro negli animali domestici e specie a vita libera, che differiscono molto
per morfologia, ciclo biologico e specie. I parassiti possono essere suddivisi in due
gruppi, bursati e non bursati in relazione alla presenza o meno di una borsa copulatrice
nel maschio. Al gruppo dei non bursati appartiene la famiglia degli ascaridi, che
comprende i generi Ascaris, Toxocara, Parascaris, Toxascaris, Ascaridia, Heterakis e
Sulcascaris.
Al genere Ascaris appartengono le specie A. suum, parassita del suino, e Ascaris
lumbricoides, parassita umano.
Ascaris suum o A scaris lumbricoides ?
Fin dalla loro prima classificazione in due specie differenti, la comunità scientifica si è
interrogata su questa scelta; l'esistenza di un antenato comune appare confermata ma
tutt'oggi, nonostante le tecniche di indagine molecolare e i numerosi studi, la precisa
7

classificazione rimane incerta. Inoltre flussi genici nelle differenti popolazioni del mondo
complicano il quadro e pongono il caso come un interessante esempio di evoluzione in
atto.
Gli ascaridi rappresentano sicuramente uno dei primi parassiti conosciuti dall'uomo, data
la loro grandezza, diffusione e abbondanza. Coproliti di iguanodonte datati 100 milioni
di anni sono risultati essere positivi a uova di ascaridi (Poinar and Boucot, 2006). Il
papiro ebers, risalente al 1500 a.C circa, descrive varie parassitosi umane tra cui la
presenza di vermi intestinali, dato confermato dal ritrovamento di uova calcificate in
mummie egizie risalenti a quel periodo (Cox, 2002). Nella letteratura romana (Galeno di
Pergamo) e greca (Celso e Ippocrate) sono presenti numerosi riferimenti ad ascaridi e
cestodi ma solo Paulus Aegineta (625-690 d.C) descriverà le caratteristiche morfologiche
e cliniche delle infestazioni da ascaridi, ulteriormente approfondite e arricchite di dettagli
solo nel 17° secolo con Francesco Redi e il suo Osservazioni intorno agli animali viventi
che si trovano negli animali viventi, uno dei primi libri di parassitologia della storia.
8

Pochi anni dopo, con le scoperte di Linneo, vennero classificati Ascaris lumbricoides
Linnaeus 1758, parassita umano e Ascaris suum Goeze 1782 che colpisce i suini; anche
se le notevoli somiglianze morfologiche e molecolari mantengono vivo il dibattito sulla
loro classificazione.
Sono state proposte diverse ipotesi sull'origine e l'evoluzione di questi parassiti:
- A. lumbricoides e A. suum sono ambedue specie valide e avrebbero avuto origine per
speciazione da una specie ancestrale, forse per la domesticazione dei maiali.
- A. lumbricoides deriverebbe direttamente dal parassita A. suum (salto d'ospite) ritrovato
nei suini, quest'ultimo rappresenterebbe quindi la specie ancestrale ancora presente.
- A. suum deriverebbe da A. lumbricoides che in questo caso rappresenterebbe la specie
ancestrale persistente, esattamente l'opposto dell'ipotesi precedente.
- A. lumbricoides e A. suum sono conspecifici viste le poche differenze morfologiche e
genetiche.
Dal punto di vista morfologico i parassiti risultano pressoché identici: le uova sono
indistinguibili tra loro, i cicli biologici sovrapponibili e gli adulti hanno differenze
9

minime per poterli di distinguere.
Studi epidemiologici hanno suggerito l'esistenza di due possibili scenari nella
popolazione mondiale: 1) un ciclo biologico specie-specifico nelle regioni altamente
endemiche come Guatemala e Cina (Anderson et al., 1993; Peng et al., 2005); 2) un ciclo
biologico condiviso da suini e umani nelle regioni non endemiche come Danimarca e
Nord America (Anderson, 1995; Nejsum et al., 2005). In altri studi effettuati in Cina, A.
suum potrebbe comportarsi da serbatoio per l'ascaridiosi umana e sono state documentate
numerose cross-infestazioni (Zhou et al., 2012).
Numerosi studi di biologia molecolare hanno confermato la stretta vicinanza filogenetica
dei due, il completo sequenziamento del DNA mitocondriale ha evidenziato che
differiscono solo per l'1,9% delle basi (Liu et al., 2012). Questo potrebbe suggerire che
esista una sola popolazione di Ascaris spp.
10

1.2 DESCRIZIONE MORFOLOGICA
1.2.1 Parassita adulto
Ascaris suum è il più grande nematode del suino e presenta forma allungata, rastremata
alle due estremità. Le femmine adulte distese possono superare i 40 cm mentre i maschi
raggiungono i 25 cm. Il corpo è ricoperto da una cuticola incolore secreta dal sottostante
ipoderma di colore bianco-giallastro, che si approfonda nella cavità celomatica formando
quattro corde: due laterali, in cui sono presenti i canali escretori, e altre due dorsali e
ventrali, in cui si collocano i nervi.
11Illustrazione 1: Esemplari adulti (maschio e femmina) di Ascaridi. http://en.wikipedia.org/wiki/File:Ascaris-suum.jpg

Il movimento è permesso grazie alla presenza di cellule muscolari, orientate
longitudinalmente al corpo, tra l'ipoderma e la cavità celomatica, e disposte in maniera
antagonista. In questo modo il movimento è effettuato per contemporanea contrazione di
un gruppo di cellule e rilassamento delle altre.
All'interno della cavità celomatica è presente un liquido che, creando una pressione
interna, permette la rigidità del corpo e favorisce la stabilità degli organi interni che
rimangono sospesi nella cavità celomatica.
Il sistema digerente è di tipo tubulare ed è composto da:
- un' apertura orale o bocca semplice, con cui il parassita si alimenta di fluidi mucosali, di
detriti cellulari e di materiale ingerito dall'ospite. E' dotata di tre labbra: una dorsale con
due papille doppie alla sua base e due ventro-laterali, ognuna con una doppia papilla
ventrale e una subventrale. La superficie interna di ogni labbro è provvista di denti.
- un faringe muscolare
- un esofago, composto da un anello muscolare che crea una pressione negativa che
permette l'ingresso di alimento nel canale alimentare e superare la tendenza dell'intestino
12

a collassare per la pressione esercitata dal liquido celomatico. Nel genere Ascaris
l'esofago ha forma di bulbo, con un evidente ingrossamento a livello posteriore.
- l'intestino, un organo tubulare deputato all'assorbimento dei nutrienti che in cui sono
presenti microvilli in modo da aumentare la superficie utile. Nelle femmine adulte
l'intestino sbocca nell'ano mentre nei maschi in una cloaca a cui afferisce anche il dotto
deferente.
Il sistema escretore è composto da due canali che decorrono all'interno di ciascuna corda
laterale e sboccano, dopo essersi uniti, in un poro escretore nella regione esofagea.
Il sistema riproduttore è composto anch'esso da strutture tubulari. L'apparato femminile è
formato da un ovaio, un ovidotto e un utero doppio (che può contenere anche ventisette
milioni di uova); sbocca in un'unica vagina che si apre all'esterno in una vulva nel terzo
medio del corpo, in una costrizione anulare tipica che facilita l'unione durante la copula.
Gli organi maschili sono costituiti da un solo testicolo che comunica tramite il dotto
deferente con la cloaca. Nella cloaca sono presenti due spiculi, organi chitinosi che
vengono introdotti durante la copula nell'apertura genitale della femmina, di forma
13

allungata e leggermente curva, della lunghezza di 1,8-3,5 mm.
I principali organi di senso sono rappresentati dalle papille cefaliche e caudali, e dagli
anphidi e phasmidi. Le papille sono organi tattili e sono collocate anteriormente alle
regione esofagea e all'estremità caudale. Gli anphidi e phasmidi sono chemiorecettori e si
trovano rispettivamente intercalati alle papille cefaliche e all'estremità caudale.
1.2.2 Uova
Le uova sono di forma ovoidali e colore bruno-giallastro con parete spessa
multistratificata, che favorisce la resistenza nell'ambiente per diversi anni anche in
condizioni sfavorevoli.
14

La porzione esterna si presenta irregolarmente mammellonata e la grandezza delle uova
può variare da 45 micron a 75 micron di lunghezza e da 35 micron a 50 micron in
larghezza.
Al microscopio ottico risulta impossibile distinguerle da quelle di A. lumbricoides.
15
Illustrazione 2: Uova larvate e non larvate.

Le uova maturano nell'ambiente in non meno di 4 settimane in condizioni ottimali (22-26
C°) e possono resistere al congelamento e all'essicazione. Inoltre non sono inattivate dai
comuni disinfettanti a base di povidone-ioduro e sali di ammonio quaternario (Labare et
al. 2013).
1.2.3 Genoma
Il genoma del parassita è stato recentemente sequenziato in modo da conoscere
maggiormente le sue caratteristiche e confrontarle con altri nematodi . Inoltre lo studio
del genoma dei nematodi, essendo abbastanza semplice, rappresenta un valido modello
per lo studio di quello di organismi superiori. Sono state sequenziate 272,782,664 bp. Il
numero di sequenze ripetute nel genoma è abbastanza basso (4,4%) a causa della
diminuzione della cromatina, un meccanismo di origine incerta che porta, una volta
completata l'embriogenesi, ad una perdita di frammenti di DNA responsabili di questo
processo (Jex et al., 2011).
Osservando le funzioni dei geni riconosciuti notiamo che rivestono particolare
importanza: 456 geni che codificano per peptidasi, tra cui alcune cistein-proteasi e serin-
16

proteasi che rivestono un ruolo chiave nell'invasione e degradazione tissutale durante la
migrazione,609 chinasi, 257 fosfatasi, geni comuni ad altri parassiti (eft-1, fzo-1, glo-1,
rho-1) che hanno ruoli essenziali nello sviluppo embrionale, larvale e riproduttivo e sono
possibili target per composti antiparassitari e vaccini e geni codificanti per recettori e
proteine canale (in particolare canali voltaggio dipendenti, target di farmaci come i
lattoni macrociclici) (Jex et al., 2011).
1.3 CICLO BIOLOGICO
I primi studi riguardanti il ciclo di questo parassita si ebbero agli inizi del secolo. In
particolare nel 1922 un pediatra giapponese, Shimesu Koino, e suo fratello minore
fornirono preziose indicazioni ingerendo rispettivamente un numero cospicuo di uova di
A. lumbricoides e di A. suum provenienti da suini. Nel giro di un mese i fratelli avevano
sviluppato sintomi influenzali e polmonari, ed entrambi riuscirono ad identificare e
descrivere larve di ascaridi in migrazione nell'espettorato, facendo così luce su una fase
di questo singolare ciclo biologico (Cox 2002).
Il ciclo biologico di questo parassita è di tipo diretto, in quanto può essere completato in
17
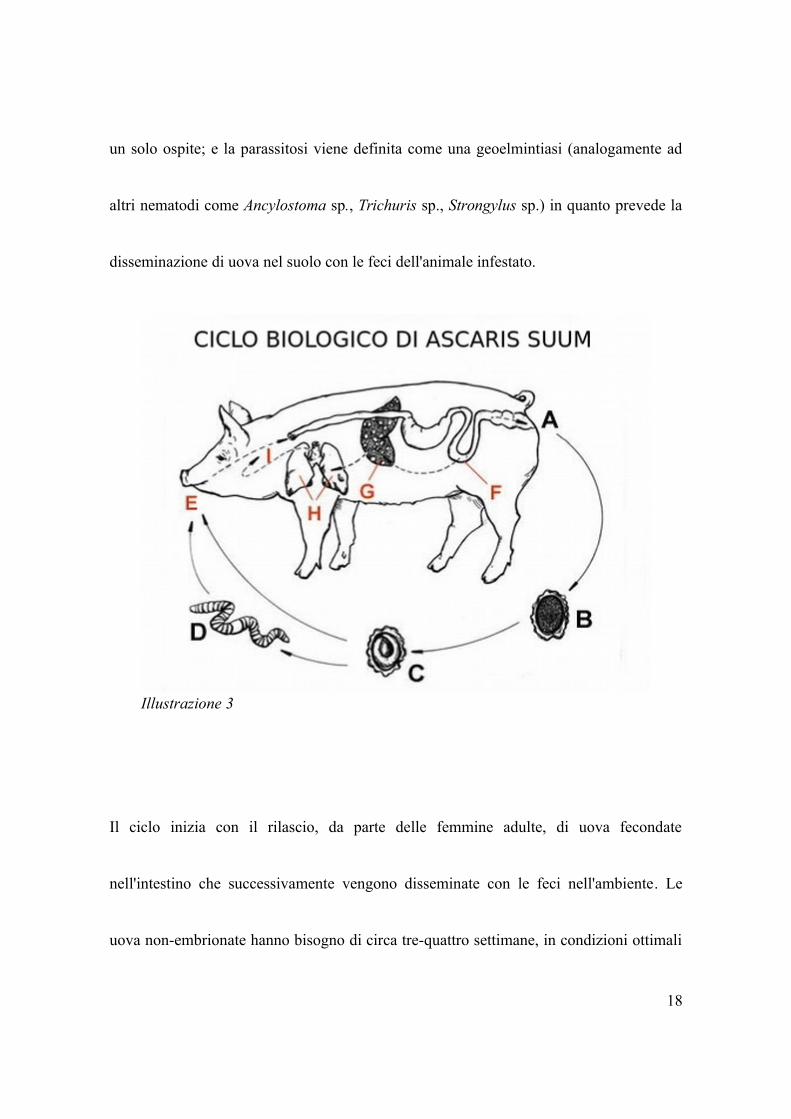
un solo ospite; e la parassitosi viene definita come una geoelmintiasi (analogamente ad
altri nematodi come Ancylostoma sp., Trichuris sp., Strongylus sp.) in quanto prevede la
disseminazione di uova nel suolo con le feci dell'animale infestato.
Il ciclo inizia con il rilascio, da parte delle femmine adulte, di uova fecondate
nell'intestino che successivamente vengono disseminate con le feci nell'ambiente. Le
uova non-embrionate hanno bisogno di circa tre-quattro settimane, in condizioni ottimali
18
Illustrazione 3

(22-26 C°) affinché maturino nell'ambiente e divengano infestanti. Le uova sono molto
resistenti e possono rimanere infestanti anche per vari anni nell'ambiente (Taylor et al.,
2010).
Il contagio avviene attraverso l'ingestione di uova infestanti, cioè contenenti una larva
L2. Se l'uovo larvato viene accidentalmente ingerito da un lombrico o un coleottero
coprofago, schiuderà al loro interno permettendo alla L2 di raggiungere i tessuti di questi
ospiti paratenici dove essa può rimanere infestante per lunghi periodi (Taylor et al.,
2010).
Le uova ingerite schiudono nel piccolo intestino per le caratteristiche chimiche e fisiche
dell'ambiente intestinale. La larva viene infatti stimolata alla produzione di proteinasi e
chitinasi che provocano la degradazione dei vari strati dell'uovo permettendole di uscire.
La larva L2 penetra nello spessore della mucosa, a livello del cieco e della parte
superiore del colon, e migra percorrendo i vasi mesenteriali verso il fegato, dove muta a
L3 (Murrell et al. 1997).
Qui attraversano il tessuto epatico alla ricerca di vasi efferenti. Seguendo il loro
19

andamento percorrono la vena epatica, la vena cava caudale e arrivano al cuore destro,
fino a raggiungere il sistema capillare successivo: il polmone.
A questo punto la larva L3 penetra nello spazio alveolare e risale l'albero respiratorio
grazie al suo movimento o a colpi di tosse fino alla faringe.
Viene ingerita e raggiunge, percorrendo il tubo digerente, il piccolo intestino, dove muta
a L4, dopo circa 14 giorni post-infezione, e poi L5. Una volta raggiunta la maturità
sessuale, dopo 6 settimane, il parassita è pronto nuovamente alla riproduzione e alla
disseminazione di nuove uova nell'ambiente. Gli adulti possono rimanere nell'intestino
per un anno anche se la maggior parte viene espulsa alla ventitreesima settimana
dall'ingestione (Pilitt et al., 1981).
1.4 IMMUNIT Ά NATURALE E ACQUISITA
L'immunità nei confronti di A. suum può essere divisa in una immunità naturale e una
immunità acquisita.
L'immunità naturale risulta ancora poco chiara anche se la componente genetica del suino
sembra essere una componente importante della resistenza naturale (Skallerup et al.,
20

2012).
In casi di esposizioni ripetute l'animale sviluppa una risposta immunitaria acquisita che
impedisce alla larva di completare il ciclo biologico.
A livello del fegato, vi è un'importante attivazione immunitaria per la migrazione larvale
caratterizzata da infiltrazione linfocitaria e eosinofilica, e dalla formazione di granulomi
e fibrosi interlobare; che si rendono visibili sulla superficie dell'organo (“white-spot” o
“milk-spot”). Alcune indagini hanno dimostrato che in suini vaccinati o che erano già
stati in contatto con il parassita, i “white spot” erano più grandi e persistenti .Inoltre
esiste una relazione tra il numero di “white spot” e il titolo di anticorpi anti-Ascaris, e nei
suini in cui l'attivazione immunitaria a livello del fegato era più elevata il numero di larve
polmonari era ridotto; a conferma dell'importanza dell'organo nello sviluppo
dell'immunità acquisita (Dold and Holland, 2011).
A livello del polmone, durante la migrazione delle L3, vengono richiamati eosinofili e
cellule mononucleate nel tessuto. Gli eosinofili sembrano avere un ruolo nello sviluppo
dell'immunità acquisita dopo esposizioni ripetute al parassita (Frontera et al., 2004).
21

1.5 PATOGENESI
I danni causati dal parassita possono essere causati dalle larve in migrazione o dagli
adulti. Le larve fuoriuscite dall'uovo possono causare piccoli danni alla mucosa
intestinale al momento della penetrazione nella stessa. A livello del fegato le larve in
migrazione si muovono nel parenchima creando tragitti e innescando una risposta
infiammatoria. A livello polmonare rompono la parete dell'alveolo causando piccole
emorragie. Le larve in migrazione possono perdersi, vagare e morire in sedi anomali
come milza, linfonodi, cervello.
Gli adulti localizzati nell'intestino, nutrendosi di fluidi contenuti nel lume; non
danneggiano l'intestino ma possono portare a fenomeni allergici ((Roberts, 2009).
1.6 SEGNI CLINICI E RILIEVI ANATOMO-PATOLOGICI
L'infestazione è difficilmente riconoscibile visto che nella maggior parte dei casi i
sintomi sono scarsi o assenti. La principale conseguenza, infatti, della presenza di
parassiti adulti è rappresentato dalla riduzione dell'indice di incremento ponderale.
Nella fase della migrazione larvale si possono riscontrare sintomi a livello polmonare e
22

epatico.
A livello epatico, le lesioni da migrazione difficilmente danno origine a sintomi evidenti
anche se provocano danni economici per il sequestro dell'organo. I tragitti nel
parenchima appaiono prima emorragici, visibili attraverso la capsula come capocchie di
spillo rosse, circondati da una sottile area rosa. Successivamente le lesioni collassano e si
risolvono in fibrosi di aspetto caratteristico (“white spot”) e facilmente individuabili al
macello dove l'organo deve essere scartato.
23
Illustrazione 4: Lesioni epatiche "white spot"-

Dipendendo dal grado dell'infestazione, l'infiltrato infiammatorio può compromettere la
funzionalità del lobulo. Focolai granulomatosi con cellule giganti, macrofagi ed
eosinofili possono concentrarsi su residui di larve intrappolate e distrutte in sede epatica.
A livello polmonare la migrazione può causare, soprattutto negli animali più giovani,
una polmonite evidente ma di facile risoluzione. Alla necroscopia si rilevano,
macroscopicamente, numerose emorragie focali e diffuse sulla superficie e nel
parenchima dell'organo, accompagnate in grado variabile da edema, congestione e
enfisema polmonare. Le larve sono di facile ritrovamento all'interno del tessuto e nel
lume dell'albero respiratorio. Microscopicamente si rileva una bronchiolite eosinofilica.
La presenza di parassiti adulti nell'intestino nella maggior parte dei casi non provoca
sintomi facilmente rilevabili. Infestazione massive possono causare ostruzioni intestinali
anche se raramente portano a rottura della parete. Occasionalmente singoli adulti possono
migrare nel dotto pancreatico o nel coledoco causando ittero post-epatico e colangiti.
Microscopicamente si riscontra ipertrofia della muscolaris esterna con allungamento
delle cripte di Lieberkhün, ipertrofia e diminuzione delle cellule caliciformi nelle
24

porzioni colpite ed infiltrazione di eosinofili e mastociti.
1.7 EPIDEMIOLOGIA E DANNI ECONOMICI
Il patrimonio suinicolo in europa ammonta 147.010.600 esemplari. La Germania è il
primo paese produttore con il (19,3%,28.331.000 capi), seguito da Spagna (17,2%,
25.250.000 capi) e Francia (9,4%, 13.778.000 capi).
L'Italia, con 8.661.000 esemplari, rappresenta l'ottavo produttore in Europa con il 5,9%
dei suini presenti nella comunità (EUROSTAT, 2013).
Nel panorama italiano Lombardia e Emilia Romagna detengono da sole il 64% del
patrimonio totale mentre la Toscana con 147.663 capi rappresenta l'1,7% del patrimonio
nazionale (Basile, 2013).
Presumibilmente A. suum è presente negli allevamenti di tutto il mondo anche se solo in
alcuni paesi sono stati effettuati studi epidemiologici e molti riguardano realtà
probabilmente non più attuali. .Tra questi vengono utilizzati parametri differenti per
determinare la presenza del parassita. Alcuni utilizzano la percentuale di fegati
sequestrati mentre altri utilizzano la presenza del parassita adulto nell'apparato intestinale
25

in sede di macellazione o l'esame quantitativo delle uova da campioni fecali (tabella 1).
26
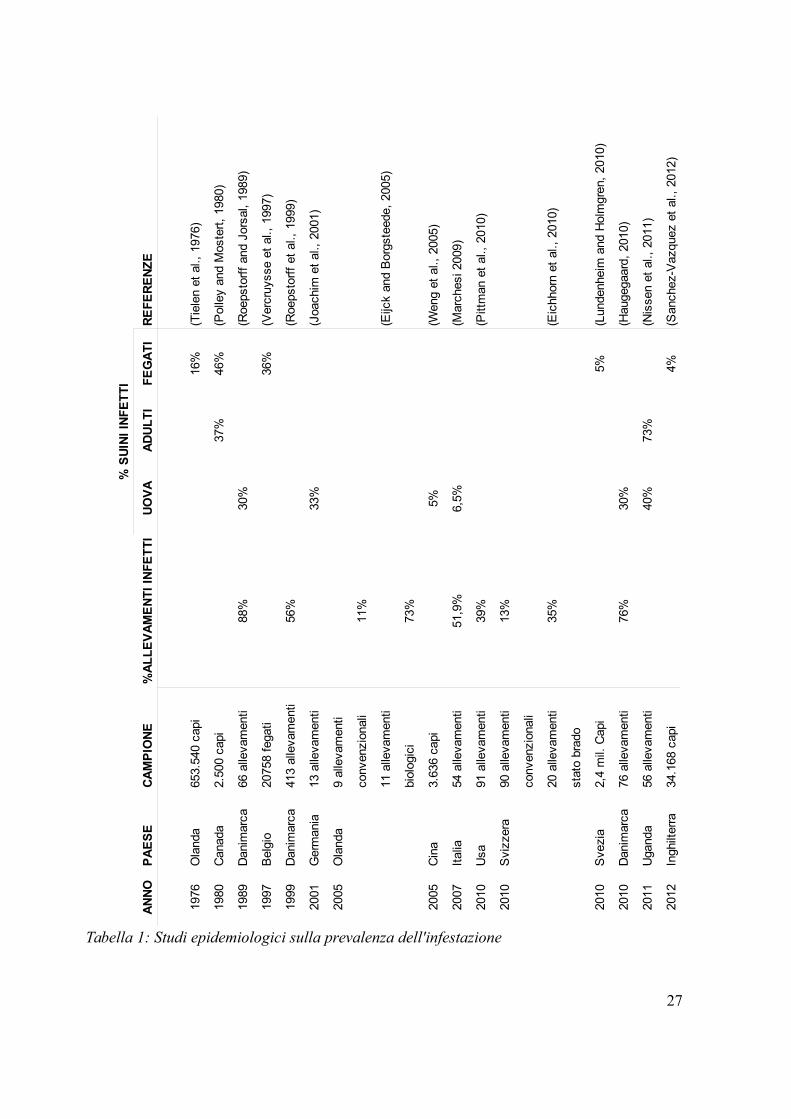
27
Tabella 1: Studi epidemiologici sulla prevalenza dell'infestazione
RE
FE
RE
NZ
E
(Tie
len
et a
l., 1
976)
(Pol
ley
and
Mos
tert
, 19
80)
(Roe
psto
rff
and
Jors
al,
1989
)
(Ver
cruy
sse
et a
l., 1
997)
(Roe
psto
rff
et a
l., 1
999)
(Joa
chim
et
al.,
200
1)
(Eijc
k an
d B
orgs
teed
e, 2
005)
(Wen
g et
al.,
200
5)
(Mar
ches
i 200
9)
(Pitt
man
et
al.,
201
0)
(Eic
hhor
n et
al.,
201
0)
(Lun
denh
eim
and
Hol
mgr
en,
2010
)
(Hau
gega
ard,
201
0)
(Nis
sen
et a
l., 2
011)
(San
chez
-Vaz
quez
et
al.,
201
2)
% S
UIN
I IN
FE
TT
I FE
GA
TI
16%
46%
36%
5% 4%
AD
UL
TI
37%
73%
UO
VA
30%
33%
5% 6,5%
30%
40%
%A
LL
EV
AM
EN
TI
INF
ET
TI
88%
56%
11%
73%
51,9
%
39%
13%
35%
76%
CA
MP
ION
E
653.
540
capi
2.50
0 ca
pi
66 a
lleva
men
ti
2075
8 fe
gati
413
alle
vam
enti
13 a
lleva
men
ti
9 al
leva
men
ti
conv
enzi
onal
i
11 a
lleva
men
ti
biol
ogic
i
3.63
6 ca
pi
54 a
lleva
men
ti
91 a
lleva
men
ti
90 a
lleva
men
ti
conv
enzi
onal
i
20 a
lleva
men
ti
stat
o br
ado
2,4
mil.
Cap
i
76 a
lleva
men
ti
56 a
lleva
men
ti
34.1
68 c
api
PA
ES
E
Ola
nda
Can
ada
Dan
imar
ca
Bel
gio
Dan
imar
ca
Ger
man
ia
Ola
nda
Cin
a
Ital
ia
Usa
Svi
zzer
a
Sve
zia
Dan
imar
ca
Uga
nda
Ingh
ilter
ra
AN
NO
1976
1980
1989
1997
1999
2001
2005
2005
2007
2010
2010
2010
2010
2011
2012

In questo quadro risulta difficile stabilire con esattezza la prevalenza dell'ascaridiosi negli
allevamenti italiani. Studi epidemiologici effettuati in Emilia Romagna rivelano che nel
51,9% degli allevamenti in esame è presente il parassita mentre la prevalenza negli
animali è del 6,5%.Negli allevamenti a ciclo chiuso la prevalenza è maggiore
(10,3%)rispetto a quelli a ciclo aperto (3,79%) (Marchesi, 2009).
Una valutazione su questo parassita può essere fatta interpretando i cambiamenti che la
zootecnia ha subito nell'ultima metà del secolo. In questo periodo abbiamo assistito
infatti ad una progressiva industrializzazione degli allevamenti e quindi a un
cambiamento delle patologie in zootecnica. Da un lato c'è stata una drastica diminuzione
delle malattie enzootiche e epidemiche, mentre dall'altro ci siamo trovati di fronte
all'aumento dell'insorgenza di malattie meno “appariscenti” , ad andamento subclinico,
come le parassitosi gastro-intestinali. Queste, nonostante sollevino gli allevatori dalla
preoccupazione immediata della morte dell'animale, interferiscono notevolmente con la
produttività potenziale dell'animale in relazione a patrimonio genetico, tipologia
d'allevamento e alimentazione e costituiscono un danno economico da non sottovalutare.
28

Possiamo individuare danni economici diretti e indiretti. Tra quelli diretti ci sono il
sequestro dei fegati parassitati e i costi per sostenere i trattamenti chemioterapici,sia
contro A. suum che nei confronti di eventuali infezioni batteriche secondarie.
Tra quelli indiretti invece ci sono quelli che derivano dal diminuito stato di salute degli
animali. Questi causano una riduzione dei parametri di produzione: l'incremento
ponderale giornaliero, l'indice di conversione degli alimenti e la qualità delle carni
(Knecht et al., 2012).
Determinare l'entità delle perdite economiche è difficile. Le numerose interazioni tra
parassita, ospite, ambiente, tipo di produzione e allevamento complicano il quadro e la
possibilità di fare valutazioni. Alcuni studi hanno attribuito alle strategie di controllo del
parassita un incremento di profitti che vanno da 3 a 12 euro ogni suino in finissaggio
all'anno (Van Meensel et al., 2010).
I motivi per cui la prevalenza di Ascaris, rimane elevata negli allevamenti intensivi,
nonostante lo sviluppo della zootecnia, sono vari. In primo luogo il ciclo biologico di tipo
diretto rende non necessaria la presenza di altri organismi per la trasmissione a nuovi
29

ospiti. Secondo, le femmine adulte sono altamente feconde e possono disseminare un
numero molto elevato di uova al giorno contaminando in poco tempo l'allevamento.
Terzo, l'elevata resistenza delle uova nell'ambiente che possono rimanere infettanti anche
per vari anni e facilitare la reinfestazione degli animali dopo trattamenti chemioterapici
in casi di ambienti molto contaminati.
Le prevalenza più elevate si riscontrano nei suini di giovane età. Secondo alcuni autori, i
suinetti neonati infatti possono contrarre l'infezione immediatamente dopo la nascita
attraverso l'assunzione di uova adese al bassoventre della scrofa (Taylor et al., 2010).
La parassitosi mostra, nelle aree con clima temperato, una precisa stagionalità. Si
manifesta con gravità nella stagione estiva mentre riduce la sua comparsa nelle stagioni
più fredde in cui la maturazione delle uova viene inibita.
30

1.8 TECNICHE DIAGNOSTICHE
La presenza di A. suum può essere investigata in differenti modi: in allevamento,
attraverso la diagnosi clinica e il riscontro di parassiti adulti nelle feci; in laboratorio, con
la diagnostica coprologica; in sede di macellazione, identificando il parassita a livello
intestinale o attraverso le lesioni presenti su polmoni e fegato.
1.8.1 Diagnosi in allevamento
Effettuare la diagnosi attraverso i segni clinici può risultare difficile. I sintomi associati
all'ascaridiosi come le difficoltà respiratorie e ridotte prestazioni zootecniche sono
aspecifici e a decorso subclinico.
1.8.2 Diagnosi di laboratorio
Per quanto riguarda la diagnosi coprologica, l'analisi delle feci può fornire importanti
informazioni valutando la presenza e il numero di uova.
Per la messa in evidenza delle uova di Ascaris, la tecnica più usata è la flottazione con
soluzione satura di Cloruro di Sodio oppure soluzioni a base di solfato di Zinco o
Magnesio. Questa permette di far affiorare le uova dal campione fecale in modo da
31

identificarle successivamente con l'esame microscopico.
Oltre alla tecnica della flottazione, la tecnica Mc Master permette di effettuare una
valutazione di tipo quantitativo espressa in UPG (uova per grammo).
Il numero di uova escrete con le feci può dare un'indicazione della carica infestante in
quanto sembra essere correlato al numero di adulti presenti nell'intestino (Nejsum et al.,
2009). Secondo alcuni autori, dobbiamo tenere in considerazione che bassi valori di UPG
potrebbero essere messi in relazione ad alcuni comportamenti fisiologici dei suini come
la coprofagia e la geofagia che determinano il semplice passaggio delle uova dal tratto
gastroenterico (Taylor et al., 2010). D'altra parte, anche l''assenza di uova nelle feci
potrebbe essere un falso negativo nel caso in cui esistono solo parassiti immaturi oppure
appartenenti a un solo sesso.
Riguardo la diagnosi sierologica invece, nonostante l'uso di test sierologici sia ormai
molto diffuso e comprenda un numero importante di malattie, attualmente non esistono
test disponibili per la diagnosi di Ascaris sp. Inoltre, possiamo rilevare che non risulta
esserci correlazione tra i livelli di anticorpi anti-Ascaris e entità dell'infestazione. Inoltre
32

sono stati studiati differenti estratti di adulti e larve, “excretory/secretory products” e
alcune proteine purificate, per la ricerca di antigeni ma nonostante siano risultati efficaci
non sono stati presi in considerazione per la diagnosi di routine (Frontera et al., 2003).
1.8.3 Diagnosi in sede di macellazione
In sede di macellazione si possono riscontrare: presenza di adulti nell'intestino, presenza
di “white spot” nel fegato, presenza di lesioni polmonari.
La presenza del parassita può derivare dal riscontro di vermi adulti in sede di
macellazione anche se necessita di un esame approfondito dell'apparato gastroenterico
che non viene fatto nella pratica lavorativa.
L'esame del fegato alla visita ispettiva può mettere in luce la presenza di lesioni
caratteristiche (“white spot”) da larve in migrazione. Il numero di lesioni nel fegato non è
correlato al numero di adulti presenti nell'intestino che possono essere anche assenti.
Questo può derivare dal fatto che l'animale è stato precedentemente immunizzato e le
larve sono state sequestrate in sede epatica.
La presenza di lesioni polmonari come pleuriti e polmoniti in sede di macellazione può
33

essere un indicatore secondario di infezione da Ascaris. Il parassita danneggia
direttamente il tessuto polmonare durante la migrazione e può essere un fattore
predisponente a infezioni secondarie. L'aspecifità di questi reperti è comunque
difficilmente associabile alla presenza di ascaridi.
1.9 PREVENZIONE E CONTROLLO
I piani di controllo di questo parassita mirano ad eliminarlo completamente
dall'allevamento; soprattutto l'eliminazione delle uova dall'ambiente costituisce il
problema cardine dell'elaborazione di piani efficaci.
I trattamenti antielmintici da soli non riescono a risolverlo e devono essere coadiuvati da
pratiche di corretta gestione, al fine di ridurre le fastidiose perdite economiche causate
dalla riduzione dei principali parametri di produttività dell'allevamento suino.
In questo paragrafo vengono affrontate le corrette pratiche igieniche, i trattamenti
antielmintici con farmaci di sintesi frequentemente utilizzati negli allevamenti e lo studio
di vaccini in grado di difendere dall'infestazione.
34

1.9.1 Prevenzione
Alcune pratiche di management in allevamento risultano di fondamentale importanza
nella messa a punto di piani di controllo efficaci. Queste hanno l'obbiettivo principale di
ridurre la presenza e limitare la diffusione delle uova nell'ambiente in quanto
rappresentano il principale problema nella lotta a questa parassitosi.
Negli allevamenti intensivi a ciclo chiuso assumono molta importanza gli interventi
igienici riguardanti l'alimentazione e la lettiera. Frequenti pulizie delle pavimentazioni,
delle pareti e dei trogoli con acqua o vapore riducono notevolmente il rischio di
infestazione.
Nei suini allevati estensivamente il problema è più complesso, e può portare
all'abbandono delle zone di pascolo per diversi anni in quanto le uova sono in grado di
resistere anche a cicli di produzioni agricole.
Dato che l'ingestione delle uova, da parte dei suinetti, dalla pelle della scrofa in sala parto
è un evento frequente, il rischio viene ridotto effettuando un lavaggio di questa prima
dell'entrata in sala parto (Taylor et al., 2010).
35

1.9.2 Trattamenti antielmintici
Dagli anni 60' con l'avvento di nuovi farmaci antielmintici ad ampio spettro il controllo
dell'ascaridiosi è diventato dipendente da questi. Questi farmaci sono disponibili in
commercio in varie forme farmaceutiche e la loro somministrazione con acqua o
mangimi rappresenta la soluzione più facile ed economica rispetto alle vie parenterali. Il
trattamento di massa però non permette di conoscere la dose di farmaco assunta da
ciascun animale.
Negli ultimi anni sono stati avanzati dubbi riguardanti l'impatto a lungo termine di questi
composti sull'ambiente. Alte percentuali di queste sostanze infatti vengono escrete
nell'ambiente dall'animale senza venire modificate (Plumb, 2008). Inoltre l'utilizzo di
escrementi di questi animali come concimi diffonde queste sostanze fuori dall'ambito
dell'allevamento con effetti a lungo termine difficilmente prevedibili (Spratt, 1997).
Nella scelta del farmaco quindi, devono essere presi in considerazione: 1) il margine di
sicurezza del composto, 2) l'efficacia nei confronti dei vari stadi del parassita, 3) lo
spettro di attività, 4) la modalità di somministrazione, 5) il costo.
36

Le pratiche di allevamento prevedono il trattamento delle scrofe prima dell'ingresso in
sala parto e dei suinetti all'atto dell'acquisto o all'ingresso nel ciclo di finissaggio e
successivamente dopo 8 settimane.
I verri vengono sottoposti a trattamento ogni 3-6 mesi.
E' buona norma allontanare le feci dal recinto 3-4 giorni post-trattamento in quanto sono
ricche di uova e di adulti in fase di disgregazione (Taylor et al., 2010).
Alcuni autori hanno dimostrato la mancanza di economicità nel trattamento di routine e
suggeriscono di trattare gli animali solo nel caso in cui i livelli di prevalenza risultino
inaccettabili. Questo programma necessita però piani di screening in allevamento e non si
adatta alle realtà più piccole (Roepstorff, 1997).
• farmaci utilizzati
Le molecole utilizzabili mostrano un'alta efficacia in test in vitro. Alcuni mostrano
attività nei confronti delle forme larvali come pyrantel, fenbendazolo, flubendazolo e
levamisole. I lattoni macrociclici (ivermectina, doramectina) hanno un effetto residuo
prolungato rispetto agli altri (tabella 2) (Vlaminck, 2013).
37

38
Tabella 2
RE
FE
RE
NZ
E
(Mart
i et al., 1978)
(Ste
ffan e
t al., 1988)
(Ste
wart
et al., 1984)
(Mart
i et al., 1978)
(Ste
wart
and R
ow
ell,
1986)
(Vanparijs
et al., 1988)
(Bra
dle
y e
t al., 1983)
(Pecheur,
1983)
(Oakle
y, 1974)
(Mart
i et al., 1978)
(Kennedy e
t al., 1987)
(Alv
a-V
ald
es e
t al., 1989)
(Lic
hte
nste
iger
et al., 1999)
(Ste
wart
et al., 1996)
(Lic
hte
nste
iger
et al., 1999)
EF
FIC
AC
IA L
AR
VE
100%
vers
o L
4
85%
vers
o L
4
92%
vers
o L
4
100%
vers
o L
4
97-1
00%
vers
o L
3 p
olm
100%
vers
o L
4
100%
vers
o L
4
100%
vers
o L
4 p
olm
100%
vers
o L
4
EF
FIC
AC
IA A
DU
LTI
100%
100%
100%
100%
92,4
%
100%
100%
100%
100%
100%
97,9
%
100%
97,7
%
100%
99%
100,0
%
97,5
%
100%
100%
100%
DO
SA
GG
IO
43 m
g/K
g 1
gg
17 m
g/K
g 1
gg
200 m
g/K
g 1
gg
3 m
g/K
g 3
gg
2,5
mg/k
g 3
gg
5m
g/K
g 1
gg
1,5
mg/K
g 5
gg
15 m
g/K
g 1
gg
40 m
g/K
g 1
0gg
7,5
mg/K
g 1
gg
8 m
g/K
g 1
gg
100m
icro
g/K
g
200 m
icro
g/K
g
300 m
icro
g/K
g 1
gg
300 m
icro
g/K
g 1
gg
300 m
icro
g/K
g 1
gg
FO
RM
UL
AZ
ION
E
ora
le
nella
razio
ne
con a
cqua
nella
razio
ne
nella
razio
ne
ora
le
nella
razio
ne
pare
nte
rale
nella
razio
ne
nella
razio
ne
nella
razio
ne
pare
nte
rale
con a
cqua
pare
nte
rale
NO
ME
Dic
hlo
rvos
Pip
era
zin
a
Fenbendazolo
Flu
bendazolo
Oxib
endazolo
Levam
isolo
Pyra
nte
l
Iverm
ectina
Dora
mectina
(Marc
hio
ndo a
nd S
zanto
,
1987)
(Marc
hio
ndo a
nd S
zanto
,
1987)
(Marc
hio
ndo a
nd S
zanto
,
1987)
(Schill
horn
van V
een a
nd
Gib
son, 1983)
(Schill
horn
van V
een a
nd
Gib
son, 1983)
99%
larv
e e
patiche e
polm
onari L
3
92,3
% v
ers
o L
4e 6
3,5
%
vers
o L
3 p
olm
onari
96g/ton
razio
ne,2
4gg

• resistenza agli antielmintici
Lo sviluppo di resistenze a farmaci antielmintici rappresenta un problema per alcune
specie che pone dei dubbi, sia sulla possibilità di utilizzo di queste sostanze, sia riguardo
le future modalità d'intervento, vista la notevole riduzione dell'attività di ricerca in nuove
classi di farmaci.
A oggi non sono riportati studi che descrivono lo sviluppo di farmaco resistenza in A.
suum. Al contrario sono diventate rilevanti le antielmintico resistenze in parassiti
filogeneticamente simili come Parascaris equorum (Laugier et al., 2012) e
Oesophagostomum spp. (Gerwert et al., 2002; Roepstorff et al., 1987).
Questo può far pensare che, proseguendo con pratiche sbagliate, anche in A. suum si
possano sviluppare questi problemi.
I farmaci in cui sono documentati questi problemi sono: benzimidazoli (tiabendazolo),
imidazotiazolo/tetraidropirimidine (levamisolo, pyrantel), avermectine-milbemicine
(ivermectina, moxidectina) (Kaplan, 2004).
39

1.9.3 Vaccinazione
La sviluppo di un vaccino contro gli ascaridi, in grado di proteggere gli animali dalle
infestazioni, viene ormai studiata da molti anni e rappresenta un possibilità concreta per
il futuro, anche se queste tecniche devono ancora essere ottimizzate e approfondite
(Vlaminck, 2013).
Numerosi studi sono stati condotti con lo scopo di rilevare quali antigeni parassitari
evocassero una risposta potente e prolungata nell'ospite. Somministrazione di antigeni di
origine larvale (Matsumoto et al., 2009) e di proteine di escrezione/secrezione (Geldhof
et al., 2004) hanno mostrato buoni risultati.
40

CAPITOLO 2: STRATEGIE ALTERNATIVE DI CONTROLLO
DELLE PARASSITOSI
Negli ultimi anni, a causa dell'insorgenza di farmaco-resistenze e alla necessità di trovare
alternative terapeutiche negli allevamenti biologici, è cresciuto l'interesse nei confronti di
metodi di lotta alle parassitosi che non prevedano l'utilizzo di molecole di sintesi.
In questo capitolo vengono descritte alcune di queste metodologie: la lotta biologica e la
fitoterapia.
2.1 LOTTA BIOLOGICA
2.1.1 Principi
La lotta biologica è una tecnica di controllo di organismi patogeni attraverso l'utilizzo di
antagonisti naturali.
In un ecosistema infatti, ogni specie è soggetta all'interazione con fattori di controllo sia
dall'ambiente che da altri organismi viventi; in particolare, nella lotta biologica si
sfruttano i rapporti di antagonismo come la predazione, il parassitismo e la competizione
interspecifica.
41

2.1.2 Storia
Questa tecnica ha le sue radici nel XVII e nel XVIII secolo quando alcuni naturalisti,
Aldrovandi, Redi, Vallisneri e altri, approfondiscono lo studio della biologia e
dell'etologia dei microrganismi inferiori, interessandosi delle relazioni fra insetti
entomofagi e fitofagi.
Nel XIX secolo questi studi vengono per le prime volte messi in pratica nell'ambito
dell'agricoltura, nella lotta contro insetti dannosi alle colture. Il primo esempio di lotta
biologica in Italia risale al 1845 quando Antonio Villa, naturalista milanese, vince una
medaglia d'oro messa in palio dalla Società d'incoraggiamento d'arti e mestieri a chi
“tentasse con qualche successo dei nuovi esperimenti tendenti a promuovere lo sviluppo
artificiale di qualche specie di insetti carnivori, onde avere con esso un efficace mezzo
per distruggere un'altra specie d'insetti riconosciuti dannosi all'agricoltura”.
In questi anni i risultati erano contrastanti e senza apprezzabili risultati, ma nel 1888 con
l'introduzione del Coccinellide Rodolia cardinalis dall'Australia alla California si ottenne
il successo che affermò le potenzialità di questa tecnica. Si riuscì infatti a debellare le
42

infestazioni di Icerya purchasi, cocciniglia originaria dell'Australasia ma che introdotta
in America stava costringendo gli agrumicolturi ad abbandonare le coltivazioni,
introducendo nell'areale il predatore Rodolia cardinalis. Questo successo rappresenta una
pietra miliare della lotta biologica (Viggiani, 1974).
Negli anni 30' si ebbero i primi studi sull'uso di funghi definiti “nematofagi” per ridurre
la presente di parassiti vegetale nel suolo (Linford, 1937).
2.1.3 Funghi nematofagi
I funghi nematofagi sono antagonisti naturali di nematodi, sono infatti in grado di
catturarli, ucciderli e digerirli a vari livelli di sviluppo e con vari meccanismi.
Questo gruppo comprende circa 200 specie diverse , a diffusione cosmopolita,
appartenenti a tutte le maggiori classi tassonomiche anche se la maggior parte fa parte
del taxon dei deuteromiceti.
Si ritrovano comunemente nei suoli ricchi di materiale organico, nello strato superficiale
fino a 20 cm di profondità (Persmark L, 1996).
La maggior parte può vivere sia come saprofita, degradando materiale organico, che
43

come parassita, utilizzando i nematodi come fonte di nutrimento (Barron, 1992).
Possono essere classificati principalmente in tre gruppi: 1) “nematode-trapping fungus”,
2) funghi endoparassiti, 3) “egg parasitic e cyst-parasitic fungi”.
Da alcuni autori viene citato anche un quarto gruppo, capace di produrre tossine che
immobilizzano il nematode (Jansson HB, Tunlid A, 1997), e un quinto che utilizza
strutture filiformi chiamati acantociti (Luo et al., 2006).
Poiché la maggior parte dei “nematode trapping” e degli “egg parasitic” sono saprofiti
del suolo risultano maggiormente utilizzabili nella lotta biologica rispetto agli
endoparassiti, strettamente dipendenti dall'ospite.
-”Nematode-trapping fungus”
L'abilità di catturare parassiti da parte di questi funghi è connessa con una specifica fase
di sviluppo del micelio in cui vengono a formarsi particolari strutture come “hyphal nets”
(Arthrobotrys oligospora), strutture tridimensionali con proprietà adesive,
“knobs”(Monacrosporium haptotylum) una singola cellula adesiva in grado di staccarsi
dal micelio e seguire il nematode seguendo il suo movimento, “branches”
44

(Monacrosporium cionopagum) o “costriction rings” (Arthrobotrys dactyloides) in cui i
nematodi sono catturati meccanicamente (Nordbring-hertz and Tunlid, 2006).
La formazione di queste strutture è legata alla specie e a fattori ambientali biotici e
abiotici. Tra quelli biotici è il nematode stesso a stimolarne la produzione venendo a
contatto con il micelio. Recentemente sono stati individuati composti secreti dai
nematodi, chiamati “ascarosidi”, che indurrebbero lo sviluppo di queste strutture
particolari (Hsueh et al., 2013). Esistono più di 100 molecole diverse identificate in vari
nematodi, generalmente formate da uno zucchero, ascarysole, legato a una catena di acidi
grassi (von Reuss et al., 2012).
Le interazioni che si creano tra il nematode e il fungo sono complesse. Il nematode è
attratto da composti rilasciati dal micelio, la cui capacità di attrazione varia secondo la
specie. L'attrazione sembra essere minore nei funghi con attitudine saprofitica.
Il micelio, contemporaneamente, avverte la presenza del nematode e l'orientamento di
particolare fibrille permette l'adesione e l'intrappolamento dello stesso. La composizione
dettagliata della superficie delle fibrille rimane sconosciuta.
45

Alcuni funghi, come A. dactyloides, riescono a catturare l'elminta attraverso strutture
chiamate “costriction rings” costituite da tre cellule disposte ad anello. Queste, nel
momento in cui la preda si trova all'interno, rispondono incrementando il loro volume in
maniera da diminuire il diametro dell'anello e immobilizzarlo. Questo singolare
46
Illustrazione 5: Penetrazione della cuticola del nematode da parte di A. oligospora al Microscopio Elettronico. Corpi Densi (DB), Rivestimento adesivo (A), cuticola nematode (NC), bulbo infettivo (IB). Veenhuis M, Nordbring-Hertz B and HarderW(1985).

meccanismo è rapido (0,1 s), irreversibile e probabilmente provocato dall'apertura di
canali che portano all'ingresso di acqua nelle cellule (Nordbring-hertz and Tunlid, 2006).
Successivamente si formano canali specifici che perforano la cuticola, probabilmente
attraverso l'attività chimica di enzimi idrolitici e, meccanica, dalla pressione generata
47
Illustrazione 6: Meccanismo di cattura “costriction ring” di A. brochopaga. Nordbring-Hertz B, Neumeister H, SjollemaKand VeenhuisM(1995) A conidial trap-forming mutant of Arthrobotrys oligospora. Mycological Research 99: 1395–1398.

dalla crescita fungina.
La penetrazione è seguita dalla digestione delle molecole organiche del nematode. Il
micelio subisce profonde modificazioni con lo sviluppo di strutture trofiche in cui
rapidamente vengono immagazzinati i nutrienti assimilati sotto forma di goccioline
lipidiche (Nordbring-hertz and Tunlid, 2006).
-Endoparassiti
In questo gruppo di funghi le spore prodotte, mobili (Catenaria anguillulae) o immobili
(Drechmeria coniospora), colonizzano il parassita aderendo alla sua superficie o
vengono ingeriti dallo stesso. In entrambi i casi la vitalità del parassita viene
irrimediabilmente compromessa. La maggior parte dei funghi endoparassiti sono parassiti
obbligati e hanno un ristretto spettro d'ospite; inoltre sono incapaci di crescere nel suolo
rendendo difficile il loro impiego nella lotta biologica ai nematodi (Braga et al., 2013).
-”Egg parasitic e cyst parasitic”
Questo gruppo di funghi parassitano le forme immobili dell'ospite o le uova attraverso la
colonizzazione del guscio, la penetrazione nello stesso e la digestione del contenuto.
48

La penetrazione avviene attraverso meccanismi meccanici, per la pressione esercitata dal
micelio (Lýsek and Krajcí, 1987), e soprattutto enzimatici, con la produzione di enzimi
come proteasi e chitinasi (Braga et al 2010).
In generale esistono due modi per poter applicare questi funghi nella lotta biologica: 1) la
diffusione dei funghi nel suolo in maniera da interagire con il parassita quando questo si
49
Illustrazione 7: Meccanismi di cattura degli elminti A) “hiphal net” di A.oligospora B)”branches” adesivi di Monacrosporium gephyropagum C)”knobs” di Monacrosporium haptotylum D) Spore adesive di Drechmeria coniospora E) “costriction ring” di A. brochopaga F) Zoospore di Catenaria anguillulae. Nordbring-Hertz B, Neumeister H, SjollemaKand VeenhuisM(1995) A conidial trap-forming mutant of Arthrobotrys oligospora. Mycological Research 99: 1395–1398.

trova nell'ambiente; 2) la somministrazione di spore agli animali in maniera che possano
attraversare l'intestino e crescere nelle feci parassitando il nematode o le sue uova.
Nei confronti delle uova di Ascaris sp. sono state saggiate, solo in vitro, varie specie
fungine.
Studi effettuati addizionando Pochonia chlamydosporia alla razione di suini, hanno
mostrato attività ovicida nei confronti delle uova di A. suum dopo il passaggio dal tratto
gastrointestinale (Ferreira et al., 2011b). Questi sono in accordo con quelli di altri autori
che avevano confermato l'attività ovicida di questo fungo in vitro nei confronti di A.
suum (Ferreira et al., 2011a; Araújo et al., 2008) e in Trichuris trichiura (Silva et al.,
2011).
Duddingtonia flagrans e Monacosporium sinense non hanno mostrato attività ovicida,
nonostante per il primo sia provata l'attività larvicida contro H. contortus (Blaszkowska
et al., 2013).
Fusarium solani, Paecilomyces variotii, Paecilomyces viridis sono anch'essi stati indicati
come possibili agenti nei confronti della lotta biologica a Ascaris sp.
50

Nella stessa ricerca Acremonium alabamense, Alternaria chlamydospora, Cladosporium
herbarum hanno mostrato attività di inibizione dello sviluppo larvale senza
colonizzazione delle uova (Blaszkowska et al., 2013).
Blaszkowska et al. (2013) ha rilevato alterazioni a carico di uova e embrioni con le
seguenti specie testate: Aspergillus fumigatus, Aspergillus terreus, , Penicillium
51
Illustrazione 8: Colonizzazione della parete esterna di uova di Ascaris suum con Paecilomyces variotii Blaszkowska, J., Kurnatowski, P., Wojcik, A., Goralska, K. Szwabe, K. (2014). In vitro evaluation of the ovistatic and ovicidal effect of the cosmopolitan filamentous fungi isolated from soil on Ascaris suum eggs. Veterinary parasitology 199, 165–71

expansum, Fusarium oxysporum Trichothecium roseum.
2.2 FITOTERAPIA
La fitoterapia è quella scienza che studia l'utilizzo di piante o estratti di piante come
trattamento medico. Le piante hanno costituito per millenni la principale fonte di
molecole medicamentose per l'uomo e gli animali e sono state progressivamente
sostituite dalle molecole di sintesi. Nel 2800 a.C veniva scritto in Cina il Pen ts'ao ching,
primo grande erbario cinese che conteneva 237 preparazioni tradizionali.
I principi attivi contenuti in un solo vegetale sono estremamente vari e la loro efficacia
può dipendere dalla loro azione sinergica. L'efficacia dell'estratto vegetale dipende quindi
dalla combinazione di più composti, detto “fitocomplesso”.
I meccanismi per cui il consumo di una pianta avrebbe effetti antiparassitari rimane
spesso sconosciuto; in alcune è stato suggerita la correlazione tra il consumo e un
incremento della risposta immunitaria, mentre nella maggior parte vengono spesso citati
composti presenti nella pianta con attività diretta sul parassita come saponine, alcaloidi,
aminoacidi non proteici, tannini, polifenoli e glicosidi.
52

Questi composti sono spesso metaboliti secondari delle piante, associati a meccanismi di
difesa nei confronti del pascolamento degli animali.
Quindi, le ricerche in vivo sull'utilizzo delle piante, dovrebbero prendere in
considerazione anche eventuali effetti anti-nutrizionali o tossici (Athanasiadou and
Kyriazakis, 2007).
Il tanaceto (Tanacetum vulgare), ad esempio, possiede una forte capacità antielmintica
grazie al contenuto di tuione. Ma è anche noto per l'effetto neurotossico e abortigeno che
lo rende inutilizzabile come principio terapeutico (Campanini, 2004).
Si ritiene che molte piante abbiano azione antielmintica anche se per molte di esse non ci
sono evidenze scientifiche, mancano infatti test in grado di definire al meglio la loro
attività (Githiori et al., 2006).
Estratti derivanti da Chenopodium ambrosoides venivano utilizzati, in passato, nel Regno
Unito per trattare infestazioni da Strongylus, Parascaris e Ascaris sia negli animali che
nell'uomo (Gibson, 1965).
Artemesia santonica, Inula helenium, Albizzia lebbek (El Garhy and Mahmoud, 2002),
53

Artemisia pallens (Nakhare and Garg, 1991), Carica papaya (Kermanshai et al.,
2001) hanno dimostrato attività ovicida e adulticida in test in vitro nei confronti A.suum.
Anche la Mentha spicata risulta molto efficace e grazie alla presenza di betasitosterolo il
quale ha dimostrato attività in vitro su Ascaris suum (Villaseñor et al., 2002).
Anche ai semi di zucca, grazie al principio attivo cucurbitina, e alla corteccia delle radici
del melograno (Punica granatum), per l'alcaloide pelletierina, viene attribuita attività
antilemintica. Queste sostanze sarebbero in grado di paralizzare il nematode (Campanini,
2004).
54

CAPITOLO 3: MATERIALI E METODI
3.1 OBBIETTIVi
Il nostro studio aveva come obbiettivo quello di saggiare in vitro l'attività dei funghi
ambientali Trichoderma sp., Beauveria sp, Acremonium sp., Trichoderma viride.,
Trichothecium sp. e di estratti vegetali nei confronti delle uova di Ascaris suum.
La scelta dei funghi è stata motivata in maniera da valutare funghi già saggiati in
letteratura nei confronti delle uova di ascaridi come Tricothecium roseum e Acremonium
sp. e funghi non ancora saggiati come Trichoderma spp, molto presente nell'ambiente, o
Beauveria sp., fungo entomopatogeno.
3.2 RACCOLTA DEI PARASSITI
La raccolta degli elminti adulti è stata effettuata presso un macello suino della provincia
di Pisa. L'apparato gastroenterico dei suini macellati, una volta isolato, è stato sottoposto
a una visita ispettiva al fine di individuare ascaridi adulti. I parassiti venivano quindi
prelevate dal lume dell'organo, posti in contenitori di vetro in ambiente umido e
trasportate nel Laboratorio di Parassitologia del Dipartimento di Scienze Veterinarie
55

Università di Pisa.
3.3 PREPARAZIONE DEI CAMPIONI
Le femmine adulte venivano individuate e dissezionate. Quindi, per le prove con i
funghi, si procedeva alla raccolta delle uova in acido solforico (soluzione 4N) (Oksanen
et al., 1990).
La concentrazione di uova veniva calcolata attraverso l'utilizzo di una camera di conta
(camera di Burker). Per le prove con funghi ambientali, le uova sono state incubate in
monostrato per 21 giorni a temperatura ambiente (18-20 C°) in piastre di Petri. Al 21°
giorno è stata controllata la corretta embrionatura prima di procedere con la fase
successiva. Mentre per le prove con estratti vegetali le uova sono state utilizzate subito
dopo essere state estratte.
3.4 PROVE IN VITRO CON FUNGHI AMBIENTALI NEI CONFRONTI DI UOVA
EMBRIONATE
Dopo 21 giorni, 1mL di sospensione di uova embrionale (contenente circa 100 uova) è
56

stato distribuito sulla superficie del terreno solido di piastre Petri contenenti agar
Sabouraud addizionato di cicloeximide e cloramfenicolo. Contestualmente si è proceduto
alla semina dei diversi funghi in ciascuna piastra, ad eccezione di una piastra lasciata
come controllo.
I funghi isolati provenivano da ceppi ambientali presenti nella micoteca del Laboratorio
di Micologia del Dipartimento di Scienze Veterinarie ed erano stati mantenuti in terreno
per dermatofiti. Dopo la semina dei funghi e la distribuzione delle uova, le piastre sono
state incubate al riparo della luce solare diretta per 15 giorni a temperatura ambiente
(circa 25°C) prima di essere esaminate, come descritto da Blaszkowska et al., (2013).
I funghi selezionati per le prove sono stati:
- Trichoderma sp..
-Beauveria sp.
-Acremonium sp.
- Trichoderma viride
57

-Trichothecium sp.
Dopo 15 giorni è stata valutata la crescita dei funghi nella piastra e sono stati allestiti
vetrini per verificare l'azione di questi sulle uova di Ascaridi. In particolare veniva
valutata la crescita del micelio, la colonizzazione del guscio da parte delle ife e eventuali
alterazioni a carico del guscio o della larva.
Lo stesso tipo di indagine è stata eseguita utilizzando 2% water agar come terreno di
coltura.
3.5 PROVE IN VITRO CON OLI ESSENZIALI
Gli estratti utilizzati per queste prove sono di varia natura, alcuni di essi (estratti di
Acacia, Acacia nilotica e Balanites aegyptiaca) sono stati forniti dal National Research
Center di Dokki (Egitto) nell'ambito di una collaborazione internazionale, e estratti dal
Dipartimento di Farmacologia dell'Università di Pisa. Mentre gli altri sono stati forniti
dalla ditta Flora s.r.l. (Lorenzana, Pisa, Italia).
58

-Lista degli estratti di oli essenziali utilizzati:
Oli diluiti al 5% in DMSO.
-Olio essenziale di Menta (proveniente da National Research Center di Dokki)
- Olio essenziale di Origano (proveniente da National Research Center di Dokki)
Oli diluiti al 5% nel mezzo di coltiv azione delle uova
- Olio essenziale di timo bianco (Thymus serpyllum) n° lotto 111898
-Olio essenziale di origano (Origanum vulgare) n° lotto 60326
-Olio essenziale di eucalipto (Eucalyptus globulus) n° lotto 129264
-Olio essenziale di maggiorana (Origanum majorana) n° lotto 112039
-Olio essenziale di menta (Mentha spicata) n° lotto 80059
-Olio essenziale di anice (Illicium verum) n° lotto 128884
-Olio essenziale di finocchio dolce (Foenicolum vulgare) n° lotto 128407
Estratti metanolici diluiti al 5% nel mezzo di coltivazione delle uova:
-Estratti di Acacia (Acacia auriculiformis)
59

-Estratti di Dattero del Deserto (Balanites aegyptiaca)
-Caratterizzazione degli oli essenziali
La composizione degli oli essenziali è stata caratterizzata utilizzando la
gascromatografia- spettrometria di massa (GS-MS); in modo da poter mettere in
relazione l'attività biologica degli oli con la loro composizione chimica. L'analisi è stata
condotta utilizzando un Varian CP-3800 gas- 3 C°/min. L'identificazione dei costituenti si
basa sul confronto tra il tempo di ritenzione ottenuto e quello di campioni puri,
confrontando i loro indici lineari con serie di n-alcani (C8-C23). Altre identificazioni
sono state possibili utilizzando dati presenti in letteratura .
Le prove sono state effettuate utilizzando Piastre Petri da 60mm.
Per ogni estratto da saggiare sono state allestite tre piastre contenenti 2 mL di
sospensione di uova non embrionate ciascuna.
Ad ogni piastra è stato aggiunto l'estratto vegetale al 5%.
Per ogni prova effettuata è stata preparata una piastra senza estratto come controllo dello
60

sviluppo larvale delle uova.
Le uova sono venivano lasciate due giorni a contatto con l'estratto e successivamente
lavate tre volte tramite centrifugazione al fine di allontanare lo stesso.
Dopo l'ultimo lavaggio, le uova venivano sospese in acido solforico e messe a
embrionare.
Dopo 21 giorni dal lavaggio, mantenendo le piastre a T ambiente, venivano allestiti
vetrini al microscopio ottico al fine di determinare la percentuale di uova embrionate e
valutare l'eventuale efficacia dell'estratto.
61

CAPITOLO 4: RISULTATI
4.1 SAGGIO DEI FUNGHI AMBIENTALI
In agar Sabouraud addizionato di cicloeximide e cloramfenicolo tutti i funghi si sono
sviluppati correttamente ma all'esame microscopico nessuno ha mostrato attività
nematofaga: le uova erano integre e le larve vitali.
Nei funghi coltivati in 2% water agar non si sono sviluppate colonie, ad eccezione di
62
Illustrazione 9: Funghi coltivati in Agar Sabouraud

Trichothecium sp. e Beauveria sp in cui abbiamo riscontrato crescita del micelio.
63
Illustrazione 10: Funghi in 2% water agar

64
Tabella 3
SPECIE ESAME MACROSCOPICO ESAME MICROSCOPICO
assenza di crescita del micelio
assenza di crescita del micelio
assenza di crescita del micelio
in agar Sabouraud
Trichoderma sp.
Colonie di colore bianco, rotonde, grandezza 2-2,5 cm. Assenza di nuove colonie dopo l'aggiunta delle uova.
Le uova sono correttamente embrionate e la larva interna vitale. Non vi sono segni di
colonizzazione o danneggiamento del guscio.
Beauveria bassiana
Colonie di colore bianco, rotonde, piane e di aspetto lanoso. Assenza di nuove colonie dopo l'aggiunta delle uova.
Trichothecium sp.Colonie di colore rosato, piane, polverose e granulose
in water-agar
Trichoderma sp.
Beauveria sp.
Scarsa crescita del micelio. Colonie piccole e poco numerose.
Non è stata individuata colonizzazione del guscio da parte delle ife. Si riscontrano molte larve fuoriuscite dal guscio e avitali
Acremonium sp.
Trichoderma viride
Trichothecium sp.
Scarsa crescita del micelio. Colonie piccole e poco numerose.
Si ritrovano uova correttamente embrionate con larva vitale e altre con segni di danneggiamento e colonizzazione del guscio e larva avitale. Presenza di larve libere avitali.

In Beauveria sp. non siamo riusciti ad individuare segni di azione diretta del fungo nei
confronti delle uova nonostante fossero presenti larve libere, segno indiretto di
danneggiamento del guscio.
In Trichothecium sp., oltre alla presenza di larve libere, abbiamo riscontrato segni di
colonizzazione delle uova nonostante difficoltà nell'identificare strutture specifiche.
Le figure numero 12 e 13 mostrano come i gusci appaiono danneggiati, con contorno
irregolare; inoltre vi sono segni di invasione nei confronti della larva.
65

66
Illustrazione 12
Illustrazione 13
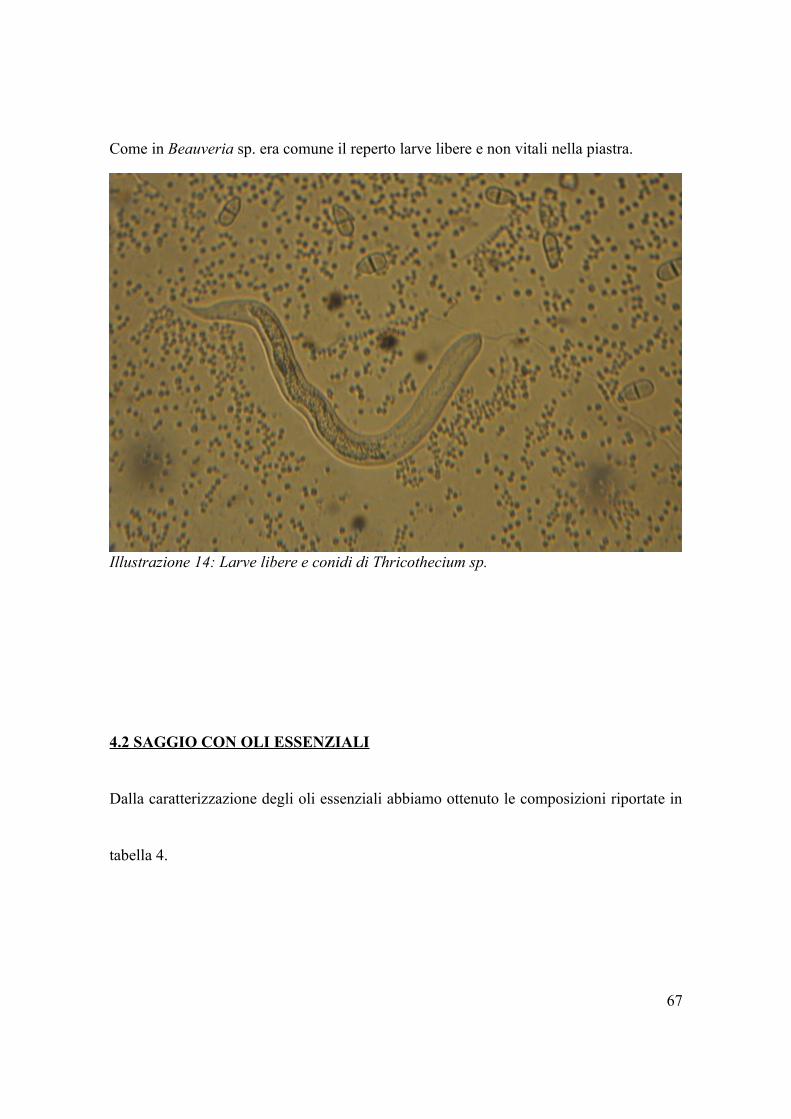
Come in Beauveria sp. era comune il reperto larve libere e non vitali nella piastra.
4.2 SAGGIO CON OLI ESSENZIALI
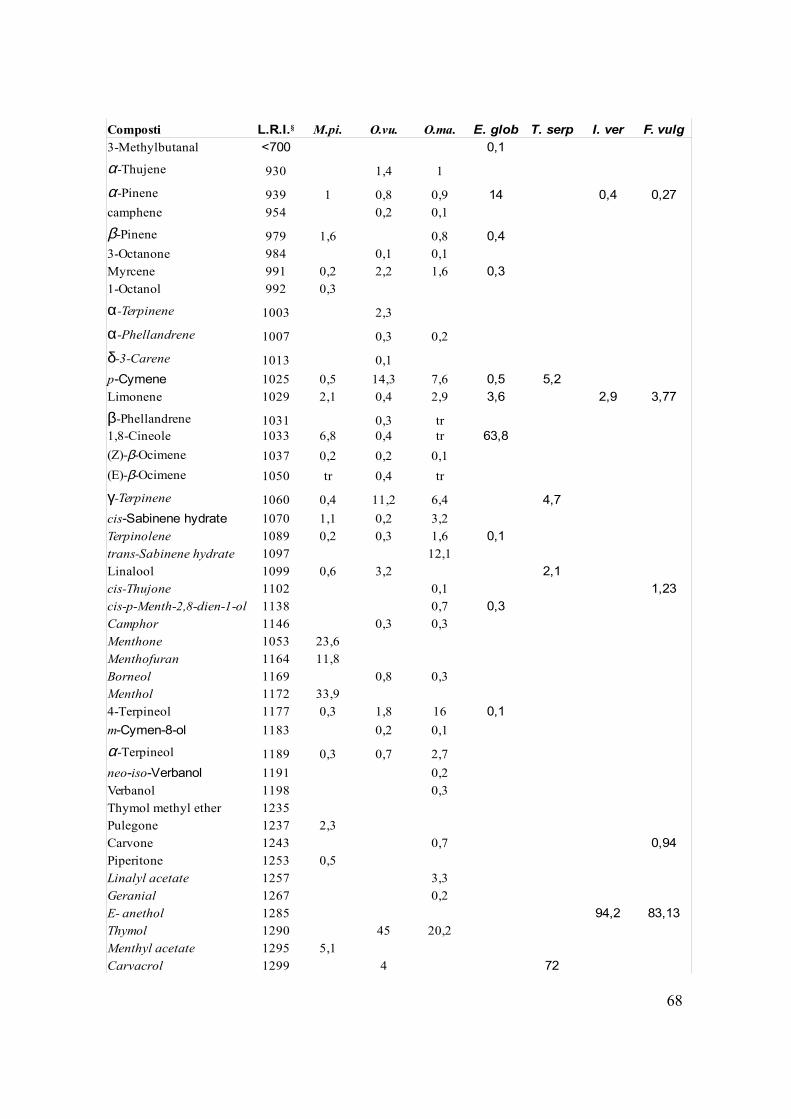
Dalla caratterizzazione degli oli essenziali abbiamo ottenuto le composizioni riportate in
tabella 4.
67
Illustrazione 14: Larve libere e conidi di Thricothecium sp.
68
Composti M.pi. O.vu. O.ma. E. glob T. serp I. ver F. vulg3-Methylbutanal <700 0,1
930 1,4 1
939 1 0,8 0,9 14 0,4 0,27camphene 954 0,2 0,1
979 1,6 0,8 0,43-Octanone 984 0,1 0,1
Myrcene 991 0,2 2,2 1,6 0,31-Octanol 992 0,3
1003 2,3
1007 0,3 0,2
1013 0,1
1025 0,5 14,3 7,6 0,5 5,2Limonene 1029 2,1 0,4 2,9 3,6 2,9 3,77
1031 0,3 tr1,8-Cineole 1033 6,8 0,4 tr 63,8
1037 0,2 0,2 0,1
1050 tr 0,4 tr
1060 0,4 11,2 6,4 4,7
1070 1,1 0,2 3,2
Terpinolene 1089 0,2 0,3 1,6 0,1trans-Sabinene hydrate 1097 12,1
Linalool 1099 0,6 3,2 2,1cis-Thujone 1102 0,1 1,23cis-p-Menth-2,8-dien-1-ol 1138 0,7 0,3Camphor 1146 0,3 0,3
Menthone 1053 23,6
Menthofuran 1164 11,8
Borneol 1169 0,8 0,3Menthol 1172 33,9
4-Terpineol 1177 0,3 1,8 16 0,1
1183 0,2 0,1
1189 0,3 0,7 2,7
1191 0,2
Verbanol 1198 0,3
Thymol methyl ether 1235Pulegone 1237 2,3
Carvone 1243 0,7 0,94Piperitone 1253 0,5
Linalyl acetate 1257 3,3
Geranial 1267 0,2
E- anethol 1285 94,2 83,13Thymol 1290 45 20,2
Menthyl acetate 1295 5,1
Carvacrol 1299 4 72
L.R.I.§
α-Thujene
α-Pinene
β-Pinene
α-Terpinene
α-Phellandrene
δ-3-Carene
p-Cymene
β-Phellandrene
(Z)-β-Ocimene
(E)-β-Ocimene
γ-Terpinene
cis-Sabinene hydrate
m-Cymen-8-ol
α-Terpineol
neo-iso-Verbanol

69
Bicyclogermacrene 1500 0,4 0,2
1502 2,6 1,4
1514 0,3 0,1
1523 0,3
1525 tr
1539 tr
1583 0,1 0,51593 0,1 0,4 0,31593 0,4 0,9
1632 0,5
1651 0,8
1654 0,6
0,3
0,4
3
5,12
2,61
β-Bisabolene
γ-Cadinene
δ-Cadinene
β-Sesquiphellandrene
α-Cadinene
Caryophyllene oxideSpathulenolViridiflorol
γ-Eudesmol
β-Eudesmol
α-Eudesmol
Guaiol
Ledol
Globulol
estragole
p-anisaldehyde
§Linear Retention Index calculated on a DB-5ms column; M.spi.= Mentha spicata; O. vu.= Origanum vulgare; O.ma.= Origanum majorana; E. glob= Eucalyptus globulus; T. serp= Thymus serpyllum; I. ver= Illicium verum; F. vulg= Foeniculum vulgare

I risultati del saggio degli estratti vegetali sono stati riportati in tabella 5.
La tabella mostra come il DMSO, in cui erano diluiti gli oli provenienti dal National
70
Tabella 5- Percentuale di embrionatura delle uova. Nella prima sezione gli estartti provenienti dal National Research Centerdi Dokki, nella seconda quelli forniti dalla ditta Flora s.r.l. e nella terza gli estratti metanolici.
ESTRATTO VEGETALE
controllo 65%DMSO 0%
Origano 0%Menta 0%
controllo 50,0%
Eucalipto 13,6%
Finocchio 31,0%
Menta 18,0%
Maggiorana 33,0%
Origano 10,8%
Timo 12,0%Anice 23,3%
controllo 50,0%
Acacia (foglie) 42,0%
17,0%
46,0%Dattero del deserto (foglie) 20,6%Dattero del deserto (foglie) 30,3%
PERCENTUALE MEDIA
Dattero del deserto (frutto maturo)
Dattero del deserto (epicarpo maturo)

Research Center di Dokki, abbia una forte azione ovicida; le uova infatti si presentavano
significativamente danneggiate, come mostrato in figura. E' stato quindi impossibile
valutare l'effetto degli oli di Origano e Menta diluiti in DMSO.
Gli oli essenziali che hanno mostrato maggiore attività nei confronti delle uova sono stati
quelli di Eucalipto, Origano e Timo inibendo di circa l'80% lo sviluppo larvale.
Gli estratti di Dattero del deserto (ad eccezione dell'estratto ottenuto dalle foglie), Acacia,
Maggiorana e Finocchio hanno avuto un'efficacia blanda verso le uova.
71
Illustrazione 15 Uova coltivate con DMSO

CAPITOLO 5: CONCLUSIONI E DISCUSSIONI
5.1 SAGGIO CON FUNGHI AMBIENTALI
I risultati del saggio sull'efficacia dei funghi ambientali nei confronti delle uova di A.
suum mostra come Trichoderma sp., Acremonium sp., Trichoderma viride, non abbiano
dimostrato attività ovicida e quindi non possono essere considerati funghi potenzialmente
nematofagi.
Beauveria sp. e Trichothecium sp. hanno mostrato attività nei confronti delle uova se
coltivati in terreno privo di sostanze organiche utilizzabili; e sono stati gli unici capaci di
formare colonie in questo tipo di terreno, probabilmente perché hanno utilizzato come
substrato organico le uova di A. suum presenti nella piastra.
Il reperto di larve libere nella piastra, in relazione al ciclo biologico degli ascaridi,
suggerisce che il guscio delle uova sia stato digerito al punto da permettere la schiusa
delle uova e la fuoriuscita delle larve.
Inoltre in Trichothecium sp. il possibile utilizzo delle uova come fonte di nutrimento da
parte del micelio, era evidenziato dallo sviluppo delle ife sulla superficie delle stesse.
72

Questi reperti preliminari dimostrano la potenziale attività nematofaga di questi due
funghi nei confronti delle uova di A. suum. Inoltre, suggeriscono che tale potenziale
attività potrebbe estrinsecarsi anche nei confronti di uova di altri ascaridi o addirittura di
altri elminti.
Blaszkowska et al.(2013) ha dimostrato la capacità di T. roseum di danneggiare
enzimaticamente uova e embrione di A. suum in in 2% Potato-dextrose agar.
Beauveria bassiana è un fungo entomopatogeno già disponibile in commercio per la lotta
biologica a artropodi parassiti delle piante, ma non ci sono studi che confermano la sua
efficacia nei confronti di elminti.
Dobbiamo sottolineare però come questa attività sia stata riscontata solo in determinate
condizioni di coltura difficilmente presenti nell'ambiente; Beauveria sp. e Trichothecium
sp. infatti non si sono comportati allo stesso modo se coltivati in terreni ricchi di sostanze
nutritive (agar Sabouraud) o in un terreno povero (2% water agar).
Questo rappresenta un forte limite al loro utilizzo in vivo; la colonizzazione delle uova
sembra infatti dovuta alle esigenze di assimilazione di sostanza organica in particolari
73

condizioni di coltura piuttosto che a una attitudine di questi funghi nei confronti delle
uova.
Ulteriori studi potrebbero inoltre valutare l'aspetto quantitativo del potere patogeno del
fungo in base alle uova parassitate, o eventuali alterazioni nei tempi di sviluppo larvale;
entrambi aspetti non presi in considerazione dalla nostra ricerca.
5.2 SAGGIO CON OLI ESSENZIALI
Il nostro studio suggerisce che gli oli essenziali di Timo, Origano, Eucalipto possono
essere considerati validi candidati per ulteriori prove nei confronti delle uova di A. suum.
Dalle gas cromatografia possiamo osservare come questi oli essenziali siano
rispettivamente composti maggiormente da monoterpeni come Carvacrolo, Timolo e 1,8-
cineolo.
Questi non sono stati testati in letteratura nei confronti delle uova di Ascaridi; tuttavia
negli ultimi anni sono stati effettuati numerosi studi riguardanti principalmente la lotta a
parassiti vegetali.
I terpeni fenolici e i terpeni ossigenati risultano essere maggiormente nematocidi rispetto
74

agli idrocarburi terpenici. Quelli che hanno mostrato maggiore attività nei confronti di
Caenorhabditis elegans sono, in ordine decrescente: carvacrolo, timolo, nerolidolo,
α-terpinene, geraniolo, citronellolo, farnesolo, limonene, pseudoionone and eugenolo
(Fawzia et al., 2013; Lei et al., 2010).
Sono inoltre presenti studi che dimostrano l'attività adulticida dell'olio essenziale di
eucalipto verso Pheretima posthuma,, nematode a vita libera simile agli ascaridi (Taur et
al., 2010), e del carvacrolo nei confronti delle uova di Haemonchus contortus (Liang et
al. , 2013).
Gli estratti dei frutti di Balanites aegyptiaca non hanno mostrato attività ovicida, in
disaccordo con Shalaby et.al. (2013) che avevano dimostrato l'attività adulticida e
ovicida di questi frutti nei confronti di Toxocara vitulorum.
L'estratto di Acacia auriculiformis non ha avuto effetti verso le uova, nonostante sia stata
riportata la capacità di inibire lo sviluppo larvale di ciatostomi da parte di altre specie di
Acacia (Payne et al., 2013), così come l'effetto verso cestodi adulti (Ghosh et al., 1996) e
verso Dirofilaria per l'azione nei confronti di Wolbachia (Datta et al., 2009).
75

Secondo il nostro studio anche la Mentha spicata potrebbe essere presa in considerazione
per ulteriori ricerche; in letteratura sono presenti studi riguardanti l'attività
antiparassitaria di Mentha piperita verso Schistosoma sp. dovuta alla presenza di
limonene (Moraes et al., 2013; Katiki et al., 2011).
Attualmente sono in corso test di sensibilità dei monoterpeni che hanno avuto maggior
effetto nei confronti delle uova di A. suum.
Inoltre è stato dimostrato come il DMSO (dimetilsolfossido), alle concentrazione da noi
utilizzata, abbia una forte azione ovicida.
Per concludere, il nostro studio preliminare ha mostrato come gli oli essenziali di
origano, timo e eucalipto possono essere presi in considerazione per ulteriori studi sulla
loro attività ovicida. In particolare dovrebbero essere testate l'attività a differenti
concentrazioni ed individuare la Dose Letale 50 e la Concentrazione Minima Inibente.
76

BIBLIOGRAFIA
Alva-Valdes, R., Wallace, D.H., Foster, A.G., Ericsson, G.F., Wooden, J.W., (1989).Efficacy of an in- feed ivermectin formulation against gastrointestinal helminths,lungworms, and sarcoptic mites in swine. American Journal of Veterinary Research50, 1392-1395.
Anderson, T. J. (1995). Ascaris infections in humans from North America: molecularevidence for cross-infection. Parasitology 110 ( Pt 2, 215–9.
Anderson, T. J., Romero-Abal, M. E. and Jaenike, J. (1993). Genetic structure andepidemiology of Ascaris populations: patterns of host affiliation in Guatemala.Parasitology 107 Pt 3, 319–34.
Araújo, J. V, Braga, F. R., Silva, a R., Araujo, J. M. and Tavela, a O. (2008). In vitroevaluation of the effect of the nematophagous fungi Duddingtonia flagrans,Monacrosporium sinense, and Pochonia chlamydosporia on Ascaris suum eggs.Parasitology research 102, 787–90. doi:10.1007/s00436-007-0852-9.
Athanasiadou, S. and Kyriazakis, I. (2007). Plant secondary metabolites: antiparasiticeffects and their role in ruminant production systems. Proceedings of the NutritionSociety 63, 631–639. doi:10.1079/PNS2004396.
Barron, G. (1992). Lignolytic and cellulolytic fungi as predators and parasites. TheFungal Community, Its Organization and Role in the Ecosystems pp. 311–326.
Basile C.G. (2013). Il mercato dei suini: produzioni e consumo. OsservatorioAgroalimentare Lombardo. 22
Blaszkowska, J., Wojcik, A., Kurnatowski, P. and Szwabe, K. (2013). Biologicalinteractions between soil saprotrophic fungi and Ascaris suum eggs. Veterinaryparasitology 196, 401–8. doi:10.1016/j.vetpar.2013.02.029.
Blaszkowska, J., Kurnatowski, P., Wojcik, A., Goralska, K. and Szwabe, K. (2014).In vitro evaluation of the ovistatic and ovicidal effect of the cosmopolitanfilamentous fungi isolated from soil on Ascaris suum eggs. Veterinary parasitology199, 165–71. doi:10.1016/j.vetpar.2013.10.026.
Bradley, R.E., Guerrero, J., Becker, H.N., Michael, B.F., Newcomb, K., (1983).Flubendazole: dose range and efficacy studies against common internal parasites ofswine. American Journal of Veterinary Research 44, 1329-1333.
Braga, F. R., Araújo, J. V. (2013). Nematophagous fungi for biological control ofgastrointestinal nematodes in domestic animals. Applied Microbiology andBiotechnology, 98(1), 71–82. doi:10.1007/s00253-013-5366-z
Campanini, E. (2004). Dizionario di fitoterapia e piante medicinali. Milano. TecnicheNuove. 2004
77

Cox, F. E. G. (2002). History of Human Parasitology. Clin. Microbiol. Rev., 15, 4,595-612 . doi:10.1128/CMR.15.4.595.
Datta, S., Maitra, S., Gayen, P., & Sinha Babu, S. P. (2009). Improved efficacy oftetracycline by acaciasides on Dirofilaria immitis. Parasitology Research, 105(3),697–702. doi:10.1007/s00436-009-1441-x
Dold, C. and Holland, C. V (2011). Investigating the underlying mechanism ofresistance to Ascaris infection. Microbes and infection / Institut Pasteur 13, 624–31.doi:10.1016/j.micinf.2010.09.013.
El Garhy, M. F. and Mahmoud, L. H. (2002). Anthelminthic efficacy of traditionalherbs on Ascaris lumbricoides. Journal of the Egyptian Society of Parasitology 32,893–900.
Eichhorn, L., Zimmermann, W., Gottstein, B., Frey, C.F., Doherr, M.G., Zeeh, F.,(2010). Gastrointestinal nematodes in Swiss pig farms - an explorative overview. In:21st IVPS Congress, Vancouver, Canada, July 18-21, 2010, p. 808.
Eijck, I.A., Borgsteede, F.H., (2005). A survey of gastrointestinal pig parasites onfree-range, organic and conventional pig farms in The Netherlands. Veterinaryresearch communications 29, 407- 414.
EUROSTAT (2013). Number of pigs. http://epp.eurostat.ec.europa.eu/tgm/table.do?tab=table&init=1&plugin=0&language=en&pcode=tag00018
Fawzia H. Abdel-Rahman, Nina M. Alaniz, Mahmoud A. Saleh. (2013) Nematicidal activity of terpenoids. Jornal of Enviromental Science and HealthVol. 48, Iss. 1,
Ferreira, S. R., Araújo, J. V, Braga, F. R., Araujo, J. M., Carvalho, R. O., Silva, A.R., Frassy, L. N. and Freitas, L. G. (2011a). Ovicidal activity of seven Pochoniachlamydosporia fungal isolates on Ascaris suum eggs. Tropical animal health andproduction 43, 639–42. doi:10.1007/s11250-010-9744-6.
Ferreira, S. R., de Araújo, J. V., Braga, F. R., Araujo, J. M., Frassy, L. N. andFerreira, A. S. (2011b). Biological control of Ascaris suum eggs by Pochoniachlamydosporia fungus. Veterinary research communications 35, 553–8.doi:10.1007/s11259-011-9494-6.
Frontera, E., Serrano, F., Reina, D., Alcaide, M., Sánchez-López, J. and Navarrete,I. (2003). Serological responses to Ascaris suum adult worm antigens in Iberianfinisher pigs. Journal of helminthology 77, 167–72. doi:10.1079/JOH2002163.
Frontera, E., Roepstorff, A., Serrano, F. J., Gázquez, A., Reina, D. and Navarrete, I.(2004). Presence of immunoglobulins and antigens in serum, lung and smallintestine in Ascaris suum infected and immunised pigs. Veterinary parasitology 119,59–71. doi:10.1016/j.vetpar.2003.09.022.
Geldhof, P., Vercauteren, I., Vercruysse, J., Knox, D. P., Van Den Broeck, W. andClaerebout, E. (2004). Validation of the protective Ostertagia ostertagi ES-thiolantigens with different adjuvantia. Parasite immunology 26, 37–43.doi:10.1111/j.0141-9838.2004.00681.x.
78

Gerwert, S., Failing, K. and Bauer, C. (2002). Prevalence of levamisole andbenzimidazole resistance in oesophagostomum populations of pig-breeding farms inNorth Rhine-Westphalia, Germany. Parasitology research 88, 63–8.
Ghosh, N. K., Babu, S. P., Sukul, N. C., & Ito, A. (1996). Cestocidal activity of Acaciaauriculiformis. Journal of Helminthology, 70(2), 171–2. Retrieved fromhttp://www.ncbi.nlm.nih.gov/pubmed/8960214
Gibson (1965). Veterinary anthelmintic medication. Technical Communication no. 33 ofthe Commonwealth Bureau of Helminthology 206.
Githiori, J. B., Athanasiadou, S. and Thamsborg, S. M. (2006). Use of plants in novelapproaches for control of gastrointestinal helminths in livestock with emphasis onsmall ruminants. Veterinary parasitology 139, 308–20.doi:10.1016/j.vetpar.2006.04.021.
Haugegaard, J., (2010). Prevalence of nematodes in Danish industrialized sow farmswith loose housed sows in dynamic groups. Vet Parasitol 168, 156-159.
Helwigh, A. B. and Nansen, P. (1999). Establishment of Ascaris suum in the pig:development of immunity following a single primary infection. Acta veterinariaScandinavica 40, 121–32.
Hsueh, Y.-P., Mahanti, P., Schroeder, F. C. and Sternberg, P. W. (2013).Nematode-trapping fungi eavesdrop on nematode pheromones. Current biology : CB 23, 83–6. doi:10.1016/j.cub.2012.11.035.
Jansson HB, Tunlid A, N.-H. B. (1997). Biological control: Nematode. Fungalbiotechnology pp 38–50.
Jex, A. R., Liu, S., Li, B., Young, N. D., Hall, R. S., Li, Y., Yang, L., Zeng, N., Xu, X.,Xiong, Z., Chen, F., Wu, X., Zhang, G., Fang, X., Kang, Y., Anderson, G. a,Harris, T. W., Campbell, B. E., Vlaminck, J., Wang, T., Cantacessi, C.,Schwarz, E. M., Ranganathan, S., Geldhof, P., Nejsum, P., Sternberg, P. W.,Yang, H., Wang, J., Wang, J. and Gasser, R. B. (2011). Ascaris suum draftgenome. Nature 479, 529–33. doi:10.1038/nature10553.
Joachim, A., Dulmer, N., Daugschies, A., Roepstorff, A., (2001). Occurrence ofhelminths in pig fattening units with different management systems in NorthernGermany. Veterinary Parasitology 96, 135-146.
Katiki LM, Chagas AC, Bizzo HR, Ferreira JF, Amarante AF. (2011) Antihelmintichactivity of Cymbopogon martinii, Cymbopogon schoenanthus and Mentha piperitaessential oils evaluated in four different in vitro test. Vet Parasitol; 183: 103-108
Kaplan, R. M. (2004). Drug resistance in nematodes of veterinary importance: a statusreport. Trends in parasitology 20, 477–81. doi:10.1016/j.pt.2004.08.001.
Kennedy, T., Lucas, M.J., Froe, D.L., (1987). Comparative Efficacy of PyrantelTartrate, Ivermectin and Fenbendazole against Experimentally Induced ImmatureAscaris-Suum in Pigs. Agri-Practice 8, 19-21.
Kermanshai, R., McCarry, B. E., Rosenfeld, J., Summers, P. S., Weretilnyk, E. a andSorger, G. J. (2001). Benzyl isothiocyanate is the chief or sole anthelmintic in
79

papaya seed extracts. Phytochemistry 57, 427–35.
Knecht, D., Jankowska, A. and Zaleśny, G. (2012). The impact of gastrointestinalparasites infection on slaughter efficiency in pigs. Veterinary parasitology 184,291–7. doi:10.1016/j.vetpar.2011.09.006.
Labare, M. P., Soohoo, H., Kim, D., Tsoi, K. yan, Liotta, J. L. and Bowman, D. D.(2013). Ineffectiveness of a quaternary ammonium salt and povidone-iodine for theinactivation of Ascaris suum eggs.
Laugier, C., Sevin, C., Ménard, S. and Maillard, K. (2012). Prevalence of Parascarisequorum infection in foals on French stud farms and first report of ivermectinresistant P equorum populations in France. Veterinary Parasitology, 188, 185-189.
Lei J., Leser M., Enan L (2010) Nematicidal activity of two monoterpenoids and SER-2tyramina receptors of Caenorhabditis elegan. Biochemical Pharmacology,79,1062-1071.
Liang Zhu, Jiali Dai, Li Yang, Jun Qiu, Anthelmintic activity of Arisaemafranchetianum and Arisaema lobatum essential oils against Haemonchus contortus,Journal of Ethnopharmacology, Volume 148, Issue 1, 21 June 2013, Pages 311-316
Lichtensteiger, C.A., DiPietro, J.A., Paul, A.J., Neumann, E.J., Thompson, L.,(1999). Persistent activity of doramectin and ivermectin against Ascaris suum inexperimentally infected pigs. Veterinary Parasitology 82, 235-241.
Linford, M. B. (1937). Stimulated activity of natural enemies of nematodes. Science(New York, N.Y.) 85, 123–4. doi:10.1126/science.85.2196.123.
Liu, G.-H., Wu, C.-Y., Song, H.-Q., Wei, S.-J., Xu, M.-J., Lin, R.-Q., Zhao, G.-H.,Huang, S.-Y. and Zhu, X.-Q. (2012). Comparative analyses of the completemitochondrial genomes of Ascaris lumbricoides and Ascaris suum from humans andpigs. Gene 492, 110–6. doi:10.1016/j.gene.2011.10.043.
Lundenheim, N., Holmgren, N., (2010). Prevalence of lesions found at slaughter amongSwedish fattening pigs. In: 21st IVPS Congress, Vancouver, Canada, July 18-21, p.925.
Luo, H., Li, X., Li, G., Pan, Y. and Zhang, K. (2006). Acanthocytes of Strophariarugosoannulata function as a nematode-attacking device. Applied and environmentalmicrobiology 72, 2982–7. doi:10.1128/AEM.72.4.2982-2987.2006.
Lýsek, H. and Krajcí, D. (1987). Penetration of ovicidal fungus Verticilliumchlamydosporium through the Ascaris lumbricoides egg-shells. Folia parasitologica34, 57–60.
Marchesi, B. (2009). Endoparassiti del suino: zoonosi e studio dei fattori di rischio. Tesidi dottorato. Corso di dottorato in Parassitologia e Malattie Parassitarie deglianimali. Università degli studi di Bologna. XXI ciclo. Discussa nell'a.a. 2008/09.Rell. Prof. Giovanni Poglayen.
Marchiondo, A.A., Szanto, J., (1987), Efficacy of dichlorvos, fenbendazole, andivermectin in swine with induced intestinal nematode infections. American Journalof Veterinary Research 48, 1233-1235.
80

Marti, O.G., Stewart, T.B., Hale, O.M., (1978), Comparative efficacy of fenbendazole,dichlorvos, and levamisole HCI against gastrointestinal nematodes of pigs. TheJournal of parasitology 64, 1028-1031.
Matsumoto, Y., Suzuki, S., Nozoye, T., Yamakawa, T., Takashima, Y., Arakawa, T.,Tsuji, N., Takaiwa, F. and Hayashi, Y. (2009). Oral immunogenicity and protectiveefficacy in mice of transgenic rice plants producing a vaccine candidate antigen(As16) of Ascaris suum fused with cholera toxin B subunit. Transgenic research 18,185–92. doi:10.1007/s11248-008-9205-4.
Moraes, J. de, Almeida, A. A. C., Brito, M. R. M., Marques, T. H. C., Lima, T. C.,Sousa, D. P. de, … Freitas, R. M. (2013). Anthelmintic activity of the naturalcompound (+)-limonene epoxide against Schistosoma mansoni. Planta Medica,79(3-4), 253–8. doi:10.1055/s-0032-1328173
Murrell, K. D., Eriksen, L., Nansen, P., Slotved, H. C. and Rasmussen, T. (1997).Ascaris suum: a revision of its early migratory path and implications for humanascariasis. The Journal of parasitology 83, 255–60.
Nakhare, S. and Garg, S. C. (1991). Anthelmintic activity of the essential oil ofartemisia pallens wall. Ancient science of life 10, 185–6.
Nejsum, P., Parker, E. D., Frydenberg, J., Roepstorff, A., Boes, J., Haque, R. andAstrup, I. (2005). Ascariasis Is a Zoonosis in Denmark. J. Clin. Microbiol. 43,1142–1148. doi:10.1128/JCM.43.3.1142.
Nejsum, P., Thamsborg, S. M., Petersen, H. H., Kringel, H., Fredholm, M. andRoepstorff, A. (2009). Population dynamics of Ascaris suum in trickle-infectedpigs. The Journal of parasitology 95, 1048–53. doi:10.1645/GE-1987.1.
Nissen, S., Poulsen, I.H., Nejsum, P., Olsen, A., Roepstorff, A., Rubaire-Akiiki, C.,Thamsborg, S.M., (2011). Prevalence of gastrointestinal nematodes in growingpigs in Kabale District in Uganda. Trop Anim Health Prod 43, 567-572.
Nordbring-hertz, B. and Tunlid, A. (2006). Nematophagous Fungi. Encyclopedia ofLife Sciences. 1–11. doi:10.1038/npg.els.0004293.
Oakley, G.A., (1974). Activity of levamisole hydrochloride administered subcutaneouslyagainst A suum infections in pigs. The Veterinary record 95, 190-193.
Oksanen, A, Eriksen, L, Roepstorff, A, Ilsøe, B, Nansen, P et al. (1990)Embryonation and infectivity of Ascaris suum eggs. A comparison of eggs collectedfrom worm uteri with eggs isolated from pig faeces. Acta veterinaria Scandinavica vol. 31 (4) p. 393-8
Payne HE., Kotze AC, Durmic Z, Vercoe PE. (2013). Australian plants showanthelmintic activity toward equine cyathostomins in vitro. Veterinary Parasitology.196. 1-2
Pecheur, M., (1983). Efficacy of Oxibendazole against Oesophagostomum dentatum andAscaris suum of Pigs. Ann Med Vet 127, 203-208.
Peng, W., Yuan, K., Hu, M., Zhou, X. and Gasser, R. B. (2005). Mutationscanning-coupled analysis of haplotypic variability in mitochondrial DNA regions
81

reveals low gene flow between human and porcine Ascaris in endemic regions ofChina. Electrophoresis 26, 4317–26. doi:10.1002/elps.200500276.
Persmark L, B. A. and J. H.-B. (1996). Population dynamics of nematophagous fungiand nematodes in an arable soil: vertical and seasonal fluctuations. Soil Biology andBiochemistry.
Pilitt, P., Lichtenfels, J., Tromba, F. and Madden, P. (1981). Differentiation of LateFourth and Early Fifth Stages of Ascaris suum Goeze, 1782 (Nematoda:Ascaridoidea) in Swine.
Pittman, J.S., Shepherd, G., Thacker, B.J., Myers, G.H., Fransisco, C.J., (2010a).Prevalence of internal parasites in a production system: part I - Sows. In: 21st IPVSCongress, Vancouver, Canada, July 18-21, 2010, p. 806.
Pittman, J.S., Shepherd, G., Thacker, B.J., Myers, G.H., Fransisco, C.J., (2010b).Prevalence of internal parasites in a production system: Part II - Finishing pigs. In:21st IPVS Congress, Vancouver, Canada, July 18-21, 2010, p. 807.
Plumb, D. (2008). Plumb’s Veterinary Drug Handbook. 6th Edition.Wiley-Blackwel.Hoboke. 2008
Poinar, G. and Boucot, A. J. (2006). Evidence of intestinal parasites of dinosaurs.Parasitology 133, 245–9. doi:10.1017/S0031182006000138.
Polley, L.R., Mostert, P.E., (1980), Ascaris suum in Saskatchewan pigs: an abattoirsurvey of prevalence and intensity of infection. The Canadian veterinary journal.21, 307-309.
Roberts, L. S. (2009). Foundation of parasitology.
Roepstorff, A. (1997). Helminth surveillance as a prerequisite for anthelmintic treatmentin intensive sow herds. Veterinary Parasitology 73, 139–151.doi:10.1016/S0304-4017(97)00062-9.
Roepstorff, A., Bjørn, H. and Nansen, P. (1987). Resistance of Oesophagostomum spp.in pigs to pyrantel citrate. Veterinary Parasitology 24, 229–239.doi:10.1016/0304-4017(87)90044-6.
Roepstorff, A., Jorsal, S.E., (1989). Prevalence of helminth infections in swine inDenmark. Veterinary Parasitology 33, 231-239.
Roepstorff, A., Nilsson, O., O'Callaghan, C.J., Oksanen, A., Gjerde, B., Richter,S.H., Ortenberg, E.O., Christensson, D., Nansen, P., Eriksen, L., Medley, G.F.,(1999), Intestinal parasites in swine in the Nordic countries: multilevel modelling ofAscaris suum infections in relation to production factors. Parasitology 119 ( Pt 5),521-534.
Sanchez-Vazquez, M.J., Nielen, M., Gunn, G.J., Lewis, F.I., (2012). Nationalmonitoring of Ascaris suum related liver pathologies in English abattoirs: atime-series analysis, 2005-2010. Veterinary Parasitology 184, 83-87.
Schillhorn van Veen, T.W., Gibson, C.D., (1983). Anthelmintic activity of ivermectin inpigs naturally infected with Ascaris and Trichuris. American Journal of Veterinary
82

Research 44, 1732-1733.
Shalaby HA, El Namaky AH, Khalil FA, Kandil OM (2012) Efficacy of methanolicextract of Balanites aegyptiaca fruits on Toxocara vitulorum. VeterinaryParasitology. 183. 3-4, 386-392.
Silva, a R., Araújo, J. V, Braga, F. R., Benjamim, L. a, Souza, D. L. and Carvalho, R.O. (2011). Comparative analysis of destruction of the infective forms of Trichuristrichiura and Haemonchus contortus by nematophagous fungi Pochoniachlamydosporia; Duddingtonia flagrans and Monacrosporium thaumasium byscanning electron microscopy. Veterinary microbiology 147, 214–9.doi:10.1016/j.vetmic.2010.06.019.
Skallerup, P., Nejsum, P., Jørgensen, C. B., Göring, H. H. H., Karlskov-Mortensen,P., Archibald, A. L., Fredholm, M. and Thamsborg, S. M. (2012). Detection of aquantitative trait locus associated with resistance to Ascaris suum infection in pigs.International Journal for Parasitology 42, 383–391.doi:10.1016/j.ijpara.2012.02.010.
Spratt, D. M. (1997). Endoparasite control strategy: implications for biodiversity ofnative fauna. International Journal for Parasitology, 27-2, 173-180,http://dx.doi.org/10.1016/S0020-7519(96)00147-6.
Steffan, P., Olaechea, F., Roepstorff, A., Bjorn, H., Nansen, P., (1988), Efficacy ofpiperazine dihydrochloride against Ascaris suum and Oesophagostomum species innaturally infected pigs. Vet Rec 123, 128-130.
Stewart, T.B., Bidner, T.D., Southern, L.L., Simmons, L.A., (1984). Efficacy offenbendazole against migrating Ascaris suum larvae in pigs. Am J Vet Res 45,984-986.
Stewart, T.B., Rowell, T.J., (1986). Susceptibility of fourth-stage Ascaris suum larvae tofenbendazole and to host response in the pig. Am J Vet Res 47, 1671-1673.
Stewart, T.B., Fox, M.C., Wiles, S.E., (1996). Doramectin efficacy againstgastrointestinal nematodes in pigs. Veterinary Parasitology 66, 101-108.
Taylor Mike, A., Coop Robert, L. and Wall Richard, L. (2010). Parassitologia emalattie parassitarie degli animali. Roma. EMSI. 2010
Taur DJ, Kulkarni VB, Patil RY. (2010). Chromatographic evaluation and anthelminticactivity of Eucalyptus globulus oil. Pharmacogn Res. 2010;2:125–7
Tielen, M.J., Truijen, W.T., Remmen, J.W., (1976). [The incidence of diseases of thelung and liver in slaughtered pigs as a criterion in the detection of herds in which thedisease is a recurrent problem (author's transl)]. Tijdschrift voor diergeneeskunde101, 962-971.
Van Meensel, J., Kanora, A., Lauwers, L., Jourquin, J., Goossens, L. and VanHuylenbroeck, G. (2010). From research to farm: ex ante evaluation of strategicdeworming in pig finishing. Veterinarni Medicina 55, 483–493.
Vanparijs, O., Hermans, L., Marsboom, R., (1988), Efficacy of flubendazole againstgastrointestinal and lung nematodes in pigs. The Veterinary record 123, 337-339.
83

Vercruysse, J., VanHoof, D., DeBie, S., (1997), Study on the prevalence of white spotsof the liver in pigs in Belgium and its relationship to management practices andanthelmintic treatment. Vlaams Diergeneeskundig Tijdschrift. 66, 28-30
Viggiani (1974). La lotta biologica di tipo convenzionale. In: Atti del X congressonazionale italiano di entomologia. Sassari. 20-25 Maggio, 1974. 161-165
Villaseñor, I. M., Angelada, J., Canlas, A. P. and Echegoyen, D. (2002). BioactivityStudies on B-Sitosterol and its Glucoside. 421, 417–421.
Vlaminck, J. (2013). Evaluation of Ascaris suum haemoglobin as a vaccine anddiagnostic antigen. Tesi di dottorato. Corso di dottorato in Virologia, Parassitoligae Immunologia. Università di Gent. Discussa nell'a.a. 2012-13. Rell. Prof. P.Geldhof Prof. Vercruysse J.
Von Reuss, S. H., Bose, N., Srinivasan, J., Yim, J. J., Judkins, J. C., Sternberg, P. W.and Schroeder, F. C. (2012). Comparative metabolomics reveals biogenesis ofascarosides, a modular library of small-molecule signals in C. elegans. Journal ofthe American Chemical Society 134, 1817–24. doi:10.1021/ja210202y.
Weng, Y.B., Hu, Y.J., Li, Y., Li, B.S., Lin, R.Q., Xie, D.H., Gasser, R.B., Zhu, X.Q.,(2005). Survey of intestinal parasites in pigs from intensive farms in GuangdongProvince, People's Republic of China. Veterinary Parasitology 127, 333-336
Zhou, C., Li, M., Yuan, K., Deng, S. and Peng, W. (2012). Pig Ascaris: an importantsource of human ascariasis in China. Infection, genetics and evolution : journal of molecular epidemiology and evolutionary genetics in infectious diseases 12,1172–7. doi:10.1016/j.meegid.2012.04.016.
84


















