Role of the β-amyloid precursor protein in Alzheimer's disease
5
REVIEWS pressing and activating domains have also been mapped for p53 (Ref. 28). Thus it appears that some proteins con- tain both activating and repressing potential, and the predominant effect depends on promoter context, in a related phenomenon, it has been observed that Kr can act either as a repressor or as an activator depending on its concentration2°, which is con- sistent with genetic evidence that Kr is a positive and negative regulator in a hierarchy of genes involved in Drosophda embryonic development. Repression of various kinds is now well embedded in our understanding of transcriptional regulation. However, there are still gaps in our knowledge, notably concerning the mechanism of action of the active transcriptional repressors. Future work to unravel this process will undoubtedly go hand in hand with a further understanding of transcriptional activation and other events surrounding transcription initiation. Acknowledgements I would like to thank H. C. Hurst, C. A. Austin and others for useful discussion and comments on this manuscript. References 1 Roede,', R. G. (1991) Trends Biochem. Sci. 16, 402-408 2 Workman, J. L. and Buchrnan, A. R. (1993) Trends Biochern. Sci. 18, 90-95 3 Morse, R. H. (1992) Trends Biochem. Sci. 17, 23-26 4 Kageyama, R. and Pastan, I. (1989) Cell 59, 815-825 5 SchOle, R. et al. (1990) Cell 61, 497-504 6 Liu, Y., Yang, N. and Teng, C. T. (1993) Mol. Cell Biol. 13, 1836-1846 7 Benezra, R. et al. (1990) Cell 61, 49-59 8 Ron, D. and Habener, J. F. (1992) Genes Dev. 6, 439-453 9 Deng, T. and Karin, M. (1993) Genes Dev. 7, 479-490 10 Bengal, E. et al. (1992) Cell 68, 507-519 11 YangYen, H. F. et al. (1990) Cell 62,1205-1215 12 Licht, J. b., Grossel, M. J., Fi~e, J. and TIBS 19 - JANUARY 1994 Hansen, U. M. (1990) Nature 346, 76-79 13 Han, K. and Manley, J. L. (1993) Genes Dev. 7,491-503 14 Jaynes, J. B and O'Farreh, P. H. (1991) EMBOJ. 10, 1427-1433 15 Han, K. and Manley, J. L. (1993) EMBO J. 12, 2723-2733 16 Shi, ¥., Seto, E., Chang, L-S. and Shenk, T. (1991) Cell 67, 377-388 17 Madden, S. L., Cook, D. M. and Rauscher, F. J., III (1993) Oncogene 8, 1713-1720 18 Cowell, I. G., Skinner, A. and Hurst, H. C. (1992) /VIOLCell Biol. 12, 3070-3077 19 Baniahmad, A., K6hne, A. C. and Renkawitz, R. (1992) EMBOJ. 11, 1015-1023 20 S~uer, F. and Jackle, H. (1993) Nature 364, 454-457 21 Saha, S., Brickman, J. M., Lehming, N. and Ptashne, M. (1993) Nature 363, 648-652 22 Hoey, T. et aL (1993) Cell 72,247-260 23 Inostroza, J. A. et al. (1992) Cell 70, 477-489 24 Hayes, J. J. and Wolffe, A. P. (1992) Bioessays 14, 597-603 25 Laurenson, P. and Rine, J. (1992) Microbiol. Rev. 56, 543-560 26 Seto, E., Shi, Y. and Shenk, T. (1991) Nature 354, 241-245 27 Wang, Z. Y., Qiu, Q. Q. and Deuel, T. F. (1993) J. Biol. Chem. 268, 9172-9175 28 Liu, X., Miller, C. W., Koeffler, P. H. and Berk, A. (1993) MoI. Cell BioL 13, 3291-3300 ALZHEIMER'SDISEASE(AD) was first described in a 51-year-old patient in 1907 by the Bavarian psychiatrist, Alois AlzheimerL The disease begins with loss of short-term memory; this is fol- lowed by progressive dementia, which in- volves memory loss, disorientation, and impairment of judgement and reason- ing. It is characterized pathologically by abnormally high levels of brain lesious ca!~cd ~,cnile p!~ques and neurofibrillary tangles (NFTs), ana by elevate~ .ua~- bets of deposits in the walls of cerebral blood vessels. In the later stages of AD, reduced levels of neurotransmitters, and extensive neuronal-cell and synap- tic loss occur 2,3. AD involves selective loss of acetyl- choline-releasing neurons ir=-~he cer- ebral cortex, and th~s has been hypoth- esized to be the cause of the early memory and learning loss. One approach to developing an AD therapy has been to search for drugs, for ex- ample cholinergic agonists, that reverse this deficit. However, few drugs signifi- cantly improve AD and those that do are marginally effective and produce unpleasant side-effects in some patients. There is an urgent need for a drug to combat the disease. F. Ashall and A. M. Goate are at the Department of Psychiatry, Washington University School of Medicine, 4940 Children's Place, St Louis, MO 63110, USA. I ' 111111111 Role of the [3.amyloidprecursor protein in Alzheimer's disease ,r I Frank Ashall and Alison M. Goate Deposition of ~amyloid peptide in the brain is an early event in Alzheimer's disease, the most common cause of dementia. Studies of the [l-amyloid precursor protein (APP), which gives rise to ~amyloid, are rapidly leading to a better understanding of the biochemical basis of the disease - a prerequisite for rational drug development. Plaques and NFTs are especially abundant in the hippocampus and sur- rounding areas - specialized regions of the cerebral cortex that are involved in memory and learning. Whilst plaques alid NFFs are numerous in AD brains, they do occur in aged normal people in lower numbers; they also occur in certain other diseases of the brain. About one person in every 20 over 65 and one in every five over 80 is afflicted with Alzheimer's disease, and all people with Down's syndrome who live into their late thirties and beyond develop a neuropathology that is indistinguish- able from non-Down's AD. Numerous advances in recent years have added substantially to our knowl- edge of the biochemical basis of AD, including the identification of the major protein constituents of plaques and NFFs 4-8, the discovery of point mu- tations that predispose individuals to familial AD and the related disease, her- editary cerebral haemorrhage with amyl- oidosis - Dutch type 0-1C~A-D)~-u, and elucidation of biochemical pathways that are central to plaque formation ~2-~6. Plaquesand tangles Senile plaques are extracellular deposits of a heterogeneous substance 42 © 1994,Elsevier Science Publishers, (UK) 0968-0004/94]$07.00
Transcript of Role of the β-amyloid precursor protein in Alzheimer's disease

REVIEWS pressing and activating domains have also been mapped for p53 (Ref. 28). Thus it appears that some proteins con- tain both activating and repressing potential, and the predominant effect depends on promoter context, in a related phenomenon, it has been observed that Kr can act either as a repressor or as an activator depending on its concentration 2°, which is con- sistent with genetic evidence that Kr is a positive and negative regulator in a hierarchy of genes involved in Drosophda embryonic development.
Repression of various kinds is now well embedded in our understanding of transcriptional regulation. However, there are still gaps in our knowledge, notably concerning the mechanism of action of the active transcriptional repressors. Future work to unravel this process will undoubtedly go hand in hand with a further understanding of transcriptional activation and other
events surrounding transcription initiation.
Acknowledgements I would like to thank H. C. Hurst, C. A.
Austin and others for useful discussion and comments on this manuscript.
References 1 Roede,', R. G. (1991) Trends Biochem. Sci. 16,
402-408 2 Workman, J. L. and Buchrnan, A. R. (1993)
Trends Biochern. Sci. 18, 90-95 3 Morse, R. H. (1992) Trends Biochem. Sci. 17,
23-26 4 Kageyama, R. and Pastan, I. (1989) Cell 59,
815-825 5 SchOle, R. et al. (1990) Cell 61, 497-504 6 Liu, Y., Yang, N. and Teng, C. T. (1993) Mol. Cell
Biol. 13, 1836-1846 7 Benezra, R. et al. (1990) Cell 61, 49-59 8 Ron, D. and Habener, J. F. (1992) Genes Dev.
6, 439-453 9 Deng, T. and Karin, M. (1993) Genes Dev. 7,
479-490 10 Bengal, E. et al. (1992) Cell 68, 507-519 11 Yang Yen, H. F. et al. (1990) Cell 62,1205-1215 12 Licht, J. b., Grossel, M. J., Fi~e, J. and
TIBS 19 - JANUARY 1994
Hansen, U. M. (1990) Nature 346, 76-79 13 Han, K. and Manley, J. L. (1993) Genes Dev.
7,491-503 14 Jaynes, J. B and O'Farreh, P. H. (1991) EMBO J.
10, 1427-1433 15 Han, K. and Manley, J. L. (1993) EMBO J. 12,
2723-2733 16 Shi, ¥., Seto, E., Chang, L-S. and Shenk, T.
(1991) Cell 67, 377-388 17 Madden, S. L., Cook, D. M. and Rauscher,
F. J., III (1993) Oncogene 8, 1713-1720 18 Cowell, I. G., Skinner, A. and Hurst, H. C. (1992)
/VIOL Cell Biol. 12, 3070-3077 19 Baniahmad, A., K6hne, A. C. and Renkawitz, R.
(1992) EMBO J. 11, 1015-1023 20 S~uer, F. and Jackle, H. (1993) Nature 364,
454-457 21 Saha, S., Brickman, J. M., Lehming, N. and
Ptashne, M. (1993) Nature 363, 648-652 22 Hoey, T. et aL (1993) Cell 72,247-260 23 Inostroza, J. A. et al. (1992) Cell 70, 477-489 24 Hayes, J. J. and Wolffe, A. P. (1992) Bioessays
14, 597-603 25 Laurenson, P. and Rine, J. (1992) Microbiol.
Rev. 56, 543-560 26 Seto, E., Shi, Y. and Shenk, T. (1991) Nature
354, 241-245 27 Wang, Z. Y., Qiu, Q. Q. and Deuel, T. F. (1993)
J. Biol. Chem. 268, 9172-9175 28 Liu, X., Miller, C. W., Koeffler, P. H. and Berk, A.
(1993) MoI. Cell BioL 13, 3291-3300
ALZHEIMER'S DISEASE (AD) was first described in a 51-year-old patient in 1907 by the Bavarian psychiatrist, Alois AlzheimerL The disease begins with loss of short-term memory; this is fol- lowed by progressive dementia, which in- volves memory loss, disorientation, and impairment of judgement and reason- ing. It is characterized pathologically by abnormally high levels of brain lesious ca!~cd ~,cnile p!~ques and neurofibrillary tangles (NFTs), ana by elevate~ .ua~- bets of deposits in the walls of cerebral blood vessels. In the later stages of AD, reduced levels of neurotransmitters, and extensive neuronal-cell and synap- tic loss occur 2,3.
AD involves selective loss of acetyl- choline-releasing neurons ir=-~he cer- ebral cortex, and th~s has been hypoth- esized to be the cause of the early memory and learning loss. One approach to developing an AD therapy has been to search for drugs, for ex- ample cholinergic agonists, that reverse this deficit. However, few drugs signifi- cantly improve AD and those that do are marginally effective and produce unpleasant side-effects in some patients. There is an urgent need for a drug to combat the disease.
F. Ashall and A. M. Goate are at the Department of Psychiatry, Washington University School of Medicine, 4940 Children's Place, St Louis, MO 63110, USA.
I '111111111
Role of the [3.amyloid precursor protein in Alzheimer's disease
,r I
Frank Ashall and Alison M. Goate
Deposition of ~amyloid peptide in the brain is an early event in Alzheimer's disease, the most common cause of dementia. Studies of the [l-amyloid precursor protein (APP), which gives rise to ~amyloid, are rapidly leading to a better understanding of the biochemical basis of the disease - a prerequisite for rational drug development.
Plaques and NFTs are especially abundant in the hippocampus and sur- rounding areas - specialized regions of the cerebral cortex that are involved in memory and learning. Whilst plaques alid NFFs are numerous in AD brains, they do occur in aged normal people in lower numbers; they also occur in certain other diseases of the brain.
About one person in every 20 over 65 and one in every five over 80 is afflicted with Alzheimer's disease, and all people with Down's syndrome who live into their late thirties and beyond develop a neuropathology that is indistinguish- able from non-Down's AD.
Numerous advances in recent years have added substantially to our knowl- edge of the biochemical basis of AD, including the identification of the major protein constituents of plaques and NFFs 4-8, the discovery of point mu- tations that predispose individuals to familial AD and the related disease, her- editary cerebral haemorrhage with amyl- oidosis - Dutch type 0-1C~A-D) ~-u, and elucidation of biochemical pathways that are central to plaque formation ~2-~6.
Plaques and tangles Senile plaques are extracellular
deposits of a heterogeneous substance
42 © 1994, Elsevier Science Publishers, (UK) 0968-0004/94]$07.00

TIBS 19 - JANUARY 1994 REVIEWS called 'amyloid'. They also contain dystrophic neurites (neural cell pro- cesses) and glial cells. Although plaques are particularly abundant in the cerebral cortical parenchyma of AD victims, other extracellular amyloid deposits occur in the walls of cerebral and meningeal blood vessels.
In 1984, Glenner and Wong 4,s isolated and partmlly sequenced the major protein constituent of cerebrovascular amyloid deposits, a 4kDa peptide. Subsequently, Masters et ai. 6 showed that this 39 to 43 amino acid molecule, called the ~-amyloid peptide, was also the principal constituent of senile plaques. Other proteins, including ubiquitin, apolipoprotein E (apoE), interleukin-1, lysosomal hydrolases and cxfantichymotrypsin, are also found in plaques.
NVI's occur within neurons and con- sist of a highly insoluble material. Ultrastructurally, they appear as paired helical filaments (PHFs). The micro- tubule-associated protein tan is a major constituent of PHFs. However, PHF tau differs in its biochemical and biological properties from normal tau (see the review by Mandelkow and Mandelkow in the December 1993 issue of TIBS). In particular, PHF tau is hyperphosphoryl- ated compared with normal tau ~7.
Studies of Down's syndrome patients indicate that NFTs develop after senile plaques have formed. Indeed, [l-amyloid deposition is the earliest sign of AD-like pathology in Down's patients; NFrs and neuro, loss occur later on, as the dis- ease progresses. Antibodies to ~pep- tide stain diffuse amyloid deposits in the brains of AD and Down's syndrome patients ~9, in addition to the more clearly developed senile plaques. These diffuse deposits may be the precursors of senile plaques.
The amylold precursor protein When the first full-length cDNA
encoding the ~-amyloid peptide was cloned and sequenced 2° it was found to encode a protein that is much larger than the 4 kDa ~-peptide. Hence, the [l- peptide is derived by proteolysis from this larger protein, called the amyloid precursor protein (APP).
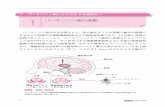
APP is a large, multidomain protein containing a single membrane-spanning region (Fig. I), with the amino terminus extending into the extracellular environ- ment, and a short region at the carboxyl terminus projecting into the cytoplasm. Of particular interest is the presence of an extracellular domain that shows
'Ox' antigen Cytoplasmic Protease Signal Negatively inhibitor ~ pepiide Cyste~e-dch charged~ Transmembrane
/ ~ - ~ ~-amyloid pep!ide regmn
O - - I~. I'~P',,. 0303 ',- ¢,O¢D ~ ¢OtlD P". KM-DAEFRHDSGYEVHHQKLVF FAEDVGSNKGA I I GLMVGGVVIAT -VIVI TLVMI,- KKK
N"~L G Q F I
Rgure 1 Domain structure of APP, showing the ~amyloid peptide region. The ~-peptide region con- tains part of the membrane-spanning domain of APP and part of the extracellular domain. Mutations that cause AD are indicated (APP-770 terminology). A double mutation of Lys670-Met671 to Asp-Leu immediately amino-terminal to the ~-peptide region occurs in a Swedish AD family; familial AD is also caused by mutation of Val717 to Phe, lie or Gly at a position two amino acid residues down from the carboxy-terminal side of the ~peptide sequence. Mutation of Glu to Gin at position 22 of the ~-peptide results in Dutch-type amyl- oidosis (HCHWA-D). Anothe~ mutation, Ala to Gly at position 21 of the ~-peptide (692 of APP), causes AD in some patients and HCHWA-D-like disease in others.
homology to the Kunitz class of pro- tease inhibitors (KPI). Indeed, KPi- containing forms of APP inhibit serine proteases known to be inactivated by Kunitz inhibitorsZL
About two thirds of the [5-amyloid peptide portion of APP is extracellular, whereas the remaining third o! the ~peptide is integrated into the membrane (Fig. 1).
Expression of APP is complex. The primary RNA transcript of APP under- goes differential splicing to generate mRNAs coding for at least five forms of APP: APP-563, APP-695, APP-714, APP- 751 and APP-770 Three of these forms, APP-563, APP-751 and APP-770, contain the KP! domain, whereas APP-695 and APP-714 do not. APP is produced in a wide variety of cell types. The relative levels of the different APP isoforms vary according to cell type and may even differ among cells within a given tissue. Whilst APP-695 mRNA is particu- larly abundant in brain, there is evi- dence that APP-751, which contains the KPI domain, is also produced in rela- tively large amounts in human brain regions involved in senile plaque formation 22.
APP is also post-translationally modi- fied: the molecule undergoes O- and
N-glycosylation, phosphorylation, and sulphation on tyrosine resid,es 23. Activators of protein kinase C, including phorbol esters, acetylcholine and inter- leukin-1, and inhibitors of phospha- tases, increase the processing and secretion of APP (Refs 24, 25). APP is also regulated compartmentally within the cell: surface APP can be secreted or internalized into the endosomal-lyso- somal system. To add to the complexity, APP is also processed by multiple pro- teolytic pathways.
The biological function of APP is not clear. The secreted forms that contain the KP! domain might be involved in regulation of protease activity at the cell surface or in the extracellular en- vironment. Indeed, the secreted form of APP-751 is identical to nexin-2, a pro- tease inhibitor that is thought to be involved in regulation of extracellular serine proteases 26. APP may have a role in tissue repair in the brain; it is se- creted by blood platelets and shares amino-terminal amino acid sequence, immunological and functional identity to platelet coagulation Factor Xi a- inhibitor, which is involved in blood clotting2!
There is also evidence that APP pro- motes cell-cell and cell--extracellular-
43

REVIEWS TIBS 19 - JANUARY 1994
N ~-amyloid ~ N deposits
APP N
Extracellular ~ ~ @ environment
Membrane ~ C ~CC "C , / Cytoplasm
•
N
13-amyloid ' deposits
Endosomes/lysosomes
Rgure 2 Proteolytic processing of APP includes pathways that lead to secretion of APP fragments, as well as intracellular routes. I~amyloid deposition is likely to be particularly dependent on cleavage in the car- boxy-terminal region of APE where the ~peptide sequence (black rectangle) is located. Crucial pro- teases include the (x-, ~- and 7-secretases ((x, 13, 7) altho.Jgh endosomal/lysosomal APP-processing proteases might also be involved in ~-amyloid peptide deposition. Question marks refer to steps that have not been unequivocally demonstrated.
matrix adhesion. Indeed, APP has a high affinity for the basement mem- brdne form of heparan sulphate 28. APP also promotes survival and growth of neurons in vitro.
APP mutations in familial AD Most cases of AD occur in elderly
people, and these are probably due mainly to a combination of environmen- tal and genetic factors. However, some rare instances of familial, genetic forms of AD exist that are due to a phenotypi- cally dominant autosomai mutation.
What has emerged from investi- gations of familial AD is that mutations that lead to AD can occur in different ge~cs ~z~ differe,~t !amilies, and that dif- ferent mutations can occur at the same genetic locus in differept families. The gene encoding APP was mapped to chromosome 21, and this suggested that the AD seen in Down's syndrome
was due to overexpression of the APP gene. This idea was further supported by the discovery that some familial forms of AD were linked to chromo- some 21.
In 1990, Levy et ai. showed that patients with Dutch-type amyloidosis (HCHWA-D) have a point mutation within the ~-peptide sequence of APP at amino acid position 22, causing a glutamate residue to be substituted for a glutamine residue ~°. Patients with HCHWA-D do not develop AD, but they do have diffuse ~-amyloid deposits within the cerebral cortex, and cer- ebrovascular ~amyloid deposits. As a result of the vascular deposits, these patients are prone to brain haemorrhage. This discovery was the first to show that a specific defect in the APP gene can cause ~-amyloid to be deposited.
Persuasive evidence that APP can have a pr mary role in AD came from
the discovery by Goate et al. 9 in 1991 that a point mutation in the APP gene (position 717, near the car- boxy-terminal amino acid of the J3-peptide) co-segre- gates with disease in a t least one early-onset AD family. Subsequently, APP mutations at the same and other codons within the APP gene were found in other AD families. None of these mutations occurs within the ~.amyloid pep- tide region of the APP molecule, but they do oc- cur very close to the ends of the ~-peptide sequence ~ig. 1).
Point mutations in the APP gene are not always the cause of familial AD. Recent research has shown that a matation in a gene on chromosome 14 can also cause familial AD 29. Chromo- some 14 mutation(s) ac- counts for the majority of cases of familial early-onset AD. The identity of the gene (or genes) involved is not yet clear, and whether or not it encodes a product that is directly involved in APP metaholi~m has yet to be determined. A gene on chromosome 19 also ap- pears to be linked to some forms of late-onset AD. This may be the apo£ gene,
which maps to the region found to be genetically linked to AD. apoE binds strongly to the ~peptide and is a con- stituent of plaques, and the apoE type 4 allele is three times more likely to occur in individuals with late-onset AD than in controls a°. Mutations in mitochondrial NADH dehydrogenase might also be a risk factor for AD al.
The finding that mutations in APP are primary genetic defects in some familial cases of AD has led to a surge in the biochemical analysis of APE particularly in its proteolytic processing, the under- standing of which is central to deter- mining how amy!did deposition is in- itiated. This work is likely to provide important information relating to sporadic forms of AD and the AD-like pathology seen in Down's syndrome, as well as familial forms of the disease, since brain pathology is very similar in all of these conditions.
44

TIBS 19 - JANUARY 1994 REVIEWS Proteolytic processing of APP
APP is post-translationally processed by numerous proteolytic pathways 12-16, which lead to secretion of various frag- ments of APP or intracellular fragmen- tation and degradation (Fig. 2).
There are at least three major spe- cific sites of protease cleavage in and around the ~amyloid sequence. One of these is the (z-secretase site, which occurs between Lysl6 and Leul7 of the 39 to 43 amino acid sequence of the lS-amyloid peptide. Hydrolysis of the peptide bond at this site precludes any 15-peptide from being formed, since it involves cleavage within the ~peptide sequence *z.
Another protease, the ~secretase, cleaves between Met671 and Asp672 (APP-770 designation), generating the free amino terminus of ~-amyloid. Hydrolysis of this Met-Asp peptide bond can occur independently of cleav- age at the carboxy-terminal end of the ~peptide, to generate an extracellular APP fragment containing none of the ~-peptide region but all of the region of APP amino-terminal to the lS-peptide, as well as a membrane-associated, poten- tially amy!oidogenic, carboxy-terminal fragment 32.
At the carboxy-terminal end of the 15- peptide region (the T-secretase site) the precise peptide bond that is cleaved is not known - there may even be several proteolytic sites - but it occurs some- where between amino acid 39 and the end of the lS-peptide sequence. The 7" scc,~tase cleavage slte is integrated into the cell membrane. Cleavage of APP at this site can occur indepen- denti, y o[ ¢!.~:wage at the oz. and lS-se- cretase sites, to generate an extra- cellular amino-terminal fragment of APP cont,~Ining a complete ~amyloid pep tide sequence 33. This fragment is, there- fore, potentially amyloidogenic. Hy- drolysis at both of the ~ and T-secretase bonds should generate the ~peptide.
Normal cells produce an extracellular soluble form of ~amyloid peptide, which can also be detected in normal cerebrospinal fluid and blood ~5,~6. The origin and function of this peptide is not clear; |~ is also not known if this sol- uble ~myloid can give rise to insol- uble lS-amyloid deposits in vivo. Cells transfected with an APP cDNA contain- ing the double .m~:tation Lys670-Met671 to Asn670-Leu671 at the amino-terminal site of the ~peptide (Fig. I) secrete five- to etghtiuld more soluble ~-peptide than control ceils 34,35. This suggests that the mutation may enhance l~mnyl-
old deposition, perhaps by providing a better substrate for lS-secretase and increasing extracellular levels of soluble ~-amyloid. However, transfectants expressing a Val to lie mutation at pos- ition 717 do not produce increased amounts of soluble ~peptide.
Cells transfected with the APP gene containing the HCHWA-D Glu693 to Gin693 mutation within the 15-peptide also do not secrete more ~-peptide than control cells. One possible explanation for the enhanced deposition of 15-amyl- old in HCHWA-D patients is that the physicochemical properties of the peptide are altered such that the pep- tide is more likely to form insoluble aggregates; this makes sense because the mutation replaces a negatively charged amino acid sidechain (Glu) with a neutral residue (Gin) within the ~-peptide. A synthetic lS-peptide con- taining the HCHWA-D mutation forms insoluble fibrils in vilro more =eadily than synthetic wild-type 15-peptide.
Familial AD may turn out to have multiple primary causes that affect a common final pathway leading to 15-amyloid deposition. Some mutations, such as Lys670-Met671 to Asn670- Leu671, might increase the levels of extracellular 15-peptide and result in deposition of 15-amyloid. Others, like the HCHWA-D mutation, might render the peptide more prone to aggregation without increasing its rate of pro- duction. In Down's syndrome, the APP gene is presumably normal but over- expressed, and this might lead to increased production of lS-peptide. Other mutations, possibly including those at position 717, might affect intra- cellular processing or trafficking of APP.
In addition to the possibility that sol- uble ~-peptide formation occur.~ at or near the cell surface by cleavage of APP at both the [5- and T-secretase cleavage sites, soluble 15-peptide might also be produced intracellularly and subsequently released into the cell's environment. Another possibility is that soluble ~amyloid is formed extracellu- larly from amino-terminal fragments ~,,, APP containing the complete 15-amylo~d region. Of course, more than one of these pathways might contribute to soluble ~amyloid formation.
Intracellularly, APP and its fragments are degraded in the endosome--lyso- some system 13,14. The 10 kDa carboxy- terminal membrane fragment formed by a-secretase, and pi, obably those formed also by the 15- and y-secretases, are taken into the lysosomes for degra-
dation. Intact APP can also be detected in lysosomes. Perhaps more relevant to ~amyloid formation, fragments of APP containing the complete ~-peptide sequence are found in endosomes/lyso- somes, suggesting that this might be a route by which l~-peptide can be gener- ated. However, ~peptide alone, without other sequences attached to it, has not been detected in lysosomes.
Carboxy-terminal fragments of APP that contain the whole ~peptide sequence in addition to regions car- boxy-terminal to the lS-peptide might also form insoluble aggregates, and these might be hydrolysed to shorter, ~- amyloid-sized fragments by carboxy- peptidases.
Ch3racterization of the o~-, 15- and 7- secretases and other proteases involved in APP processing will be an important component of research aimed at rational design of anti-AD drugs. Protease inhibitors aimed at one or more of these enzymes might retard amyloid deposition and slow the dis- ease process. Other drugs might be tar- geted to the trafficking of APP or the deposition of 15-amyloid. Transgenic models of AD are also an important i-esearch goal: they will provide much needed animal models for tes' '=lg drugs and investigating the appearance of AD pathology in vivo.
References I Alzheimer, A. (1907) All. Z. Psychia'.r. 64,
146-148 2 Katzman, R. (1986) New EngL I. Meal. 314,
964-973 3 Holtzman, D. M. and Mobley, W. C. (1991j
Trends Blochem. ScL 16, 140-144 4 Glenner, G. G. and Wong, C. W. (1984)
Biochem. Biophys. Res. Commun. !22, 11~1-1135
5 GPenner, G. G. and Wong, C. W. (1984) Biochern. P, iophys. Res. Cemmun. 120, 885-890
6 Masters, C. L. et al. (1985) Proc. Nat/Acad. ScL USA 82, 4245-4249
7 Lee, V. M-Y. et al. (1991) Science 251, 675-678
8 Rament, S. et aL (1989) J. Neuro/. Sci. 92, 133-141
9 Goate, A. M. et aL (1991) Nature 349, 704-706
10 Levy, E. eta/. (1990) Science 248, 1124-1126 . 11 Murrell, J. et al. (1991) Science 254, 97-99 12 Esch, F. S. et al. (1990) Science 248,
1122-1124 . 13 Haas, C. et al. (1992) Nature 357, 500-502 14 Estus, S. et al. (1992) Science 255, 726-728 15 Haas, C. et al. (1992) Nature 359, 322-325 16 Shoji, M. st aL (!o92) Science 258, 126-129 17 Grunde-iqbal, I. et al. (1986)Proc. Natl Acad.
ScL USA 83, 4913-4917 18 Kosik, K. S. (1991) Trcnds Neurosci. 14,
218-219 19 Yamaguchi, H. et aL (1988) Acta NeuropathoL
(Berlin) 77, 11.~-119 20 Kang, J. et ~1. (1987) Nature 325, 733-736
45

COMPUTER CORNER 21 Kitaguchi, N. et al. (1988) Nature 33!,
5~.0 532 22 Anderson, J. P. et aL (1989) EMBO J. 8,
3627-3632 23 Weidemann, A. et aL (1989) Cell 57, 115-126 24 Buxbaum, J. et al. (1992) Proc. Natl Acad. Sci.
USA 89, 1.007-~r~-10078 25 Buxbaum, J. etal. (1990) Proc. Natl Acad. Sci.
USA 87, 6003-6006
26 Oltersdorf, T. et al. (1989) Nature 341, 144-147
27 Smith~ R. P., Higuchi, D. A. and Broze, G. J. (1990) Science 248, 1126-1128
28 Narindrasorasak, S. et aL (1991) J. Biol. Chem. 266, 12878-12883
29 Schellenberg, G. D. et al. (1992) Science 258, 668-671
3 0 Strittmatter, W. J. et al. (!993) Proc. Natl Acad.
TIBS 1 9 - J A N U A R Y 1 9 9 4
ScL USA 90, 1977-1981 31 Lin, F-H. et aL (1992) Biochem. Biophys. Res.
Cornmun. 182, 238-246 32 Seubert, P. et aL (1993) Nature 361, 260-263 33 Anderson, J. P. et aL (1992) J. Neurochem.
59, 2328-2331 34 Citron, M, ot al. (1992) Nature 360, 672-674 35 Cai, X-D., Golde, T. E. and Younkin, S. G. (1993)
Scienca 259, 514-516
Besides the standard ligation and trans- formation questions that appear routinely, many ideas resurface when new people join the newsgroup. One recurring theme is the pros and cons of using prepackaged reagent kits sold by commercial companies.
Kits are designed to be a quick and easy alternative for anything from mini- prep plasmid DNA extractions to removal of mineral oil from PCR tubes. There are now kits available for just about every routine procedure used in molecular biology laboratories.
The battle over kits i.~ ongoing on methds-reagnts. Mention the use of a kit and you are bound to start trouble between the 'kit' and 'no-kit' advocates. This inevitably spurs 'Kit Wars'. Sides are qui,~kly drawn and opinions start flying, usually falling just short of name calling. Recently, a huge fight erupted over kits, which prompted me to search [or past articles on the topic. By using Gopher (see TIBS 18, 107-108), ! found more than 150 archived articles posted to bionet.molbio.methds-reagnts related to the use of kits. Of ',hese, nearly half are devoted to the discussion of 'kits' versus "no kits'.
The kit advocates are in search of speed. They prefer the fastest and safest route to the end of their exper- iment. These researchers are cons: antly looking for the best kit to get their experiments to work. For many, kits are
In the USA BIOSCI is supported by a grant from the National Science Foundation with important contributions from the Department of Energy, the National Institute of General Medical Sciences and the National Center for Human Genome Research. In the UK, BIOSCI is supported at the Government-funded Science and Engineering Research Council's Daresbury Laboratory. For further information on BIOSCt, please contact one of the addresses below. For more details,
b see also "fIBS 18, pp. 446-448.
i Address Serving ! i biosci The Americas I C~net.bio.net and Pacific Rim . biosci t
@o~resbury.ac.uk Eurupe, Africa and Central A.~ia
46
i
Methods and reagents
Kit Wars
a highly useful way of completing diffi- cult procedures that would otherwise cost large amounts of time iFi trying to locate and collect all the necessary components. Kits are useful for diag- nostic labs that do a large number oi routine procedures in a single day, but they can also be a tremendous help in a one-off experiment. In addition, use of kits can avoid some dangerous but necessary steps involved in making your own reagents.
Unfortunately, the attitude of some is 'if the kit works, use it'. These people are thus accused by the 'no-kit' advo- cates of being 'kit-robots', having little or no knowledge of the biochemical processes involved in the procedure. Kit-robots frequently get burned when the kit does not work in their h~.qds, and generaUy do not question why it doesn't work, but rather discard it for a different version.
Altt~ough the cost of kits is relatively high, some kit advocates claim that they are sometimes cheaper than the materials needed to make up all the components. For example, one netter wrote that both the fmoF M DNA sequencing system and Magic TM maxi- prep DNA purification system from Promega are much cheaper than homc~ made versions based on the cost ol ingredients alone.
The no-kit advocates are those who believe in making almost everything used in their experiments from scratch. This has several advantages, including the fact that the user knows the exact components of the cocktail, their con- centrations and the reasons for includ- ing them. It is also more flexible because the system can be customized to suit a particular need, and any prob- lems can be in,vestigated.
The major drawbacks of kits include the expense, wasted leftover materials and proprietary or hidden components. When companies hide the true compo- nents of their kits under the term 'pro- prietary reagent', many researchers fee' it is their right to know what the com- ponents are, and some people who object strongly to this practice refuse to buy the kit, Perhaps one of the main reasons why the no-kit advocates do not like these mystery components is the lack of flexibility this bestows on the prescribed method, which requires knowledge of all the components to make subtle adjustments. Hidden com- ponents are therefore sometimes seen as an impediment to the advancement of scientific knowledge. For example, both Promega and Qiagen Inc., as well as many other companies, sell excellent products for the binding and clean-up of PCR products. However, each brand of spin column has a proprietary resin for binding DNA, which means that researchers wanting to use this tech- nique do not have the option of making up the recipe in their own laboratories.
Perhaps the most outspoken and provocative no-kit person in the newsgroup is Jim Grail~,a~ (]graham @bronze.ucs.indiana.edu), who has spent much ot his time arguing against the use of kits and searching for alterna- tive ways to make his own reagents. Originally prompted by postings with less exchange of technical information, he feels compelled to keep himself from succumbing to pro.made kits because 'current advances in molecular biology techniques are no longer available to the general science public via the litera- ture, but are being sold to us through commercial companies at inflated prices'.
© I994, Elsevier Science Publishers. (UK) 0968--0004/94/$07.00