
Research Article Collagen V Is a Potential Substrate for ...
9
Research Article Collagen V Is a Potential Substrate for Clostridial Collagenase G in Pancreatic Islet Isolation Hiroki Shima, 1,2 Akiko Inagaki, 3 Takehiro Imura, 3,4 Youhei Yamagata, 5 Kimiko Watanabe, 4 Kazuhiko Igarashi, 1,2,6 Masafumi Goto, 3 and Kazutaka Murayama 7 1 Department of Biochemistry, Tohoku University Graduate School of Medicine, Sendai 980-8575, Japan 2 AMED-CREST, Japan Agency for Medical Research and Development, Tokyo 100-0004, Japan 3 Division of Transplantation and Regenerative Medicine, Tohoku University School of Medicine, Sendai 980-8575, Japan 4 New Industry Creation Hatchery Center, Tohoku University, Sendai 980-8579, Japan 5 Graduate School of Agricultural Science, Tokyo University of Agriculture and Technology, Fuchu 183-8509, Japan 6 Center for Regulatory Epigenome and Diseases, Tohoku University Graduate School of Medicine, Sendai 980-8575, Japan 7 Graduate School of Biomedical Engineering, Tohoku University, Sendai 980-8575, Japan Correspondence should be addressed to Kazutaka Murayama; [email protected] Received 19 February 2016; Accepted 27 March 2016 Academic Editor: Pedro M. Geraldes Copyright © 2016 Hiroki Shima et al. is is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. e clostridial collagenases, H and G, play key roles in pancreatic islet isolation. Collagenases digest the peptide bond between Yaa and the subsequent Gly in Gly-Xaa-Yaa repeats. To fully understand the pancreatic islet isolation process, identification of the collagenase substrates in the tissue is very important. Although collagen types I and III were reported as possible substrates for collagenase H, the substrate for collagenase G remains unknown. In this study, collagen type V was focused upon as the target for collagenases. In vitro digestion experiments for collagen type V were performed and analyzed by SDS-PAGE and mass spectrometry. Porcine pancreatic tissues were digested in vitro under three conditions and observed during digestion. e results revealed that collagen type V was only digested by collagenase G and that the digestion was initiated from the N-terminal part. Tissue degradation during porcine islet isolation was only observed in the presence of both collagenases H and G. ese findings suggest that collagen type V is one of the substrates for collagenase G. e enzymatic activity of collagenase G appears to be more important for pancreatic islet isolation in large mammals such as pigs and humans. 1. Introduction e clostridial collagenases, collagenase H (ColH) and col- lagenase G (ColG), are used for pancreatic islet isolation in combination with neutral proteases [1]. e collagenases possess a multimodular domain structure [2], and both collagenases have five domains (Figure 1). Activator and peptidase domains are involved in the collagenolytic activity [3] and are commonly contained in ColH and ColG. e active center comprises an HEXXH zinc-binding motif [4]. Polycystic kidney disease-like domains (PKDs) are located at the middle part of the enzyme, with two PKDs in ColH and one PKD in ColG. At the C-terminal end, both collagenases possess a collagenase binding domain (CBD), which is responsible for collagen triple helix binding [5, 6]. ColH has one CBD and ColG has two CBDs located tandemly. Molecular structures of clostridial collagenases have been revealed for the collagenase module of ColG [7], peptidase domain of ColH [8] (JBC, 288, 20184) and ColT [8], PKD of ColH [9] and ColG [7–10], and CBD of ColH [11] and ColG [11, 12]. Regarding the regulation of enzymatic activity, calcium ions play a critical role. e calcium ion- binding sites are contained within the PKD and CBD, and calcium ion binding enhances the stability of the protein molecule and maintains the domain conformations [13, 14]. e collagenases generally digest the bond between Yaa and Hindawi Publishing Corporation Journal of Diabetes Research Volume 2016, Article ID 4396756, 8 pages http://dx.doi.org/10.1155/2016/4396756
Transcript of Research Article Collagen V Is a Potential Substrate for ...

Research ArticleCollagen V Is a Potential Substrate for ClostridialCollagenase G in Pancreatic Islet Isolation
Hiroki Shima12 Akiko Inagaki3 Takehiro Imura34 Youhei Yamagata5 Kimiko Watanabe4
Kazuhiko Igarashi126 Masafumi Goto3 and Kazutaka Murayama7
1Department of Biochemistry Tohoku University Graduate School of Medicine Sendai 980-8575 Japan2AMED-CREST Japan Agency for Medical Research and Development Tokyo 100-0004 Japan3Division of Transplantation and Regenerative Medicine Tohoku University School of Medicine Sendai 980-8575 Japan4New Industry Creation Hatchery Center Tohoku University Sendai 980-8579 Japan5Graduate School of Agricultural Science Tokyo University of Agriculture and Technology Fuchu 183-8509 Japan6Center for Regulatory Epigenome and Diseases Tohoku University Graduate School of Medicine Sendai 980-8575 Japan7Graduate School of Biomedical Engineering Tohoku University Sendai 980-8575 Japan
Correspondence should be addressed to Kazutaka Murayama kmurabmetohokuacjp
Received 19 February 2016 Accepted 27 March 2016
Academic Editor Pedro M Geraldes
Copyright copy 2016 Hiroki Shima et al This is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
The clostridial collagenases H and G play key roles in pancreatic islet isolation Collagenases digest the peptide bond betweenYaa and the subsequent Gly in Gly-Xaa-Yaa repeats To fully understand the pancreatic islet isolation process identification of thecollagenase substrates in the tissue is very important Although collagen types I and III were reported as possible substrates forcollagenase H the substrate for collagenase G remains unknown In this study collagen type V was focused upon as the target forcollagenases In vitro digestion experiments for collagen typeVwere performed and analyzed by SDS-PAGE andmass spectrometryPorcine pancreatic tissues were digested in vitro under three conditions and observed during digestion The results revealed thatcollagen typeVwas only digested by collagenaseG and that the digestionwas initiated from theN-terminal part Tissue degradationduring porcine islet isolation was only observed in the presence of both collagenases H and GThese findings suggest that collagentypeV is one of the substrates for collagenaseGThe enzymatic activity of collagenaseG appears to bemore important for pancreaticislet isolation in large mammals such as pigs and humans
1 Introduction
The clostridial collagenases collagenase H (ColH) and col-lagenase G (ColG) are used for pancreatic islet isolationin combination with neutral proteases [1] The collagenasespossess a multimodular domain structure [2] and bothcollagenases have five domains (Figure 1) Activator andpeptidase domains are involved in the collagenolytic activity[3] and are commonly contained in ColH and ColG Theactive center comprises an HEXXH zinc-binding motif [4]Polycystic kidney disease-like domains (PKDs) are locatedat the middle part of the enzyme with two PKDs inColH and one PKD in ColG At the C-terminal end both
collagenases possess a collagenase binding domain (CBD)which is responsible for collagen triple helix binding [56] ColH has one CBD and ColG has two CBDs locatedtandemly Molecular structures of clostridial collagenaseshave been revealed for the collagenase module of ColG [7]peptidase domain of ColH [8] (JBC 288 20184) and ColT[8] PKD of ColH [9] and ColG [7ndash10] and CBD of ColH[11] and ColG [11 12] Regarding the regulation of enzymaticactivity calcium ions play a critical role The calcium ion-binding sites are contained within the PKD and CBD andcalcium ion binding enhances the stability of the proteinmolecule and maintains the domain conformations [13 14]The collagenases generally digest the bond between Yaa and
Hindawi Publishing CorporationJournal of Diabetes ResearchVolume 2016 Article ID 4396756 8 pageshttpdxdoiorg10115520164396756
2 Journal of Diabetes Research
ColH
ColG
PepAct
CBDCBD
CBD
PKD
PKD PKD
Act Pep
N
N C
C
Figure 1 Schematic diagram of the domain architectures ofColG and ColH Act activator domain Pep peptidase domainPKD polycystic kidney disease-like domain CBD collagen-bindingdomain The collagenase module is composed of two functionaldomains an activator domain and a peptidase domain
Gly in the Gly-Xaa-Yaa(-Gly-) sequence repeats of fibrillarcollagens [15 16] Degradation of collagen type I (Col-I) iswell-known for collagenolytic activity of ColH and ColGColH also plays a crucial role during rat pancreatic isletisolation [17] In vitro pancreatic tissue digestion experimentsand mass spectrometric analyses revealed that Col-I andcollagen type III (Col-III) are substrates for ColH [17]Although the coexistence of ColG can increase the yield ofisolated islets it has been difficult to determine the substratespecificity for ColG in rat experiments Meanwhile bothColH and ColG are used for pancreatic tissue dissociation inlarge mammals such as pigs Identification of the substratefor ColG in pancreatic tissue is one of the key factors towardunderstanding the pancreatic islet isolation process
The fibrillar collagens are targets for many collagenases[2] The commonly found fibrillar collagens are types I IIIII and V [18] A previous study showed that Col-I collagentype II (Col-II) and Col-III were fully digested by ColG [17]while collagen type V (Col-V) was relatively less digested(manuscript in preparation) Col-V is a fibrillar collagenwith low abundance Several isoforms are found for Col-Vdepending on the chain components [19]Themost commonisoform is 1205721(V)
21205722(V) However other isoforms such as
1205721(V)3and 1205721(V)1205722(V)1205723(V) have also been reported [20]
Col-V exists in tissues colocalized with Col-I and appears toregulate the assembly of heterotypic fibers containing Col-Iand Col-V [20ndash22]
In the present study Col-V digestion byColGwas focusedupon to investigate the substrate specificity of ColG Thedigestions were analyzed by SDS-PAGE and mass spec-trometry The differences in Col-V digestion by ColH andColG were evaluated The results revealed that Col-V is aputative substrate for ColG in pancreatic islet isolation inlarge mammals
2 Materials and Methods
21 Collagenases and Porcine Pancreatic Tissue Recombinantenzymes of ColG and ColH which were kindly providedby Meiji Seika Pharma Co Ltd were produced by anEscherichia coli protein expression systemwith ColG or ColHgenes isolated from Clostridium histolyticum as describedpreviously [17]
Porcine pancreatic tissue samples were obtained from2ndash4-year-old adult retired sows weighing 200ndash300 kg Allanimal experiments were performed according to the guide-lines for animal experiments and related activities at TohokuUniversity
22 Digestion of Col-V Col-V (from human placenta) waspurchased from BD Biosciences (Bedford MA) Enzymesolutions of ColH and ColG were prepared at 01mgmLwith 100mM HEPES (pH 80) and 1mM CaCl
2 Col-V
(087mgmL) and enzyme solution (ColH or ColG) weremixed at a ratio of 1 1 The reactions were conducted at 30∘Cand sampled at 3 6 9 12 15 30 60 120 and 240min forsodium dodecyl sulfate-polyacrylamide gel electrophoresis(SDS-PAGE) and mass spectrometry The sampling solu-tions were quenched by adding one-fifth volume of 50mMethylenediaminetetraacetic acid for SDS-PAGE and 1 tri-fluoroacetic acid for mass spectrometry The SDS-PAGE gelswere stained with Imperial Protein Stain (Thermo FisherScientific Inc Waltham MA) SDS-PAGE gel images forthe lanes sampled at 60 120 and 240min were analyzedusing Atto CS Analyzer 3 software (Atto Corporation TokyoJapan)
The sampling solution at 60min for ESI-MS was incu-bated at 60∘C for 30min and then digested (overnight) at37∘C with 10 120583gmL trypsin adjusted to pH gt 7 by sodiumbicarbonate before digestion
23 MALDI-tof-MS Analysis MALDI-tof-MS analyses werecarried out using anAXIMAperformancemass spectrometer(Shimadzu Kyoto Japan) equipped with a 337 nm nitrogenlaser operating in the positive-ion mode with an acceleratingvoltage of +20 kV A timed ion selector was used to deflections of low mz (lt500) from the detector The spectra wereacquired by averaging the data from 200 laser shots toimprove the data quality and ion statistics The mass spectrawere calibrated with angiotensin II and ACTH fragment18ndash39 (Sigma) as external mass standards using 120572-cyano-4-hydroxycinnamic acid as a matrix The spectra were pro-cessed using Biotech Launchpad software (Shimadzu)
24 Digestion of Porcine Pancreatic Tissue Small pieces ofporcine pancreatic tissue (sim60mg) were incubated with01mgmL of thermolysin (Hampton Research Aliso ViejoCA) in 20mM HEPES (pH 80) and 1mM CaCl
2contain-
ing a protease inhibitor cocktail (complete Roche BaselSwitzerland) at 30∘C overnight The pancreatic tissues werethen washed with the same buffer and further digested at30∘C with 01mgmL of ColH ColG or ColH + ColG for9 h The digestions were performed for three times for eachcondition Each digested sample (100120583L) was precipitatedwith ice-cold 10 trichloroacetic acid in acetone (400120583L)and dried after centrifugation at 13000timesgThe dried sampleswere digested with trypsin (10 120583gmL) at 37∘C in 100mMammonium bicarbonate solution overnight
25 Peptide Identification by ESI-MS The digested peptidesfrom the Col-V and tissue samples were treated with a ZipTip(Millipore Billerica MA) for mass spectrometry analysis
Journal of Diabetes Research 3
and loaded onto a 75 120583m diameter 10 cm length fused-silicacapillary column containing C18 resin (Nikkyo TechnosTokyo Japan) The peptides were eluted with an acetonitrilegradient (typically 24 to 22 in 46min then to 33in 49min) in 01 formic acid and analyzed by an LTQOrbitrap Velos mass spectrometer (Thermo Fisher ScientificInc) Database searches against the NCBInr and Swissprotdatabases were performed using the MASCOT ver 251search engine (Matrix Science London UK) The MASCOTsearches were carried out without protease specificity using5 ppm peptide mass tolerance and 05 Da MSMS toleranceOxidation (+159949Da) was considered as a variable mod-ification for Met Cys Pro and Lys The peptides identifiedwith MASCOT expectation values lt 005 were considered assignificant hits
3 Results
31 In Vitro Digestion of Col-V by Collagenases
311 SDS-PAGE Analysis of Col-V Digestion In vitro diges-tion of Col-V was performed to investigate the differences indigestion patterns by the collagenases Figure 2(a) shows anSDS-PAGE analysis of the Col-V digestion patterns by ColHand ColG from 15 to 60min The results showed distinctdifferences between the Col-V digestion patterns by ColHand ColG On the SDS-PAGE gel no band changes wereobserved in the lanes for ColHwithin 1 h while ColG showedsignificant digestion of Col-V within this time frame
312 MALDI Analysis of Col-V Digestion The SDS-PAGEanalysis showed clear differences in the Col-V digestionpatterns between ColH and ColG Therefore to sensitivelydetect the low molecular weight peptide fragments MALDI-tof-MS analyses were conducted for the digestion solutionsat 15 30 and 60min The Col-V digestion samples by ColHindicated only a few peaks aroundmz 700ndash800 which couldreflect impurities arising from thematrix or originally presentwithin the enzymeCol-V solutions (Figure 2(b)) In contrastthe samples digested by ColG exhibited many peaks aroundmz 1000ndash3000 These peaks were considered to be Col-Vpeptide fragments digested by ColG Although the individualpeak intensities could not be compared quantitatively inthe MALDI spectra the relative intensity patterns changedindicating that the digestion sites by ColG changed over time
32 Peptide Fragment Analysis by ESI-MS To investigatethe Col-V cleavage sites by ColG the digested sample at1 h was further fragmented by trypsin and analyzed byLC-MS Peptide sequences and modifications (hydroxylatedproline and lysine) were identified by MSMS analysis TheMS experiments were performed in duplicate The analyzedpeptides included trypticsemitryptic peptides (digested atthe C-terminus of LysArg) collagenolytic peptides (digestedat the N-terminus of Gly-Xaa-Yaa repeats) and other pep-tides (neither tryptic nor collagenolytic peptides) The pep-tides were categorized tryptic peptides semitryptic peptidesincludingnot including Gly-Xaa-Yaa sequences at either
205
116974
6955
365
Mar
ker
Col
-V
Col
G
Col
H
V +
H15
min
V +
H30
min
V +
H60
min
V +
G15
min
V +
G30
min
V +
G60
min
(a)
ColG 15min
ColG 30min
ColG 60min
ColH 15min
ColH 30min
ColH 60min
1000 1500 2000 2500 3000
(b)
Figure 2 In vitro Col-V digestion by ColH and ColG (a) SDS-PAGE analysis of digested Col-V (b) MALDI-tof-MS analysis of thedigested samples at 15 30 and 60minThepeak intensities are scaledto 100 for the most intense peak in each chart
side and nontryptic peptides includingnot including Gly-Xaa-Yaa sequences at either side or both sides Consideringthis categorization the total number of peptides was countedas the sum of the number of tryptic peptides + number ofall semitryptic peptides + number of all nontryptic peptidesThe collagenolytic peptides were calculated as the sum of thesemitryptic peptides and nontryptic peptides including Gly-Xaa-Yaa sequences at either side or both sidesThe results aresummarized in Table 1The total number of detected peptideswas in the order of ColGtrypsin digestion gt ColHtrypsingt trypsin It was considered that the fragmentation of Col-V proceeded by double enzyme digestions especially for thecombination of ColGtrypsin The Col-V digested by trypsinconsisted of mainly tryptic and semitryptic peptides Sometryptic peptides were shared with collagenolytic peptidesbecause the Col-V sequence included lysine or arginine atthe Yaa position of the Gly-Xaa-Yaa repeats The double-digested samples included both tryptic and collagenolytic
4 Journal of Diabetes Research
Table 1 Peptide numbers detected by ESI-MS
pep tryp semitryp nontryp col pepGxx-G non-Gxx-G Gxx-G non-Gxx-G
V1205721 29 31 20 33 1 2 8 6 0 0 0 0 3 61205722 45 38 30 25 4 6 11 7 0 0 0 0 9 161205723 17 17 15 15 0 0 2 2 0 0 0 0 0 0
V + H1205721 39 33 21 20 9 6 4 6 3 0 0 0 31 271205722 46 47 26 30 9 7 10 9 1 1 0 0 22 171205723 21 18 15 16 4 1 2 1 0 0 0 0 19 6
V + G1205721 64 56 17 15 17 16 8 5 22 20 0 0 61 641205722 71 62 22 19 28 26 7 2 14 15 0 0 59 661205723 34 24 10 7 12 9 1 1 11 7 0 0 68 67
pep tryp semitryp and nontryp indicate the number of total peptides number of tryptic peptides (RK)darrxxxsdot sdot sdot xxx(RK)darrxxxsdot sdot sdot number ofsemitryptic peptides (RK)darrxxxsdot sdot sdot or sdot sdot sdot xxx(RK)darrxxxsdot sdot sdot and number of nontryptic peptides respectivelyGxx-G is the preferred substrate sequence for collagenase at position P3-P2-P1-P11015840Type V collagen was digested by trypsin (V) trypsin + collagenase H (V + H) and trypsin + collagenase G (V + G)The percentage of collagenolytic peptides was calculated by the following equation col pep = (number of semitryptic Gxx-G + number of nontryptic Gxx-G)(number of total peptides)
peptides In the ColHtrypsin digestion the percentage ofcollagenolytic peptides was around 30 for 1205721 and 20 for1205722 In contrast the percentage of collagenolytic peptides forColGtrypsin digestion exceeded 60ndash70 for all chain typesThese results suggested that collagenolytic peptides were themain components in the ColGtrypsin digestion sample andtherefore that ColG digested Col-V
33 Initial Digestion of Col-V by ColG
331 SDS-PAGEAnalysis In this study in vitro digestionwasperformed for 15ndash60min as described above However theCol-V digestion had already started by 15min To analyze theinitial stage of the digestion the Col-V digestion experimentwas performed for a shorter time (3ndash15min) and the digestedsamples were analyzed by SDS-PAGE (Figure 3) The mainbands shifted to lower molecular weights over time Further-more below themain bands some new bands appeared suchas the band at sim95 kDa
332 Cleavage Site Distribution Analysis The above resultsindicated that Col-V was degraded by ColG and thereforethat the sample solutions within 15min included peptidefragments digested by ColG The digestion solutions at 3 69 12 and 15min were mixed in equal amounts and measuredby LC-MS The detected peptides contained a Gly-Xaa-Yaasequence at the N- or C-terminus The cleavage sites weremapped on the Col-V sequences (1205721 1205722 and 1205723) (Supple-mentary Figure 1 red triangles see Supplementary Materialavailable online at httpdxdoiorg10115520164396756)The cleavage sites were distributed in the whole region of theGly-Xaa-Yaa repeats
The initial digestionwas evaluated by analyzing the short-term digestion In this digestion experiment the shortestdigestion time was 3min The 3min digestion solutionwas subjected to LC-MS analysis and the cleavage sitesequences (containing Gly-Xaa-Yaa) were mapped on thesame sequences described above (Supplementary Figure 1
205
116974
6955
365
Mar
ker
Col
-V
Col
G3
min
Col
G6
min
Col
G9
min
Col
G12
min
Col
G15
min
Figure 3 SDS-PAGE analysis of Col-V digestion by ColG within15min
blue triangles) The cleavage sites within the 3-min digestionsample were distributed on the N-terminal side of the 1205721 and1205722 chains In this experiment Col-V from human placentawas used Human placenta contains Col-V in the form of1205721(V)1205722(V)1205723(V) [23] Thus the 1205723 chain could be detectedHowever the distribution in the 1205723 chain was not clearbecause the relative abundance of the detected peptides wasmuch lower than those of the other two chains
For the 1205721 chain the modification positions for hydroxylgroups onto proline have been reported [24] The ColGpreference for hydroxyproline (Hyp) at the Yaa position canbe calculated by (number of Hyp at the Yaa position within
Journal of Diabetes Research 5
cleavage sites)(number of cleavage sites) = 2953 = 547Meanwhile the average distribution of Hyp within the Gly-Xaa-Yaa repeats can be calculated by (number of Hyp atthe Yaa position)(number of Gly-Xaa-Yaa repeats) = 97338= 287 These results indicated that Hyp residues werepreferentially involved at theYaa position in the cleavage sites
34 Long-Term Digestion of Col-V by ColG The short-termdigestion of Col-V by ColG revealed an interesting digestionpattern Therefore we conducted a long-term digestionexperiment to investigate the digestion pattern from 1 to4 h The digestion samples were analyzed by SDS-PAGE(Figure 4(a)) On the SDS-PAGE gel the main bands atgt120 kDa disappeared and several bands appeared in thelower molecular weight region These bands were numbered1ndash8 Of these bands the intensities of bands 1ndash6 decreasedover time (Figure 4(b)) Meanwhile the lower molecularweight bands (7 and 8) increased in intensity over timeAlthough the cleavage sites cannot be identified by SDS-PAGE analysis the appearance of distinct bands indicatedthe existence of region selectivity for ColG digestion withinthe Gly-Xaa-Yaa repeats of Col-V Furthermore the newlyappearing bands also decreased in the long-term diges-tion suggesting that there were secondary digestions of thedigested fragments
35 Tissue Digestion and MS Analysis
351 Porcine Pancreatic Tissue Digestion During pancre-atic islet isolation the exogenous enzymes used are ColHColG and thermolysin [1] Of these enzymes thermolysindoes not digest collagens under physiological conditions(manuscript in preparation) The porcine pancreatic tissuedigestion experiments were carried out in two steps Thefirst step involved digestion by thermolysin and the secondstep involved digestion by ColH ColG or ColH + ColG(double digestion) These two-step digestions had significanteffects on detecting the collagen peptides in the subsequentmass spectrometric analyses because the digested peptidesby thermolysin could be removed from the solution beforethe second digestion Figure 5 shows the observed pancreatictissue digestion patterns over time The tissue samples werenot digested by thermolysin alone To exclude any influenceof the proteins digested by thermolysin the sample solu-tions were changed after the thermolysin digestion and theobservations were started after addition of the collagenasesInterestingly the pancreatic tissues were not digested byColH or ColG alone Only the solution including both ColHand ColG exhibited tissue degradation These observationsare consistent with our previous findings that both ColGand ColH are necessary for dissociating porcine pancreatictissues Although the observations were carried out until 9 hall three tissue pieces were digested within 3 h
352 Col-V Fragments Produced by ColG Digestion of Pancre-atic Tissue The digested samples by ColH and ColG wereapplied to LC-MS analysis Database searches focused onidentifying collagen peptides were carried outThe identifiedcollagen peptides were picked up from the search results
205
116974
69
55
365
ColGlowast1
lowast2
lowast3
lowast4
lowast5
lowast6
lowast7
lowast8
Col
-V
Mar
ker
V +
G1
hr
V +
G2
hrs
V +
G4
hrs
(a)
ColGColG ColG
Inte
nsity
(au
)
lowast1
lowast1
lowast1
lowast2
lowast2
lowast2
lowast3 lowast
3
lowast3
lowast4
lowast4
lowast4
lowast5
lowast5
lowast5
lowast6
lowast6
lowast6
lowast7
lowast7
lowast7
lowast8
lowast8
lowast8
V + G 1hr V + G 2hrs V + G 4hrs
(b)
Figure 4 SDS-PAGE analysis of Col-V digestion by ColG over along time period (sim4 h) (a) SDS-PAGE analysis of Col-V digestionsColG and the main bands are numbered (b) Band intensity analysisby densitometry
and are summarized in Table 2 Most of the identifiedpeptides were trypticsemitryptic peptides Col-V peptideswere hardly detected and their sequences were shared bytrypticsemitryptic peptides It was speculated that Col-Vdigestion proceeded to unsuitably small fragments in the
6 Journal of Diabetes Research
Col
G
n2
n1
n3
n2
n1
n3
n2
n1
n3
Col
HC
olH
+ G
0hr 3hrs 6hrs 9hrs
Figure 5 In vitro porcine pancreatic tissue digestions by proteasesThe digestions were observed at 0 3 6 and 9 h Each digestion wasconducted in triplicate
presence of ColG together with other endogenous proteasesfor detection of the mass spectrometric analyses
4 Discussion
Our in vitro digestion experiments revealed that the Col-Vdigestion patterns were distinctly different betweenColH andColG
Col-V digestion by ColG was clearly shown by SDS-PAGE and mass spectrometric analyses Meanwhile ColHdid not digest Col-V under the same conditions used forColGAlthough this study could not investigate the differencein binding ability toward Col-V between ColH and ColG itis considered that the difference in domain construction is
Table 2 Collagen peptide identification from tissue digestion sam-ples
Col type tryp + semitryp nontryp Gxx-G
Col-V 1205721 2 0 0 21205722 4 0 0 3
Col-IV 1205721 1 2 3 21205722 1 1 1 1
Col-VI1205721 5 8 0 01205722 9 1 1 01205723 3 2 1 0
tryp semitryp and nontryp indicate the number of tryptic peptides (RK)darrxxxsdot sdot sdot xxx(RK)darrxxxsdot sdot sdot number of semitryptic peptides (RK)darrxxxsdot sdot sdotor sdot sdot sdot xxx(RK)darrxxxsdot sdot sdot and number of nontryptic peptides respectivelyThe total number of detected peptides is shown by tryp + semitryp + nontryp Gxx-G can be shared in tryp semitryp and nontrypDatabase searches for porcine collagens were based on the following NCBIaccession numbers 1205721(V) 62461592 1205722(V) 157427707 1205721(IV) 9271916811205722(IV) 927191683 1205721(VI) 92020086 92020094 and 545895133 1205722(VI)927203606 1205723(VI) 92020131 and 927211912
one of the possible reasons for the different collagenolyticactivities of ColH and ColG for Col-V ColH has onlyone CBD while ColG contains tandem CBDs (Figure 1)The CBDs of clostridial collagenases have been investigatedin detail A structural comparison indicated that the twostructures were similar (rmsd for C120572 = 15 A) despite therelatively low sequence similarity [11] The substrate-bindingspecificity of CBDs is broad for collagen types IndashIV [6] TheCBDs of ColH and ColG can bind to triple helical collagensbut not to telopeptides [5 25] However tandem CDBs canbind more efficiently to Col-I than a monomeric CBD [25]Consequently the domain construction especially the CBDarrangement plays a very important role in the digestioncharacteristics for Col-V
In the present study Col-V digestion by ColG produceddistinct digestion characteristics within a short time period(within minutes) and a long time period (within hours)The mass spectrometric analyses revealed that the identifiedpeptide cleavage sites were distributed in the N-terminalregion of theGly-Xaa-Yaa repeat region of Col-VThis findingsuggests that the digestion of Col-V is initiated from the N-terminal region According to the ColG digestion for Col-I [26] Col-I is hydrolyzed near the C-terminal end of thetriple helix region and a second cleavage occurs at the N-terminal side Although a digestion tendency for the C-terminal side was not found in our results digestion fromthe N-terminal region is a common digestion property ofColG for both Col-I and Col-V The long-term digestionexperiment indicated several residual parts of Col-V thatappeared as distinct bands at 1 h on SDS-PAGE These partswere also hydrolyzed by subsequent digestion (sim4 h) It issuggested that the Col-V sequence includes some parts thatare resistant to collagenolysis
The tissue digestion experiments clearly showed that bothColH and ColG were needed to degrade the pancreatic tissuestructure For human tissue degradation it was reported thatthe ratio of ColGH influenced the islet isolation outcome
Journal of Diabetes Research 7
[27 28] These results indicate that ColG may play moreimportant roles in digestion procedure of pig and humantissues than that of rat tissues The mass spectrometricanalysis for the digested samples identified collagen peptidesof Col-IV Col-V and collagen type VI Our previous studyrevealed that ColH is crucial for rat pancreatic islet isolationand the most abundantly detected collagen peptides in themass spectrometric analysis were Col-I and Col-III [17]These results strongly suggested the presence of markeddifferences between rat and porcine pancreatic tissue degra-dations Although it is difficult to identify the exact reasonfor the discrepancies fibril formation could be a possibleexplanation Collagen molecules are differently involved invarious tissues according to animal species [29] Col-V isassociated with Col-I in the microfibrillar structure [3031] Furthermore Col-V can be involved in heparinheparinsulfate modifications [32 33] and glycosylation [34] aswell as the well-known hydroxyl group modifications atproline and lysine residues [35] Therefore the differences incircumstances of the Col-IV fibrillar structures in the ratand porcine pancreatic tissue may influence the pancreaticislet isolation efficiency Although the similarities of thecircumstances of the Col-IV fibrillar structures among ratspigs and humans for pancreatic islet isolation are difficult tocompare ColG appears to be more important in porcine andhuman islet isolation processes and Col-V may be one of thekey substrates for ColG
5 Conclusions
In this study we focused on Col-V digestion by clostridialcollagenases Col-V was distinctively digested by ColG Massspectrometric analyses revealed that Col-V digestion byColG proceeded from the N-terminal region Although bothclostridial collagenases ColH and ColG include commondomains such as the activator domain peptidase domainPKD and CBD their domain construction is different Thedomain construction may play a crucial role for determiningthe collagen digestion patterns by the clostridial collagenasesThe porcine pancreatic tissue digestion experiments demon-strated that both collagenases are needed to degrade the tissuestructure These observations are completely different fromthose in rat experiments in which ColG was not cruciallyneeded Consequently the present study reveals that ColG aswell as ColH plays a crucial role in pancreatic islet isolationin large mammals and that Col-V can be one of the keysubstrates for ColG
Competing Interests
The authors of this paper received partial financial supportfromMeiji Seika PharmaCo Ltd and ShimadzuCorporationaccording to a collaborative research contract
Authorsrsquo Contributions
Kazutaka Murayama designed the experiments performedthe sample digestion andMALDI-tof-MS analyses andwrote
the paper Akiko Inagaki and Takehiro Imura performed thetissue digestion experiments Hiroki Shima performed theLC-MS spectrometric analyses and wrote part of the paperYouhei Yamagata and Kimiko Watanabe participated in thepaper corrections and critical review Kazuhiko Igarashi andMasafumi Goto participated in discussions during the paperpreparation and paper corrections
Acknowledgments
The authors acknowledge the support of the BiomedicalResearch Core and TAMRIC (Tohoku Advanced MedicalResearch and IncubationCenter) of TohokuUniversityGrad-uate School of Medicine Part of this research was supportedby the Tohoku University Business Incubation Program
References
[1] R C McCarthy A G Breite M L Green and F E DwuletldquoTissue dissociation enzymes for isolating human islets fortransplantation Factors to consider in setting enzyme accep-tance criteriardquo Transplantation vol 91 no 2 pp 137ndash145 2011
[2] K Watanabe ldquoCollagenolytic proteases from bacteriardquo AppliedMicrobiology and Biotechnology vol 63 no 5 pp 520ndash5262004
[3] U Eckhard E Schonauer P Ducka P Briza D Nuss andH Brandstetter ldquoBiochemical characterization of the catalyticdomains of three different clostridial collagenasesrdquo BiologicalChemistry vol 390 no 1 pp 11ndash18 2009
[4] C-M Jung O Matsushita S Katayama J Minami J Sakuraiand A Okabe ldquoIdentification of metal ligands in the Clostrid-ium histolyticumColH collagenaserdquo Journal of Bacteriology vol181 no 9 pp 2816ndash2822 1999
[5] O Matsushita C-M Jung J Minami S Katayama N Nishiand A Okabe ldquoA study of the collagen-binding domain of a116-kDa Clostridium histolyticum collagenaserdquo The Journal ofBiological Chemistry vol 273 no 6 pp 3643ndash3648 1998
[6] T Toyoshima O Matsushita J Minami N Nishi A Okabeand T Itano ldquoCollagen-binding domain of a Clostridium his-tolyticum collagenase exhibits a broad substrate spectrum bothin vitro and in vivordquo Connective Tissue Research vol 42 no 4pp 281ndash290 2001
[7] U Eckhard E Schonauer D Nuss and H Brandstetter ldquoStruc-ture of collagenase G reveals a chew-and-digest mechanismof bacterial collagenolysisrdquo Nature Structural and MolecularBiology vol 18 no 10 pp 1109ndash1114 2010
[8] U Eckhard E Schonauer andH Brandstetter ldquoStructural basisfor activity regulation and substrate preference of clostridialcollagenases G H and Trdquo The Journal of Biological Chemistryvol 288 no 28 pp 20184ndash20194 2013
[9] R Bauer K Janowska K Taylor et al ldquoStructures of threepolycystic kidney disease-like domains from Clostridium his-tolyticum collagenases ColG and ColHrdquo Acta CrystallographicaSection D Biological Crystallography vol 71 pp 565ndash577 2015
[10] U Eckhard and H Brandstetter ldquoPolycystic kidney disease-likedomains of clostridial collagenases and their role in collagenrecruitmentrdquo Biological Chemistry vol 392 no 11 pp 1039ndash1045 2011
8 Journal of Diabetes Research
[11] R Bauer J J Wilson S T L Philominathan D Davis OMatsushita and J Sakona ldquoStructural comparison of ColHand ColG collagen-binding domains from Clostridium his-tolyticumrdquo Journal of Bacteriology vol 195 no 2 pp 318ndash3272013
[12] J J Wilson O Matsushita A Okabe and J Sakon ldquoA bacterialcollagen-binding domain with novel calcium-binding motifcontrols domain orientationrdquoThe EMBO Journal vol 22 no 8pp 1743ndash1752 2003
[13] N Ohbayashi T Matsumoto H Shima et al ldquoSolution struc-ture of clostridial collagenase H and its calcium-dependentglobal conformation changerdquo Biophysical Journal vol 104 no7 pp 1538ndash1545 2013
[14] N Ohbayashi N Yamagata M Goto K Watanabe Y Yam-agata and K Murayama ldquoEnhancement of the structuralstability of full-length clostridial collagenase by calcium ionsrdquoApplied and Environmental Microbiology vol 78 no 16 pp5839ndash5844 2012
[15] U Eckhard P F Huesgen H Brandstetter and C M OverallldquoProteomic protease specificity profiling of clostridial collage-nases reveals their intrinsic nature as dedicated degraders ofcollagenrdquo Journal of Proteomics vol 100 pp 102ndash114 2014
[16] O Matsushita and A Okabe ldquoClostridial hydrolytic enzymesdegrading extracellular componentsrdquoToxicon vol 39 no 11 pp1769ndash1780 2001
[17] A Fujio K Murayama Y Yamagata et al ldquoCollagenase H iscrucial for isolation of rat pancreatic isletsrdquo Cell Transplanta-tion vol 23 no 10 pp 1187ndash1198 2014
[18] M K Gordon and R A Hahn ldquoCollagensrdquo Cell and TissueResearch vol 339 no 1 pp 247ndash257 2010
[19] MD Shoulders andR T Raines ldquoCollagen structure and stabil-ityrdquo Annual Review of Biochemistry vol 78 pp 929ndash958 2009
[20] D E Birk ldquoType V collagen heterotypic type IV collageninteractions in the regulation of fibril assemblyrdquoMicron vol 32no 3 pp 223ndash237 2001
[21] M A Weis D M Hudson L Kim M Scott J-J Wu and D REyre ldquoLocation of 3-Hydroxyproline residues in collagen typesI II III and VXI implies a role in fibril supramolecularassemblyrdquo Journal of Biological Chemistry vol 285 no 4 pp2580ndash2590 2010
[22] R J Wenstrup J B Florer E W Brunskill S M Bell I Cher-voneva and D E Birk ldquoType V collagen controls the initiationof collagen fibril assemblyrdquo Journal of Biological Chemistry vol279 no 51 pp 53331ndash53337 2004
[23] C Niyibizi P P Fietzek and M Van der Rest ldquoHumanplacenta type V collagens Evidence for the existence of an1205721(V)1205722(V)1205723(V) collagen moleculerdquo Journal of BiologicalChemistry vol 259 no 22 pp 14170ndash14174 1984
[24] C Yang A C Park N A Davis et al ldquoComprehensive massspectrometricmapping of the hydroxylated amino acid residuesof the1205721(V) collagen chainrdquoThe Journal of Biological Chemistryvol 287 no 48 pp 40598ndash40610 2012
[25] O Matsushita T Koide R Kobayashi K Nagata and AOkabe ldquoSubstrate recognition by the collagen-binding domainof Clostridium histolyticum class I collagenaserdquo The Journal ofBiological Chemistry vol 276 no 12 pp 8761ndash8770 2001
[26] M F French K A Mookhtiar and H E Van Wart ldquoLimitedproteolysis of type I collagen at hyperreactive sites by class Iand II Clostridium histolyticum collagenases complementarydigestion patternsrdquo Biochemistry vol 26 no 3 pp 681ndash6871987
[27] T Kin X Zhai T B Murdoch A Salam A M J Shapiro andJ R T Lakey ldquoEnhancing the success of human islet isolationthrough optimization and characterization of pancreas dissoci-ation enzymerdquo American Journal of Transplantation vol 7 no5 pp 1233ndash1241 2007
[28] T Kin X Zhai D OrsquoGorman and A M J Shapiro ldquoDetri-mental effect of excessive collagenase class II on human isletisolation outcomerdquo Transplant International vol 21 no 11 pp1059ndash1065 2008
[29] J H M Van Deijnen P T R Van Suylichem G H J Woltersand R Van Schilfgaarde ldquoDistribution of collagens type I typeIII and type V in the pancreas of rat dog pig andmanrdquoCell andTissue Research vol 277 no 1 pp 115ndash121 1994
[30] T F Linsenmayer E Gibney F Igoe et al ldquoType V collagenmolecular structure and fibrillar organization of the chicken1205721(V) NH2-terminal domain a putative regulator of cornealfibrillogenesisrdquo Journal of Cell Biology vol 121 no 5 pp 1181ndash1189 1993
[31] J K Mouw G Ou and V M Weaver ldquoExtracellular matrixassembly a multiscale deconstructionrdquo Nature Reviews Molec-ular Cell Biology vol 15 no 12 pp 771ndash785 2014
[32] F Delacoux A Fichard C Geourjon R Garrone and FRuggiero ldquoMolecular features of the collagenVheparin bindingsiterdquo The Journal of Biological Chemistry vol 273 no 24 pp15069ndash15076 1998
[33] S Ricard-Blum M Beraud N Raynal R W Farndale and FRuggiero ldquoStructural requirements for heparinheparan sulfatebinding to type V collagenrdquoThe Journal of Biological Chemistryvol 281 no 35 pp 25195ndash25204 2006
[34] W Henkel and K Dreisewerd ldquoCyanogen bromide peptides ofthe fibrillar collagens I III and V and their mass spectrometriccharacterization detection of linear peptides peptide glycosyla-tion and cross-linking peptides involved in formation of homo-and heterotypic fibrilsrdquo Journal of Proteome Research vol 6 no11 pp 4269ndash4289 2007
[35] K Gelse E Poschl and T Aigner ldquoCollagensmdashstructurefunction and biosynthesisrdquo Advanced Drug Delivery Reviewsvol 55 no 12 pp 1531ndash1546 2003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

2 Journal of Diabetes Research
ColH
ColG
PepAct
CBDCBD
CBD
PKD
PKD PKD
Act Pep
N
N C
C
Figure 1 Schematic diagram of the domain architectures ofColG and ColH Act activator domain Pep peptidase domainPKD polycystic kidney disease-like domain CBD collagen-bindingdomain The collagenase module is composed of two functionaldomains an activator domain and a peptidase domain
Gly in the Gly-Xaa-Yaa(-Gly-) sequence repeats of fibrillarcollagens [15 16] Degradation of collagen type I (Col-I) iswell-known for collagenolytic activity of ColH and ColGColH also plays a crucial role during rat pancreatic isletisolation [17] In vitro pancreatic tissue digestion experimentsand mass spectrometric analyses revealed that Col-I andcollagen type III (Col-III) are substrates for ColH [17]Although the coexistence of ColG can increase the yield ofisolated islets it has been difficult to determine the substratespecificity for ColG in rat experiments Meanwhile bothColH and ColG are used for pancreatic tissue dissociation inlarge mammals such as pigs Identification of the substratefor ColG in pancreatic tissue is one of the key factors towardunderstanding the pancreatic islet isolation process
The fibrillar collagens are targets for many collagenases[2] The commonly found fibrillar collagens are types I IIIII and V [18] A previous study showed that Col-I collagentype II (Col-II) and Col-III were fully digested by ColG [17]while collagen type V (Col-V) was relatively less digested(manuscript in preparation) Col-V is a fibrillar collagenwith low abundance Several isoforms are found for Col-Vdepending on the chain components [19]Themost commonisoform is 1205721(V)
21205722(V) However other isoforms such as
1205721(V)3and 1205721(V)1205722(V)1205723(V) have also been reported [20]
Col-V exists in tissues colocalized with Col-I and appears toregulate the assembly of heterotypic fibers containing Col-Iand Col-V [20ndash22]
In the present study Col-V digestion byColGwas focusedupon to investigate the substrate specificity of ColG Thedigestions were analyzed by SDS-PAGE and mass spec-trometry The differences in Col-V digestion by ColH andColG were evaluated The results revealed that Col-V is aputative substrate for ColG in pancreatic islet isolation inlarge mammals
2 Materials and Methods
21 Collagenases and Porcine Pancreatic Tissue Recombinantenzymes of ColG and ColH which were kindly providedby Meiji Seika Pharma Co Ltd were produced by anEscherichia coli protein expression systemwith ColG or ColHgenes isolated from Clostridium histolyticum as describedpreviously [17]
Porcine pancreatic tissue samples were obtained from2ndash4-year-old adult retired sows weighing 200ndash300 kg Allanimal experiments were performed according to the guide-lines for animal experiments and related activities at TohokuUniversity
22 Digestion of Col-V Col-V (from human placenta) waspurchased from BD Biosciences (Bedford MA) Enzymesolutions of ColH and ColG were prepared at 01mgmLwith 100mM HEPES (pH 80) and 1mM CaCl
2 Col-V
(087mgmL) and enzyme solution (ColH or ColG) weremixed at a ratio of 1 1 The reactions were conducted at 30∘Cand sampled at 3 6 9 12 15 30 60 120 and 240min forsodium dodecyl sulfate-polyacrylamide gel electrophoresis(SDS-PAGE) and mass spectrometry The sampling solu-tions were quenched by adding one-fifth volume of 50mMethylenediaminetetraacetic acid for SDS-PAGE and 1 tri-fluoroacetic acid for mass spectrometry The SDS-PAGE gelswere stained with Imperial Protein Stain (Thermo FisherScientific Inc Waltham MA) SDS-PAGE gel images forthe lanes sampled at 60 120 and 240min were analyzedusing Atto CS Analyzer 3 software (Atto Corporation TokyoJapan)
The sampling solution at 60min for ESI-MS was incu-bated at 60∘C for 30min and then digested (overnight) at37∘C with 10 120583gmL trypsin adjusted to pH gt 7 by sodiumbicarbonate before digestion
23 MALDI-tof-MS Analysis MALDI-tof-MS analyses werecarried out using anAXIMAperformancemass spectrometer(Shimadzu Kyoto Japan) equipped with a 337 nm nitrogenlaser operating in the positive-ion mode with an acceleratingvoltage of +20 kV A timed ion selector was used to deflections of low mz (lt500) from the detector The spectra wereacquired by averaging the data from 200 laser shots toimprove the data quality and ion statistics The mass spectrawere calibrated with angiotensin II and ACTH fragment18ndash39 (Sigma) as external mass standards using 120572-cyano-4-hydroxycinnamic acid as a matrix The spectra were pro-cessed using Biotech Launchpad software (Shimadzu)
24 Digestion of Porcine Pancreatic Tissue Small pieces ofporcine pancreatic tissue (sim60mg) were incubated with01mgmL of thermolysin (Hampton Research Aliso ViejoCA) in 20mM HEPES (pH 80) and 1mM CaCl
2contain-
ing a protease inhibitor cocktail (complete Roche BaselSwitzerland) at 30∘C overnight The pancreatic tissues werethen washed with the same buffer and further digested at30∘C with 01mgmL of ColH ColG or ColH + ColG for9 h The digestions were performed for three times for eachcondition Each digested sample (100120583L) was precipitatedwith ice-cold 10 trichloroacetic acid in acetone (400120583L)and dried after centrifugation at 13000timesgThe dried sampleswere digested with trypsin (10 120583gmL) at 37∘C in 100mMammonium bicarbonate solution overnight
25 Peptide Identification by ESI-MS The digested peptidesfrom the Col-V and tissue samples were treated with a ZipTip(Millipore Billerica MA) for mass spectrometry analysis
Journal of Diabetes Research 3
and loaded onto a 75 120583m diameter 10 cm length fused-silicacapillary column containing C18 resin (Nikkyo TechnosTokyo Japan) The peptides were eluted with an acetonitrilegradient (typically 24 to 22 in 46min then to 33in 49min) in 01 formic acid and analyzed by an LTQOrbitrap Velos mass spectrometer (Thermo Fisher ScientificInc) Database searches against the NCBInr and Swissprotdatabases were performed using the MASCOT ver 251search engine (Matrix Science London UK) The MASCOTsearches were carried out without protease specificity using5 ppm peptide mass tolerance and 05 Da MSMS toleranceOxidation (+159949Da) was considered as a variable mod-ification for Met Cys Pro and Lys The peptides identifiedwith MASCOT expectation values lt 005 were considered assignificant hits
3 Results
31 In Vitro Digestion of Col-V by Collagenases
311 SDS-PAGE Analysis of Col-V Digestion In vitro diges-tion of Col-V was performed to investigate the differences indigestion patterns by the collagenases Figure 2(a) shows anSDS-PAGE analysis of the Col-V digestion patterns by ColHand ColG from 15 to 60min The results showed distinctdifferences between the Col-V digestion patterns by ColHand ColG On the SDS-PAGE gel no band changes wereobserved in the lanes for ColHwithin 1 h while ColG showedsignificant digestion of Col-V within this time frame
312 MALDI Analysis of Col-V Digestion The SDS-PAGEanalysis showed clear differences in the Col-V digestionpatterns between ColH and ColG Therefore to sensitivelydetect the low molecular weight peptide fragments MALDI-tof-MS analyses were conducted for the digestion solutionsat 15 30 and 60min The Col-V digestion samples by ColHindicated only a few peaks aroundmz 700ndash800 which couldreflect impurities arising from thematrix or originally presentwithin the enzymeCol-V solutions (Figure 2(b)) In contrastthe samples digested by ColG exhibited many peaks aroundmz 1000ndash3000 These peaks were considered to be Col-Vpeptide fragments digested by ColG Although the individualpeak intensities could not be compared quantitatively inthe MALDI spectra the relative intensity patterns changedindicating that the digestion sites by ColG changed over time
32 Peptide Fragment Analysis by ESI-MS To investigatethe Col-V cleavage sites by ColG the digested sample at1 h was further fragmented by trypsin and analyzed byLC-MS Peptide sequences and modifications (hydroxylatedproline and lysine) were identified by MSMS analysis TheMS experiments were performed in duplicate The analyzedpeptides included trypticsemitryptic peptides (digested atthe C-terminus of LysArg) collagenolytic peptides (digestedat the N-terminus of Gly-Xaa-Yaa repeats) and other pep-tides (neither tryptic nor collagenolytic peptides) The pep-tides were categorized tryptic peptides semitryptic peptidesincludingnot including Gly-Xaa-Yaa sequences at either
205
116974
6955
365
Mar
ker
Col
-V
Col
G
Col
H
V +
H15
min
V +
H30
min
V +
H60
min
V +
G15
min
V +
G30
min
V +
G60
min
(a)
ColG 15min
ColG 30min
ColG 60min
ColH 15min
ColH 30min
ColH 60min
1000 1500 2000 2500 3000
(b)
Figure 2 In vitro Col-V digestion by ColH and ColG (a) SDS-PAGE analysis of digested Col-V (b) MALDI-tof-MS analysis of thedigested samples at 15 30 and 60minThepeak intensities are scaledto 100 for the most intense peak in each chart
side and nontryptic peptides includingnot including Gly-Xaa-Yaa sequences at either side or both sides Consideringthis categorization the total number of peptides was countedas the sum of the number of tryptic peptides + number ofall semitryptic peptides + number of all nontryptic peptidesThe collagenolytic peptides were calculated as the sum of thesemitryptic peptides and nontryptic peptides including Gly-Xaa-Yaa sequences at either side or both sidesThe results aresummarized in Table 1The total number of detected peptideswas in the order of ColGtrypsin digestion gt ColHtrypsingt trypsin It was considered that the fragmentation of Col-V proceeded by double enzyme digestions especially for thecombination of ColGtrypsin The Col-V digested by trypsinconsisted of mainly tryptic and semitryptic peptides Sometryptic peptides were shared with collagenolytic peptidesbecause the Col-V sequence included lysine or arginine atthe Yaa position of the Gly-Xaa-Yaa repeats The double-digested samples included both tryptic and collagenolytic
4 Journal of Diabetes Research
Table 1 Peptide numbers detected by ESI-MS
pep tryp semitryp nontryp col pepGxx-G non-Gxx-G Gxx-G non-Gxx-G
V1205721 29 31 20 33 1 2 8 6 0 0 0 0 3 61205722 45 38 30 25 4 6 11 7 0 0 0 0 9 161205723 17 17 15 15 0 0 2 2 0 0 0 0 0 0
V + H1205721 39 33 21 20 9 6 4 6 3 0 0 0 31 271205722 46 47 26 30 9 7 10 9 1 1 0 0 22 171205723 21 18 15 16 4 1 2 1 0 0 0 0 19 6
V + G1205721 64 56 17 15 17 16 8 5 22 20 0 0 61 641205722 71 62 22 19 28 26 7 2 14 15 0 0 59 661205723 34 24 10 7 12 9 1 1 11 7 0 0 68 67
pep tryp semitryp and nontryp indicate the number of total peptides number of tryptic peptides (RK)darrxxxsdot sdot sdot xxx(RK)darrxxxsdot sdot sdot number ofsemitryptic peptides (RK)darrxxxsdot sdot sdot or sdot sdot sdot xxx(RK)darrxxxsdot sdot sdot and number of nontryptic peptides respectivelyGxx-G is the preferred substrate sequence for collagenase at position P3-P2-P1-P11015840Type V collagen was digested by trypsin (V) trypsin + collagenase H (V + H) and trypsin + collagenase G (V + G)The percentage of collagenolytic peptides was calculated by the following equation col pep = (number of semitryptic Gxx-G + number of nontryptic Gxx-G)(number of total peptides)
peptides In the ColHtrypsin digestion the percentage ofcollagenolytic peptides was around 30 for 1205721 and 20 for1205722 In contrast the percentage of collagenolytic peptides forColGtrypsin digestion exceeded 60ndash70 for all chain typesThese results suggested that collagenolytic peptides were themain components in the ColGtrypsin digestion sample andtherefore that ColG digested Col-V
33 Initial Digestion of Col-V by ColG
331 SDS-PAGEAnalysis In this study in vitro digestionwasperformed for 15ndash60min as described above However theCol-V digestion had already started by 15min To analyze theinitial stage of the digestion the Col-V digestion experimentwas performed for a shorter time (3ndash15min) and the digestedsamples were analyzed by SDS-PAGE (Figure 3) The mainbands shifted to lower molecular weights over time Further-more below themain bands some new bands appeared suchas the band at sim95 kDa
332 Cleavage Site Distribution Analysis The above resultsindicated that Col-V was degraded by ColG and thereforethat the sample solutions within 15min included peptidefragments digested by ColG The digestion solutions at 3 69 12 and 15min were mixed in equal amounts and measuredby LC-MS The detected peptides contained a Gly-Xaa-Yaasequence at the N- or C-terminus The cleavage sites weremapped on the Col-V sequences (1205721 1205722 and 1205723) (Supple-mentary Figure 1 red triangles see Supplementary Materialavailable online at httpdxdoiorg10115520164396756)The cleavage sites were distributed in the whole region of theGly-Xaa-Yaa repeats
The initial digestionwas evaluated by analyzing the short-term digestion In this digestion experiment the shortestdigestion time was 3min The 3min digestion solutionwas subjected to LC-MS analysis and the cleavage sitesequences (containing Gly-Xaa-Yaa) were mapped on thesame sequences described above (Supplementary Figure 1
205
116974
6955
365
Mar
ker
Col
-V
Col
G3
min
Col
G6
min
Col
G9
min
Col
G12
min
Col
G15
min
Figure 3 SDS-PAGE analysis of Col-V digestion by ColG within15min
blue triangles) The cleavage sites within the 3-min digestionsample were distributed on the N-terminal side of the 1205721 and1205722 chains In this experiment Col-V from human placentawas used Human placenta contains Col-V in the form of1205721(V)1205722(V)1205723(V) [23] Thus the 1205723 chain could be detectedHowever the distribution in the 1205723 chain was not clearbecause the relative abundance of the detected peptides wasmuch lower than those of the other two chains
For the 1205721 chain the modification positions for hydroxylgroups onto proline have been reported [24] The ColGpreference for hydroxyproline (Hyp) at the Yaa position canbe calculated by (number of Hyp at the Yaa position within
Journal of Diabetes Research 5
cleavage sites)(number of cleavage sites) = 2953 = 547Meanwhile the average distribution of Hyp within the Gly-Xaa-Yaa repeats can be calculated by (number of Hyp atthe Yaa position)(number of Gly-Xaa-Yaa repeats) = 97338= 287 These results indicated that Hyp residues werepreferentially involved at theYaa position in the cleavage sites
34 Long-Term Digestion of Col-V by ColG The short-termdigestion of Col-V by ColG revealed an interesting digestionpattern Therefore we conducted a long-term digestionexperiment to investigate the digestion pattern from 1 to4 h The digestion samples were analyzed by SDS-PAGE(Figure 4(a)) On the SDS-PAGE gel the main bands atgt120 kDa disappeared and several bands appeared in thelower molecular weight region These bands were numbered1ndash8 Of these bands the intensities of bands 1ndash6 decreasedover time (Figure 4(b)) Meanwhile the lower molecularweight bands (7 and 8) increased in intensity over timeAlthough the cleavage sites cannot be identified by SDS-PAGE analysis the appearance of distinct bands indicatedthe existence of region selectivity for ColG digestion withinthe Gly-Xaa-Yaa repeats of Col-V Furthermore the newlyappearing bands also decreased in the long-term diges-tion suggesting that there were secondary digestions of thedigested fragments
35 Tissue Digestion and MS Analysis
351 Porcine Pancreatic Tissue Digestion During pancre-atic islet isolation the exogenous enzymes used are ColHColG and thermolysin [1] Of these enzymes thermolysindoes not digest collagens under physiological conditions(manuscript in preparation) The porcine pancreatic tissuedigestion experiments were carried out in two steps Thefirst step involved digestion by thermolysin and the secondstep involved digestion by ColH ColG or ColH + ColG(double digestion) These two-step digestions had significanteffects on detecting the collagen peptides in the subsequentmass spectrometric analyses because the digested peptidesby thermolysin could be removed from the solution beforethe second digestion Figure 5 shows the observed pancreatictissue digestion patterns over time The tissue samples werenot digested by thermolysin alone To exclude any influenceof the proteins digested by thermolysin the sample solu-tions were changed after the thermolysin digestion and theobservations were started after addition of the collagenasesInterestingly the pancreatic tissues were not digested byColH or ColG alone Only the solution including both ColHand ColG exhibited tissue degradation These observationsare consistent with our previous findings that both ColGand ColH are necessary for dissociating porcine pancreatictissues Although the observations were carried out until 9 hall three tissue pieces were digested within 3 h
352 Col-V Fragments Produced by ColG Digestion of Pancre-atic Tissue The digested samples by ColH and ColG wereapplied to LC-MS analysis Database searches focused onidentifying collagen peptides were carried outThe identifiedcollagen peptides were picked up from the search results
205
116974
69
55
365
ColGlowast1
lowast2
lowast3
lowast4
lowast5
lowast6
lowast7
lowast8
Col
-V
Mar
ker
V +
G1
hr
V +
G2
hrs
V +
G4
hrs
(a)
ColGColG ColG
Inte
nsity
(au
)
lowast1
lowast1
lowast1
lowast2
lowast2
lowast2
lowast3 lowast
3
lowast3
lowast4
lowast4
lowast4
lowast5
lowast5
lowast5
lowast6
lowast6
lowast6
lowast7
lowast7
lowast7
lowast8
lowast8
lowast8
V + G 1hr V + G 2hrs V + G 4hrs
(b)
Figure 4 SDS-PAGE analysis of Col-V digestion by ColG over along time period (sim4 h) (a) SDS-PAGE analysis of Col-V digestionsColG and the main bands are numbered (b) Band intensity analysisby densitometry
and are summarized in Table 2 Most of the identifiedpeptides were trypticsemitryptic peptides Col-V peptideswere hardly detected and their sequences were shared bytrypticsemitryptic peptides It was speculated that Col-Vdigestion proceeded to unsuitably small fragments in the
6 Journal of Diabetes Research
Col
G
n2
n1
n3
n2
n1
n3
n2
n1
n3
Col
HC
olH
+ G
0hr 3hrs 6hrs 9hrs
Figure 5 In vitro porcine pancreatic tissue digestions by proteasesThe digestions were observed at 0 3 6 and 9 h Each digestion wasconducted in triplicate
presence of ColG together with other endogenous proteasesfor detection of the mass spectrometric analyses
4 Discussion
Our in vitro digestion experiments revealed that the Col-Vdigestion patterns were distinctly different betweenColH andColG
Col-V digestion by ColG was clearly shown by SDS-PAGE and mass spectrometric analyses Meanwhile ColHdid not digest Col-V under the same conditions used forColGAlthough this study could not investigate the differencein binding ability toward Col-V between ColH and ColG itis considered that the difference in domain construction is
Table 2 Collagen peptide identification from tissue digestion sam-ples
Col type tryp + semitryp nontryp Gxx-G
Col-V 1205721 2 0 0 21205722 4 0 0 3
Col-IV 1205721 1 2 3 21205722 1 1 1 1
Col-VI1205721 5 8 0 01205722 9 1 1 01205723 3 2 1 0
tryp semitryp and nontryp indicate the number of tryptic peptides (RK)darrxxxsdot sdot sdot xxx(RK)darrxxxsdot sdot sdot number of semitryptic peptides (RK)darrxxxsdot sdot sdotor sdot sdot sdot xxx(RK)darrxxxsdot sdot sdot and number of nontryptic peptides respectivelyThe total number of detected peptides is shown by tryp + semitryp + nontryp Gxx-G can be shared in tryp semitryp and nontrypDatabase searches for porcine collagens were based on the following NCBIaccession numbers 1205721(V) 62461592 1205722(V) 157427707 1205721(IV) 9271916811205722(IV) 927191683 1205721(VI) 92020086 92020094 and 545895133 1205722(VI)927203606 1205723(VI) 92020131 and 927211912
one of the possible reasons for the different collagenolyticactivities of ColH and ColG for Col-V ColH has onlyone CBD while ColG contains tandem CBDs (Figure 1)The CBDs of clostridial collagenases have been investigatedin detail A structural comparison indicated that the twostructures were similar (rmsd for C120572 = 15 A) despite therelatively low sequence similarity [11] The substrate-bindingspecificity of CBDs is broad for collagen types IndashIV [6] TheCBDs of ColH and ColG can bind to triple helical collagensbut not to telopeptides [5 25] However tandem CDBs canbind more efficiently to Col-I than a monomeric CBD [25]Consequently the domain construction especially the CBDarrangement plays a very important role in the digestioncharacteristics for Col-V
In the present study Col-V digestion by ColG produceddistinct digestion characteristics within a short time period(within minutes) and a long time period (within hours)The mass spectrometric analyses revealed that the identifiedpeptide cleavage sites were distributed in the N-terminalregion of theGly-Xaa-Yaa repeat region of Col-VThis findingsuggests that the digestion of Col-V is initiated from the N-terminal region According to the ColG digestion for Col-I [26] Col-I is hydrolyzed near the C-terminal end of thetriple helix region and a second cleavage occurs at the N-terminal side Although a digestion tendency for the C-terminal side was not found in our results digestion fromthe N-terminal region is a common digestion property ofColG for both Col-I and Col-V The long-term digestionexperiment indicated several residual parts of Col-V thatappeared as distinct bands at 1 h on SDS-PAGE These partswere also hydrolyzed by subsequent digestion (sim4 h) It issuggested that the Col-V sequence includes some parts thatare resistant to collagenolysis
The tissue digestion experiments clearly showed that bothColH and ColG were needed to degrade the pancreatic tissuestructure For human tissue degradation it was reported thatthe ratio of ColGH influenced the islet isolation outcome
Journal of Diabetes Research 7
[27 28] These results indicate that ColG may play moreimportant roles in digestion procedure of pig and humantissues than that of rat tissues The mass spectrometricanalysis for the digested samples identified collagen peptidesof Col-IV Col-V and collagen type VI Our previous studyrevealed that ColH is crucial for rat pancreatic islet isolationand the most abundantly detected collagen peptides in themass spectrometric analysis were Col-I and Col-III [17]These results strongly suggested the presence of markeddifferences between rat and porcine pancreatic tissue degra-dations Although it is difficult to identify the exact reasonfor the discrepancies fibril formation could be a possibleexplanation Collagen molecules are differently involved invarious tissues according to animal species [29] Col-V isassociated with Col-I in the microfibrillar structure [3031] Furthermore Col-V can be involved in heparinheparinsulfate modifications [32 33] and glycosylation [34] aswell as the well-known hydroxyl group modifications atproline and lysine residues [35] Therefore the differences incircumstances of the Col-IV fibrillar structures in the ratand porcine pancreatic tissue may influence the pancreaticislet isolation efficiency Although the similarities of thecircumstances of the Col-IV fibrillar structures among ratspigs and humans for pancreatic islet isolation are difficult tocompare ColG appears to be more important in porcine andhuman islet isolation processes and Col-V may be one of thekey substrates for ColG
5 Conclusions
In this study we focused on Col-V digestion by clostridialcollagenases Col-V was distinctively digested by ColG Massspectrometric analyses revealed that Col-V digestion byColG proceeded from the N-terminal region Although bothclostridial collagenases ColH and ColG include commondomains such as the activator domain peptidase domainPKD and CBD their domain construction is different Thedomain construction may play a crucial role for determiningthe collagen digestion patterns by the clostridial collagenasesThe porcine pancreatic tissue digestion experiments demon-strated that both collagenases are needed to degrade the tissuestructure These observations are completely different fromthose in rat experiments in which ColG was not cruciallyneeded Consequently the present study reveals that ColG aswell as ColH plays a crucial role in pancreatic islet isolationin large mammals and that Col-V can be one of the keysubstrates for ColG
Competing Interests
The authors of this paper received partial financial supportfromMeiji Seika PharmaCo Ltd and ShimadzuCorporationaccording to a collaborative research contract
Authorsrsquo Contributions
Kazutaka Murayama designed the experiments performedthe sample digestion andMALDI-tof-MS analyses andwrote
the paper Akiko Inagaki and Takehiro Imura performed thetissue digestion experiments Hiroki Shima performed theLC-MS spectrometric analyses and wrote part of the paperYouhei Yamagata and Kimiko Watanabe participated in thepaper corrections and critical review Kazuhiko Igarashi andMasafumi Goto participated in discussions during the paperpreparation and paper corrections
Acknowledgments
The authors acknowledge the support of the BiomedicalResearch Core and TAMRIC (Tohoku Advanced MedicalResearch and IncubationCenter) of TohokuUniversityGrad-uate School of Medicine Part of this research was supportedby the Tohoku University Business Incubation Program
References
[1] R C McCarthy A G Breite M L Green and F E DwuletldquoTissue dissociation enzymes for isolating human islets fortransplantation Factors to consider in setting enzyme accep-tance criteriardquo Transplantation vol 91 no 2 pp 137ndash145 2011
[2] K Watanabe ldquoCollagenolytic proteases from bacteriardquo AppliedMicrobiology and Biotechnology vol 63 no 5 pp 520ndash5262004
[3] U Eckhard E Schonauer P Ducka P Briza D Nuss andH Brandstetter ldquoBiochemical characterization of the catalyticdomains of three different clostridial collagenasesrdquo BiologicalChemistry vol 390 no 1 pp 11ndash18 2009
[4] C-M Jung O Matsushita S Katayama J Minami J Sakuraiand A Okabe ldquoIdentification of metal ligands in the Clostrid-ium histolyticumColH collagenaserdquo Journal of Bacteriology vol181 no 9 pp 2816ndash2822 1999
[5] O Matsushita C-M Jung J Minami S Katayama N Nishiand A Okabe ldquoA study of the collagen-binding domain of a116-kDa Clostridium histolyticum collagenaserdquo The Journal ofBiological Chemistry vol 273 no 6 pp 3643ndash3648 1998
[6] T Toyoshima O Matsushita J Minami N Nishi A Okabeand T Itano ldquoCollagen-binding domain of a Clostridium his-tolyticum collagenase exhibits a broad substrate spectrum bothin vitro and in vivordquo Connective Tissue Research vol 42 no 4pp 281ndash290 2001
[7] U Eckhard E Schonauer D Nuss and H Brandstetter ldquoStruc-ture of collagenase G reveals a chew-and-digest mechanismof bacterial collagenolysisrdquo Nature Structural and MolecularBiology vol 18 no 10 pp 1109ndash1114 2010
[8] U Eckhard E Schonauer andH Brandstetter ldquoStructural basisfor activity regulation and substrate preference of clostridialcollagenases G H and Trdquo The Journal of Biological Chemistryvol 288 no 28 pp 20184ndash20194 2013
[9] R Bauer K Janowska K Taylor et al ldquoStructures of threepolycystic kidney disease-like domains from Clostridium his-tolyticum collagenases ColG and ColHrdquo Acta CrystallographicaSection D Biological Crystallography vol 71 pp 565ndash577 2015
[10] U Eckhard and H Brandstetter ldquoPolycystic kidney disease-likedomains of clostridial collagenases and their role in collagenrecruitmentrdquo Biological Chemistry vol 392 no 11 pp 1039ndash1045 2011
8 Journal of Diabetes Research
[11] R Bauer J J Wilson S T L Philominathan D Davis OMatsushita and J Sakona ldquoStructural comparison of ColHand ColG collagen-binding domains from Clostridium his-tolyticumrdquo Journal of Bacteriology vol 195 no 2 pp 318ndash3272013
[12] J J Wilson O Matsushita A Okabe and J Sakon ldquoA bacterialcollagen-binding domain with novel calcium-binding motifcontrols domain orientationrdquoThe EMBO Journal vol 22 no 8pp 1743ndash1752 2003
[13] N Ohbayashi T Matsumoto H Shima et al ldquoSolution struc-ture of clostridial collagenase H and its calcium-dependentglobal conformation changerdquo Biophysical Journal vol 104 no7 pp 1538ndash1545 2013
[14] N Ohbayashi N Yamagata M Goto K Watanabe Y Yam-agata and K Murayama ldquoEnhancement of the structuralstability of full-length clostridial collagenase by calcium ionsrdquoApplied and Environmental Microbiology vol 78 no 16 pp5839ndash5844 2012
[15] U Eckhard P F Huesgen H Brandstetter and C M OverallldquoProteomic protease specificity profiling of clostridial collage-nases reveals their intrinsic nature as dedicated degraders ofcollagenrdquo Journal of Proteomics vol 100 pp 102ndash114 2014
[16] O Matsushita and A Okabe ldquoClostridial hydrolytic enzymesdegrading extracellular componentsrdquoToxicon vol 39 no 11 pp1769ndash1780 2001
[17] A Fujio K Murayama Y Yamagata et al ldquoCollagenase H iscrucial for isolation of rat pancreatic isletsrdquo Cell Transplanta-tion vol 23 no 10 pp 1187ndash1198 2014
[18] M K Gordon and R A Hahn ldquoCollagensrdquo Cell and TissueResearch vol 339 no 1 pp 247ndash257 2010
[19] MD Shoulders andR T Raines ldquoCollagen structure and stabil-ityrdquo Annual Review of Biochemistry vol 78 pp 929ndash958 2009
[20] D E Birk ldquoType V collagen heterotypic type IV collageninteractions in the regulation of fibril assemblyrdquoMicron vol 32no 3 pp 223ndash237 2001
[21] M A Weis D M Hudson L Kim M Scott J-J Wu and D REyre ldquoLocation of 3-Hydroxyproline residues in collagen typesI II III and VXI implies a role in fibril supramolecularassemblyrdquo Journal of Biological Chemistry vol 285 no 4 pp2580ndash2590 2010
[22] R J Wenstrup J B Florer E W Brunskill S M Bell I Cher-voneva and D E Birk ldquoType V collagen controls the initiationof collagen fibril assemblyrdquo Journal of Biological Chemistry vol279 no 51 pp 53331ndash53337 2004
[23] C Niyibizi P P Fietzek and M Van der Rest ldquoHumanplacenta type V collagens Evidence for the existence of an1205721(V)1205722(V)1205723(V) collagen moleculerdquo Journal of BiologicalChemistry vol 259 no 22 pp 14170ndash14174 1984
[24] C Yang A C Park N A Davis et al ldquoComprehensive massspectrometricmapping of the hydroxylated amino acid residuesof the1205721(V) collagen chainrdquoThe Journal of Biological Chemistryvol 287 no 48 pp 40598ndash40610 2012
[25] O Matsushita T Koide R Kobayashi K Nagata and AOkabe ldquoSubstrate recognition by the collagen-binding domainof Clostridium histolyticum class I collagenaserdquo The Journal ofBiological Chemistry vol 276 no 12 pp 8761ndash8770 2001
[26] M F French K A Mookhtiar and H E Van Wart ldquoLimitedproteolysis of type I collagen at hyperreactive sites by class Iand II Clostridium histolyticum collagenases complementarydigestion patternsrdquo Biochemistry vol 26 no 3 pp 681ndash6871987
[27] T Kin X Zhai T B Murdoch A Salam A M J Shapiro andJ R T Lakey ldquoEnhancing the success of human islet isolationthrough optimization and characterization of pancreas dissoci-ation enzymerdquo American Journal of Transplantation vol 7 no5 pp 1233ndash1241 2007
[28] T Kin X Zhai D OrsquoGorman and A M J Shapiro ldquoDetri-mental effect of excessive collagenase class II on human isletisolation outcomerdquo Transplant International vol 21 no 11 pp1059ndash1065 2008
[29] J H M Van Deijnen P T R Van Suylichem G H J Woltersand R Van Schilfgaarde ldquoDistribution of collagens type I typeIII and type V in the pancreas of rat dog pig andmanrdquoCell andTissue Research vol 277 no 1 pp 115ndash121 1994
[30] T F Linsenmayer E Gibney F Igoe et al ldquoType V collagenmolecular structure and fibrillar organization of the chicken1205721(V) NH2-terminal domain a putative regulator of cornealfibrillogenesisrdquo Journal of Cell Biology vol 121 no 5 pp 1181ndash1189 1993
[31] J K Mouw G Ou and V M Weaver ldquoExtracellular matrixassembly a multiscale deconstructionrdquo Nature Reviews Molec-ular Cell Biology vol 15 no 12 pp 771ndash785 2014
[32] F Delacoux A Fichard C Geourjon R Garrone and FRuggiero ldquoMolecular features of the collagenVheparin bindingsiterdquo The Journal of Biological Chemistry vol 273 no 24 pp15069ndash15076 1998
[33] S Ricard-Blum M Beraud N Raynal R W Farndale and FRuggiero ldquoStructural requirements for heparinheparan sulfatebinding to type V collagenrdquoThe Journal of Biological Chemistryvol 281 no 35 pp 25195ndash25204 2006
[34] W Henkel and K Dreisewerd ldquoCyanogen bromide peptides ofthe fibrillar collagens I III and V and their mass spectrometriccharacterization detection of linear peptides peptide glycosyla-tion and cross-linking peptides involved in formation of homo-and heterotypic fibrilsrdquo Journal of Proteome Research vol 6 no11 pp 4269ndash4289 2007
[35] K Gelse E Poschl and T Aigner ldquoCollagensmdashstructurefunction and biosynthesisrdquo Advanced Drug Delivery Reviewsvol 55 no 12 pp 1531ndash1546 2003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Journal of Diabetes Research 3
and loaded onto a 75 120583m diameter 10 cm length fused-silicacapillary column containing C18 resin (Nikkyo TechnosTokyo Japan) The peptides were eluted with an acetonitrilegradient (typically 24 to 22 in 46min then to 33in 49min) in 01 formic acid and analyzed by an LTQOrbitrap Velos mass spectrometer (Thermo Fisher ScientificInc) Database searches against the NCBInr and Swissprotdatabases were performed using the MASCOT ver 251search engine (Matrix Science London UK) The MASCOTsearches were carried out without protease specificity using5 ppm peptide mass tolerance and 05 Da MSMS toleranceOxidation (+159949Da) was considered as a variable mod-ification for Met Cys Pro and Lys The peptides identifiedwith MASCOT expectation values lt 005 were considered assignificant hits
3 Results
31 In Vitro Digestion of Col-V by Collagenases
311 SDS-PAGE Analysis of Col-V Digestion In vitro diges-tion of Col-V was performed to investigate the differences indigestion patterns by the collagenases Figure 2(a) shows anSDS-PAGE analysis of the Col-V digestion patterns by ColHand ColG from 15 to 60min The results showed distinctdifferences between the Col-V digestion patterns by ColHand ColG On the SDS-PAGE gel no band changes wereobserved in the lanes for ColHwithin 1 h while ColG showedsignificant digestion of Col-V within this time frame
312 MALDI Analysis of Col-V Digestion The SDS-PAGEanalysis showed clear differences in the Col-V digestionpatterns between ColH and ColG Therefore to sensitivelydetect the low molecular weight peptide fragments MALDI-tof-MS analyses were conducted for the digestion solutionsat 15 30 and 60min The Col-V digestion samples by ColHindicated only a few peaks aroundmz 700ndash800 which couldreflect impurities arising from thematrix or originally presentwithin the enzymeCol-V solutions (Figure 2(b)) In contrastthe samples digested by ColG exhibited many peaks aroundmz 1000ndash3000 These peaks were considered to be Col-Vpeptide fragments digested by ColG Although the individualpeak intensities could not be compared quantitatively inthe MALDI spectra the relative intensity patterns changedindicating that the digestion sites by ColG changed over time
32 Peptide Fragment Analysis by ESI-MS To investigatethe Col-V cleavage sites by ColG the digested sample at1 h was further fragmented by trypsin and analyzed byLC-MS Peptide sequences and modifications (hydroxylatedproline and lysine) were identified by MSMS analysis TheMS experiments were performed in duplicate The analyzedpeptides included trypticsemitryptic peptides (digested atthe C-terminus of LysArg) collagenolytic peptides (digestedat the N-terminus of Gly-Xaa-Yaa repeats) and other pep-tides (neither tryptic nor collagenolytic peptides) The pep-tides were categorized tryptic peptides semitryptic peptidesincludingnot including Gly-Xaa-Yaa sequences at either
205
116974
6955
365
Mar
ker
Col
-V
Col
G
Col
H
V +
H15
min
V +
H30
min
V +
H60
min
V +
G15
min
V +
G30
min
V +
G60
min
(a)
ColG 15min
ColG 30min
ColG 60min
ColH 15min
ColH 30min
ColH 60min
1000 1500 2000 2500 3000
(b)
Figure 2 In vitro Col-V digestion by ColH and ColG (a) SDS-PAGE analysis of digested Col-V (b) MALDI-tof-MS analysis of thedigested samples at 15 30 and 60minThepeak intensities are scaledto 100 for the most intense peak in each chart
side and nontryptic peptides includingnot including Gly-Xaa-Yaa sequences at either side or both sides Consideringthis categorization the total number of peptides was countedas the sum of the number of tryptic peptides + number ofall semitryptic peptides + number of all nontryptic peptidesThe collagenolytic peptides were calculated as the sum of thesemitryptic peptides and nontryptic peptides including Gly-Xaa-Yaa sequences at either side or both sidesThe results aresummarized in Table 1The total number of detected peptideswas in the order of ColGtrypsin digestion gt ColHtrypsingt trypsin It was considered that the fragmentation of Col-V proceeded by double enzyme digestions especially for thecombination of ColGtrypsin The Col-V digested by trypsinconsisted of mainly tryptic and semitryptic peptides Sometryptic peptides were shared with collagenolytic peptidesbecause the Col-V sequence included lysine or arginine atthe Yaa position of the Gly-Xaa-Yaa repeats The double-digested samples included both tryptic and collagenolytic
4 Journal of Diabetes Research
Table 1 Peptide numbers detected by ESI-MS
pep tryp semitryp nontryp col pepGxx-G non-Gxx-G Gxx-G non-Gxx-G
V1205721 29 31 20 33 1 2 8 6 0 0 0 0 3 61205722 45 38 30 25 4 6 11 7 0 0 0 0 9 161205723 17 17 15 15 0 0 2 2 0 0 0 0 0 0
V + H1205721 39 33 21 20 9 6 4 6 3 0 0 0 31 271205722 46 47 26 30 9 7 10 9 1 1 0 0 22 171205723 21 18 15 16 4 1 2 1 0 0 0 0 19 6
V + G1205721 64 56 17 15 17 16 8 5 22 20 0 0 61 641205722 71 62 22 19 28 26 7 2 14 15 0 0 59 661205723 34 24 10 7 12 9 1 1 11 7 0 0 68 67
pep tryp semitryp and nontryp indicate the number of total peptides number of tryptic peptides (RK)darrxxxsdot sdot sdot xxx(RK)darrxxxsdot sdot sdot number ofsemitryptic peptides (RK)darrxxxsdot sdot sdot or sdot sdot sdot xxx(RK)darrxxxsdot sdot sdot and number of nontryptic peptides respectivelyGxx-G is the preferred substrate sequence for collagenase at position P3-P2-P1-P11015840Type V collagen was digested by trypsin (V) trypsin + collagenase H (V + H) and trypsin + collagenase G (V + G)The percentage of collagenolytic peptides was calculated by the following equation col pep = (number of semitryptic Gxx-G + number of nontryptic Gxx-G)(number of total peptides)
peptides In the ColHtrypsin digestion the percentage ofcollagenolytic peptides was around 30 for 1205721 and 20 for1205722 In contrast the percentage of collagenolytic peptides forColGtrypsin digestion exceeded 60ndash70 for all chain typesThese results suggested that collagenolytic peptides were themain components in the ColGtrypsin digestion sample andtherefore that ColG digested Col-V
33 Initial Digestion of Col-V by ColG
331 SDS-PAGEAnalysis In this study in vitro digestionwasperformed for 15ndash60min as described above However theCol-V digestion had already started by 15min To analyze theinitial stage of the digestion the Col-V digestion experimentwas performed for a shorter time (3ndash15min) and the digestedsamples were analyzed by SDS-PAGE (Figure 3) The mainbands shifted to lower molecular weights over time Further-more below themain bands some new bands appeared suchas the band at sim95 kDa
332 Cleavage Site Distribution Analysis The above resultsindicated that Col-V was degraded by ColG and thereforethat the sample solutions within 15min included peptidefragments digested by ColG The digestion solutions at 3 69 12 and 15min were mixed in equal amounts and measuredby LC-MS The detected peptides contained a Gly-Xaa-Yaasequence at the N- or C-terminus The cleavage sites weremapped on the Col-V sequences (1205721 1205722 and 1205723) (Supple-mentary Figure 1 red triangles see Supplementary Materialavailable online at httpdxdoiorg10115520164396756)The cleavage sites were distributed in the whole region of theGly-Xaa-Yaa repeats
The initial digestionwas evaluated by analyzing the short-term digestion In this digestion experiment the shortestdigestion time was 3min The 3min digestion solutionwas subjected to LC-MS analysis and the cleavage sitesequences (containing Gly-Xaa-Yaa) were mapped on thesame sequences described above (Supplementary Figure 1
205
116974
6955
365
Mar
ker
Col
-V
Col
G3
min
Col
G6
min
Col
G9
min
Col
G12
min
Col
G15
min
Figure 3 SDS-PAGE analysis of Col-V digestion by ColG within15min
blue triangles) The cleavage sites within the 3-min digestionsample were distributed on the N-terminal side of the 1205721 and1205722 chains In this experiment Col-V from human placentawas used Human placenta contains Col-V in the form of1205721(V)1205722(V)1205723(V) [23] Thus the 1205723 chain could be detectedHowever the distribution in the 1205723 chain was not clearbecause the relative abundance of the detected peptides wasmuch lower than those of the other two chains
For the 1205721 chain the modification positions for hydroxylgroups onto proline have been reported [24] The ColGpreference for hydroxyproline (Hyp) at the Yaa position canbe calculated by (number of Hyp at the Yaa position within
Journal of Diabetes Research 5
cleavage sites)(number of cleavage sites) = 2953 = 547Meanwhile the average distribution of Hyp within the Gly-Xaa-Yaa repeats can be calculated by (number of Hyp atthe Yaa position)(number of Gly-Xaa-Yaa repeats) = 97338= 287 These results indicated that Hyp residues werepreferentially involved at theYaa position in the cleavage sites
34 Long-Term Digestion of Col-V by ColG The short-termdigestion of Col-V by ColG revealed an interesting digestionpattern Therefore we conducted a long-term digestionexperiment to investigate the digestion pattern from 1 to4 h The digestion samples were analyzed by SDS-PAGE(Figure 4(a)) On the SDS-PAGE gel the main bands atgt120 kDa disappeared and several bands appeared in thelower molecular weight region These bands were numbered1ndash8 Of these bands the intensities of bands 1ndash6 decreasedover time (Figure 4(b)) Meanwhile the lower molecularweight bands (7 and 8) increased in intensity over timeAlthough the cleavage sites cannot be identified by SDS-PAGE analysis the appearance of distinct bands indicatedthe existence of region selectivity for ColG digestion withinthe Gly-Xaa-Yaa repeats of Col-V Furthermore the newlyappearing bands also decreased in the long-term diges-tion suggesting that there were secondary digestions of thedigested fragments
35 Tissue Digestion and MS Analysis
351 Porcine Pancreatic Tissue Digestion During pancre-atic islet isolation the exogenous enzymes used are ColHColG and thermolysin [1] Of these enzymes thermolysindoes not digest collagens under physiological conditions(manuscript in preparation) The porcine pancreatic tissuedigestion experiments were carried out in two steps Thefirst step involved digestion by thermolysin and the secondstep involved digestion by ColH ColG or ColH + ColG(double digestion) These two-step digestions had significanteffects on detecting the collagen peptides in the subsequentmass spectrometric analyses because the digested peptidesby thermolysin could be removed from the solution beforethe second digestion Figure 5 shows the observed pancreatictissue digestion patterns over time The tissue samples werenot digested by thermolysin alone To exclude any influenceof the proteins digested by thermolysin the sample solu-tions were changed after the thermolysin digestion and theobservations were started after addition of the collagenasesInterestingly the pancreatic tissues were not digested byColH or ColG alone Only the solution including both ColHand ColG exhibited tissue degradation These observationsare consistent with our previous findings that both ColGand ColH are necessary for dissociating porcine pancreatictissues Although the observations were carried out until 9 hall three tissue pieces were digested within 3 h
352 Col-V Fragments Produced by ColG Digestion of Pancre-atic Tissue The digested samples by ColH and ColG wereapplied to LC-MS analysis Database searches focused onidentifying collagen peptides were carried outThe identifiedcollagen peptides were picked up from the search results
205
116974
69
55
365
ColGlowast1
lowast2
lowast3
lowast4
lowast5
lowast6
lowast7
lowast8
Col
-V
Mar
ker
V +
G1
hr
V +
G2
hrs
V +
G4
hrs
(a)
ColGColG ColG
Inte
nsity
(au
)
lowast1
lowast1
lowast1
lowast2
lowast2
lowast2
lowast3 lowast
3
lowast3
lowast4
lowast4
lowast4
lowast5
lowast5
lowast5
lowast6
lowast6
lowast6
lowast7
lowast7
lowast7
lowast8
lowast8
lowast8
V + G 1hr V + G 2hrs V + G 4hrs
(b)
Figure 4 SDS-PAGE analysis of Col-V digestion by ColG over along time period (sim4 h) (a) SDS-PAGE analysis of Col-V digestionsColG and the main bands are numbered (b) Band intensity analysisby densitometry
and are summarized in Table 2 Most of the identifiedpeptides were trypticsemitryptic peptides Col-V peptideswere hardly detected and their sequences were shared bytrypticsemitryptic peptides It was speculated that Col-Vdigestion proceeded to unsuitably small fragments in the
6 Journal of Diabetes Research
Col
G
n2
n1
n3
n2
n1
n3
n2
n1
n3
Col
HC
olH
+ G
0hr 3hrs 6hrs 9hrs
Figure 5 In vitro porcine pancreatic tissue digestions by proteasesThe digestions were observed at 0 3 6 and 9 h Each digestion wasconducted in triplicate
presence of ColG together with other endogenous proteasesfor detection of the mass spectrometric analyses
4 Discussion
Our in vitro digestion experiments revealed that the Col-Vdigestion patterns were distinctly different betweenColH andColG
Col-V digestion by ColG was clearly shown by SDS-PAGE and mass spectrometric analyses Meanwhile ColHdid not digest Col-V under the same conditions used forColGAlthough this study could not investigate the differencein binding ability toward Col-V between ColH and ColG itis considered that the difference in domain construction is
Table 2 Collagen peptide identification from tissue digestion sam-ples
Col type tryp + semitryp nontryp Gxx-G
Col-V 1205721 2 0 0 21205722 4 0 0 3
Col-IV 1205721 1 2 3 21205722 1 1 1 1
Col-VI1205721 5 8 0 01205722 9 1 1 01205723 3 2 1 0
tryp semitryp and nontryp indicate the number of tryptic peptides (RK)darrxxxsdot sdot sdot xxx(RK)darrxxxsdot sdot sdot number of semitryptic peptides (RK)darrxxxsdot sdot sdotor sdot sdot sdot xxx(RK)darrxxxsdot sdot sdot and number of nontryptic peptides respectivelyThe total number of detected peptides is shown by tryp + semitryp + nontryp Gxx-G can be shared in tryp semitryp and nontrypDatabase searches for porcine collagens were based on the following NCBIaccession numbers 1205721(V) 62461592 1205722(V) 157427707 1205721(IV) 9271916811205722(IV) 927191683 1205721(VI) 92020086 92020094 and 545895133 1205722(VI)927203606 1205723(VI) 92020131 and 927211912
one of the possible reasons for the different collagenolyticactivities of ColH and ColG for Col-V ColH has onlyone CBD while ColG contains tandem CBDs (Figure 1)The CBDs of clostridial collagenases have been investigatedin detail A structural comparison indicated that the twostructures were similar (rmsd for C120572 = 15 A) despite therelatively low sequence similarity [11] The substrate-bindingspecificity of CBDs is broad for collagen types IndashIV [6] TheCBDs of ColH and ColG can bind to triple helical collagensbut not to telopeptides [5 25] However tandem CDBs canbind more efficiently to Col-I than a monomeric CBD [25]Consequently the domain construction especially the CBDarrangement plays a very important role in the digestioncharacteristics for Col-V
In the present study Col-V digestion by ColG produceddistinct digestion characteristics within a short time period(within minutes) and a long time period (within hours)The mass spectrometric analyses revealed that the identifiedpeptide cleavage sites were distributed in the N-terminalregion of theGly-Xaa-Yaa repeat region of Col-VThis findingsuggests that the digestion of Col-V is initiated from the N-terminal region According to the ColG digestion for Col-I [26] Col-I is hydrolyzed near the C-terminal end of thetriple helix region and a second cleavage occurs at the N-terminal side Although a digestion tendency for the C-terminal side was not found in our results digestion fromthe N-terminal region is a common digestion property ofColG for both Col-I and Col-V The long-term digestionexperiment indicated several residual parts of Col-V thatappeared as distinct bands at 1 h on SDS-PAGE These partswere also hydrolyzed by subsequent digestion (sim4 h) It issuggested that the Col-V sequence includes some parts thatare resistant to collagenolysis
The tissue digestion experiments clearly showed that bothColH and ColG were needed to degrade the pancreatic tissuestructure For human tissue degradation it was reported thatthe ratio of ColGH influenced the islet isolation outcome
Journal of Diabetes Research 7
[27 28] These results indicate that ColG may play moreimportant roles in digestion procedure of pig and humantissues than that of rat tissues The mass spectrometricanalysis for the digested samples identified collagen peptidesof Col-IV Col-V and collagen type VI Our previous studyrevealed that ColH is crucial for rat pancreatic islet isolationand the most abundantly detected collagen peptides in themass spectrometric analysis were Col-I and Col-III [17]These results strongly suggested the presence of markeddifferences between rat and porcine pancreatic tissue degra-dations Although it is difficult to identify the exact reasonfor the discrepancies fibril formation could be a possibleexplanation Collagen molecules are differently involved invarious tissues according to animal species [29] Col-V isassociated with Col-I in the microfibrillar structure [3031] Furthermore Col-V can be involved in heparinheparinsulfate modifications [32 33] and glycosylation [34] aswell as the well-known hydroxyl group modifications atproline and lysine residues [35] Therefore the differences incircumstances of the Col-IV fibrillar structures in the ratand porcine pancreatic tissue may influence the pancreaticislet isolation efficiency Although the similarities of thecircumstances of the Col-IV fibrillar structures among ratspigs and humans for pancreatic islet isolation are difficult tocompare ColG appears to be more important in porcine andhuman islet isolation processes and Col-V may be one of thekey substrates for ColG
5 Conclusions
In this study we focused on Col-V digestion by clostridialcollagenases Col-V was distinctively digested by ColG Massspectrometric analyses revealed that Col-V digestion byColG proceeded from the N-terminal region Although bothclostridial collagenases ColH and ColG include commondomains such as the activator domain peptidase domainPKD and CBD their domain construction is different Thedomain construction may play a crucial role for determiningthe collagen digestion patterns by the clostridial collagenasesThe porcine pancreatic tissue digestion experiments demon-strated that both collagenases are needed to degrade the tissuestructure These observations are completely different fromthose in rat experiments in which ColG was not cruciallyneeded Consequently the present study reveals that ColG aswell as ColH plays a crucial role in pancreatic islet isolationin large mammals and that Col-V can be one of the keysubstrates for ColG
Competing Interests
The authors of this paper received partial financial supportfromMeiji Seika PharmaCo Ltd and ShimadzuCorporationaccording to a collaborative research contract
Authorsrsquo Contributions
Kazutaka Murayama designed the experiments performedthe sample digestion andMALDI-tof-MS analyses andwrote
the paper Akiko Inagaki and Takehiro Imura performed thetissue digestion experiments Hiroki Shima performed theLC-MS spectrometric analyses and wrote part of the paperYouhei Yamagata and Kimiko Watanabe participated in thepaper corrections and critical review Kazuhiko Igarashi andMasafumi Goto participated in discussions during the paperpreparation and paper corrections
Acknowledgments
The authors acknowledge the support of the BiomedicalResearch Core and TAMRIC (Tohoku Advanced MedicalResearch and IncubationCenter) of TohokuUniversityGrad-uate School of Medicine Part of this research was supportedby the Tohoku University Business Incubation Program
References
[1] R C McCarthy A G Breite M L Green and F E DwuletldquoTissue dissociation enzymes for isolating human islets fortransplantation Factors to consider in setting enzyme accep-tance criteriardquo Transplantation vol 91 no 2 pp 137ndash145 2011
[2] K Watanabe ldquoCollagenolytic proteases from bacteriardquo AppliedMicrobiology and Biotechnology vol 63 no 5 pp 520ndash5262004
[3] U Eckhard E Schonauer P Ducka P Briza D Nuss andH Brandstetter ldquoBiochemical characterization of the catalyticdomains of three different clostridial collagenasesrdquo BiologicalChemistry vol 390 no 1 pp 11ndash18 2009
[4] C-M Jung O Matsushita S Katayama J Minami J Sakuraiand A Okabe ldquoIdentification of metal ligands in the Clostrid-ium histolyticumColH collagenaserdquo Journal of Bacteriology vol181 no 9 pp 2816ndash2822 1999
[5] O Matsushita C-M Jung J Minami S Katayama N Nishiand A Okabe ldquoA study of the collagen-binding domain of a116-kDa Clostridium histolyticum collagenaserdquo The Journal ofBiological Chemistry vol 273 no 6 pp 3643ndash3648 1998
[6] T Toyoshima O Matsushita J Minami N Nishi A Okabeand T Itano ldquoCollagen-binding domain of a Clostridium his-tolyticum collagenase exhibits a broad substrate spectrum bothin vitro and in vivordquo Connective Tissue Research vol 42 no 4pp 281ndash290 2001
[7] U Eckhard E Schonauer D Nuss and H Brandstetter ldquoStruc-ture of collagenase G reveals a chew-and-digest mechanismof bacterial collagenolysisrdquo Nature Structural and MolecularBiology vol 18 no 10 pp 1109ndash1114 2010
[8] U Eckhard E Schonauer andH Brandstetter ldquoStructural basisfor activity regulation and substrate preference of clostridialcollagenases G H and Trdquo The Journal of Biological Chemistryvol 288 no 28 pp 20184ndash20194 2013
[9] R Bauer K Janowska K Taylor et al ldquoStructures of threepolycystic kidney disease-like domains from Clostridium his-tolyticum collagenases ColG and ColHrdquo Acta CrystallographicaSection D Biological Crystallography vol 71 pp 565ndash577 2015
[10] U Eckhard and H Brandstetter ldquoPolycystic kidney disease-likedomains of clostridial collagenases and their role in collagenrecruitmentrdquo Biological Chemistry vol 392 no 11 pp 1039ndash1045 2011
8 Journal of Diabetes Research
[11] R Bauer J J Wilson S T L Philominathan D Davis OMatsushita and J Sakona ldquoStructural comparison of ColHand ColG collagen-binding domains from Clostridium his-tolyticumrdquo Journal of Bacteriology vol 195 no 2 pp 318ndash3272013
[12] J J Wilson O Matsushita A Okabe and J Sakon ldquoA bacterialcollagen-binding domain with novel calcium-binding motifcontrols domain orientationrdquoThe EMBO Journal vol 22 no 8pp 1743ndash1752 2003
[13] N Ohbayashi T Matsumoto H Shima et al ldquoSolution struc-ture of clostridial collagenase H and its calcium-dependentglobal conformation changerdquo Biophysical Journal vol 104 no7 pp 1538ndash1545 2013
[14] N Ohbayashi N Yamagata M Goto K Watanabe Y Yam-agata and K Murayama ldquoEnhancement of the structuralstability of full-length clostridial collagenase by calcium ionsrdquoApplied and Environmental Microbiology vol 78 no 16 pp5839ndash5844 2012
[15] U Eckhard P F Huesgen H Brandstetter and C M OverallldquoProteomic protease specificity profiling of clostridial collage-nases reveals their intrinsic nature as dedicated degraders ofcollagenrdquo Journal of Proteomics vol 100 pp 102ndash114 2014
[16] O Matsushita and A Okabe ldquoClostridial hydrolytic enzymesdegrading extracellular componentsrdquoToxicon vol 39 no 11 pp1769ndash1780 2001
[17] A Fujio K Murayama Y Yamagata et al ldquoCollagenase H iscrucial for isolation of rat pancreatic isletsrdquo Cell Transplanta-tion vol 23 no 10 pp 1187ndash1198 2014
[18] M K Gordon and R A Hahn ldquoCollagensrdquo Cell and TissueResearch vol 339 no 1 pp 247ndash257 2010
[19] MD Shoulders andR T Raines ldquoCollagen structure and stabil-ityrdquo Annual Review of Biochemistry vol 78 pp 929ndash958 2009
[20] D E Birk ldquoType V collagen heterotypic type IV collageninteractions in the regulation of fibril assemblyrdquoMicron vol 32no 3 pp 223ndash237 2001
[21] M A Weis D M Hudson L Kim M Scott J-J Wu and D REyre ldquoLocation of 3-Hydroxyproline residues in collagen typesI II III and VXI implies a role in fibril supramolecularassemblyrdquo Journal of Biological Chemistry vol 285 no 4 pp2580ndash2590 2010
[22] R J Wenstrup J B Florer E W Brunskill S M Bell I Cher-voneva and D E Birk ldquoType V collagen controls the initiationof collagen fibril assemblyrdquo Journal of Biological Chemistry vol279 no 51 pp 53331ndash53337 2004
[23] C Niyibizi P P Fietzek and M Van der Rest ldquoHumanplacenta type V collagens Evidence for the existence of an1205721(V)1205722(V)1205723(V) collagen moleculerdquo Journal of BiologicalChemistry vol 259 no 22 pp 14170ndash14174 1984
[24] C Yang A C Park N A Davis et al ldquoComprehensive massspectrometricmapping of the hydroxylated amino acid residuesof the1205721(V) collagen chainrdquoThe Journal of Biological Chemistryvol 287 no 48 pp 40598ndash40610 2012
[25] O Matsushita T Koide R Kobayashi K Nagata and AOkabe ldquoSubstrate recognition by the collagen-binding domainof Clostridium histolyticum class I collagenaserdquo The Journal ofBiological Chemistry vol 276 no 12 pp 8761ndash8770 2001
[26] M F French K A Mookhtiar and H E Van Wart ldquoLimitedproteolysis of type I collagen at hyperreactive sites by class Iand II Clostridium histolyticum collagenases complementarydigestion patternsrdquo Biochemistry vol 26 no 3 pp 681ndash6871987
[27] T Kin X Zhai T B Murdoch A Salam A M J Shapiro andJ R T Lakey ldquoEnhancing the success of human islet isolationthrough optimization and characterization of pancreas dissoci-ation enzymerdquo American Journal of Transplantation vol 7 no5 pp 1233ndash1241 2007
[28] T Kin X Zhai D OrsquoGorman and A M J Shapiro ldquoDetri-mental effect of excessive collagenase class II on human isletisolation outcomerdquo Transplant International vol 21 no 11 pp1059ndash1065 2008
[29] J H M Van Deijnen P T R Van Suylichem G H J Woltersand R Van Schilfgaarde ldquoDistribution of collagens type I typeIII and type V in the pancreas of rat dog pig andmanrdquoCell andTissue Research vol 277 no 1 pp 115ndash121 1994
[30] T F Linsenmayer E Gibney F Igoe et al ldquoType V collagenmolecular structure and fibrillar organization of the chicken1205721(V) NH2-terminal domain a putative regulator of cornealfibrillogenesisrdquo Journal of Cell Biology vol 121 no 5 pp 1181ndash1189 1993
[31] J K Mouw G Ou and V M Weaver ldquoExtracellular matrixassembly a multiscale deconstructionrdquo Nature Reviews Molec-ular Cell Biology vol 15 no 12 pp 771ndash785 2014
[32] F Delacoux A Fichard C Geourjon R Garrone and FRuggiero ldquoMolecular features of the collagenVheparin bindingsiterdquo The Journal of Biological Chemistry vol 273 no 24 pp15069ndash15076 1998
[33] S Ricard-Blum M Beraud N Raynal R W Farndale and FRuggiero ldquoStructural requirements for heparinheparan sulfatebinding to type V collagenrdquoThe Journal of Biological Chemistryvol 281 no 35 pp 25195ndash25204 2006
[34] W Henkel and K Dreisewerd ldquoCyanogen bromide peptides ofthe fibrillar collagens I III and V and their mass spectrometriccharacterization detection of linear peptides peptide glycosyla-tion and cross-linking peptides involved in formation of homo-and heterotypic fibrilsrdquo Journal of Proteome Research vol 6 no11 pp 4269ndash4289 2007
[35] K Gelse E Poschl and T Aigner ldquoCollagensmdashstructurefunction and biosynthesisrdquo Advanced Drug Delivery Reviewsvol 55 no 12 pp 1531ndash1546 2003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

4 Journal of Diabetes Research
Table 1 Peptide numbers detected by ESI-MS
pep tryp semitryp nontryp col pepGxx-G non-Gxx-G Gxx-G non-Gxx-G
V1205721 29 31 20 33 1 2 8 6 0 0 0 0 3 61205722 45 38 30 25 4 6 11 7 0 0 0 0 9 161205723 17 17 15 15 0 0 2 2 0 0 0 0 0 0
V + H1205721 39 33 21 20 9 6 4 6 3 0 0 0 31 271205722 46 47 26 30 9 7 10 9 1 1 0 0 22 171205723 21 18 15 16 4 1 2 1 0 0 0 0 19 6
V + G1205721 64 56 17 15 17 16 8 5 22 20 0 0 61 641205722 71 62 22 19 28 26 7 2 14 15 0 0 59 661205723 34 24 10 7 12 9 1 1 11 7 0 0 68 67
pep tryp semitryp and nontryp indicate the number of total peptides number of tryptic peptides (RK)darrxxxsdot sdot sdot xxx(RK)darrxxxsdot sdot sdot number ofsemitryptic peptides (RK)darrxxxsdot sdot sdot or sdot sdot sdot xxx(RK)darrxxxsdot sdot sdot and number of nontryptic peptides respectivelyGxx-G is the preferred substrate sequence for collagenase at position P3-P2-P1-P11015840Type V collagen was digested by trypsin (V) trypsin + collagenase H (V + H) and trypsin + collagenase G (V + G)The percentage of collagenolytic peptides was calculated by the following equation col pep = (number of semitryptic Gxx-G + number of nontryptic Gxx-G)(number of total peptides)
peptides In the ColHtrypsin digestion the percentage ofcollagenolytic peptides was around 30 for 1205721 and 20 for1205722 In contrast the percentage of collagenolytic peptides forColGtrypsin digestion exceeded 60ndash70 for all chain typesThese results suggested that collagenolytic peptides were themain components in the ColGtrypsin digestion sample andtherefore that ColG digested Col-V
33 Initial Digestion of Col-V by ColG
331 SDS-PAGEAnalysis In this study in vitro digestionwasperformed for 15ndash60min as described above However theCol-V digestion had already started by 15min To analyze theinitial stage of the digestion the Col-V digestion experimentwas performed for a shorter time (3ndash15min) and the digestedsamples were analyzed by SDS-PAGE (Figure 3) The mainbands shifted to lower molecular weights over time Further-more below themain bands some new bands appeared suchas the band at sim95 kDa
332 Cleavage Site Distribution Analysis The above resultsindicated that Col-V was degraded by ColG and thereforethat the sample solutions within 15min included peptidefragments digested by ColG The digestion solutions at 3 69 12 and 15min were mixed in equal amounts and measuredby LC-MS The detected peptides contained a Gly-Xaa-Yaasequence at the N- or C-terminus The cleavage sites weremapped on the Col-V sequences (1205721 1205722 and 1205723) (Supple-mentary Figure 1 red triangles see Supplementary Materialavailable online at httpdxdoiorg10115520164396756)The cleavage sites were distributed in the whole region of theGly-Xaa-Yaa repeats
The initial digestionwas evaluated by analyzing the short-term digestion In this digestion experiment the shortestdigestion time was 3min The 3min digestion solutionwas subjected to LC-MS analysis and the cleavage sitesequences (containing Gly-Xaa-Yaa) were mapped on thesame sequences described above (Supplementary Figure 1
205
116974
6955
365
Mar
ker
Col
-V
Col
G3
min
Col
G6
min
Col
G9
min
Col
G12
min
Col
G15
min
Figure 3 SDS-PAGE analysis of Col-V digestion by ColG within15min
blue triangles) The cleavage sites within the 3-min digestionsample were distributed on the N-terminal side of the 1205721 and1205722 chains In this experiment Col-V from human placentawas used Human placenta contains Col-V in the form of1205721(V)1205722(V)1205723(V) [23] Thus the 1205723 chain could be detectedHowever the distribution in the 1205723 chain was not clearbecause the relative abundance of the detected peptides wasmuch lower than those of the other two chains
For the 1205721 chain the modification positions for hydroxylgroups onto proline have been reported [24] The ColGpreference for hydroxyproline (Hyp) at the Yaa position canbe calculated by (number of Hyp at the Yaa position within
Journal of Diabetes Research 5
cleavage sites)(number of cleavage sites) = 2953 = 547Meanwhile the average distribution of Hyp within the Gly-Xaa-Yaa repeats can be calculated by (number of Hyp atthe Yaa position)(number of Gly-Xaa-Yaa repeats) = 97338= 287 These results indicated that Hyp residues werepreferentially involved at theYaa position in the cleavage sites
34 Long-Term Digestion of Col-V by ColG The short-termdigestion of Col-V by ColG revealed an interesting digestionpattern Therefore we conducted a long-term digestionexperiment to investigate the digestion pattern from 1 to4 h The digestion samples were analyzed by SDS-PAGE(Figure 4(a)) On the SDS-PAGE gel the main bands atgt120 kDa disappeared and several bands appeared in thelower molecular weight region These bands were numbered1ndash8 Of these bands the intensities of bands 1ndash6 decreasedover time (Figure 4(b)) Meanwhile the lower molecularweight bands (7 and 8) increased in intensity over timeAlthough the cleavage sites cannot be identified by SDS-PAGE analysis the appearance of distinct bands indicatedthe existence of region selectivity for ColG digestion withinthe Gly-Xaa-Yaa repeats of Col-V Furthermore the newlyappearing bands also decreased in the long-term diges-tion suggesting that there were secondary digestions of thedigested fragments
35 Tissue Digestion and MS Analysis
351 Porcine Pancreatic Tissue Digestion During pancre-atic islet isolation the exogenous enzymes used are ColHColG and thermolysin [1] Of these enzymes thermolysindoes not digest collagens under physiological conditions(manuscript in preparation) The porcine pancreatic tissuedigestion experiments were carried out in two steps Thefirst step involved digestion by thermolysin and the secondstep involved digestion by ColH ColG or ColH + ColG(double digestion) These two-step digestions had significanteffects on detecting the collagen peptides in the subsequentmass spectrometric analyses because the digested peptidesby thermolysin could be removed from the solution beforethe second digestion Figure 5 shows the observed pancreatictissue digestion patterns over time The tissue samples werenot digested by thermolysin alone To exclude any influenceof the proteins digested by thermolysin the sample solu-tions were changed after the thermolysin digestion and theobservations were started after addition of the collagenasesInterestingly the pancreatic tissues were not digested byColH or ColG alone Only the solution including both ColHand ColG exhibited tissue degradation These observationsare consistent with our previous findings that both ColGand ColH are necessary for dissociating porcine pancreatictissues Although the observations were carried out until 9 hall three tissue pieces were digested within 3 h
352 Col-V Fragments Produced by ColG Digestion of Pancre-atic Tissue The digested samples by ColH and ColG wereapplied to LC-MS analysis Database searches focused onidentifying collagen peptides were carried outThe identifiedcollagen peptides were picked up from the search results
205
116974
69
55
365
ColGlowast1
lowast2
lowast3
lowast4
lowast5
lowast6
lowast7
lowast8
Col
-V
Mar
ker
V +
G1
hr
V +
G2
hrs
V +
G4
hrs
(a)
ColGColG ColG
Inte
nsity
(au
)
lowast1
lowast1
lowast1
lowast2
lowast2
lowast2
lowast3 lowast
3
lowast3
lowast4
lowast4
lowast4
lowast5
lowast5
lowast5
lowast6
lowast6
lowast6
lowast7
lowast7
lowast7
lowast8
lowast8
lowast8
V + G 1hr V + G 2hrs V + G 4hrs
(b)
Figure 4 SDS-PAGE analysis of Col-V digestion by ColG over along time period (sim4 h) (a) SDS-PAGE analysis of Col-V digestionsColG and the main bands are numbered (b) Band intensity analysisby densitometry
and are summarized in Table 2 Most of the identifiedpeptides were trypticsemitryptic peptides Col-V peptideswere hardly detected and their sequences were shared bytrypticsemitryptic peptides It was speculated that Col-Vdigestion proceeded to unsuitably small fragments in the
6 Journal of Diabetes Research
Col
G
n2
n1
n3
n2
n1
n3
n2
n1
n3
Col
HC
olH
+ G
0hr 3hrs 6hrs 9hrs
Figure 5 In vitro porcine pancreatic tissue digestions by proteasesThe digestions were observed at 0 3 6 and 9 h Each digestion wasconducted in triplicate
presence of ColG together with other endogenous proteasesfor detection of the mass spectrometric analyses
4 Discussion
Our in vitro digestion experiments revealed that the Col-Vdigestion patterns were distinctly different betweenColH andColG
Col-V digestion by ColG was clearly shown by SDS-PAGE and mass spectrometric analyses Meanwhile ColHdid not digest Col-V under the same conditions used forColGAlthough this study could not investigate the differencein binding ability toward Col-V between ColH and ColG itis considered that the difference in domain construction is
Table 2 Collagen peptide identification from tissue digestion sam-ples
Col type tryp + semitryp nontryp Gxx-G
Col-V 1205721 2 0 0 21205722 4 0 0 3
Col-IV 1205721 1 2 3 21205722 1 1 1 1
Col-VI1205721 5 8 0 01205722 9 1 1 01205723 3 2 1 0
tryp semitryp and nontryp indicate the number of tryptic peptides (RK)darrxxxsdot sdot sdot xxx(RK)darrxxxsdot sdot sdot number of semitryptic peptides (RK)darrxxxsdot sdot sdotor sdot sdot sdot xxx(RK)darrxxxsdot sdot sdot and number of nontryptic peptides respectivelyThe total number of detected peptides is shown by tryp + semitryp + nontryp Gxx-G can be shared in tryp semitryp and nontrypDatabase searches for porcine collagens were based on the following NCBIaccession numbers 1205721(V) 62461592 1205722(V) 157427707 1205721(IV) 9271916811205722(IV) 927191683 1205721(VI) 92020086 92020094 and 545895133 1205722(VI)927203606 1205723(VI) 92020131 and 927211912
one of the possible reasons for the different collagenolyticactivities of ColH and ColG for Col-V ColH has onlyone CBD while ColG contains tandem CBDs (Figure 1)The CBDs of clostridial collagenases have been investigatedin detail A structural comparison indicated that the twostructures were similar (rmsd for C120572 = 15 A) despite therelatively low sequence similarity [11] The substrate-bindingspecificity of CBDs is broad for collagen types IndashIV [6] TheCBDs of ColH and ColG can bind to triple helical collagensbut not to telopeptides [5 25] However tandem CDBs canbind more efficiently to Col-I than a monomeric CBD [25]Consequently the domain construction especially the CBDarrangement plays a very important role in the digestioncharacteristics for Col-V
In the present study Col-V digestion by ColG produceddistinct digestion characteristics within a short time period(within minutes) and a long time period (within hours)The mass spectrometric analyses revealed that the identifiedpeptide cleavage sites were distributed in the N-terminalregion of theGly-Xaa-Yaa repeat region of Col-VThis findingsuggests that the digestion of Col-V is initiated from the N-terminal region According to the ColG digestion for Col-I [26] Col-I is hydrolyzed near the C-terminal end of thetriple helix region and a second cleavage occurs at the N-terminal side Although a digestion tendency for the C-terminal side was not found in our results digestion fromthe N-terminal region is a common digestion property ofColG for both Col-I and Col-V The long-term digestionexperiment indicated several residual parts of Col-V thatappeared as distinct bands at 1 h on SDS-PAGE These partswere also hydrolyzed by subsequent digestion (sim4 h) It issuggested that the Col-V sequence includes some parts thatare resistant to collagenolysis
The tissue digestion experiments clearly showed that bothColH and ColG were needed to degrade the pancreatic tissuestructure For human tissue degradation it was reported thatthe ratio of ColGH influenced the islet isolation outcome
Journal of Diabetes Research 7
[27 28] These results indicate that ColG may play moreimportant roles in digestion procedure of pig and humantissues than that of rat tissues The mass spectrometricanalysis for the digested samples identified collagen peptidesof Col-IV Col-V and collagen type VI Our previous studyrevealed that ColH is crucial for rat pancreatic islet isolationand the most abundantly detected collagen peptides in themass spectrometric analysis were Col-I and Col-III [17]These results strongly suggested the presence of markeddifferences between rat and porcine pancreatic tissue degra-dations Although it is difficult to identify the exact reasonfor the discrepancies fibril formation could be a possibleexplanation Collagen molecules are differently involved invarious tissues according to animal species [29] Col-V isassociated with Col-I in the microfibrillar structure [3031] Furthermore Col-V can be involved in heparinheparinsulfate modifications [32 33] and glycosylation [34] aswell as the well-known hydroxyl group modifications atproline and lysine residues [35] Therefore the differences incircumstances of the Col-IV fibrillar structures in the ratand porcine pancreatic tissue may influence the pancreaticislet isolation efficiency Although the similarities of thecircumstances of the Col-IV fibrillar structures among ratspigs and humans for pancreatic islet isolation are difficult tocompare ColG appears to be more important in porcine andhuman islet isolation processes and Col-V may be one of thekey substrates for ColG
5 Conclusions
In this study we focused on Col-V digestion by clostridialcollagenases Col-V was distinctively digested by ColG Massspectrometric analyses revealed that Col-V digestion byColG proceeded from the N-terminal region Although bothclostridial collagenases ColH and ColG include commondomains such as the activator domain peptidase domainPKD and CBD their domain construction is different Thedomain construction may play a crucial role for determiningthe collagen digestion patterns by the clostridial collagenasesThe porcine pancreatic tissue digestion experiments demon-strated that both collagenases are needed to degrade the tissuestructure These observations are completely different fromthose in rat experiments in which ColG was not cruciallyneeded Consequently the present study reveals that ColG aswell as ColH plays a crucial role in pancreatic islet isolationin large mammals and that Col-V can be one of the keysubstrates for ColG
Competing Interests
The authors of this paper received partial financial supportfromMeiji Seika PharmaCo Ltd and ShimadzuCorporationaccording to a collaborative research contract
Authorsrsquo Contributions
Kazutaka Murayama designed the experiments performedthe sample digestion andMALDI-tof-MS analyses andwrote
the paper Akiko Inagaki and Takehiro Imura performed thetissue digestion experiments Hiroki Shima performed theLC-MS spectrometric analyses and wrote part of the paperYouhei Yamagata and Kimiko Watanabe participated in thepaper corrections and critical review Kazuhiko Igarashi andMasafumi Goto participated in discussions during the paperpreparation and paper corrections
Acknowledgments
The authors acknowledge the support of the BiomedicalResearch Core and TAMRIC (Tohoku Advanced MedicalResearch and IncubationCenter) of TohokuUniversityGrad-uate School of Medicine Part of this research was supportedby the Tohoku University Business Incubation Program
References
[1] R C McCarthy A G Breite M L Green and F E DwuletldquoTissue dissociation enzymes for isolating human islets fortransplantation Factors to consider in setting enzyme accep-tance criteriardquo Transplantation vol 91 no 2 pp 137ndash145 2011
[2] K Watanabe ldquoCollagenolytic proteases from bacteriardquo AppliedMicrobiology and Biotechnology vol 63 no 5 pp 520ndash5262004
[3] U Eckhard E Schonauer P Ducka P Briza D Nuss andH Brandstetter ldquoBiochemical characterization of the catalyticdomains of three different clostridial collagenasesrdquo BiologicalChemistry vol 390 no 1 pp 11ndash18 2009
[4] C-M Jung O Matsushita S Katayama J Minami J Sakuraiand A Okabe ldquoIdentification of metal ligands in the Clostrid-ium histolyticumColH collagenaserdquo Journal of Bacteriology vol181 no 9 pp 2816ndash2822 1999
[5] O Matsushita C-M Jung J Minami S Katayama N Nishiand A Okabe ldquoA study of the collagen-binding domain of a116-kDa Clostridium histolyticum collagenaserdquo The Journal ofBiological Chemistry vol 273 no 6 pp 3643ndash3648 1998
[6] T Toyoshima O Matsushita J Minami N Nishi A Okabeand T Itano ldquoCollagen-binding domain of a Clostridium his-tolyticum collagenase exhibits a broad substrate spectrum bothin vitro and in vivordquo Connective Tissue Research vol 42 no 4pp 281ndash290 2001
[7] U Eckhard E Schonauer D Nuss and H Brandstetter ldquoStruc-ture of collagenase G reveals a chew-and-digest mechanismof bacterial collagenolysisrdquo Nature Structural and MolecularBiology vol 18 no 10 pp 1109ndash1114 2010
[8] U Eckhard E Schonauer andH Brandstetter ldquoStructural basisfor activity regulation and substrate preference of clostridialcollagenases G H and Trdquo The Journal of Biological Chemistryvol 288 no 28 pp 20184ndash20194 2013
[9] R Bauer K Janowska K Taylor et al ldquoStructures of threepolycystic kidney disease-like domains from Clostridium his-tolyticum collagenases ColG and ColHrdquo Acta CrystallographicaSection D Biological Crystallography vol 71 pp 565ndash577 2015
[10] U Eckhard and H Brandstetter ldquoPolycystic kidney disease-likedomains of clostridial collagenases and their role in collagenrecruitmentrdquo Biological Chemistry vol 392 no 11 pp 1039ndash1045 2011
8 Journal of Diabetes Research
[11] R Bauer J J Wilson S T L Philominathan D Davis OMatsushita and J Sakona ldquoStructural comparison of ColHand ColG collagen-binding domains from Clostridium his-tolyticumrdquo Journal of Bacteriology vol 195 no 2 pp 318ndash3272013
[12] J J Wilson O Matsushita A Okabe and J Sakon ldquoA bacterialcollagen-binding domain with novel calcium-binding motifcontrols domain orientationrdquoThe EMBO Journal vol 22 no 8pp 1743ndash1752 2003
[13] N Ohbayashi T Matsumoto H Shima et al ldquoSolution struc-ture of clostridial collagenase H and its calcium-dependentglobal conformation changerdquo Biophysical Journal vol 104 no7 pp 1538ndash1545 2013
[14] N Ohbayashi N Yamagata M Goto K Watanabe Y Yam-agata and K Murayama ldquoEnhancement of the structuralstability of full-length clostridial collagenase by calcium ionsrdquoApplied and Environmental Microbiology vol 78 no 16 pp5839ndash5844 2012
[15] U Eckhard P F Huesgen H Brandstetter and C M OverallldquoProteomic protease specificity profiling of clostridial collage-nases reveals their intrinsic nature as dedicated degraders ofcollagenrdquo Journal of Proteomics vol 100 pp 102ndash114 2014
[16] O Matsushita and A Okabe ldquoClostridial hydrolytic enzymesdegrading extracellular componentsrdquoToxicon vol 39 no 11 pp1769ndash1780 2001
[17] A Fujio K Murayama Y Yamagata et al ldquoCollagenase H iscrucial for isolation of rat pancreatic isletsrdquo Cell Transplanta-tion vol 23 no 10 pp 1187ndash1198 2014
[18] M K Gordon and R A Hahn ldquoCollagensrdquo Cell and TissueResearch vol 339 no 1 pp 247ndash257 2010
[19] MD Shoulders andR T Raines ldquoCollagen structure and stabil-ityrdquo Annual Review of Biochemistry vol 78 pp 929ndash958 2009
[20] D E Birk ldquoType V collagen heterotypic type IV collageninteractions in the regulation of fibril assemblyrdquoMicron vol 32no 3 pp 223ndash237 2001
[21] M A Weis D M Hudson L Kim M Scott J-J Wu and D REyre ldquoLocation of 3-Hydroxyproline residues in collagen typesI II III and VXI implies a role in fibril supramolecularassemblyrdquo Journal of Biological Chemistry vol 285 no 4 pp2580ndash2590 2010
[22] R J Wenstrup J B Florer E W Brunskill S M Bell I Cher-voneva and D E Birk ldquoType V collagen controls the initiationof collagen fibril assemblyrdquo Journal of Biological Chemistry vol279 no 51 pp 53331ndash53337 2004
[23] C Niyibizi P P Fietzek and M Van der Rest ldquoHumanplacenta type V collagens Evidence for the existence of an1205721(V)1205722(V)1205723(V) collagen moleculerdquo Journal of BiologicalChemistry vol 259 no 22 pp 14170ndash14174 1984
[24] C Yang A C Park N A Davis et al ldquoComprehensive massspectrometricmapping of the hydroxylated amino acid residuesof the1205721(V) collagen chainrdquoThe Journal of Biological Chemistryvol 287 no 48 pp 40598ndash40610 2012
[25] O Matsushita T Koide R Kobayashi K Nagata and AOkabe ldquoSubstrate recognition by the collagen-binding domainof Clostridium histolyticum class I collagenaserdquo The Journal ofBiological Chemistry vol 276 no 12 pp 8761ndash8770 2001
[26] M F French K A Mookhtiar and H E Van Wart ldquoLimitedproteolysis of type I collagen at hyperreactive sites by class Iand II Clostridium histolyticum collagenases complementarydigestion patternsrdquo Biochemistry vol 26 no 3 pp 681ndash6871987
[27] T Kin X Zhai T B Murdoch A Salam A M J Shapiro andJ R T Lakey ldquoEnhancing the success of human islet isolationthrough optimization and characterization of pancreas dissoci-ation enzymerdquo American Journal of Transplantation vol 7 no5 pp 1233ndash1241 2007
[28] T Kin X Zhai D OrsquoGorman and A M J Shapiro ldquoDetri-mental effect of excessive collagenase class II on human isletisolation outcomerdquo Transplant International vol 21 no 11 pp1059ndash1065 2008
[29] J H M Van Deijnen P T R Van Suylichem G H J Woltersand R Van Schilfgaarde ldquoDistribution of collagens type I typeIII and type V in the pancreas of rat dog pig andmanrdquoCell andTissue Research vol 277 no 1 pp 115ndash121 1994
[30] T F Linsenmayer E Gibney F Igoe et al ldquoType V collagenmolecular structure and fibrillar organization of the chicken1205721(V) NH2-terminal domain a putative regulator of cornealfibrillogenesisrdquo Journal of Cell Biology vol 121 no 5 pp 1181ndash1189 1993
[31] J K Mouw G Ou and V M Weaver ldquoExtracellular matrixassembly a multiscale deconstructionrdquo Nature Reviews Molec-ular Cell Biology vol 15 no 12 pp 771ndash785 2014
[32] F Delacoux A Fichard C Geourjon R Garrone and FRuggiero ldquoMolecular features of the collagenVheparin bindingsiterdquo The Journal of Biological Chemistry vol 273 no 24 pp15069ndash15076 1998
[33] S Ricard-Blum M Beraud N Raynal R W Farndale and FRuggiero ldquoStructural requirements for heparinheparan sulfatebinding to type V collagenrdquoThe Journal of Biological Chemistryvol 281 no 35 pp 25195ndash25204 2006
[34] W Henkel and K Dreisewerd ldquoCyanogen bromide peptides ofthe fibrillar collagens I III and V and their mass spectrometriccharacterization detection of linear peptides peptide glycosyla-tion and cross-linking peptides involved in formation of homo-and heterotypic fibrilsrdquo Journal of Proteome Research vol 6 no11 pp 4269ndash4289 2007
[35] K Gelse E Poschl and T Aigner ldquoCollagensmdashstructurefunction and biosynthesisrdquo Advanced Drug Delivery Reviewsvol 55 no 12 pp 1531ndash1546 2003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
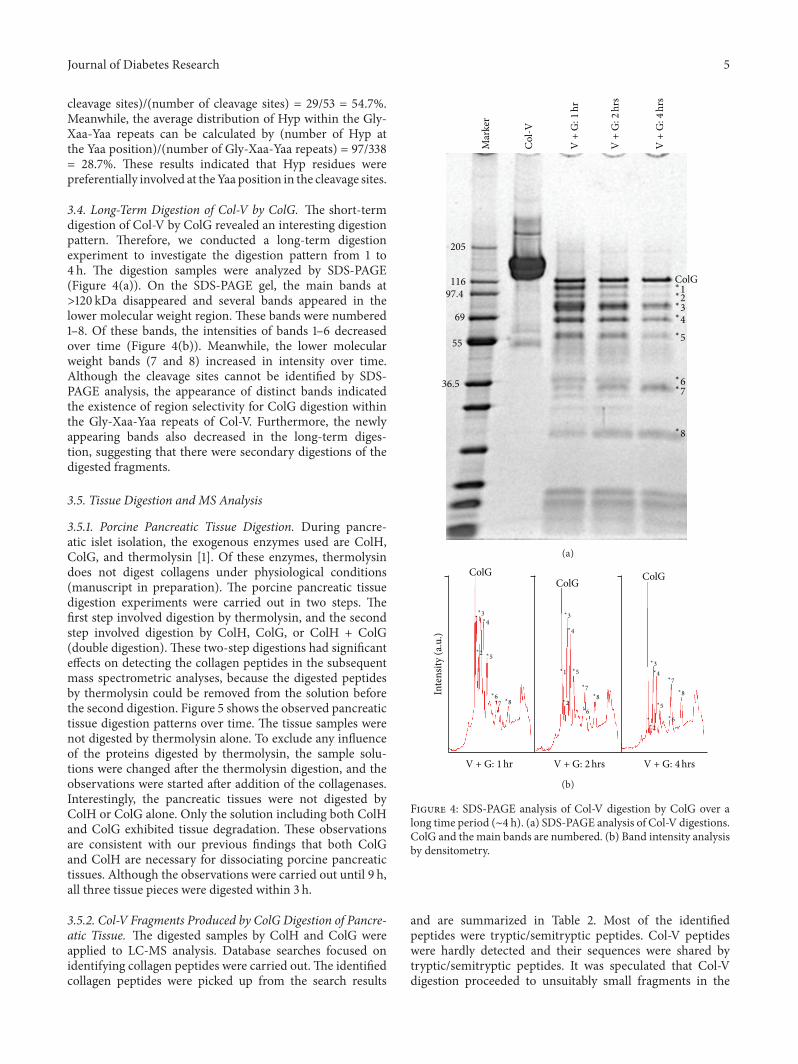
Journal of Diabetes Research 5
cleavage sites)(number of cleavage sites) = 2953 = 547Meanwhile the average distribution of Hyp within the Gly-Xaa-Yaa repeats can be calculated by (number of Hyp atthe Yaa position)(number of Gly-Xaa-Yaa repeats) = 97338= 287 These results indicated that Hyp residues werepreferentially involved at theYaa position in the cleavage sites
34 Long-Term Digestion of Col-V by ColG The short-termdigestion of Col-V by ColG revealed an interesting digestionpattern Therefore we conducted a long-term digestionexperiment to investigate the digestion pattern from 1 to4 h The digestion samples were analyzed by SDS-PAGE(Figure 4(a)) On the SDS-PAGE gel the main bands atgt120 kDa disappeared and several bands appeared in thelower molecular weight region These bands were numbered1ndash8 Of these bands the intensities of bands 1ndash6 decreasedover time (Figure 4(b)) Meanwhile the lower molecularweight bands (7 and 8) increased in intensity over timeAlthough the cleavage sites cannot be identified by SDS-PAGE analysis the appearance of distinct bands indicatedthe existence of region selectivity for ColG digestion withinthe Gly-Xaa-Yaa repeats of Col-V Furthermore the newlyappearing bands also decreased in the long-term diges-tion suggesting that there were secondary digestions of thedigested fragments
35 Tissue Digestion and MS Analysis
351 Porcine Pancreatic Tissue Digestion During pancre-atic islet isolation the exogenous enzymes used are ColHColG and thermolysin [1] Of these enzymes thermolysindoes not digest collagens under physiological conditions(manuscript in preparation) The porcine pancreatic tissuedigestion experiments were carried out in two steps Thefirst step involved digestion by thermolysin and the secondstep involved digestion by ColH ColG or ColH + ColG(double digestion) These two-step digestions had significanteffects on detecting the collagen peptides in the subsequentmass spectrometric analyses because the digested peptidesby thermolysin could be removed from the solution beforethe second digestion Figure 5 shows the observed pancreatictissue digestion patterns over time The tissue samples werenot digested by thermolysin alone To exclude any influenceof the proteins digested by thermolysin the sample solu-tions were changed after the thermolysin digestion and theobservations were started after addition of the collagenasesInterestingly the pancreatic tissues were not digested byColH or ColG alone Only the solution including both ColHand ColG exhibited tissue degradation These observationsare consistent with our previous findings that both ColGand ColH are necessary for dissociating porcine pancreatictissues Although the observations were carried out until 9 hall three tissue pieces were digested within 3 h
352 Col-V Fragments Produced by ColG Digestion of Pancre-atic Tissue The digested samples by ColH and ColG wereapplied to LC-MS analysis Database searches focused onidentifying collagen peptides were carried outThe identifiedcollagen peptides were picked up from the search results
205
116974
69
55
365
ColGlowast1
lowast2
lowast3
lowast4
lowast5
lowast6
lowast7
lowast8
Col
-V
Mar
ker
V +
G1
hr
V +
G2
hrs
V +
G4
hrs
(a)
ColGColG ColG
Inte
nsity
(au
)
lowast1
lowast1
lowast1
lowast2
lowast2
lowast2
lowast3 lowast
3
lowast3
lowast4
lowast4
lowast4
lowast5
lowast5
lowast5
lowast6
lowast6
lowast6
lowast7
lowast7
lowast7
lowast8
lowast8
lowast8
V + G 1hr V + G 2hrs V + G 4hrs
(b)
Figure 4 SDS-PAGE analysis of Col-V digestion by ColG over along time period (sim4 h) (a) SDS-PAGE analysis of Col-V digestionsColG and the main bands are numbered (b) Band intensity analysisby densitometry
and are summarized in Table 2 Most of the identifiedpeptides were trypticsemitryptic peptides Col-V peptideswere hardly detected and their sequences were shared bytrypticsemitryptic peptides It was speculated that Col-Vdigestion proceeded to unsuitably small fragments in the
6 Journal of Diabetes Research
Col
G
n2
n1
n3
n2
n1
n3
n2
n1
n3
Col
HC
olH
+ G
0hr 3hrs 6hrs 9hrs
Figure 5 In vitro porcine pancreatic tissue digestions by proteasesThe digestions were observed at 0 3 6 and 9 h Each digestion wasconducted in triplicate
presence of ColG together with other endogenous proteasesfor detection of the mass spectrometric analyses
4 Discussion
Our in vitro digestion experiments revealed that the Col-Vdigestion patterns were distinctly different betweenColH andColG
Col-V digestion by ColG was clearly shown by SDS-PAGE and mass spectrometric analyses Meanwhile ColHdid not digest Col-V under the same conditions used forColGAlthough this study could not investigate the differencein binding ability toward Col-V between ColH and ColG itis considered that the difference in domain construction is
Table 2 Collagen peptide identification from tissue digestion sam-ples
Col type tryp + semitryp nontryp Gxx-G
Col-V 1205721 2 0 0 21205722 4 0 0 3
Col-IV 1205721 1 2 3 21205722 1 1 1 1
Col-VI1205721 5 8 0 01205722 9 1 1 01205723 3 2 1 0
tryp semitryp and nontryp indicate the number of tryptic peptides (RK)darrxxxsdot sdot sdot xxx(RK)darrxxxsdot sdot sdot number of semitryptic peptides (RK)darrxxxsdot sdot sdotor sdot sdot sdot xxx(RK)darrxxxsdot sdot sdot and number of nontryptic peptides respectivelyThe total number of detected peptides is shown by tryp + semitryp + nontryp Gxx-G can be shared in tryp semitryp and nontrypDatabase searches for porcine collagens were based on the following NCBIaccession numbers 1205721(V) 62461592 1205722(V) 157427707 1205721(IV) 9271916811205722(IV) 927191683 1205721(VI) 92020086 92020094 and 545895133 1205722(VI)927203606 1205723(VI) 92020131 and 927211912
one of the possible reasons for the different collagenolyticactivities of ColH and ColG for Col-V ColH has onlyone CBD while ColG contains tandem CBDs (Figure 1)The CBDs of clostridial collagenases have been investigatedin detail A structural comparison indicated that the twostructures were similar (rmsd for C120572 = 15 A) despite therelatively low sequence similarity [11] The substrate-bindingspecificity of CBDs is broad for collagen types IndashIV [6] TheCBDs of ColH and ColG can bind to triple helical collagensbut not to telopeptides [5 25] However tandem CDBs canbind more efficiently to Col-I than a monomeric CBD [25]Consequently the domain construction especially the CBDarrangement plays a very important role in the digestioncharacteristics for Col-V
In the present study Col-V digestion by ColG produceddistinct digestion characteristics within a short time period(within minutes) and a long time period (within hours)The mass spectrometric analyses revealed that the identifiedpeptide cleavage sites were distributed in the N-terminalregion of theGly-Xaa-Yaa repeat region of Col-VThis findingsuggests that the digestion of Col-V is initiated from the N-terminal region According to the ColG digestion for Col-I [26] Col-I is hydrolyzed near the C-terminal end of thetriple helix region and a second cleavage occurs at the N-terminal side Although a digestion tendency for the C-terminal side was not found in our results digestion fromthe N-terminal region is a common digestion property ofColG for both Col-I and Col-V The long-term digestionexperiment indicated several residual parts of Col-V thatappeared as distinct bands at 1 h on SDS-PAGE These partswere also hydrolyzed by subsequent digestion (sim4 h) It issuggested that the Col-V sequence includes some parts thatare resistant to collagenolysis
The tissue digestion experiments clearly showed that bothColH and ColG were needed to degrade the pancreatic tissuestructure For human tissue degradation it was reported thatthe ratio of ColGH influenced the islet isolation outcome
Journal of Diabetes Research 7
[27 28] These results indicate that ColG may play moreimportant roles in digestion procedure of pig and humantissues than that of rat tissues The mass spectrometricanalysis for the digested samples identified collagen peptidesof Col-IV Col-V and collagen type VI Our previous studyrevealed that ColH is crucial for rat pancreatic islet isolationand the most abundantly detected collagen peptides in themass spectrometric analysis were Col-I and Col-III [17]These results strongly suggested the presence of markeddifferences between rat and porcine pancreatic tissue degra-dations Although it is difficult to identify the exact reasonfor the discrepancies fibril formation could be a possibleexplanation Collagen molecules are differently involved invarious tissues according to animal species [29] Col-V isassociated with Col-I in the microfibrillar structure [3031] Furthermore Col-V can be involved in heparinheparinsulfate modifications [32 33] and glycosylation [34] aswell as the well-known hydroxyl group modifications atproline and lysine residues [35] Therefore the differences incircumstances of the Col-IV fibrillar structures in the ratand porcine pancreatic tissue may influence the pancreaticislet isolation efficiency Although the similarities of thecircumstances of the Col-IV fibrillar structures among ratspigs and humans for pancreatic islet isolation are difficult tocompare ColG appears to be more important in porcine andhuman islet isolation processes and Col-V may be one of thekey substrates for ColG
5 Conclusions
In this study we focused on Col-V digestion by clostridialcollagenases Col-V was distinctively digested by ColG Massspectrometric analyses revealed that Col-V digestion byColG proceeded from the N-terminal region Although bothclostridial collagenases ColH and ColG include commondomains such as the activator domain peptidase domainPKD and CBD their domain construction is different Thedomain construction may play a crucial role for determiningthe collagen digestion patterns by the clostridial collagenasesThe porcine pancreatic tissue digestion experiments demon-strated that both collagenases are needed to degrade the tissuestructure These observations are completely different fromthose in rat experiments in which ColG was not cruciallyneeded Consequently the present study reveals that ColG aswell as ColH plays a crucial role in pancreatic islet isolationin large mammals and that Col-V can be one of the keysubstrates for ColG
Competing Interests
The authors of this paper received partial financial supportfromMeiji Seika PharmaCo Ltd and ShimadzuCorporationaccording to a collaborative research contract
Authorsrsquo Contributions
Kazutaka Murayama designed the experiments performedthe sample digestion andMALDI-tof-MS analyses andwrote
the paper Akiko Inagaki and Takehiro Imura performed thetissue digestion experiments Hiroki Shima performed theLC-MS spectrometric analyses and wrote part of the paperYouhei Yamagata and Kimiko Watanabe participated in thepaper corrections and critical review Kazuhiko Igarashi andMasafumi Goto participated in discussions during the paperpreparation and paper corrections
Acknowledgments
The authors acknowledge the support of the BiomedicalResearch Core and TAMRIC (Tohoku Advanced MedicalResearch and IncubationCenter) of TohokuUniversityGrad-uate School of Medicine Part of this research was supportedby the Tohoku University Business Incubation Program
References
[1] R C McCarthy A G Breite M L Green and F E DwuletldquoTissue dissociation enzymes for isolating human islets fortransplantation Factors to consider in setting enzyme accep-tance criteriardquo Transplantation vol 91 no 2 pp 137ndash145 2011
[2] K Watanabe ldquoCollagenolytic proteases from bacteriardquo AppliedMicrobiology and Biotechnology vol 63 no 5 pp 520ndash5262004
[3] U Eckhard E Schonauer P Ducka P Briza D Nuss andH Brandstetter ldquoBiochemical characterization of the catalyticdomains of three different clostridial collagenasesrdquo BiologicalChemistry vol 390 no 1 pp 11ndash18 2009
[4] C-M Jung O Matsushita S Katayama J Minami J Sakuraiand A Okabe ldquoIdentification of metal ligands in the Clostrid-ium histolyticumColH collagenaserdquo Journal of Bacteriology vol181 no 9 pp 2816ndash2822 1999
[5] O Matsushita C-M Jung J Minami S Katayama N Nishiand A Okabe ldquoA study of the collagen-binding domain of a116-kDa Clostridium histolyticum collagenaserdquo The Journal ofBiological Chemistry vol 273 no 6 pp 3643ndash3648 1998
[6] T Toyoshima O Matsushita J Minami N Nishi A Okabeand T Itano ldquoCollagen-binding domain of a Clostridium his-tolyticum collagenase exhibits a broad substrate spectrum bothin vitro and in vivordquo Connective Tissue Research vol 42 no 4pp 281ndash290 2001
[7] U Eckhard E Schonauer D Nuss and H Brandstetter ldquoStruc-ture of collagenase G reveals a chew-and-digest mechanismof bacterial collagenolysisrdquo Nature Structural and MolecularBiology vol 18 no 10 pp 1109ndash1114 2010
[8] U Eckhard E Schonauer andH Brandstetter ldquoStructural basisfor activity regulation and substrate preference of clostridialcollagenases G H and Trdquo The Journal of Biological Chemistryvol 288 no 28 pp 20184ndash20194 2013
[9] R Bauer K Janowska K Taylor et al ldquoStructures of threepolycystic kidney disease-like domains from Clostridium his-tolyticum collagenases ColG and ColHrdquo Acta CrystallographicaSection D Biological Crystallography vol 71 pp 565ndash577 2015
[10] U Eckhard and H Brandstetter ldquoPolycystic kidney disease-likedomains of clostridial collagenases and their role in collagenrecruitmentrdquo Biological Chemistry vol 392 no 11 pp 1039ndash1045 2011
8 Journal of Diabetes Research
[11] R Bauer J J Wilson S T L Philominathan D Davis OMatsushita and J Sakona ldquoStructural comparison of ColHand ColG collagen-binding domains from Clostridium his-tolyticumrdquo Journal of Bacteriology vol 195 no 2 pp 318ndash3272013
[12] J J Wilson O Matsushita A Okabe and J Sakon ldquoA bacterialcollagen-binding domain with novel calcium-binding motifcontrols domain orientationrdquoThe EMBO Journal vol 22 no 8pp 1743ndash1752 2003
[13] N Ohbayashi T Matsumoto H Shima et al ldquoSolution struc-ture of clostridial collagenase H and its calcium-dependentglobal conformation changerdquo Biophysical Journal vol 104 no7 pp 1538ndash1545 2013
[14] N Ohbayashi N Yamagata M Goto K Watanabe Y Yam-agata and K Murayama ldquoEnhancement of the structuralstability of full-length clostridial collagenase by calcium ionsrdquoApplied and Environmental Microbiology vol 78 no 16 pp5839ndash5844 2012
[15] U Eckhard P F Huesgen H Brandstetter and C M OverallldquoProteomic protease specificity profiling of clostridial collage-nases reveals their intrinsic nature as dedicated degraders ofcollagenrdquo Journal of Proteomics vol 100 pp 102ndash114 2014
[16] O Matsushita and A Okabe ldquoClostridial hydrolytic enzymesdegrading extracellular componentsrdquoToxicon vol 39 no 11 pp1769ndash1780 2001
[17] A Fujio K Murayama Y Yamagata et al ldquoCollagenase H iscrucial for isolation of rat pancreatic isletsrdquo Cell Transplanta-tion vol 23 no 10 pp 1187ndash1198 2014
[18] M K Gordon and R A Hahn ldquoCollagensrdquo Cell and TissueResearch vol 339 no 1 pp 247ndash257 2010
[19] MD Shoulders andR T Raines ldquoCollagen structure and stabil-ityrdquo Annual Review of Biochemistry vol 78 pp 929ndash958 2009
[20] D E Birk ldquoType V collagen heterotypic type IV collageninteractions in the regulation of fibril assemblyrdquoMicron vol 32no 3 pp 223ndash237 2001
[21] M A Weis D M Hudson L Kim M Scott J-J Wu and D REyre ldquoLocation of 3-Hydroxyproline residues in collagen typesI II III and VXI implies a role in fibril supramolecularassemblyrdquo Journal of Biological Chemistry vol 285 no 4 pp2580ndash2590 2010
[22] R J Wenstrup J B Florer E W Brunskill S M Bell I Cher-voneva and D E Birk ldquoType V collagen controls the initiationof collagen fibril assemblyrdquo Journal of Biological Chemistry vol279 no 51 pp 53331ndash53337 2004
[23] C Niyibizi P P Fietzek and M Van der Rest ldquoHumanplacenta type V collagens Evidence for the existence of an1205721(V)1205722(V)1205723(V) collagen moleculerdquo Journal of BiologicalChemistry vol 259 no 22 pp 14170ndash14174 1984
[24] C Yang A C Park N A Davis et al ldquoComprehensive massspectrometricmapping of the hydroxylated amino acid residuesof the1205721(V) collagen chainrdquoThe Journal of Biological Chemistryvol 287 no 48 pp 40598ndash40610 2012
[25] O Matsushita T Koide R Kobayashi K Nagata and AOkabe ldquoSubstrate recognition by the collagen-binding domainof Clostridium histolyticum class I collagenaserdquo The Journal ofBiological Chemistry vol 276 no 12 pp 8761ndash8770 2001
[26] M F French K A Mookhtiar and H E Van Wart ldquoLimitedproteolysis of type I collagen at hyperreactive sites by class Iand II Clostridium histolyticum collagenases complementarydigestion patternsrdquo Biochemistry vol 26 no 3 pp 681ndash6871987
[27] T Kin X Zhai T B Murdoch A Salam A M J Shapiro andJ R T Lakey ldquoEnhancing the success of human islet isolationthrough optimization and characterization of pancreas dissoci-ation enzymerdquo American Journal of Transplantation vol 7 no5 pp 1233ndash1241 2007
[28] T Kin X Zhai D OrsquoGorman and A M J Shapiro ldquoDetri-mental effect of excessive collagenase class II on human isletisolation outcomerdquo Transplant International vol 21 no 11 pp1059ndash1065 2008
[29] J H M Van Deijnen P T R Van Suylichem G H J Woltersand R Van Schilfgaarde ldquoDistribution of collagens type I typeIII and type V in the pancreas of rat dog pig andmanrdquoCell andTissue Research vol 277 no 1 pp 115ndash121 1994
[30] T F Linsenmayer E Gibney F Igoe et al ldquoType V collagenmolecular structure and fibrillar organization of the chicken1205721(V) NH2-terminal domain a putative regulator of cornealfibrillogenesisrdquo Journal of Cell Biology vol 121 no 5 pp 1181ndash1189 1993
[31] J K Mouw G Ou and V M Weaver ldquoExtracellular matrixassembly a multiscale deconstructionrdquo Nature Reviews Molec-ular Cell Biology vol 15 no 12 pp 771ndash785 2014
[32] F Delacoux A Fichard C Geourjon R Garrone and FRuggiero ldquoMolecular features of the collagenVheparin bindingsiterdquo The Journal of Biological Chemistry vol 273 no 24 pp15069ndash15076 1998
[33] S Ricard-Blum M Beraud N Raynal R W Farndale and FRuggiero ldquoStructural requirements for heparinheparan sulfatebinding to type V collagenrdquoThe Journal of Biological Chemistryvol 281 no 35 pp 25195ndash25204 2006
[34] W Henkel and K Dreisewerd ldquoCyanogen bromide peptides ofthe fibrillar collagens I III and V and their mass spectrometriccharacterization detection of linear peptides peptide glycosyla-tion and cross-linking peptides involved in formation of homo-and heterotypic fibrilsrdquo Journal of Proteome Research vol 6 no11 pp 4269ndash4289 2007
[35] K Gelse E Poschl and T Aigner ldquoCollagensmdashstructurefunction and biosynthesisrdquo Advanced Drug Delivery Reviewsvol 55 no 12 pp 1531ndash1546 2003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

6 Journal of Diabetes Research
Col
G
n2
n1
n3
n2
n1
n3
n2
n1
n3
Col
HC
olH
+ G
0hr 3hrs 6hrs 9hrs
Figure 5 In vitro porcine pancreatic tissue digestions by proteasesThe digestions were observed at 0 3 6 and 9 h Each digestion wasconducted in triplicate
presence of ColG together with other endogenous proteasesfor detection of the mass spectrometric analyses
4 Discussion
Our in vitro digestion experiments revealed that the Col-Vdigestion patterns were distinctly different betweenColH andColG
Col-V digestion by ColG was clearly shown by SDS-PAGE and mass spectrometric analyses Meanwhile ColHdid not digest Col-V under the same conditions used forColGAlthough this study could not investigate the differencein binding ability toward Col-V between ColH and ColG itis considered that the difference in domain construction is
Table 2 Collagen peptide identification from tissue digestion sam-ples
Col type tryp + semitryp nontryp Gxx-G
Col-V 1205721 2 0 0 21205722 4 0 0 3
Col-IV 1205721 1 2 3 21205722 1 1 1 1
Col-VI1205721 5 8 0 01205722 9 1 1 01205723 3 2 1 0
tryp semitryp and nontryp indicate the number of tryptic peptides (RK)darrxxxsdot sdot sdot xxx(RK)darrxxxsdot sdot sdot number of semitryptic peptides (RK)darrxxxsdot sdot sdotor sdot sdot sdot xxx(RK)darrxxxsdot sdot sdot and number of nontryptic peptides respectivelyThe total number of detected peptides is shown by tryp + semitryp + nontryp Gxx-G can be shared in tryp semitryp and nontrypDatabase searches for porcine collagens were based on the following NCBIaccession numbers 1205721(V) 62461592 1205722(V) 157427707 1205721(IV) 9271916811205722(IV) 927191683 1205721(VI) 92020086 92020094 and 545895133 1205722(VI)927203606 1205723(VI) 92020131 and 927211912
one of the possible reasons for the different collagenolyticactivities of ColH and ColG for Col-V ColH has onlyone CBD while ColG contains tandem CBDs (Figure 1)The CBDs of clostridial collagenases have been investigatedin detail A structural comparison indicated that the twostructures were similar (rmsd for C120572 = 15 A) despite therelatively low sequence similarity [11] The substrate-bindingspecificity of CBDs is broad for collagen types IndashIV [6] TheCBDs of ColH and ColG can bind to triple helical collagensbut not to telopeptides [5 25] However tandem CDBs canbind more efficiently to Col-I than a monomeric CBD [25]Consequently the domain construction especially the CBDarrangement plays a very important role in the digestioncharacteristics for Col-V
In the present study Col-V digestion by ColG produceddistinct digestion characteristics within a short time period(within minutes) and a long time period (within hours)The mass spectrometric analyses revealed that the identifiedpeptide cleavage sites were distributed in the N-terminalregion of theGly-Xaa-Yaa repeat region of Col-VThis findingsuggests that the digestion of Col-V is initiated from the N-terminal region According to the ColG digestion for Col-I [26] Col-I is hydrolyzed near the C-terminal end of thetriple helix region and a second cleavage occurs at the N-terminal side Although a digestion tendency for the C-terminal side was not found in our results digestion fromthe N-terminal region is a common digestion property ofColG for both Col-I and Col-V The long-term digestionexperiment indicated several residual parts of Col-V thatappeared as distinct bands at 1 h on SDS-PAGE These partswere also hydrolyzed by subsequent digestion (sim4 h) It issuggested that the Col-V sequence includes some parts thatare resistant to collagenolysis
The tissue digestion experiments clearly showed that bothColH and ColG were needed to degrade the pancreatic tissuestructure For human tissue degradation it was reported thatthe ratio of ColGH influenced the islet isolation outcome
Journal of Diabetes Research 7
[27 28] These results indicate that ColG may play moreimportant roles in digestion procedure of pig and humantissues than that of rat tissues The mass spectrometricanalysis for the digested samples identified collagen peptidesof Col-IV Col-V and collagen type VI Our previous studyrevealed that ColH is crucial for rat pancreatic islet isolationand the most abundantly detected collagen peptides in themass spectrometric analysis were Col-I and Col-III [17]These results strongly suggested the presence of markeddifferences between rat and porcine pancreatic tissue degra-dations Although it is difficult to identify the exact reasonfor the discrepancies fibril formation could be a possibleexplanation Collagen molecules are differently involved invarious tissues according to animal species [29] Col-V isassociated with Col-I in the microfibrillar structure [3031] Furthermore Col-V can be involved in heparinheparinsulfate modifications [32 33] and glycosylation [34] aswell as the well-known hydroxyl group modifications atproline and lysine residues [35] Therefore the differences incircumstances of the Col-IV fibrillar structures in the ratand porcine pancreatic tissue may influence the pancreaticislet isolation efficiency Although the similarities of thecircumstances of the Col-IV fibrillar structures among ratspigs and humans for pancreatic islet isolation are difficult tocompare ColG appears to be more important in porcine andhuman islet isolation processes and Col-V may be one of thekey substrates for ColG
5 Conclusions
In this study we focused on Col-V digestion by clostridialcollagenases Col-V was distinctively digested by ColG Massspectrometric analyses revealed that Col-V digestion byColG proceeded from the N-terminal region Although bothclostridial collagenases ColH and ColG include commondomains such as the activator domain peptidase domainPKD and CBD their domain construction is different Thedomain construction may play a crucial role for determiningthe collagen digestion patterns by the clostridial collagenasesThe porcine pancreatic tissue digestion experiments demon-strated that both collagenases are needed to degrade the tissuestructure These observations are completely different fromthose in rat experiments in which ColG was not cruciallyneeded Consequently the present study reveals that ColG aswell as ColH plays a crucial role in pancreatic islet isolationin large mammals and that Col-V can be one of the keysubstrates for ColG
Competing Interests
The authors of this paper received partial financial supportfromMeiji Seika PharmaCo Ltd and ShimadzuCorporationaccording to a collaborative research contract
Authorsrsquo Contributions
Kazutaka Murayama designed the experiments performedthe sample digestion andMALDI-tof-MS analyses andwrote
the paper Akiko Inagaki and Takehiro Imura performed thetissue digestion experiments Hiroki Shima performed theLC-MS spectrometric analyses and wrote part of the paperYouhei Yamagata and Kimiko Watanabe participated in thepaper corrections and critical review Kazuhiko Igarashi andMasafumi Goto participated in discussions during the paperpreparation and paper corrections
Acknowledgments
The authors acknowledge the support of the BiomedicalResearch Core and TAMRIC (Tohoku Advanced MedicalResearch and IncubationCenter) of TohokuUniversityGrad-uate School of Medicine Part of this research was supportedby the Tohoku University Business Incubation Program
References
[1] R C McCarthy A G Breite M L Green and F E DwuletldquoTissue dissociation enzymes for isolating human islets fortransplantation Factors to consider in setting enzyme accep-tance criteriardquo Transplantation vol 91 no 2 pp 137ndash145 2011
[2] K Watanabe ldquoCollagenolytic proteases from bacteriardquo AppliedMicrobiology and Biotechnology vol 63 no 5 pp 520ndash5262004
[3] U Eckhard E Schonauer P Ducka P Briza D Nuss andH Brandstetter ldquoBiochemical characterization of the catalyticdomains of three different clostridial collagenasesrdquo BiologicalChemistry vol 390 no 1 pp 11ndash18 2009
[4] C-M Jung O Matsushita S Katayama J Minami J Sakuraiand A Okabe ldquoIdentification of metal ligands in the Clostrid-ium histolyticumColH collagenaserdquo Journal of Bacteriology vol181 no 9 pp 2816ndash2822 1999
[5] O Matsushita C-M Jung J Minami S Katayama N Nishiand A Okabe ldquoA study of the collagen-binding domain of a116-kDa Clostridium histolyticum collagenaserdquo The Journal ofBiological Chemistry vol 273 no 6 pp 3643ndash3648 1998
[6] T Toyoshima O Matsushita J Minami N Nishi A Okabeand T Itano ldquoCollagen-binding domain of a Clostridium his-tolyticum collagenase exhibits a broad substrate spectrum bothin vitro and in vivordquo Connective Tissue Research vol 42 no 4pp 281ndash290 2001
[7] U Eckhard E Schonauer D Nuss and H Brandstetter ldquoStruc-ture of collagenase G reveals a chew-and-digest mechanismof bacterial collagenolysisrdquo Nature Structural and MolecularBiology vol 18 no 10 pp 1109ndash1114 2010
[8] U Eckhard E Schonauer andH Brandstetter ldquoStructural basisfor activity regulation and substrate preference of clostridialcollagenases G H and Trdquo The Journal of Biological Chemistryvol 288 no 28 pp 20184ndash20194 2013
[9] R Bauer K Janowska K Taylor et al ldquoStructures of threepolycystic kidney disease-like domains from Clostridium his-tolyticum collagenases ColG and ColHrdquo Acta CrystallographicaSection D Biological Crystallography vol 71 pp 565ndash577 2015
[10] U Eckhard and H Brandstetter ldquoPolycystic kidney disease-likedomains of clostridial collagenases and their role in collagenrecruitmentrdquo Biological Chemistry vol 392 no 11 pp 1039ndash1045 2011
8 Journal of Diabetes Research
[11] R Bauer J J Wilson S T L Philominathan D Davis OMatsushita and J Sakona ldquoStructural comparison of ColHand ColG collagen-binding domains from Clostridium his-tolyticumrdquo Journal of Bacteriology vol 195 no 2 pp 318ndash3272013
[12] J J Wilson O Matsushita A Okabe and J Sakon ldquoA bacterialcollagen-binding domain with novel calcium-binding motifcontrols domain orientationrdquoThe EMBO Journal vol 22 no 8pp 1743ndash1752 2003
[13] N Ohbayashi T Matsumoto H Shima et al ldquoSolution struc-ture of clostridial collagenase H and its calcium-dependentglobal conformation changerdquo Biophysical Journal vol 104 no7 pp 1538ndash1545 2013
[14] N Ohbayashi N Yamagata M Goto K Watanabe Y Yam-agata and K Murayama ldquoEnhancement of the structuralstability of full-length clostridial collagenase by calcium ionsrdquoApplied and Environmental Microbiology vol 78 no 16 pp5839ndash5844 2012
[15] U Eckhard P F Huesgen H Brandstetter and C M OverallldquoProteomic protease specificity profiling of clostridial collage-nases reveals their intrinsic nature as dedicated degraders ofcollagenrdquo Journal of Proteomics vol 100 pp 102ndash114 2014
[16] O Matsushita and A Okabe ldquoClostridial hydrolytic enzymesdegrading extracellular componentsrdquoToxicon vol 39 no 11 pp1769ndash1780 2001
[17] A Fujio K Murayama Y Yamagata et al ldquoCollagenase H iscrucial for isolation of rat pancreatic isletsrdquo Cell Transplanta-tion vol 23 no 10 pp 1187ndash1198 2014
[18] M K Gordon and R A Hahn ldquoCollagensrdquo Cell and TissueResearch vol 339 no 1 pp 247ndash257 2010
[19] MD Shoulders andR T Raines ldquoCollagen structure and stabil-ityrdquo Annual Review of Biochemistry vol 78 pp 929ndash958 2009
[20] D E Birk ldquoType V collagen heterotypic type IV collageninteractions in the regulation of fibril assemblyrdquoMicron vol 32no 3 pp 223ndash237 2001
[21] M A Weis D M Hudson L Kim M Scott J-J Wu and D REyre ldquoLocation of 3-Hydroxyproline residues in collagen typesI II III and VXI implies a role in fibril supramolecularassemblyrdquo Journal of Biological Chemistry vol 285 no 4 pp2580ndash2590 2010
[22] R J Wenstrup J B Florer E W Brunskill S M Bell I Cher-voneva and D E Birk ldquoType V collagen controls the initiationof collagen fibril assemblyrdquo Journal of Biological Chemistry vol279 no 51 pp 53331ndash53337 2004
[23] C Niyibizi P P Fietzek and M Van der Rest ldquoHumanplacenta type V collagens Evidence for the existence of an1205721(V)1205722(V)1205723(V) collagen moleculerdquo Journal of BiologicalChemistry vol 259 no 22 pp 14170ndash14174 1984
[24] C Yang A C Park N A Davis et al ldquoComprehensive massspectrometricmapping of the hydroxylated amino acid residuesof the1205721(V) collagen chainrdquoThe Journal of Biological Chemistryvol 287 no 48 pp 40598ndash40610 2012
[25] O Matsushita T Koide R Kobayashi K Nagata and AOkabe ldquoSubstrate recognition by the collagen-binding domainof Clostridium histolyticum class I collagenaserdquo The Journal ofBiological Chemistry vol 276 no 12 pp 8761ndash8770 2001
[26] M F French K A Mookhtiar and H E Van Wart ldquoLimitedproteolysis of type I collagen at hyperreactive sites by class Iand II Clostridium histolyticum collagenases complementarydigestion patternsrdquo Biochemistry vol 26 no 3 pp 681ndash6871987
[27] T Kin X Zhai T B Murdoch A Salam A M J Shapiro andJ R T Lakey ldquoEnhancing the success of human islet isolationthrough optimization and characterization of pancreas dissoci-ation enzymerdquo American Journal of Transplantation vol 7 no5 pp 1233ndash1241 2007
[28] T Kin X Zhai D OrsquoGorman and A M J Shapiro ldquoDetri-mental effect of excessive collagenase class II on human isletisolation outcomerdquo Transplant International vol 21 no 11 pp1059ndash1065 2008
[29] J H M Van Deijnen P T R Van Suylichem G H J Woltersand R Van Schilfgaarde ldquoDistribution of collagens type I typeIII and type V in the pancreas of rat dog pig andmanrdquoCell andTissue Research vol 277 no 1 pp 115ndash121 1994
[30] T F Linsenmayer E Gibney F Igoe et al ldquoType V collagenmolecular structure and fibrillar organization of the chicken1205721(V) NH2-terminal domain a putative regulator of cornealfibrillogenesisrdquo Journal of Cell Biology vol 121 no 5 pp 1181ndash1189 1993
[31] J K Mouw G Ou and V M Weaver ldquoExtracellular matrixassembly a multiscale deconstructionrdquo Nature Reviews Molec-ular Cell Biology vol 15 no 12 pp 771ndash785 2014
[32] F Delacoux A Fichard C Geourjon R Garrone and FRuggiero ldquoMolecular features of the collagenVheparin bindingsiterdquo The Journal of Biological Chemistry vol 273 no 24 pp15069ndash15076 1998
[33] S Ricard-Blum M Beraud N Raynal R W Farndale and FRuggiero ldquoStructural requirements for heparinheparan sulfatebinding to type V collagenrdquoThe Journal of Biological Chemistryvol 281 no 35 pp 25195ndash25204 2006
[34] W Henkel and K Dreisewerd ldquoCyanogen bromide peptides ofthe fibrillar collagens I III and V and their mass spectrometriccharacterization detection of linear peptides peptide glycosyla-tion and cross-linking peptides involved in formation of homo-and heterotypic fibrilsrdquo Journal of Proteome Research vol 6 no11 pp 4269ndash4289 2007
[35] K Gelse E Poschl and T Aigner ldquoCollagensmdashstructurefunction and biosynthesisrdquo Advanced Drug Delivery Reviewsvol 55 no 12 pp 1531ndash1546 2003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Journal of Diabetes Research 7
[27 28] These results indicate that ColG may play moreimportant roles in digestion procedure of pig and humantissues than that of rat tissues The mass spectrometricanalysis for the digested samples identified collagen peptidesof Col-IV Col-V and collagen type VI Our previous studyrevealed that ColH is crucial for rat pancreatic islet isolationand the most abundantly detected collagen peptides in themass spectrometric analysis were Col-I and Col-III [17]These results strongly suggested the presence of markeddifferences between rat and porcine pancreatic tissue degra-dations Although it is difficult to identify the exact reasonfor the discrepancies fibril formation could be a possibleexplanation Collagen molecules are differently involved invarious tissues according to animal species [29] Col-V isassociated with Col-I in the microfibrillar structure [3031] Furthermore Col-V can be involved in heparinheparinsulfate modifications [32 33] and glycosylation [34] aswell as the well-known hydroxyl group modifications atproline and lysine residues [35] Therefore the differences incircumstances of the Col-IV fibrillar structures in the ratand porcine pancreatic tissue may influence the pancreaticislet isolation efficiency Although the similarities of thecircumstances of the Col-IV fibrillar structures among ratspigs and humans for pancreatic islet isolation are difficult tocompare ColG appears to be more important in porcine andhuman islet isolation processes and Col-V may be one of thekey substrates for ColG
5 Conclusions
In this study we focused on Col-V digestion by clostridialcollagenases Col-V was distinctively digested by ColG Massspectrometric analyses revealed that Col-V digestion byColG proceeded from the N-terminal region Although bothclostridial collagenases ColH and ColG include commondomains such as the activator domain peptidase domainPKD and CBD their domain construction is different Thedomain construction may play a crucial role for determiningthe collagen digestion patterns by the clostridial collagenasesThe porcine pancreatic tissue digestion experiments demon-strated that both collagenases are needed to degrade the tissuestructure These observations are completely different fromthose in rat experiments in which ColG was not cruciallyneeded Consequently the present study reveals that ColG aswell as ColH plays a crucial role in pancreatic islet isolationin large mammals and that Col-V can be one of the keysubstrates for ColG
Competing Interests
The authors of this paper received partial financial supportfromMeiji Seika PharmaCo Ltd and ShimadzuCorporationaccording to a collaborative research contract
Authorsrsquo Contributions
Kazutaka Murayama designed the experiments performedthe sample digestion andMALDI-tof-MS analyses andwrote
the paper Akiko Inagaki and Takehiro Imura performed thetissue digestion experiments Hiroki Shima performed theLC-MS spectrometric analyses and wrote part of the paperYouhei Yamagata and Kimiko Watanabe participated in thepaper corrections and critical review Kazuhiko Igarashi andMasafumi Goto participated in discussions during the paperpreparation and paper corrections
Acknowledgments
The authors acknowledge the support of the BiomedicalResearch Core and TAMRIC (Tohoku Advanced MedicalResearch and IncubationCenter) of TohokuUniversityGrad-uate School of Medicine Part of this research was supportedby the Tohoku University Business Incubation Program
References
[1] R C McCarthy A G Breite M L Green and F E DwuletldquoTissue dissociation enzymes for isolating human islets fortransplantation Factors to consider in setting enzyme accep-tance criteriardquo Transplantation vol 91 no 2 pp 137ndash145 2011
[2] K Watanabe ldquoCollagenolytic proteases from bacteriardquo AppliedMicrobiology and Biotechnology vol 63 no 5 pp 520ndash5262004
[3] U Eckhard E Schonauer P Ducka P Briza D Nuss andH Brandstetter ldquoBiochemical characterization of the catalyticdomains of three different clostridial collagenasesrdquo BiologicalChemistry vol 390 no 1 pp 11ndash18 2009
[4] C-M Jung O Matsushita S Katayama J Minami J Sakuraiand A Okabe ldquoIdentification of metal ligands in the Clostrid-ium histolyticumColH collagenaserdquo Journal of Bacteriology vol181 no 9 pp 2816ndash2822 1999
[5] O Matsushita C-M Jung J Minami S Katayama N Nishiand A Okabe ldquoA study of the collagen-binding domain of a116-kDa Clostridium histolyticum collagenaserdquo The Journal ofBiological Chemistry vol 273 no 6 pp 3643ndash3648 1998
[6] T Toyoshima O Matsushita J Minami N Nishi A Okabeand T Itano ldquoCollagen-binding domain of a Clostridium his-tolyticum collagenase exhibits a broad substrate spectrum bothin vitro and in vivordquo Connective Tissue Research vol 42 no 4pp 281ndash290 2001
[7] U Eckhard E Schonauer D Nuss and H Brandstetter ldquoStruc-ture of collagenase G reveals a chew-and-digest mechanismof bacterial collagenolysisrdquo Nature Structural and MolecularBiology vol 18 no 10 pp 1109ndash1114 2010
[8] U Eckhard E Schonauer andH Brandstetter ldquoStructural basisfor activity regulation and substrate preference of clostridialcollagenases G H and Trdquo The Journal of Biological Chemistryvol 288 no 28 pp 20184ndash20194 2013
[9] R Bauer K Janowska K Taylor et al ldquoStructures of threepolycystic kidney disease-like domains from Clostridium his-tolyticum collagenases ColG and ColHrdquo Acta CrystallographicaSection D Biological Crystallography vol 71 pp 565ndash577 2015
[10] U Eckhard and H Brandstetter ldquoPolycystic kidney disease-likedomains of clostridial collagenases and their role in collagenrecruitmentrdquo Biological Chemistry vol 392 no 11 pp 1039ndash1045 2011
8 Journal of Diabetes Research
[11] R Bauer J J Wilson S T L Philominathan D Davis OMatsushita and J Sakona ldquoStructural comparison of ColHand ColG collagen-binding domains from Clostridium his-tolyticumrdquo Journal of Bacteriology vol 195 no 2 pp 318ndash3272013
[12] J J Wilson O Matsushita A Okabe and J Sakon ldquoA bacterialcollagen-binding domain with novel calcium-binding motifcontrols domain orientationrdquoThe EMBO Journal vol 22 no 8pp 1743ndash1752 2003
[13] N Ohbayashi T Matsumoto H Shima et al ldquoSolution struc-ture of clostridial collagenase H and its calcium-dependentglobal conformation changerdquo Biophysical Journal vol 104 no7 pp 1538ndash1545 2013
[14] N Ohbayashi N Yamagata M Goto K Watanabe Y Yam-agata and K Murayama ldquoEnhancement of the structuralstability of full-length clostridial collagenase by calcium ionsrdquoApplied and Environmental Microbiology vol 78 no 16 pp5839ndash5844 2012
[15] U Eckhard P F Huesgen H Brandstetter and C M OverallldquoProteomic protease specificity profiling of clostridial collage-nases reveals their intrinsic nature as dedicated degraders ofcollagenrdquo Journal of Proteomics vol 100 pp 102ndash114 2014
[16] O Matsushita and A Okabe ldquoClostridial hydrolytic enzymesdegrading extracellular componentsrdquoToxicon vol 39 no 11 pp1769ndash1780 2001
[17] A Fujio K Murayama Y Yamagata et al ldquoCollagenase H iscrucial for isolation of rat pancreatic isletsrdquo Cell Transplanta-tion vol 23 no 10 pp 1187ndash1198 2014
[18] M K Gordon and R A Hahn ldquoCollagensrdquo Cell and TissueResearch vol 339 no 1 pp 247ndash257 2010
[19] MD Shoulders andR T Raines ldquoCollagen structure and stabil-ityrdquo Annual Review of Biochemistry vol 78 pp 929ndash958 2009
[20] D E Birk ldquoType V collagen heterotypic type IV collageninteractions in the regulation of fibril assemblyrdquoMicron vol 32no 3 pp 223ndash237 2001
[21] M A Weis D M Hudson L Kim M Scott J-J Wu and D REyre ldquoLocation of 3-Hydroxyproline residues in collagen typesI II III and VXI implies a role in fibril supramolecularassemblyrdquo Journal of Biological Chemistry vol 285 no 4 pp2580ndash2590 2010
[22] R J Wenstrup J B Florer E W Brunskill S M Bell I Cher-voneva and D E Birk ldquoType V collagen controls the initiationof collagen fibril assemblyrdquo Journal of Biological Chemistry vol279 no 51 pp 53331ndash53337 2004
[23] C Niyibizi P P Fietzek and M Van der Rest ldquoHumanplacenta type V collagens Evidence for the existence of an1205721(V)1205722(V)1205723(V) collagen moleculerdquo Journal of BiologicalChemistry vol 259 no 22 pp 14170ndash14174 1984
[24] C Yang A C Park N A Davis et al ldquoComprehensive massspectrometricmapping of the hydroxylated amino acid residuesof the1205721(V) collagen chainrdquoThe Journal of Biological Chemistryvol 287 no 48 pp 40598ndash40610 2012
[25] O Matsushita T Koide R Kobayashi K Nagata and AOkabe ldquoSubstrate recognition by the collagen-binding domainof Clostridium histolyticum class I collagenaserdquo The Journal ofBiological Chemistry vol 276 no 12 pp 8761ndash8770 2001
[26] M F French K A Mookhtiar and H E Van Wart ldquoLimitedproteolysis of type I collagen at hyperreactive sites by class Iand II Clostridium histolyticum collagenases complementarydigestion patternsrdquo Biochemistry vol 26 no 3 pp 681ndash6871987
[27] T Kin X Zhai T B Murdoch A Salam A M J Shapiro andJ R T Lakey ldquoEnhancing the success of human islet isolationthrough optimization and characterization of pancreas dissoci-ation enzymerdquo American Journal of Transplantation vol 7 no5 pp 1233ndash1241 2007
[28] T Kin X Zhai D OrsquoGorman and A M J Shapiro ldquoDetri-mental effect of excessive collagenase class II on human isletisolation outcomerdquo Transplant International vol 21 no 11 pp1059ndash1065 2008
[29] J H M Van Deijnen P T R Van Suylichem G H J Woltersand R Van Schilfgaarde ldquoDistribution of collagens type I typeIII and type V in the pancreas of rat dog pig andmanrdquoCell andTissue Research vol 277 no 1 pp 115ndash121 1994
[30] T F Linsenmayer E Gibney F Igoe et al ldquoType V collagenmolecular structure and fibrillar organization of the chicken1205721(V) NH2-terminal domain a putative regulator of cornealfibrillogenesisrdquo Journal of Cell Biology vol 121 no 5 pp 1181ndash1189 1993
[31] J K Mouw G Ou and V M Weaver ldquoExtracellular matrixassembly a multiscale deconstructionrdquo Nature Reviews Molec-ular Cell Biology vol 15 no 12 pp 771ndash785 2014
[32] F Delacoux A Fichard C Geourjon R Garrone and FRuggiero ldquoMolecular features of the collagenVheparin bindingsiterdquo The Journal of Biological Chemistry vol 273 no 24 pp15069ndash15076 1998
[33] S Ricard-Blum M Beraud N Raynal R W Farndale and FRuggiero ldquoStructural requirements for heparinheparan sulfatebinding to type V collagenrdquoThe Journal of Biological Chemistryvol 281 no 35 pp 25195ndash25204 2006
[34] W Henkel and K Dreisewerd ldquoCyanogen bromide peptides ofthe fibrillar collagens I III and V and their mass spectrometriccharacterization detection of linear peptides peptide glycosyla-tion and cross-linking peptides involved in formation of homo-and heterotypic fibrilsrdquo Journal of Proteome Research vol 6 no11 pp 4269ndash4289 2007
[35] K Gelse E Poschl and T Aigner ldquoCollagensmdashstructurefunction and biosynthesisrdquo Advanced Drug Delivery Reviewsvol 55 no 12 pp 1531ndash1546 2003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

8 Journal of Diabetes Research
[11] R Bauer J J Wilson S T L Philominathan D Davis OMatsushita and J Sakona ldquoStructural comparison of ColHand ColG collagen-binding domains from Clostridium his-tolyticumrdquo Journal of Bacteriology vol 195 no 2 pp 318ndash3272013
[12] J J Wilson O Matsushita A Okabe and J Sakon ldquoA bacterialcollagen-binding domain with novel calcium-binding motifcontrols domain orientationrdquoThe EMBO Journal vol 22 no 8pp 1743ndash1752 2003
[13] N Ohbayashi T Matsumoto H Shima et al ldquoSolution struc-ture of clostridial collagenase H and its calcium-dependentglobal conformation changerdquo Biophysical Journal vol 104 no7 pp 1538ndash1545 2013
[14] N Ohbayashi N Yamagata M Goto K Watanabe Y Yam-agata and K Murayama ldquoEnhancement of the structuralstability of full-length clostridial collagenase by calcium ionsrdquoApplied and Environmental Microbiology vol 78 no 16 pp5839ndash5844 2012
[15] U Eckhard P F Huesgen H Brandstetter and C M OverallldquoProteomic protease specificity profiling of clostridial collage-nases reveals their intrinsic nature as dedicated degraders ofcollagenrdquo Journal of Proteomics vol 100 pp 102ndash114 2014
[16] O Matsushita and A Okabe ldquoClostridial hydrolytic enzymesdegrading extracellular componentsrdquoToxicon vol 39 no 11 pp1769ndash1780 2001
[17] A Fujio K Murayama Y Yamagata et al ldquoCollagenase H iscrucial for isolation of rat pancreatic isletsrdquo Cell Transplanta-tion vol 23 no 10 pp 1187ndash1198 2014
[18] M K Gordon and R A Hahn ldquoCollagensrdquo Cell and TissueResearch vol 339 no 1 pp 247ndash257 2010
[19] MD Shoulders andR T Raines ldquoCollagen structure and stabil-ityrdquo Annual Review of Biochemistry vol 78 pp 929ndash958 2009
[20] D E Birk ldquoType V collagen heterotypic type IV collageninteractions in the regulation of fibril assemblyrdquoMicron vol 32no 3 pp 223ndash237 2001
[21] M A Weis D M Hudson L Kim M Scott J-J Wu and D REyre ldquoLocation of 3-Hydroxyproline residues in collagen typesI II III and VXI implies a role in fibril supramolecularassemblyrdquo Journal of Biological Chemistry vol 285 no 4 pp2580ndash2590 2010
[22] R J Wenstrup J B Florer E W Brunskill S M Bell I Cher-voneva and D E Birk ldquoType V collagen controls the initiationof collagen fibril assemblyrdquo Journal of Biological Chemistry vol279 no 51 pp 53331ndash53337 2004
[23] C Niyibizi P P Fietzek and M Van der Rest ldquoHumanplacenta type V collagens Evidence for the existence of an1205721(V)1205722(V)1205723(V) collagen moleculerdquo Journal of BiologicalChemistry vol 259 no 22 pp 14170ndash14174 1984
[24] C Yang A C Park N A Davis et al ldquoComprehensive massspectrometricmapping of the hydroxylated amino acid residuesof the1205721(V) collagen chainrdquoThe Journal of Biological Chemistryvol 287 no 48 pp 40598ndash40610 2012
[25] O Matsushita T Koide R Kobayashi K Nagata and AOkabe ldquoSubstrate recognition by the collagen-binding domainof Clostridium histolyticum class I collagenaserdquo The Journal ofBiological Chemistry vol 276 no 12 pp 8761ndash8770 2001
[26] M F French K A Mookhtiar and H E Van Wart ldquoLimitedproteolysis of type I collagen at hyperreactive sites by class Iand II Clostridium histolyticum collagenases complementarydigestion patternsrdquo Biochemistry vol 26 no 3 pp 681ndash6871987
[27] T Kin X Zhai T B Murdoch A Salam A M J Shapiro andJ R T Lakey ldquoEnhancing the success of human islet isolationthrough optimization and characterization of pancreas dissoci-ation enzymerdquo American Journal of Transplantation vol 7 no5 pp 1233ndash1241 2007
[28] T Kin X Zhai D OrsquoGorman and A M J Shapiro ldquoDetri-mental effect of excessive collagenase class II on human isletisolation outcomerdquo Transplant International vol 21 no 11 pp1059ndash1065 2008
[29] J H M Van Deijnen P T R Van Suylichem G H J Woltersand R Van Schilfgaarde ldquoDistribution of collagens type I typeIII and type V in the pancreas of rat dog pig andmanrdquoCell andTissue Research vol 277 no 1 pp 115ndash121 1994
[30] T F Linsenmayer E Gibney F Igoe et al ldquoType V collagenmolecular structure and fibrillar organization of the chicken1205721(V) NH2-terminal domain a putative regulator of cornealfibrillogenesisrdquo Journal of Cell Biology vol 121 no 5 pp 1181ndash1189 1993
[31] J K Mouw G Ou and V M Weaver ldquoExtracellular matrixassembly a multiscale deconstructionrdquo Nature Reviews Molec-ular Cell Biology vol 15 no 12 pp 771ndash785 2014
[32] F Delacoux A Fichard C Geourjon R Garrone and FRuggiero ldquoMolecular features of the collagenVheparin bindingsiterdquo The Journal of Biological Chemistry vol 273 no 24 pp15069ndash15076 1998
[33] S Ricard-Blum M Beraud N Raynal R W Farndale and FRuggiero ldquoStructural requirements for heparinheparan sulfatebinding to type V collagenrdquoThe Journal of Biological Chemistryvol 281 no 35 pp 25195ndash25204 2006
[34] W Henkel and K Dreisewerd ldquoCyanogen bromide peptides ofthe fibrillar collagens I III and V and their mass spectrometriccharacterization detection of linear peptides peptide glycosyla-tion and cross-linking peptides involved in formation of homo-and heterotypic fibrilsrdquo Journal of Proteome Research vol 6 no11 pp 4269ndash4289 2007
[35] K Gelse E Poschl and T Aigner ldquoCollagensmdashstructurefunction and biosynthesisrdquo Advanced Drug Delivery Reviewsvol 55 no 12 pp 1531ndash1546 2003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom


















