
PROGRAMA DE PÓS-GRADUAÇÃO EM GENÉTICA · 2019-10-25 · Marcela e Cadu, que mesmo não...
120
PROGRAMA DE PÓS-GRADUAÇÃO EM GENÉTICA UNIVERSIDADE FEDERAL DE PERNAMBUCO CENTRO DE CIÊNCIAS BIOLÓGICAS DEPARTAMENTO DE GENÉTICA Análise da expressão e investigação de mecanismos envolvidos com o controle da atividade de homólogos do fator de iniciação da tradução eIF4E ao longo do ciclo de vida de Leishmania amazonensis. MARIANA MARQUES COUTELO PEREIRA Recife, 2008
Transcript of PROGRAMA DE PÓS-GRADUAÇÃO EM GENÉTICA · 2019-10-25 · Marcela e Cadu, que mesmo não...

PPRROOGGRRAAMMAA DDEE PPÓÓSS--GGRRAADDUUAAÇÇÃÃOO EEMM GGEENNÉÉTTIICCAA UUNNIIVVEERRSSIIDDAADDEE FFEEDDEERRAALL DDEE PPEERRNNAAMMBBUUCCOO CCEENNTTRROO DDEE CCIIÊÊNNCCIIAASS BBIIOOLLÓÓGGIICCAASS
DDEEPPAARRTTAAMMEENNTTOO DDEE GGEENNÉÉTTIICCAA
Análise da expressão e investigação de mecanismos envolvidos com o controle da
atividade de homólogos do fator de iniciação da tradução eIF4E ao longo do ciclo de vida de
Leishmania amazonensis.
MMAARRIIAANNAA MMAARRQQUUEESS CCOOUUTTEELLOO PPEERREEIIRRAA
Recife, 2008

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
1
PPRROOGGRRAAMMAA DDEE PPÓÓSS--GGRRAADDUUAAÇÇÃÃOO EEMM GGEENNÉÉTTIICCAA UUNNIIVVEERRSSIIDDAADDEE FFEEDDEERRAALL DDEE PPEERRNNAAMMBBUUCCOO CCEENNTTRROO DDEE CCIIÊÊNNCCIIAASS BBIIOOLLÓÓGGIICCAASS
DDEEPPAARRTTAAMMEENNTTOO DDEE GGEENNÉÉTTIICCAA
Análise da expressão e investigação de mecanismos
envolvidos com o controle da atividade de homólogos do fator de iniciação da tradução eIF4E
ao longo do ciclo de vida de Leishmania amazonensis.
MMAARRIIAANNAA MMAARRQQUUEESS CCOOUUTTEELLOO PPEERREEIIRRAA
Dissertação apresentada ao
Programa de Pós-Graduação
em Genética da
Universidade Federal de
Pernambuco, como parte dos
requisitos necessários para a
obtenção do grau de Mestre
em Genética.
Orientador: Dr. Osvaldo
Pompílio de Melo Neto
Recife - PE 2008

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
2
UUNNIIVVEERRSSIIDDAADDEE FFEEDDEERRAALL DDEE PPEERRNNAAMMBBUUCCOO PPRROOGGRRAAMMAA DDEE PPÓÓSS--GGRRAADDUUAAÇÇÃÃOO EEMM GGEENNÉÉTTIICCAA
PARECER DA COMISSÃO EXAMINADORA DA DEFESA DE DISSERTAÇÃO DE MESTRADO DE
MMAARRIIAANNAA MMAARRQQUUEESS CCOOUUTTEELLOO PPEERREEIIRRAA
“Análise da expressão e investigação de mecanismos envolvidos com o controle da atividade de homólogos do fator de iniciação da
tradução eIF4E ao longo do ciclo de vida de Leishmania amazonensis.”
Área de Concentração: BIOLOGIA MOLECULAR
Recife - PE 2008

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
3
"Não existe triunfo sem perda,
não há vitória sem sofrimento,
não há liberdade sem
sacrifício."
(Senhor dos Anéis - O Retorno do Rei)

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
4
Ao meu marido, minha família e meus amigos...

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
5
AGRADECIMENTOS
Gostaria de agradecer primeiramente a Deus, por mais uma jornada conquistada, por mais essa vitória em minha vida, por mais um obstáculo vencido.
Ao meu Orientador, Dr. Osvaldo Pompílio de Melo Neto, pela confiança depositada e orientação para realização deste trabalho.
Aos meus melhores amigos do laboratório, Danona, Taminha, Christian, Franklin,
Mari Palma, Cariri, Mari Azevedo, Julie, (que também é da Microbiologia!!) e Cássia, pela experiência partilhada no dia a dia, por tornar o ambiente de trabalho tão agradável e pelo companheirismo fora do laboratório!! ADORO VOCÊS!!!! E não esquecendo minha amiga Poprilas, minha “mestra” das culturas, que era pra quem eu recorria quando comecei a manipular com células!!!!!
A todos os companheiros do CPqAM: Ronnie, Éden, Rodrigo, Marília, Rafaela,
Eduardo, Bruna, Ana Paula, Wladimir, Wellington, Marcelinho, Duschinka, Taty, Paloma e a todos os funcionários que proporcionaram a realização deste trabalho.
À Drª Regina Bressan e a Drª Cássia Docena, pela ajuda na microscopia confocal.
Aos Meus amigos da Faculdade Januana Teixeira, Cynthia Maria, Emanuel Borges e Ana Ruth Sampaio. A distância traz saudades, mas nunca o esquecimento... Obrigada pela amizade verdadeira e sincera...
Aos meus “compadres” e “comadres”, eternos amigos, Mariluce e Henrique e Marcela e Cadu, que mesmo não entendendo o meu trabalho, sempre torcem pelo meu sucesso.
Ao pessoal do 14º Batalhão de Infantaria Motorizado, literalmente a minha
segunda casa, pelos momentos maravilhosos no Clube dos Oficiais, onde eu passava fins de semanas relaxando as idéias!!! Em especial a Rose, Dani e Nastassja, por serem tão batalhadoras e por me darem o apoio necessário. Valeu meninas!!!
Em especial:
À minha família. Minha mãe Denise, meu pai José Augusto, meu irmão Daniel e
minha irmã Gabriela, pelo apoio e conforto dado nos momentos difíceis, pelas horas de alegrias e também pela torcida que eu sei que não é pouca. Agradeço também a minha tia Alba, meu tio Zé Carlos, meus primos e meu padrinho Breno, pelos momentos de descontração, afinal, a vida não é só trabalho!!!!!!
À minha outra família, minha sogra Dora, minha cunhada Mariana, meu cunhado
Paulo e minha cunhada Cristiane pelo apoio, confiança e carinho dados a mim. Muito obrigada!!!!!
E principalmente, queria agradecer ao meu marido e melhor amigo Expedito
Júnior, pelo amor, carinho, dedicação, compreensão, companheirismo, pelo interesse de querer entender e aprender o meu trabalho e principalmente pela paciência nestes quase 8 anos juntos. TE AMO MINHA VIDA!!!!!

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
6
SUMÁRIO
LISTA DE ABREVIATURAS .............................................................08
LISTA DE FIGURAS E TABELA .......................................................11
RESUMO........................................................................................13
1. INTRODUÇÃO............................................................................14
2. OBJETIVOS ...............................................................................17
2.1 Geral ...................................................................................17
2.2 Específicos ...........................................................................17
3. REVISÃO DA LITERATURA ........................................................18
3.1. Família Trypanosomatidae .....................................................18
3.2 O gênero Leishmania e a leishmaniose .....................................18
3.2.1 Taxonomia e transmissão ..................................................20
3.2.2 Aspectos clínicos da leishmaniose .......................................22
3.2.3 Impacto da doença ...........................................................24
3.2.4 Ciclo evolutivo..................................................................26
3.2.5 Cultivo dos parasitas.........................................................29
3.3 Características da biologia molecular dos tripanossomatídeos ......30
3.3.1 RNAs polimerases, promotores e Transcrição policistrônica ....31
3.3.2 Processamento de mRNAs – “trans-splicing e poliadenilação ..33
3.3.3 Cap4...............................................................................35 3.4 Controle da expressão gênica nos tripanossomatídeos................36
3.5 Biossíntese protéica ...............................................................38
3.5.1 Iniciação da tradução ........................................................38
3.5.1.1 Complexo eIF4F e PABP...............................................39
3.5.1.2 eIF4E: estrutura e função ............................................41
3.5.1.3 Modificações pós-traducionais – fosforilação do eIF4E .....43
3.5.2 Complexo EIF4F e PABP em tripanossomatídeos .................45
3.5.3 Eventos de fosforilação em tripanossomatídeos ..................48

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
7
4. REFERÊNCIAS BIBLIOGRÁFICAS...............................................50
5. MANUSCRITO DO ARTIGO CIENTÍFICO.....................................67
6. CONCLUSÕES ..........................................................................105
7. ABSTRACT...............................................................................106
8. ANEXOS ..................................................................................107
8.1 Instruções para autores......................................................107

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
8
LISTA DE ABREVIATURAS
EPB’s - Proteínas ligadoras ao 4E (“4E Binding Protein”)
AIDS/SIDA – Síndrome da imunodeficiência adquirida
(“Acquired Immunodeficiency Syndrome”)
AUG – Códon de iniciação da tradução
BSA – Albumina sérica bovina
DNA – Ácido Desoxirribonucléico
ECL – Detecção por quimioluminescência
(“Enhanced Chemioluminescence”)
eIFs – Fatores de iniciação da tradução eucarióticos
(“Eukaryotic Initiation factors”)
EPs - Prociclinas presente na forma procíclica
GEETs - Prociclinas presente na forma procíclica
GMP – Guanosina monofosfato
GP-63 - Glicoproteína de 63KDa
GST – Glutationa S Transferase
GTP – Guanosina Trifosfato
HIV – Vírus da imunodeficiência humana
(“Human Immunodeficiency Virus”)
IL-12 – Interleucina 12
IPTG – Isopropil-tio-β-D-galactopiranosídeo
kDa – Kilodalton
LB – Meio de cultura Luria Bertani
LC– Leishmaniose Cutânea
LCD – Leishmaniose cutâneo difusa
LMC– Leishmaniose Muco-Cutânea
LPG - complexo glicofosforoglicano
LV– Leishmaniose Visceral
m7GTP - cap monometilado

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
9
Map quinases - Mitogen-activated protein
Mnk1 – Quinase responsável pela fosforilação do eIF4E
Mnk2 – Quinase responsável pela fosforilação do eIF4E
mRNA – ácido ribonucléico mensageiro
mtHsp70 – proteína de choque térmico HSP70 mitocondrial
ORF – Seqüência aberta de leitura (“Open Reading Frame”)
PABP – Proteína de ligação a cauda poli-A (“Poly-A binding protein”)
PBS – Solução salina tamponada com fosfato
(“Phosphate Buffered Saline”)
PCR – Reação em cadeia da polimerase (“Polymerase Chain Reaction”)
PKDL – Leishmaniose dérmica pós Calazart
(“post Kala-azar dermal leishmaniasis”)
PVDF – Membrana de nitrocelulose
RNA – Ácido ribonucléico
RNAP - RNA polimerase
RNAPI - RNA polimerase I
RNAPII - RNA polimerase II
RNAPIII - RNA polimerase III
RRMs – Motivos de reconhecimento de RNA (“RNA recognition motif”)
rRNAs – RNAs ribossomais
SDS-PAGE – Gel de poliacrilamida em condições desnaturantes
(“Sodium Disulfate – Polyacrilamide Gel Eletrophoresis”)
SFB – Soro Fetal Bovino
SL – Seqüência spliced leader ou mini-éxon
snRNAs - RNAs nucleares
TBS – Solução salina tamponada com TRIS (“Tris buffered Saline”)
TCA – Ácido tricloroacético
TGFα - fator de transformação do crescimento
tRNA – Ácido ribonucléico transportador
tRNAi - tRNA iniciador

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
10
tRNAs - RNAs transportadores
UTR – Região não traduzível (“Untranslated Region”)
VSG – Glicoproteína Variante de Superficie
(“Variant Surface Glycoprotein”)

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
11
LISTA DE FIGURAS E TABELAS
Figuras da Revisão Bibliográfica
Figura 1. Vetores invertebrados da leishmaniose. ..............................19
Figura 2. Formas evolutivas do gênero Leishmania. ...........................20
Figura 3. Manifestações clínicas da leishmaniose. ...............................24
Figura 4. Mapa da distribuição geográfica dos países com maior
prevalência da leishmaniose. ............................................................25
Figura 5. Mapa da distribuição geográfica da Leishmaniose e da co-
infecção Leishmaniose/HIV. ..............................................................26
Figura 6. Ciclo de vida da Leishmania.. .............................................27
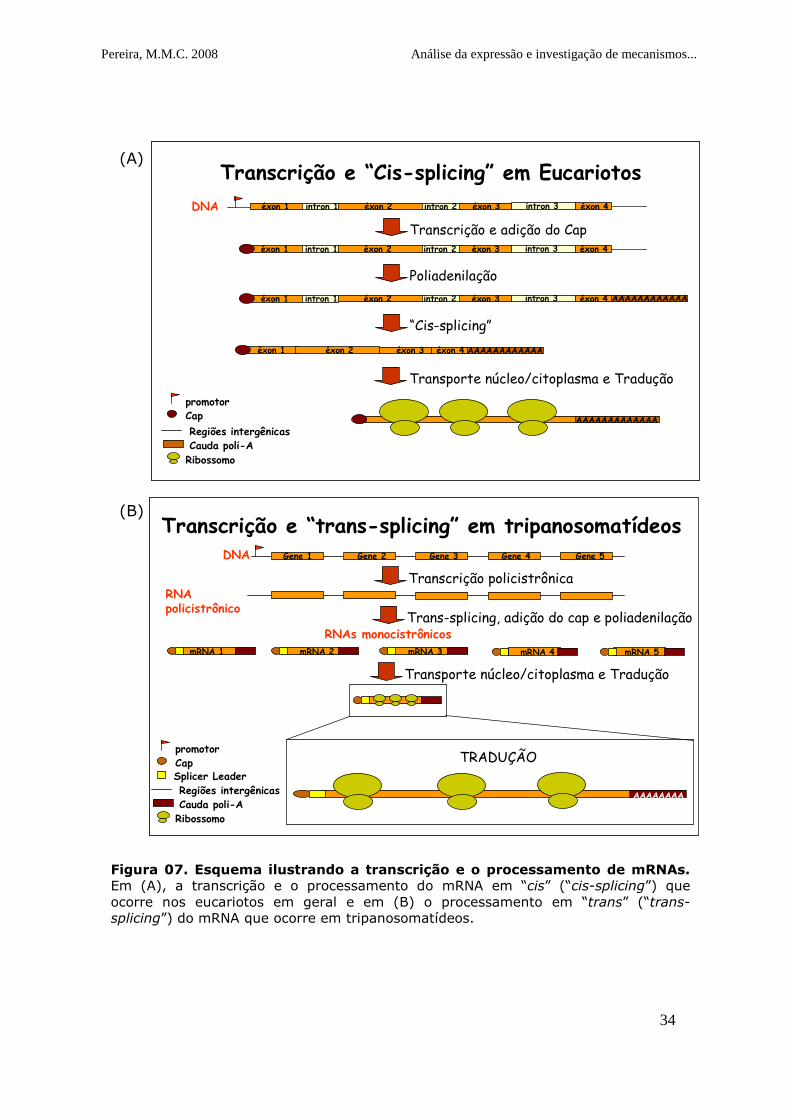
Figura 7. Esquema ilustrando a transcrição e o processamento de mRNAs
.....................................................................................................34
Figura 8. Esquema da iniciação da tradução cap-dependente. ......... ....41
Figuras do Artigo Científico
Figura 1. Padrão de crescimento e diferenciação de L. amazonensis em
cultivo in vitro .................................................................................98

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
12
Figura 2. Expressão dos quatro homológos do fator de iniciação da
tradução EIF4E e da proteína PABP1 durante as curvas de crescimento e
diferenciação de L. amazonensis........................................................99
Figura 3. Localização subcelular dos fatores EIF4E3 e EIF4E4 ............100
Figura 4. Avaliação do crescimento da L. amazonensis em 24h na
presença de meio novo ou condicionado, com e sem soro fetal bovino
(SFB)....... ...................................................................................101
Figura 5. Expressão dos fatores EIF4E3, EIF4E4 e a PABP1 nas culturas
crescidas por 24 horas na presença de meio novo ou condicionado, com e
sem soro fetal bovino (SFB)............................................................102
Figura 6. Análise do perfil de síntese protéica em células de L.
amazonensis em diferentes estágios de crescimento.. ........................103
Figura 7. Expressão dos fatores EIF4E3 e EIF4E4 em curvas de
crescimento realizadas na presença de inibidores de transcrição
(actinomicina D) ou tradução (cicloheximida)....................................104
Tabelas da Revisão Bibliográfica
Tabela 1. Quantificação dos homólogos eIF4E de Leishmania major na
fase promastigota............................................................................46

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
13
RESUMO
Os tripanossomatídeos são protozoários que causam manifestações
clínicas de impacto mundial e possuem características bioquímicas
peculiares que os diferem dos demais eucariotos. Sua expressão gênica é
regulada em nível pós-transcricional, provavelmente no estágio de
iniciação da síntese protéica. A tradução cap-dependente é iniciada
através da ligação do fator de iniciação eIF4E, pertencente ao complexo
eIF4F, ao cap presente na extremidade 5’ dos mRNAs. Foram identificados
múltiplos homólogos do fator EIF4E nestes parasitas, que provavelmente
estão associados ao seu complexo ciclo de vida. Neste trabalho, nós
buscamos entender como ocorre a expressão de quatro homólogos deste
fator (EIF4E1-4) durante o ciclo de vida de Leishmania amazonensis. Para
tal, foram realizadas curvas a fim de se obter as três principais formas
evolutivas do parasita. Todos os homólogos estão presentes ao longo da
curva e os EIF4E2-4 apresentam múltiplas isoformas, sugerindo
modificações pós-traducionais por fosforilação. Destes, as isoformas dos
EIF4E3 e 4 apresentaram expressão constrastante que não interferem
com sua localização subcelular, citoplasmática. A ausência de soro fetal
bovino no meio induziu uma desfosforilação prematura do EIF4E4 e uma
“hiperfosforilação” do EIF4E3, associados à falta de crescimento celular.
EIF4E3 “desfosforilado” e EIF4E4 “fosforilado” estão associados a uma
maior atividade de tradução. A presença de um inibidor de transcrição
influenciou no padrão de expressão dos dois homólogos, porém um
inibidor de tradução afetou apenas o EIF4E3. Estes resultados indicam que
os homólogos de eIF4E atuam de forma distinta na célula e que a
fosforilação deve ter um papel significante na regulação destes fatores em
Leishmania e na regulação da síntese de proteínas como um todo.
Palavras-chave: Leishmania amazonensis, EIF4E, fosforilação

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
14
1. INTRODUÇÃO
Os tripanosomatídeos, pertencentes à ordem Kinetoplastida, são
protozoários eucariotos flagelados cujos gêneros Leishmania e
Trypanosoma são responsáveis por doenças de impacto mundial como a
Doença do Sono (T. brucei), Doença de Chagas (T. cruzi) e várias formas
de leishmaniose (Leishmania sp.) (www.who.int). As formas clínicas da
leishmaniose são particularmente diversas e representam um complexo de
doenças que atingem anualmente 1,5 a 2 milhões de pessoas no mundo
todo, e cerca de 350 milhões de pessoas encontram-se em áreas de risco.
Este quadro é preocupante e desperta o interesse para pesquisas mais
aprofundadas sobre estes parasitas, tanto em nível celular como
molecular.
As espécies patogênicas de tripanosomatídeos possuem um
complicado ciclo de vida, alternando entre o hospedeiro mamífero e o
inseto vetor hematófago. No gênero Leishmania, a forma promastigota,
flagelada extracelular, é a forma predominante no trato digestivo dos
vetores invertebrados, fêmeas de dípteros flebotomíneos. Durante o
repasto sanguíneo, estas formas são transmitidas ao hospedeiro
vertebrado, quando são fagocitados por células do sistema fagocítico
mononuclear, e se transformam em amastigotas, parasitas intracelulares
obrigatórios sem flagelo aparente.
Do ponto de vista da sua biologia, os tripanosomatideos são
eucariotos que possuem características bioquímicas e moleculares
relevantes que os diferenciam dos demais eucariotos. Em especial se
destaca a síntese dos seus mRNAs, realizada de forma policistrônica, onde
dezenas de genes podem ser transcritos em um único e longo mRNA
precursor cuja transcrição é controlada por um único promotor. Estes pré-
mRNAs são processados pelo mecanismo de trans-splicing, que divide este

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
15
transcrito único em múltiplos mRNAs maduros cada um representando
uma seqüência codificadora individual. A transcrição policistrônica e o
escasso número de promotores nestes parasitas impede uma regulação
individual da síntese de mRNAs, como ocorre em outros eucariotos. Desta
forma, acredita-se que boa parte dos mecanismos de regulação da sua
expressão gênica ocorram em nível pós-transcricional, como durante a
tradução dos mRNAs em proteínas (Clayton, 2002).
A biossíntese protéica ou tradução é um dos processos mais
importantes para a sobrevivência de todos os organismos. Esse processo
nos tripanosomatídeos ainda é pouco conhecido e precisa de mais estudos
para seu total entendimento. A etapa de iniciação é a fase mais crítica
deste processo, sujeita a diferentes mecanismos de regulação, e onde se
define quando e com que intensidade os diferentes mRNAs celulares serão
utilizados na síntese de proteínas. Esta etapa compreende o
reconhecimento do mRNA maduro pela subunidade 40S do ribossomo e a
identificação do códon AUG, que indica o início da seqüência que codifica
para as proteínas propriamente, sendo mediada por proteínas conhecidas
como fatores de iniciação da tradução (eIFs). Entre esses fatores destaca-
se o eIF4E, a proteína que reconhece a estrutura cap na extremidade 5’
do mRNA. Como parte do complexo heterotrimérico eIF4F, junto com o
eIF4A e o eIF4G, esta proteína é a principal responsável pelo
reconhecimento dos mRNAs para a tradução (Gingras et al., 1999).
A iniciação da tradução é bem estudada em leveduras, plantas e
vertebrados, porém pouco se sabe sobre este processo nos
tripanosomatídeos. Vários homólogos de fatores de iniciação da tradução
já foram identificados e suas seqüências se encontram disponíveis em
bancos de dados de acesso público. Para entender a iniciação da tradução
nos tripanosomatídeos, nosso grupo buscou identificar e caracterizar
nestes organismos homólogos para as várias subunidades do eIF4F.
Múltiplos homólogos em potencial foram identificados para as subunidades

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
16
eIF4E e eIF4G em bancos de seqüências genômicas. Iniciando sua
caracterização funcional, observou-se, no caso dos homólogos de eIF4E,
que estes variam em diferentes aspectos como a afinidade de ligação ao
cap, níveis de expressão e interação com homólogos específicos de eIF4G.
Os resultados obtidos até o momento sugerem um alto grau de
complexidade na iniciação da tradução desses parasitas, o que pode
refletir uma adaptação ao seu complexo ciclo de vida (Dhalia et al., 2005).
Com a identificação de múltiplos homólogos da subunidade eIF4E
em Leishmania, é de se esperar que nem todos atuem diretamente na
iniciação da tradução. Alternativamente, é possível que um ou mais
possam estar atuando em fases específicas do complexo ciclo de vida do
parasita. Neste caso sua expressão seria variável de acordo com estas
diferentes fases. Este trabalho teve como ponto principal, então,
investigar a expressão de quatro homólogos de eIF4E de L. amazonensis
(EIF4E1-4) em formas representativas de diferentes fases do seu ciclo de
vida e também em células submetidas a diferentes condições de estresse.
Os resultados obtidos sugerem um novo elemento de complexidade no
controle da síntese protéica nos tripanosomatídeos que envolve
modificações pós-traducionais em fatores de tradução selecionados.

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
17
2. OBJETIVOS
GERAL
Analisar e comparar a expressão de quatro homólogos do fator de
iniciação da tradução eIF4E durante o ciclo e vida de Leishmania
amazonensis, bem como investigar mecanismos que estejam envolvidos
com o controle de expressão/atividade destas proteínas.
ESPECÍFICOS
1. Padronizar condições de crescimento e diferenciação de Leishmania
amazonensis em cultura e obtenção de extratos protéicos de diferentes
fases do seu ciclo de vida;
2. Realizar ensaios de “western-blot” para comparar a expressão dos
homólogos EIF4E1-4, utilizando como controle o fator de iniciação EIF4A1
e as proteínas ribossomais P0 e L19 (validando quantidade celular) e a
proteína mtHsp70 (validando a diferenciação celular);
3. Realizar ensaios de imunocitoquímica a fim de observar mudanças de
localização subcelular dos homólogos ao longo do ciclo de vida do
parasita;
4. Avaliar padrões de expressão/modificação dos homólogos em células
submetidas a condições especiais de cultura, como ausência de soro fetal
bovino, induzindo estresse celular.
5. Realizar ensaios de marcação metabólica a fim de comparar mudanças
no padrão de expressão protéica do parasita com alterações no padrão de
expressão/modificação de homólogos específicos;
6. Analisar o efeito de inibidores de transcrição (actinomicina D) e
tradução (cicloheximida) no padrão de expressão/modificação dos
homólogos em estudo;

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
18
3. REVISÃO DA LITERATURA
3.1 Família Trypanosomatidae
A família Tripanosomatidae é formada por organismos unicelulares
que divergiram muito cedo na linhagem evolutiva dos demais eucariotos,
caracterizados por apresentarem corpo alongado dotado de um único
flagelo terminal e um único núcleo, geralmente central (Schmidt e
Roberts, 1996). A presença de uma estrutura em forma de disco achatado
contendo o DNA mitocondrial, conhecida como cinetoplasto, é uma
característica destes protozoários que permite agrupá-los na ordem
Kinetoplastida (Vickerman and Coombs, 1999). Esta família possui nove
gêneros, onde todos os membros são exclusivamente parasitas que
apresentam um complexo ciclo de vida passando por vários estágios
morfológicos e envolvendo mais de um hospedeiro (Singh, 2006).
Os gêneros Trypanosoma e Leishmania merecem destaque devido à
sua importância médica e veterinária, pois são responsáveis por
enfermidades de impacto mundial, como a Doença de Chagas
(Trypanosoma cruzi), Doença do Sono (Trypanosoma brucei) e as várias
formas de Leishmaniose (Leishmania sp.). Em todo o mundo, cerca de
350 milhões de pessoas se encontram em áreas de risco e milhares de
pessoas morrem anualmente devido a essas enfermidades (www.who.int).
3.2 O Gênero Leishmania e a Leishmaniose
O gênero Leishmania é representado por pelo menos 20 agentes
etiológicos causadores de leishmaniose, enfermidade que afeta o mundo
todo, sendo mais frequente na América do Sul, América Central, África
Central, Oriente Médio e no sudoeste da Ásia (www.who.int). A

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
19
leishmaniose compreende um grupo de doenças que expõe amplas
manifestações clínicas em humanos, cujo tipo não vai depender apenas da
espécie que causou a infecção, mas também da saúde geral e da
maquinaria genética do indivíduo infectado (Alexander et al., 1999).
A leishmaniose é transmitida por cerca de 30 espécies de
flebotomíneos, insetos dípteros pertencentes aos gêneros Phlebotomus e
Lutzomya, encontrados no Velho e Novo Mundo respectivamente (FIGURA
1). As espécies de Leishmania apresentam ciclo digenético com duas
formas vegetativas diferentes: a forma extracelular móvel, promastigota,
que é encontrada no vetor invertebrado e a forma obrigatoriamente
intracelular, amastigota, capaz de se multiplicar em vários hospedeiros
vertebrados, infectando células da linhagem fagocítica. A forma
promastigota é dividida em várias formas de desenvolvimento conhecidas
como promastigotas procíclica, nectomona, leptomona, haptomona,
paramastigota e metacíclica. Porém, as formas observadas em meio de
cultura incluem a forma procíclica, que é a forma não infectiva, mas
apresenta uma alta taxa de multiplicação, e a forma metacíclica, que é a
forma infectiva, mas que não apresenta divisão celular (Shaw, 1994;
Alexander et al., 1999; Gossage et al., 2003; Neves, 2005; Besteiro et al.,
2007) (FIGURA 2).
Na sua forma metacíclica o parasita tem sido descrito como
(A) (B)(A) (B)
Figura 1. Vetores invertebrados da leishmaniose. Em (A) o inseto do gênero Phlebotomus causador da doença no Velho Mundo e em (B) o gênero Luztomya, causador da doença no Novo Mundo. Fonte: http://www.who.int/leishmaniasis/disease_epidemiology/en/index.html

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
20
pequeno, com um flagelo relativamente grande, aparecendo em maior
número “in vitro” na fase estacionária. Esta forma é tida como a fase
infectante da Leishmania, embora as formas promastigota procíclica e
amastigota também sejam capazes de infectar o hospedeiro (Teixeira et
al., 2002). A forma procíclica é considerada uma célula ovóide, bastante
móvel e que possui atividade replicativa. As formas promastigotas
possuem uma variabilidade muito grande nas suas dimensões, variando
entre 10 µm a 40 µm de comprimento por 1,5 µm a 3,0 µm de diâmetro.
Já a forma amastigota intracelular é aflagelada, arredondada e possui
aproximadamente de 1,5 a 3 µm de comprimento por 3 a 6 µm de
diâmetro (Neves, 2005) (FIGURA 2). Vários genes específicos da
metaciclogênese têm sido mostrados, contudo, os mecanismos
moleculares que levam à iniciação e ao controle deste mecanismo vital
ainda permanecem indefinidos (Cunningham et al., 2001).
3.2.1 Taxonomia e Transmissão
As espécies de Leishmania foram divididas em dois subgêneros
taxonomicamente distintos, Viannia e Leishmania, baseadas em
similaridades moleculares (análise dos fragmentos do DNA do
Figura 2. Formas evolutivas do gênero Leishmania. Em (A e B) se encontra a forma promastigota encontrada no trato digestivo do hospedeiro invertebrado, onde (A) é a forma procíclica e (B) é a forma metacíclica. Em (C) se encontra a forma amastigota presente no hospedeiro vertebrado. Fonte: Besteiro et al., 2007.
(A)
(B) (C)

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
21
cinetoplasto, cortado por enzimas de restrição e PCR com "primers"
espécie-específicos), bioquímicas (análise isoenzimática) e imunológicas
(separação empregando anticorpos monoclonais) (Pearson et al., 2000).
As espécies foram agrupadas em complexos que recebem o nome da
espécie mais importante e possuem distribuição geográfica, epidemiologia
e tendência a visceralização que lhe são peculiares (Pearson et al., 2000).
O subgênero Viannia compreende o complexo L. brasiliensis, sendo as
principais espécies L. brasiliensis, L. guyanensis, L. panamensis e L.
peruviana. Já o subgênero Leishmania compreende os complexos L.
donovani (L. donovani, L. infantum e L. chagasi), L. mexicana (sendo as
espécies principais L. mexicana, L. amazonensis e L. venezuelensis) e L.
hertigi (este último complexo não parasita o homem). O subgênero
Leishmania ainda possui três espécies de importância clínica, mas que não
foram agrupadas em complexos: L. major, L. tropica e L. aethiopica
(Neves, 2005).
A transmissão mais comum da leishmaniose é através da picada da
fêmea do inseto hematófago, que ao fazer o repasto sanguíneo, passa a
forma promastigota da Leishmania para o hospedeiro mamífero, seja ele o
homem, o cão ou algum animal silvestre. Os hospedeiros vertebrados são
primariamente mamíferos, tais como cães e várias espécies de roedores
(Schmidt e Roberts, 1996). Porém são conhecidas também entre
edentados (tatu, tamanduá, preguiça), marsupiais (gambá), primatas e o
homem (Neves, 2005). Estes, por sua vez, atuam como reservatório de
infecção, contaminando insetos que não carregam o parasita, tornando
cíclico o seu modo de transmissão.
Outros modos de transmissão também podem ocorrer, mas com
menor freqüência, tais como: transfusão sanguínea, compartilhamento de
seringas através do uso de drogas injetáveis, transmissão congênita,
transmissão sexual e acidentes laboratoriais.
(a) Transfusão sanguínea: têm sido reportados casos em muitos

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
22
países, inclusive no Brasil. O parasita precisa estar no sangue periférico do
doador, sobreviver ao processo de estocagem do banco de sangue e
infectar o receptor (Neves, 2005).
(b) Compartilhamento de seringas: Em 1993, foram detectados
200 casos de HIV associado com leishmaniose na Espanha, e mais de
85% ocorreu com usuários de drogas (Alvar et al., 1997).
(c) Transmissão congênita: pelo menos 10 casos deste tipo de
transmissão foram reportados na literatura, sendo a maior parte dos
casos na Índia. A infecção deve ocorrer durante a troca do sangue
materno no tempo da passagem do feto no momento do nascimento
(Singh, 2006).
(d) Transmissão sexual: pode ser transmitida pela urina e fluidos
prostáticos através de pacientes contaminados (Singh, 2006).
(e) Acidentes laboratoriais: foram reportados casos de
contaminação através da auto-inoculação de materiais perfuro-cortantes
tanto com a forma promastigota como a forma amastigota (Neves, 2005).
3.2.2 Aspectos Clínicos da Leishmaniose
A leishmaniose pode ser uma doença lentamente progressiva e
demorar até sete anos para se tornar clinicamente aparente. Nesse caso,
seus sinais clínicos (tosse, diarréia, icterícia e sangramentos) são
freqüentemente confundidos com outras inúmeras doenças, dificultando o
diagnóstico da leishmaniose e retardando a sua identificação (Slappendel
e Ferrer, 1998; McConkey et al., 2002). Dependendo da espécie de
Leishmania, da localização geográfica e da resposta imune do hospedeiro,
a infecção em humanos pode resultar em várias formas clínicas de
leishmaniose particularmente diversas (www.dpd.cdc.gov).
No Homem a leishmaniose ocorre em pelo menos quatro grandes

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
23
formas: leishmaniose cutânea, difuso cutânea, mucocutânea e visceral.
(a) Leishmaniose Cutânea ou tegumentar (LC): considerada a
forma mais comum da doença, é caracterizada por lesões múltiplas
localizadas na pele, principalmente na face, braços e pernas, sendo elas
agudas ou crônicas. Embora as lesões possam freqüentemente se curar
espontaneamente, estas podem criar graves incapacidades permanentes e
cicatrizes. É causada pelas espécies L. major, L. tropica, L. mexicana, L.
amazonensis e espécies do subgênero Viannia, principalmente a L.
brasiliensis (Neves, 2005; Singh, 2006) (Figura 3A).
(b) Leishmaniose Mucocutânea (LMC): inicialmente começa como
uma lesão cutânea, que pode então se espalhar causando lesões
mucocutâneas que são frequentemente resistentes ao tratamento ou a
cura. Esta infecção é causada pelas espécies do subgênero Viannia, como
L. brasiliensis, L. panamensis ou L. guyanensis. (Cohen-Freue et al.,
2007). (Figura 3B).
(c) Leishmaniose cutâneo-difusa (LCD): ocorre em indivíduos com
deficiência na resposta imune celular. Severamente, causa lesões
disseminadas que se assemelham a lesões de lepra, nunca se curam
espontaneamente e estão sujeitas a recaídas após o tratamento (Alvar et
al., 1997; Desjeux, 2004) (Figura 3C).
(d) Leishmaniose Visceral (LV): Também conhecida como “Kala
azar” ou febre dumdum. Tem um período de incubação de vários meses a
vários anos, sendo a forma mais severa da leishmaniose, tem uma taxa
de mortalidade de quase 100%, se não tratada, e difere da leishmaniose
cutânea por envolver os órgãos internos. É causada por espécies do
complexo L. donovani, caracterizando-se por febres ondulantes, perda de
peso, esplenomegalia, hepatomegalia e/ou linfoadenopatias e anemia

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
24
(Figura 3D). Após a recuperação, os pacientes podem desenvolver uma
forma crônica chamada leishmaniose dérmica pós-calazar (PKDL – “post
Kala-azar dermal leishmaniasis”), que normalmente requer um tratamento
longo e caro (Desjeux, 2004; Neves, 2005) (Figura 3E).
(A) (B) (C) (D) (E)
L. infantum é considerado o principal agente associado à
leishmaniose canina, e por isso, as infecções em cães são freqüentemente
consideradas como visceral. Porém, os cães podem adquirir tanto a
leishmaniose visceral quanto a cutânea (Slappendel e Ferrer, 1998).
3.2.3 Impacto da Doença
A leishmaniose é considerada a segunda principal doença causada
por protozoários no mundo, perdendo em incidência apenas para a Malária
(www.vet.uga.edu). Atualmente, a leishmaniose é considerada endêmica
em 88 países, 72 dos quais são países em desenvolvimento, e, dentro
destes, 13 estão entre os países menos desenvolvidos do mundo
(Desjeux, 2004) (Figura 4).
Figura 3. Manifestações clínicas da leishmaniose. (A) Leishmaniose cutânea ou tegumentar; (B) Leishmaniose mucocutânea; (C) Leishmaniose cutânea-difusa; (D) Leishmaniose visceral; (E) Leishmaniose dérmica pós-calazar (PKDL). Fonte: http:// www.who.int/tdr

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
25
A leishmaniose é uma das infecções oportunistas que atacam os
indivíduos infectados por HIV, e a maioria das co-infecções envolve a
forma de leishmaniose visceral. Ela acelera a manifestação da AIDS
acumulada por imuno-supressão e pelo estímulo à replicação do vírus.
Também pode alterar infecções que são assintomáticas em sintomáticas.
Além disso, a leishmaniose visceral pode ser propagada junto com o vírus
da AIDS por via endovenosa, a partir do uso de agulhas por drogas
intravenosas. Até 1999, 31 países relataram a existência de uma co-
infecção Leishmania/ HIV. (www.who.int) (Figura 05).
(A) (B)
No Brasil, já foram descritas seis espécies de Leishmania causadoras
de leishmaniose cutânea, sendo L. brasiliensis a mais freqüente. No
período de 1970 a 2001, o número de casos variou de 3.000 para 37.000
e a partir da década de 90, os casos apresentaram um pequeno aumento,
com grande variação anual. (FUNASA, 2002a). A Leishmaniose visceral,
Leishmaniose CutâneaPaíses mais endêmicos (90% dos casos)
Países mais endêmicos - Leishmaniose cutânea
Leishmaniose CutâneaPaíses mais endêmicos (90% dos casos)
Países mais endêmicos - Leishmaniose cutânea
Leishmaniose VisceralPaíses mais endêmicos (90% dos casos)
Países mais endêmicos - Leishmaniose visceral
Leishmaniose VisceralPaíses mais endêmicos (90% dos casos)
Países mais endêmicos - Leishmaniose visceral
Figura 4. Mapa da distribuição geográfica dos países com maior prevalência da leishmaniose. (A) ilustra países que possuem maior prevalência da leishmaniose cutânea. Em (B), os países com maior prevalência em leishmaniose visceral. Fonte: http://www.who.int/leishmaniasis/leishmaniasis_maps/en/index2.html

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
26
causada pela L. chagasi, atinge 19 estados especialmente da região
Nordeste, onde se concentram mais de 90% dos casos humanos da
doença. Há focos importantes também nas regiões Norte, Sudeste e
Centro-Oeste, sendo que nas duas últimas décadas tem havido um
crescente aumento da incidência, com uma média anual nos últimos cinco
anos de 3500 casos (FUNASA, 2002b).
3.2.4 Ciclo Evolutivo
O ciclo de vida da Leishmania envolve uma alternância entre o
hospedeiro vertebrado e o inseto vetor. E como resultado o parasita
encontra extremas mudanças ambientais e reponde a essas mudanças
através de diferenciação em formas altamente adaptadas que o permite
LeishmanioseCo-infecçãoLeishmanioseCo-infecção
Figura 5. Mapa da distribuição geográfica da Leishmaniose e da co-infecção Leishmaniose/HIV. O azul claro ilustra a distribuição da leishmaniose no mundo, enquanto que o azul escuro mostra os pontos onde ocorre a co-infecção Leishmaniose/HIV. Fonte: http://www.who.int/csr/resources/figure231.gif

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
27
invadir e proliferar dentro de seus hospedeiros (Barak et al., 2005). As
formas amastigotas de Leishmania são encontradas parasitando células do
sistema mononuclear do hospedeiro vertebrado, principalmente
macrófagos residentes na pele, sobrevivendo e se multiplicando nestas
células. O inseto vetor ao fazer o repasto sanguíneo ingere, junto com o
sangue, macrófagos parasitados por estas formas. Ao chegarem ao
estômago, os macrófagos se rompem e liberam as amastigotas, que
sofrem uma divisão binária e se transformam rapidamente em
promastigotas dentro do sangue que está envolto por uma membrana
peritrófica secretada pelas células do estômago do inseto. Este membrana
se rompe entre o terceiro e quarto dia, liberando as formas promastigotas
(Neves, 2005) (Figura 6).
Um padrão diferente de desenvolvimento de promastigotas é
Figura 6. Ciclo de vida da Leishmania. Foto ilustrando as formas encontradas nos hospedeiros invertebrados (promastigotas) e vertebrados (amastigotas) e como ocorre a transmissão entre os hospedeiros. Fonte: http:// www.who.int/tdr/diseases/leish/lifecycle.html

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
28
observado nos dois subgêneros, Leishmania e Viannia, onde os membros
do primeiro subgênero desenvolvem-se exclusivamente no intestino médio
e posterior dos vetores (desenvolvimento suprapilário), enquanto que no
segundo desenvolvem-se primeiramente no intestino posterior dos
vetores, e depois migram para o intestino médio e anterior
(desenvolvimento peripilário). (Lainson e Shaw, 1987; Gossage et al.,
2003). Após esta etapa, em ambos os subgêneros, o parasita se
transforma em paramastigotas, e a partir deste ponto ocorre a
metaciclogênese, onde as células migram para a parte anterior do tubo
digestivo do vetor e atingem o estágio infectivo, se transformando em
promastigotas metacíclicas. O tempo necessário para a ocorrência destas
mudanças varia de três a cinco dias, dependendo da espécie de
Leishmania (Neves, 2005).
A transmissão para o hospedeiro vertebrado se dá quando o inseto
infectado realiza o repasto sanguíneo. Ao ingerir o sangue do hospedeiro
vertebrado, as formas metacíclicas são introduzidas no local e em quatro
a oito horas os parasitas são englobados pelos macrófagos teciduais. As
formas amastigotas são encontradas 24 horas após a fagocitose, se
dividem por divisão binária e rompem a membrana do macrófago, sendo
liberadas novas amastigotas que são novamente fagocitadas, iniciando no
local uma reação inflamatória (Neves, 2005) (Figura 6).
As formas promastigotas metacíclicas utilizam mecanismos que
permitem resistir à ação de elementos do soro do vertebrado,
principalmente do sistema complemento, onde proteínas de superfície do
parasita como a GP-63 (Glicoproteína de 63KDa) e a LPG (complexo
lipofosfoglicano) possuem papéis fundamentais (Neves, 2005). Além
disso, eles são injetados no local da picada juntamente com potentes
vasodilatadores que produzem eritremas de longa duração, facilitando a
sua penetração na corrente sanguínea (Singh, 2006). O pH desempenha
um papel central na transformação de promastigotas em amastigotas, e

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
29
as formas amastigotas apresentam uma maior atividade metabólica
estando em ambiente ácido, embora o citoplasma da amastigota seja
regulado para se manter em um pH próximo do neutro (Burchmore e
Barrett, 2001).
3.2.5 Cultivo dos Parasitas
A maior parte das informações metabólicas, bioquímicas e biológicas
da Leishmania é devido a inúmeros estudos na forma promastigota, pois
suas culturas são de fácil manutenção in vitro e esta pode ser obtida
facilmente em grande quantidade. Contudo, a forma amastigota é
responsável pelas manifestações clínicas nos hospedeiros vertebrados e os
quimioterápicos e vacinas precisam ser desenvolvidos contra esse estágio
do ciclo de vida do parasita. A forma amastigota, entretanto, tem seu
estudo restrito devido a limitações no seu cultivo in vitro e a dificuldades
na obtenção de grandes quantidades de células viáveis, e livres de
contaminação, a partir de hospedeiros de laboratório. Além disso,
amastigotas isoladas a partir de tecidos infectados representam
populações heterogêneas e, durante o processo de infecção,
presumivelmente, diferem em relação ao seu estágio de desenvolvimento
(Joshi et al., 1993; Debrabant et al., 2004).
A diferenciação de promastigotas em amastigotas em cultura
(axênicas) tem sido reportada utilizando apenas uma temperatura mais
elevada (32-37ºC) ou em combinação com uma diminuição do pH do meio
(Bates, 1993; Zilberstein e Shapira, 1994). Em um estudo com curvas de
crescimento em promastigotas para observação do processo de
diferenciação celular, foi visto que quanto maior o número de células
metacíclicas presentes no meio, maior a percentagem de diferenciação
das células em amastigotas axênicas. Neste mesmo estudo foi observado

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
30
também que o número de repiques realizado está diretamente ligado com
a diminuição do aparecimento de células infecivas metacíclicas. Numa
cultura com 73% de células metacíclicas, a diferenciação em amastigotas
axênicas conseguida foi de 100% (Cysne-Finkelstein et al., 1998). Outros
estudos mostraram que amastigotas axênicas podem ser virulentas “in
vitro” e “in vivo” e infectar células de hamsters, camundongos e
macrófagos, porém, a diferenciação em meio de cultura diminui a
virulência a longo prazo e produz células atenuadas (Gupta et al., 2001;
Debrabant et al., 2004). Alguns estudos abordam vários critérios no
intuito de mostrar similaridades entre a amastigota in vitro e in vivo,
porém, ainda restam controvérsias sobre a reprodutibilidade e/ou o tempo
certo da transformação do parasita (Gupta et al., 2001; Teixeira et al.,
2002, Debrabant et al., 2004; Cohen-Freue et al., 2007).
3.3 Características da biologia molecular dos Tripanosomatídeos
Os tripanossomatídeos são organismos bastante atípicos que
divergiram precocemente na evolução dos eucariotos e apresentam
características moleculares únicas nos mecanismos de síntese e
processamento de seus mRNAs. Nestes organismos os genes codificadores
de proteínas são organizados em blocos numa mesma fita de DNA, não
possuem íntrons (com exceção do gene da poli-A polimerase) e são
transcritos de forma policistrônica a partir de sequencias promotoras não
definidas. Esses policístrons formam mRNAs monocistrônicos maduros
através do mecanismo de processamento em “trans” (“trans-splicing”),
que difere do processamento em “cis” (“cis-splicing”), típico da maioria
dos eucariotos (Myler e Stuart, 2000; Clayton, 2002; Campbell et al.,
2003; Liang et al., 2003).
O processamento em “trans” do pré-mRNA policistrônico acrescenta,

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
31
na região 5’, uma sequência denominada spliced leader (SL), de 39
nucleotídeos, que é característica apenas de mRNAs dos
tripanosomatídeos tratando-se de um substrato comum utilizado em todas
as reações desse processamento (Liang et al., 2003). Na extremidade 5’
do spliced leader é encontrada uma estrutura cap modificada, denominado
de cap4. Em outros organismos, foi observado que esta estrutura possui
um importante papel na estabilidade e transporte dos mRNAs e na
iniciação da tradução (Liang et al., 2003; Gu e Lima, 2005).
Os tripanossomatídeos possuem escassez de promotores para a RNA
polimerase II (RNAPII), e, com isso, não há nenhuma regulação
quantitativa ao nível de transcrição para a maioria dos genes que
codificam proteínas (Campbell et al., 2003). A conseqüência principal
destes mecanismos atípicos de síntese e processamento dos mRNAs nos
tripanosomatídeos é que a regulação da expressão de genes é
predominantemente pós transcricional, ou seja, ao nível de controle da
estabilidade dos mRNAs e controle da sua utilização na síntese de
proteínas (Clayton, 2002).
3.3.1 RNA Polimerases, Promotores e Transcrição Policistrônica
Os tripanosomatídeos possuem cópias das três RNA polimerases
(RNAP) eucarióticas, altamente conservadas. Foi observado que a RNAPI
atua caracteristicamente transcrevendo os genes dos pré-RNAs
ribossomais (pré-rRNAs) 18S, 5,8S e 28S. Em T. brucei, porém, esta
enzima também transcreve dois tipos de mRNAs que são estágio
específicos e codificam proteínas de superfíce: a variante protéica da
glicoproteína, VSG, da forma sanguínea; e as prociclinas EPs e GEETs da
forma procíclica (Gunzl et al., 2003). Portanto, T. brucei é o único
organismo reportado até agora que utiliza a RNAPI para transcrever genes

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
32
que codificam proteínas (Devaux et al., 2006).
A RNAPII atua na transcrição dos mRNAs, como em outros
eucariotos, e também na transcrição do mRNA precursor que codifica a
seqüência SL, também chamada de mini-éxon (Gilinger e Bellofatto,
2001). Já a RNAPIII transcreve RNAs transportadores (tRNAs), o rRNA 5S
e pequenos RNAs nucleares (snRNAs) ricos em uridina (Palenchar e
Bellofatto, 2006). Ainda não foram caracterizados promotores para a
RNAPII, com exceção do promotor da SL. Com isso, o conhecimento sobre
a transcrição de proteínas dependentes da RNAPII em tripanossomatídeos
se dá através da comparação com o que ocorre na transcrição do gene da
SL (Clayton, 2002). Mais recentemente, contudo, estudos realizados no
cromossomo 1 de Leishmania major mostram uma transcrição
policistrônica bidirecional de 79 genes, onde ocorre a transcrição de 50
ORFs (Open Reading Frame) em uma fita do DNA e de 29 ORFs restantes
na fita oposta. Ao que parece, a transcrição pela RNAP II se inicia em um
elemento promotor, próximo ao centro, e procede ininterruptamente em
direção às extremidades, transcrevendo as múltiplas ORFs (Myler et al.,
1999; Martinez-Calvillo et al., 2003).
Na transcrição policistrônica, então, os pré-mRNAs são sintetizados,
a partir de um único promotor, ao longo de centenas de kilobases,
gerando transcritos independentes através de reações acopladas de
processamento em “trans” e poliadenilação (Liang et al., 2003). Todavia,
diferentemente dos operons bacterianos, estas diferentes seqüências
codificadoras não estão relacionados em termos de funcionalidade ou
expressão temporal das proteínas codificadas (Campbell et al., 2003).

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
33
3.3.2 Processamento de mRNAs – “Trans-splicing” e
Poliadenilação
O processamento do mRNA é um mecanismo chave para a
transformação de precursores em mRNAs maduros prontos para serem
utilizados na síntese protéica. Este processo pode exercer um papel crítico
na regulação da expressão gênica em uma variedade de eucariotos. Duas
formas de processamento têm sido reportadas em eucariotos. A maioria
dos eucariotos utiliza o processamento em “cis”, processo que remove os
íntrons de mRNAs precursores, gerando mRNAs maduros (FIGURA 7A). O
processamento em “trans” é um processo menos conhecido e aparece
mais comumente no grupo de protozoários pertencentes a ordem
Kinetoplastida, o que inclui os tripanossomatídeos (Hastings, 2005)
(FIGURA 7B).
O “trans-splicing” foi primeiramente descoberto em T. brucei e já foi
confirmado em outros tripanossomatídeos, incluindo membros dos
gêneros Leishmania, Leptomonas, Trypanosoma, Crithidia e Phytomonas.
Este também é muito freqüente em nematóides, alguns trematóides e
Euglena spp. (Nilsen, 1993). O processamento em “trans” ocorre através
de uma transesterificação formando uma estrutura em “Y” em lugar do
laço intermediário que ocorre no processamento em “cis” (Liang et al.,
2003). Essa reação envolve a adição da sequência SL, junto com o cap4,
na extremidade 5’ dos mRNAs (Zick et al., 2005). Acoplado à
poliadenilação dos RNAs na sua região 3´, esse processamento leva à
formação dos mRNAs maduros (Liang et al., 2003).
Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
34
DNA
Transcrição e adição do Cap
Poliadenilaçãoéxon 1 intron 1 éxon 2 intron 2 intron 3 éxon 4
éxon 1 intron 1 éxon 2 intron 2 intron 3 éxon 4éxon 3
éxon 1 intron 1 éxon 2 intron 2 intron 3 éxon 4éxon 3
AAAAAAAAAAAA
“Cis-splicing”
éxon 1 éxon 2 éxon 3 AAAAAAAAAAAAéxon 4
éxon 3
Transporte núcleo/citoplasma e Tradução
AAAAAAAAAAAAA
Transcrição e “Cis-splicing” em Eucariotos
promotorCap
Regiões intergênicasCauda poli-A
Ribossomo
DNA
Transcrição e adição do Cap
Poliadenilaçãoéxon 1 intron 1 éxon 2 intron 2 intron 3 éxon 4
éxon 1 intron 1 éxon 2 intron 2 intron 3 éxon 4éxon 3
éxon 1 intron 1 éxon 2 intron 2 intron 3 éxon 4éxon 3
AAAAAAAAAAAA
“Cis-splicing”
éxon 1 éxon 2 éxon 3 AAAAAAAAAAAAéxon 4
éxon 3
Transporte núcleo/citoplasma e Tradução
AAAAAAAAAAAAA
Transcrição e “Cis-splicing” em Eucariotos
promotorCap
Regiões intergênicasCauda poli-A
Ribossomo
(A)
RNA policistrônico
Transcrição policistrônica
DNA Gene 1 Gene 2 Gene 3 Gene 4 Gene 5
Trans-splicing, adição do cap e poliadenilaçãoRNAs monocistrônicos
Transporte núcleo/citoplasma e Tradução
TRADUÇÃO
AAAAAAAA
promotorCapSplicer LeaderRegiões intergênicasCauda poli-ARibossomo
mRNA 1 mRNA 2 mRNA 3 mRNA 4 mRNA 5
Transcrição e “trans-splicing” em tripanosomatídeos
RNA policistrônico
Transcrição policistrônica
DNA Gene 1 Gene 2 Gene 3 Gene 4 Gene 5
Trans-splicing, adição do cap e poliadenilaçãoRNAs monocistrônicos
Transporte núcleo/citoplasma e Tradução
TRADUÇÃO
AAAAAAAA
promotorCapSplicer LeaderRegiões intergênicasCauda poli-ARibossomo
mRNA 1 mRNA 2 mRNA 3 mRNA 4 mRNA 5
Transcrição e “trans-splicing” em tripanosomatídeos(B)
Figura 07. Esquema ilustrando a transcrição e o processamento de mRNAs. Em (A), a transcrição e o processamento do mRNA em “cis” (“cis-splicing”) que ocorre nos eucariotos em geral e em (B) o processamento em “trans” (“trans-splicing”) do mRNA que ocorre em tripanosomatídeos.

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
35
O “trans-splicing” e a poliadenilação são processos acoplados que
aparentemente são regulados por sinais hierárquicos que ocorrem em
seqüências de polipirimidina intergênicas (Hug et al., 1994). Nenhuma
região específica clássica, tais como os sinais de poliadenilação (AAUAAA)
em eucariotos superiores, parece estar presente nos RNAs mensageiros
dos tripanossomatídeos. Resíduos de adenosina, quando repetidos
principalmente, parecem ser alvos preferenciais da poliadenilação (Liang
et al., 2003).
3.3.3 Cap4
Todos os mRNAs de células eucarióticas (exceto os das organelas)
possuem uma estrutura conhecida como cap na região 5’ dos mRNAs,
composta por um resíduo de 7-metil-guanosina (m7GTP) ligado ao
nucleosídeo inicial do transcrito por uma ponte trifosfato (Gingras et al.,
1999). A estrutura do cap no mRNA de eucariotos é formado pela ação
seqüencial de três enzimas. Um dos três fosfatos presente na região 5’
dos pré-mRNAs é clivado pela ação da RNA trifosfatase, na presença de
cátions divalentes (Ho e Shuman 2001). Esta reação permite a
transferência do GMP para GTP através da enzima guanililtransferase, na
porção 5’ difosfato para formar o GpppN (Silva et al., 1998). Ocorre então
a metilação na posição N-7 da guanosina pela ação da (guanina N-7)
metiltransferase (Shuman, 2001).
Nos tripanosomatídeos o cap é modificado por sete metilações,
composta de 2’-O-metilações na ribose dos primeiros quatro nucleotídeos
(AACU), com metilações adicionais sobre as bases do primeiro (m6,6A) e
do quarto (m3U) nucleotídeos, formando a estrutura altamente complexa
m7 guanosina (5’) ppp-N6,N6,2’-O-tri-metil-adenosina-p-2’-O-metil-

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
36
adenosina-p’-2’-O-metil-citosina-p-3,2’-O-di-metil-uridina, que foi
denominada de Cap4 (Bangs et al., 1992; Mair Ullu and Tschudi, 2000).
Ao contrário dos demais eucariotos, a sua adição aos mRNAs ocorre em
uma etapa pós-transcricional, quando da inserção da sequência SL pelo
“trans-splicing” (Mair, Ullu e Tschudi, 2000). Foi sugerido que a enzima
cap metiltransferase poderia ser alvo de promissoras drogas contra
tripanossomatídeos, uma vez que o mecanismo de formação e estrutura
do cap são fundamentalmente diferentes das do hospedeiro (Tschudi e
Ullu 2002), mas apenas as enzimas RNA trifosfatase e guanililtransferase
foram bem caracterizadas bioquimicamente (Silva et al., 1998; Ho e
Shuman, 2001). No entanto, um trabalho recente foi publicado
caracterizando também a (guanina N-7) metiltransferase de T. brucei
(Hall e Ho, 2006). Apesar das evidências mostrarem que há uma
necessidade da presença do cap durante o processamento em “trans”, sua
função ainda não foi totalmente esclarecida. Porém, a presença da
estrutura cap4, juntamente com os elementos da seqüência SL pode ter
uma função importante na tradução dos tripanosomatídeos (Zamudio et
al., 2006).
3.4 Controle da Expressão Gênica nos Tripanossomatídeos
O controle da expressão gênica pode ser realizado em vários níveis
como, por exemplo, na transcrição, no processamento pós-transcricional
do mRNA, na sua degradação, na tradução, via modificações pós-
traducionais e na degradação de proteínas (Meijer e Thomas, 2002). Nos
eucariotos, a regulação da expressão gênica é principalmente regulada ao
nível transcricional, pela modulação da ação da RNA polimerase II. Mais
de trinta genes de regulação estágio-específica foram estudados em
tripanossomatídeos, e nenhuma evidência de regulação de sua

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
37
transcrição, pela polimerase II, ou de regulação de seu processamento, ao
nível de poliadenilação ou trans-splicing, foi observada. Pelo contrário, a
regulação da expressão de genes estágio-específicos parece ocorrer
principalmente no nível pós-transcricional, mediada por seqüências na
região 3’ UTR dos mRNAs, que determinam a sua abundância, modulando
sua degradação. As mesmas seqüências regulatórias, com freqüência,
também modulam a eficiência da tradução dos mRNAs (Clayton, 2002).
A regulação da tradução desempenha um papel crítico no
crescimento, na proliferação e no desenvolvimento celular. Vantagens no
controle da tradução incluem resposta rápida e um modo de atingir o nível
de um produto gênico na ausência de transcrição. Todos os elementos do
mRNA maduro, no sentido 5’-3’, a estrutura cap, a região 5’ UTR, a ORF, a
região 3’ UTR e a cauda poli-A, contribuem para uma tradução eficiente do
mRNA. Quando a sequência 5’ UTR é analisada, certas características
podem sugerir um controle deste mRNA. Por exemplo, as regiões 5’ UTR
que possuem níveis de tradução eficientes são curtas, possuem baixo teor
das bases GC, são relativamente não estruturadas e não contem códons
AUG (Kochetov et al., 1998). A região 3’ UTR também parece participar da
regulação da expressão gênica. A tradução da proteína amastina de
Leishmania se mostrou mediada por seqüências dentro da região 3’ UTR
do seus diferentes mRNAs que variam em tamanho e composição (Wu et
al., 2000; Boucher et al., 2002). Uma característica da região regulatória
destes mRNAs é a presença de uma seqüência de aproximadamente 450
nucleotídeos que é altamente conservada, não apenas na maioria dos
transcritos da amastina (Rochette et al., 2005), mas também está
presente em vários outros mRNAs de Leishmania, incluindo conhecidos
mRNAs específicos de amastigotas (Boucher et al., 2002). Acredita-se que
esta seqüência atue no nível de regulação da tradução dos respectivos
mRNAs.

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
38
3.5 Biossíntese Protéica
A síntese protéica, também conhecida como tradução, é um ponto
importante do controle da expressão gênica. Este processo é responsável
pelas estruturas básicas dos seres vivos, sendo um dos mais importantes
para a sobrevivência tanto de organismos procariontes como de
eucariontes. A tradução do mRNA em proteínas se inicia após a montagem
do ribossomo 80S através da interação entre tRNA iniciador (tRNAi),
mRNA e as subunidades 40S e 60S. As taxas de tradução podem ser
controladas em cada um dos seus três estágios: iniciação, elongação e
terminação. Enquanto que as fases de elongação e terminação são
assistidas por um grupo limitado de fatores, a iniciação da tradução é um
evento complexo, sendo auxiliado por mais de 25 polipeptídeos (Gerbauer
e Hentze, 2004). Dentre estes três estágios, a iniciação parece ser o
principal alvo da regulação da tradução, uma vez que é nessa fase que o
ribossomo é recrutado ao RNA mensageiro e posicionado corretamente no
códon AUG de iniciação da tradução (Prevôt, Darlix e Ohlmann, 2003).
3.5.1 Iniciação da Tradução
O complexo processo de iniciação da tradução que leva a montagem
do ribossomo 80S consiste em vários estágios relacionados que são
mediados por proteínas conhecidas como fatores de iniciação da tradução
eucarióticos (eIFs - eukaryotic initiation factors) (Pestova et al., 2001).
Estes estágios incluem:
(1) Seleção do RNA transportador inicial (tRNAi), a partir do
conjunto de tRNAs celulares, pelo fator de iniciação eIF2 e montagem do
complexo ternário (eIF2 / GTP / tRNAi). Ligação deste complexo a outros
eIFs e a subunidade 40S para formar o complexo de pré-iniciação 43S;

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
39
(2) Ligação do complexo 43S ao mRNA, o que na maioria das vezes
ocorre por um mecanismo que envolve o reconhecimento inicial do cap na
extremidade 5' do mRNA pela subunidade eIF4E (cap-binding-protein) do
complexo eIF4F;
(3) Movimento do complexo ribossomal ligado ao mRNA ao longo da
região 5' não traduzida (5' UTR), a partir do sítio inicial de ligação até o
códon AUG de iniciação da tradução. O complexo inicial 48S é formado e o
códon AUG tem suas bases pareadas com as bases do anticódon do tRNAi;
(4) Dissociação dos fatores de iniciação da tradução do complexo
48S e junção da subunidade 60S para formar o ribossomo 80S, deixando
o tRNAi no sítio P ribossomal (Hershey e Merrick, 2000; He et al., 2003;
Prevôt, Darlix e Ohlmann, 2003; Gerbauer e Hentze, 2004).
3.5.1.1 Complexo eIF4F e PABP
Dentre os inúmeros fatores de iniciação da tradução que participam
do processo de biossíntese protéica, o complexo eIF4F se destaca por ter
papel crucial no recrutamento do ribossomo para interação com o mRNA e
no posicionamento correto no códon AUG. O papel proposto ao complexo
eIF4F é se ligar a extremidade 5’ do mRNA, desfazer quaisquer estruturas
secundárias aí encontradas, e então facilitar a associação do complexo
43S na agora desestruturada extremidade 5’. O complexo eIF4F também
auxilia a subunidade menor ribosomal na sua migração ao longo do mRNA
até encontrar o códon AUG (Kapp e Lorsch, 2004).
O eIF4F é composto de três subunidades conhecidas como eIF4A,
eIF4E e eIF4G. O fator eIF4E, também conhecido por proteína de ligação
ao cap, é responsável pelo reconhecimento do cap presente nos mRNAs
eucarióticos, recrutamento dos outros fatores de iniciação e por último da
subunidade 40S para o mRNA (Prevôt, Darlix e Ohlmann, 2003). O eIF4A

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
40
é uma RNA helicase, de 46 kDa que age removendo regiões de estrutura
secundária dentro da região 5' não traduzida (5' UTR) do mRNA,
facilitando a ligação da subunidade menor ribossomal e permitindo sua
migração até o códon de iniciação da tradução (Sonenberg e Dever,
2003). O eIF4G é uma proteína de alto peso molecular que interage com
diferentes proteínas celulares e virais, incluindo os fatores de iniciação da
tradução (Gingras et al., 1999). Este fator possui sítios de ligação para o
eIF4E, eIF4A, eIF3 e PABP (Proteína de ligação á cauda poli-A). Sua
interação com o eIF3, associado à subunidade menor ribossomal, e com o
complexo eIF4E/cap-mRNA, promove o reconhecimento do mRNA por esta
subunidade e a formação do complexo 48S (Gross et al., 2003).
O eIF4G possui também na sua região N-terminal um sítio de
ligação com a PABP. A ligação simultânea do eIF4G, com a proteína de
ligação ao cap, eIF4E, e a PABP associada com a cauda poli-A pode trazer
a porção 3’ do RNA para perto da região 5’ e fazer com que o mRNA
circularize (FIGURA 8). Esta circularização seria responsável tanto pela
“reciclagem” dos ribossomos, que após o término da síntese da cadeia
polipeptídica voltariam à região 5´, quanto por estimular a interação entre
o eIF4E e o cap (Wells et al., 1998; Lloyd, 2006).
A PABP de eucariotos possui 70KDa e é a maior proteína
citoplasmática que se liga ao mRNA. É uma proteína conservada
filogeneticamente, interage com a cauda poli-A dos mRNAs eucarióticos e
com várias outras proteínas, incluindo o eIF4G. Por isso, participa da
iniciação da tradução e tem sido reconhecida como um verdadeiro fator de
iniciação da tradução (Sachs e Varani, 2000; Kuhn e Wahle, 2004). A
PABP contém quatro motivos de reconhecimento do RNA (RRM) no seu
domínio N-terminal que estão envolvidos tanto na ligação com o mRNA
quanto com o eIF4G. Além de participar da iniciação da tradução, outros
estudos realizados mostraram que a PABP interage com o fator eRF3,
envolvido na terminação da tradução (Bruzik et al., 1988). Esta proteína

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
41
também possui um papel na poliadenilação de transcritos nucleares e na
estabilidade e decaimento dos mRNAs (Couttet et al., 1997; Gallie, 1998;
Mangus, Amrani e Jacobson, 1998; Hoshino et al., 1999).
3.5.1.2 eIF4E : Estrutura e Função
A biossíntese protéica em eucariotos é altamente dependente da
presença da estrutura m7GTP-cap, e o reconhecimento desta estrutura na
iniciação da tradução é feito pelo eIF4E. Este fator é uma fosfoproteína de
24 kDa que possui uma estrutura tridimensional filogeneticamente
conservada desde leveduras até vertebrados (Hershey e Merrick, 2000).
Ela possui na sua organização uma superfície côncava, rica em resíduos
hidrofóbicos, constituída de oito fitas β antiparalelas (Sonenberg e Dever,
AAAAAAAAAAAAAAAA
Cap4
eIF4GeIF4G
PABPPABP
40S40S
Met.eIF3eIF3
PABPPABP PABPPABP
eIF4AeIF4A
eIF4EeIF4E AUG
AAU
5’ UTR
3’ UTRmRNA
AAAAAAAAAAAAAAAA
Cap4
eIF4GeIF4G
PABPPABP
40S40S
Met.eIF3eIF3
PABPPABP PABPPABP
eIF4AeIF4A
eIF4EeIF4EeIF4EeIF4E AUG
AAU
5’ UTR
3’ UTRmRNA
Figura 8. Esquema da iniciação da tradução cap-dependente. Esquema ilustrando a associação do complexo de iniciação da tradução EIF4F (formado pelas subunidades eIF4A, eIF4E e eIF4G) com a PAPB, ligada a cauda poli-A na extremidade 3’ do mRNA. Essa associação causa a circularização do mRNA, e facilita o recrutamento do complexo 43S para o local, iniciando a tradução.

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
42
2003). O reconhecimento do cap é mediado pelo empacotamento entre os
triptofanos W56 e W102, formando 3 pontes de hidrogênio com a cadeia
lateral do ácido glutâmico E103, e interação por forças de van der Waals
com o grupo metil N(7) do triptofano W166 (aminoácidos referentes ao
eIF4E humano). Estes quatro resíduos fazem contato com a 7-metil
guanosina e são conservados entre a maioria dos eIF4Es conhecidos
(Marcotrigiano et al., 1997).
A atividade do eIF4E é regulada em vários níveis: via modulação de
sua transcrição, através de sua interação com a família de proteínas
ligadoras ao 4E (4EPB’s) e por fosforilação (Raught e Gingras, 1999). O
controle transcricional da síntese do eIF4E ainda não está bem
esclarecido, mas sabe-se que o seu promotor contém dois sítios de ligação
myc, sendo ambos necessários para a expressão do gene em sistemas
heterólogos (Jones et al., 1996). A atividade do eIF4E no complexo eIF4F
é limitada por proteínas da família 4EPB (4EPB1, 4EPB2 e 4EPB3) que
funcionam como sequestradores do eIF4E e assim inibem a tradução cap-
dependente (Sonenberg, 1996). O sítio de ligação das 4E-BPs ao eIF4E é
praticamente o mesmo encontrado no eIF4G, portanto, a função destas
proteínas seria mimetizar o fator eIF4G, competindo pela ligação com o
eIF4E (Sachs e Varani, 2000). A ligação destas proteínas aumenta a
afinidade do eIF4E pelo cap, mas impede a formação do complexo eIF4F
por impedir a associação do eIF4E com o eIF4G e, com isso, impede a
iniciação da tradução (Haghighat et al., 1995). O eIF4E também participa
de forma crucial no controle do ciclo celular (Strudwick e Borden, 2002).
Sua superexpressão causa perda do controle do ciclo celular e está
associada a uma série de tumores humanos (De Benedetti e Rhoads,
1990).
O fator eIF4E tem sido estudado numa variedade de organismos, em
muitos casos revelando a existência de múltiplos homólogos. Células de
mamíferos contêm três diferentes isoformas do eIF4E. Todas estas

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
43
proteínas ligam-se ao m7GTP, mas elas variam na habilidade de interagir
com eIF4G e com membros da família de repressores eIF4E-BP. A exata
função de todas as diferentes isoformas do eIF4E não foi totalmente
esclarecida (von der Haar et al., 2004), porém o eIF4E possui
considerável conservação filogenética, especialmente no sítio de ligação
ao cap (Tomoo et al., 2002) e demonstrou ser capaz de se ligar a
análogos da estrutura cap, principalmente no cap monometilado (m7GTP)
e, em alguns casos, na sua variante tri-metilada em nematóides (Keiper
et al., 2000).
3.5.1.3 Modificações Pós-Traducionais - Fosforilação do eIF4E
As modificações pós-traducionais, como fosforilação, glicosilação,
acetilação, metilação, entre outras, são encontradas com bastante
frequência nas proteínas em geral. Eventos de fosforilação, observados
em fatores de iniciação da tradução, atuam através de mecanismos ainda
pouco conhecidos que ajudam na regulação da biossíntese protéica.
Dentre estes fatores, já vem sendo estudada a fosforilação do fator eIF4E,
principalmente em humanos (Hiremath et al., 1989; Joshi et al., 1995;
Wang et al., 1998; Scheper et al., 2002; Zuberek et al., 2003). Durante o
ciclo celular, a fosforilação do eIF4E é baixa no período G0, aumenta
através das fases G1 e S e é radicalmente reduzida na fase M, em direta
correlação com a atividade de tradução (Bonneau e Sonenberg, 1987). O
eIF4E também começa a desfosforilar depois de choques de temperatura,
juntamente com a diminuição dos níveis de tradução, porém, nem sempre
a fosforilação do eIF4E e o aumento dos níveis de tradução estão
correlacionados. Estudos mostraram que quando a célula recebe algum
tipo de estresse (como, por exemplo, com arsênio ou anisomicina), induz
um aumento na fosforilação do eIF4E, enquanto que os níveis de tradução

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
44
diminuem drasticamente (Morley e McKendrick, 1997; Wang et al., 1998;
Dolniak et al., 2008). Neste caso, a diminuição da síntese protéica parece
causar um efeito negativo em outros componentes da maquinaria de
tradução. É possível que a fosforilação do eIF4E seja de fato um
mecanismo compensatório na indução do estresse tentando estimular a
biossíntese protéica (Gingras et al., 1999; Dolniak et al., 2008).
O eIF4E de mamíferos é fosforilado em um resíduo único, a serina
209 (S209) (Joshi et al., 1995), pelas quinases Mnk1 e Mnk2, que estão
associadas ao eIF4G, e essa fosforilação ocorre em resposta a vários
estímulos extracelulares (Gingras et al., 1999). Ensaios de fosforilação
com eIF4Es mutantes incapazes de se ligar ao eIF4G mostraram que eles
eram ineficientes para a fosforilação pela Mnk1 “in vivo”, sugerindo que
essa fosforilação só deve ocorrer quando o eIF4E se encontra ligado ao
eIF4G (Dever, 1999). Dados obtidos em 1994 sugerem que essa
fosforilação no eIF4E causa um aumento na afinidade pelo cap (Minich et
al., 1994), porém, dois estudos mostraram contradições a respeito destes
resultados, onde foi observado que a fosforilação causa a diminuição da
afinidade tanto do cap, como de alguns análogos ao cap (Scheper et al.,
2002; Zuberek et al., 2003). Baseado em estudos de cristalografia do
eIF4E de camundongo, acredita-se que a S209 fosforilada forma uma
ponte salina com a K159 e estabiliza a ligação do eIF4E com o cap
(Marcotrigiano et al., 1997). Esse resultado estaria de acordo com os
dados anteriores de que o eIF4E fosforilado possuiria mais afinidade ao
cap.
Dois mecanismos de ativação de quinases em cascata têm sido
caracterizados na regulação por fosforilação da atividade do eIF4E. O
primeiro provém da quinase PI3 e envolve as proteínas Akt e TOR. Esse
mecanismo influencia a fosforilação do ativador traducional rpS6 via
quinase ribossomal S6K1 que se liga e bloqueia a proteína repressora do
eIF4E (4E-BP), liberando a atividade do eIF4E (Kaufman, 2000; Zhang et

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
45
al., 2002; Wang e Proud, 2007). A proteína TOR integra sinais de fatores
de crescimento, nutrientes, stress e os níveis de energia para o controle
do crescimento celular (Inoki et al., 2005). O segundo mecanismo ocorre
através das quinases Mnk e envolve uma família de MAP quinases (MAPKs
- Mitogen-activated protein kinases), especificamente ERK e p38 que
ativam a quinase Mnk e esta, por sua vez, fosforila o eIF4E (Gingras et
al., 1999). A proteína repressora do eIF4E (4E-BP) também é regulada
por fosforilação através das MAPKs. A fosforilação de serinas e treoninas
específicas nestas proteínas modulam a sua afinidade pelo eIF4E (Averous
e Proud, 2006). A eIF4E-BP hipofosforilada se liga eficientemente ao
eIF4E, mas a fosforilação de um número crítico de resíduos anula esta
ligação (Gingras et al., 1999). Maiores estudos sobre a regulação da
atividade destas proteínas devem ser feitos, porém, já se sabe que a
fosforilação da serina 65 da eIF4E-BP1 provoca uma repulsão eletrostática
entre esta proteína e o eIF4E, impedindo a sua ligação (Marcotrigiano et
al., 1999).
O efeito fisiológico do aumento da fosforilação do eIF4E permanence
um enigma. Alguns estudos têm sugerido que esse aumento é requerido
para estimulação da tradução global por favorecer a forte associação do
eIF4E com a estrutura cap dos mRNAs (Morley e Naegele, 2002).
3.5.2 Complexo eIF4F e PABP em Tripanosomatídeos
Até o momento pouco se sabe a respeito de mecanismos de
tradução em tripanosomatídeos e apenas alguns trabalhos caracterizaram
fatores envolvidos neste processo. Dados mais relevantes foram obtidos
apenas com a PABP, identificada de diferentes espécies de
tripanosomatídeos (Batista et al., 1994; Pitula, Ruyechan e Williams,
1998; Bates et al., 2000). Em L. major foi descrito um gene que codifica

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
46
para a PABP (PABP1) que demonstrou ter um significativo grau de
homologia com outras PABPs identificadas na literatura, porém baixa
similaridade com PABPs descritas em outros tripanosomatídeos (Bates et
al ,2000). Estudos com a incorporação de fósforo radioativo (32P), mostrou
que uma das isoformas desta proteína se apresenta na forma fosforilada
(Bates et al., 2000). Só a partir do término do sequenciamento dos
genomas de L. major, T. cruzi e T. brucei (Berriman et al., 2005; El Sayed
et al., 2005; Ivens et al., 2005), entretanto, e com as novas ferramentas
de bioinformática e de biologia molecular, é que avanços significativos
começaram a ser feitos no estudo de outros fatores de tradução em
tripanosomatídeos, como descrito a seguir.
O primeiro homólogo de fator de iniciação da tradução identificado
em tripanosomatídeos foi um homólogo de eIF4A de Leishmania (LeIF),
identificado e caracterizado como indutor de IL-12 e resposta Th1 em
BALB/c mediante sua porção N-terminal (Skeiky et al., 1998). Com o
avanço no sequenciamento de genomas, e a partir de uma busca
sistemática por subunidades do complexo eIF4F, dois homólogos distintos
de eIF4A de L. major foram identificados, que diferiam em abundância e
na sua capacidade de interagir com homólogos de eIF4G (Dhalia et al.,
2005). Cinco homólogos de eIF4G e quatro homólogos de eIF4E (ver a
seguir), também foram identificados, conservados em diferentes espécies
de tripanosomatídeos e sugerindo uma elevada complexidade na iniciação
da sua síntese protéica. Em continuidade, com o estudo dos respectivos
ortólogos de eIF4A de T. brucei, foram investigados aspectos como níveis
celulares, localização subcelular e requisitos para viabilidade celular. Os
resultados obtidos indicam que apenas o TbEIF4AI é ortólogo funcional do
eIF4A com função na tradução. Já o TbEIF4AIII parece ser o ortólogo do
eIF4AIII de mamíferos, cuja função é predominantemente nuclear, ao
nível de processamento de mRNAs (Dhalia et al., 2006).
No que diz respeito a homólogos de eIF4E, um primeiro candidato

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
47
foi inicialmente identificado e caracterizado de L. major (LeishIF4E-1).
Esta proteína possui considerável conservação filogenética, especialmente
no sítio de ligação ao cap, localização citoplasmática e demonstrou ser
capaz de se ligar a análogos da estrutura cap, principalmente o cap
monometilado e o cap4 sintético (Yoffe et al., 2004). A identificação de
quatro homólogos em um trabalho paralelo, mostrou uma maior
complexidade em relação a função dessa proteína do que se esperava
originalmente. Ensaios de afinidade ao cap clássico de 7-metil-guanosina,
com três destas proteínas sintetizadas na presença de metionina marcada
com 35S, mostraram que apenas a LmEIF4E1 foi capaz de se ligar ao cap.
A modelagem molecular do LmEIF4E1 foi então realizada, confirmando
que o mesmo possui características suficientes para a ligação ao cap
(Dhalia et al., 2005). As três proteínas tiveram sua expressão investigada
em extratos de L. major onde se observou que seus níveis variam
significantemente, com LmEIF4E3 sendo muito abundante e ambas
LmEIF4E1 e 2 presentes em níveis muito mais baixos (TABELA 1) (Dhalia
et al., 2005).
Tabela 1. Quantificação dos homólogos eIF4E de Leishmania major na fase promastigota. Homólogos Peso Molecular Nº de moléculas/célula LmEIF4E1 23,8 kDa 3,2 x 103 LmEIF4E2 31,4 kDa 1 x 103
LmEIF4E3 37,8 kDa 7,1 x 104
Fonte : Dhalia et al., 2005.
Os resultados descritos para o LmEIF4E1 estão de acordo com
aqueles descritos para o LeishIF4E-1, a mesma proteína identificada
independentemente por um segundo grupo (Yoffe et al., 2004). Mais
recentemente, ensaios de interação dos quatro homólogos de eIF4E de L.
major com a protéina eIF4E-BP1 de mamíferos também foram realizados,
e foi observado que as proteínas LeishIF4E1 e LeishIF4E4 se ligaram à

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
48
proteína GST-eIF4E-BP1 imobilizada em resina. Neste mesmo estudo, foi
observada, através de fracionamento subcelular, que a localização das
quatro isoformas do eIF4E é citoplasmática (Yoffe et al., 2006).
3.5.3 Eventos de Fosforilação em Tripanosomatídeos
Em eucariotos, a fosforilação de proteínas é, dentre as modificações
bioquímicas regulatórias, a mais importante, e já foi descrita como
atuando numa série de eventos, tais como o “turnover” de proteínas,
localização, interação e atividade de várias moléculas. Nos
tripanossomatídeos, o número de fosfoproteínas encontradas é
consideravelmente maior do que o observado para outros parasitas
intracelulares que transitam em ambientes distintos (Parsons et al.,
2005). Durante os diferentes estágios do ciclo celular de Leishmania e
Trypanossoma foram observados padrões diferenciados de fosforilação de
proteína, sugerindo que as quinases e fosfatases estejam envolvidas nos
processos de diferenciação (Bengs et al., 2005). Análise das proteínas
quinases nos recém-completos genomas de L. major, T.brucei e T. cruzi
mostram que eles codificam, respectivamente, 179, 156 e 171 proteínas
kinases que são cataliticamente ativas e ainda 17, 20 e 19 genes de
proteínas quinases atípicas (Naula, Parsons e Mottram, 2005). Especula-
se que a modulação de proteínas através dos eventos de
fosforilação/desfoforilação esteja associada ao desenvolvimento do
processo de infecção, no momento da interação parasito-hospedeiro.
Foi observado em T. cruzi que o fator de transformação do
crescimento (TGFα) produzido por macrófagos, induziu fosforilação na
tirosina de duas proteínas de superfície que são específicas de
amastigotas, aumentando assim a síntese de DNA e o crescimento celular
(Alexander et al., 2003). Esse estudo é importante visto que o macrófago

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
49
tem papel crucial na imunomodulação da infecção com T. cruzi e também
em Leishmania. Recentemente foi determinada a presença de atividade da
proteína tirosina-fosfatase em extratos protéicos da forma promastigota
de L. major (Guirre-Garcia et al., 2006). Essas descobertas são bastante
promissoras e significativas, contudo, muito pouco ainda se sabe sobre os
eventos de fosforilação e sobre as proteínas quinases em
tripanosomatídeos, necessitando de estudos mais aprofundados nesta
área.

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
50
4. REFERÊNCIAS BIBLIOGRÁFICAS
Alexander J, Satoskar AR, Russell DG. (1999) Leishmania species: models
of intracellular parasitism. J Cell Sci. 112 Pt 18:2993-3002.
Alexander AD, Villalta F and Lima MF (2003) Transforming growth factor a
binds to Tripanosoma cruzi amastigotes to induce signaling ans
cellular proliferation. Infect Immun. 71(7):4201-5.
Alvar J, Canavate C, Gutierrez-Solar B, et al. (1997) Leishmania and
human immunodeficiency virus coinfection: the first 10 years. Clin
Microbiol Rev. 10:298-319.
Averous J, Proud CG. (2006) When translation meets transformation: the
mTOR story. Oncogene. 25 (48): 6423-35.
Bangs JD, Crain PF, Hashizume T, McCloskey JA and Boothroyd JC (1992)
Mass spectrometry of mRNA cap 4 from trypanosomatids reveals
two novel nucleosides. J Biol Chem. 267: 9805-9815.
Barak E, Amin-Spector S, Gerliak E, Goyard S, Holland N, Zilberstein D.
(2005) Differentiation of Leishmania donovani in host-free system:
analysis of signal perception and response. Mol Biochem Parasitol.
141: 99– 108.
Bates EJ, Knuepfer E and Smith DF (2000) Poly(A)-binding protein I of
Leishmania: functional analysis and localisation in trypanosomatid
parasites. Nucleic Acids Res. 28: 1211-1220.
Bates PA. (1993) Axenic culture of Leishmania amastigotes. Parasitol

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
51
Today. (4): 143-6.
Batista JA, Teixeira SM, Donelson JE, Kirchhoff LV and de Sa CM (1994)
Characterization of a Trypanosoma cruzi poly(A)-binding protein and
its genes. Mol Biochem Parasitol. 67: 301-312.
Bengs F, Scholz A, Kuhn D, Wiese M. (2005) LmxMPK9, a mitogen-
activated protein kinase homologue affects flagellar length in
Leishmania mexicana. Mol Microbiol. 55:1606-15.
Berriman M, Ghedin E, Hertz-Fowler C, Blandin G, Renauld H,
Bartholomeu DC, Lennard NJ, Caler E, Hamlin NE, Haas B, Böhme U,
Hannick L, Aslett MA, Shallom J, Marcello L, et al. (2005) The
genome of the African trypanosome Trypanosoma brucei. 309
(5733): 416-22.
Besteiro S, Williams RAM, Coombs GH, Mottram JC. (2007) Protein
turnover and differentiation in Leishmania. Int J Parasitol.
37(10):1063-75.
Bonneau AM, Sonenberg N. (1987) Involvement of the 24-kDa cap-
binding protein in regulation of protein synthesis in mitosis. J Biol
Chem. 262 (23): 11134-9
Boucher N, Wu Y, Dumas C, Dube M, Sereno D, Breton M, Papadopoulou
B. (2002) A common mechanism of stage-regulated gene expression
in Leishmania mediated by a conserved 3'-untranslated region
element. J Biol Chem. 277 (22): 19511-20.
Bruzik JP, Van DK, Hirsh D and Steitz JA (1988) Trans splicing involves a

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
52
novel form of small nuclear ribonucleoprotein particles. Nature 335:
559-562.
Burchmore RJ, Barrett MP. (2001) Life in vacuoles – nutrient acquisition
by Leishmania amastigotes. Int J Parasitol. 31(12): 1311-20.
Campbell DA, Thomas S, Sturm NR. (2003) Transcription in kinetoplastid
protozoa: why be normal? Microbes Infect. 5:1231-40.
Clayton CE. (2002) Life without transcriptional control? From fly to man
and back again. EMBO J. 21:1881-8.
Cohen-Freue G, Holzer TR, Forney JD, McMaster WR. (2007) Global gene
expression in Leishmania. Int J Parasitol. 37:1077-86.
Couttet P, Fromont-Racine M, Steel D, Pictet R and Grange T (1997)
Messenger RNA deadenylylation precedes decapping in mammalian
cells. Proc.Natl.Acad.Sci.U.S.A 94: 5628-5633.
Cunningham ML, Titus RG, Turco SJ, Beverley SM. (2001) Regulation of
differentiation to the infective stage of the protozoan parasite
Leishmania major by tetrahydrobiopterin. Science. 13; 292 (5515):
285-7
Cysne-Finkelstein L, Temporal RM, Alves FA, Leon LL. (1998) Long-term
cultivation of axenic amastigotes is associated to metacyclogenesis
of promatigotes. Exp Parasitol. 89 (1): 58-62
De Benedetti A, Rhoads RE (1990) Overexpression of eukaryotic protein
synthesis initiation factor 4E in HeLa cells results in aberrant growth

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
53
and morphology. Proc. Natl. Acad. Sci. U.S.A 87(21): 8212-6
Debrabant A, Joshi MB, Pimenta PF, Dwyer DM. (2004) Generation of
Leishmania donovani axenic amastigotes: their growth and
biological characteristics. Int J Parasitol. 34 (2): 205-17.
Desjeux P. (2004) Leishmaniasis: current situation and new perspectives.
Comp Immunol Microbiol Infect Dis. 27:305-18.
Devaux S, Lecordier L, Uzureau P, Walgraffe D, Dierick JF, Poelvoorde P,
Pays E, Vanhamme L. (2006) Characterization of RNA polymerase II
subunits of Trypanosoma brucei. Mol Biochem Parasitol. 148 (1):
60-8.
Dever TE (1999) Translation initiation: adept at adapting. Trends
Biochem.Sci. 24(10); 398-403
Dhalia R, Marinsek N, Reis CR, Katz R, Muniz JR, Standart N, Carrington M
and de Melo Neto OP (2006) The two eIF4A helicases in
Trypanosoma brucei are functionally distinct. Nucleic Acids Res. 34:
2495-2507.
Dhalia R, Reis CR, Freire ER, et al. (2005) Translation initiation in
Leishmania major: characterisation of multiple eIF4F subunit
homologues. Mol Biochem Parasitol. 140: 23-41.
Dolniak B, Katsoulidis E, Carayol N, Altman JK, Redig AJ, Tallman MS,
Ueda T, Watanabe-Fukunaga R, Fukunaga R, Platanias LC. (2008)
Regulation of arsenic trioxide-induced cellular responses by Mnk1
and Mnk2. J Biol Chem. [In press].

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
54
El-Sayed NM, Myler PJ, Bartholomeu DC, Nilsson D, Aggarwal G, Tran AN,
Ghedin E, Worthey EA, Delcher AL, Blandin G, Westenberger SJ,
Caler E, Cerqueira GC, Branche C, Haas B, Anupama A, et al. (2005)
The genome sequence of Trypanosoma cruzi, etiologic agent of
Chagas disease. 309 (5733): 409-15.
FUNASA, 2002(a). Vigilância e monitoramento da leishmaniose
tegumentar americana em unidades territoriais - Brasil, 1994-2001.
Boletim Eletrônico Epidemiológico. Ano 02, N°05 (13/12/2002).
FUNASA 2002(b). Leishmaniose visceral no Brasil: situação atual,
principais aspectos epidemiológicos, clínicos e medidas de controle.
Boletim Eletrônico Epidemiológico. Ano 02, N°06 (13/12/2002).
Gallie DR (1998) A tale of two termini: A functional interaction between
the termini of an mRNA is a prerequisite for efficient translation
initiation. Gene 216: 1-11.
Gebauer F and Hentze MW (2004) Molecular mechanisms of translational
control. Nat.Rev.Mol.Cell Biol. 5: 827-835.
Gilinger G and Bellofatto V (2001) Trypanosome spliced leader RNA genes
contain the first identified RNA polymerase II gene promoter in
these organisms. Nucleic Acids Res. 29: 1556-1564.
Gingras AC, Raught B and Sonenberg N (1999) eIF4 initiation factors:
effectors of mRNA recruitment to ribosomes and regulators of
translation. Annu.Rev.Biochem. 68: 913-963.

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
55
Gossage SM, Rogers ME, Bates PA. (2003) Two separate growth phases
during the development of Leishmania in sand flies: implications for
understanding the life cycle. Int J Parasitol. 33:1027-34.
Gross JD, Moerke NJ, Von der Haar T, Lugovskoy AA, Sachs AB, McCarthy
JEG and Wagner G (2003) Ribosome Loading onto the mRNA Cap is
driven by conformacional coupling between eIF4G and eIF4E. Cell,
115: 739-750.
Gu M., Lima C.D. (2005) Processing the message: structural insights into
capping and decapping mRNA. Curr. Opin, Struct. Biol. 15(1):99-
106.
Guirre-Garcia MM, Escalona-Montano AR, Bakalara N, et al. (2006)
Leishmania major : detection of membrane-bound protein tyrosine
phosphatase. Parasitology;132:641-9.
Gunzl A, Bruderer T, Laufer G, Schimanski B, Tu LC, Chung HM, Lee PT,
Lee MG. (2003) RNA polymerase I transcribes procyclin genes and
variant surface glycoprotein gene expression sites in Trypanosoma
brucei. Eukaryot. Cell. 2(3):542-51.
Gupta N., Goyal N. and Rastogi AK. (2001) In vitro cultivation and
characterization of axenic amastigotes of Leishmania. Trends
Parasitol. 17(3):150-3.
Haghighat A, Mader S, Pause A and Sonenberg N (1995) Repression of
cap-dependent translation by 4E-binding protein 1: competition with
p220 for binding to eukaryotic initiation factor-4E. EMBO J. 14:
5701-5709.

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
56
Hall MP, Ho CK. (2006) Functional characterization of a 48 kDa
Trypanosoma brucei cap 2 RNA methyltransferase. Nucleic Acids
Res. 34(19):5594-602.
Hastings K.E. (2005) SL trans-splicing: easy come or easy go? Trends
Genet. 21(4):240-7.
He H, Haar T, Singh C, Ii M, Li B, Hinnebusch A, McCarthy J and Asano KT
(2003) The yeast initiation factor 4G (eIF4G) HEAT domain interacts
with eIF1and eIF5 and is involved in stringent AUG selection. Mol
Cell Biol 23:5431-5445.
Hershey JWB and Merrick WC (2000) The pathway and mechanism of
initiation of protein synthesis. Translational Control of Gene
Expression (Cold Spring Habor Laboratory Press); pp. 33-38.
Hiremath LS, Hiremath ST, Rychlik W, Joshi S, Domier LL, Rhoads RE.
(1989) In vitro synthesis, phosphorylation, and localization on 48 S
initiation complexes of human protein synthesis initiation factor 4E.
J Biol Chem. 264 (2): 1132-8.
Ho, C.K. and Shuman, S. (2001) Trypanosoma brucei RNA triphosphatase.
Antiprotozoal drug target and guide to eukaryotic phylogeny. J. Biol.
Chem. 276: 46182–46186.
Hoshino S., Imai M., Kobayashi T., Uchida N., Natada T. (1999) The
eucariotic polypeptide chain releasing factor (eRF3/GSPT) carryng
translation termination signal to the 3’-poly(A) tailof mRNA. Direct
association of eRF3/GSPT with polyadenilate-binding protein. J. Biol.

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
57
Chem. 24(24); 16677-80
Hug M., Hotz HR., Hartman C., Clayton C. (1994) Hierarchies f RNA-
processing signals in a trypanosome surface antigen mRNA
precursor. Mol. Cell. Biol. 14(11); 7428-7435.
Inoki K, Ouyang H, Li Y, Guan KL. (2005) Signaling by target of rapamycin
proteins in cell growth control. Microbiol Mol Biol Rev. 69 (1): 79-
100.
Ivens AC, Peacock CS, Worthey EA, et al. (2005) The genome of the
kinetoplastid parasite, Leishmania major. Science. 309: 436-442.
Jones RM, Branda J, Johnston KA, Polymenis M, Gadd M, Rustgi A,
Callanan L, Schmidt EV. (1996). An essential E box in the promoter
of the gene encoding the mRNA cap-binding protein (eukaryotic
initiation factor 4E) is a target for activation by c-myc. Mol Cell Biol.
16(9): 4754-64.
Joshi B, Cai AL, Keiper BD, Minich WB, Mendez R, Beach CM, Stepinski J,
Stolaeski R, Darzynkiewicz E and Rhoads RE (1995) Phosphorilation
of eukaryotic protein synthesis factor 4E at Ser-209. J. Biol Chem
270: 14597-14603.
Joshi M, Dwyer DM, Nakhasi HL. (1993) Cloning and characterization of
differentially expressed genes from in vitro-grown 'amastigotes' of
Leishmania donovani. 58 (2): 345-54.
Kapp LD, Lorsch JR (2004) The molecular mechanics of eukaryotic
translation. Annu.Rev.Biochem. 73: 657-704.

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
58
Kaufman RJ. (2000) Overview of vector design for mammalian gene
expression. Mol Biotechnol. 16(2):151-60.
Keiper BD, Lamphear BJ, Deshpande AM Jankowska-Anyszka M, Aamodt
EJ, Blumenthal T, Rhoads RE (2000) Functional characterization of
five eIF4E isoforms in Caernorhabditis elegans. J. Biol. Chem; 275;
10590-10596.
Kochetov AV, Ischenko IV, Vorobiev DG, Kel AE, Babenko VN, Kisselev LL,
Kolchanov NA. (1998) Eukaryotic mRNAs encoding abundant and
scarce proteins are statistically dissimilar in many structural
features. 4; 440 (3): 351-5.
Kuhn U. and Wahle E. (2004) Structure and function of poly(A) binding
proteins. 25; 1678 (2-3): 67-84.
Lainson, R, Shaw, JJ, (1987). Evolution, classification and geographical
distribution. In: Peters, W., Killick-Kendrick, R. (Eds.), The
Leishmaniases in Biology and Medicine, Vol. 1. Academic Press,
London, 1–120.
Liang XH, Haritan A, Uliel S and Michaeli S. (2003) trans and cis splicing in
trypanosomatids: mechanism, factors, and regulation. Eukaryot.Cell
2: 830-840.
Lloyd RE (2006) Translational control by viral proteinases. Virus Res. 119
(1): 76-88.
Mair G, Ullu E, Tschudi C. (2000) Cotranscriptional cap 4 formation on the

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
59
Trypanosoma brucei spliced leader RNA. J Biol Chem. 15; 275(37):
28994-9.
Mangus DA, Amrani N, Jacobson A. 1998 Pbp1p, a factor interacting with
Saccharomyces cerevisiae poly(A)-binding protein, regulates
polyadenylation. 18 (12): 7383-96.
Marcotrigiano J, Gingras AC, Sonenberg N, Burley SK. (1997) Cocrystal
structure of the messenger RNA 5' cap-binding protein (eIF4E)
bound to 7-methyl-GDP. Cell 13; 89(6): 951-61.
Marcotrigiano J, Gingras AC, Sonenberg N and Burley Sk (1999) Cap-
dependent translation initiation in eukaryotes is regulated by a
molecular mimic of eIF4G. Mol Cell 3(6): 707-16.
Martinez-Calvillo S, Yan S, Nguyen D, Fox M, Stuart K and Myler PJ (2003)
Transcription of Leishmania major Friedlin chromosome 1 initiates in
both directions within a single region. Mol Cell 11: 1291-1299.
McConkey SE, López A, Shaw D, Calder J (2002) Leishmanial polyarthritis
in a dog. Canine Vet J 43(8): 607-609.
Meijer HA and Thomas AA (2002) Control of eukaryotic protein synthesis
by upstream open reading frames in the 5'-untranslated region of an
mRNA. Biochem J. 367: 1-11.
Minich WB, Balastra ML, Goss DJ, Rhoads RE (1994). Chromatographic
resolution of in vivo phosphorylated and nonphosphorylated
eucariotic translarion initiation factor eIF-4E: increase cap affinity of
the phosphorylated form. Proc Natl Acad Sci USA; 91(16); 7668-

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
60
7672.
Morley SJ, Naegele S. (2002) Phosphorylation of eukaryotic initiation
factor (eIF) 4E is not required for de novo protein synthesis
following recovery from hypertonic stress in human kidney cells. 277
(36): 32855-9.
Morley SJ, McKendrick L. (1997) Involvement of stress-activated protein
kinase and p38/RK mitogen-activated protein kinase signaling
pathways in the enhanced phosphorylation of initiation factor 4E in
NIH 3T3 cells. 272 (28): 17887-93.
Myler PJ, Audleman L, Devos T, Hixson G, Kiser P, Lemley C, Magness C,
Rickel E, Sisk E et al. (1999) Leishmania major Friedlin chromosome
1 has an unusual distribution of protein-coding genes. Proc. Natl.
Acad. Sci. U.S.A 96: 2902-2906.
Myler PJ, Stuart KD. (2000) Recent developments from the Leishmania
genome project. Curr Opin Microbiol. (4): 412-6.
Naula C, Parsons M, and Mottram JC (2005) Protein kinases as drug
targets in trypanosomes and Leishmania. Biochim. Biophys. Acta
1754(1-2):151-9.
Neves D (2005) Parasitologia Humana. 10ª edição. Ed. Atheneu, São
Paulo, 521p.
Nilsen TW. (1993) Trans-splicing of nematode premessenger RNA. Annu
Rev Microbiol. 47: 413-40.

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
61
Palenchar JB e Bellofatto V (2006) Gene transcription in trypanosomes.
Mol Biochem Parasitol. 146(2): 135-41.
Parsons M, Worthey EA, Ward PN, Mottram JC (2005). Comparative
analysis of the kinomes of three pathogenic trypanosomatids:
Leishmania major, Trypanosoma brucei and Trypanosoma cruzi.
BMC Genomics; 15; 6: 127.
Pearson RD, Sousa AQ, Jeronimo SMB. (2000) Leishmania species:
Visceral (Kala-Azar), Cutaneous, and Mucosal Leishmaniasis. In:
Mandell, G.L., Bennett, J.E., Dolin, R.D. Mandell, Douglas and
Bennett’s Principles and Practice of Infectious Diseases, 5th ed.,
Philadelphia, Churchill-Livingstone.
Pestova TV, Kolupaeva VG, Lomakin IB, et al. (2001) Molecular
mechanisms of translation initiation in eukaryotes. Proc. Natl. Acad.
Sci. USA. 98: 7029-7036.
Pitula J, Ruyechan WT and Williams N (1998) Trypanosoma brucei:
identification and purification of a poly(A)-binding protein. Exp.Parasitol.
88: 157-160.
Prévôt D, Darlix JL, Ohlmann T (2003) Conducting the initiation of protein
synthesis: the role of eIF4G. Biol Cell 95(3-4):141-56.
Raught B and Gingras AC (1999) eIF4E activity is regulated at multiple
levels. Int J Biochem Cell Biol 31: 43-57.
Rochette A, McNicoll F, Girard J, Breton M, Leblanc E, Bergeron MG,
Papadopoulou B. (2005) Characterization and developmental gene

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
62
regulation of a large gene family encoding amastin surface proteins
in Leishmania spp. Mol Biochem Parasitol. 140 (2): 205-20.
Sachs AB and Varani G (2000) Eukaryotic translation initiation: there are
(at least) two sides to every story. Nat. Struct. Biol. 7: 356-361.
Scheper GC, van Kollenburg B, Hu J, Luo Y, Goss DJ, Proud CG (2002)
Phosphorylation of eucariotic initiation factor 4E markedly reduces
its affinity for capped mRNA. J. Biol. Chem. 277(5); 3303-3309.
Schmidt G and Roberts L (1996) Foundations of Parasitology. 5ª edição.
Times Mirror High Education Group, Iowa, USA, 659p.
Shaw JJ (1994) Taxonomy of the genus Leishmania: present and future
trends and their implications. Mem. Inst. Oswaldo Cruz 89: 471-
478.
Shuman S. (2001) Structure, mechanism, and evolution of the mRNA
capping apparatus. Prog Nucleic Acid Res Mol Biol. 66: 1-40.
Silva E, Ullu E, Kobayashi R, Tschudi C. (1998) Trypanosome capping
enzymes display a novel two-domain structure. Mol. Cell. Biol. 18:
4612–4619.
Singh S. (2006) New developments in diagnosis of leishmaniasis. Indian J
Med Res. 123:311-30.
Skeiky YA, Kennedy M, Kaufman D, et al. (1998) LeIF: a recombinant
Leishmania protein that induces an IL-12-mediated Th1 cytokine
profile. J. Immunol. 161: 6171-6179.

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
63
Slappendel RJ, Ferrer L. (1998) In: Greene CE: Infectious Diseases of the
Dog and Cat. WB Saunders Co, Philadelphia, 450-458.
Sonenberg N (1996). Translational control. In: Stabilization of eukaryotic
initiation factor 4E binding to the mRNA 5’-cap by domains of eIF4G.
Biol. Chem. 39: 30551-30555.
Sonenberg N and Dever TE (2003) Eukaryotic translation initiation factors
and regulators. Curr. Opin. Struct. Biol. 13: 56-63.
Strudwick S, Borden KL. (2002) The emerging roles of translation factor
eIF4E in the nucleus. Differentiation. 70(1): 10-22.
Teixeira MCA, Santos RJ, Sampaio RB, Pontes-de-Carvalho L, dos-Santos
WLC. (2002) A simple and reproducible method to obtain large
numbers of axenic amastigotes of different Leishmania species.
Parasitol Res. 88: 963-968.
Tomoo K, Shen X, Okabe K, Nozoe Y, Fukuhara S, Morino S, Ishida T,
Taniguchi T, Hasegawa H, Terashima A, Sasaki M, Katsuya Y,
Kitamura K, Miyoshi H, Ishikawa M, Miura K. (2002) Crystal
structures of 7-methylguanosine 5'-triphosphate (m(7)GTP)- and
P(1)-7-methylguanosine-P(3)-adenosine-5',5'-triphosphate
(m(7)GpppA)-bound human full-length eukaryotic initiation factor
4E: biological importance of the C-terminal flexible region. Biochem
J. 362 (Pt 3): 539-44.
Tschudi C, Ullu E. (2002) Unconventional rules of small nuclear RNA
transcription and cap modification in trypanosomatids. Gene Expr.

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
64
10: 3–16.
Vickerman K., Coombs GH. (1999) Protozoan paradigms for cell biology. J
Cell Sci. 112 (Pt 17) 2797-98.
von der Haar T, Gross JD, Wagner G, McCarthy JE (2004) The mRNA cap-
binding protein eIF4E in post-transcriptional gene expression. Nat
Struct Mol Biol. 11(6): 503-11.
Wang X, Flynn A, Waskiewicz AJ, Webb BL, Vries RG, Baines IA, Cooper
JA, Proud CG. (1998) The phosphorylation of eukaryotic initiation
factor eIF4E in response to phorbol esters, cell stresses, and
cytokines is mediated by distinct MAP kinase pathways. J Biol Chem.
273 (16): 9373-7.
Wang X, Proud CG. (2007) Methods for studying signal-dependent
regulation of translation factor activity. Methods Enzymol. 431: 113-
42
Wells SE, Hillner PE, Vale RD and Sachs AB (1998) Circularization of
mRNA by eukaryotic translation initiation factors. Mol.Cell 2: 135-
140.
Wu Y, El Fakhry Y, Sereno D, Tamar S, Papadopolou B. (2000) A new
developmentally regulated gene family in Leishmania amastigotes
ecoding a homolog of amastin surface proteins. Mol. Biol. Parasit.
110(2):345-57.
Yoffe Y, Zuberek J, Lerer A, et al. (2006) Binding specificities and potential
roles of isoforms of eukaryotic initiation factor 4E in Leishmania.

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
65
Eukaryot.Cell. 5: 1969-1979.
Yoffe Y, Zuberek J, Lewdorowicz M, Zeira Z, Keasar C, Orr-Dahan I,
Jankowska-Anyszka M, Stepinski J, Darzynkiewicz E, Shapira M.
(2004) Cap-binding activity of an homolog from Leishmania. RNA.
10(11):1764-75.
Zamudio JR, Mittra B, Zeiner GM, Feder M, Bujnicki JM, Sturm NR and
Campbell DA (2006) Complete cap 4 formation is not required for
viability in Trypanosoma brucei. Eucaryot Cell. 5: 905-915.
Zhang X, Shu L, Hosoi H, Murti KG, Houghton PJ. (2002) Predominant
nuclear localization of mammalian target of rapamycin in normal
and malignant cells in culture. 277 (31): 28127-34.
Zick A, Onn I, Bezalel R, Margalit H, Shlomai J. (2005) Assigning functions
to genes: identification of S-phase expressed genes in Leishmania
major based on post-transcriptional control elements. Nucleic Acids
Res. 33 (13): 4235-42.
Zilberstein D, Shapira M. (1994) The role of pH and temperature in the
development of Leishmania parasites. Annu Rev Microbiol. 48: 449-
70.
Zuberek J, Wyslouch-Cieszynska A, Niedzwiecka A, Dadlez M, Stepinski J,
Augustyniak W, Gingras AC, Zhang Z, Burley SK, Sonenberg N,
Stolarski R, Darzynkiewicz E. (2003) Phospholylation of eIF4E
attenuates its interation with mRNA 5’ cap analogs by electrostatic
repulsion: inten-mediated protein ligation strategy to obtain
phosphorylated protein. RNA. 9(1):52-61.

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
66
Referências Eletrônicas:
CDC - Center for Disease Control and Prevention
http://www.dpd.cdc.gov/DPDx (Dezembro, 2007).
UGA - University of Georgia, College of Veterinary Medicine.
http://www.vet.uga.edu (Dezembro, 2007).
WHO – World Health Organization.
http://www.who.int (Dezembro, 2007).

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
67
5. MANUSCRITO DE ARTIGO CIENTÍFICO
Análise da expressão e investigação de mecanismos envolvidos com o controle da atividade de homólogos do fator de iniciação da tradução EIF4E ao longo do ciclo de vida de Leishmania amazonensis.
Pereira, M.M.P. 1,2 ; da Costa Lima,T.D.C.1,2 and de Melo Neto,O.P 1,2,*
1 Programa de Pós-Graduação em Genética, Departamento de Genética, Universidade Federal de Pernambuco, Av. Professor Moraes Rego s/n, Cidade Universitária, 50670-
901, Recife, PE, Brasil. 2 Centro de Pesquisas Aggeu Magalhães, Fundação Osvaldo Cruz, Av. Professor Moraes
Rego s/n, Cidade Universitária, 50670-420, Recife, PE, Brasil.
*Autor para correspondência: Fax: 00558121012636 E-mail: [email protected]
Artigo a ser enviado para a revista Molecular and Biochemical
Parasitology. ISSN 0166-6851. Fator de impacto (JCR, 2005): 2,733.

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
68
RESUMO
A iniciação da tradução em eucariotos é um ponto crucial no processo de
biossíntese protéica, onde a estrutura cap dos mRNAs têm papel
fundamental. O cap é reconhecido pelo eIF4E, que participa do complexo
de iniciação da tradução eIF4F e permite a ligação do ribosoma ao mRNA.
Em Leishmania, onde a regulação da expressão gênica é mediada em boa
parte pelo controle da sua tradução, vários homólogos de eIF4E foram
identificados. Nosso trabalho visou comparar o padrão de expressão de
quatro homólogos (EIF4E1-4) desse fator e identificar que mecanismos
poderiam estar agindo no controle da sua expressão e atividade. Curvas
de crescimento e diferenciação foram realizadas no intuito de se obter as
três formas principais de L. amazonensis. Foi observado que todos os
homólogos estão presentes ao longo das curvas e possuem padrões de
expressão distintos. Os EIF4E2-4 apresentaram-se com duas isoformas,
sugerindo modificações pós-traducionais como fosforilação. Destes, os
EIF4E3 e 4 apresentaram variações na expressão destas isoformas
durante o ciclo de vida do parasita e foram investigados mais
detalhadamente. Experimentos de localização subcelular indicaram que as
duas isoformas de ambas as proteínas são encontradas no citoplasma
tendo a ausência de soro fetal bovino no meio induzindo modificações
pós-traducionais associadas à falta de crescimento celular. Por marcação
metabólica observou-se que a isoforma “desfosforilada” do EIF4E3 e a
isoforma “fosforilada” do EIF4E4 estão associadas a uma maior atividade
de tradução. A incubação com inibidores de transcrição e tradução sugeriu
que a desfosforilação do EIF4E3 é dependente da tradução, e que a
fosforilação do EIF4E4 é dependente da transcrição. Estes resultados
indicam uma possível regulação da biossíntese protéica através de
mecanismos pós-traducionais, como a fosforilação, porém maiores
estudos são necessários para o entendimento desse processo.
Palavras-chave: Leishmania, EIF4E, fosforilação.

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
69
1. INTRODUÇÃO
O gênero Leishmania é um dos nove gêneros pertencentes à família
Trypanosomatidae, formada por um grupo de parasitas que causam uma
variedade de doenças de importância médica e veterinária. A leishmaniose
representa um elevado problema de saúde pública, sendo endêmica em
88 países e afetando cerca de 1,5 a 2 milhões de pessoas atualmente,
com cerca de 400 mil novos casos por ano [1]. Esta doença apresenta
uma importante diversidade clínica e epidemiológica que vai desde lesões
cutâneas localizadas até infecções viscerais que podem ser fatais [2].
A Leishmania é transmitida aos mamíferos pela picada do inseto
hematófago e, por estar presente em dois hospedeiros, possui um
complexo ciclo de vida envolvendo várias formas de desenvolvimento,
apresentando uma rápida adaptação a mudanças de condições ambientais
[3]. Alterna entre duas formas morfológicas significativas, onde a forma
intracelular obrigatória amastigota infecta hospedeiros mamíferos e a
forma flagelada promastigota está presente no inseto vetor.
Nos tripanosomatídeos encontramos algumas particularidades que
os diferem dos demais eucariotos, como a transcrição policistrônica e o
trans-splicing. Além disso, esses protozoários se caracterizam pela
importância da regulação pós-transcricional no controle da sua expressão
gênica, onde a síntese de proteínas parece ser um componente
importante dessa regulação [4]. O processo de biossíntese protéica,
conhecido também como tradução, tem um papel fundamental para a
sobrevivência de todos os organismos. Esse mecanismo nos
tripanosomatídeos ainda é pouco conhecido e precisa de mais estudos
para seu total entendimento. A tradução representa o passo final para a
via de expressão gênica, que media a formação do proteoma a partir da
informação genômica e pode ser dividida em três principais etapas:
iniciação, elongação e terminação [5]. Participam desse processo

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
70
inúmeros fatores protéicos que auxiliam o ribossomo nestas três etapas.
Enquanto que na elongação e terminação o número destes fatores é
limitado, na iniciação da tradução participam mais de 25 polipeptídeos,
sendo a etapa mais crítica do processo, sujeita a diferentes mecanismos
de regulação. Esta etapa compreende o reconhecimento do mRNA maduro
pela subunidade 40S do ribossomo e a identificação do códon AUG [6].
A iniciação da tradução é mediada por proteínas conhecidas como
fatores de iniciação da tradução (eIFs). Entre esses fatores destaca-se o
complexo eIF4F, composto pelas subunidades: eIF4E, a proteína de
reconhecimento da estrutura cap; o eIF4A, que possui uma atividade de
helicase responsável pela remoção de estruturas secundárias da
extremidade 5’ do mRNA; e o eIF4G, a proteína estruturadora do
complexo que possui sítios de ligação para o eIF4E, eIF4A, eIF3 e PABP
(Proteína de ligação á cauda poli-A) [7]. Para que ocorra a síntese
protéica, o complexo eIF4F se liga a estrutura cap presente na região 5’
dos mRNAs e permite o recrutamento da subunidade menor ribossomal. A
PABP se liga ao fator eIF4G, promovendo uma interação entre as
extremidade 3’ e 5’ do mRNA, causando uma circularização da molécula
do mensageiro e facilitando a sua tradução [8, 9].
O eIF4E é uma fosfoproteína de 24 kDa, responsável pelo
reconhecimento do cap presente nos mRNAs eucarióticos, a primeira
etapa do recrutamento para o mRNA dos outros fatores de iniciação e por
fim da subunidade 40S [6]. É um fator essencial para a estabilidade do
complexo eIF4F na ligação ao cap e assim possui papel crucial na iniciação
da tradução [10]. É uma proteína conservada desde leveduras até o
homem e um número variado de isoformas deste fator tem sido
encontrado em diversos organismos, dentre estes: três em humanos
[11,12]; três em Arabidopsis thaliana [13] e cinco em Caernorhabdtis
elegans [14]. Em mamíferos a atividade do fator eIF4E é limitada
principalmente por proteínas da família eIF4E-BP, que funcionam como

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
71
seqüestradoras do eIF4E e assim inibem a tradução e controlam as taxas
globais de produção de proteínas [15]. Além disso, o eIF4E é ainda
controlado diretamente por fosforilação pela enzima MnK, aumentando a
sua taxa de tradução [16].
Até o momento, pouco se sabe a respeito do mecanismo de
tradução em tripanosomatídeos e até recentemente apenas alguns poucos
trabalhos tinham enfocado proteínas envolvidas neste processo, dentre
estes alguns fatores de elongação e a proteína PABP [17,18]. Só em 2004
foi identificado e caracterizado um primeiro homólogo do fator eIF4E de
Leishmania major (LeishIF4E-1), que possui conservação no sítio de
ligação ao cap, localização citoplasmática e demonstrou ser capaz de se
ligar tanto ao cap monometilado como ao cap4, característico dos
tripanosomatídeos [19]. Já a partir do término do sequenciamento do
genoma de L. major [20] e outros tripanosomatídeos, foi possível a
identificação e caracterização preliminar de quatro homólogos de fator
eIF4E (LmEIF4E1-4); cinco de eIF4G (LmEIF4G1-5) e dois de EIF4A
(LmEIF4A1-2) [21].
O número elevado de homólogos de eIF4E e eIF4G encontrados em
tripanosomatídeos, organismos unicelulares, levantou a possibilidade de
que nem todos possam estar atuando diretamente como fator de iniciação
da tradução. Alternativamente, como estes parasitas possuem um
complexo ciclo de vida, estas proteínas poderiam estar atuando nas
diferentes fases deste ciclo controlando diferencialmente a síntese de
proteínas. Neste estudo, analisamos a expressão de quatro homólogos do
fator eIF4E (LmEIF4E1-4) em L. amazonensis durante diferentes fases do
seu ciclo de vida, onde observamos a expressão diferenciada entre os
quatro fatores. Duas destas proteínas apresentaram expressão modificada
durante o ciclo de vida, tendo sido detalhadamente avaliadas para um
melhor entendimento das causas das modificações observadas em seu
comportamento.

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
72
2. METODOLOGIA
2.1. Obtenção e armazenamento dos parasitas.
Formas promastigotas de Leishmania amazonensis
(MHOM/77/LTB0016), foram mantidas em meio Schneider pH 7,2 e
temperatura de 26ºC, suplementado com 10% de soro fetal bovino (SFB),
100UI de penicilina, 100 µg/ml de estreptomicina e 1 mM de L-glutamina.
Esta forma foi utilizada para inoculação no coxim plantar de camundongos
Balb/c numa concentração de 1 x 107 células diluída em 25 µl de PBS (137
mM NaCl, 2,7 mM KCl , 4,3 mM Na2HPO4, 1,47 mM KH2PO4, pH 7.3).
Quatro a oito semanas após o inóculo, a lesão se tornava aparente e
então os camundongos eram sacrificados para obtenção das formas
amastigotas infectivas [22]. Destas, formas amastigotas foram obtidas
tanto para a produção de extratos protéicos quanto para a realização da
diferenciação em forma promastigota infectivas, estocadas através do
congelamento em nitrogênio líquido. As células utilizadas para o
procedimento de congelamento sofreram apenas uma passagem, a fim de
não perderem sua infectividade, sendo congeladas utilizando glicerol 60%
numa diluição de 1:1.
2.2. Curvas de crescimento e diferenciação de L. amazonensis.
Curvas de crescimento da forma promastigota de L. amazonensis
foram estabelecidas a partir do descongelamento de células promastigotas
infectivas em meio Schneider pH 7,2. Estas foram cultivadas até a fase
estacionária, sendo repassadas ao mesmo meio numa diluição de 106
células/ml para dar início à curva padrão de crescimento, propriamente.
Essa curva foi monitorada diariamente e o crescimento dos parasitas

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
73
quantificado para detecção da fase estacionária. Nesta fase, a partir do
quinto dia, se observava a presença de uma elevada proporção de células
metacíclicas. As células neste ponto eram repassadas, também numa
diluição de 106 células/ml, para meio Schneider pH 5,5, suplementado
com 20% de SFB e colocadas a 32ºC, a fim de induzir sua diferenciação
em nova curva de crescimento. As células foram monitoradas durante
toda a curva, quantificadas e alíquotas padronizadas foram retiradas nos
tempos de 0h, 2h, 6h e a cada 24h até a sua morte. Essas alíquotas foram
centrifugadas e ressuspendidas em tampão de amostra para SDS-PAGE
para análise posterior através de ensaios de Western-blot.
Curvas com quatro condições diferentes no meio Schneider pH 7,2
também foram realizadas, onde foi utilizado um meio condicionado de
uma cultura crescida até o 8º dia. Esse meio condicionado foi
centrifugado, filtrado para ser reutilizado no ensaio e colocado em duas
garrafas de cultura, onde em uma delas foi acrescentado SFB. Também foi
utilizado neste ensaio meio Schneider pH 7,2 novo, em duas condições
distintas, com e sem SFB. Células estacionárias foram repassadas, numa
diluição de 106 células/ml, para as quatro garrafas de cultura. Alíquotas
foram retiradas nos tempos de 0h, 15min, 30min, 1h, 2h, 6h e em 24h,
onde as células foram quantificadas para observação do seu crescimento e
processadas para Western-blot.
2.3. Expressão e purificação das proteínas recombinantes.
A expressão das proteínas recombinantes foi realizada através da
transformação em células BL21 de Escherichia coli, crescida em meio LB,
de construções plasmidiais contendo os vários genes em estudo clonados
em dois vetores de expressão distintos: pGEX4T3, para expressão de
proteínas fusionadas a Glutationa S-transferase (GST); pET21D, para
expressão de proteínas fusionadas a uma cauda de poli-histidinas. As

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
74
bactérias transformadas foram crescidas até a densidade ótica (OD600) de
0,5, e posteriormente induzidas, por 4 horas a 30ºC, com IPTG numa
concentração final de 0,1 mM. Após a indução, as bactérias foram
centrifugadas (10.000 g / 4ºC / 10 min), ressuspendidas em 20ml de PBS
(137 mM NaCl, 2,7 mM KCl , 4,3 mM Na2HPO4 ,1,47 mM KH2PO4, pH 7.3),
e lisadas por ultrasonicação. A purificação das proteínas recombinantes foi
realizada de acordo com protocolo previamente estabelecido [23], através
de cromatografia de afinidade tanto para proteínas fusionadas com cauda
de poli-histidina, utilizando a resina Ni-NTA agarose (Qiagen) quanto com
GST, com a resina glutationa-sefarose (GE Biosciences). A purificação das
proteínas foi analisada em gel SDS-PAGE 15%, submetido a uma solução
corante contendo Azul de Comassie R-250, e quantificadas através de
uma curva de diluição de BSA com quantidades pré-definidas.
2.4. Purificação do anticorpo por Imunoadsorção.
Soros produzidos contra as diferentes proteínas em estudo foram
purificados através da técnica de imunoadsorção, onde as respectivas
proteínas recombinantes foram fracionadas em gel SDS-PAGE 15% e
transferidas para membrana PVDF (Immobilon-P – Millipore®). Após a
transferência, a membrana foi corada com 0,2% Rouge Ponceau / 1% TCA
onde a banda correspondente à proteína de interesse foi excisada da
membrana e cortada em pequenos pedaços colocados em tubo de 1,6 ml.
Após o bloqueio de 30 minutos em solução de PBS com leite 1% e Tween-
20 a 0,05%, o soro foi adicionado numa diluição de 1:1 com PBS e
deixado sob agitação durante aproximadamente 18 horas, a 4ºC. A
eluição dos anticorpos foi conseguida através da utilização de 200µl da
solução de glicina ácida 0,1 M (pH 2,5) com vigorosa agitação do tubo
durante 5 minutos, seguida da neutralização do pH por adição de 20 µl de

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
75
TRIS 1M pH8,0 e 200 µl de PBS duas vezes concentrado. Os anticorpos
purificados foram então utilizados em ensaios de “Western-Blot” para o
teste de sensibilidade e especificidade, utilizando diluições seriadas das
respectivas proteínas recombinantes quantificadas.
2.5. Ensaios de “Western-Blot”.
Os extratos obtidos através das curvas realizadas com os parasitas
foram fracionados em gel SDS-PAGE 15% e transferidos para membranas
PVDF (Immobilon-P – Millipore®) para a realização dos ensaios de
“Western-blot”. As membranas foram bloqueadas em solução de TBS (20
mM Tris, 500 mM NaCl, pH 7,5), leite desnatado 5% e Tween-20 a 0,05%
em temperatura ambiente por uma hora. Em seguida, a membrana foi
incubada com soro policlonal, na diluição pré-definida, durante uma hora
em solução de TBS, leite 5% e Tween-20 a 0,05%. Após lavagem com
TBS, a membrana foi incubada com o segundo anticorpo (anti-IgG de
coelho conjugado com peroxidase) (Jackson ImmunoResearch
Laboratories), numa diluição de 1:10.000, na mesma solução que a
incubação anterior, seguida de nova lavagem com TBS. O sinal foi
visualizado através de quimioluminescência (técnica ECL – “Enhanced
Chemioluminecency”) em filme Kodak Biomax Light.
2.6. Imunocitoquímica.
A imunocitoquímica foi realizada utilizando aproximadamente 5 x
106 células em cada reação. As células foram centrifugadas (1600 g /
25ºC / 10 min), lavadas com PBS e fixadas com paraformaldeído a 3%
por 10 minutos a temperatura ambiente. As células foram novamente

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
76
centrifugadas e colocadas em placa já contendo lamínula fixada com poli-
L-lisina (Sigma). Após 15 minutos, foram feitas as lavagens com PBS-
glicina 20 mM e a permeabilização com 0,1% de Triton 100x (Sigma) por
10 minutos, seguida das lavagens e do bloqueio com PBS-Glicina + BSA
1% por 20 minutos. Posteriormente, foi colocado o primeiro anticorpo
purificado na diluição pré-definida por 1h a 37ºC. Após lavagens, foi
colocada RNAse A 10 mM por 20 minutos, para degradar todo RNA
presente na célula. Foram feitas novas lavagens e colocado o segundo
anticorpo Alexa Fluor 488 conjugado com o segundo anticorpo diluído
1:500. Para a marcação do DNA foi utilizado o TOTO 3 (Molecular Probes)
diluído 1:1000.As células foram deixadas no escuro por 1h a 37ºC. Ainda
no escuro, foram lavadas, secas, as lamínulas fixadas em lâminas com 10
µl de Prolong-gold e guardadas protegidas da luz em freezer. As células
foram visualizadas em microscópio confocal utilizando o programa de
microscopia Leica SPII-AOBS.
2.7. Marcação Metabólica
A marcação metabólica foi realizada utilizando 1 x 107 células para
cada reação. As células foram crescidas como padronizado nas curvas de
crescimento de promastigotas, e no tempo pré-determinado, foram
retiradas e colocadas em tubos de centrífuga de 15ml. Adicionava-se
então metionina marcada com [35S] para uma concentração final de 10
mCi/ml e deixado a 27ºC durante 1 hora sob agitação. Após esse período,
as células foram centrifugadas (14000RPM/25ºC/10 min) e
ressuspendidas em tampão de amostra para SDS-PAGE para análise
através de eletroforese em gel 15% e exposição em filme Kodak Biomax
MR por 5 dias.

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
77
2.8 Curvas de inibição de transcrição e tradução.
Para realização das curvas de inibição, as células foram repassadas
para meio Schneider pH 7,2, também na diluição de 106 células/ml, onde
foi adicionado cicloheximida, para inibição da tradução, ou actinomicina D,
para inibição da transcrição. Os compostos foram utilizados nas
concentrações finais de 50 µg/ml e 10 µg/ml respectivamente. Alíquotas
foram retiradas nos tempos de 0h, 15min, 30min, 1h, 2h, 6h e em 24h.
3. RESULTADOS
3.1 Curvas de crescimento e diferenciação.
Para analisar a expressão dos homólogos de eIF4E durante o ciclo
de vida da Leishmania amazonensis, foram realizadas diferentes curvas de
crescimento e diferenciação de forma a maximizar a obtenção das três
principais formas de Leishmania (promastigotas procíclica e metacíclica e
amastigota). Nestas curvas, foram utilizadas células recém isoladas de
lesão de camundongos, e/ou derivadas de alíquotas congeladas, de forma
que ao início dos experimentos não tenham sido submetidas a mais do
que 5 repiques contados do seu isolamento do animal. Este procedimento
foi realizado para manter estas células altamente infectivas e com alta
capacidade de diferenciação. De forma a sincronizar, da melhor maneira
possível, a fase de crescimento celular, padronizou-se iniciar as curvas
com alíquotas de culturas já em fase estacionária. Inicialmente, curvas
foram estabelecidas em meio com pH 7,2 e a 26ºC, visando a obtenção de
formas procíclicas (crescimento exponencial e estacionário) e metacíclicas
da Leishmania. Estas foram monitoradas diariamente, onde foram

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
78
observados diversos fatores como morfologia, motilidade e crescimento
celular. A partir do 5º dia de crescimento celular, as células se
encontravam em fase estacionária, onde eram observadas, em 40 a 50%
das células da cultura, as formas metacíclicas com flagelo alongado. Para
a diferenciação em amastigotas axênicos (em contrapartida às
amastigotas obtidas diretamente de animais), curvas semelhantes foram
estabelecidas no mesmo meio de cultura, porém com o pH acidificado (pH
5,5) e a incubação em temperatura mais elevada (32ºC). A monitoração
das células permitiu a observação, já a partir do primeiro dia da curva, da
presença de formas redondas semelhantes à amastigota.
Todas as curvas realizadas apresentaram o mesmo padrão de
crescimento tanto nas formas promastigotas (FIGURA 1A) como na forma
amastigota “like” (semelhante à amastigota) (FIGURA 1C). A forma
promastigota foi encontrada em fase exponencial até o quarto dia, e no
quinto dia as células se encontravam na fase estacionária. No sexto dia da
curva, as células apresentavam-se na fase de declínio, onde foi observada
pouca motilidade, múltiplas rosetas e várias células deformadas. A cultura
em pH 5,5 e a 32ºC apresentou crescimento até o terceiro dia, a partir do
qual as células começaram a entrar em fase estacionária. A morfologia
arredondada característica de amastigota já foi observada a partir do
primeiro dia da curva, em menor proporção, e passou a ser predominante
no final da curva. A partir destas culturas foram retiradas alíquotas em
intervalos selecionados para a realização dos demais experimentos
descritos neste trabalho.
3.2 Validação das curvas de crescimento e diferenciação.
Uma vez estabelecidas as condições de cultivo, partiu-se para
confirmar a expressão, nas respectivas curvas, de proteínas que

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
79
participam na tradução e cuja expressão é sabidamente constitutiva.
Assim, alíquotas contendo quantidades equivalentes de células de cada
curva foram inicialmente avaliadas por SDS-PAGE e em seguida em
ensaios de Western-blot com soro contra proteínas selecionadas. Assim, a
proteína ribosomal P0 e o fator de iniciação da tradução eIF4A1 foram
utilizados como controle, para padronizar quantidade celular e confirmar a
qualidade das amostras protéicas, visto que sabidamente apresentam
expressão constitutiva durante todo o ciclo de vida de espécies de
Leishmania [24] (FIGURA 1B e 1D). Uma segunda proteína ribosomal, a
L19, também teve sua expressão investigada e produziu o mesmo padrão
que a proteína P0.
Também foi utilizado um controle a fim de validar a curva de
diferenciação celular em amastigotas axênicos, a proteína HSP70
mitocondrial (mtHSP70), que possui uma expressão diferenciada em
células amastigotas. Em estudos prévios foram identificados dois
homólogos da mtHSP70 em L. chagasi e L. major, denominados de Lc2.1
e Lc2.2. Soros produzidos contra esses dois homólogos reconhecem as
duas proteínas visto que são altamente conservadas. Em extratos de
promastigotas de várias espécies de Leishmania, foi observado que a
proteína predominante é a de maior peso molecular, provavelmente a
Lc2.2. Porém, em amastigotas provenientes de lesão em camundongos
Balb/c apenas uma banda de menor peso molecular é reconhecida
(provavelmente a Lc2.1).
Nas curvas realizadas, três perfis de expressão foram observados de
mtHSP70. O primeiro perfil foi visto em promastigotas crescidos a 26ºC e
no começo da curva de diferenciação a 32ºC. Neste foram detectadas
duas bandas, onde a banda com peso molecular mais alto (Lc2.2) se
mostrou bem mais abundante que a segunda, menor (Lc2.1). Um perfil
intermediário foi visto no meio da curva de diferenciação onde a
expressão da Lc2.1 foi aumentada enquanto a Lc2.2 diminuiu até ficarem

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
80
aproximadamente com o mesmo nível de expressão. No final da curva de
diferenciação, foi observado que os níveis de expressão de Lc2.2
diminuíram drasticamente, permanecendo a Lc2.1. Como em amastigotas
de lesão foi observada apenas a expressão da banda de menor peso
molecular (Lc2.1), este resultado valida a curva de diferenciação em
amastigotas axênicas, visto que, ao final da curva a 32ºC, quando
praticamente todas as células alteraram sua morfologia e ficaram
arredondadas, o perfil de expressão delas é semelhante ao da amastigota
de lesão (FIGURA 1E) [25].
3.3 Expressão de quatro homólogos do fator eIF4E durante ciclo
de vida de L. amazonensis.
Em seguida, partiu-se para a análise da expressão dos quatro
homólogos de eIF4E de L. amazonensis. Foram realizados ensaios de
“Western-blot” com anticorpos contra os fatores de iniciação da tradução
EIF4E1-4. Foi observado que os fatores EIF4E1 e EIF4E2 estão presentes
durante toda a curva de forma constitutiva. O EIF4E1 só conseguiu ser
detectado no “Western-blot” utilizando uma concentração de células cinco
vezes maior do que a utilizada nos demais ensaios, visto que essa
proteína é pouco abundante. Já o EIF4E2, mesmo estando em menor
quantidade na célula do que o EIF4E1, conseguiu ser detectado na forma
promastigota com 106 células, porém só foi detectado na forma
amastigota com cinco vezes mais células (FIGURA 2A e 2B).
No que concerne aos fatores EIF4E3 e EIF4E4, os resultados foram
mais complexos. Ambos são expressos durante todo o ciclo de vida e
aparecem sob a forma de duas bandas, se comportando de forma
contrastante. Enquanto que no EIF4E3 a banda de maior peso molecular
diminui de intensidade e depois reaparece forte sozinha no final da curva,
no EIF4E4 a banda de maior peso molecular aumenta e depois desaparece

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
81
(FIGURA 2A). O mesmo acontece no final da curva de diferenciação do
EIF4E2, onde é também observada a presença de duas bandas. Já na
curva de diferenciação em meio Schneider pH 5,5, os fatores EIF4E3 e
EIF4E4 são expressos da mesma forma que na curva de crescimento em
meio Schneider pH 7,2, porém, ocorre uma diminuição brusca da
expressão das duas proteínas no final da curva (FIGURA 2B).
Em estudos prévios, a expressão de um dos homólogos da proteína
de ligação a cauda poli-A (PABP1) em L. amazonensis, mostrou o mesmo
padrão de duas bandas nas primeiras horas após o repique celular [26]
(FIGURA 2C). Já foi comprovado que esta segunda banda reconhecida
representa a forma fosforilada desta proteína [18]. Também já foi
comprovada a regulação de fatores de iniciação da tradução através de
fosforilação em outros organismos e há vários estudos sobre a ocorrência
de regulação por fosforilação do fator eIF4E de humano. Tudo isso nos
leva a supor que a presença de uma segunda banda, de maior peso
molecular, nos fatores EIF4E observados de L. amazonensis, poderia ser o
reconhecimento da proteína na sua forma fosforilada. Outro ponto
interessante observado nestas curvas foi a velocidade com que ocorreu a
modificação no perfil de expressão das isoformas dos fatores EIF4E3,
EIF4E4 e na PABP1, concluída em grande parte nas primeiras duas horas
após o repique e precedendo a fase de crescimento celular acelerado.
3.4 Localização subcelular dos fatores de iniciação da tradução
EIF4E3 e EIF4E4.
Com o intuito de saber se as duas bandas, ou isoformas, observadas
para os EIF4E3 e EIF4E4, estariam exercendo funções em locais diferentes
na célula, realizamos o ensaio de imunocitoquímica utilizando células
coletadas em pontos estratégicos da curva de crescimento. No ponto 6h

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
82
da curva, encontramos o fator EIF4E3 apenas com a banda de menor peso
molecular, que seria a proteína na sua forma “não fosforilada” e o EIF4E4
apenas com a banda de maior peso molecular, que seria a forma
“fosforilada” da proteína. Já no ponto de 144h, o contrário foi observado.
Portanto, estes dois pontos foram escolhidos para verificar se a
“fosforilação”/“desfosforilação” das proteínas estariam modificando sua
localização dentro da célula. Os resultados da imunocitoquímica mostram
que os dois fatores apresentaram localização citoplasmática tanto no
ponto de 6h como no ponto de 144h, mostrando que não ocorreu
modificação de localização nas isoformas encontradas destes dois fatores
(FIGURA 3).
3.5 Análise do crescimento celular na indução de estresse em
quatro condições de meio Schneider pH 7,2.
A análise dos resultados de expressão dos EIF4E3 e E4 nas curvas
de crescimento chamou a atenção para a rápida mudança no perfil de
modificação destas proteínas induzida pelo repique celular. Em um
primeiro momento especulou-se que estas modificações poderiam estar
ocorrendo devido a três fatores: (1) densidade celular – ocorre uma
diluição no começo da curva e ao final são observadas células em
abundância. Esse efeito “diluição” celular poderia servir de sinal para se
ativar/desativar eventos pós traducionais; (2) utilização de meio novo -
através do repique do meio esgotado de nutrientes para um meio rico as
células poderiam modificar sua expressão; (3) soro fetal bovino (SFB) -
altamente necessário para o crescimento celular e que poderia estar
limitado no final da curva. Para se identificar qual(is) dentre estes fatores
poderiam estar influenciando nestas modificações pós-traducionais,
elaboramos um ensaio onde quatro condições diferentes de meio foram
testadas. A primeira condição, controle (N+, meio novo com SFB),

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
83
correspondeu ao repique normal para dar início a uma nova curva. Nas
duas condições seguintes foi utilizado um meio previamente condicionado,
ou seja, já utilizado em uma curva de crescimento anterior mantida até o
estágio de morte celular. Este meio, teoricamente esgotado de nutrientes,
foi reciclado e utilizado para se estabelecer duas novas curvas, com e sem
SFB (C+ e C-, meio condicionado com e sem SFB). Na quarta condição o
repique foi realizado em meio novo, mas se evitou adicionar o SFB (N-).
Após 24 horas, as células nas quatro condições testadas foram
observadas e quantificadas (Figura 4A), onde se observou que o meio
novo sem o SFB (N-) obteve o menor crescimento, seis vezes menor que
o controle (N+). Nesta condição as células já estavam bastante
deformadas e com pouca motilidade, característico de células em fase de
declínio. O meio condicionado ainda foi capaz de permitir a multiplicação
dos parasitas, tanto na ausência de SFB (C-), que embora tenha sido o
segundo pior desempenho ainda assim foi melhor que o (N-), quanto com
suplementação pelo soro (C+), quando o crescimento celular foi de 70 a
80% do observado para o controle. Acredita-se que as células
conseguiram crescer mais na condição (C-) em relação a (N-) devido à
presença de resquícios de SFB no meio condicionado. Estes resultados
sugerem que a densidade celular é um ponto importante que leva à
parada de crescimento e sua retomada quando ocorre a diluição no meio
condicionado, mas o ponto mais relevante parece ser a presença de SFB
no meio de cultura, tendo em vista o resultado obtido na condição (N-).
3.6 Expressão dos fatores EIF4E3 e EIF4E4 nas quatro condições
de meio Schneider pH 7,2.
A Figura 5 mostra os resultados obtidos com a análise da expressão
dos EIF4E3 e 4, e também da PABP1, nas quatro condições de

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
84
crescimento testadas, imediatamente após o repique (tempo 0) e nos
períodos sucessivos de 15 minutos, 30 minutos, 1, 2, 6 e 24 horas. A
proteína PABP1 foi utilizada neste experimento como parâmetro de
comparação, visto que também sofre modificações pós-traducionais,
nesse caso comprovadamente por fosforilação, nas primeiras horas após o
repique celular. Os intervalos mais curtos para a análise foram
selecionados para se permitir uma melhor definição quanto ao período de
início das mudanças observadas após o repique. Foi observado que no
meio novo sem SFB, onde a célula é exposta a maior condição de
estresse, a fosforilação da PABP já começa a partir dos primeiros 15
minutos após o repique, em contraste com o controle, que só apresenta
fosforilação após 2h. No tempo de 24h o padrão de modificação da PABP1
é semelhante em todas as condições (FIGURA 5A). Já com o EIF4E3 a
proteína na condição (N-) de estresse começa a “desfosforilar” a partir de
1h após o repique, diferente da condição do controle, onde foi observado
o começo da “desfosforilação” com 15 minutos (FIGURA 5B). No caso do
EIF4E4 não parece haver diferenças significativas entre as várias
condições testadas nas primeiras 6 horas após o repique. Em termos de
cinética das modificações nas três proteínas, observou-se que estas
estavam presentes após 1h do repique, em qualquer condição, mas seu
efeito maior se dava em até 6h.
Chama a atenção no caso da curva (N-) para o EIF4E3 a presença
de uma terceira banda após 24h de crescimento, que poderia ser uma
“hiperfosforilação” da proteína. Este padrão é totalmente distinto do
observado no controle, ou mesmo em células em fase estacionária de
crescimento, entretanto nas células crescidas em meio condicionado, que
apresentam fenótipos intermediários entre (N-) e (N+), esta terceira
banda aparece em menor intensidade. No caso do homólogo EIF4E4
ocorre uma “desfosforilação” precoce em 24h de crescimento no meio
novo sem SFB (FIGURA 5C), compatível com o fenótipo observado na fase

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
85
estacionária de crescimento. Tanto no caso do EIF4E3 como do EIF4E4,
então, e ao contrário da PABP1, o fenótipo observado às 24h sem SFB é
compatível com uma entrada precoce na fase estacionária de crescimento,
em especial no caso do EIF4E3, onde este fenótipo é exacerbado com o
aparecimento da banda “hiperfosforilada”.
3.7 Análise de síntese de proteínas através de marcação
metabólica com metionina marcada com [35S].
A fim de se correlacionar níveis qualitativos e quantitativos de
síntese protéica com padrões de modificação dos fatores EIF4E3 e EIF4E4,
foi realizado o ensaio de marcação metabólica com 35S de alíquotas de
células retiradas de pontos representativos das curvas de crescimento em
pH 7,2. Para realizar esta marcação, escolhemos pontos onde se observou
padrões contrastantes de expressão dos EIF4E3 e EIF4E4, como nos
pontos de 6h e 144h da curva padrão, e o ponto 24h da curva gerada na
condição (N-) do item anterior. Os resultados mostraram que houve
diferença no padrão de expressão entre os três pontos citados (FIGURA
6), onde as bandas diferenciais de cada tempo estão marcadas com setas
vermelhas. É claramente observado também que a incorporação da
metionina marcada no ponto de 6h é maior do que nos outros dois
tempos, visto que as células já estão em fase de declínio e a síntese de
proteínas não está mais ocorrendo no mesmo modo. Outro fato
interessante é que apenas nos tempos de 24h e 144h observamos a
presença de uma expressão forte de uma banda de aproximadamente 70
KDa (seta verde). No geral é possível correlacionar níveis mais elevados
de síntese protéica com a presença da banda “defosforilada” do EIF4E3 e
da banda fosforilada do EIF4E4.

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
86
3.8 Expressão dos fatores EIF4E3 e EIF4E4 na presença de
inibidores de transcrição e tradução.
Curvas de crescimento foram realizadas sob condição de inibição de
transcrição e tradução de forma a verificar se estes inibidores poderiam
afetar os padrões de modificação pós-traducional destas proteínas. Para a
inibição da transcrição, foi utilizada a actinomicina D (derivada de
Streptomyces), que inibe a transcrição por ligar-se fortemente e
especificamente ao DNA dúplex e, assim, impedir que ele seja molde para
a transcrição [27]. Já para a inibição da tradução, foi utilizada a
cicloheximida, que inibe a atividade de peptidil transferase dos ribossomos
de eucariotos (80S) [28]. A curva de inibição da transcrição com
actinomicina D interferiu no padrão de expressão do fator EIF4E3, pois foi
observada uma “desfosforilação” tardia em 24h. Já no ponto 6h são
encontradas as duas isoformas em níveis equivalentes, diferentemente do
controle que no ponto 6h a proteína aparece totalmente “desfosforilada”
(FIGURA 7). Já no fator EIF4E4, o inibidor provocou uma “desfosforilação”
no ponto de 6h, que deveria estar “fosforilado”, e continuou
“desfosforilado” em 24h, onde deveria apresentar as duas isoformas. Já na
curva utilizando cicloheximida, a inibição da tradução não interfere na
resposta do fator EIF4E4. Porém o contrário é observado com o fator
EIF4E3, pois a expressão da proteína não varia e é observado um bloqueio
da sua “desfosforilação”.
4. DISCUSSÃO
Através deste estudo, foi possível identificar em L. amazonensis que,
entre os quatro homólogos de eIF4E analisados, os EIF4E3 e EIF4E4
apresentaram ao menos duas isoformas claramente identificáveis, com

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
87
um padrão de expressão que varia durante suas diferentes formas de
vida. Até agora, não foi realizado nenhum estudo sobre a fosforilação do
fator eIF4E em Leishmania. Entretanto, fortes evidências nos levam a
pensar que a isoforma de maior peso molecular nestes homólogos seja
conseqüência desse mecanismo pós-traducional, em especial a ocorrência
de estudos sobre fosforilação do fator eIF4E em outros organismos como
Drosophila [29] e humanos [30, 31, 32, 33, 34]. Em L. major, o evento
de fosforilação foi confirmado na PABP1 através da incorporação “in vivo”
de [32P] [18]. Esta fosforilação também foi observada em L. amazonensis,
nas primeiras horas após o repique na curva de crescimento e
diferenciação [26].
Além de apresentarem variação na expressão e duas isoformas,
observamos também que a “fosforilação/desfosforilação” dos fatores
EIF4E3 e 4 se apresentou invertida, isto é, quando um está “fosforilado” o
outro está “desfosforilado” e vice-versa, sendo que no meio da curva as
duas isoformas estão presentes em ambas as proteínas. Isso nos leva a
supor que estes homólogos poderiam estar atuando na célula e na
iniciação da tradução alternadamente, ou na tradução de mRNAs
específicos que sejam expressos em diferentes fases do complexo ciclo de
vida do parasita. Estas mudanças transientes da expressão gênica durante
a diferenciação, desenvolvimento, ou progressão do ciclo celular também
já foram descritas em outros organismos como Drosophilla [35],
Plasmodium falciparum [36], Saccharomyces cerevisiae [37] e humanos
[38].
Devido ao número elevado de fosfoproteínas encontradas em
tripanossomatídeos, eventos de fosforilação podem ser o mecanismo
chave para a regulação, visto que este número é consideravelmente maior
do que o encontrado para outros parasitas intracelulares que transitam
em ambientes distintos [39]. A quantidade de proteínas quinases
presentes nos tripanosomatídeos é cerca de 33% maior do que em S.

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
88
cerevisiae e duas vezes maior que em P. falciparum [40, 41]. Um estudo
recente mostra a participação de proteínas da família MAP quinases que
estão envolvidas na regulação do tamanho do flagelo em L. mexicana. A
proteína LmxMKK não é detectada em amastigotas, é altamente regulada
durante a diferenciação para promastigotas com níveis reduzidos durante
o começo desta diferenciação. Além disso, a proteína LmxMPK3 é capaz de
fosforilar LmxMKK, indicando uma possível regulação por “feedback” [42].
A diferenciação de promastigotas para amastigotas em Leishmania é
um processo complexo, porém as mudanças morfológicas que ocorrem no
parasita parecem ser bem coordenadas e reguladas [43]. Um estudo
recente foi realizado investigando mudanças na abundância do RNA
durante a diferenciação promastigota-amastigota de L. donovani em meio
de cultura. Este estudo revelou que vários genes sofrem regulações
transientes e permanentes durante a diferenciação e que a maioria das
mudanças ocorre no final deste processo, quando a diferenciação
morfológica é praticamente completa. Genes que possuem regulação
transiente durante a diferenciação incluem aqueles que codificam
proteínas de choque térmico, proteínas que interagem com o RNA,
proteínas quinases, entre outros [44]. Outro estudo recente com L. major
mostrou, através de PCR em tempo real e da técnica de microarranjos
(“Microarrays”), que a maioria dos mRNAs de L. major são
constitutivamente expressos e a presença de diferentes níveis de mRNA
encontrados nas formas amastigotas e promastigotas ocorre
provavelmente por eventos pós-transcricionais que leva a uma diferença
na estabilidade dos mRNAs [45].
Nossos resultados mostraram uma expressão diferenciada nos
primeiros 15 minutos do repique celular, sugerindo que o perfil de
expressão observado é devido realmente a modificações pós-traducionais,
e não a eventos que dependam de transcrição ou tradução. No caso da
expressão das proteínas nas curvas de inibição, observamos que o padrão

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
89
de expressão do fator EIF4E4 não é modificado na presença do inibidor de
tradução, o que ocorre também com a proteína PABP1 (dados não
mostrados). No caso do fator EIF4E3 há mudanças no comportamento
pós-traducional da proteína, onde ocorre um bloqueio da “desfosforilação”
em 6h. Já com a presença do inibidor de transcrição, ocorreu mudanças
no perfil de expressão de ambos os homólogos. Ocorre uma
“desfosforilação” tardia do EIF4E3 em 24h, e, no caso do EIF4E4, uma
“desfosforilação” que começa a partir de 6h, ponto onde a proteína
deveria estar apenas na sua forma “fosforilada”. Na PABP1 em L. major, a
actinomicina D modificou tanto seu padrão de expressão, como sua
localização subcelular, sugerindo que a expressão desta proteína poderia
ser regulada ao nível pós-transcricional em resposta a abundância de
mRNA presente na célula [18].
Através do ensaio de marcação metabólica, foi possível observar que
a incorporação da metionina praticamente não ocorreu nos tempos de 24h
sem SFB e 144h, pontos onde a célula está em fase de declínio e, por
isso, a biossíntese protéica é baixa. Devido a isso, podemos sugerir que a
atividade dos fatores EIF4E3 e 4 ocorre quando estão nas isoformas
“desfosforilada” e “fosforilada” respectivamente, visto que no ponto de
elevada biossíntese protéica (6h), os fatores EIF4E3 e 4 encontram-se
nestas isoformas. Além disso, na inibição da tradução, o fator EIF4E3 não
consegue desfosforilar totalmente no ponto de 6h, indicando que se não
está ocorrendo tradução, não precisa haver a modificação da proteína
para a sua forma ativa. Através de uma análise da associação de
proteínas com os poliribossomos, por exemplo, poderíamos observar a
presença de alguma isoforma, que seria um indicativo da participação da
proteína na tradução. Isso confirmaria nossas suposições de que a
“desfoforilação” do EIF4E3 e a “fosforilação” do EIF4E4 estariam ativando
a proteína para participação na biossíntese protéica.
Com todos os dados obtidos neste trabalho, juntamente com os

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
90
outros dados, seria possível sugerir que os fatores EIF4E3 e EIF4E4
poderiam estar atuando como fatores de iniciação da tradução na
Leishmania. Já os outros fatores poderiam estar envolvidos, através de
funções modulatórias, indiretamente na regulação da tradução. Para
confirmar essas suposições, é necessária a realização de maiores estudos
sobre estes fatores e como ocorre seu mecanismo de regulação pós-
traducional, que parece ser de extrema importância para o mecanismo de
biossíntese protéica e crucial na sobrevida do parasita.
5. Referências
[1] TDR - Special Programme for Research and Training in Tropical
Diseases. http://www.who.int/tdr (Dezembro, 2007).
[2] Handman E. (2001) Leishmania virulence: it's a knock out! Trends
Parasitol. 17 (2): 60.
[3] Besteiro S, Williams RAM, Coombs GH, Mottram JC. (2007) Protein
turnover and differentiation in Leishmania. Int J Parasitol.
37(10):1063-75.
[4] Clayton CE. (2002) Life without transcriptional control? From fly to
man and back again. EMBO J. 21:1881-8.
[5] Gebauer F and Hentze MW (2004) Molecular mechanisms of
translational control. Nat.Rev.Mol.Cell Biol. 5: 827-835.
[6] Prévôt D, Darlix JL, Ohlmann T (2003) Conducting the initiation of
protein synthesis: the role of eIF4G. Biol Cell 95(3-4):141-56.

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
91
[7] Hershey JWB and Merrick WC (2000) The pathway and mechanism of
initiation of protein synthesis. Translational Control of Gene
Expression (Cold Spring Habor Laboratory Press); pp. 33-38.
[8] Sonenberg N and Dever TE (2003) Eukaryotic translation initiation
factors and regulators. Curr Opin Struct Biol. 13: 56-63.
[9] Lloyd RE (2006) Translational control by viral proteinases. Virus Res.
119 (1): 76-88.
[10] von der Haar T, Ball PD and McCarthy JE (2000) Stabilization of
eukaryotic initiation factor 4E binding to the mRNA 5'-Cap by
domains of eIF4G. J Biol Chem. 275: 30551-30555.
[11] Rom E, Kim HC, Gingras AC, Marcotrigiano J, Favre D, Olsen H,
Burley SK, Sonenberg N. (1998) Cloning and characterization of
4EHP, a novel mammalian eIF4E-related cap-binding protein. J Biol
Chem. 273 (21): 13104-9.
[12] Goodfellow IG, Roberts LO. (2007) Eukaryotic initiation factor 4E. Int
J Biochem Cell Biol. [in press].
[13] Jankowska-Anyszka M, Lamphear BJ, Aamodt EJ, Harrington T,
Darzynkiewicz E, Stolarski R, Rhoads RE. (1998) Multiple isoforms of
eukaryotic protein synthesis initiation factor 4E in Caenorhabditis
elegans can distinguish between mono- and trimethylated mRNA
cap structures. J Biol Chem. 273 (17): 10538-42.
[14] Keiper BD, Lamphear BJ, Deshpande AM Jankowska-Anyszka M,

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
92
Aamodt EJ, Blumenthal T, Rhoads RE (2000) Functional
characterization of five eIF4E isoforms in Caernorhabditis elegans. J
Biol Chem. 275; 10590-10596.
[15] Sonenberg N (1996). Translational control. In: Stabilization of
eukaryotic initiation factor 4E binding to the mRNA 5’-cap by
domains of eIF4G. Biol Chem. 39: 30551-30555.
[16] Gingras AC, Raught B and Sonenberg N (1999) eIF4 initiation
factors: effectors of mRNA recruitment to ribosomes and regulators
of translation. Annu Rev Biochem. 68: 913-963.
[17] Batista JA, Teixeira SM, Donelson JE, Kirchhoff LV and de Sa CM
(1994) Characterization of a Trypanosoma cruzi poly(A)-binding
protein and its genes. Mol Biochem Parasitol. 67: 301-312.
[18] Bates EJ, Knuepfer E and Smith DF (2000) Poly(A)-binding protein I
of Leishmania: functional analysis and localisation in trypanosomatid
parasites. Nucleic Acids Res. 28: 1211-1220.
[19] Yoffe Y, Zuberek J, Lewdorowicz M, Zeira Z, Keasar C, Orr-Dahan I,
Jankowska-Anyszka M, Stepinski J, Darzynkiewicz E, Shapira M.
(2004) Cap-binding activity of an homolog from Leishmania. RNA.
10 (11): 1764-75.
[20] Ivens AC, Peacock CS, Worthey EA, et al. (2005) The genome of the
kinetoplastid parasite, Leishmania major. Science. 309: 436-442.
[21] Dhalia R, Reis CR, Freire ER, et al. (2005) Translation initiation in
Leishmania major: characterisation of multiple eIF4F subunit

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
93
homologues. Mol Biochem Parasitol. 140: 23-41.
[22] Uliana SRB, Goyal N, Freymüller E and Smith DF (1999) Leishmania:
overexpression ad comparative structural analysis of the stage-
regulated meta 1 gene. Experimental Parasitology. 92: 183-191.
[23] de Melo Neto OP, Standart N, and Martins de, SC (1995)
Autoregulation of poly(A)-binding protein synthesis in vitro. Nucleic
Acids Res. 23 (12): 2198-2205
[24] Rocha PO (2006) Análise da expressão de fatores envolvidos na
síntese de proteínas durante o ciclo de vida de Leishmania sp.
Dissertação de Mestrado do Programa de Pós Graduação em
Genética da Universidade Federal de Pernambuco, Recife-PE, Brasil.
[25] Campos RM, Nascimento M, Ferraz JC, Pereira MMC, Rocha PO,
Thompson GM, Cysne-Finkelstein L, Figueiredo RCBQ and de Melo Neto
OP (2008) Distinct mitochondrial HSP70 homologues conserved in
various Leishmania species suggest novel biological functions. Artigo
Submetido à revista Molecular and Biochemical Parasitology.
[26] da Costa Lima TDC (2007) Caracterização funcional de homólogos da
proteína de ligação a cauda poli-A (PABP) de Leishmania major.
Dissertação de Mestrado do Programa de Pós Graduação em
Genética da Universidade Federal de Pernambuco, Recife-PE, Brasil.
[27] ICB – Intituto de Ciências biológicas
http://www.icb.ufmg.br/prodabi/prodabi3/grupos/grupo1/inibidores.h
tm (Dezembro, 2007).

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
94
[28] FIOCRUZ – Fundação Oswaldo Cruz
http://www.dbbm.fiocruz.br/helpdesk/mbiology/aula_Traducao.pdf
(Dezembro, 2007).
[29] Lachance PE, Miron M, Raught B, Sonenberg N, Lasko P. (2002)
Phosphorylation of eukaryotic translation initiation factor 4E is
critical for growth. Mol Cell Biol. 22 (6): 1656-63.
[30] Hiremath LS, Hiremath ST, Rychlik W, Joshi S, Domier LL, Rhoads
RE. (1989) In vitro synthesis, phosphorylation, and localization on
48 S initiation complexes of human protein synthesis initiation factor
4E. J Biol Chem. 264 (2): 1132-8.
[31] Joshi B, Cai AL, Keiper BD, Minich WB, Mendez R, Beach CM,
Stepinski J, Stolaeski R, Darzynkiewicz E and Rhoads RE (1995)
Phosphorilation of eukaryotic protein synthesis factor 4E at Ser-209.
J Biol Chem 270: 14597-14603.
[32] Wang X, Flynn A, Waskiewicz AJ, Webb BL, Vries RG, Baines IA,
Cooper JA, Proud CG. (1998) The phosphorylation of eukaryotic
initiation factor eIF4E in response to phorbol esters, cell stresses,
and cytokines is mediated by distinct MAP kinase pathways. J Biol
Chem. 273 (16): 9373-7.
[33] Scheper GC, van Kollenburg B, Hu J, Luo Y, Goss DJ, Proud CG
(2002) Phosphorylation of eucariotic initiation factor 4E markedly
reduces its affinity for capped mRNA. J Biol Chem. 277(5); 3303-
3309.

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
95
[34] Zuberek J, Wyslouch-Cieszynska A, Niedzwiecka A, Dadlez M,
Stepinski J, Augustyniak W, Gingras AC, Zhang Z, Burley SK,
Sonenberg N, Stolarski R, Darzynkiewicz E. (2003) Phospholylation
of eIF4E attenuates its interation with mRNA 5’ cap analogs by
electrostatic repulsion: inten-mediated protein ligation strategy to
obtain phosphorylated protein. RNA. 9 (1): 52-61.
[35] Wagner RA, Tabibiazar R, Liao A, Quertermous T. (2005) Genome-
wide expression dynamics during mouse embryonic development
reveal similarities to Drosophila development. Dev Biol. 288: 595–
611.
[36] Bozdech Z, Llinas M, Pulliam B, Wong E, Zhu J, DeRisi J. (2003) The
transcriptome of the intraerythrocytic developmental cycle of
Plasmodium falciparum. PLOS. 1:E5.
[37] Spellman PT, Sherlock G, Zhang MQ, et al. (1998) Comprehensive
identification of cell cycle-regulated genes of the yeast
Saccharomyces cerevisiae by microarray hybridization. Mol Biol Cell.
9: 3273–97.
[38] Whitfield ML, Sherlock G, Saldanha AJ, et al. (2002) Identification of
genes periodically expressed in the human cell cycle and their
expression in tumors. Mol Biol Cell. 13: 1977–2000.
[39] Parsons M, Worthey EA, Ward PN, Mottram JC (2005). Comparative
analysis of the kinomes of three pathogenic trypanosomatids:
Leishmania major, Trypanosoma brucei and Trypanosoma cruzi.
BMC Genomics; 15; 6: 127.

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
96
[40] Ward P, Equinet L, Packer J, Doerig C. (2004) Protein kinases of the
human malaria parasite Plasmodium falciparum: the kinome of a
divergent eukaryote. BMC Genomics. 5 (1): 79.
[41] Naula C, Parsons M, and Mottram JC (2005) Protein kinases as drug
targets in trypanosomes and Leishmania. Biochim Biophys Acta
1754 (1-2): 151-9.
[42] Erdmann M, Scholz A, Melzer IM, Schmetz C, Wiese M. (2006)
Interacting protein kinases involved in the regulation of flagellar
length. Mol Biol Cell. 17 (4): 2035-45.
[43] Barak E, Amin-Spector S, Gerliak E, Goyard S, Holland N, Zilberstein
D. (2005) Differentiation of Leishmania donovani in host-free
system: analysis of signal perception and response. Mol Biochem
Parasitol. 141: 99– 108.
[44] Saxena A, Lahav T, Holland N, Aggarwal G, Anupama A, Huang Y,
Volpin H, Myler PJ, Zilberstein D. (2007) Analysis of the Leishmania
donovani transcriptome reveals an ordered progression of transient
and permanent changes in gene expression during differentiation.
Mol Biochem Parasitol. 152 (1): 53-65.
[45] Leifso K, Cohen-Freue G, Dogra N, Murray A, McMaster WR. (2007)
Genomic and proteomic expression analysis of Leishmania
promastigote and amastigote life stages: the Leishmania genome is
constitutively expressed. Mol Biochem Parasitol. 152 (1): 35-46.

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
97
(B)Curva de Crescimento em Meio Schneider 7,2
0
10
20
30
40
50
60
0h 2h 6h 24h 48h 72h 96h 120h 144h
Tempo (horas)
Nº d
e c
élul
as (
106 )
Fase logestacionária
declínio
Fase lag
Formas metacíclicas
Curva de Crescimento – Meio Schneider pH 7,2 (B)Curva de Crescimento em Meio Schneider 7,2
0
10
20
30
40
50
60
0h 2h 6h 24h 48h 72h 96h 120h 144h
Tempo (horas)
Nº d
e c
élul
as (
106 )
Fase logestacionária
declínio
Fase lag
Formas metacíclicas
Curva de Crescimento – Meio Schneider pH 7,2
Amastigota de lesão
26ºC pH 7,2 32ºC pH 5,5
0h 24h 72h 120h 0h 24h 72h 120h 144h
(E)
Amastigota de lesão
26ºC pH 7,2 32ºC pH 5,5
0h 24h 72h 120h 0h 24h 72h 120h 144h
(E)
(A) (B)
pH 7,2 – 26ºC
0h 2h 6h 24h 48h 72h 96h 120h 14 4h
EIF4A1
P0
L19
pH 7,2 – 26ºC
0h 2h 6h 24h 48h 72h 96h 120h 14 4h
EIF4A1
P0
L19
(C) (D)
Curva de Crescimento em Meio Schneider 5,5
0
3
6
9
12
15
0h 2h 6h 24h 48h 72h 96h 120h 144h
Tempo (horas)
Nº
de c
élul
as (
x10
6 ) Fase logestacionária
declínioFase lag
Amastigotas “like”
Curva de Crescimento – Meio Schneider pH 5,5Curva de Crescimento em Meio Schneider 5,5
0
3
6
9
12
15
0h 2h 6h 24h 48h 72h 96h 120h 144h
Tempo (horas)
Nº
de c
élul
as (
x10
6 ) Fase logestacionária
declínioFase lag
Amastigotas “like”
Curva de Crescimento – Meio Schneider pH 5,5pH 5,5 – 32ºC
0h 2h 6h 24h 48h 72h 96h 120h
EIF4A1
P0
L19
pH 5,5 – 32ºC
0h 2h 6h 24h 48h 72h 96h 120h
EIF4A1
P0
L19
Figura 1. Padrão de crescimento e diferenciação de L. amazonensis em cultivo in vitro. (A) Curva de crescimento de células promastigotas procíclicas e metacíclicas em meio Schneider pH 7,2, a 26oC. (B) Expressão das proteínas EIF4A1, P0 e L19 utilizadas como controle da curva de crescimento em pH 7,2. (C) Curva de diferenciação de células amastigotas “like” em meio Schneider pH 5,5, a 32oC. (D) Expressão das proteínas EIF4A1, P0 e L19 utilizadas como controle da curva de diferenciação em pH 5,5. (E) Expressão da proteína mtHSP70 em pontos representativos das curva de crescimento e diferenciação e em extrato de amastigotas retiradas de lesão de camundongo Balb/c. (Figura modificada do artigo Campos et al., 2008, submetido a revista Molecular and Biochemical Parasitology). As alíquotas foram fracionadas em SDS-PAGE 15% e analisados por “Western-Blot”. Todas as alíquotas contém 106 células, com exceção da amastigota de lesão que contém 2,5 x 106 células.
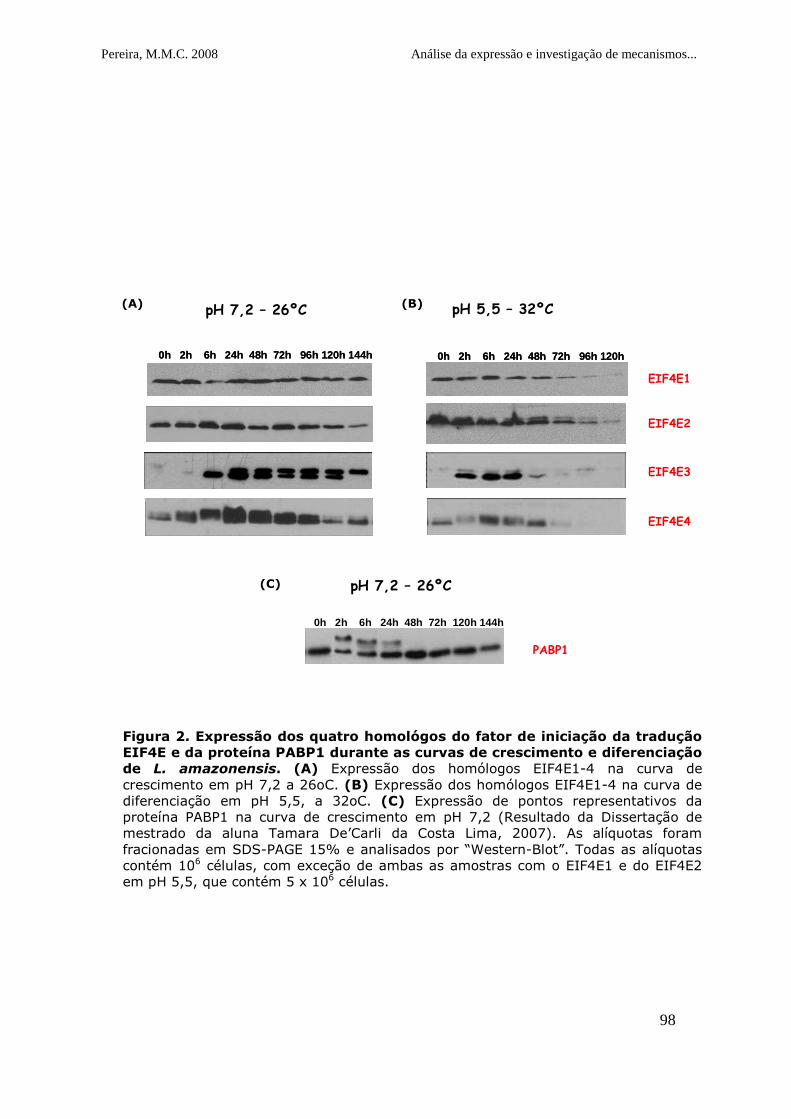
Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
98
EIF4E3
EIF4E4
EIF4E1
EIF4E2
(A)
(C)
PABP1
pH 7,2 – 26ºC
(B)pH 7,2 – 26ºC
0h 2h 6h 24h 48h 72h 96h 120h 144h0h 2h 6h 24h 48h 72h 96h 120h 144h
pH 5,5 – 32ºC
0h 2h 6h 24h 48h 72h 96h 120h0h 2h 6h 24h 48h 72h 96h 120h
0h 2h 6h 24h 48h 72h 120h 144h
Figura 2. Expressão dos quatro homológos do fator de iniciação da tradução EIF4E e da proteína PABP1 durante as curvas de crescimento e diferenciação de L. amazonensis. (A) Expressão dos homólogos EIF4E1-4 na curva de crescimento em pH 7,2 a 26oC. (B) Expressão dos homólogos EIF4E1-4 na curva de diferenciação em pH 5,5, a 32oC. (C) Expressão de pontos representativos da proteína PABP1 na curva de crescimento em pH 7,2 (Resultado da Dissertação de mestrado da aluna Tamara De’Carli da Costa Lima, 2007). As alíquotas foram fracionadas em SDS-PAGE 15% e analisados por “Western-Blot”. Todas as alíquotas contém 106 células, com exceção de ambas as amostras com o EIF4E1 e do EIF4E2 em pH 5,5, que contém 5 x 106 células.

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
99
144 horas
6 horas
(A)
(B)
EIF4E3 EIF4E4
EIF4E3 EIF4E4
Figura 3. Localização subcelular dos fatores EIF4E3 e EIF4E4. (A) Células com 6h de crescimento em meio com pH 7,2, a 26º C. (B) Células com 144h de crescimento em meio com pH 7,2, a 26º C. Mesmo com o marcador de núcleo e cinetoplasto não estando evidente, nota-se que os dois homólogos apresentaram localização citoplasmática.
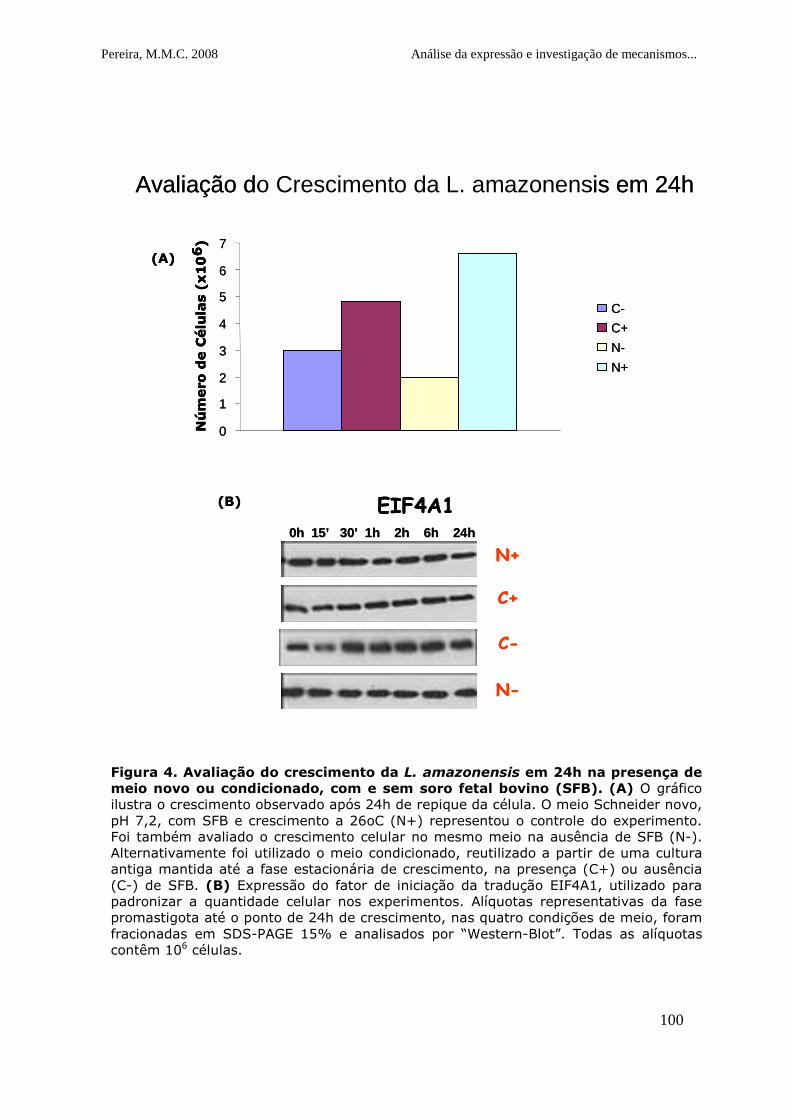
Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
100
Figura 4. Avaliação do crescimento da L. amazonensis em 24h na presença de meio novo ou condicionado, com e sem soro fetal bovino (SFB). (A) O gráfico ilustra o crescimento observado após 24h de repique da célula. O meio Schneider novo, pH 7,2, com SFB e crescimento a 26oC (N+) representou o controle do experimento. Foi também avaliado o crescimento celular no mesmo meio na ausência de SFB (N-). Alternativamente foi utilizado o meio condicionado, reutilizado a partir de uma cultura antiga mantida até a fase estacionária de crescimento, na presença (C+) ou ausência (C-) de SFB. (B) Expressão do fator de iniciação da tradução EIF4A1, utilizado para padronizar a quantidade celular nos experimentos. Alíquotas representativas da fase promastigota até o ponto de 24h de crescimento, nas quatro condições de meio, foram fracionadas em SDS-PAGE 15% e analisados por “Western-Blot”. Todas as alíquotas contêm 106 células.
Avaliação do Crescimento da L. amazonensis em 24h
0
1
2
3
4
5
6
7
Número de Células (x106)
C-
C+
N-
N+
(A)
Avaliação do Crescimento da L. amazonensis em 24h
0
1
2
3
4
5
6
7
Número de Células (x106)
Número de Células (x106)
C-
C+
N-
N+
(A)
(B)
0h 15’ 30’ 1h 2h 6h 24h
EIF4A1
N-
N+
C-
C+
(B)
0h 15’ 30’ 1h 2h 6h 24h
EIF4A1
N-
N+
C-
C+

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
101
(A) (B)
EIF4E30h 15’ 30’ 1h 2h 6h 24h
N-
N+
C-
C+
N-
N+
C-
C+
PABP10h 15’ 30’ 1h 2h 6h 24h
(C)
EIF4E40h 15’ 30’ 1h 2h 6h 24h
N-
N+
C-
C+
Figura 5. Expressão dos fatores EIF4E3, EIF4E4 e a PABP1 nas culturas crescidas por 24 horas na presença de meio novo ou condicionado, com e sem soro fetal bovino (SFB). (A) PABP1, (B) EIF4E3, (C) EIF4E4. As culturas foram definidas como descrito na Figura 4. Alíquotas representativas da fase promastigota até o ponto de 24h de crescimento, nas quatro condições de meio, foram fracionadas em SDS-PAGE 15% e analisados por “Western-Blot”. Todas as alíquotas contêm 106 células.

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
102
pH 7,2 – 26ºC
97KDa
66KDa
45KDa
30KDa
20KDa
14,4KDa
6h 144h 24h s/ SFB
97KDa
66KDa
45KDa
30KDa
20KDa
14,4KDa
6h 144h 24h s/ SFB
Figura 6. Análise do perfil de síntese protéica em células de L. amazonensis em diferentes estágios de crescimento. Marcação metabólica com metionina marcada com 35S, comparando mudanças no padrão de expressão protéica de células de L. amazonensis derivadas de fases representativas de sua curva padrão de crescimento (6h e 144h) ou de células retiradas da curva (N-, ausência de soro fetal bovino), da Figura 4, no tempo de 24h. As setas vermelhas indicam proteínas presentes apenas na fase exponencial do crescimento do parasita, enquanto que as setas verdes indicam proteínas presentes apenas na estacionárias. Maiores mudanças não puderam ser observadas devido a pouca incorporação de metionina no tempo de 24h sem soro fetal. Todas as alíquotas contém 106 células.
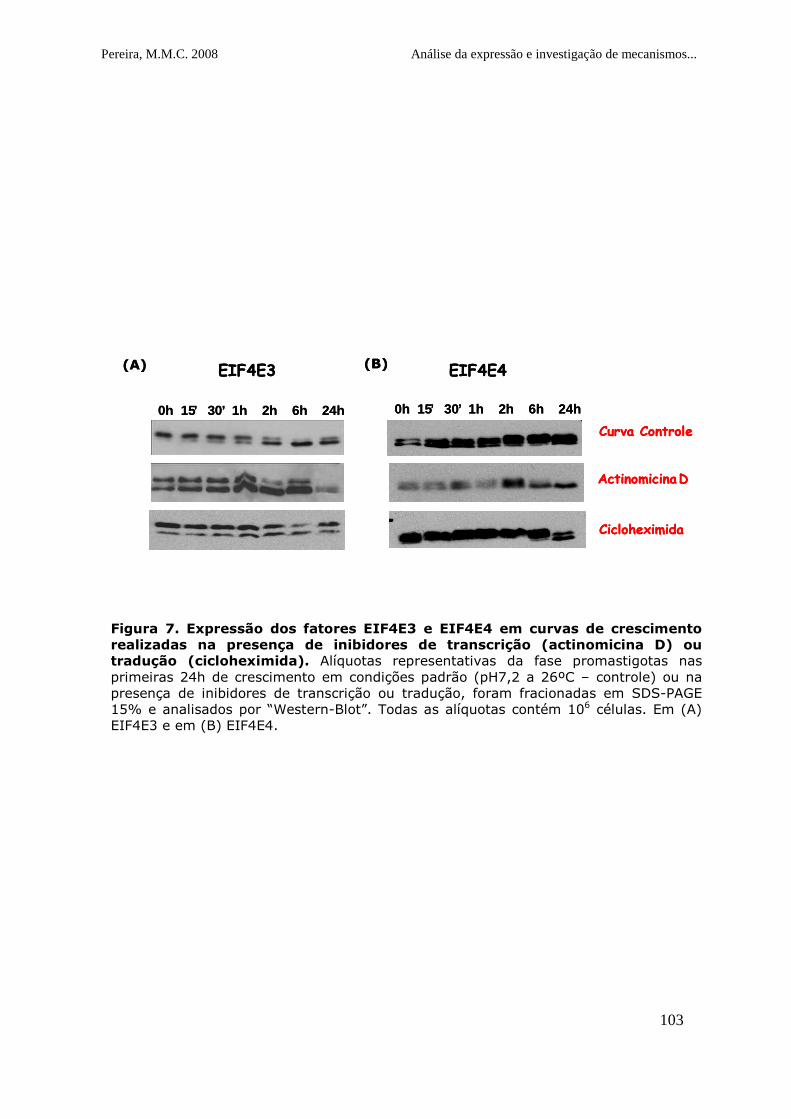
Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
103
Figura 7. Expressão dos fatores EIF4E3 e EIF4E4 em curvas de crescimento realizadas na presença de inibidores de transcrição (actinomicina D) ou tradução (cicloheximida). Alíquotas representativas da fase promastigotas nas primeiras 24h de crescimento em condições padrão (pH7,2 a 26ºC – controle) ou na presença de inibidores de transcrição ou tradução, foram fracionadas em SDS-PAGE 15% e analisados por “Western-Blot”. Todas as alíquotas contém 106 células. Em (A) EIF4E3 e em (B) EIF4E4.
EIF4E3
0h 15’ 30’ 1h 2h 6h 24h
Curva Controle
ActinomicinaD
Cicloheximida
EIF4E4
0h 15’ 30’ 1h 2h 6h 24h
(A) (B)EIF4E3
0h 15’ 30’ 1h 2h 6h 24h
Curva Controle
ActinomicinaD
Cicloheximida
EIF4E4
0h 15’ 30’ 1h 2h 6h 24h
(A) (B)

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
104
6. CONCLUSÕES •••• Os quatro homólogos do EIF4E estão presentes ao longo da curva de
crescimento de L. amazonensis e possuem padrões de expressão
diferenciados, indicando que os homólogos atuam de forma distinta na
célula durante seu ciclo de vida;
• Os fatores EIF4E2-4 apresentaram um padrão de expressão com duas
isoformas, onde a banda de maior peso molecular deve ser provocada por
modificações pós-traducionais, provavelmente a fosforilação;
•••• Os fatores EIF4E3 e 4 são citoplasmáticos e apresentaram padrões de
expressão contrastante e variáveis de acordo com a fase de crescimento,
indicando uma possível complementaridade funcional destas proteínas na
iniciação da tradução;
• O repique modifica o padrão de expressão das células, que é induzido
pela diluição da célula no meio e pelo soro fetal bovino;
•••• A biossíntese protéica alta no ponto de 6h e baixa no ponto de 144h,
juntamente com o bloqueio da desfosforilação do EIF4E3 na curva de
inibição de tradução é um forte indício de que a atividade do EIF4E3 e 4
ocorre com as proteínas na sua forma "desfosforilada" e fosforilada
respectivamente.

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
105
7. ABSTRACT The trypanosomatid protozoans cause clinical diseases of global impact
and have peculiar biochemical characteristics that differ from the others
eukaryotes. Gene expression in these parasites is regulated at the post-
transcriptional level, probably at the translation initiation step. The cap-
dependent translation begins with the binding of the translation initiation
factor eIF4E, belonging to the eIF4F complex, to the cap present in the 5'
end of the mRNAs. Multiple homologues to EIF4E were identified in
trypanosomatids, which are probably associated to their complex life
cycle. In this study, our aim was to understand the expression pattern of
four homologues (EIF4E1-4) during the life cycle of Leishmania
amazonensis. Growth curves were performed in order to obtain the three
main growth forms of the parasite. All homologues were expressed
throughout the curve and EIF4E2-4 display multiple isoforms, probably
due to post-translational modifications through phosphorylation. The
EIF4E3 and 4 isoforms display a contrasting pattern of expression which
do not interfere with their subcellular cytoplasmic localization. Lack of fetal
calf serum in the medium induced a premature “dephosphorylation” of
EIF4E4 and a "hyperphosphorylation" of EIF4E3, associated with lack of
cell growth. “Non-phosphorylated” EIF4E3 and “phosphorylated” EIF4E4
were associated with high protein synthesis levels. The presence of a
transcription inhibitor influenced the expression pattern of the two
homologues, but a translation inhibitor affected only EIF4E3. These results
indicate that the eIF4E homologues act in distinct ways in the cell and that
phosphorylation may have a significant role in the regulation of these
factors in Leishmania and in the control of global protein biosynthesis.
Key-Words: Leishmania amazonensis, EIF4E, phospholylation.

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
106
8. ANEXOS
8.1 Instruções para Autores
RREEVVIISSTTAA:: Molecular and Biochemical Parasitology
Guide for Authors
Submission of a paper to Molecular and Biochemical Parasitology,
including a revised version, implies the transfer of copyright from the
author(s) to the publisher and therefore that the corresponding author has
obtained the approval of all other authors to the text and that it does not
contain information previously published (except as a meeting abstract or
by submission of sequence data to an electronic database) and is not
under consideration for publication elsewhere. Publication in Molecular and
Biochemical Parasitology is taken to imply the authors' willingness to
comply with reasonable requests to supply reagents such as recombinant
clones and monoclonal antibodies, and sequence data in electronic form to
persons lacking access to computer databases.
Manuscripts returned for revision should be returned to the editor within 3
months. Papers accepted for publication should be as concise as possible
and should be no longer than 14 printed pages. In exceptional cases the
editors will consider longer papers (never exceeding 20 printed pages) if
the authors of such complex papers show to the satisfaction of the editors
that the limitation in length would result in subdivision of the material into
several papers and hence in an increase in the total number of pages
necessary for the presentation of the work.
Online Submission Submission to this journal is now totally online. Please

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
107
use the following guidelines to prepare your article. Via the online
submission page of this journal (http://ees.elsevier.com/molbio/) you will
be guided stepwise through the creation and uploading of the various files.
The system automatically converts source files to a single Adobe Acrobat
PDF version of the article, which is used in the peer review process. Please
note that even though manuscript source files are converted to PDF at
submission for the review process, these source files are needed for
further processing after acceptance. All correspondence, including
notification of the Editor's decision and request for revision, takes place by
email and via the author's homepage, removing the need for a hard-copy
paper trail.
The above represents a brief outline of this form of submission. It can be
advantageous to print this "guide for authors" from the site
(http://authors.elsevier.com/) for reference in the subsequent stages of
article preparation.
Authors' rights
As an author you (or your employer) may do the following:
- make copies (print or electronic of the article for your own personal use,
including for your own classroom teaching use
- make copies and distribute such copies (including through email) of the
article to research colleagues, for the personal use by such colleagues(but
not commercially or systematiccally, e.g., via an email list or list servier)
- post a pre-print version of the article on Internet websites including
electronic pre-print servers, and to retain indefinitely such versions on
such servers or sites

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
108
- post a revised version of the final text of the article (to reflect changes
made in the peer review and editing process) on your personal or
institutional website or server, with a link to the journal homepage on
www.elsevier.com
- present the article at a meeting or conference and to distribute copies of
the article to delegates attending such a meeting
- for your employer, if the article is a 'work for hire', made within the
scope of your employment, your employer may use all or part of the
information in the article for other intra-company use (e.g., training)
- retain patent and trademark rights and rights to any processes or
procedure described in the article
- include the article in full or in part in a thesis or dissertation (provided
that this is not to be published commercially)
- use the article or any part thereof in a printed compilation of your works,
such as collected writings or lecture notes (subsequent to publication of
your article in the journal)
- prepare other derivative works, to extend the article into book-length
form, or to otherwise re-use portions or excerpts in other works, with full
acknowledgement of its original publication in the journal.
Protein and Nucleic Acid Sequences. Novel nucleotide or protein sequence
data must be deposited in the GenBank™, EMBL or DDBJ databases and
an accession number obtained before the paper can be accepted for

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
109
publication. Submission to any one of the collaborating databanks is
sufficient to ensure entry in all. The accession number should be included
as a footnote on the title page of the manuscript:
'Note: Nucleotide sequence data reported in this paper are available in the
GenBank™, EMBL and DDBJ databases under the accession number(s)----
'. If requested the database will withhold release of data until publication.
The usual method for submitting sequence data is by World Wide Web to
either GenBank™ (via Banklt: http://www.ncbi.nlm.nih.gov/BankIt/),
EMBL (via Webln: http://www.ebi.ac.uk/subs/allsubs.html) or to DDBJ
(via SAKURA: http://sakura.ddbj.nig.ac.jp/). Special types of submissions
such, as genomes, bulk submissions, segmented sets, and
population/phylogenetic/mutation studies, can be more easily prepared
with the Sequin programme (available from the above Web sites). Files
generated by the Sequin programme may be sent via e-mail to GenBank™
(submissions: e-mail: [email protected]; enquiries: e-mail:
[email protected], EMBL (submissions: e-mail: [email protected];
enquiries: e-mail: [email protected]) or DDBJ (submissions: e-mail:
[email protected]; enquiries: e-mail: [email protected]).
Submitters without Web or e-mail access should write to one of the
following addresses to obtain a hard copy submission form (GenBank
Submissions, National Center for Biotechnology Information, National
Library of Medicine, 8600 Rockville Pike, Building 38A, Room 8N-805,
Bethesda, MD 20894, USA. EMBL Nucleotide Sequence Submissions,
European Bioinformatics Institute, Hinxton Hall, Hinxton, Cambridge,
CB10 1SD, UK. DNA Data Bank of Japan, Center for Information Biology,
National Institute of Genetics, Mishima, Shizuoka 411-8540, Japan).
Authors are encouraged by the databases to update their entries as the
need arises.

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
110
DNA sequences and GenBank Accession numbers
Many Elsevier journals cite "gene accession numbers" in their running text
and footnotes. Gene accession numbers refer to genes or DNA sequences
about which further information can be found in the databases at the
National Center for Biotechnical Information (NCBI) at the National Library
of Medicine. Elsevier authors wishing to enable other scientists to use the
accession numbers cited in their papers via links to these sources, should
type this information in the following manner:
For each and every accession number cited in an article, authors should
type the accession number in bold, underlined text. Letters in the
accession number should always be capitalised. (See Example 1 below).
This combination of letters and format will enable Elsevier's typesetters to
recognize the relevant texts as accession numbers and add the required
link to GenBank's sequences.
Example 1: "Note:GenBank accession nos. AI631510 , AI631511 ,
AI632198 , and BF223228) , a B-cell tumor from a chronic lymphatic
leukemia (GenBank accession no. BE675048 ), and a T-cell lymphoma
(GenBank accession no. AA361117 )".
Authors are encouraged to check accession numbers used very carefully.
An error in a letter or number can result in a dead link.
In the final version of the printed article , the accession number text will
not appear bold or underlined (see Example 2 below).
Example 2: "Note:GenBank accession nos. AI631510, AI631511,
AI632198, and BF223228), a B-cell tumor from a chronic lymphatic
leukemia (GenBank accession no. BE675048), and a T-cell lymphoma
(GenBank accession no. AA361117)".

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
111
In the final version of the electronic copy , the accession number text will
be linked to the appropriate source in the NCBI databases enabling
readers to go directly to that source from the article (see Example 3
below).
Example 3: "GenBank accession nos. AI631510, AI631511, AI632198, and
BF223228), a B-cell tumor from a chronic lymphatic leukemia (GenBank
accession no. BE675048), and a T-cell lymphoma (GenBank accession no.
AA361117)".
Manuscripts Manuscripts should be in English on numbered pages with
double-spaced typing throughout (including tables, legends and reference
lists) on one side of the paper only with margins of a least 3cm all round.
They should be divided into: (1) title page - include a succinct title (which
should not normally exceed 100 characters and should not contain any
subtitles or abbreviations), the names of all authors, including a given
name for each, the institutions with city, state and country where the
work was performed, the name and complete address (including
telephone, telefax and e-mail) of the corresponding author, a list of
abbreviations and a list of addresses of authors who have moved from the
institutions where the work was performed. (2) abstract - maximum 250
words, (3) keywords (3-6 indexing terms), (4)introduction, (5) materials
and methods, (6) results, (7) discussion, (8) acknowledgements (grant
support and technical support to be listed here), (9) references, (10)
tables and (11) figure legends. A recent issue of the journal should be
consulted for details. In the interests of clarity and brevity, it may
sometimes be advantageous to combine the results and discussion into a
single section. Everyone makes minor modifications to standard methods.
Do not describe standard materials and methods or modifications unless

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
112
they have significant and demonstrable utility. Do not duplicate
descriptions of methodology in the figure legends. Generic and species
names should be typed out in full the first time mentioned - in the title,
the summary and the text - and thereafter the generic name should be
abbreviated. Words or letters to be printed in italics should either be in
italics or underlined. The metric system should be used throughout.
Short communications These are intended for the publication of brief
definitive reports, primarily to complete DNA sequence data, methods,
biochemical or immunochemical data, that do not merit a full-length
publication. Short communications are no more than four pages long
including everything, with maximally two figures, one table and a
maximum of 20 references . A single page contains about 900 words. Only
the salient points of a long DNA sequence should be published, as the
whole sequence will be available for a computer database. The title,
authorship and affiliations will be in the standard format of the journal.
The text should not be sectioned, except for references. Essential
experimental details may be incorporated into a figure legend. To facilitate
rapid publication, authors will be expected to supply high-quality copy and
expedite any necessary revisions, although decisions will normally be yes
or no, based on the quality and appropriateness of the initial submission.
Minireviews. Minireviews are by invitation only. Potential topics of general
current interest should be submitted to the senior editor for consideration.
Reviews should be short, current, specific and potentially provocative.
They should provide a balanced synthesis from the available data rather
than a comprehensive regurgitation of the literature. If possible, they
should provide new concepts and ideas extending across different parasite
systems. Reviews are restricted to about 4000 words, at most three
display items including figures and tables and a list of references of not

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
113
more than 50. The text can be divided into simple subsections with a
succinct abstract. Minireviews will undergo the established review process
at MBP, and will be published by an accelerated schedule if accepted.
References In the text, references should be numbered singly in square
brackets in order of their citation, e.g. [2,3,5-7]. In thelist, references
should be numbered in the order of citation in the text, not in alphabetical
order. Unpublished data, personal communications and papers in
preparation or 'submitted' should not be listed in the references (but may
be incorporated at the appropriate place in the text); work 'in press' may
be listed only if it has been accepted for publication. Personal
communications must be accompanied by a letter from the named
person(s) giving permission to quote such information. Abstracts (whether
published or not), theses and similar material are not to be quoted in the
list. If necessary, they can be referred to in the text in parentheses.
Periodicals [1], books [2] and edited books [3] should accord with the
following examples:
[1] Furuya T, Zhong L, Meyer-Fernandes JR, Lu H, Moreno SNJ, Docampo
R. Ecto-protein tyrosine phosphatase activity in Trypanosoma cruzi
infective stages. Mol Biochem Parasitol 1988;92:339-48.
[2] Sambrook J, Fritsch EF, Maniatis T. Molecular Cloning: a Laboratory
Manual. 2nd ed. Cold Spring Harbor: Cold Spring Harbor Laboratory Press
1989.
[3] Borst P, Bitter W, Blundell PA, et al. The expression sites for variant
surface glycoproteins of Trypanosoma brucei. In: Hide G, Mottram JC,
Coombs GH, Holmes PH, editors. Trypanosomiasis and Leishmaniasis:
Biology and Control. Oxford: CAB International, 1997;7:109-31.

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
114
Abbreviations of journal titles should conform to those adopted by the List
of Serial Title Word abbreviations, ISDS International Centre, 20, rue
Bachaumont, 75002 Paris, France (ISBN 2-904938-02-8).
Tables Each table should be typed double-spaced on a separate sheet and
have a short descriptive title. A legend may be placed under table.
Footnotes should be identified in the table by a, b, c, etc.
Figures Figures must be in a form and condition suitable for high quality
reproduction. Lettering should be clear and of adequate size to be legible
after reduction. Consider the printed page and column proportions when
preparing figures. If figures are not to be reduced their format should not
exceed 16 x 20 cm. Multiple panels of a single figure must be mounted
together. Each DNA sequence figure must fit on a single sheet of paper.
Place numbering at one end of each line, not on separate lines, and avoid
excessive line spacing. Consider placing nucleotide and protein data in
separate panels, using single-letter amino acid abbreviations for the
protein sequence and grouping nucleotides either continuously or in blocks
of ten separated by one space (90 to 120 nt per line). Over 10 000 bp can
legibly fit on each journal page in this format (see, e.g., Mol. Biochem.
Parasitol. 95:141-146). Preferably use a sans-serif font. Upper case is
standard, except that introns or other features can be usefully
distinguished by lower case. Provide sharp laser-printer or imagesetter
copy. Nucleotide sequences of long coding regions, where the amino acid
sequence is the primary feature, and long DNA sequences, may, at the
editor's discretion, be omitted from the printed paper. They can be
obtained from electronic databases or from the authors. Half-tone
illustrations may be included. They should be submitted as black-and-
white prints on glossy paper and have as much contrast as possible. A

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
115
scale should appear on photomicrographs. Colour plates will be published
free of charge if colour contributes to the understanding of the
information. In all other cases, the author should be prepared to pay the
extra costs of 635 EUR for the first page and 318 EUR for following pages
of colour. Figures legends should be typed double spaced at the end of the
text, not on the figures. Figures should be checked extremely carefully,
particularly after revisions. No changes to figures will be possible after
acceptance of the manuscript.
Detailed instructions Abbreviations, symbols, chemical and biochemical
nomenclature, etc., should follow the recommendations given in the
Journal of Biological Chemistry (Vol. 272, pp. 28165-28170;
http://www.jbc.org). Avoid abbreviations which are not in common use
across the field of molecular and biochemical parasitology. Those used
should be defined in the text on first usage and listed as a footnote on the
title page. Do not introduce abbreviations unless they are used at least 4
times.
Genetic nomenclature for Trypanosoma and Leishmania should follow the
guidelines proposed by Clayton et al (1998), Mol Biochem Parasitol
1998;97:221-224
(http://www.elsevier.nl/cas/tree/store/molbio/free/1998/97/12/3178.pdf)
Author enquiries For enquiries relating to the submission of articles please
visit Elsevier's Author Gateway at http://authors.elsevier.com . The
Author Gateway also provides the facility to track accepted articles and set
up email alerts to inform you of when an article's status has changed, as
well as detailed artwork guidelines, copyright information, frequently
asked questions and more.

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
116
Contact details for questions arising after acceptance of an article,
especially those related to proofs, are provided after registration of an
article for publication.
ProofreadingOne set of page proofs in PDF format will be sent by e-mail to
the corresponding Author. (If we do not have an e-mail address then
paper proofs will be sent by post). Elsevier now sends PDF proofs which
can be annotated; for this you will need to download Adobe Reader
version 7 available free from
http://www.adobe.com/products/acrobat/readstep2.html. Instructions on
how to annotate PDF files will accompany the proofs. If you do not wish to
use the PDF annotations function, you may list the corrections (including
replies to the Query Form) and return to Elsevier in an e-mail. Please list
your corrections quoting line number. If, for any reason, this is not
possible, then mark the corrections and any other comments (including
replies to the Query Form) on a printout of your proof and return by fax,
or scan the pages and e-mail, or send by post. Proofs should be read
carefully and returned within 2 days of receipt. Please use this proof only
for checking the typesetting, editing, completeness and correctness of the
text, tables and figures. Significant changes to the article as accepted for
publication will only be considered at this stage with permission from the
Editor. We will do everything possible to get your article published quickly
and accurately. Therefore, it is important to ensure that all of your
corrections are sent back to us in one communication: please check
carefully before replying, as inclusion of any subsequent corrections
cannot be guaranteed. Proofreading is solely your responsibility. Note that
Elsevier may proceed with the publication of your article if no response is
received.
Language Polishing For authors who require information about language
editing and copyediting services pre- and post-submission, please visit

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
117
http://www.elsevier.com/wps/find/authorshome.authors/languagepolishin
g or contact [email protected] for more information. Please
note that Elsevier neither endorses nor takes responsibility for any
products, gooods or services offered by outside vendors through our
services or in any advertising. For more information, please refer to our
terms and conditions
http://www.elsevier.com/wps/find/termsconditions.cws_home/termscondi
tions.
US National Institutes of Health (NIH) voluntary posting/ "Public Access
Policy"
Elsevier facilitates author posting in connection with the voluntary posting
request of the NIH (referred to as the NIH "Public Access Policy"; see
http://publicaccess.nih.gov/) by submitting the peer-reviewed author's
manuscript directly to PubMed Central on request from the author,
immediately after formal publication. Please e-mail us at
[email protected]) that your work has received NIH funding
(with the NIH grant/project number(s), as well as the name and e-mail
address of the Principal Investigator(s)) and that you intend to respond to
the NIH request. Upon such confirmation, Elsevier will submit to PubMed
Central on your behalf a version of your manuscript that will include peer-
review comments, for public access posting 12 months after the final
publication date. This will ensure that you will have responded fully to the
NIH request policy. There will be no need for you to post your manuscript
directly to PubMed Central, and any such posting is prohibited (although
Elsevier will not request that manuscripts authored and posted by US
government employees should be taken down from PubMed Central).
Individual modifications to this general policy may apply to some Elsevier
journals and its society publishing partners.

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
118
Editors
C.E. Clayton Zentum für Molekulare Biologie, Universität Heidelberg, Im
Neuerheimer Feld 282, D-69120 Heidelberg, Germany Tel: +49 6221
546876 Fax: +49 6221 545894 E-mail:[email protected]
A.P. Waters Department of Parasitology, L4-Q, Leiden University Medical
Center, Albinusdreef 2, 2333 ZA Leiden, The Netherlands. Tel.: +31 71
5265069; Fax +31 71 52668907; e-mail:[email protected]
P.T. LoVerde, Southwest Foundation for Biomedical Research, PO BOX
760549, USA; e-mail: [email protected]
B. Ullman Department of Biochemistry, Oregon Health Science University,
3181 SW Sam Jackson Park Road, Portland OR 97201, USA Tel: +1 503
494 8437 Fax: +1 503 494 8393 E-mail:[email protected]
Reviews Editor
Alister Craig, Liverpool School of Tropical Medicine, Pembroke Place,
Liverpool L3 5QA, UK. Tel: +44 151 705 3161; Fax: +44 151 705 3371;
email:[email protected]
Sponsored Articles:
Molecular and Biochemical Parasitology offers authors or their institutions
the option to sponsor non-subscriber access to their articles on Elsevier's
electronic publishing platforms.

Pereira, M.M.C. 2008 Análise da expressão e investigação de mecanismos...
119
Pereira, Mariana Marques Coutelo
Análise da expressão e investigação de mecanismos
envolvidos com o controle da atividade de homólogos do
fator de iniciação da tradução eIF4E ao longo do ciclo de
vida de Leismania amazonensis. / Mariana Marques Coutelo
Pereira. – Recife: A Autora, 2008.
118 fls. .: il.
Dissertação (Mestrado em Genética) – UFPE. CCB
1. Leishmania amazonensis 2. Fosforilação 3. Protozoa
I.Título
593.1 CDU (2ª. Ed.) UFPE
561.99 CDD (22ª. Ed.) CCB – 2008 – 124






![Clave de Cotización: CADU Fecha: 2017-12-31 [411000-AR ...cdn.investorcloud.net/cadu/InformacionFinanciera/... · Clave de Cotización: CADU Fecha: 2017-12-31 1 de 132 [411000-AR]](https://static.fdocument.pub/doc/165x107/6006959ea83e8f036a41a1c8/clave-de-cotizacin-cadu-fecha-2017-12-31-411000-ar-cdn-clave-de-cotizacin.jpg)




![Clave de Cotización: CADU Fecha: 2019-12-31 [411000-AR] Datos …cdn.investorcloud.net/cadu/InformacionFinanciera/Reporte... · 2020-07-02 · Clave de Cotización: CADU Fecha: 2019-12-31](https://static.fdocument.pub/doc/165x107/5f297480deedb36ecb588823/clave-de-cotizacin-cadu-fecha-2019-12-31-411000-ar-datos-cdn-2020-07-02.jpg)






