
POST. MIKROBIOL., PROKARIOTY REDUKUJ¥CE Fe(III): … · 2017-03-02 · sz„ odkryt„ bakteri„...
20
1. Wprowadzenie flelazo (Fe) jest dominuj„cym pierwiastkiem naszej Planety, uwa¿a siŒ, ¿e stanowi 45% jej masy. Jest g‡ów- nym sk‡adnikiem (ok. 90%) j„dra Ziemii, gdzie wystŒ- puje w stanie rodzimym. Natomiast w skorupie ziem- skiej, której gruboæ wynosi 570 km (rednio 45 km), zawartoæ ¿elaza szacuje siŒ na ok. 5% i jest ono czwartym z kolei pierwiastkiem, po tlenie (O), krze- mie (Si) i glinie (Al). W skorupie ziemskiej ¿elazo wystŒpuje g‡ównie na +2 lub +3 stopniu utlenienia w postaci ró¿nych minera‡ów (rud ¿elaza) przedsta- wionych w Tabeli I. flelazo wystŒpuje równie¿ w nie- wielkich ilociach na stopniu utlenienia 0, w stanie ro- dzimym w ska‡ach magmowych oraz meteorytach ¿elaznych. flelazo jest bardzo reaktywne i jego forma Fe(II) jon ¿elazawy w rodowisku neutralnym i wa- runkach tlenowych ‡atwo ulega utlenieniu do formy Fe(III) jon ¿elazowy [20]. flelazo (III) mo¿e ulegaæ mikrobiologicznej redukcji do formy dwuwartocio- wej w wyniku dwóch procesów: (i) redukcji asymila- cyjnej zwi„zanej z przyswajaniem ¿elaza i wbudowy- waniem go w enzymy, kofaktory [20] oraz (ii) redukcji dysymilacyjnej zwi„zanej z wykorzystaniem ¿elaza Fe(III) jako ostatecznego akceptora elektronów w pro- cesie oddychania w warunkach beztlenowych. Taki typ oddychania nazywany oddychaniem ¿elazowym lub oddychaniem Fe(III) (ferric ion respiration lub Fe(III) respiration) co oznacza, ¿e ca‡y proces zwi„- zany jest z produkcj„ energii (61). Nie ka¿dy proces POST. MIKROBIOL., 2009, 48, 2, 105124 http://www.pm.microbiology.pl PROKARIOTY REDUKUJ¥CE Fe(III): KLASYFIKACJA, WYST˚POWANIE, MECHANIZMY REDUKCJI Fe(III), ROLA EKOLOGICZNA I ZNACZENIE BIOTECHNOLOGICZNE Anna Sikora 1 * i Mieczys‡aw K. B‡aszczyk 2 1 Zak‡ad Biologii Molekularnej, Instytut Biochemii i Biofizyki PAN, ul. Pawiæskiego 5a, 02-106 Warszawa 2 Samodzielny Zak‡ad Biologii Mikroorganizmów, Szko‡a G‡ówna Gospodarstwa Wiejskiego, ul. Nowoursynowska 159, 02-776 Warszawa Wp‡ynŒ‡o w styczniu 2009 r. 1. Wprowadzenie. 2. Charakterystyka fizjologiczno-biochemiczna grup mikroorganizów redukuj„cych Fe(III). 3. Mechanizmy re- dukcji Fe(III). 4. Rola ekologiczna prokariotów redukuj„cych Fe(III) w rodowisku. 5. Znaczenie prokariotów redukuj„cych Fe(III) w biotechnologii. 5.1. Mikrobiologiczne ogniwa paliwowe. 5.2. Bioremediacja. 6. Podsumowanie Fe(III)-reducing prokaryotes: classification, distribution, mechanisms of Fe(III) reduction, ecological role and biotechnological significance Abstract: FRM, ferric ion-respiring microorganisms, are a group of prokaryotes that use iron (III) and other metals as terminal electron acceptors in the process of anaerobic respiration. The list of culturable species of FRM is relatively short and the species belong to different phylogenetic groups. The article presents a physiological and biochemical characteristics of FRM, their eco- logical role and contribution to the circulation of elements in nature. Special attention is paid to the biotechnological significance of FRM, because of their potential role in (i) bioremediation of anaerobic environments contaminated by organic compounds and toxic heavy metals or radionuclides; (ii) electricity production in microbial fuel cells (MFC). The mechanisms of Fe(III) reduction are also described. 1. Introduction. 2. Physiological and biochemical characteristics of ferric respiring microorganisms. 3. Mechanisms of Fe(III) reduc- tion. 4. Ecological role of FRM. 5. Biotechnological significance of FRM. 5.1. Microbial fuel cells. 5.2. Bioremediation. 6. Summary S‡owa kluczowe: prokarioty redukuj„ce Fe(III), mechanizmy redukcji Fe(III), mikrobiologiczne ogniwa paliwowe, bioremediacja Key words: Fe(III)-respiring microorganisms (FRM), mechanisms of Fe(III) reduction, microbial fuel cells, bioremediation * Autor do korespondencji: tel. (22) 592 3337, e-mail:[email protected] Magnetyt Fe 3 O 4 lub FeO×Fe 2 O 3 Hematyt Fe 2 O 3 Limonit Fe 2 O 3 × nH 2 O lub FeOOH Syderyt FeCO 3 Getyt Fe 2 O 3 × H 2 O Ilmenit FeO × TiO 2 Wiwianit [Fe 3 (PO 4 ) × 8H 2 O] Piryt (markasyt) FeS 2 Pirotyn Fe n S n+1 (n=56) Arsenopiryt FeAsS Tabela I Minera‡y (rudy) ¿elaza wystŒpuj„ce w skorupie ziemskiej [20, 61, 62] Ruda ¿elaza Wzór chemiczny
Transcript of POST. MIKROBIOL., PROKARIOTY REDUKUJ¥CE Fe(III): … · 2017-03-02 · sz„ odkryt„ bakteri„...

1. Wprowadzenie
¯elazo (Fe) jest dominuj¹cym pierwiastkiem naszejPlanety, uwa¿a siê, ¿e stanowi 45% jej masy. Jest g³ów-nym sk³adnikiem (ok. 90%) j¹dra Ziemii, gdzie wystê-puje w stanie rodzimym. Natomiast w skorupie ziem-skiej, której grubo�æ wynosi 5�70 km (�rednio 45 km),zawarto�æ ¿elaza szacuje siê na ok. 5% i jest onoczwartym z kolei pierwiastkiem, po tlenie (O), krze-mie (Si) i glinie (Al). W skorupie ziemskiej ¿elazowystêpuje g³ównie na +2 lub +3 stopniu utlenieniaw postaci ró¿nych minera³ów (rud ¿elaza) przedsta-wionych w Tabeli I. ¯elazo wystêpuje równie¿ w nie-wielkich ilo�ciach na stopniu utlenienia 0, w stanie ro-dzimym w ska³ach magmowych oraz meteorytach¿elaznych. ¯elazo jest bardzo reaktywne i jego formaFe(II) � jon ¿elazawy w �rodowisku neutralnym i wa-runkach tlenowych ³atwo ulega utlenieniu do formyFe(III) � jon ¿elazowy [20]. ¯elazo (III) mo¿e ulegaæmikrobiologicznej redukcji do formy dwuwarto�cio-wej w wyniku dwóch procesów: (i) redukcji asymila-cyjnej zwi¹zanej z przyswajaniem ¿elaza i wbudowy-waniem go w enzymy, kofaktory [20] oraz (ii) redukcji
dysymilacyjnej zwi¹zanej z wykorzystaniem ¿elazaFe(III) jako ostatecznego akceptora elektronów w pro-cesie oddychania w warunkach beztlenowych. Takityp oddychania nazywany oddychaniem ¿elazowymlub oddychaniem Fe(III) (ferric ion respiration lubFe(III) respiration) co oznacza, ¿e ca³y proces zwi¹-zany jest z produkcj¹ energii (61). Nie ka¿dy proces
POST. MIKROBIOL.,2009, 48, 2, 105�124http://www.pm.microbiology.pl
PROKARIOTY REDUKUJ¥CE Fe(III): KLASYFIKACJA,WYSTÊPOWANIE, MECHANIZMY REDUKCJI Fe(III),
ROLA EKOLOGICZNA I ZNACZENIE BIOTECHNOLOGICZNE
Anna Sikora1* i Mieczys³aw K. B³aszczyk2
1 Zak³ad Biologii Molekularnej, Instytut Biochemii i Biofizyki PAN, ul. Pawiñskiego 5a, 02-106 Warszawa2 Samodzielny Zak³ad Biologii Mikroorganizmów, Szko³a G³ówna Gospodarstwa Wiejskiego,
ul. Nowoursynowska 159, 02-776 Warszawa
Wp³ynê³o w styczniu 2009 r.
1. Wprowadzenie. 2. Charakterystyka fizjologiczno-biochemiczna grup mikroorganizów redukuj¹cych Fe(III). 3. Mechanizmy re-dukcji Fe(III). 4. Rola ekologiczna prokariotów redukuj¹cych Fe(III) w �rodowisku. 5. Znaczenie prokariotów redukuj¹cych Fe(III)w biotechnologii. 5.1. Mikrobiologiczne ogniwa paliwowe. 5.2. Bioremediacja. 6. Podsumowanie
Fe(III)-reducing prokaryotes:classification, distribution, mechanisms of Fe(III) reduction, ecological role and biotechnological significance
Abstract: FRM, ferric ion-respiring microorganisms, are a group of prokaryotes that use iron (III) and other metals as terminalelectron acceptors in the process of anaerobic respiration. The list of culturable species of FRM is relatively short and the speciesbelong to different phylogenetic groups. The article presents a physiological and biochemical characteristics of FRM, their eco-logical role and contribution to the circulation of elements in nature. Special attention is paid to the biotechnological significanceof FRM, because of their potential role in (i) bioremediation of anaerobic environments contaminated by organic compounds andtoxic heavy metals or radionuclides; (ii) electricity production in microbial fuel cells (MFC). The mechanisms of Fe(III) reductionare also described.
1. Introduction. 2. Physiological and biochemical characteristics of ferric respiring microorganisms. 3. Mechanisms of Fe(III) reduc-tion. 4. Ecological role of FRM. 5. Biotechnological significance of FRM. 5.1. Microbial fuel cells. 5.2. Bioremediation. 6. Summary
S³owa kluczowe: prokarioty redukuj¹ce Fe(III), mechanizmy redukcji Fe(III), mikrobiologiczne ogniwa paliwowe, bioremediacjaKey words: Fe(III)-respiring microorganisms (FRM), mechanisms of Fe(III) reduction, microbial fuel cells, bioremediation
* Autor do korespondencji: tel. (22) 592 3337, e-mail:[email protected]
Magnetyt Fe3O
4 lub FeO×Fe
2O
3
Hematyt Fe2O
3
Limonit Fe2O
3 × nH
2O lub FeOOH
Syderyt FeCO3
Getyt Fe2O
3 × H
2O
Ilmenit FeO × TiO2
Wiwianit [Fe3(PO
4) × 8H
2O]
Piryt (markasyt) FeS2
Pirotyn FenS
n+1 (n=5�6)
Arsenopiryt FeAsS
Tabela IMinera³y (rudy) ¿elaza wystêpuj¹ce w skorupie ziemskiej
[20, 61, 62]
Ruda ¿elaza Wzór chemiczny

106 ANNA SIKORA I MIECZYS£AW K. B£ASZCZYK
Acidobacteria Geothrix G. fermentans
Actinobacteria Acidimicrobium Am. ferrooxidans
Aquificae Sulfurihydrogenibium S. azorense, S. subterraneum
DeferribacteresDeferribacter D. abyssi, D. thermophilus
Geovibrio G. ferrireducens
Deinococcus-ThermusDeinococcus D. geothermalis
Thermus T. scotoductus
Alkaliphilus A. metalliredigens
Anaerobranca A. californensis, A. gottschalkii, A. horicoshi
Bacillus B. infernus, B. subterraneus
Carboxydothermus C. ferrireducens
Firmicutes Sulfobacillus S. acidophilus, S. thermosulfidooxidans
Thermoanaerobacter T. acetoethylicus, T. brokii, T. siderophilus, T. sulfurophilus, T. wiegelii
Thermolithobacter T. ferrireducens, T. carboxydivorans
Termoterrabacterium T. ferrireducens
Thermosinus T. carboxydovorans
Thermovenabulum T. ferrioganovorum
Nitrospira Magnetobacterium M. magnetitacticum
"-Proteobacteria Acidophilium A. cryptum
Ferribacter F. limneticum, F. thermoautotrophicum
$-Proteobacteria Ferribacterium F. limneticum
Rhodoferax R. ferrireducens
Acidithiobacillus A. ferrooxidans
Aeromonas A. hydrophila
(-Proteobacteria Ferrimonas F. balearica
Pantoea P. agglomerans
Shewanella S. alga, S. baltica, S. frigidimarina, S. oneidensis, S. olleyana,S. paeleana, S. potomacii, S. putrefaciens, S. saccharophila
Anaeromyxobacter A. dehalogenans
Desulfobulbus D. propionicus
Desulfotomaculum D. reducens
Desulfitobacterium D. frappieri, D. hafniense, D. metallireducens
Desulfuromonas D. acetexigenes, D. acetoxidans, D. chloroethenica, D. michiganensis,D. palmitatis, D. thiophila, D. svalbardensis
Desulfuromusa D. bakii, D. kysingii, D. ferrireducens, D. succinoxidans
Geoalkalibacter G. ferrihydriticus
*-Proteobacteria Geobacter G. bemidjiensis, G. bremensis, G. chapelleii, G. grbiciae,G. humireducens, G. hydrogenophilus, G. metallireducens,G. pelophilus, G. psychrophilus, G. pickeringii, G. sulfurreducens,G. thiogenes, G. uraniireducens
Geopsychrobacter G. electrodiphilus, G. multivorans
Geothermobacter G. ehrlichii
Malonomonas M. rubra
Pelobacter P. acetylenicus, P. acidigalici, P. carbinolicus, P. massiliensis,P. propionicus, P. venetianus
g-Proteobacteria Sulfurispirillum S. barnessi
ThermodesulfobacteriaGeothermobacterium G. ferrireducens
Thermodesulfobacterium T. commune
Thermotoga Thermotoga T. lettingae, T. maritima, T. subterranea
T a b e l a I ILista mikroorganizmów redukuj¹cych ¿elazo (III) z domeny Bacteria zdolnych do oddychania ¿elazowego [5, 8, 9, 13, 15, 16, 20,
24, 3, 34, 35, 37, 38, 45, 48, 61, 62, 66, 67, 82, 83, 91, 95, 98, 100]
Typ/klasa RodzajGatunki przyk³adowe

107PROKARIOTY REDUKUJ¥CE FE(III): KLASYFIKACJA, WYSTÊPOWANIE, MECHANIZMY REDUKCJI FE(III)...
dysymilacyjnej redukcji ¿elaza zwi¹zany jest z wytwa-rzaniem energii. Ma to miejsce wtedy, gdy redukcja¿elaza towarzyszy innym typom oddychania np. fer-mentacji, czy redukcji siarczanów a ¿elazo Fe(III) sta-nowi dodatkowy akceptor elektronów. Uwa¿a siê, ¿ejest to sposób na pozbycie siê nadmiaru elektronówi protonów z komórki (si³a redukcyjna), przy niedo-statku zasadniczego akceptora [20, 62].
2. Charakterystyka fizjologiczno-biochemicznagrup mikroorganizmów redukuj¹cych Fe(III)
Dla okre�lenia prokariotów wykorzystuj¹cych Fe(III)jako ostateczny akceptor elektronów w oddychaniubeztlenowym stosuje siê skrót FRM (Fe(III)-respiringmicroorganisms). Oprócz Fe(III) redukuj¹ one Mn(IV)do Mn(II) oraz kwasy humusowe a tak¿e inne metale,pierwiastki promieniotwórcze, np. U(VI) i zwi¹zki jakazotany, fumaran czy siarkê S0. FRM zdolne do oddy-chania Fe(III) nie tworz¹ specyficznej filogenetycznejgrupy. Nale¿¹ one do obu domen prokariotów: Bacte-ria i Archaea. Obecnie znanych jest znacznie ponad100 gatunków FRM. Ponad po³owa gatunków stale re-zyduje w �rodowiskach psychro-mezofilnych (choæ nies¹ psychrofilami, ale mog¹ byæ psychrotolerantami),reszta zajmuje �rodowiska termofilne i hypertermofil-ne (�ród³a gor¹ce, wody geotermalne, kominy hydro-termalne). Jedna grupa FRM ca³kowicie utlenianiamateriê organiczn¹ do dwutlenku wêgla, druga nie-kompletnie, zwykle do octanu. Liczne FRM, zw³asz-cza hypertermofilne, wykorzystuj¹ wodór jako �ród³oelektronów do redukcji Fe(III). W naturalnych �rodo-wiskach prokarioty FRM w warunkach beztlenowychutleniaj¹ produkty rozk³adu materii organicznej, g³ów-nie produkty fermentacji glikolitycznych i nieglikoli-tycznych. W zwi¹zku z tym, ¿e wodór i octan s¹ dwo-ma g³ównymi po�rednimi produktami fermentacjimaterii organicznej to uwa¿a siê, ¿e w naturalnych �ro-dowiskach o pH obojêtnym bogatych w zwi¹zki Fe(III)czy Mn(IV) stanowi¹ one istotne �ród³o elektronóww procesach redukcji metali. Dlatego przy charakterys-tyce FRM w badaniach laboratoryjnych zawsze okre�lasiê ich zdolno�æ do utleniania octanu i wodoru. Octanstosuje siê jako �ród³o wêgla i energii do selekcji pro-kariotów redukuj¹cych Fe(III). Utlenianie octanu prze-biega zgodnie z reakcj¹ 1, a wodoru zgodnie z reak-cj¹ 2 [61, 62, 67, 95]:
CH3COO� (octan) + 8Fe(III) ++ 4H2O → 2HCO3
� + 8Fe(II) + 9H+ (reakcja 1)H2 + 2Fe(III) → 2H+ + 2Fe(II) (reakcja 2)
Tabela II pokazuje listê prokariotów FRM z dome-ny Bacteria a tabela III z domeny Archaea zdolnychdo dysymilacyjnej redukcji ¿elaza, z wytworzeniem
energii w procesie oddychania ¿elazowego. Najlepiejznanymi i zbadanymi z FRM [5, 20, 34, 35, 38, 48,61, 62, 66, 67, 91, 98] jest rodzina Geobacteraceaeoraz rodzina Desulfuromonadaceae, obie z klasy *-Pro-teobacteria, typu Proteobacteria. Wg Bergey Manual[5] do Geobacteraceae nale¿y rodzaj Geobacter, któryjest jak dotychczas stosunkowo dobrze zbadany. Pierw-sz¹ odkryt¹ bakteri¹ z rodzaju Geobacter by³ G. metalli-reducens (pocz¹tkowo oznaczony jako szczep GS-15),wyizolowany z osadu rzeki Potomac w USA w 1987 ro-ku. Do Geobacteraceae nale¿¹ gatunki bakterii mezo-filnych, rosn¹ce w zakresie temperatur 25�40°C a tak¿epsychrotolerancyjny Geopsychrobacter electrodiphilus,rosn¹cy w zakresie temperatur 4�30°C oraz termofilnyGeothermobacter ehrlichii, wyizolowany z hydroter-malnego �ród³a, rosn¹cy w temperaturach 35�65°Cprzy optimum 55°C. W�ród Geobacteraceae s¹ te¿inne bakterie ekstremofilne, np. Geoalkalibacter ferri-hydriticus, bakteria alkalofilna o optymalnym pH = 8,6.Bakterie z rodziny Geobacteraceae wystêpuj¹ w po-wierzchniowych warstwach osadów dennych ró¿no-rodnych zbiorników wodnych p³ytkich i g³êbokich,zw³aszcza piaszczystych, bogatych w zwi¹zki Fe(III).Bakterie te wystêpuj¹ szczególnie licznie w miejscachska¿onych rop¹ naftow¹. Izolacja chromosomalnegoDNA i amplifikacja na jego matrycy genów koduj¹-cych 16SrRNA wykaza³a, ¿e w wymienionych �rodo-wiskach jest to dominuj¹ca grupa bakterii. Wg BergeyManual [5] do nastêpnej do�æ dobrze poznanej rodzinyDesulfuromonadaceae nale¿¹ gatunki z rodzaju Desul-furomonas, Desulfuromusa, Malonomonas i Pelo-bacter, wystêpuj¹ce w osadach morskich. PsychrofilnyDesulfuromonas svalbardensis i psychrotolerancyjnyDesulfuromusa ferrireducens by³y izolowane z mor-skich osadów dennych Arktyki, z optymaln¹ tempera-tur¹ wzrostu odpowiednio 14°C i 14�17°C i maksy-maln¹ 20°C i 23°C.
Archeoglobus A. fulgidus
FerroglobusF. indicus, F. placidus,
Euryarchaeota F. pacificus
Geoglobus G. ahangari
Pyrococcus P. furiosus
Thermococcus T. celer, T. sibiricus
Geogemma G. barosi, G. indica,G. pacifica
Crenarchaeota Hyperthermus H. butylicus
Pyrobaculum P. aerophilum, P. islandicum
Pyrodictium P. abyssi, P. occultum
Sulfolobus S. acidocaldarius, S. shibatae
T a b e l a I I ILista mikroorganizmów redukuj¹cych ¿elazo (III) z domenyArchaea zdolnych do oddychania ¿elazowego [5, 39, 40, 95]
Typ Rodzaj Gatunki przyk³adowe

108 ANNA SIKORA I MIECZYS£AW K. B£ASZCZYK
Oprócz rodzaju Pelobacter, pozosta³e bakterie nale-¿¹ce do dwóch powy¿szych rodzin s¹ zdolne do ca³ko-witego utleniania octanu do dwutlenku wêgla z jedno-czesn¹ redukcj¹ Fe(III). Wiêkszo�æ z nich ca³kowicieutlenia kwasy organiczne, zwi¹zki aromatyczne (np.toluen, benzoesan, fenol, krezol), niektóre etanol, d³u-go³añcuchowe kwasy t³uszczowe, cukry oraz amino-kwasy. Du¿a czê�æ wykorzystuje wodór jako dawcêelektronów do procesów redukcji. Reakcje 3�6 przed-stawiaj¹ kolejno utlenianie benzoesanu, toluenu, fenolui krezolu z jednoczesn¹ redukcj¹ Fe(III):
C6H5COO� (benzoesan) + 30Fe(III) ++ 19H2O → 7HCO3
� + 30Fe(II) + 36H+ (reakcja 3)C6H5CH3 (toluen) + 36Fe(III) ++ 21H2O → 7HCO3
� + 36Fe(II) + 43H+ (reakcja 4)C6H5OH (fenol) + 28Fe(III) ++ 17H2O → 6HCO3
� + 28Fe(II) + 34H+ (reakcja 5)C7H8O (krezol) + 34Fe(III) ++ 20H2O → 7HCO3
� + 34Fe(II) + 41H+ (reakcja 6)
Przed odkryciem zdolno�ci do redukcji Fe(III) uwa-¿ano, ¿e bakterie z rodzaju Pelobacter uzyskuj¹ energiêwy³¹cznie w wyniku procesów fermentacji. Fermentuj¹one 2,3-butanediol, acetoinê, glikol etylenowy i poli-etylenowy, acetylen oraz kwas galusowy. RodzajDesulfuromusa oraz Malonomonas rubra prowadz¹fermentacjê jab³czanu i fumaranu. Czê�æ gatunków De-sulfuromonas, Desulfuromusa, Geoalkalibacter ferri-hydriticus, Pelobacter jest �ci�le beztlenowa, inne s¹zdolne tak¿e do oddychania tlenowego (Geobacter).Odkrycie, ¿e G. sulfurreducens prze¿ywa w warunkachatmosferycznych oraz jest zdolny do wzrostu przyobni¿onym stê¿eniu tlenu w stosunku do powietrzat³umaczy fakt, ¿e rodzaj Geobacter dominuje w �ro-dowiskach beztlenowych, które czasami mog¹ ulegaænatlenieniu. Rodzaje Desulfuromonas, Desulfuromusa,Malonomonas, Pelobacter oraz G. sulfurreducens mog¹redukowaæ tak¿e S0.
Kolejne FRM, które s¹ intensywnie badane i do�ædobrze poznane to gatunki z rodzaju Shewanella, z ro-dziny Shewanellaceae, klasy *-Proteobacteria, typuProteobacteria wyizolowane z osadów dennych i warstwpodpowierzchniowych ró¿norodnych zbiorników wod-nych oraz gleb. Nale¿¹ tu bakterie s³odkowodne i mor-skie. S¹ równie¿ szczepy patogenne. W �rodowiskachbeztlenowych, gdzie redukowane jest ¿elazo, stanowi¹one zaledwie do 2% ogólnej ilo�ci bakterii, a nawetnie wykrywa siê ich tam wcale. Prawdopodobn¹ przy-czyn¹ niewielkiej ilo�ci bakterii z rodzaju Shewanellaw ogólnej puli FRM jest prawdopodobnie niewielkailo�æ dostêpnych substratów w �rodowisku oraz fakt,¿e prowadz¹ one niepe³ne utlenianie bardziej z³o¿o-nych substratów. Jako �ród³o elektronów do redukcjiFe(III) bakterie te wykorzystuj¹ wodór oraz zwi¹zkiorganiczne jak kwasy organiczne (mleczan, pirogro-
nian, mrówczan) i wêglowodany (glukoza). S¹ to fakul-tatywne beztlenowce; ruchliwe Gram-ujemne pa³eczki,które posiadaj¹ wici [5, 61, 62, 67]. Utlenianie mlecza-nu, pirogronianu i mrówczanu z jednoczesn¹ redukcj¹Fe(III) przebiega odpowiednio wed³ug reakcji 7�9.
C2H4OHCOO� (mleczan) + 4Fe(III) ++ 2H2O → CH3COO� (octan) + HCO3
�
+ 4Fe(II) + 5H+ (reakcja 7)CH3COCOO� (pirogronian)+ 2Fe(III) + 2H2O → CH3COO� (octan)+ HCO3
� + 2Fe(II) + 3H+ (reakcja 8)HCOO� (mrówczan) + 2Fe(III)+ H2O → HCO3
� + 2Fe(II) + 2H+ (reakcja 9)
Fakultatywne beztlenowce z rodzaju Ferrimonasi Aeromonas to kolejne bakterie z klasy (-Proteobac-teria redukuj¹ce Fe(III) z jednoczesnym utlenianiemzwi¹zków organicznych [20, 83].
W�ród FRM zidentyfikowano pojedyncze gatunki,filogenetycznie odleg³e od siebie oraz od pozosta³ychznanych redukuj¹cych Fe(III). Geothrix fermentansto Gram-ujemna nieruchliwa pa³eczka, wyizolowanaze zbiornika wodnego zaneczyszczonego rop¹ naf-tow¹. Jest to �cis³y beztlenowiec zdolny do redukcjiFe(III) z jednoczesnym utlenianiem propionianu, mle-czanu, fumaranu, bursztynianu oraz d³ugo³añcucho-wych kwasów t³uszczowych jak kwas palmitynowy.Energiê mo¿e uzyskiwaæ tak¿e w wyniku fermentacjicytrynianu czy fumaranu. Jest gatunkiem nale¿¹cymdo typu Acidobacteria [8].
Geovibrio ferrireducens zosta³ wyizolowany z te-renów ska¿onych wêglowodorami. Jest to ruchli-wy Gram-ujemny przecinkowiec, bakteria mezofilna,�ci�le beztlenowa, niefermentuj¹ca. Elektrony do re-dukcji Fe(III) uzyskuje w wyniku utleniania octanu,wodoru, mleczanu, propionianu, bursztynianu, fumara-nu, pirogronianu oraz, co jest rzadkie, aminokwasów.Reakcja utleniania proliny w po³¹czeniu z redukcj¹¿elaza przebiega zgodnie z reakcj¹ 10:
C5H9O2N (prolina) + 8H2O + 22 [Fe3+]→ 5CO2 + NH4+ + 21H+ + 22 [Fe2+] (reakcja 10)
G. ferrireducens potrafi te¿ redukowaæ siarkê S0.Najwiêksze podobieñstwo wykazuje do halofilnej,beztlenowej, heterotroficznej pa³eczki Flexistipes sinus-arabici, z któr¹ tworzy odrêbn¹ grupê filogenetyczn¹w obrêbie domeny Bacteria [13].
Ferribacterium limneticum jest ruchliw¹ pa³ecz-k¹ nale¿¹c¹ do klasy $-Proteobacteria, wyizolowan¹z osadów dennych jeziora zanieczyszczonego odpada-mi kopalnianymi. Jest to bakteria �ci�le beztlenowa,redukuje Fe(III) utleniaj¹c octan i inne kwasy orga-niczne, niezdolna do fermentacji. Najbardziej spo-krewniona jest z bakteri¹ fotosyntetyzuj¹c¹ Rhodocyc-lus tenuis, która nie jest heterotrofem i nie jest zdolna

109PROKARIOTY REDUKUJ¥CE FE(III): KLASYFIKACJA, WYSTÊPOWANIE, MECHANIZMY REDUKCJI FE(III)...
do redukcji Fe(III), ale F. limneticum nie jest bakteri¹fotosyntetyzuj¹c¹ [16].
Rhodoferax ferrireducens jest krótk¹ ruchliw¹Gram-ujemn¹ pa³eczk¹ z pojedyncz¹ wici¹, z klasy$-Proteobacteria, wyizolowan¹ z osadów dennychzatoki Oyster Bay w stanie Wirginia w USA, rosn¹c¹w temperaturze 4�30°C. Jest fakultatywnym beztle-nowcem wykazuj¹cym unikaln¹ zdolno�æ ca³kowitegoutleniania glukozy do dwutlenku wêgla z jednoczesn¹redukcj¹ Fe(III) zgodnie z reakcj¹ 11:
C6H12O6 (glukoza) + 6 H2O ++ 24 Fe(III) → 6 CO2 + 24 Fe(II) ++ 24 H+ (reakcja 11)
Ro�nie te¿ na pod³o¿u zawieraj¹cym octan, mle-czan, jab³czan, propionian, pirogronian, bursztynian,benzoesan. Mimo ¿e bakteria ta jest najbardziej spo-krewniona z rodzajem Rhodoferax to nie jest ani bak-teri¹ fotosyntetyzuj¹c¹ ani fermentuj¹c¹ jak inni znaniprzedstawiciele tego rodzaju. Podobnie nie wykryto¿adnej innej bakterii z rodzaju Rhodoferax zdolnej dopozyskiwania energii z utleniania zwi¹zków organicz-nych w po³¹czeniu z redukcj¹ Fe(III) [15].
Procesy redukcji Fe(III) zachodz¹ równie¿ w �ro-dowiskach kwa�nych. Wyizolowano bakteriê Acido-philium cryptum, z klasy "-Proteobacteria, z osadówkwa�nego jeziora pokopalnianego. Jest to Gram-ujem-na pa³eczka, bezwzglêdny beztlenowiec zdolny doutleniania cukrów, kwasów organicznych i wodoruw po³¹czeniu z redukcj¹ ¿elaza [45]. Ze �rodowiskkwa�nych izoluje siê tak¿e bakterie uwa¿ane za neutro-file. Jedna z nich to Desulfitobacterium metallireducensjedyny przedstawiciel rodzaju Desulfitobacterium,Gram-dodatni beztlenowiec zdolny do redukcji Fe(III),Mn(IV), U(VI), Co(III) oraz kwasów humusowych.Redukuje te¿ siarkê S0 i chlorozwi¹zki a utlenia kwa-sy organiczne i alkohole. Inna to Anaeromyxobacterdehalogenans z rzêdu Myxococcales z *-Proteobac-teria, Gram-ujemny fakultatywny beztlenowiec utle-niaj¹cy octan i redukuj¹cy Fe(III), azotany, fumarani zwi¹zki chlorofenolowe [82]. Bakterie autotroficznejak Acidithiobacillus thiooxidans, A. ferrooxidans czySulfolobus acidocaldarius te¿ mog¹ redukowaæ Fe(III)do Fe(II) w kwa�nym pH utleniaj¹c siarkê, co opisujereakcja 12. Proces ten zachodzi w warunkach tleno-wych lub beztlenowych i uwa¿a siê, ¿e nie s³u¿y onprodukcji energii [62].
S0 + 6Fe(III) + 4H2O → HSO4� +
+ 6Fe(II) + 7H+ (reakcja 12)
Oprócz alkalofilnych przedstawicieli rodziny Geo-bacteraceae znane s¹ tak¿e inne gatunki FRM izo-lowane ze �rodowisk o odczynie zasadowym. Nale¿¹tu Gram-dodatnie bezwzglêdne beztlenowce Alkali-philus metalliredigens, którego optymalne pH wzrostu
wynosi 9,5, redukuj¹cy Fe(III), Co(III), Cr(VI) orazkwasy humusowe czy Tindallia magadiensis, bakteriafermentuj¹ca aminokwasy o optymalnym pH wzros-tu 8,5 [100].
Kolejn¹ grup¹ FRM, choæ s³abo poznan¹, s¹ hyper-termofilne archeony ¿yj¹ce w �rodowiskach gdzie pa-nuj¹ temperatury 80�110°C i gdzie znajduje siê du¿ozwi¹zków Fe(III). Utleniaj¹ one wodór, octan, d³ugo-³añcuchowe kwasy t³uszczowe oraz zwi¹zki aromatycz-ne do dwutlenku wêgla. Redukuj¹ równie¿ inne metale,Mn(IV), Au(III), Cr(VI), Co(III), U(VI), Tc(VII). Nale-¿¹ tu przedstawiciele nastêpuj¹cych gatunków Archaea:Pyrobaculum aerophilum, P. islandicum, Pyrodictiumabyssi, Methanopyrus kandleri, Ferroglobus placidus,Geoglobus ahangari, Archeoglobus fulgidus, Pyrococ-cus furiosus, Geogemma barosii (szczep 121) rosn¹cyw temperaturze 121°C oraz bakterii: Thermotoga ma-ritima, Geothermobacterium ferrireducens, Thermo-anaerobacter siderophilus, Thermus sp., [5, 39, 40, 95].
Redukcjê ¿elaza jako proces uboczny towarzysz¹cyprocesowi fermentacji obserwowano dla nastepuj¹cychgatunków i rodzajów bakterii: Paenibacillus polymyxa,Bacillus circulans, B. megaterium, Escherichia coli,Enterobacter aerogenes, Clostridium beijerinckii, C. pas-teurianum, C. sporogenes, Lactobacillus lactis, Pseudo-monas spp., Vibrio sp., Citrobacter freundii [20, 62].W�ród bakterii fermentuj¹cych wykrywa siê te¿ takie,które równie¿ mog¹ pozyskiwaæ energiê w wyniku od-dychania ¿elazowego. Przyk³adem mo¿e byæ Pantoneaagglomerans nale¿¹ca do rodziny Enterobacteriaceae,która wystêpuje g³ównie w glebie oraz w zbiornikachwodnych. Jak wszystkie bakterie nale¿¹ce do tej ro-dziny jest fakultatywnym beztlenowcem i prowadziprocesy fermentacji. Ale wykazano, ¿e tak¿e oddychabeztlenowo redukuj¹c Fe(III) oraz Mn(IV), Cr(VI)i kwasy humusowe utleniaj¹c octan i wodór [24].
Redukcja rozpuszczalnych zwi¹zków Fe(III) orazkwasów humusowych i Cr(VI) jest te¿ procesem ubocz-nym towarzysz¹cym fermentacji, jako podstawowejformie zdobywania energii u �cis³ego beztlenowcaDeinococcus radiodurans. Bakteria ta jest najbardziejopornym organizmem na promieniowanie jonizuj¹cespo�ród dotychczas odkrytych. Prze¿ywa dawki pro-mieniowania o warto�ci do 1500 Gy (grej) [25].
Niektóre gatunki rodzaju Bacillus równie¿ mog¹pozyskiwaæ energiê w wyniku oddychania ¿elazo-wego. Nale¿¹ do nich B. infernus czy B. subterraneus.B. infernus zosta³ wyizolowany z niecki triasowejz g³êboko�ci 2.7 km. Jest to sci�le beztlenowa, termo-filna, halotolerancyjna, lekko alkalofilna bakteria,która fermentuje glukozê a tak¿e redukuje Fe(III),Mn(IV), azotany utleniaj¹c mrówczan i mleczan [9].B. subterraneus wyizolowano z odwiertu g³êbokichwód geotermalnych w basenie artezyjskim w Australii,jest to mezofilny fakultatywny beztlenowiec. Redukuje

110 ANNA SIKORA I MIECZYS£AW K. B£ASZCZYK
Fe(III), Mn(IV), fumaran, azotany i azotyny utlenia-j¹c cukry, glicerol, etanol i mleczan [37].
Redukcja Fe(III) mo¿e towarzyszyæ utlenianiu siar-czanów przez bakterie redukuj¹ce siarczany. Jest toproces uboczny nie s³u¿¹cy produkcji energii obser-wowany u np. rodzaju Desulfovibrio, Desulfobac-terium autotrophicum czy Desulfobacter postgatei[61, 62]. Ale niektórzy przedstawiciele bakterii redu-kuj¹cych siarczany te¿ mog¹ oddychaæ przez reduk-cjê Fe(III), Mn(IV) czy kwasów humusowych. Nale¿¹tu Desulfobulbus propionicus z rodziny Desulfobul-baceae wystepuj¹cej w morskich osadach dennych czyDesulfotomaculum reducens [33, 34].
3. Mechanizmy redukcji Fe(III)
Rozwa¿aj¹c mechanizmy transferu elektronów miê-dzy komórk¹ bakterii a zwi¹zkiem Fe(III) w �rodowis-ku zewnêtrznym nale¿y wzi¹æ pod uwagê nastêpuj¹cezagadnienia: bezpo�redni kontakt mikroorganizmuz nierozpuszczalnym zwi¹zkiem Fe(III); udzia³ prze-no�ników elektronów, zw³aszcza kwasów humusowychznajduj¹cych siê w �rodowisku; produkcjê przeno�-ników elektronów oraz substancji rozpuszczaj¹cychnierozpuszczalne zwi¹zki Fe(III) przez komórkê;bezpo�redni kontakt mikroorganizmu z rozpuszczal-nym zwi¹zkiem Fe(III). W wiêkszo�ci naturalnych�rodowisk Fe(III) wystêpuje najczê�ciej w postacinierozpuszczalnych tlenków. Filogenetycznie odleg³ebakterie z grupy FRM wykszta³ci³y ró¿ne sposobyprzenoszenia elektronów na nierozpuszczalne zwi¹zki¿elaza Fe(III) [79].
Naturalnie w �rodowisku wystêpuj¹ przeno�nikielektronów oraz zwi¹zki chelatuj¹ce, które u³atwiaj¹procesy mikrobiologicznej redukcji zw³aszcza nieroz-puszczalnych zwi¹zków Fe(III). Przeno�nik elektronówwychwytuje elektrony produkowane przez komórkê,w wyniku czego sam ulega redukcji, a nastêpnie abio-tycznie redukuje Fe(III). W efekcie reakcji utlenienianastêpuje regeneracja przeno�nika (Rys. 1). Najlepiejpoznanymi przeno�nikami elektronów s¹ kwasy hu-musowe, które wchodz¹ w sk³ad próchnicy gleboweja tak¿e naturalnych wód. Zawieraj¹ one chinony, zwi¹z-ki o w³a�ciwo�ciach aromatycznych zawieraj¹ce dwiegrupy ketonowe (C = O), których atomy wêgla wcho-dz¹ w sk³ad pier�cienia aromatycznego. Pobranie przeznie elektronów powoduje powstanie wolnych rodnikówchinonowych. Kwasy humusowe mog¹ byæ koñcowymakceptorem elektronów w oddychaniu FRM tak samojak zwi¹zki Fe(III). W �rodowiskach bogatych w kwa-sy humusowe oraz zwi¹zki Fe(III) dochodzi do abio-tycznej redukcji tego ostatniego. Mikrobiologiczniezredukowane kwasy humusowe mog¹ abiotycznie redu-kowaæ zwi¹zki Fe(III) do Fe(II) i same ulegaæ utlenie-
niu w wyniku czego nastêpuje ich regeneracja. W tensposób te same kwasy mog¹ wielokrotnie ulegaæ pro-cesom redukcji i utleniania. Obecno�æ kwasów humu-sowych przyspiesza redukcjê ¿elaza a tak¿e utlenianiematerii organicznej [61, 69]. Oprócz kwasów humu-sowych tak¿e inne zwi¹zki maj¹ zdolno�æ przenosze-nia elektronów. Nale¿¹ do nich zwi¹zki fenolowe znaj-duj¹ce siê w li�ciach drzew np. klonu, dêbu, orzechaw³oskiego. Uwa¿a siê, ¿e substancje humusowe wy-stêpuj¹ce w naturalnych �rodowiskach posiadaj¹ te¿w³a�ciwo�ci chelatuj¹ce (chelacyjne), czyli odznacza-j¹ siê zdolno�ci¹ do chemicznego wi¹zania z metalamii minera³ami, przez co zwiêkszaj¹ ich rozpuszczal-no�æ. Pewne �rodowiska obftuj¹ w naturalne przeno�-niki elektronów oraz zwi¹zki chelatuj¹ce, w innychjest ich deficyt [79]. W badaniach laboratoryjnych dobadania mechanizmów redukcji Fe(III) przy udzialeprzeno�ników elektronów jako analogów kwasów hu-musowych stosuje siê chinony, najczê�ciej dwusiar-czan 2,6-antrachinonu (AQDS) [69].
Mechanizmy transferu elektronów z komórki bak-terii na zewn¹trzkomórkowe akceptory jakimi s¹zwi¹zki Fe(III) zosta³y do�æ dobrze poznane u G. sul-furreducens, poniewa¿ jego genom zosta³ ca³kowiciezsekwencjonowany. Ciekaw¹ cech¹ tej bakterii jestobecno�æ 111 genów koduj¹cych bia³ka uwa¿ane zacytochromy typu c na podstawie posiadanej charakte-rystycznej sekwencji CXXCH wi¹¿¹cej hem. Szacujesiê, ¿e 31 tych bia³ek zlokalizowanych jest w zew-nêtrznej b³onie komórkowej [75]. Mechanizmy reduk-cji nierozpuszczalnych i rozpuszczalnych zwi¹zków¿elaza czê�ciowo ró¿ni¹ siê. Schemat mechanizmu re-
Rys. 1. Mechanizm dzia³ania przeno�ników elektronóww procesie redukcji zwi¹zków ¿elaza (III) przez mikroorganizmy
redukuj¹ce ¿elazo
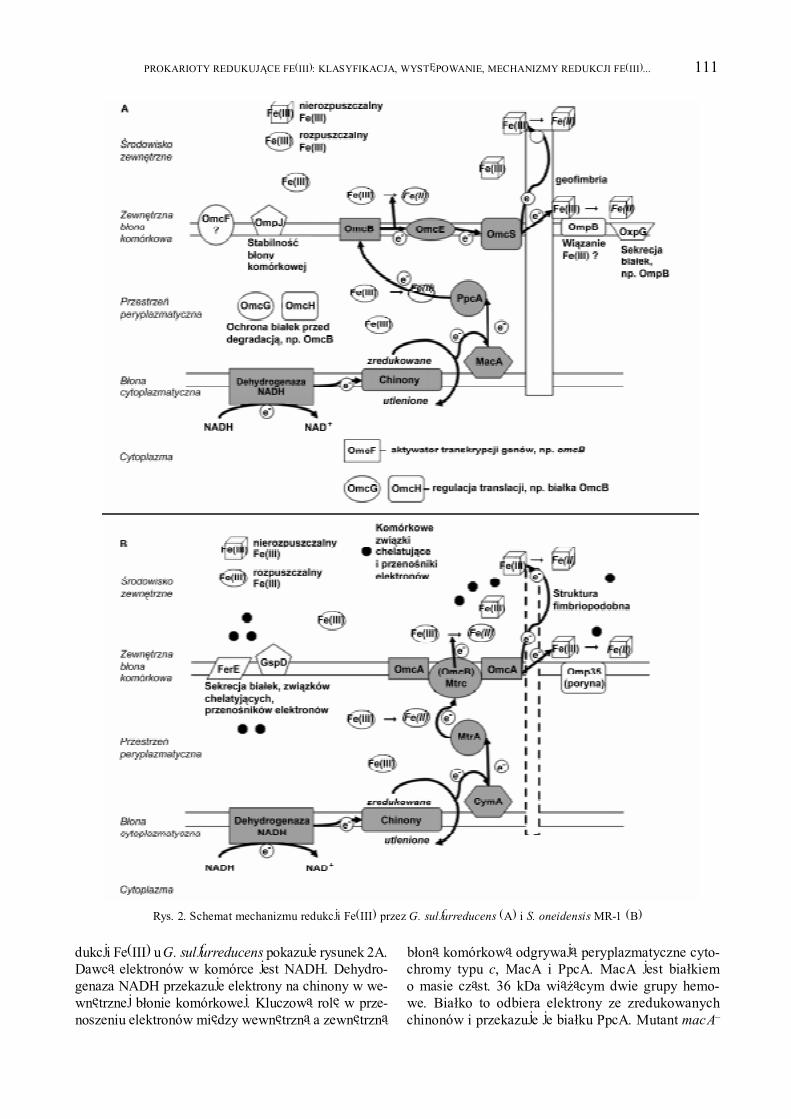
111PROKARIOTY REDUKUJ¥CE FE(III): KLASYFIKACJA, WYSTÊPOWANIE, MECHANIZMY REDUKCJI FE(III)...
dukcji Fe(III) u G. sulfurreducens pokazuje rysunek 2A.Dawc¹ elektronów w komórce jest NADH. Dehydro-genaza NADH przekazuje elektrony na chinony w we-wnêtrznej b³onie komórkowej. Kluczow¹ rolê w prze-noszeniu elektronów miêdzy wewnêtrzn¹ a zewnêtrzn¹
b³on¹ komórkow¹ odgrywaj¹ peryplazmatyczne cyto-chromy typu c, MacA i PpcA. MacA jest bia³kiemo masie cz¹st. 36 kDa wi¹¿¹cym dwie grupy hemo-we. Bia³ko to odbiera elektrony ze zredukowanychchinonów i przekazuje je bia³ku PpcA. Mutant macA�
Rys. 2. Schemat mechanizmu redukcji Fe(III) przez G. sulfurreducens (A) i S. oneidensis MR-1 (B)

112 ANNA SIKORA I MIECZYS£AW K. B£ASZCZYK
wykazuje znaczne ni¿sz¹ aktywno�æ redukcji Fe(III),gdy dostarczone jest ono w postaci rozpuszczalnegocytrynianu Fe(III) [12]. PpcA jest bia³kiem o masiecz¹st. 9,6 kDa wi¹¿¹cym trzy grupy hemowe. MutacjappcA obni¿a redukcjê cytrynianu Fe(III) o 60% w po-równaniu ze szczepem dzikim, gdy dawc¹ elektronówjest octan. Bia³ko to nie ma znaczenia, gdy �ród³oelektronów stanowi wodór, bo utlenianie wodoru ka-talizowane jest przez peryplazmatyczn¹ hydrogenazê.PpcA jest niezbêdne do procesów redukcji uranu (VI)oraz kwasów humusowych [50]. PpcA przenosi elek-trony na bia³ko OmcB, które jest cytochromem typu cwystêpuj¹cym w zewnêtrznej b³onie komórkowej.Wi¹¿e ono wiele grup hemowych i pe³ni bardzo istotn¹rolê w transporcie elektronów do zewnêtrznej b³onykomórkowej, do bia³ek OmcE i OmcS. Mutant omcB�
G. sulfurreducens jest niezdolny do wzrostu na pod³o-¿u zawieraj¹cym Fe(III) w postaci rozpuszczalnegocytrynianu. Bia³ko OmcB konieczne jest te¿ do reduk-cji nierozpuszczalnych tlenków Fe(III) oraz wymaga-ne jest, gdy dawc¹ elektronów jest zarówno octan jaki wodór. W komórce G. sulfurreducens wystêpuje jesz-cze bia³ko OmcC wykazuj¹ce a¿ 79% identyczno�cido OmcB, ale w mutancie omcC� proces redukcji ¿e-laza w postaci cytrynianu ¿elazowego przebiega po-dobnie jak w szczepie dzikim. Funkcja bia³ka OmCnie jest znana [46].
OmcS, bia³ko o masie cz¹st. 50 kDa wi¹¿¹ce sze�ægrup hemowych, i OmcE, bia³ko o masie cz¹st. 30 kDawi¹¿¹ce cztery grupy hemowe, to cytochromy typu czlokalizowane na powierzchni zewnêtrznej b³ony ko-mórkowej, które s¹ konieczne do wzrostu na pod³o¿uzawieraj¹cym nierozpuszczalny tlenek Fe(III) jako je-dyny akceptor elektronów. Zaobserwowano, ¿e mutantomcE� wykazywa³ niewielk¹ zdolno�æ do redukcjitlenków Fe(III) i Mn(IV), natomiast omcS� nie, cowskazuje na wa¿niejsz¹ rolê bia³ka OmcS w proce-sach redukcji nierozpuszczalnych zwi¹zków Fe(III) czyMn(IV). Mutanty omcE� i omcS� rosn¹ na pod³o¿uz cytrynianem Fe(III) jak szczep dziki. Uwa¿a siê, ¿erol¹ bia³ek OmcS i OmcE jest przekazywanie elektro-nów na geofimbrie (opis ni¿ej). Gen omcS prawdopo-dobnie tworzy operon z genem omcT koduj¹cym bia³-ko OmcT, które wykazuje 62.6% homologii do bia³kaOmcS. Jego funkcja jest nieznana [74].
Badania pokazuj¹, ¿e redukcja Fe(III) wymagawspó³dzia³ania wielu cytochromów typu c. Rol¹ jed-nych jest transfer elektronów, inne pe³ni¹ funkcje re-gulatorowe. Do tej drugiej grupy nale¿¹ bia³ka OmcF,OmcG i OmcH. Bia³ko OmcF wi¹¿¹ce tylko jedn¹ gru-pê hemow¹ jest najmniejszym z cytochromów typu cG. sulfurreducens, jego masa cz¹st. wynosi 9.4 kDa.Mutacja omcF powoduje znaczn¹ utratê zdolno�ci re-dukcji rozpuszczalnego cytrynianu Fe(III). W mutan-tach omcF� nie wykrywa siê mRNA dla bia³ek OmcB
i OmcC oraz obserwuje siê zmiany ilo�ciowe innychcytochromów typu c. Bia³ko OmcF jest uwa¿ane zaaktywator transkrypcji genów, g³ównie omcB, a tak¿eomcC i prawdopodobnie innych. Ciekawym zjawis-kiem jest fakt, ¿e mutacji omcF towarzyszy nadekspre-sja bia³ka OmcS, co umo¿liwia bakterii spowolnionywzrost na pod³o¿u z cytrynianem ¿elaza. Bia³ko OmcFwykrywa siê te¿ w zewnêtrznej b³onie komórkowejbakterii [42].
OmcG, bia³ko o masie cz¹st. 78.7 kDa, i OmcH,bia³ko o masie cz¹st. 103 kDa, to dwa homologicznecytochromy typu c wi¹¿¹ce wiele grup hemowych.Wykazuj¹ 84.4% identyczno�ci. Nie okre�lano ich lo-kalizacji w komórce. W mutantach omcG� i omcH� niewykrywa siê bia³ka OmcB, chocia¿ transkrypcja genuomcB zachodzi bez zak³óceñ. Ekspresja tylko jednegoz dwóch bia³ek OmcG lub OmcH z plazmidu znosiefekty podwójnej mutacji omcG omcH. Przypuszczal-n¹ rol¹ bia³ek OmcG i OmcH jest regulacja translacjilub/i stabilizacja bia³ka OmcB oraz ochrona przed de-gradacj¹ w przestrzeni peryplazmatycznej [41].
Ponadto przy deficycie ostatecznych akceptorówelektronów w pod³o¿u, ich rolê mog¹ przej�ciowo pe³-niæ cytochromy typu c, znajduj¹ce siê w periplazmiei zewnêtrznej b³onie komórkowej. Szacuje siê, ¿e takimechanizm dostarcza komórce G. sulfurreducens ener-gii wystarczaj¹cej do przetrwania przez 8 minut lubzapewnia ruch wici na pokonanie odleg³o�ci kilkusetd³ugo�ci komórki, aby przemie�ciæ siê do miejsca gdziewystêpuj¹ akceptory elektronów [22].
Do prawid³owego przebiegu procesów redukcjiFe(III) niezbêdnie konieczne s¹ równie¿ bia³ka zew-nêtrznej b³ony komórkowej nie bêd¹ce przeno�nikamielektronów. Nale¿¹ do nich bia³ka: OmpJ, OxpG, OmpB.Pe³ni¹ one rolê po�redni¹ w procesie transferu elektro-nów. OmpJ jest najobficiej wystêpuj¹cym bia³kiemw zewnêtrznej b³onie komórkowej. Jest to bia³ko spe-cyficzne dla rodziny Geobacteraceae. Nale¿y ono doporyn, grupy bia³ek b³onowych buduj¹cych kana³ydyfuzyjne (pory) w zewnêtrznej b³onie komórkowej.Poryny maj¹ strukturê $-harmonijki. Pory umo¿liwiaj¹biern¹ dyfuzjê ró¿nych roztworów wodnych. Obecno�æbia³ka OmpJ wymagana jest do redukcji rozpuszczalne-go cytrynianu Fe(III) oraz nierozpuszczalnych tlenkówFe(III) i Mn(IV), poniewa¿ odgrywa ono wa¿n¹ rolêw utrzymaniu fizycznej integralno�ci b³ony komórko-wej, zapewniaj¹c jednocze�nie jej prawid³owe funkcjo-nowanie. Integralno�æ b³ony komórkowej jest prawdo-podobnie niezbêdnym czynnikiem dojrzewania bia³ekprzenosz¹cych elektrony i ca³ego procesu przenoszeniaelektronów. W mutancie ompJ� G. sulfurreducens na-stêpuje ok. 50% obni¿enie ilo�ci bia³ek cytochromówtypu c w przestrzeni peryplazmatycznej oraz w ze-wnêtrznej b³onie komórkowej. Obserwuje siê równie¿powiêkszenie przestrzeni peryplazmatycznej [1].

113PROKARIOTY REDUKUJ¥CE FE(III): KLASYFIKACJA, WYSTÊPOWANIE, MECHANIZMY REDUKCJI FE(III)...
OmpB, bia³ko o masie cz¹st. 139.6 kDa, jest praw-dopodobnie oksydaz¹ miedziow¹, gdy¿ posiada czterymiejsca wi¹zania jonów miedzi, dwa przy N-terminal-nym i dwa przy C-terminalnym koñcu. Wykazuje naj-wiêksze podobieñstwo do oksydazy manganowej MofALeptothrix discophora, bakterii utleniaj¹cej zwi¹zkiMn(II). Bia³ko OmpB ulega ekspresji na tym samympoziomie zarówno przy rozpuszczalnych jak i nieroz-puszczalnych zwi¹zkach Fe(III) jako ostatecznych ak-ceptorach elektronów. Jest to bia³ko wystêpuj¹ce napowierzchni zewnêtrznej b³ony komórkowej, wykry-wa siê go równie¿ w p³ynach zewn¹trzkomórkowych.Mutant ompB- G. sulfurreducens redukuje rozpusz-czalny cytrynian ¿elaza oraz fumaran jak szczep dziki,natomiast wykazuje znacznie obni¿on¹ redukcjê nie-rozpuszczalnych tlenków Fe(III) oraz Mn(IV). Niewiadomo czy bia³ko OmpB jest zaanga¿owane w re-dukcjê Fe(III) i Mn(IV) czy j¹ po�rednio u³atwia pe³-ni¹c funkcjê pomocnicz¹. Mimo ¿e bia³ko to jest uwa-¿ane za oksydazê miedziow¹, to posiada te¿ domenêodpowiedzialn¹ za wi¹zanie fibronektyny, charaktery-styczn¹ dla bakteryjnych hydrolaz, zw³aszcza celulazi chitynaz, odpowiedzialn¹ za adhezjê bia³ka do poli-sacharydu i za zachowanie prawid³owej konformacjicentrum aktywnego enzymu. W bia³ku OmpB wykry-to równie¿ sekwencjê, która mo¿e byæ potencjalnymmiejscem wi¹zania ¿elaza. Uwa¿a siê, ¿e domeny od-powiedzialne za wi¹zanie ¿elaza i fibronektyny odgry-waj¹ rolê w odzia³ywaniu miêdzy bia³kiem OmpBa tlenkami Fe(III) oraz Mn(IV). Za sekrecjê bia³kaOmpB odpowiada bia³ko OxpG, które jest pseudo-pilin¹, nale¿¹c¹ do bia³ek ogólnego systemu sekrecjitypu II. Pseudopiliny tworz¹ fimbriopodobne strukturyperyplazmatyczne, umo¿liwiaj¹ce sekrecjê bia³ek przeztworzenie kana³u dla wydzielanego bia³ka lub wy-pychanie go na zewn¹trz b³ony. Przypuszcza siê, ¿erol¹ bia³ka OxpG jest tak¿e sekrecja innych bia³ekodpowiedzialnych za redukcjê nierozpuszczalnychtlenków Fe(III) i Mn(IV). Mutant oxpG� nie jest zdol-ny do redukcji nierozpuszczalnych tlenków Fe(III)oraz Mn(IV). Natomiast redukcja rozpuszczalnegocytrynianu ¿elazowego zachodzi bez zmian, tak jaku szczepu dzikiego [73].
Pocz¹tkowo uwa¿ano, ¿e w redukcji Fe(III) bior¹udzia³ tylko cytochromy typu c. I tak jest, gdy akcep-torem elektronów s¹ rozpuszczalne zwi¹zki ¿elaza,które odbieraj¹ elektrony bezpo�rednio od bia³ek w ze-wnêtrznej b³onie komórkowej lub w przestrzeni pery-plazmatycznej. Jako ciekawostkê nale¿y tu podaæ, ¿eu bakterii z rodzaju Pelobacter nie wykryto cytochro-mów typu c [66]. Ostatnio u G. sulfurreducens wykrytofimbrie, nazwane geofimbriamii, które maj¹ zdolno�æprzenoszenia elektronów. Zbadano przewodnictwoelektryczne bia³ek fimbrii i innych bia³ek zewn¹trzko-mórkowych i wykazano, ¿e bia³ka geofimbrii s¹ do-
brymi przewodnikami elektronów i nazwano je �na-noprzewodami� (microbial nanowires). Geofimbrietworz¹ siê, gdy bakterie rosn¹ na pod³o¿u zawieraj¹-cym nierozpuszczalne lub s³abo rozpuszczalne zwi¹zkitrójwarto�ciowego ¿elaza, np. tlenki ¿elaza a tak¿eprzy innym ni¿ Fe(III) akceptorze elektronów, np.fumaranie w warunkach suboptymalnych. Przy roz-puszczalnym cytrynianie ¿elazowym nie stwierdzonoich obecno�ci. Przypomnijmy, ¿e w wiêkszo�ci �rodo-wisk dominuj¹ nierozpuszczalne tlenki Fe(III). Bia³ko,z którego zbudowane s¹ geofimbrie to produkt genupilA [84]. PilA jest bia³kiem strukturalnym fimbriitypu IV. Fimbrie typu IV odpowiedzialne s¹ za poru-szanie siê komórki niezale¿ne od wici, kolonizacjêi tworzenie biofilmów, zapewniaj¹ kontakt bakteriiz pod³o¿em. Najlepiej poznane s¹ one u bakterii pato-gennych, np. z rodzaju Neisseria czy Pseudomonas[72]. W przypadku G. sulfurreducens bia³ko PilA jestkrótsze ni¿ u innych bakterii, ale zawiera silnie kon-serwowan¹ N-terminaln¹ domenê fimbrii typu IV.Mutant pilA� Geobacter sulfurreducens nie rós³ napod³o¿u, gdzie jedynym akceptorem elektronów by³ynierozpuszczalne tlenki Fe(III). Natomiast, gdy dopod³o¿a dodano inny akceptor elektronów, np. fuma-ran, to zarówno szczep dziki jak i mutant pilA� wyka-zywa³y normalny wzrost. Na zdjêciach spod mikro-skopu elektronowego widaæ by³o, ¿e kryszta³y tlenku¿elaza otacza³y geofimbrie w przypadku dzikiegoszczepu a w mutancie pilA� tworzy³y skupiska przykomórkach. To wskazuje, ¿e funkcj¹ fimbrii nie jestzapewnienie fizycznego kontaktu z kryszta³ami zwi¹z-ku ¿elaza, ale bezpo�redni udzia³ w przenoszeniu elek-tronów. Dok³adny mechanizm tego zjawiska nie jestjeszcze poznany. Wprowadzenie plazmidu z dzikimgenem pilA na plazmidzie do mutanta pilA� powodo-wa³o zniesienie efektów mutacji i powstanie funkcjo-nalnych geofimbrii. Uwa¿a siê, ¿e geofimbrie odgry-waj¹ wa¿n¹ rolê w �rodowiskach, gdzie przewa¿aj¹nierozpuszczalne zwi¹zki ¿elaza trójwarto�ciowego[84]. Pokazano, ze fimbrie G. sulfurreducens s¹ ko-nieczne zarówno do tworzenia biofilmu jak i przeno-szenia elektronów na anodê w mikrobiologicznychogniwach paliwowych [85, 86].
Procesy redukcji Fe(III) zosta³y te¿ do�æ dobrze po-znane dla Shewanella oneidensis szczep MR-1 (daw-niej S. putrefaciens szczep MR-1) (rys. 2B), którejgenom równie¿ zosta³ zsekwencjonowany. GenomS. oneidensis zawiera 42 geny koduj¹ce bia³ka cyto-chromy typu c, 15 z nich zwi¹zanych jest z b³onami,w tym 5 z b³on¹ zewnetrzn¹ [31]. Podobnie jaku G. sulfurreducens bia³kowe przeno�niki elektronówodbieraj¹ je ze zredukowanych chinonów w wewnêtrz-nej b³onie komórkowej. Pierwszym jest CymA, bia³kob³ony cytoplazmatycznej o masie cz¹st. 20,8 kDa,które jest cytochromem typu c, wi¹¿¹cym cztery grupy

114 ANNA SIKORA I MIECZYS£AW K. B£ASZCZYK
hemowe. Mutant cymA� jest defektywny w redukcjizwi¹zków Fe(III) i Mn(IV) a tak¿e azotanów i fuma-ranu [77]. Bia³ko Cym A przenosi elektrony na bia³koMtrA o masie cz¹st. 36 kDa, które jest cytochromemtypu c wi¹¿¹cym 10 grup hemowych zlokalizowanymw peryplazmie. Mutant mtrA� wykazuje brak zdol-no�ci redukcji Fe (III). Rol¹ bia³ka MtrA jest przeka-zanie elektronów cytochromom typu c zewnêtrznejb³ony komórkowej, bia³kom MtrC (OmcB) i OmcA.Byæ mo¿e ³añcuszek bia³ek przenosz¹cych elektronyjest d³u¿szy, ale nie zosta³y one jeszcze poznane. Masacz¹st. bia³ka MtrC wynosi 75 kDa, a bia³ka OmcA83 kDa, oba wi¹¿¹ 10 grup hemowych, s¹ lipopro-teinami i pe³ni¹ funkcjê reduktaz Fe(III). Tworz¹ sta-bilny kompleks bia³kowy, heterotrimer zbudowanyz dwóch cz¹steczek bia³ka OmcA i jednej cz¹steczkiMtrC. Pojedyncze mutanty mtrC� i omcA� zachowuj¹czê�ciow¹ zdolno�æ redukcji rozpuszczalnych i nieroz-puszczalnych zwi¹zków Fe (III) [3, 92].
Podobnie jak u G. sulfurreducens w komórkachS. oneidensis inne bia³ka ni¿ tylko cytochromy typu cuczestnicz¹ w procesie redukcji ¿elaza. W zewnêtrznejb³onie komórkowej znajduje siê bia³ko MtrB o masiecz¹st. 75 kDa, które posiada charakterystyczn¹ se-kwencjê CXXC wi¹¿ac¹ metale. Prawdopodobn¹ rol¹bia³ka MtrB jest wi¹zanie bakterii z jonem metaluw trakcie procesu redukcji zwi¹zków Fe(III) i Mn(IV).Bia³ko MtrB mo¿e te¿ odpowiadaæ za w³a�ciw¹ loka-lizacjê bia³ek MtrC i OmcA w b³onie zewnêtrznej.Mutant mtrB� jest niezdolny do redukcji Fe(III)i Mn(IV) [4, 93]. Innymi bia³kami s¹ bia³ka zewnêtrz-nej b³ony komórkowej nale¿¹ce do ogólnego systemusekrecji typu II: produkty genów ferE (gspE) orazgspD odpowiedzialne za translokacjê bia³ek MtrCi OmcA do i przez zewnêtrzn¹ b³onê komórkow¹. Mu-tanty ferE i gspD s¹ niezdolne do wzrostu na pod³o¿uzawieraj¹cym zwi¹zki Fe(III) i Mn(IV). Istnieje hipo-teza, ¿e ten system sekrecji bierze te¿ udzia³ w uwal-nianiu na zewn¹trz komórki substancji bêd¹cych prze-no�nikami elektronów oraz zwi¹zków chelatuj¹cych(patrz ni¿ej) [19, 93]. Kolejnym bia³kiem wp³ywaj¹cymna proces redukcji Fe (III) jest bia³ko z grupy poryn,Omp35, o masie cz¹st. 35 kDa wystêpuj¹ce w zewnêtrz-nej b³onie komórkowej. Mutanty omp35� wykazuj¹zaburzenia w procesie redukcji fumaranu, azotanówi zwi¹zków Fe(III) oraz Mn(IV), ale dok³adny mecha-nizm tego zjawiska nie zosta³ poznany [71].
U S. oneidensis wykryto te¿ fimbriopodobne, jakto okre�lili autorzy, struktury pe³ni¹ce funkcjê nano-drutów, ale nie okre�lono z jakich bia³ek s¹ one zbu-dowane. Stwierdzono, ¿e tworz¹ siê one w warunkachbeztlenowych lub przy ograniczonej ilo�ci tlenu w �ro-dowisku. Wykazano równie¿, ¿e przewodzenie elek-tronów przez nanodruty u S. oneidensis wymaga funk-cjonalnych cytochromów typu c MtrC i OmcA [28].
We wnêtrzu komórek S. oneidensis wykryto gra-nule zawieraj¹ce tlenki Fe(II) i Fe(III) oraz tlenkiMn(IV). Uwa¿a siê, ¿e bakterie te przechowuj¹ mine-ra³y zawieraj¹ce Fe(III) lub Mn(IV) w komórce jakozapas ostatecznych akceptorów elektronów, które s¹wykorzystywane w warunkach beztlenowych przybraku tych zwi¹zków w �rodowisku [26, 27].
U gatunków z rodzaju Shewanella oraz u Geothrixfermentans opisano strategiê umo¿liwiaj¹c¹ redukcjênierozpuszczalnych zwi¹zków Fe(III), bez bezpo�red-niego kontaktu miêdzy komórk¹ bakterii a akcepto-rem elektronów. Polega ona na uwalnianiu zwi¹zkówpe³ni¹cych funkcjê przeno�nika elektronów (flawinyi chinony) oraz zwi¹zków chelatuj¹cych (np. chinony).U rodzaju Shewanella zidentyfikowano przeno�nikielektronów, którymi s¹ flawiny, ryboflawina oraz mo-nonukleotyd flawinowy (FMN) [14]. Uwa¿a siê, ¿eprzeno�nikiem elektronów u G. fermentans jest roz-puszczalny w wodzie zwi¹zek z grupy chinonów o ma-sie cz¹st. mniejszej ni¿ 12 kDa, chocia¿ nie zosta³ onzidentyfikowany. Produkcja przeno�ników elektronówjest wydatkiem energetycznym dla komórki i op³acasiê, gdy bakterie ¿yj¹ w populacjach o du¿ych gês-to�ciach, przede wszystkim, gdy tworz¹ biofilm [79].W badaniach nad mechanizmami redukcji nierozpusz-czalnych zwi¹zków Fe(III) u S. oneidensis stwierdzo-no, ¿e substancje pe³ni¹ce funkcje przeno�ników elek-tronów wydzielane s¹ tylko do matriks biofilmuutworzonego przez te bakterie [47].
W obecno�ci azotanów czy fumaranu jako akcep-torów elektronów w pod³o¿u bakterie redukuj¹ce ¿e-lazo (G. metalireducens, G. sulfurreducens, G. fermen-tans i S. alga) redukuj¹ je, wykorzystuj¹c jako �ród³oelektronów zredukowane kwasy humusowe a tak¿ezredukowane zwi¹zki Fe(II). Zjawisko wykorzystan-nia zredukowanych kwasów humusowych jako �ród³aelektronów do redukcji azotanów obserwuje siê te¿ dlabakterii denitryfikacyjnych [23, 68].
Redukcja Fe(III), która nie s³u¿y produkcji energii,u bakterii fermentuj¹cych jest wynikiem dzia³aniahydrogenaz. Dawc¹ elektronów do procesów redukcjijest wodór. Pokazano, ¿e w warunkach beztlenowychE. coli mo¿e redukowaæ rozpuszczalne zwi¹zki tech-netu Tc(VII) do nierozpuszczalnych form ulegaj¹cychwytr¹caniu. Reakcja ta jest katalizowana przez hydro-genazê Hyc, która wchodzi w sk³ad kompleksu FHL,odpowiedzialn¹ za produkcjê wodoru z mrówczanuw fermentacji kwasów mieszanych (fermentacji kwasumrówkowego). Dawcê elektronów do redukcji Tc(VII)stanowi mrówczan lub wodór [52]. Okazuje siê rów-nie¿, ¿e hydrogenaza I Clostridium pasteurianum posia-da aktywno�æ reduktazy selenianu (IV) SeO3
2� → S0
a tak¿e zwi¹zków telluru (IV) TeO32�. Elektrony po-
chodz¹ z utleniania cz¹steczek wodoru, a po�rednikiemw ich przenoszeniu jest ferredoksyna [99]. Z kolei

115PROKARIOTY REDUKUJ¥CE FE(III): KLASYFIKACJA, WYSTÊPOWANIE, MECHANIZMY REDUKCJI FE(III)...
u prokariotów redukuj¹cych siarczany rolê reduktazymetali pe³ni cytochrom c3 [65].
Fe(III) mo¿e byæ te¿ redukowane do Fe(II) abiotycz-nie, np. w reakcji z siarkowodorem produkowanymprzez prokarioty redukuj¹ce siarczany zgodnie z reak-cjami 13�15 [20]:
2Fe3+ + H2S → 2Fe2+ + 2H+ + S0 (reakcja 13)Fe2+ + H2S → FeS + 2H+ (reakcja 14)2Fe3+ + 2H2S → FeS2 + Fe2+ + 4H+ (reakcja 15)
Podobn¹ redukcjê Fe(III) do Fe(II) obserwuje siêw reakcji z innym metabolitem jakim jest kwas mrów-kowy zgodnie z reakcj¹ 16 [62]:
CH3COO� + 8Fe(III) + 4H2O V 2HCO3� +
+ 8Fe(II) + 9H+ (reakcja 16)
4. Rola ekologiczna prokariotówredukuj¹cych Fe(III) w �rodowisku
Redukcja Fe(III) uwa¿ana jest za jedn¹ z pierwot-nych, obok redukcji S0, a nawet wrêcz najbardziejpierwotn¹ form¹ oddychania mikroorganizmów. Uwa-¿a siê, ¿e pierwsze organizmy ¿ywe to przedstawicieleFRM. Prawdopodobnie utlenianie wodoru po³¹czonez redukcj¹ Fe(III) by³o prametabolizmem, który da³pocz¹tek ewolucji ¿ycia na Ziemi. Warunki �rodowi-skowe wczesnego okresu pocz¹tków ¿ycia na Ziemi,3�5 miliardów lat temu, czyli wysoka temperatura,du¿a ilo�æ wodoru oraz zwi¹zków ¿elaza pocho-dz¹cych z wnêtrza Ziemi a wystêpuj¹cych na jejpowierzchni w formie utlenionej Fe(III), g³ównie naskutek intensywnego dzia³ania promieniowania UV,sprzyja³a produkcji energii w wyniku utleniania wo-doru i redukcji zwi¹zków Fe(III). Istnieje wiele dowo-dów na to, ¿e z³o¿a magnetytu z okresu wczesnegoprekambru s¹ pochodzenia mikrobiologicznego a do-k³adnie wynikiem aktywno�ci ¿yj¹cych ówcze�niehypertermofilnych FRM. Metod¹ badania stosunkuzawarto�ci dwóch izotopów wêgla 12C do 13C stwier-dzono, ¿e wiek materii organicznej towarzysz¹cejprekambryjskim z³o¿om magnetytu jest �ci�le sko-relowany z wiekiem utlenionej materii organicznejz tamtego okresu. Jak dotychczas nie zosta³ poznany¿aden abiotyczny proces zwi¹zany z jednoczesnymutlenianiem materii organicznej i redukcj¹ Fe(III) domagnetytu [61, 62].
Uwa¿a siê, ¿e analogiczne procesy mikrobiologicz-nej redukcji Fe(III) w po³¹czeniu z utlenianiem wo-doru jakie mia³y miejsce w okresie pocz¹tków ¿y-cia na Ziemi obecnie odbywaj¹ siê w �rodowiskach,w których panuj¹ wysokie temperatury (80�110°C),np. w gor¹cych �ród³ach. Gor¹ce �ród³a obfituj¹ w du¿eilo�ci Fe(II), które ulega utlenieniu do formy Fe(III)
w zetkniêciu siê z powietrzem. Wokó³ �róde³ gro-madz¹ siê osady bogate w zwi¹zki Fe(III). Z gor¹-cych �róde³ wydobywaja siê te¿ du¿e ilo�ci wodoru.Wszystko to sprawia, ¿e tworz¹ siê idealne warunkido rozwoju termofilnych i hypertermofilnych beztle-nowych mikroorganizmów redukuj¹cych Fe(III), dlaktórych �ród³em elektronów jest utlenianie wodoru.Oprócz wodoru wspó³czesne termofilne i hypertermo-filne FRM prawdopodobnie utleniaj¹ równie¿ zwi¹zkiorganiczne jak octan, zwi¹zki aromatyczne czy d³ugo-³añcuchowe kwasy t³uszczowe. Dowodem na aktyw-no�æ tych mikroorganizmów w gor¹cych �rodowiskachjest wykrywany tam magnetyt w postaci ziarnistychstruktur [39, 61, 62, 95]. Równie¿ meteoryty pocho-dz¹ce z Marsa zawieraj¹ ziarniste struktury magnety-tu. Istnieje przypuszczenie, ¿e powsta³y one w wynikuaktywo�ci FRM na Marsie [61]. Jak wiadomo badanianad magnetytem pozwoli³y na odkrycie zasad mag-netyzmu i elektryczno�ci. Jako ciekawostkê warto tudodaæ, ¿e pad³o nawet stwierdzenie, ¿e gdyby nieG. metallireducens (GS-15) i inne FRM nie mieli-by�my dzi� radia i telewizji [61].
Innymi minera³ami powsta³ymi w wyniku aktyw-no�ci bakterii redukuj¹cych Fe(III) i Mn(IV) s¹ syde-ryt (FeCO3), wiwianit [Fe3(PO4) × 8H2O] czy rodo-chrosyt (MnCO3). FRM mog¹ te¿ redukowaæ zwi¹zkiuranu U(VI), w wyniku czego powstaj¹ z³o¿a U(IV)w postaci uraninitu (UO2) [61].
Analogiczne procesy mikrobiologicznej redukcjiU(VI) do tych zachodz¹cych w warunkach mezo-filnych maj¹ te¿ miejsce w �rodowiskach termo-i hypertermofilnych. Pokazano, ¿e hypertermofilnyarcheon Pyrobaculum islandicum jest zdolny do re-dukcji U(VI) do U(IV) w temperaturze 100°C, w wy-niku czego powstaje nierozpuszczalny uraninit. Re-dukcja U(VI) wymaga obecno�ci wodoru jako dawcyelektronów [40].
Z mikrobiologiczn¹ redukcj¹ Fe(III) w glebachzwi¹zane jest zjawisko powstawania gleb glejowych.Polega ono na tym, ¿e gleba staje siê kleista i przyj-muje szar¹ lub zielonkawo-niebiesk¹ barwê. Jest towynik powstania zwi¹zku Fe3(OH)8 zawieraj¹cego¿elazo na stopniu utlenienia +2 i +3 w wyniku reakcji17 i 18 [20]:
Fe(OH)3 + 3H+ + e � Fe2+ + 3H2O (reakcja 17)2 Fe(OH)3 + Fe2+ ++ 2OH� → Fe3(OH)8 (reakcja 18)
Innym zjawiskiem jest lateryzacja gleby w wilgot-nych i gor¹cych klimatach. W wyniku intensywnejaktywno�ci prokariotów redukuj¹cych Fe(III) cz¹stkiorganiczne ulegaj¹ prawie ca³kowitej mineralizacjii jednocze�nie tworz¹ siê du¿e ilo�ci tlenków Fe(II),które ulegaj¹ wytr¹caniu i powoduj¹ jakby cemento-wanie cz¹steczek gleby. W efekcie tworzy siê twardy
116 ANNA SIKORA I MIECZYS£AW K. B£ASZCZYK
czerwony lub ciemnobr¹zowy lateryt. Jego sk³adnikiems¹ tak¿e tlenki glinu [20].
Redukcja Fe(III) oraz Mn(IV) przez bakterie wp³y-wa na stan wód, g³ównie wód gruntowych. W wyni-ku reakcji redukcji niektóre nierozpuszczalne tlenkiFe(III) i Mn(IV) ulegaj¹ przekszta³ceniu do rozpusz-czalnych zwi¹zków Fe(II) i Mn(II). W takiej formiedostaj¹ siê do wód gruntowych, gdzie w wyniku ze-tkniêcia siê z powietrzem ulegaj¹ utlenieniu do nie-rozpuszczalnych tlenków Fe(III) i Mn(IV). Powodujeto z³¹ jako�æ wody pitnej, jej nieodpowiedni smaki kolor, problemy typu zatykanie siê rur wodno-kana-lizacyjnych, itp. Ponadto wraz z rozpuszczalnymi zre-dukowanymi zwi¹zkami Fe(II) i Mn(II) do wód grun-towych mog¹ przedostaæ siê fosforany i �ladowe ilo�citoksycznych metali, które zwykle adsorbuj¹ siê na nie-rozpuszczalnych tlenkach Fe(III) i Mn(IV). W efekcietych procesów nastêpuje wzrost si³y jonowej, wzroststê¿enia kationów, tak¿e wzrost pH i ska¿enie wódmetalami toksycznymi. To wszystko z kolei ma nie-korzystny wp³yw na wzrost ro�lin [61, 62].
Jak ju¿ wspomniano przy omawianiu mechaniz-mów redukcji Fe(III) koñcowym akceptorem elektro-nów w oddychaniu FRM mog¹ byæ kwasy humusowe.Zredukowane kwasy humusowe w �rodowisku mo-g¹ stanowiæ �ród³o elektronów do innych procesówmikrobiologicznej redukcji np. azotanów, selenianów,arsenianów [61].
Utlenianie materii organicznej z jednoczesn¹ re-dukcj¹ Fe(III) lub Mn(IV) stanowi wa¿ny elementw obiegu wêgla w beztlenowych osadach dennychs³odkowodnych zbiorników wodnych, w osadach mor-skich, na terenach podmok³ych, w podmok³ych glebach,np. pola ry¿owe, w p³ytkich zbiornikach wodnych za-nieczyszczonych zwi¹zkami organicznymi. Niezbêd-nym warunkiem jest dostêpno�æ zwi¹zków Fe(III) orazMn(IV) i ³atwo�æ utleniania ich form zredukowanych[61]. Oddychanie przez redukcjê Fe(III) oraz Mn(IV)odgrywa istotn¹ rolê w �rodowiskach ubogich w azota-ny i siarczany, gdzie nie zachodzi metanogeneza [20].
Jednocze�nie mikrobiologiczna redukcja siarczanówjak i procesy metanogenezy s¹ zahamowane w �rodo-wiskach bogatych w Fe(III). Dodanie tlenków Fe(III)lub Mn(IV) do �rodowisk, gdzie intensywnie zachodz¹procesy redukcji siarczanów oraz procesy metanoge-nezy, powoduje ich wyra�ne spowolnienie. Nie wynikato z toksyczno�ci zwi¹zków ¿elaza, tylko ze zmianysk³adu mikroflory i zdominowania jej przez organizmyFRM. Podobny efekt obserwuje siê po wprowadzeniudo pod³o¿a wodoru i/lub octanów. Zjawisko to jestwynikiem wspó³zawodnictwa miêdzy poszczególnymigrupami mikroorganizmów o substraty organiczne bê-d¹ce donorami elektronów. Decyduje o tym potencja³oksydoredukcyjny reakcji redukcji zwi¹zków nieor-ganicznych i utleniania materii organicznej [89]. Jakwidaæ z danych w Tabeli IV prokarioty redukuj¹ceFe(III) i Mn(IV) ¿yj¹ na pograniczu dodatniego i ujem-nego potencja³u oksydoredukcyjnego, uzyskuj¹c przytym dwukrotnie wiêcej energii przy utlenieniu molasubstratu organicznego ni¿ prokarioty redukuj¹ce siar-czany (PRS) oraz archeony metanogenne. FRM s¹ wiêcbardziej istotne ekologicznie.
Ogromne znaczenie dla �rodowiska ma usuwaniezanieczyszczeñ organicznych z terenów ska¿onych rop¹naftow¹ czy wysypisk �mieci przez FRM (rozdz. 5.2).
Mikrobiologiczna transformacja form ¿elaza Fe(II)i Fe(III) pe³ni wa¿n¹ rolê w obiegu ¿elaza w przyro-dzie. Do procesów mikrobiologicznych wp³ywaj¹cychna kr¹¿enie ¿elaza oprócz oddychania Fe(III) i innychform dysymilacyjnej redukcji ¿elaza nale¿¹ oddzia³y-wanie koñcowych produktów metabolizmu bakterii naminera³y zawierajace ¿elazo oraz procesy mikrobiolo-gicznego utleniania jonu ¿elazawego do jonu ¿elazo-wego [20]. FRM maj¹ te¿ istotne znaczenie w kr¹¿e-niu manganu w przyrodzie. Uwa¿a siê, ¿e ilo�æzwi¹zków manganu w �rodowisku stanowi 10% ilo�cizwi¹zków ¿elaza, ale za to Mn(IV) jest chêtniej wyko-rzystwany jako akceptor elektronów ni¿ Fe(III), corównie¿ spowodowane jest potecja³em redoks reakcjiredukcji Fe(III) i Mn(IV) (Tabela IV) [61, 89].
Redukcja tlenu: O2 + 4H+ + 4e� � 2H
2O 0,812 �29,9
Redukcja jonów azotanowych: NO3
� + 6H+ + 6e_ � N2 + 3H
2O 0,747 �28,4
Redukcja jonów Mn+4 do Mn2+: MnO2 + 4H+ + 2e� � Mn2+ + 2H
2O 0,526 �23,3
Redukcja jonów Fe3+ do Fe2+: Fe(OH)3 + 3H+ + e� � Fe2+ + 3H
2O �0,047 �10,1
Redukcja jonów SO4
2� do H2S: SO
42� + 10H+ + 8e� � H
2S + 4H
2O �0,221 �5,9
Redukcja CO2 do CH
4: CO
2 + 8H+ + 8e� � CH
4 + 2H
2O �0,244 �5,6
T a b e l a I VWarto�ci termodynamiczne redukcji nieorganicznych akceptorów elektronów
w obecno�ci materii organicznej [89]
ReakcjaWarto�ci Eh
(V))G
(kcal/mol)

117PROKARIOTY REDUKUJ¥CE FE(III): KLASYFIKACJA, WYSTÊPOWANIE, MECHANIZMY REDUKCJI FE(III)...
5. Znaczenie prokariotów redukuj¹cych Fe(III)w biotechnologii
Biotechnologiê okre�la siê jako integracjê naukprzyrodniczych i in¿ynieryjnych w celu zastosowaniaczynników biologicznych (organizmów, komórek, en-zymów) do pozyskania dóbr i us³ug. W�ród czynni-ków biologicznych wykorzystywanych w biotechno-logii dominuj¹c¹ rolê odgrywaj¹ bakterie. Prokariotyredukuj¹ce Fe(III) oraz inne metale stanowi¹ bardzociekaw¹ grupê drobnoustrojów z punktu widzeniabiotechnologii. Wyizolowane i zidentyfikowane nie-dawno, w przysz³o�ci mog¹ odegraæ du¿¹ rolê w pro-cesach bioremediacji oraz znale�æ zastosowanie w mi-krobiologicznych ogniwach paliwowych do produkcjipr¹du z jednoczesnym usuwaniem ró¿norodnych od-padów organicznych.
5.1. Mikrobiologiczne ogniwa paliwowe
Mikrobiologiczne ogniwa paliwowe (MFC, micro-bial fuel cell) to uk³ady w których wykorzystuje siêmikroorganizmy beztlenowe do produkcji energiielektrycznej. W wyniku oddychania komórkowegonastêpuje przekszta³cenie energii chemicznej ukrytejw wi¹zaniach chemicznych w zwi¹zkach organicznych
w energiê elektryczn¹. MFC dzia³aj¹ na zasadzie che-micznych ogniw paliwowych. W tych ostatnich pro-dukcja energii elektrycznej nastêpuje w wyniku elek-trochemicznej reakcji spalania paliwa (wodoru lubprostego zwi¹zku chemicznego bogatego w wodór, jaknp. metanol czy lekkie wêglowodory) doprowadza-nego do anody w utleniaczu, np. tlenie z powietrzadoprowadzanym do katody. Mikrobiologiczne ogniwopaliwowe sk³ada siê z przedzia³ów anodowego i kato-dowego oddzielonych od siebie membran¹ jonoselek-tywn¹ przepuszczaj¹c¹ jony, zwykle protony. W prze-dziale anodowym maj¹ miejsce reakcje utleniania.Protony przenoszone s¹ z przedzia³u anodowego doprzedzia³u katodowego przez membranê. Elektronyprzenoszone s¹ na katodê przez zewnêtrzny obwódelektryczny, w którym wydziela siê energia elektrycz-na. W przedziale katodowym nastêpuj¹ reakcje reduk-cji. Jak do tej pory zaproponowano kilka typów MFC[11, 55, 94]. Jedna z koncepcji zak³ada, ¿e w przedzialeanodowym MFC hodowane s¹ bakterie redukuj¹ceFe(III), które tworz¹ biofilm na anodzie. W wynikuutleniania przez te bakterie zwi¹zków organicznychw procesie oddychania powstaj¹ elektrony, które prze-noszone s¹ bezpo�rednio na anodê MFC zamiast nanaturalne akceptory znajduj¹ce siê w �rodowiskuzewn¹trzkomórkowym. W efekcie tego procesu pro-dukowana jest energia elektryczna (rys. 3). Bakterie
Rys. 3. Schemat mikrobiologicznego ogni-wa paliwowego z udzia³em bakterii redu-kuj¹cych ¿elazo (III).Na zdjêciach pokazano anodê MFC w po-staci tkaniny wêglowej poro�niêtej bakte-riami z hodowli selekcjonuj¹cej bakterieredukuj¹ce ¿elazo (badania w³asne).

118 ANNA SIKORA I MIECZYS£AW K. B£ASZCZYK
zdolne do produkcji energii w wyniku bezpo�redniegotransferu elektronów pochodz¹cych z utleniania materiiorganicznej na anodê mikrobiologicznego ogniwa pa-liwowego zosta³y nazwane przez jednych autorówelektricigenami (electricigens) [57�60] a przez innychegzoelektrogenami (exoelectrogens) [55]. Do tej poryopisano dzia³anie MFC z udzia³em czystych kulturnastêpuj¹cych gatunków bakterii: Rhodoferax ferrire-ducens [15], Geothrix fermentans [8], Desulfobulbuspropionicus [33], Aeromonas hydrophila [83], Clostri-dium butyricum szczep EG3 [81], Geopsychrobacterelectrodiphilus [35], Geobacter sulfurreducens [6],G. metallireducens, Desulfuromonas acetoxidans [7],S. putrefaciens szczep MR-1 [43]. Najczê�ciej stoso-wanym materia³em, z którego wykonuje siê anodyw tego typu ogniwach jest grafit. W eksperymentachz Geobacter sulfurreducens pokazano, ¿e bardzodobrym materia³em anodowym mo¿e byæ z³oto. Toodkrycie jest wa¿ne dla zastosowañ MFC z udzia³embakterii redukuj¹cych ¿elazo na mikroskalê [88].
Buduje siê równie¿ osadowe mikrobiologiczneogniwa paliwowe (SMFC, sediment MFC). Anoda tegotypu ogniwa umieszczana jest bezpo�rednio w beztle-nowych osadach dennych zbiorników wodnych a ka-toda w miejscach, do których dociera tlen. Osi¹ganagêsto�æ mocy produkowanego pr¹du wynosi od kilkudo kilkudziesiêciu mW/m2. Takie uk³ady zwane te¿BUGs (Benthic Unattened Generators) mog¹ mieæ za-stosowanie jako systemy przeznaczone do zasilaniaurz¹dzeñ elektrycznych w odleg³ych miejscach jakg³êbiny oceanu, gdzie wymiana baterii jest technicz-nie trudna i kosztowna. Osadowe MFC mo¿na te¿ zbu-dowaæ w warunkach laboratoryjnych umieszczaj¹canodê ogniwa w próbkach osadów dennych. Analizybiofilmów pokrywaj¹cych anody osadowych MFCpokaza³y, ¿e ich dominuj¹cym sk³adnikiem s¹ bakte-rie z rodziny Geobacteraceae, rodzaj Desulfuromonasw przypadku s³onych osadów morskich i rodzajGeobacter w przypadku osadów s³odkowodnych.W ró¿nych warunkach znacz¹cy udzia³ maj¹ te¿ innebakterie, np. z rodziny Desulfobulbaceace czy rodzajGeothrix [7, 17, 33, 55, 56, 57, 97]. Osadowe mikro-biologiczne ogniwa paliwowe pozwalaj¹ na odkrywa-nie i izolacjê nowych elektrochemicznie aktywnychgatunków i szczepów bakterii [35, 81, 83].
Obecnie rozwa¿a siê trzy kierunki rozwoju i zasto-sowañ MFC z udzia³em elektricigenów. Pierwszy tousuwanie odpadów organicznych z jednoczesn¹ pro-dukcj¹ energii elektrycznej. Substratami mia³yby tubyæ �cieki pochodz¹ce z gospodarstw domowych czyodpady z produkcji rolnej. Rozwa¿a siê stworze-nie uk³adów, odpowiedników systemów BUGs, którepokrywa³yby czê�ciowo zapotrzebowanie na energiê,zw³aszcza na du¿ych obszarach rolnych oraz w krajachrozwijaj¹cych siê. Drugi kierunek to samowystarczalne
roboty pobieraj¹ce substraty organiczne z otoczenia.Trzeci kierunek to zastosowania na wspomnian¹ ju¿mikroskalê, g³ównie biosensory, czyli systemy, którewykorzystuj¹ reakcje biologiczne do wykrywania ró¿-nych zwi¹zków. Obecno�æ danego zwi¹zku w prze-dziale anodowym uruchamia przep³yw pr¹du, co re-jestrowane jest metodami elektronicznymi [6, 44]. Tepotencjalne zastosowania wymagaj¹ zwiêkszenia wy-dajno�ci i optymalizacji MFC. Konieczne jest wiêcdog³êbne poznanie mechanizmów transferu elektronówna anodê bioogniwa przez znane ju¿ mikroorganizmyzwane elektricigenami, ich fizjologii, ekologii orazposzukiwanie nowych gatunków. Narzêdzi ku temudostarcza obecny burzliwy rozwój genomiki i ró¿no-rodnych metod biologii molekularnej. Szuka siê spo-sobu zwiêkszenia aktywno�ci oddechowej bakteriipokrywajacych anodê. Rozwa¿ana jest koncepcja mo-dyfikacji genetycznych bakterii w kierunku konstruk-cji mutantów np. nadprodukuj¹cych bia³ka, odpowie-dzialnych za kontakt z powierzchni¹ anody [56�59].
Wydaje siê, ¿e nie by³o i nie istnieje ¿adna presjaewolucyjna w kierunku produkcji energii przez elek-tricigeny w naturalnych �rodowiskach. Nie wiadomoczy bia³ka odpowiedzialne za przenoszenie elektro-nów na akceptory znajduj¹ce siê na zewn¹trz komórkiw naturalnym �rodowisku i w warunkach laboratoryj-nych dzia³aj¹ równie wydajnie, gdy bakteria ro�nie namateriale, z którego zbudowana jest anoda MFC [59].Stosuj¹c technikê mikroczipów DNA oraz hybrydy-zacjê northern porównywano ekspresjê genów G. sul-furreducens rosn¹cych na pod³o¿u z cytrynianem ¿e-laza oraz na anodzie grafitowej w mikrobiologicznymogniwie paliwowym. Stwierdzono zmienion¹ ekspre-sjê 474 genów, gdy elektroda by³a jedynym akcepto-rem elektronów. 197 genów by³o transkrybowanychsilniej a 277 s³abiej. Najwieksz¹ ró¿nicê, bo a¿ 19-krot-ny wzrost ekspresji, obserwowano dla genu koduj¹-cego bia³ko OmcS, cytochrom typu c w zewnêtrznejb³onie komórkowej. Co wiêcej, mutant z delecj¹ omcSnie by³ zdolny do produkcji energii. Podwy¿szonejekspresji podlega³y te¿ geny omcE i omcT. Przypom-nijmy, ¿e bia³ka OmcS i OmcE s¹ odpowiedzialne zaredukcjê nierozpuszczalnego tlenku ¿elazowego. ¯ad-nych ró¿nic nie obserwowano natomiast dla transkryp-tów omcB, ompB, ompJ oraz piliny buduj¹cej nanodru-ty. Ponadto mutacje w genach omcB i pilA nie zmieni³yprodukcji energii elektrycznej. Cytochromy typu c zlo-kalizowane w zewnêtrznej b³onie komórkowej prawdo-podobnie odgrywaj¹ istotn¹ rolê w kontaktach komórkibakterii z elektrod¹ [36].
Wyniki innych badañ sugeruj¹, ¿e równie¿ innebia³ka mog¹ byæ istotne w transferze elektronów nanaturalne akceptory w �rodowisku ni¿ na anodêmikrobiologicznego ogniwa paliwowego. Otrzyma-no mutanty S. oneidensis MR-1, które produkowa³y

119PROKARIOTY REDUKUJ¥CE FE(III): KLASYFIKACJA, WYSTÊPOWANIE, MECHANIZMY REDUKCJI FE(III)...
pr¹d w MFC, ale nie by³y w stanie redukowaæ tlenkuFe(III) [10]. Z kolei inna bakteria z rodziny Geobac-teraceae, Pelobacter carbinolicus jest zdolna do re-dukcji ¿elaza (III), ale w MFC z jej udzia³em nie by³produkowany pr¹d [87].
5.2. Bioremediacja
Terminem bioremediacja okre�la siê technologiêleczenia �rodowiska naturalnego ze ska¿eñ, g³ówniepowodowanych przez metale ciê¿kie, produkty ropynaftowej i pierwiastki radioaktywne, za pomoc¹ ¿y-wych mikroorganizmów w celu ich rozk³adu lub prze-prowadzenia w formy mniej szkodliwe. Bioremediacjain situ w³¹cza procesy degradacji, detoksyfikacji orazimmobilizacji zanieczyszczeñ przez mikroorganizmy,które naturalnie ¿yj¹ w strefach ska¿onych [60].
FRM mog¹ znale�æ zastosowanie w procesach bio-remediacji na dwa sposoby. Pierwszy to utlenianie dodwutlenku wêgla w warunkach beztlenowych zwi¹z-ków organicznych, jak np. wêglowodory aromatycznew miejscach ska¿onych rop¹ naftow¹, po³¹czone z re-dukcj¹ ¿elaza. Tereny ska¿one przez ropê naftow¹ obfi-tuj¹ w FRM, g³ównie rodzaj Geobacter. Istnieje pomys³zastosowania uk³adów opartych na mikrobiologicz-nych ogniwach paliwowych do oczyszczania terenówska¿onych ropopochodnymi zwi¹zkami organicznymi.W wyniku utleniania tych zwi¹zków przez bakteriez grupy FRM produkowana by³aby energia elektryczna[76]. Dodanie substancji chelatuj¹cych nierozpuszczal-ne tlenki Fe(III) do próbek osadów zanieczyszczonychrop¹ naftow¹ powodowa³o przyspieszenie procesuutleniania ropopochodnych zwi¹zków aromatycznychi procesów redukcji Fe(III). Mo¿e to byæ strategi¹ sto-sowan¹ w bioremediacji terenów ska¿onych rop¹ naf-tow¹ przy udziale bakterii z grupy FRM [70].
Drugi sposób zastosowania bakterii redukuj¹cych¿elazo w procesach bioremediacji to wykorzystanieich zdolno�ci do redukcji innych metali ni¿ Fe(III) czyMn (IV), zw³aszcza tych, które s¹ toksyczne i nie-bezpieczne dla cz³owieka i �rodowiska. Wiêkszo�æpierwiastków mo¿e wystêpowaæ na ró¿nym stopniuutlenienia w ró¿nych zwi¹zkach chemicznych. Stopieñutlenienia, na którym znajduje siê dany pierwiastekwp³ywa na w³asno�ci fizyko-chemiczne (np. rozpusz-czalno�æ w wodzie) zwi¹zków, które tworzy. Szcze-gólne znaczenie z punktu widzenia bioremediacji maj¹procesy (czêsto s¹ to procesy redukcji) przeprowadza-nia form toksycznych pierwiastków rozpuszczalnychw wodzie w formy nierozpuszczalne, które ulegaj¹wytr¹caniu i w ten sposób ³atwo je usun¹æ z miejscska¿onych.
FRM mog¹ byæ skutecznym narzêdziem do bio-remediacji terenów ska¿onych uranem pochodz¹cymz odpadów radioaktywnych. Okres pó³trwania uranu
w zale¿no�ci od izotopu wynosi od 247 tysiêcy do4.5 miliardów lat. Zwi¹zki uranu (VI) s¹ rozpuszczal-ne w wodzie, st¹d bardzo trudne do usuniêcia ze �ro-dowiska. Natomiast uran (IV) tworzy nierozpuszczal-ne tlenki � uraninit, które ³atwo ulegaj¹ wytr¹ceniu.Zatem mikrobiologiczna redukcja U (VI) do U (IV)mo¿e przyczyniæ siê do usuwania uranu ze �rodo-wiska, zw³aszcza ze ska¿onych wód [2, 63, 64, 96].Czyste kultury bakterii z grupy FRM (rodzaj Geobac-ter, Shewanella) redukuj¹ U(VI) do U(IV) i w ten spo-sób pozyskuj¹ energiê niezbêdn¹ do procesów ¿ycio-wych. Reakcje 19 i 20 opisuj¹ redukcjê U(VI) doU(IV) przy udziale elektronów pochodz¹cych z utle-niania odpowiednio octanu i wodoru.
CH3COO� (octan) + 4U(VI) ++ 4H2O → 2HCO3
� + 4U(IV) + 9H+ + (reakcja 19)H2 + U(VI) → 2H+ + U(IV) (reakcja 20)
Do redukcji uranu (VI) zdolne te¿ s¹ prokariotyredukuj¹ce siarczany, jednak proces ten nie s³u¿y po-zyskiwaniu energii. Dotychczas stwierdzono, ¿e zdol-no�æ redukcji uranu (VI) do uranu (IV) posiadaj¹ bak-terie z rodziny Desufovibrionaceae, np. Desulfovibriodesulfuricans, a tak¿e nieliczni przedstawiciele z ro-dziny Peptococcaceae, np. rodzaj Desulfotomaculum.Filogenetyczna analiza populacji bakterii w próbkachpobranych z osadów ska¿onych uranem pokazuje, ¿edominuj¹ tam przedstawiciele rodzin Geobacteraceaei Desufovibrionaceae [96].
W badaniach laboratoryjnych pokazano, ¿e prost¹metod¹ zwiêkszania procesów redukcji uranu (VI)w ska¿onych próbkach pobranych z naturalnych �ro-dowisk wodnych jest dodanie octanu jako dawcy elek-tronów. W takich warunkach nastêpuje silny rozwójFRM, g³ównie z rodziny Geobacteraceae, które sta-nowi¹ nawet do 40% ca³ego zespo³u mikroorgani-zmów (microbial community) [32]. Tak¹ sam¹ strate-giê (wzbogacanie w octan) zastosowano w badaniachin situ w zbiorniku wodnym zanieczyszczonym zwi¹z-kami uranu. Do�wiadczenie trwa³o 3 miesi¹ce. Pocz¹t-kowo równie¿ obserwowano rozwój bakterii z rodzi-ny Geobacteraceae i wzmo¿one wytr¹canie uraninitu.Stwierdzono 70% obni¿enie zawarto�ci U(VI). Jednakpo pewnym czasie dominuj¹c¹ grup¹ sta³y siê bakterieredukuj¹ce siarczany, które utlenia³y octan. Spowo-dowane to by³o g³ównie wyczerpaniem siê zwi¹zków¿elaza Fe(III). W efekcie nast¹pi³ wzrost ilo�ci uranu(VI) w badanym �rodowisku. Bakterie redukuj¹cesiarczany równie¿ redukowa³y uran (VI) do U(IV), aleznacznie mniej efektywnie w porównaniu do stanu, gdyw badanym �rodowisku dominowa³a rodzina Geobac-teraceae [2]. W trzecim miesi¹cu opisanego ekspery-mentu pobrano próbkê osadu dennego i wyizolowanoz niej nowy gatunek nale¿¹cy do rodzaju Geobacter� Geobacter uraniireducens [91].

120 ANNA SIKORA I MIECZYS£AW K. B£ASZCZYK
W obecno�ci azotanów nie obserwuje siê reduk-cji U(VI) do U(IV) zarówno w badaniach in situ jaki w warunkach laboratoryjnych w czystych hodowlachG. metallireducens oraz w próbkach pobranych z ró¿-nych miejsc ska¿onych uranem. We wszystkich tychuk³adach redukowany jest jon azotanowy [21, 23, 90].Istnieje kilka przyczyn tego zjawiska. Po pierwsze, re-akcja redukcji jonu azotanowego jest bardziej korzyst-na termodynamicznie dla bakterii redukuj¹cych ¿ela-zo zdolnych te¿ do redukcji NO3
� ni¿ reakcja redukcjiuranu (VI). Po drugie, w naturalnych �rodowiskachredukcja metali jest energetycznie mniej korzystn¹ for-m¹ oddychania beztlenowego w porównaniu z reduk-cj¹ azotanów i konkurencjê wygrywaj¹ bakterie deni-tryfikacyjne. Po trzecie, zredukowane formy U(IV)podobnie jak Fe(II) mog¹ byæ utleniane w obecno�ciazotanów przez bakterie denitryfikacyjne, a nawet przezsame FRM. Poza tym utlenianie zarówno U (IV) oraz¿elaza Fe(II) mo¿e zachodziæ abiotycznie [23].
Bardzo wa¿nym odkryciem z punktu widzenia za-stosowania rodzaju Geobacter do bioremediacji tere-nów ska¿onych uranem (VI) by³o wykazanie, ¿e dawc¹elektronów do procesów redukcji prowadzonych przezte bakterie mo¿e byæ grafitowa elektroda [29]. Takieelektrody pokryte biofilmem bakteryjnym umieszczanow miejscach ska¿onych uranem (VI). Stanowi³y one�ród³o elektronów do mikrobiologicznej reakcji re-dukcji U(VI) do U(IV). Takie rozwi¹zanie znacznieu³atwia usuwanie wytr¹conego uraninitu ze �rodowiska,gdy¿ osadza siê on na powierzchni elektrod [30].
Produktem rozszczepienia uranu jest technet 99Tc.Wystêpuje w du¿ych ilo�ciach w miejscach testowaniabroni j¹drowej oraz tam, gdzie sk³adowane s¹ odpadyradioaktywne. Okres pó³trwania technetu 99Tc jest bar-dzo d³ugi i wynosi 213 tysiêcy lat. W �rodowisku Tcwystêpuje na +7 stopniu utlenienia w postaci jonuTcO4
�, który mo¿e byæ w³¹czany w ³añcuch pokar-mowy jako analog siarczanów. Technet Tc(VII) mo¿eulegaæ mikrobiologicznej redukcji, w wyniku czegopowstaje nierozpuszczalny tlenek TcO2, w którym tech-net wystêpuje na +4 stopniu utlenienia. W takiej posta-ci technet mo¿e byæ stosunkowo ³atwo usuniêty ze �ro-dowiska. Pokazano dwa mechanizmy redukcji Tc(VII)do Tc(IV) dla G. sulfurreducens. W pierwszym technetredukowany jest enzymatycznie przy jednoczesnymutlenianiu wodoru. Uwa¿a siê, ¿e funkcj¹ reduktazyTc(VII) pe³ni hydrogenaza. Drugi mechanizm zak³a-da, ¿e Tc(VII) mo¿e ulegaæ redukcji do Tc(IV) po�red-nio w sposób abiotyczny przez zredukowane kwasyhumusowe oraz magnetyt. Wytr¹cony TcO2 osadza siêna powierzchni kryszta³ów magnetytu. Do redukcjitechnetu oprócz G. sulfurreducens zdolny jest tak¿eG. metallireducens i inny przedstawiciel grupy FRM,S. putrefaciens [51], a tak¿e E. coli (mechanizm reduk-cji opisany w rozdz. 3) [52, 53] czy D. desulfuricans.
W przypadku tej ostatniej, gdy donorem elektronówby³ wodór lub mrówczan, a akceptorem elektronówjon TcO4
� zamiast siarczanu w warunkach beztleno-wych, nastêpowa³o utlenienie wodoru lub mrówczanuz jednoczesn¹ redukcj¹ TcO4
� do nierozpuszczalnegosiarczku Tc. Wynik ten wskazywa³ na enzymatyczn¹redukcjê Tc(VII), któr¹ przeprowadza peryplazma-tyczna hydrogenaza. Oprócz tego jon TcO4
� mo¿e byæredukowany abiotycznie do siarczku technetu w reak-cji z siarkowodorem, powsta³ym w wyniku aktywno�cibakterii redukuj¹cych siarczany [53, 54].
Cz³onkowie rodzin Geobacteraceae oraz Shewa-nellaceae s¹ modelowymi organizmami w badaniachnad zastosowaniem bakterii redukuj¹cych ¿elazo dobioremediacji terenów ska¿onych uranem czy metala-mi ciê¿kimi. Jak wiadomo bakterie z rodzin Geobac-teraceae oraz Shewanellacae s¹ neutrofilami. Podda-no zatem analizie konsorcja redukuj¹ce ¿elazo i uran,z podpowierzchniowych osadów z terenów ska¿onychuranem i azotanami, o niskim pH. Metod¹ analizysekwencji genu koduj¹cego 16S rRNA z DNA po-chodz¹cego z pobranych próbek osadów wykazanoznaczny udzia³ fakultatywnych beztlenowców zdol-nych do redukcji ¿elaza nale¿¹cych do rodzaju Anaero-myxobacter [82].
Wykazano równie¿, ¿e przedstawiciel rodziny Geo-bacteraceae, G. metallireducens, jest zdolny do pozys-kania energii w wyniku utleniania materii organicznejw po³¹czeniu z redukcj¹ wanadu V(V) do V(IV). Wa-nad jest naturalnie wystêpuj¹cym metalem skorupyziemskiej w ilo�ciach podobnych do cynku. Do �rodo-wiska dostaje siê równie¿, w ilo�ciach powoduj¹cychjego ska¿enie, wraz z odpadami z przemys³u wydobyw-czego, metalurgicznego, farmaceutycznego oraz nowo-czesnych technologii. Wanad mo¿e wystêpowaæ w �ro-dowisku na +3, +4 i +5 stopniu utlenienia. Najbardziejtoksyczn¹ form¹ jest V(V). Zwi¹zki V(V) s¹ rozpusz-czalne w wodzie. Wanad na stopniu utlenienie +4 jestnierozpuszczalny w wodzie. Wystêpuje w postaci ka-tionów VO2+, które ulegaj¹ adsorpcji na cz¹steczkachkwasów humusowych i tworz¹ z nimi trwa³e kom-pleksy. Zatem przekszta³cenie formy V(V) do V(IV)mo¿e stanowiæ metodê immobilizacji wanadu a na-stêpnie usuwanie go ze ska¿onych terenów [80].
Bakterie z rodzaju Geobacter oraz bakterie ekstre-mofilne jak Pyrobaculum islandicum mog¹ te¿ redu-kowaæ Hg(II) do Hg(0), Cr(VI) do Cr(III), czy Co(III)do Co(II) [49, 64].
Redukcja metali niezwi¹zana z pozyskiwaniemenergii niezbêdnej do ¿ycia jest do�æ szeroko roz-powszechniona w�ród bakterii i odgrywa du¿¹ rolêw kr¹¿eniu ró¿nych metali w przyrodzie. Procesy temaj¹ szczególnie istotne znaczenie dla �rodowiskaz powodu przekszta³cania ró¿nych toksycznych metaliw formy mniej toksyczne, ³atwe do usuniêcia. Do

121PROKARIOTY REDUKUJ¥CE FE(III): KLASYFIKACJA, WYSTÊPOWANIE, MECHANIZMY REDUKCJI FE(III)...
takich metali nale¿¹: selen [redukcja Se(VI), Se(IV),Se(0) przez ró¿ne gatunki Clostridium, Citrobacter,Flavobacterium, Pseudomonas], chrom [redukcjaCr(VI) do Cr (III) przez ró¿ne gatunki Pseudomonas,Bacillus oraz Aeromonas dechromatica, Achromobac-ter eurydice, Micrococcus roseus, Escherichia coli,Enterobacter cloacae, Desulfovibrio desulfuricans],rtêæ [redukcja Hg(II) do Hg(0)], technet [redukcjaTc(VII) do Tc(IV) przez rodzaj Moraxella i Planococ-cus], wanad [redukcja V(V) do V(IV) przez gatunkiPseudomonas], molibden [redukcja Mo(V) do Mo(IV)przez Pseudomonas guillermondii, Micrococcus sp.,Acidithiobacillus ferrooxidans, gatunki Sulfolobus],mied� [redukcja Cu(II) do Cu(I) przez A. ferrooxidans],palad [redukcja Pd(II) do Pd(0) przez Desulfovibriodesulfuricans] [64].
Zdolno�æ mikroorganizmów do redukcji metali za-równo maj¹cej na celu produkcjê energii (G. metalli-reducens) jak i bêd¹cej procesem ubocznym (E. coli,D. desulfuricans, B. subtilis, P. aeruginosa) mo¿nate¿ wykorzystaæ do otrzymywania cennych kruszcówjakimi s¹ z³oto i srebro. W wyniku redukcji rozpusz-czalnych form Au(III) lub Au(I) powstaje z³oto meta-liczne, a Ag(I) metaliczne srebro [18, 64].
6. Podsumowanie
Wyniki dotychczasowych badañ nad FRM wska-zuj¹, ¿e energia pozyskiwana do wzrostu i rozwojuz dysymilacyjnej redukcji Fe(III) oraz Mn(IV) i innychmetali wp³ywa istotnie na cykle biogeochemicznew �rodowiskach beztlenowych. FRM tworz¹ �super-rodzinê�, do której nale¿¹ ró¿norodne taksony z do-meny Bacteria jak i domeny Archaea. FRM s¹ tak¿ewa¿nym czynnikiem w procesie bioremediacji za-równo zanieczyszczeñ organicznych oraz metalamiciê¿kimi i pierwiastkami promieniotwórczymi w bez-tlenowych warstwach �rodowisk mezofilnych jak i psy-chrofilnych, termofilnych i hypertermofilnych. FRMznalaz³y równie¿ zastosowanie w MFC jako czynnikprodukuj¹cy energiê elektryczn¹. FRM wykszta³ci³yszereg mechanizmów umo¿liwiaj¹cych redukcjê roz-puszczalnych i nierozpuszczalnych zwi¹zków Fe(III),Mn(IV) i innych metali.
7. Pi�miennictwo
1. Afkar E., Reguera G., Schiffer M., Lovley D.R.: A novelGeobacteraceae-specific outer membrane protein J (OmpJ) isessential for electron transport to Fe(III) and Mn(IV) oxidesin Geobacter sulfurreducens. BMC Microbiol. 5, 41 (2005)
2. Anderson R.T., Lovley D.R. i wsp.: Stimulating the in situactivity of Geobacter species to remove uranium fromthe groundwater of a uranium-contaminated aquifer. Appl.
Environ. Microbiol. 69, 5884�5891 (2003) (wy¿ej cytowanapraca jest dzie³em 13 autorów)
3. Beliaev A.S., Saffarini D.A., McLaughlin J.M., Hunnicut D.:MtrC, an outer membrane decahaem c cytochrome requiredfor metal reduction in Shewanella putrefaciens MR-1. Mol.Microbiol. 39, 722�730 (2001)
4. Beliaev A.S., Saffarini D.A.: Shewanella putrefaciens mtrBencodes an outer membrane protein required for Fe(III) andMn(IV) reduction. J. Bacteriol. 180, 6292�6297 (1998)
5. Bergey�s Manual of Systematic Bacteriology, 2nd Edition,Springer, New York 2001/2005
6. Bond D.R., Lovley D.R.: Electricity production by Geobac-ter sulfurreducens attached to electrodes. Appl. Environ.Microbiol. 69, 1548�1555 (2003)
7. Bond D.R., Holmes D.E., Tender L.M., Lovley D.R.: Elec-trode-reducing microorganisms that harvest energy frommarine sediments. Science, 295, 483�485 (2002)
8. Bond D.R., Lovley D.R.: Evidence for involvement of anelectron shuttle in electricity generation by Geothrix fermen-tas. Appl. Environ. Microbiol. 71, 2186�2189 (2005)
9. Boone D.R., Liu Y., Zhao Z.J., Balkwill D.L., Drake G.R.,Stevens T.O., Aldrich H.C.: Bacillus infernus sp. nov.,an Fe(III)- and Mn(IV)-reducing anaerobe from the deep ter-restrial subsurface. Int. J. System. Bacteriol. 45, 441�448(1995)
10. Bretschger O., K.H. Nealson i wsp.: Current production andmetal oxide reduction by Shewanella oneidensis MR-1 wildtype and mutants. Appl. Environ. Microbiol. 73, 7003�7011(2007) (wy¿ej cytowana praca jest dzie³em 18 autorów)
11. Bullen R.A., Arnot T.C., Lakeman J.B., Walsh F.C.: Bio-fuel cells and their development. Biosens. Bioelectron. 21,2015�2045 (2006)
12. Butler J.E., Kaufmann F., Coppi M.V., Nunez C., Lovley D.R.:MacA, A diheme c-type cytochrome involved in Fe(III)reduction by Geobacter sulfurreducens. J. Bacteriol. 186,4042�4045 (2004)
13. Caccavo F., Coates J.D., Rossello-Mora R.A., Ludwig W.,Schleifer K.H., Lovley D.R., McInerney M.J.: Geovibrio ferri-reducens, a phylogenetically distinct dissimilatory Fe(III)-re-ducing bacterium. Arch. Microbiol. 165, 370�376 (1996)
14. von Canstein H., Ogawa J., Shimizu S., Lloyd J.R.: Secretionof flavins by Shewanella species and their role in extracellularelectron transfer. Appl. Environ. Microbiol. 74, 615�623(2008)
15. Chaudhuri S.K., Lovley D.R.: Electricity generation by directoxidation of glucose in mediatorless microbial fuel cells.Nature Biotechnol. 21, 1229�1232 (2003)
16. Cummings D.E., Caccavo F., Spring S., Rosenzweig R.F.:Ferribacterium limneticum, gen. nov., sp. nov., an Fe(III)-reducing microorganism isolated from mining-impactedfreshwater lake sediments. Arch. Microbiol. 171, 183�188(1999)
17. DeLong E.F., Chandler P.: Power from the deep. Nature Bio-technol. 20, 788�789 (2002)
18. Deplanche K., Macaskie L.E.: Biorecovery of gold by Esche-richia coli and Desulfovibrio desulfuricans. Biotechnol.Bioeng. 99, 1055�1064 (2008)
19. DiChristina T.J., Moore C.M., Haller C.A.: DissimilatoryFe(III) and Mn(IV) reduction by Shewanella putrefaciensrequires ferA, a homolog of the pulE (gspE) type II proteinsecretion gene. J. Bacteriol. 184, 142�151 (2002)
20. Ehrlich H.L., Geomicrobiology of iron (w) Geomicrobio-logy, red. H.L. Ehrlich, Marcel Dekker, Inc. New York, Basel,2002, s. 345.

122 ANNA SIKORA I MIECZYS£AW K. B£ASZCZYK
21. Elias D.A., Krumholz L.R., Wong D., Long P.E., Suflita J.M.:Characterization of microbial activities and U reduction ina shallow aquifer contaminated by uranium mill tailings.Microb. Ecol. 46, 83�91 (2003)
22. Esteve-Nunez A., Sosnik J, Visconti P., Lovley D.R.:Fluorescent properties of c-type cytochromes reveal theirpotential role as an extracytoplasmic electron sink in Geo-bacter sulfurreducens. Environ. Microbiol. 10, 497�505(2008)
23. Finneran K.T., Housewright M.E., Lovley D.R.: Multipleinfluences of nitrate on uranium solubility during bioreme-diation of uranium-contaminated subsurface sediments. Env.Microbiol. 4, 510�516 (2002)
24. Francis C.A., Obraztsova A.Y., Tebo B.M.: Dissimilatorymetal reduction by the facultative anaerobe Pantoea agglo-merans SP1. Appl. Environ. Microbiol. 66, 543�548 (2000)
25. Fredrickson J.K., Kostandarithes H.M., Li S.W., Plymale A.E.,Daly M.J.: Reduction of Fe(III), Cr(VI), U(VI), and Tc(VII)by Deinococcus radiodurans R1. Appl. Environ. Microbiol.66, 2006�2011 (2000)
26. Glasauer S., Langley S., Boyanov M., Lai B., Kemner K.,Beveridge T.J.: Mixed-valence cytoplasmic iron granules arelinked to anaerobic respiration. Appl. Environ. Microbiol. 73,993�996 (2007)
27. Glasauer S., Langley S., Beveridge T.J.: Intracellular man-ganese granules formed by a subsurface bacterium. Environ.Microbiol. 6, 1042�1048 (2004)
28. Gorby Y.A., Fredrickson J.K. i wsp.: Electrically conductivebacterial nanowires produced by Shewanella oneidensisstrain MR-1 and other microorganisms. Proc. Natl. Acad. Sci.USA, 103, 11358�11363 (2006) (wy¿ej cytowana praca jestdzie³em 24 autorów)
29. Gregory K.B., Bond D.R., Lovley D.R.: Graphite electrodesas electron donors for anaerobic respiration. Environ. Micro-biol. 6, 596�604 (2004)
30. Gregory K.B., Lovley D.R.: Remediation and recovery ofuranium from contaminated subsurface environments withelectrodes. Environ. Sci. Technol. 39, 8943�8947 (2005)
31. Heidelberg J.F. C.M. Fraser i wsp.: Genome sequence ofthe dissimilatory metal iron-reducing bacterium Shewanellaoneidensis. Nature Biotechnol. 20, 1118�1123 (2002) (wy¿ejcytowana praca jest dzie³em 43 autorów)
32. Holmes D.E., Finneran K.T., O�Neil R.A., Lovley D.R.:Enrichment of Geobacteraceae associated with stimulationof dissimilatory metal reduction in uranium-contaminatedaquifer sediments. Appl. Environ. Microbiol. 68, 2300�2306(2002)
33. Holmes D.E., Bond D.R., Lovley D.R.: Electron transfer byDesulfobulbus propionicus to Fe(III) and graphite electrodes.Appl. Environ. Microbiol. 70, 1234�1237 (2004)
34. Holmes D.E., Bond D.R., O�Neil R.A., Reimers C.E., Ten-der L.R., Lovley D.R.: Microbial communities associatedwith electrodes harvesting electricity from a variety of aquaticsediments. Microb. Ecol. 48, 178�190 (2004)
35. Holmes D.E., Nicoll J.S., Bond D.R., Lovley D.R.: Potentialrole of a novel psychrotolerant member of the family Geobac-teraceae, Geopsychrobacter electrodiphilus gen. nov., sp.nov., in electricity production by a marine sediment fuel cell.Appl. Environ. Microbiol. 70, 6023�6030 (2004)
36. Holmes D.E., Chaudhuri S.K., Nevin K.P., Mehta T.,Methe B.A., Liu A., Ward J.E., Woodard T.L., Webster J.,Lovley D.R.: Microarray and genetic analysis of electrontransfer to electrodes in Geobacter sulfurreducens. Environ.Microbiol. 8, 1805�1815 (2006)
37. Kanso S., Greene A.C., Patel B.K.C.: Bacillus subterraneussp. nov., an iron- and manganese-reducing bacterium froma deep subsurface Australian thermal aquifer. Int. J. System.Evol. Microbiol. 52, 869�874 (2002)
38. Kashefi K., Holmes D.E., Baross J.A., Lovley D.R.: Ther-mophily in the Geobacteraceae: Geothermobacter ehrlichiigen. nov., sp. nov., a novel thermophilic member of theGeobacteraceae from the �Bag City� hydrothermal vent.Appl. Environ. Microbiol. 69, 2985�2993 (2003)
39. Kashefi K., Holmes D.E., Lovley D.R., Tor J.M.: Potentialimportance of dissimilatory Fe(III)-reducing microorganismsin hot sedimentary environments (w) The Subseafloor Bio-sphere at Mid-Ocean Ridges, Geophysical Monograph Series144, 2004, str. 199.
40. Kashefi K., Moskowitz B.M., Lovley D.R.: Characterizationof extracellular minerals produced during dissimilatoryFe(III) and U(VI) reduction at 100ºC by Pyrobaculum islan-dicum. Geobiology, 6, 147�154 (2008)
41. Kim B.H., Qian X., Leang C., Coppi M.V., Lovley D.R.: Twoputative c-type multiheme cytochromes required for theexpression of OmcB, an outer membrane protein essentialfor optimal Fe(III) reduction in Geobacter sulfurreducens.J. Bacteriol. 188, 3138�3142 (2006)
42. Kim B.H., Leang C., Ding Y.-H. R., Glaven R.H., Coppi M.V.,Lovley D.R.: OmcF, a putative c-type monoheme outer mem-brane cytochrome required for the expression of outer mem-brane cytochromes in Geobacter sulfurreducens. J. Bacteriol.187, 4505�4513 (2005)
43. Kim H.J, Park H.S., Hyun M.S., Chang I.S., Kim M.,Kim B.H.: A mediator-less microbial fuel cell using a metalreducing bacterium, Shewanella putrefaciens. Enzyme Microb.Technol. 30, 145�152 (2002)
44. Kim H.J, Hyun M.S., Chang I.S., Kim B.H.: A MicrobialFuel Cell Type Lactate Biosensor Using a Metal-ReducingBacterium, Shewanella putrefaciens. J. Microbiol. Biotechnol.9, 365�367 (1998)
45. Kusel K., Dorsch T., Acker G., Stackebrandt E.: Microbialreduction of Fe(III) in acidic sediments: isolation of Acidi-philum cryptum JF-5 capable of coupling the reduction ofFe(III) to the oxidation of glucose. Appl. Environ. Microbiol.65, 3633�3640 (1999)
46. Leang C, Coppi M.V., Lovley D.R.: OmcB, a c-type poly-heme cytochrome, involved in Fe(III) reduction in Geobactersulfurreducens. J. Bacteriol. 185, 2096�2103 (2003)
47. Lies D.P., Hernandez M.E., Kappler A., Mielke R.E.,Gralnick J.A., Newman D.K.: Shewanella oneidensis MR-1uses overlapping pathways for iron reduction at a distanceand by direct contact under conditions relevant for biofilms.Appl. Environ. Microbiol. 71, 4414�4426 (2005)
48. Lin W.C., Coppi M.V., Lovley D.R.: Geobacter sulfur-reducens can grow with oxygen as a terminal electron accep-tor. Appl. Environ. Microbiol. 70, 2525�2528 (2004)
49. Lloyd J.R., Lovley D.R.: Microbial detoxification of metalsand radionuclides. Curr. Opin. Biotechnol. 12, 248�253(2001)
50. Lloyd J.R., Leang C., Hodges-Myerson A.L., Coppi M.V.,Cuifo S., Methe B., Sandler S.J., Lovley D.R.: Biochemicaland genetic characterization of PPcA, a periplasmic c-typecytochrome in Geobacter sulfurreducens. Biochem. J. 369,153�161 (2003)
51. Lloyd J.R., Sole V.A., van Praagh C.V.G., Lovley D.R.:Direct and Fe(II)-mediated reduction of technetium byFe(III)-reducing bacteria. Appl. Environ. Microbiol. 66,3743�3749 (2000)

123PROKARIOTY REDUKUJ¥CE FE(III): KLASYFIKACJA, WYSTÊPOWANIE, MECHANIZMY REDUKCJI FE(III)...
52. Lloyd J.R., Cole J.A., Macaskie L.E.: Reduction and removalof heptavalent technetium from solution by Escherichia coli.J. Bacteriol. 179, 2014�2021 (1997)
53. Lloyd J.R., Thomas G.H., Finlay J.A., Cole J.A., MacaskieL.E.: Microbial reduction of technetium by Escherichia coliand Desulfovibrio desulfuricans: Enhancement via the useof high-activity strains and effect of process parameters. Bio-technol. Bioeng. 66, 122�130 (1999)
54. Lloyd J.R., Nolting H.-F., Sole V.A., Bosecker K., MacaskieL.E.: Technetium reduction and precipitation by sulfate-reducing bacteria. Geomicrobiol. J. 15, 45�58 (1998)
55. Logan B.E.: Microbial fuel cells, red. B.E. Logan, John Wiley& Sons, Hoboken, New Jersey, USA, 2008.
56. Lovley D.R.: Microbial energizers:fuel cells that keep ongoing. Microbe, 1, 323�329 (2006)
57. Lovley D.R.: Bug juice: harvesting electricity with micro-organisms. Nat. Rev. Microbiol. 4, 497�508 (2006)
58. Lovley D.R.: Microbial fuel cells: novel microbial physio-logies and engineering approaches. Curr Opin. Biotechnol. 17,327�332 (2006)
59. Lovley D.R.: Taming electricigens. The Scientist, July 2006,46
60. Lovley D.R.: Anaerobes to the rescue. Science, 293, 1444�1446(2001)
61. Lovley D.R., Fe(III) and Mn(IV) reduction (w) Environmen-tal Microbe-Metal Interactions, red. D.R. Lovley. ASM PressWashington, D.C., 2000, s. 3.
62. Lovley D.R.: Dissimilatory Fe(III) and Mn(IV) reduction.Microbiol. Rev. 55, 259�287 (1991)
63. Lovley D.R.: Dissimilatory reduction of iron and uranium(w) Trends in Microbial Ecology, red. R. Guerrero i C. Pe-dros-Alio. Spanish Society for Microbiology Barcelona,1993, s. 71.
64. Lovley D.R.: Dissimilatory metal reduction. Annu. Rev.Microbiol. 47, 263�290 (1993)
65. Lovley D.R., Philips E.P.: Reduction of chromate by Desulfo-vibrio vulgaris and its c
3 cytochrome. Appl. Environ. Micro-
biol. 60, 726�728 (1994)66. Lovley D.R., Philips E.J.P., Lonergan D.J., Widman P.K.:
Fe(III) and S0 reduction by Pelobacter carbinolicus. Appl.Environ. Microbiol. 61, 2132�2138 (1995)
67. Lovley D.R., Holmes D.E., Nevin K.P.: Dissimilatory Fe(III)and Mn(IV) reduction. Adv. Microbiol. Physiol. 49, 219�286(2004)
68. Lovley D.R., Fraga J.L., Coates J.D., Blunt-Harris E.L.:Humics as an electron donor for anaerobic respiration. Envi-ron. Microbiol. 1, 89�98 (1999)
69. Lovley D.R., Fraga J.L., Blunt-Harris E.L., Hayes L.A.,Phillips E.J., Coates D.J.: Humic substances as a mediatorfor microbially catalyzed metal reduction. Acta Hydrochim.Hydrobiol. 26, 152�157 (1998)
70. Lovley D.R., Woodward J.C., Chapelle F.H.: Rapid anaerobicbenzene oxidation with a variety of chelated Fe(III) forms.Appl. Environ. Microbiol. 62, 288�291 (1996)
71. Maier T.M., Myers C.R.: The outer membrane proteinOmp35 affects the reduction of Fe(III), nitrate, and fumarateby Shewanella oneidensis MR-I. BMC Microbiol. 4, 23 (2004)
72. Mattick J.S.: Type IV pili and twiching motility. Annu. Rev.Microbiol. 56, 289�314 (2002)
73. Mehta T., Childers S.E., Glaven R., Lovley D.R., Mester T.:A putative multicopper protein secreted by an atypical typeII secretion system involved in the reduction of insolubleelectron acceptors in Geobacter sulfurreducens. Micro-biology, 152, 2257�2264 (2006)
74. Mehta T., Coppi M.V., Childers S.E., Lovley D.R.: Outermembrane c-type cytochromes required for Fe(III) andMn(IV) oxide reduction in Geobacter sulfurreducens. Appl.Environ. Microbiol. 71, 8634�8641 (2005)
75. Methe B.A., C.M. Fraser I wsp.: Genom of Geobacter sul-furreducens: metal reduction in subsurface environments.Science, 302, 1967�1969 (2003) (wy¿ej cytowana praca jestdzie³em 34 autorów)
76. Morris J.M., Jin S.: Feasibility of using microbial fuel celltechnology for bioremediation of hydrocarbons in ground-water. J. Env. Sci. Health, 43, 18�23 (2008)
77. Myers C.R., Myers J.: Cloning and sequence of cymA, a geneencoding a tetraheme cytochrome c required for reduction ofiron (III), fumarate and nitrate by Shewanella putrefaciensMR-1. J. Bacteriol. 179, 1143�1152 (1997)
78. Nevin K.P., Lovley D.R.: Mechanisms for accessing insolubleFe(III) oxide during dissimilatory Fe(III) reduction by Geothrixfermentas. Appl. Environ. Microbiol. 68, 2294�2299 (2002)
79. Nevin K.P., Lovley D.R.: Mechanisms for Fe(III) oxidereduction in sedimentary environments. Geomicrobiol. J. 19,141�159 (2002)
80. Ortiz-Bernard I., Anderson R.T., Vrionis H.A., Lovley D.R.:Vanadium respiration by Geobacter metallireducens: novelstrategy for in situ removal of vanadium from groundwater.Appl. Environ. Microbiol. 70, 3091�3095 (2004)
81. Park H.S., Kim B.H., Kim H.S., Kim H.J., Kim G.T.,Kim M., Ch I.S., Park Y.K., Chang H.I.: A novel electro-chemically active and Fe(III)-reducing bacterium phylo-genetically related to Clostridium butyricum isolated froma microbial fuel cell. Anaerobe, 7, 297�306 (2001)
82. Petrie L., North N.N., Dollhopf S.L., Balkwill D.L., KostkaJ.E.: Enumeration and characterization of iron(III)-reducingmicrobial communities from acidic subsurface sedimentscontaminated with uranium. Appl. Environ. Microbiol. 69,7467�7479 (2003)
83. Pham C.A., Jung S.J., Phung N.T., Lee J., Chang I.S.,Kim B.H., Yi H., Chun J.: A novel electrochemically activeand Fe(III)-reducing bacterium phylogenetically related toAeromonas hydrophila, isolated from a microbial fuel cell.FEMS Microbiol. Lett. 223, 129�134 (2003)
84. Reguera G., McCarthy K.D., Mehta T., Nicoll J.S., Tuomi-nen M.T., Lovley D.R.: Extracellular electron transfer viamicrobial nanowires. Nature, 435, 1098�1101 (2005)
85. Reguera G., Nevin K.P., Nicoll J.S., Covalla S.F., WoodardT.L., Lovley D.R.: Biofilm and nanowire production leads toincreased current in Geobacter sulfurreducens fuel cells.Appl. Environ. Microbiol. 72, 7345�7348 (2006)
86. Reguera G., Pollina R.B., Nicoll J.S., Lovley D.R.: Possiblenonconductive role of Geobacter sulfurreducens pilus nano-wires in biofilm formation. J. Bacteriol. 189, 2125�2127 (2007)
87. Richter H., Lanthier M., Nevin K.P., Lovley D.R.: Lack ofelectricity production by Pelobacter carbinolicus indicatesthat the capacity for Fe(III) oxide reduction does not neces-sarily confer electron transfer ability to fuel cell anodes. Appl.Environ. Microbiol. 73, 5347�5353 (2007)
88. Richter H., McCarthy K., Nevin K.P., Johnson J.P., RotelloV.M., Lovley D.R.: Electricity generation by Geobacter sul-furreducens attached to gold electrodes. Langmuir, 24,4376�4379 (2008)
89. Schlesinger W.H.: Biogeochemistry: an analysis of globalchange. Academic Press, San Diego, 1997
90. Senko J.M., Istok J.D., Suflita J.M., Krumholz L.R.: In-situevidence for uranium immobilization and remobilization.Environ. Sci. Technol. 36, 1491�1496 (2002)

124 ANNA SIKORA I MIECZYS£AW K. B£ASZCZYK
91. Shelobolina E.S., Vrionis H.A., Findlay R.H., Lovley D.R.:Geobacter uraniireducens sp. nov., isolated from subsurfacesediment undergoing uranium bioremediation. Int. J. Syst.Evol. Microbiol. 58, 1075�1078 (2008)
92. Shi L, T.C. Squier i wsp.: Isolation of a high-affinity functio-nal protein complex between OmcA and MtrC: two outermembrane decaheme c-type cytochromes of S. oneidensisMR-1. J. Bacteriol. 188, 4705�4714 (2006) (wy¿ej cytowa-na praca jest dzie³em 17 autorów)
93. Shi L., Squier T.C., Zachara J.M., Fredrickson J.K.: Respira-tion of metal (hydr)oxides by Shewanella and Geobacter:a key role for multihaem c-type cytochromes. Mol. Micro-biol. 65, 12�20 (2007)
94. Sikora A., Sikora R.: Mikrobiologiczne ogniwa paliwowe(Microbial fuel cells). Biotechnologia Monografie, 2(2),68�77 (2005)
95. Slobodkin A.I.: Thermophilic microbial metal reduction.Microbiology, 74, 501�514 (2005)
96. Suzuki Y., Kelly S.D., Kemner K.M., Banfield J.F.: Directmicrobial reduction and subsequent preservation of uraniumin natural near-surface sediment. Appl. Environ. Microbiol.71, 1790�1797 (2005)
97. Tender L.M., Reimers C.E., Stecher H.A., Holmes D.E.,Bond D.R., Lowy D.A., Pilobello K., Fertig S.J., LovleyD.R.: Harnessing microbially generated power on theseafloor. Nature Biotechnol. 20, 821�825 (2002)
98. Vandieken V., Muâmann M., Niemann H., Jorgensen B.B.:Desulfuromonas svalbardensis sp. Nov. And Desulfuromusaferrireducens sp. Nov., psychrophilic, Fe(III)-reducing bac-teria isolated from Arctic sediments, Svalbard. Int. J. Sys-tem. Evol. Microbiol. 56, 1133�1139 (2006)
99. Yanke L.J., Bryant R.D., Laishley E.J.: Hydrogenase I ofClostridium pasteurianum functions as a novel selenitereductase. Anaerobe, 1, 61�67 (1995)
100. Ye Q., Roh Y., Carroll S.L., Blair B., Zhou J., Zhang C.L.,Fields M.W.: Alkaline anaerobic respiration: isolationand characterization of a novel alkaliphilic and metal-redu-cing bacterium. Appl. Environ. Microbiol. 70, 5595�5602(2004)
Praca powsta³a w ramach grantu Ministerstwa Nauki i Szkol-nictwa Wy¿szego nr 2P04B00429












