
Podstawy genetyki populacji -...
59
Podstawy genetyki populacji Populacje o skończonej liczebności. Dryf. Modele wielogenowe.
Transcript of Podstawy genetyki populacji -...

Podstawy genetyki populacji
Populacje o skończonej liczebności. Dryf. Modele wielogenowe.

Dryf genetyczny a ewolucja
2
} Dobór naturalny nie jest jedynym mechanizmem kształtującym zmiany ewolucyjne
} Losowe procesy w populacjach o skończonej liczebności – dryf genetyczny

Dryf genetyczny
3
} W populacjach o skończonej liczebności może dochodzić do zmian częstości alleli nawet jeżeli nie działa na nie dobór
} Nowy allel (mutacja) może się utrwalić w populacji nawet bez selekcji } częściowo (polimorfizm) } całkowicie

Model dryfu
} Populacja reprezentowana przez kulki w worku } 50 brązowych i 50 zielonych (allele)
} Losujemy 10 kulek } Uzupełniamy liczbę kulek znowu do 100
} w takiej samej proporcji, jak wylosowane 10 (model losowego sukcesu reprodukcyjnego)
} Efekt:

Działanie dryfu 1. Zmiana częstości alleli w populacji, może
zredukować zróżnicowanie populacji. 2. Działa szybciej w małych populacjach. 3. Może przyczynić się do specjacji
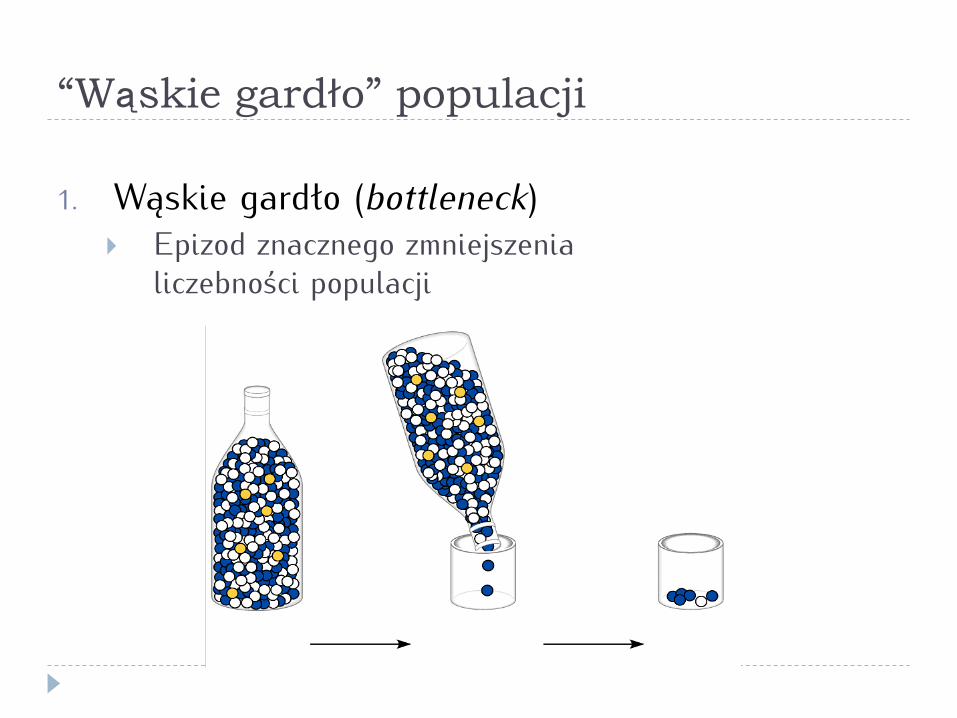
“Wąskie gardło” populacji
1. Wąskie gardło (bottleneck) } Epizod znacznego zmniejszenia
liczebności populacji

Znaczenie dla gatunku
7
} Wąskie gardło znacznie zmniejsza różnorodność genetyczną populacji przez dryf i wsobność
} Ogranicza to możliwości adaptacji do środowiska i stwarza zagrożenie dla populacji } choroby i pasożyty } zmiany środowiskowe } konkurencja
} Gdy liczebność populacji spadnie poniżej wartości krytycznej, gatunku nie da się utrzymać } główne zagrożenie – choroby zakaźne
Słoń morski północny
Gepard

Słoń morski północny
8
} Polowania w XVIII-XIX wieku zmniejszyły liczebność do <100 sztuk } Na początku XX wieku jedna kolonia u wybrzeży Meksyku } W XX wieku pod ochroną
} Obecnie >100 000 sztuk } Małe zróżnicowanie genetyczne

Inne przykłady
9
} Gepard } Zróżnicowanie na tyle małe, że przeszczepy od niespokrewnionych osobników nie
są odrzucane } Pierwsze wąskie gardło w epoce zlodowaceń
} Żubr } Obecnie ok. 3000 osobników, potomstwo 12 sztuk } Duża wrażliwość na choroby (np. pryszczyca)
} Wiele zwierząt domowych i hodowlanych } Chomik syryjski – wszystkie hodowlane osobniki wywodzą się z jednego miotu
znalezionego w Syrii ok. 1930 r. } W naturze gatunek rzadki i zagrożony
} Człowiek
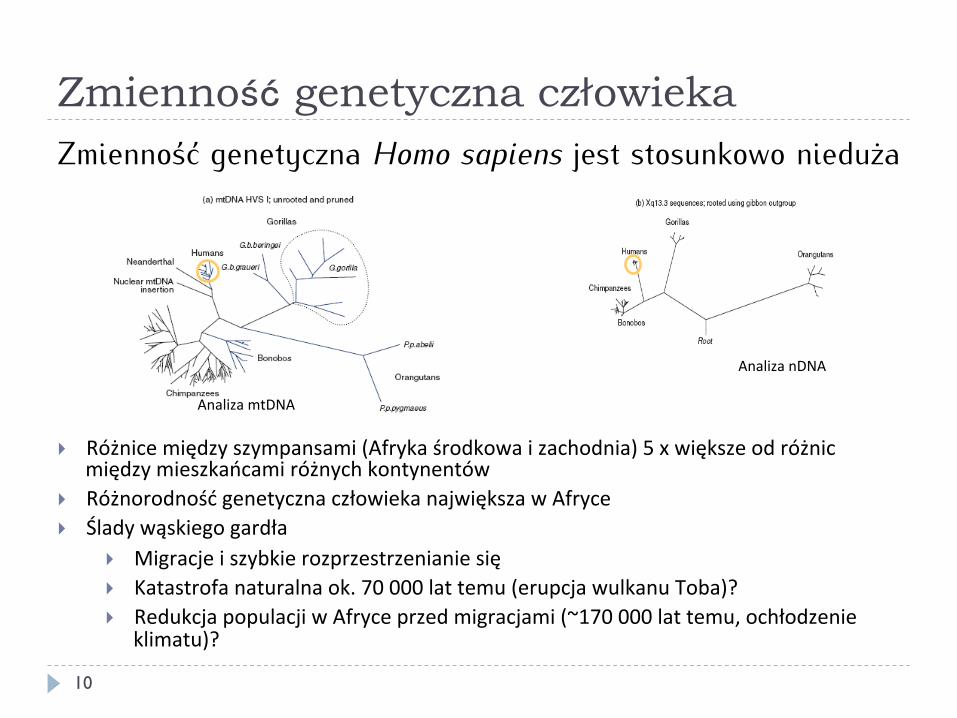
Zmienność genetyczna człowieka
10
Zmienność genetyczna Homo sapiens jest stosunkowo nieduża
} Różnice między szympansami (Afryka środkowa i zachodnia) 5 x większe od różnic między mieszkańcami różnych kontynentów
} Różnorodność genetyczna człowieka największa w Afryce } Ślady wąskiego gardła
} Migracje i szybkie rozprzestrzenianie się } Katastrofa naturalna ok. 70 000 lat temu (erupcja wulkanu Toba)? } Redukcja populacji w Afryce przed migracjami (~170 000 lat temu, ochłodzenie
klimatu)?
Analiza mtDNA
Analiza nDNA

Efekt założyciela
11
} Mała grupa oderwana od macierzystej populacji – efekt wąskiego gardła } kolonizacja nowych siedlisk } fragmentacja zasięgu
} Redukcja różnorodności allelicznej } Powstaje populacja, w której częstości alleli mogą się
znacznie różnić od obserwowanych w populacji macierzystej

Efekt założyciela
12
} Nowa populacja powstająca z niewielkiej liczby osobników może znacząco różnić się częstościami alleli od populacji wyjściowej
} U człowieka – niektore rzadkie choroby genetyczne występują częściej w pewnych grupach etnicznych
} Utrata różnorodności genetycznej człowieka – seria efektów założycielskich } Im dalej od Afryki, tym mniejsza
różnorodność

Wyspa niewidzących kolorów
13
} W 1775 wyspę Pingelap spustoszył tajfun, zginęło 90% ludności, ocalało ~20 osób
} Wśród ocalałych był władca Nahnmwarki Mwanenised, który był nosicielem rzadkiej recesywnej mutacji powodującej achromatopsję
} Obecnie 10% ludności wyspy nie widzi barw, a 30% to nosiciele } Dla porównania, w USA choroba występuje z częstością 1:33 000 osób
} Achromatopsja to nie to samo, co daltonizm!
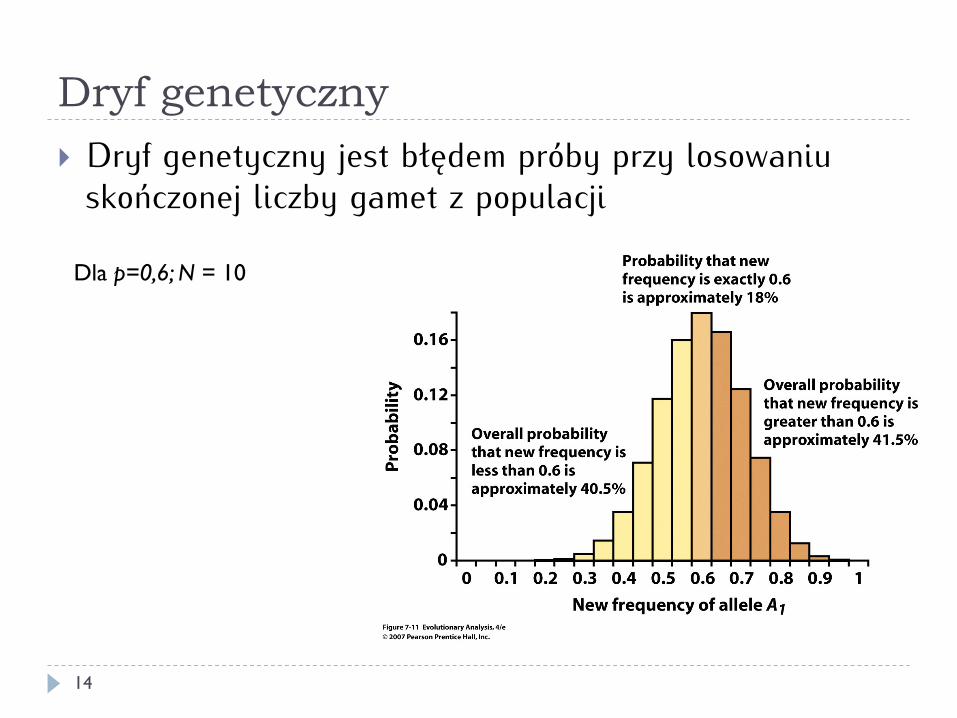
Dryf genetyczny
14
} Dryf genetyczny jest błędem próby przy losowaniu skończonej liczby gamet z populacji
Dla p=0,6; N = 10

Dryf a wielkość populacji
15
} Efekty dryfu genetycznego są wyraźniejsze w populacjach o mniejszej wielkości
} Z czasem dryf doprowadzi do utraty jednego z alleli i utrwalenia drugiego – utrata heterozygotyczności
16
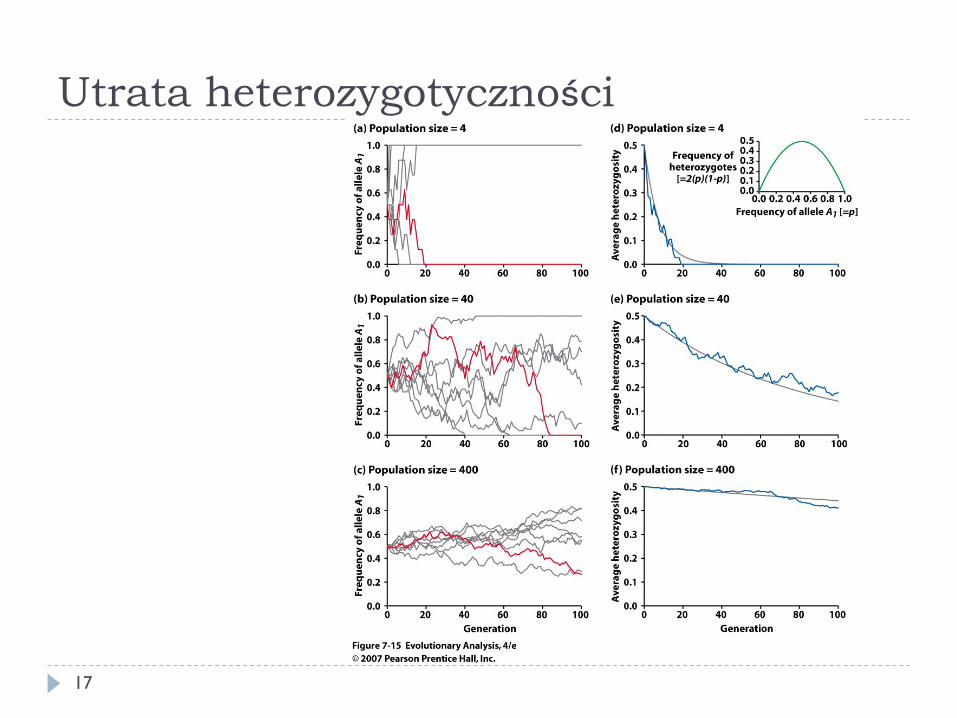
Utrata heterozygotyczności
17

Utrata heterozygotyczności
18
} Przy braku działania doboru dryf doprowadzi do utraty jednego allelu i utrwalenia (fiksacji) drugiego
} Może powodować powstanie populacji odmiennych genetycznie, bez udziału doboru
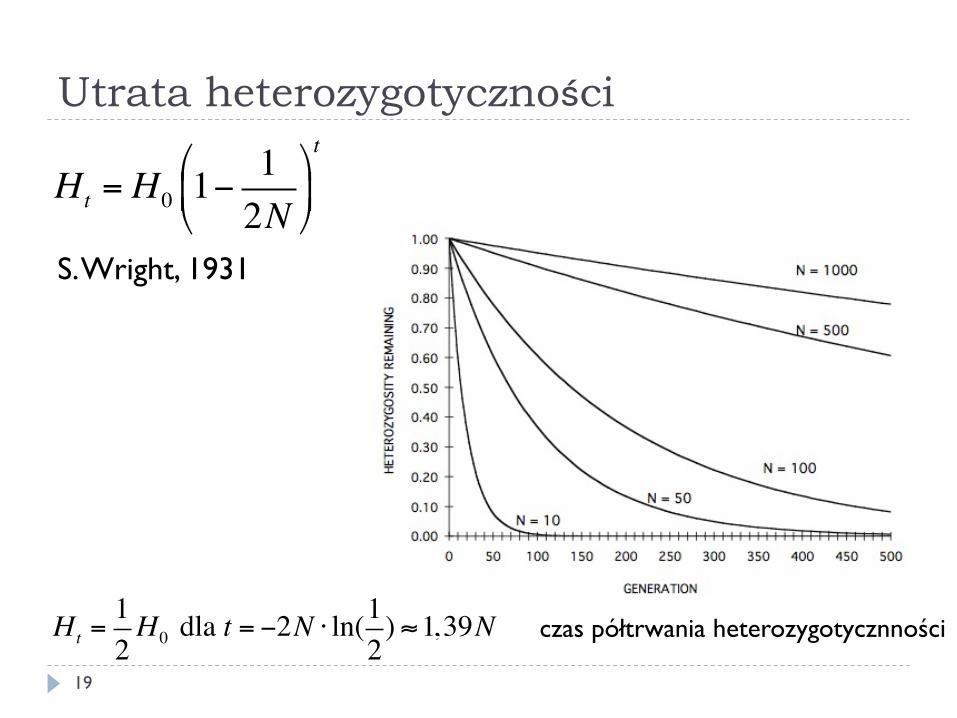
Utrata heterozygotyczności
19
Ht = H0 1−12N
"
#$
%
&'t
S. Wright, 1931
Ht =12H0 dla t = −2N ⋅ ln(1
2) ≈1,39N czas półtrwania heterozygotycznności

Efektywna wielkość populacji
20
} We wszystkich modelach zakładaliśmy panmiksję – jednakowe prawdopodobieństwo wydania potomstwa przez każdego osobnika
} Rzeczywiste populacje nie spełniają tego warunku } nierównomierne stosunki płci (haremy) } zróżnicowanie sukcesu reprodukcyjnego

Efektywna wielkość populacji
21
} Efektywna wielkość populacji Ne jest to liczebność idealnej populacji panmiktycznej, w której tempo dryfu byłoby takie same, jak w badanej populacji o rzeczywistej liczebności N
} We wszystkich dotychczasowych rozważaniach podając N tak naprawdę mieliśmy na myśli Ne

Efektywna wielkość populacji
22
} Zależność Ne (N) nie jest ewidentna, zależy od biologii danej populacji
} Niektóre proste modele: } różna liczba samców i samic uczestniczących w rozrodzie
} wsobność
} zmienna liczba osobników w różnych pokoleniach
Ne =4N f Nm
N f + Nm
Ne =N1+ f
Ne =1t
1Nii=1
t
∑

Efektywna wielkość populacji
23
} Ne można zbadać analizując neutralne polimorfizmy w populacji i porównać z N (zliczeniem osobników)
} Przykłady Ne/N } kot domowy: 0,4 } traszka grzebieniasta: 0,16 } grizzly: 0,27
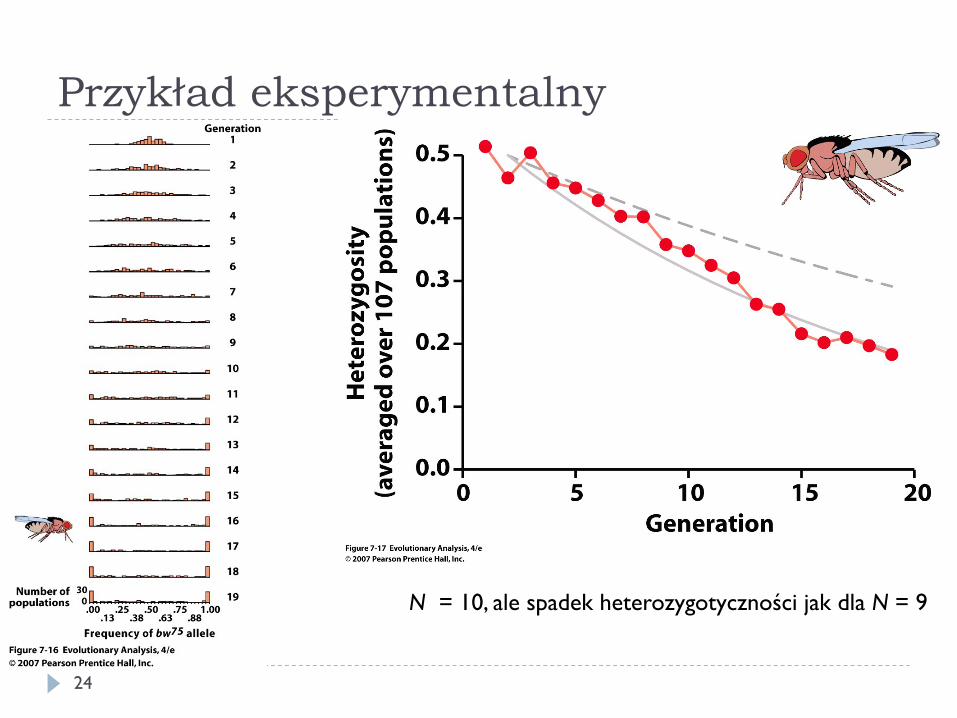
Przykład eksperymentalny
24
N = 10, ale spadek heterozygotyczności jak dla N = 9

Utrwalenie allelu
25
Prawdopodobieństwo utrwalenia konkretnego allelu } W populacji N osobników diploidalnych jest 2N alleli } Utrwalenie oznacza, że wszystkie allele obecne w
populacji pochodzą od jednego } Prawdopodobieństwo tego jest 1/2N } Jeżeli częstość allelu jest p, to wyjściowo jest 2Np
kopii } Czyli prawdopodobieństwo utrwalenia wynosi:
2Np×1/2N = p

Dryf i mutacje
26
} Mutacja powoduje powstanie nowego allelu } Przy założeniu braku doboru (neutralność) } Prawdopodobieństwo, że nowy allel się utrwali wynosi
1/2N } Utrwalanie się kolejnych mutacji powoduje ewolucję
populacji – ewolucja neutralna

Tempo ewolucji neutralnej
27
Prawdopodobieństwo utrwalenia mutacji neutralnej: 1/2N Prawdopodobieństwo powstania zmutowanego allelu: 2Nµ (µ - tempo mutacji) Prawdopodobieństwo powstania i utrwalenia się zmutowanego allelu (tempo ewolucji neutralnej):
2Nµ ⋅ 12N
= µ
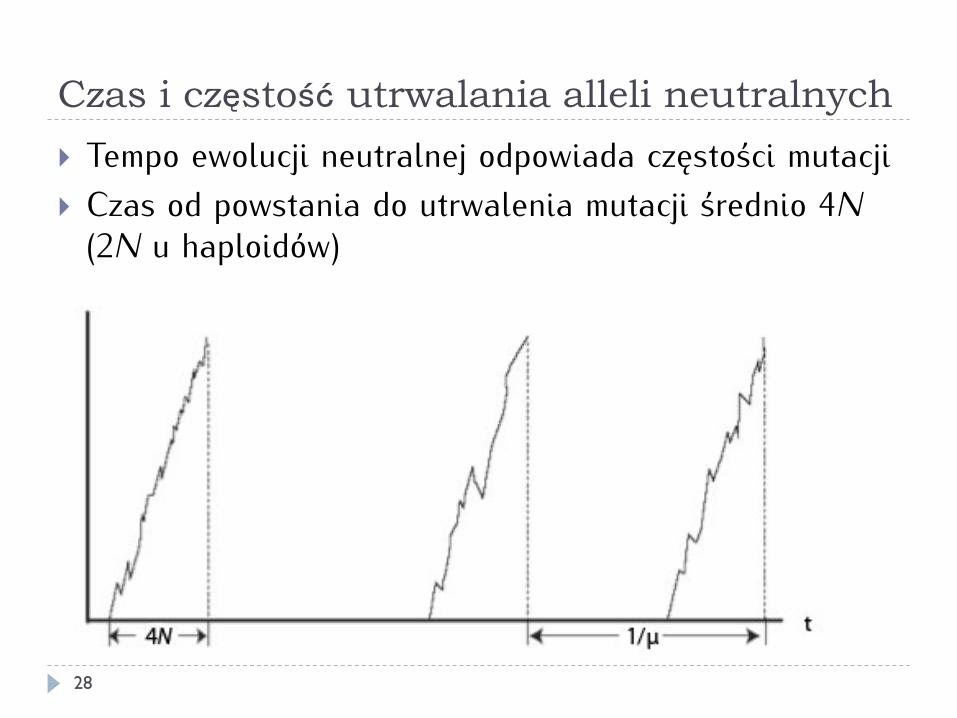
Czas i częstość utrwalania alleli neutralnych
28
} Tempo ewolucji neutralnej odpowiada częstości mutacji } Czas od powstania do utrwalenia mutacji średnio 4N
(2N u haploidów)

Ewolucja neutralna
29
} Dryf zmniejsza różnorodność alleli (prowadzi do utrwalania jedneo z alleli)
} Mutacje powodują powstawanie nowych alleli } Dzięki temu różnorodność zostanie zachowana, ale
skład konkretnych alleli się będzie zmieniał

Dryf i dobór
30
} Dryf może doprowadzić do utraty allelu korzystnego, albo do utrwalenia allelu niekorzystnego
} Równowaga między dryfem a doborem zależy od wielkości populacji i siły (współczynnika) selekcji
} Prosty model (kodominacja) A1A2 A1A2 A2A2
w 1 1+s 1+2s

Dryf i dobór
31
Prosty model (kodominacja) A1A2 A1A2 A2A2
w 1 1+s 1+2s Model nie jest trywialny do wyprowadzenia (Kimura 1962) Rezultat: P = 1− e
−4Nesq
1− e−4Nes

Dryf i dobór
32
Gdy s ≈ 0 to P ≈ q (prawdopodobieństwo utrwalenia allelu neutralnego jest równe jego częstości)
P = 1− e−4Nesq
1− e−4Nes

Dryf i dobór
33
Gdy s ≈ 0 to P ≈ q (prawdopodobieństwo utrwalenia allelu neutralnego jest równe jego częstości)
P = 1− e−4Nesq
1− e−4Nes

Dryf i dobór – allele nieznacznie korzystne
34
} Jeżeli s > 0 i N jest duże to P ≈ 2s } 98% mutacji o s = 0,01 się nie utrwali
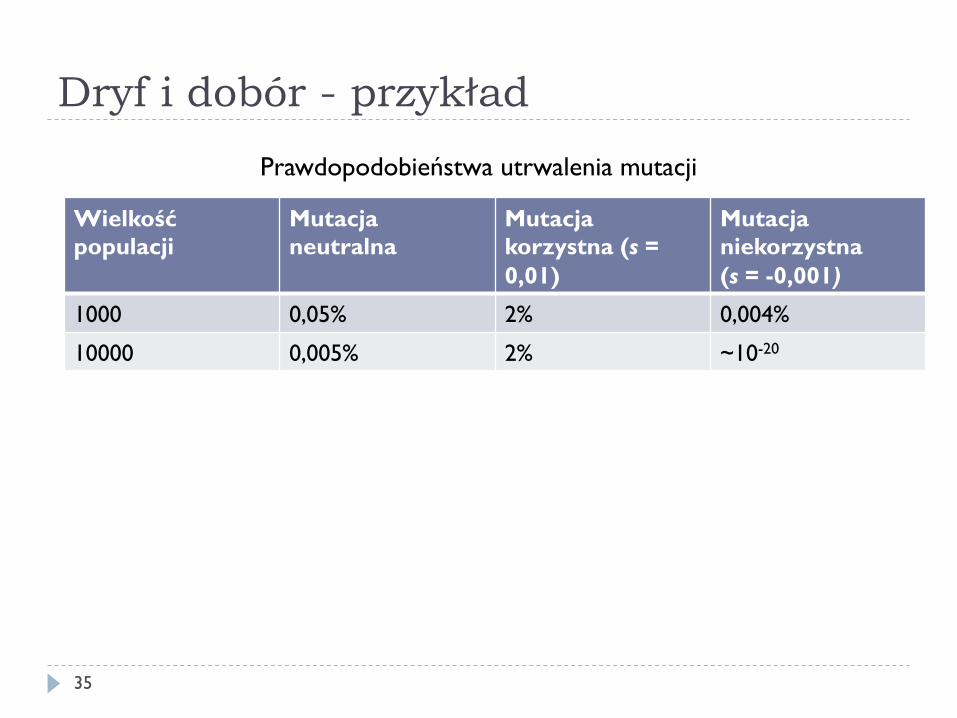
Dryf i dobór - przykład
35
Wielkość populacji
Mutacja neutralna
Mutacja korzystna (s = 0,01)
Mutacja niekorzystna (s = -0,001)
1000 0,05% 2% 0,004%
10000 0,005% 2% ~10-20
Prawdopodobieństwa utrwalenia mutacji

Dryf i dobór
36
} Czas utrwalenia się mutacji neutralnej
} Czas utrwalenia się mutacji korzystnej t = 4N
t = 2 ln(2N )s

Dryf i dobór
37
} Tempo utrwalania mutacji neutralnych
} Tempo utrwalania mutacji korzystnych
2Nµ ⋅ 12N
= µ
2Nµ ⋅2s = 4Nsµ
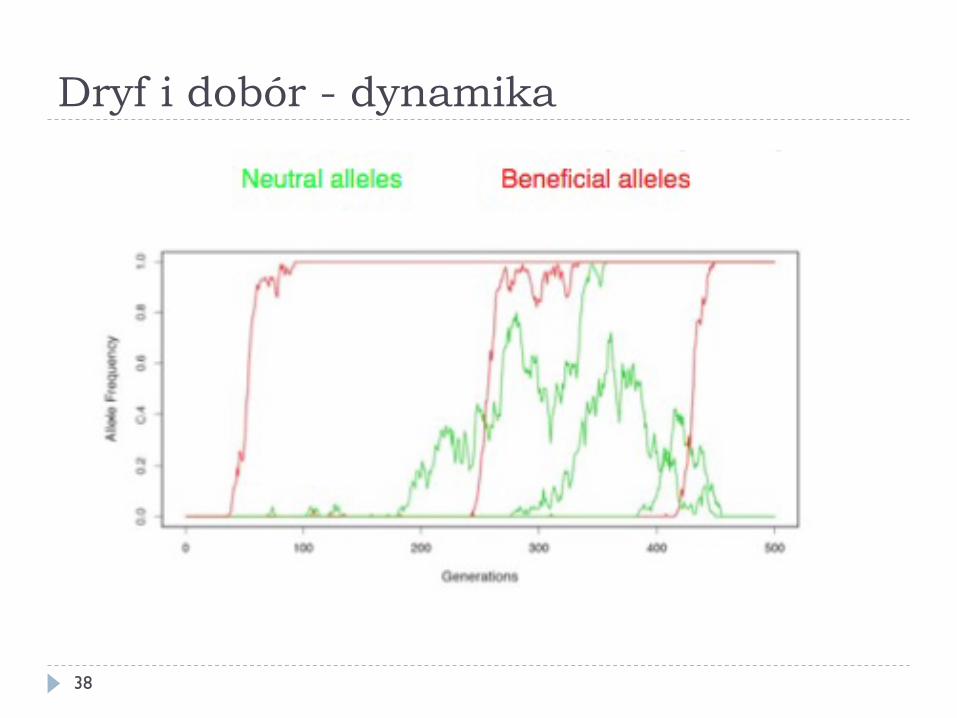
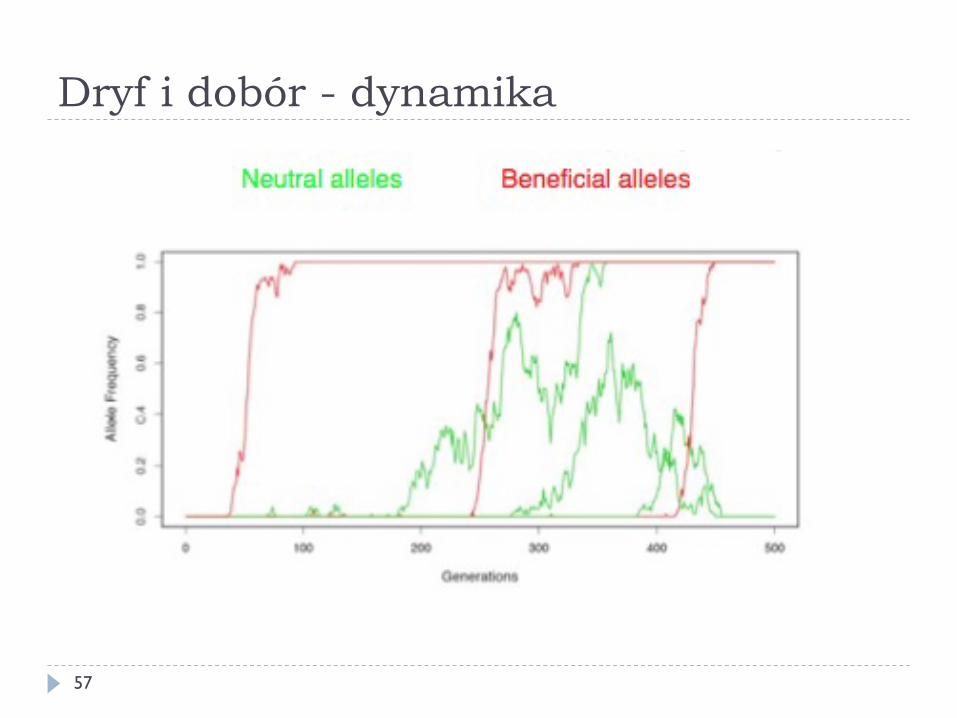
Dryf i dobór - dynamika
38

Dryf i dobór - podsumowanie
39
} Większość mutacji (korzystnych, neutralnych i niekorzystnych) nie utrwali się w populacji
} Gdy dobór przeciwko allelowi niekorzystnemu jest nieznaczny mutacja szkodliwa jest efektywnie neutralna – zostanie utrwalona z prawdopodobieństwem takim, jak neutralna
} Dobór jest nieznaczny gdy:
s ≤ 14Ne

Dryf i dobór – podsumowanie
40
} Gdy Ne jest duże, mutacje szkodliwe są skutecznie usuwane
} Nawet gdy Ne jest duże, wiele mutacji korzystnych jest traconych, jeżeli s nie jest bardzo duże

Modele wielogenowe
41

Modele wielogenowe
42
} Efekty związane ze sprzężeniem loci } równowaga i nierównowaga sprzężeń } zmiatanie selekcyjne i genetic hitchhiking
} Cechy wielogenowe i wieloczynnikowe } loci cech ilościowych (QTL) } supergeny

Sprzężenia i równowaga sprzężeń
43
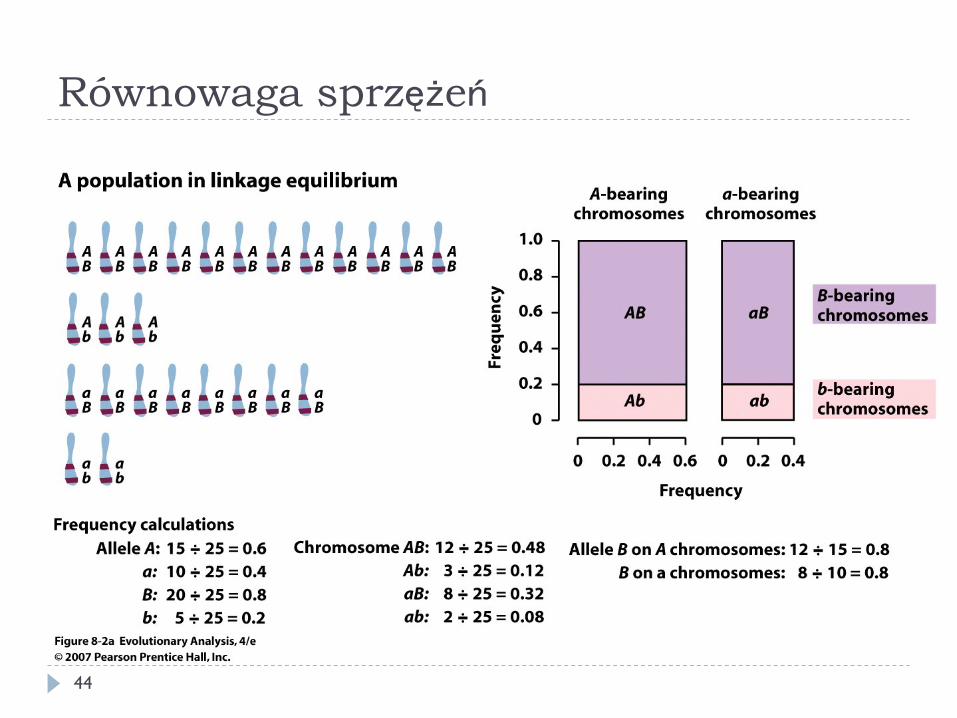
} Równowaga sprzężeń – genotyp w jednym locus jest niezależny od genotypu w drugim
} Haplotyp – genotyp (zbiór alleli) dla wielu loci danego chromosomu (lub gamety)
Równowaga sprzężeń
44

Równowaga sprzężeń
45
} W populacji będącej w stanie równowagi częstość haplotypu to iloczyn częstości alleli
A a B b p q s t Haplotypy AB Ab aB ab ps pt qs qt
Nierównowaga sprzężęń
46

Nierównowaga sprzężeń
47
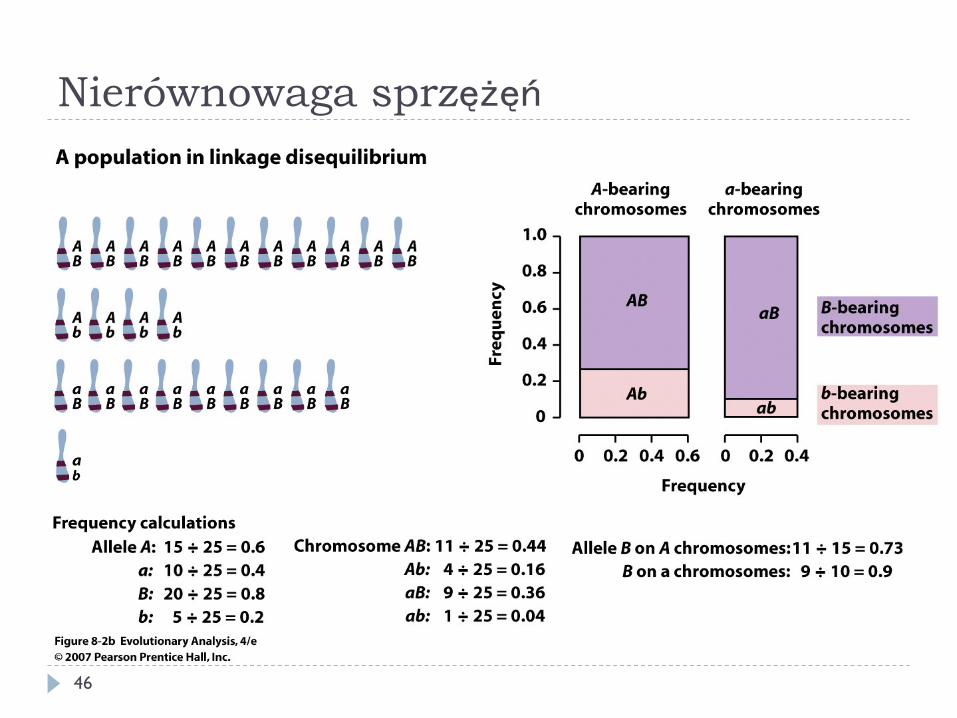
} Nielosowa korelacja genotypu (allelu) w jednym locus z allelem w drugim locus
} Współczynnik nierównowagi D
gdzie gAB to częstość haplotypu AB itd.
} D przyjmuje wartości od -1/4 do +1/4, dla populacji w równowadze sprzężeń D = 0
D = gABgab − gAbgaB

Inna miara nierównowago sprzężeń
48
Korelacja alleliczna r gdzie p, q, s, t to częstości alleli (p i q w locus A, s i t w locus B). Wartości od -1 do +1. Czasmi podawana też wartość r2
r = Dpqst

Skąd bierze się nierównowaga sprzężeń
49
} Migracje
} Dobór na genotyp wielu loci } efekty kumulatywne } supergeny
} Kombinacja doboru w jednym z loci i dryfu

Hitch-hiking i zmiatanie selekcyjne
50
} W jednym locus pojawia się korzystna mutacja } Dobór naturalny szybko utrwala ten korzystny allel } Wraz z nim utrwalają się neutralne (a nawet
niekorzystne) allele w loci blisko sprzężonych – genetic hitch-hiking
} W sąsiedztwie niedawno utrwalonego korzystnego allelu obserwuje się zmniejszoną różnorodność alleliczną – zmiatanie selekcyjne (selective sweep)

Zmiatanie selekcyjne
51 © Nature Edutaction, 2008
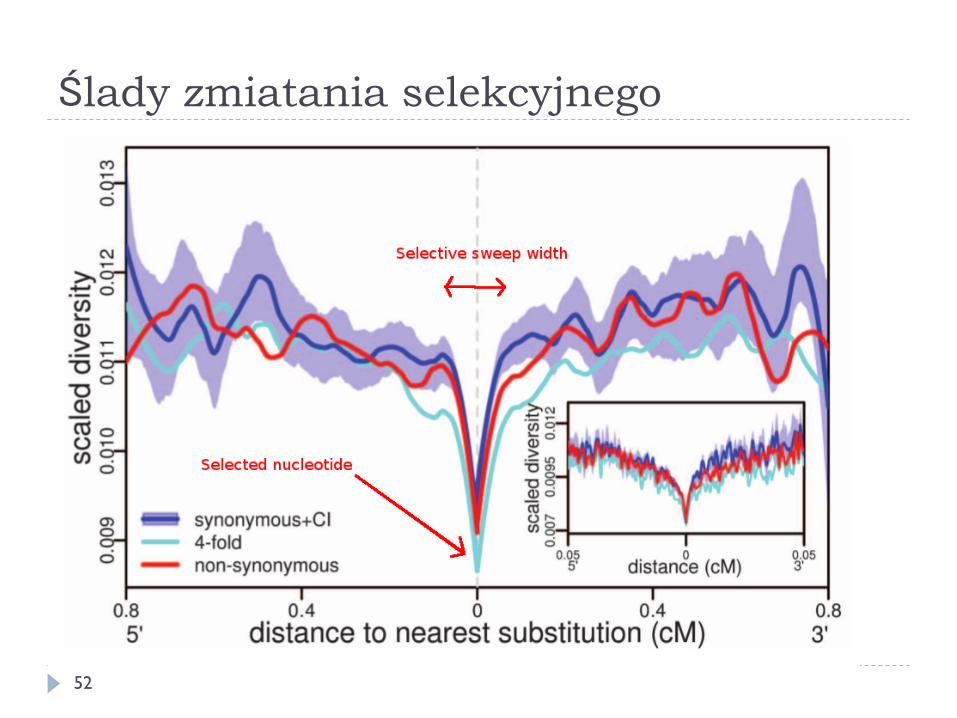
Ślady zmiatania selekcyjnego
52
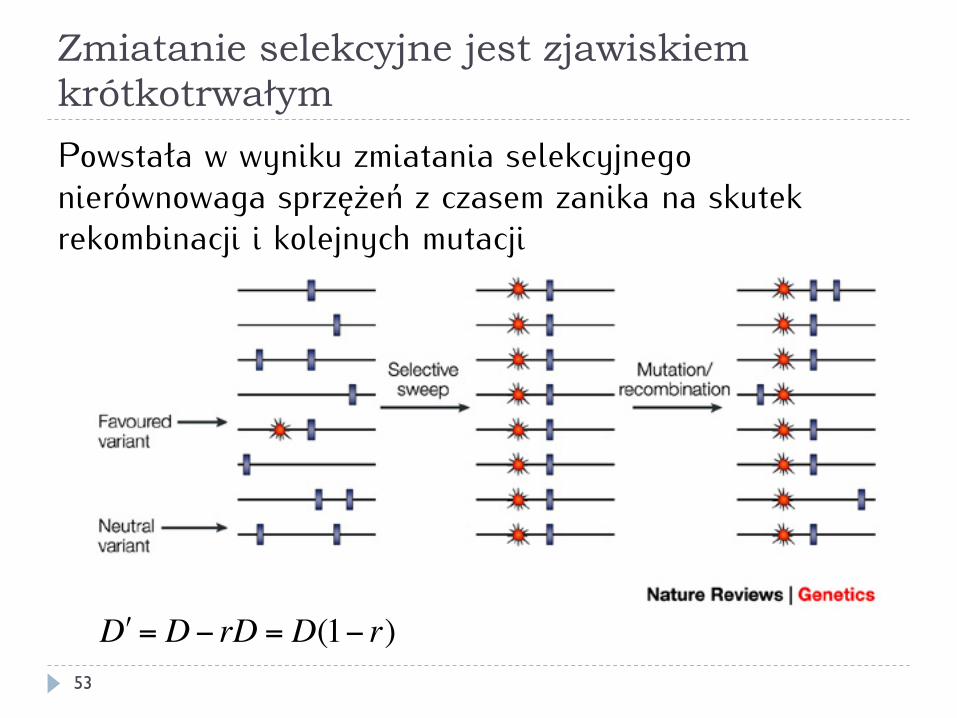
Zmiatanie selekcyjne jest zjawiskiem krótkotrwałym
53
Powstała w wyniku zmiatania selekcyjnego nierównowaga sprzężeń z czasem zanika na skutek rekombinacji i kolejnych mutacji
!D = D− rD = D(1− r)
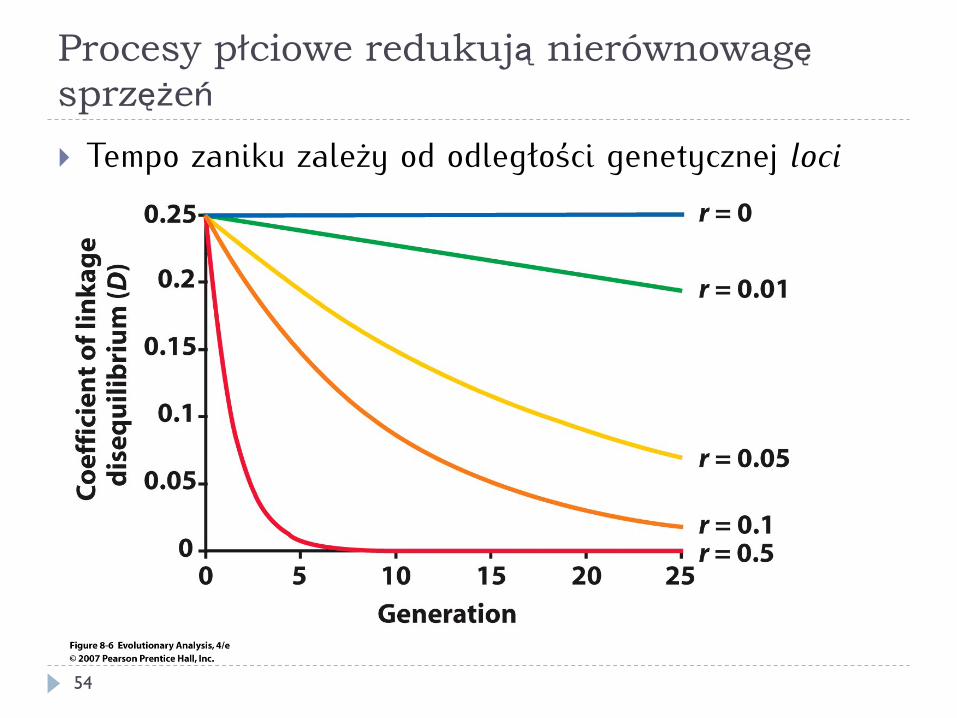
Procesy płciowe redukują nierównowagę sprzężeń
54
} Tempo zaniku zależy od odległości genetycznej loci

Zastosowanie
55
} Zakładając, że mutacja korzystna pojawiła się w populacji jednokrotnie, badanie różnorodności neutralnych alleli (markerów) w pobliżu locus mutacji pozwoli oszacować wiek mutacji.
} Podobny efekt daje też epizod wąskiego gardła liczebności populacji – badając nierównowagę sprzężeń można badać historię demograficzną populacji

Wykrywanie doboru dodatniego
56
} Pojawienie się nowej mutacji tworzy nierównowagę sprzężeń
} Jeżeli nierównowaga jest wyraźna, to oznacza to, że mutacja pojawiła się niedawno (rekombinacja nie zdążyła jej zaburzyć)
} Jeżeli taka niedawna mutacja ma wysoką częstość w populacji, to znaczy, że utrwalał ją dobór dodatni
Dryf i dobór - dynamika
57
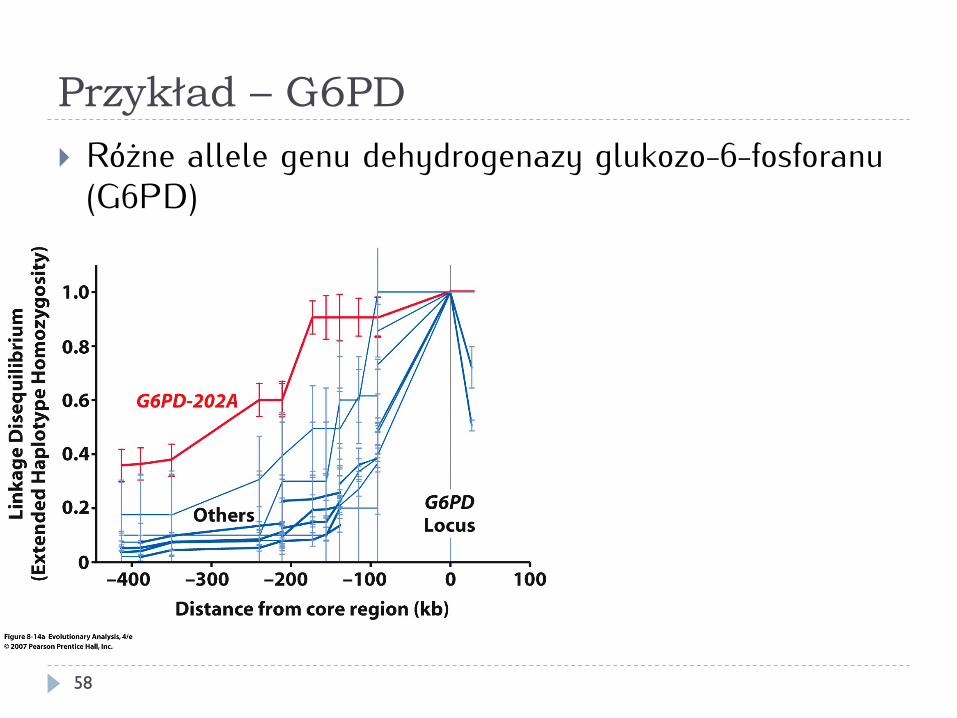
Przykład – G6PD
58
} Różne allele genu dehydrogenazy glukozo-6-fosforanu (G6PD)

Przykład – G6PD
59
} Allel G6PD-202A wykazuje ślady doboru dodatniego (zmiatanie selekcyjne)
} Oporność na malarię


















