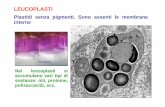
Plastidi, fotosintesi!
33
Processo che converte Energia luminosa in Energia chimica e coinvolge molte molecole diverse: pigmenti enzimi accettori e trasportatori di e - Componenti funzionali dei cloroplasti Sono quelli legati al processo più importante che ha sede nei cloroplasti: la fotosintesi. La fotosintesi è effettuata dagli organismi autotrofi che possiedono strutture, pigmenti e enzimi in grado di catturare l’energia luminosa.
-
Upload
jennifer-pradelli -
Category
Education
-
view
687 -
download
0
Transcript of Plastidi, fotosintesi!

Processo che converte
Energia luminosa in
Energia chimica e
coinvolge molte molecole
diverse:
pigmenti
enzimi
accettori e trasportatori di
e-
Componenti funzionali dei cloroplasti
Sono quelli legati al processo
più importante che ha sede
nei cloroplasti: la fotosintesi.
La fotosintesi è effettuata dagli
organismi autotrofi che possiedono
strutture, pigmenti e enzimi in grado
di catturare l’energia luminosa.

Con questo processo gli organismi sono in grado di (1) trasformare l’energia luminosa in energia chimica sotto forma di energia di legame (ATP) e di energia di potenziale di ossidoriduzione (NADPH) e di (2) organicare e ridurre la CO2.
luce (h)
6 CO2 + 6 H2O C6H12O6 + 6O2 (-684 kcal)
clorofilla
Consta di due fasi:
1) Fase luminosa (o fotochimica)
2) Fase di carbonificazione

• Sui tilacoidi sono presenti:
• Complessi antenna: assorbono e trasferiscono l’Energia
luminosa
• Centri di reazione: trasformano l’energia luminosa in
Energia di tipo ossidoriduttivo
• Enzimi: es. ATPasi
• Accettori e trasportatori di elettroni
FASE LUMINOSA (reazioni fotochimiche)
ha luogo nei tilacoidi
-cattura dell’energia luminosa e foto-eccitazione della clorofilla
-fotolisi dell’acqua e produzione di potere ossidante O2
-produzione di potere riducente NADPH
-produzione di energia chimica ATP

• FASE di carbonificazione
• ha luogo nello stroma
• cattura e fissazione (riduzione) della CO2
organicazione del C
• consumo di ATP e di NADPH
Nello stroma sono presenti:
Enzimi: convertono l’Energia accumulata come ATP e NADPH in
carboidrati e altri composti
-enzima RuBisCO
-enzimi del Ciclo di Calvin-Benson (o ciclo C3)
-enzimi per la sintesi e la degradazione dell’amido
-enzimi per la riduzione e l’organicazione dell’azoto

The yellow loops indicate the positions of the
active-site.
è particolarmente lento: fissa solo 3
molecole di CO2 al secondo; in
compenso è l’enzima più
abbondante sulla Terra
l’enzima Rubisco,
ribulosio bifosfato carbossilasi/ossigenasi,
è il ponte tra sistema vivente e sistema non-vivente
è composto da 8 copie di una grossa proteina (blue & light
blue), codificata dal DNA cloroplastidiale e sintetizzata
nello stroma, e da 8 copie di una proteina più piccola (red &
orange), codificata dal DNA nucleare, sintetizzata nel
citoplasma e poi importata.

I pigmenti sono sostanze che
assorbono specifiche lunghezze
d’onda e hanno un colore
corrispondente alle lunghezze
d’onda che non assorbono e che
riflettono.
PIGMENTI FOTOSINTETICI
*
Le clorofille sono
metallocomposti della
famiglia delle porfirine.
L’anello porfirinico, simile
all’eme dell’emoglobina, ha
una serie di doppi legami
coniugati (cioè un legame C-
C alternato a C=C)
Al centro dell’anello: atomo
di Mg coordinato con i 4
atomi N

In natura quattro clorofille:
Clorofilla a: in tutti gli organismi fotosintetici
(procarioti e eucarioti)
Clorofilla b: Archeoplastidia, Proclorofita
(anche alghe euglenoidi)
Clorofilla c: in molte alghe eucariote (non appartenenti
agli Archeoplastidia)
Clorofilla d: rara (in poche alghe rosse e in un
solo cianobatterio)

*
Clorofilla a: coda di fitolo,
alcol alifatico esterificato
all’anello D, che rende le
molecole idrofobiche;
presenza di un gruppo
metilico CH3 all’anello B.
Clorofilla b: identica,
ma con gruppo aldeidico
(-CHO) al posto del
metilico
Clorofilla c: manca la coda
di Fitolo, al suo posto H
Clorofilla d: con -CHO al
posto del gruppo H2C=CH

Carotenoidi: pigmenti liposolubili dal
giallo all’arancio; presenti in tutti gli
organismi fotosintetici. Formati da molte
unità isoprenoidi che possono formare
agli estremi della molecola un anello
esagonale. Si dividono in 2 categorie:
Caroteni: es. Β-carotene (carote) e
licopene (pomodoro)
Xantofille: derivati ossigenati, es.
zeaxantina (cariossidi mais)
Pigmenti accessori

le clorofille
presentano un
minimo di
assorbimento
nella regione del
verde.
Per le piante, la
luce
monocromatica
di questo
colore equivale
a buio
completo



Tornando al processo fotosintetico….
Nei cloroplasti i pigmenti fotosintetici sono legati alle
membrane tilacoidali.
I pigmenti sono associati tra loro e ad altre molecole a
formare dei complessi chiamati FOTOSISTEMI: ognuno è
costituito da 250 a 400 molecole di pigmenti.

Ogni fotosistema è costituito da più di 200 molecole di clorofilla a,
piccole quantità di clorofilla b, carotenoidi legati a proteine, e
molecole di clorofilla speciale, chiamate P, presenti nel centro di
reazione.
I pigmenti del fotosistema assorbono E luminosa (fotoni) e si
eccitano, trasferendo poi E ad un’altra molecola per risonanza.
Parte idrofobica
Anello tetrapirrolico
Fosfolipidi
di
membrana
CENTRO DI REAZIONE
Da Museth, 2006
fotone
fotone
I FOTOSISTEMI


e- : da
ossidazione
acqua
e- :vanno →
NADP+ che si
riduce a
NADPH
I Centri di Reazione sono due, indicati come P 680 e P 700.

P 700: consiste di una clorofilla a dimerica (ma una
sola si ossida)
P 680: è compresa nelle proteine integrali di membrana
tilacoidale D1 e D2, precisamente in quella D1.
P 680 fa parte del Fotosistema II
P 700: fa parte del Fotosistema I
Il primo Fotosistema ad entrare in funzione è il
Fotosistema II, che tuttavia è stato scoperto dopo il
Fotosistema I, da cui il nome.

Clorofilla a Clorofilla a
FOTOSISTEMA II FOTOSISTEMA I
Fotolisi
dell’acqua
L’O2 è un
sottoprodotto
della
fotosintesi
O2 O2
O2
FOTOSISTEMA I: localizzato
prevalentemente nei tilacoidi intergrana
e sui margini dei tilacoidi dei grana.
FOTOSISTEMA II: localizzato
prevalentemente nelle zone
appressate dei grana (partizioni)

PSII e PSI lavorano in serie a trasferire elettroni dalla fotolisi dell’acqua a una B-
vitamina nota come NADP+. Dalla ossidazione delle clorofille si ottengono
elettroni eccitati che vengono trasferiti tramite appositi transfer. L’e- dal PII
passa al PI, che, in seguito a eccitazione, ha ceduto il proprio e- al NADP+. L’e-
perso dal PII è subito rimpiazzato attraverso fotolisi dell’acqua.

PSI * e PSII* sono i centri di reazione ossidati: sono ossidanti fortissimi che devono
essere ridotti molto velocemente (nanosecondi 10-9). Per ridurre il PSII, gli elettroni
vengono strappati all’acqua tramite il “complesso evolvente ossigeno”, una parte
del PSII che sporge nel lume tilacoidale, tramite fotolisi dell’acqua. Per ridurre il PSI,
esso riceve tramite trasportatori l’elettrone perso dal PSII.

La produzione di ATP, benchè endoergonica, è possibile grazie al gradiente di H+
presente tra stroma e lume.
accumulo di protoni H+

Il complesso evolvente ossigeno

Principalmente come
conseguenza della
fotolisi dell’acqua, nel
lume si accumulano
protoni e si crea un
gradiente tra lume e
stroma.
I protoni passano
allo stroma attivando
l’ ATP sintetasi con
guadagno di Energia
e fosforilazione di
ADP ad ATP

The ATP synthase complex transports protons across the thylakoid
from lumen to stroma. These protons flow down an electrochemical
gradient and the energy released by that flow is trapped on the stroma
side of the membrane by ATP synthesis.

PSI non è un ossidante abbastanza forte da strappare elettroni attraverso
la fotolisi dell’acqua (difficilissima da ossidare). D’altra parte il PSII
“energizzato” non è un riduttore abbastanza forte da donare elettroni
al NADP+.
Perché due fotosistemi ?
Quindi ambedue i Fotosistemi sono necessari per
l’ossidazione dell’acqua e per la riduzione del NADP+.

Il sistema di trasporto elettroni (ETS) è un gruppo di proteine carrier di
membrana o periferiche, interagenti nel trasporto di elettroni. L’ETS fra i
due fotosistemi è dato da feofitina, plastochinone, citocromi b ed f e
plastocianina. L’ETS dopo il PSI consiste di quinoni, proteine con zolfo e
ferro, ferrodossina e una flavoproteina che riduce il NADP+. Questa
successione è chiamata trasporto aciclico.

I prodotti della
fase luminosa

Fase di carbonificazione Consiste di una serie di reazioni
enzimatiche che portano alla
formazione di carboidrati partendo da
CO2, ATP e NADPH. Si svolge nello
stroma.
Le reazioni sono catalizzate da enzimi
presenti nello stroma e organizzate nel
“CICLO di CALVIN” o “ciclo riduttivo dei
pentoso fosfati” (PCR)
1) carbossilazione: è
catalizzata da Rubisco;
aggiunge 1 molecola di
CO2 ad uno zucchero a 5
atomi di C (ribulosio 1,5
difosfato)
2 molecole stabili di
un composto a 3
atomi di C = 3 fosfo
gligerato =3-PGA

2) Reazioni di riduzione che portano alla formazione della gliceraldeide 3-
fosfato = GAP, un trioso che farà da mattone per la costruzione di tutti gli altri
zuccheri e di molte altre molecole organiche. L’energia necessaria è fornita
da NADPH e ATP (fase luminosa)
3) Reazioni di rigenerazione che portano a ricostruire il ribulosio 1,5
bifosfato in modo che sia disponibile il nuovo substrato su cui agirà la
Rubisco
L’organicazione di 36
molecole di CO2
porta alla sintesi un
esoso (6C) e 6
molecole del pentoso
(5x6 = 30 C)
necessarie all’azione
della Rubisco.
Per 1 molecola di
CO2 sono necessarie:
3 molecole di ATP e 2
di NADPH

+ Ribulosio-1,5- bifosfato
2 molecole di G3P Il GAP può essere usato nel metabolismo
plastidiale (es. amido primario) o essere
esportato nel citosol mediante uno specifico
traslocatore dei trioso fosfati, inserito nella
membrana interna del cloroplasto, che scambia
i triosi con P (da utilizzare nella sintesi di ATP).

Durante il giorno la quantità di zuccheri prodotta può essere superiore rispetto
alla quantità consumata o esportata nel citosol per il metabolismo della cellula.
All’interno del cloroplasto l’eccesso viene polimerizzato a amido, detto amido
primario, e deposto sotto forma di piccoli granuli all’interno del cloroplasto.
Durante la notte, questo amido
viene idrolizzato in zuccheri solubili,
sia per essere usato nel
metabolismo cellulare, sia per
essere traslocato ai plastidi di
riserva , gli amiloplasti, dove si
accumula come amido secondario.
Cloroplasto con
granuli di amido
primario
Giovane amiloplasto con
residui del sistema
lamellare e grandi granuli di
amido secondario


I prodotti della
fase di
carbonificazione
Non solo zuccheri,
anche aminoacidi,
grassi, ecc.