
Phytochrome Signaling Is Mediated by PHYTOCHROME ... · Phytochrome Signaling Is Mediated by...
17
Phytochrome Signaling Is Mediated by PHYTOCHROME INTERACTING FACTOR in the Liverwort Marchantia polymorpha Keisuke Inoue, a Ryuichi Nishihama, a Hideo Kataoka, a Masashi Hosaka, a Ryo Manabe, a Mika Nomoto, b Yasuomi Tada, c Kimitsune Ishizaki, a,d and Takayuki Kohchi a,1 a Graduate School of Biostudies, Kyoto University, Kyoto 606-8502, Japan b Division of Biological Science, Graduate School of Science, Nagoya University, Chikusa, Nagoya 464-8602, Japan c Center for Gene Research, Nagoya University, Chikusa, Nagoya 464-8602, Japan d Graduate School of Science, Kobe University, Kobe 657-8501, Japan ORCID ID: 0000-0003-0504-8196 (K. Ishizaki) Phytochromes are red light (R) and far-red light (FR) receptors that play important roles in many aspects of plant growth and development. Phytochromes mainly function in the nucleus and regulate sets of genes by inhibiting negatively acting basic helix-loop-helix transcription factors named PHYTOCHROME INTERACTING FACTORs (PIFs) in Arabidopsis thaliana. Although R/FR photoreversible responses and phytochrome genes are well documented in diverse lineages of plants, the extent to which phytochrome signaling is mediated by gene regulation beyond angiosperms remains largely unclear. Here, we show that the liverwort Marchantia polymorpha, an emerging model basal land plant, has only one phytochrome gene, Mp-PHY, and only one PIF gene, Mp-PIF. These genes mediate typical low fluence responses, which are reversibly elicited by R and FR, and regulate gene expression. Mp-phy is light-stable and translocates into the nucleus upon irradiation with either R or FR, demonstrating that the single phytochrome Mp-phy exhibits combined biochemical and cell-biological characteristics of type I and type II phytochromes. Mp-phy photoreversibly regulates gemma germination and downstream gene expression by interacting with Mp-PIF and targeting it for degradation in an R-dependent manner. Our findings suggest that the molecular mechanisms for light-dependent transcriptional regulation mediated by PIF transcription factors were established early in land plant evolution. INTRODUCTION Light is important for sessile plants not only as a source of energy for photosynthesis but also as a major source of environmental information that triggers adaptations in their growth and de- velopment. Land plants have acquired multiple photoreceptors, such as phytochromes, cryptochromes, phototropins, ZEITLUPE/ FLAVIN BINDING, KELCH REPEAT, F-BOX1/LOV KELCH PROTEIN2, and UV RESISTANCE LOCUS8, to perceive the light environment precisely (Kami et al., 2010; Ito et al., 2012; Jenkins, 2014; Christie et al., 2015). Among these photoreceptors, phy- tochromes are the sole receptors for red light (R) and far-red light (FR). Phytochromes exist in two different forms, the R-absorbing Pr form and the FR-absorbing Pfr form. R triggers activation of phytochromes by converting the Pr form to the Pfr form, whereas FR inactivates them by converting the Pfr form back to the Pr form. Phytochromes are grouped into two types, type I and type II, which have distinct biochemical and physiological properties in Arabidopsis thaliana. The type I phytochrome A (phyA) is highly abundant in etiolated seedlings and is rapidly degraded upon the conversion to the Pfr form, whereas the type II phytochromes phyB to phyE are relatively light-stable (Sharrock and Clack, 2002). Monocots such as rice (Oryza sativa) and maize (Zea mays) also have light-labile phyA and light-stable phyB (Reddy and Sharma, 1998; Xie et al., 2014). The rapid decline of light-labile phyto- chromes is observed within 2 to 4 h after R irradiation, although the degradation rate depends on the plant species. In Arabidopsis, both types of phytochromes translocate from the cytosol into the nucleus in response to light and mainly function in the nucleus (Kircher et al., 1999; Yamaguchi et al., 1999; Huq et al., 2003; Klose et al., 2015). Hence, light-dependent nuclear translocation is especially important for phytochrome signaling in Arabidopsis. The molecular mechanism for translocation is also different be- tween type I and type II phytochromes. Nuclear translocation of phyA is triggered by either R or FR and requires binding to FAR- RED ELONGATED HYPOCOTYL1 (FHY1) or FHY1-LIKE (FHL), which transports phyA to the nucleus (Hiltbrunner et al., 2006; Genoud et al., 2008; Rausenberger et al., 2011). By contrast, phyB efficiently translocates into the nucleus only in response to R (Yamaguchi et al., 1999). phyB does not require FHY1 or FHL for nuclear translocation and is thought to enter the nucleus via its own nuclear localization signal or by binding to transcription factors involved in phytochrome signaling (Chen et al., 2005; Pfeiffer et al., 2012). Physiological responses mediated by phytochromes are mainly classified into three modes of action, namely, low fluence response (LFR), very low fluence response (VLFR), and high irradiance re- sponse (HIR), according to fluence requirements, photoreversibility, 1 Address correspondence to [email protected]. The author responsible for distribution of materials integral to the findings presented in this article in accordance with the policy described in the Instructions for Authors (www.plantcell.org) is: Takayuki Kohchi ([email protected]). www.plantcell.org/cgi/doi/10.1105/tpc.15.01063 The Plant Cell, Vol. 28: 1406–1421, June 2016, www.plantcell.org ã 2016 American Society of Plant Biologists. All rights reserved.
Transcript of Phytochrome Signaling Is Mediated by PHYTOCHROME ... · Phytochrome Signaling Is Mediated by...

Phytochrome Signaling Is Mediated by PHYTOCHROMEINTERACTING FACTOR in the LiverwortMarchantia polymorpha
Keisuke Inoue,a Ryuichi Nishihama,a Hideo Kataoka,a Masashi Hosaka,a Ryo Manabe,a Mika Nomoto,b
Yasuomi Tada,c Kimitsune Ishizaki,a,d and Takayuki Kohchia,1
aGraduate School of Biostudies, Kyoto University, Kyoto 606-8502, JapanbDivision of Biological Science, Graduate School of Science, Nagoya University, Chikusa, Nagoya 464-8602, JapancCenter for Gene Research, Nagoya University, Chikusa, Nagoya 464-8602, JapandGraduate School of Science, Kobe University, Kobe 657-8501, Japan
ORCID ID: 0000-0003-0504-8196 (K. Ishizaki)
Phytochromes are red light (R) and far-red light (FR) receptors that play important roles in many aspects of plant growth anddevelopment. Phytochromes mainly function in the nucleus and regulate sets of genes by inhibiting negatively acting basichelix-loop-helix transcription factors named PHYTOCHROME INTERACTING FACTORs (PIFs) in Arabidopsis thaliana.Although R/FR photoreversible responses and phytochrome genes are well documented in diverse lineages of plants, theextent to which phytochrome signaling is mediated by gene regulation beyond angiosperms remains largely unclear. Here, weshow that the liverwort Marchantia polymorpha, an emerging model basal land plant, has only one phytochrome gene,Mp-PHY, and only one PIF gene, Mp-PIF. These genes mediate typical low fluence responses, which are reversibly elicited byR and FR, and regulate gene expression. Mp-phy is light-stable and translocates into the nucleus upon irradiation with eitherR or FR, demonstrating that the single phytochrome Mp-phy exhibits combined biochemical and cell-biological characteristicsof type I and type II phytochromes. Mp-phy photoreversibly regulates gemma germination and downstream gene expression byinteracting with Mp-PIF and targeting it for degradation in an R-dependent manner. Our findings suggest that the molecularmechanisms for light-dependent transcriptional regulation mediated by PIF transcription factors were established early in landplant evolution.
INTRODUCTION
Light is important for sessile plants not only as a source of energyfor photosynthesis but also as a major source of environmentalinformation that triggers adaptations in their growth and de-velopment. Land plants have acquired multiple photoreceptors,such as phytochromes, cryptochromes, phototropins, ZEITLUPE/FLAVIN BINDING, KELCH REPEAT, F-BOX1/LOV KELCHPROTEIN2, and UV RESISTANCE LOCUS8, to perceive the lightenvironment precisely (Kami et al., 2010; Ito et al., 2012; Jenkins,2014; Christie et al., 2015). Among these photoreceptors, phy-tochromes are the sole receptors for red light (R) and far-red light(FR). Phytochromes exist in two different forms, the R-absorbingPr form and the FR-absorbing Pfr form. R triggers activation ofphytochromes by converting the Pr form to the Pfr form, whereasFR inactivates thembyconverting thePfr formback to thePr form.
Phytochromes are grouped into two types, type I and type II,which have distinct biochemical and physiological properties inArabidopsis thaliana. The type I phytochrome A (phyA) is highlyabundant in etiolated seedlings and is rapidly degraded upon theconversion to thePfr form,whereas the type IIphytochromesphyB
to phyE are relatively light-stable (Sharrock and Clack, 2002).Monocots such as rice (Oryza sativa) and maize (Zea mays) alsohave light-labile phyA and light-stable phyB (Reddy and Sharma,1998; Xie et al., 2014). The rapid decline of light-labile phyto-chromes is observedwithin2 to4hafterR irradiation, although thedegradation rate depends on the plant species. In Arabidopsis,both types of phytochromes translocate from the cytosol into thenucleus in response to light and mainly function in the nucleus(Kircher et al., 1999;Yamaguchi etal., 1999;Huqetal., 2003;Kloseet al., 2015). Hence, light-dependent nuclear translocation isespecially important for phytochrome signaling in Arabidopsis.The molecular mechanism for translocation is also different be-tween type I and type II phytochromes. Nuclear translocation ofphyA is triggered by either R or FR and requires binding to FAR-RED ELONGATED HYPOCOTYL1 (FHY1) or FHY1-LIKE (FHL),which transports phyA to the nucleus (Hiltbrunner et al., 2006;Genoudet al., 2008; Rausenberger et al., 2011). By contrast, phyBefficiently translocates into the nucleus only in response to R(Yamaguchi et al., 1999). phyB does not require FHY1 or FHL fornuclear translocation and is thought to enter the nucleus via itsown nuclear localization signal or by binding to transcriptionfactors involved in phytochrome signaling (Chen et al., 2005;Pfeiffer et al., 2012).Physiological responses mediated by phytochromes are mainly
classified into three modes of action, namely, low fluence response(LFR), very low fluence response (VLFR), and high irradiance re-sponse (HIR), according to fluence requirements, photoreversibility,
1 Address correspondence to [email protected] author responsible for distribution of materials integral to the findingspresented in this article in accordance with the policy described in theInstructions for Authors (www.plantcell.org) is: Takayuki Kohchi([email protected]).www.plantcell.org/cgi/doi/10.1105/tpc.15.01063
The Plant Cell, Vol. 28: 1406–1421, June 2016, www.plantcell.org ã 2016 American Society of Plant Biologists. All rights reserved.

and reciprocity (Mancinelli, 1994; Casal et al., 1998). Among theseresponses, LFR is a well-documented response, which showsreciprocity and photoreversibility: It is induced by R and reversiblyinhibitedbyFR. VLFR is inducedby very-low-intensity visible light ofany wavelength and does not show photoreversibility, as a very lowratio of the Pfr form of phytochrome is sufficient to trigger the re-sponse (Shinomuraetal., 1996).HIR requiresprolongedexposure torelatively high-intensity light and shows neither reciprocity nor R/FRphotoreversibility. Under carefully designed experimental con-ditions, however, FR-HIR shows reciprocity and FR/R (not R/FR)photoreversibility (Shinomura et al., 2000). In Arabidopsis, phyAcontributes to VLFR and FR-HIR (Nagatani et al., 1993; Parks andQuail, 1993; Whitelam et al., 1993; Shinomura et al., 1996), whilephyBmainly regulates LFR andR-HIR (Mancinelli, 1994; Shinomuraet al., 1996).
Since the phytochromes in Arabidopsis mainly function in thenucleus as described above, most of the phytochrome responsesresult from regulation of gene expression via transcription factors.Collectively, numerous studies have demonstrated that there aretwo main pathways in phytochrome signaling. In the first pathway,PHYTOCHROME INTERACTINGFACTORs (PIFs), belonging to thebasic helix-loop-helix (bHLH) transcription factor family, were thefirst transcription factors to be identified and implicated in phyto-chromesignaling (Nietal.,1998;LeivarandMonte,2014).PIFsactasnegative regulators of phytochrome signaling by modulating theexpression of a large set of genes in the dark. Light-activatedphytochromes interact with PIFs and trigger phosphorylation andsubsequent proteasomal degradation of PIFs in a light-dependentmanner. The changes in protein abundance of PIFs result in vari-ous physiological responses, such as seed germination, seed-ling deetiolation, inhibition of hypocotyl elongation, and shadeavoidance responses. In the second pathway, CONSTITUTIVEPHOTOMORPHOGENIC1 (COP1)andSUPPRESSOROFPHYA-105(SPA) family proteins represent central negative regulators of lightsignaling (Huang et al., 2014). In the dark, a COP1/SPA complextargets a set of photomorphogenesis-promoting factors, suchas ELONGATED HYPOCOTYL5, for proteasomal degradationto inhibit photomorphogenesis. Recent analysis has shown thatlight-activatedphytochromes interactwith theSPA family in a light-dependent manner and promote the dissociation of the COP1/SPA complex (Lu et al., 2015; Sheerin et al., 2015). Phytochrome-dependent regulation of gene expression has also been reported inseveral angiosperms (Sawers et al., 2002; Takano et al., 2005;Kebrom et al., 2006; Takano et al., 2009; Kebrom et al., 2010) andgymnosperms (Alosi and Neale, 1992; Christensen et al., 2002).However, themolecularmechanismsofR-mediated transcriptionalregulation in these species are still poorly understood.
In contrast to the phytochromes in Arabidopsis, the phyto-chromes in the fern Adiantum capillus-veneris and the mossPhyscomitrella patens reportedly function largely in the cytosol or atthe plasma membrane and regulate phototropism and chloroplastphotorelocation movement, which are mainly regulated by thephototropins in Arabidopsis (Rösler et al., 2010; Jaedicke et al.,2012). Recently, nuclear functions of phytochromes in Ad. capillus-veneris and P. patens have been suggested. InAd. capillus-veneris,microbeam irradiation in the nucleus to selectively activate nucleus-localized phytochrome was able to induce spore germination(Tsuboi et al., 2012). In P. patens, PHY1 and PHY3 translocate from
the cytosol into the nucleus via identical mechanisms to those inseedplants, and they regulateHIR-like responses under continuousFR (Possart and Hiltbrunner, 2013). However, whether phyto-chromes regulate gene expression outside of angiosperms has notbeen directly confirmed.The liverwort Marchantia polymorpha is an emerging model
plantamong thebasal landplants,owing toseveral advantages forgenetic studies: a low-genetic redundancy, a haploid-dominantlife cycle, and tractable genetic tools, such as high-efficiencytransformation methods and genetic technologies (Ishizaki et al.,2008, 2013, 2016; Kubota et al., 2013; Sugano et al., 2014; Eklundet al., 2015; Kato et al., 2015; Flores-Sandoval et al., 2016;Nishihama et al., 2016). Since liverworts represent an early di-vergent lineage of extant land plants (Qiu et al., 2006), inves-tigations using liverworts are of particular interest in terms of landplant evolution. In addition, genome and transcriptome projectsare ongoing by the Community Sequencing Program in the U.S.Department of Energy Joint Genome Institute.Several studies of R responses in M. polymorpha have been
reported (Ninnemann and Halbsguth, 1965; Fredericq and DeGreef, 1966; De Greef et al., 1971; Otto and Halbsguth, 1976;Hartmann and Jenkins, 1984). A pulse of FR at the end of thephotoperiod induces vertical upward growth of thalli and a de-crease in theamountof chlorophyll, and theeffect ofR is cancelledby FR followed by a pulse of R, suggesting that a phytochrome isinvolved in these responses (Fredericq and De Greef, 1966;Hartmann and Jenkins, 1984). Experiments using pulses of R andFR demonstrate that senescence of thalli and germination ofgemmae may be regulated by phytochromes (Ninnemann andHalbsguth, 1965;DeGreef et al., 1971;Otto andHalbsguth, 1976).Recently, a phytochrome in M. polymorpha has been shown toregulate cell cycle reentry of differentiated cells in thallus explants,as well as cell shapes in newly regenerating tissues (Nishihamaet al., 2015). In this study, we discovered the lone, single-copygenes for phytochrome and PIF inM. polymorpha. We examinedthe biochemical and cell biological characteristics of the singlephytochrome and the involvement of the single PIF in R signaling.Our data show that M. polymorpha has a light-dependent tran-scriptional regulatory mechanism mediated by a module con-sisting of the phytochrome and PIF transcription factor.
RESULTS
Mp-PHY, the Only Phytochrome Gene in M. polymorpha
To identify the R and FR receptor phytochrome inM. polymorpha,we screened cDNA and genomic libraries and cloned a phyto-chrome gene (see Methods). BLAST searches against M. poly-morpha transcriptome and genome databases revealed that thisgene,namedMp-PHY, is theonlycopy resemblingaphytochromegene in the genome, which is consistent with previous reportsshowing the existence of one phytochromegene in other liverwortspecies (Suzuki et al., 2001; Mathews, 2006; Li et al., 2015). Theamino acid sequence of Mp-phy differs by only six residues fromthat of M. paleacea phytochrome, which has been reported toreside in a clade sister to that of phytochromes in seed plants(Supplemental Figure 1; Li et al., 2015). The deduced amino acidsequence of Mp-PHY included the typical domains of the
Phytochrome Signaling in Liverwort 1407

canonical phytochromes in land plants (Supplemental Figure 2).The genomic DNA sequence corresponding to Mp-PHY revealedthat all of the three intron-insertion sites in the coding sequencewere identical to those of most phytochrome genes in land plants(Supplemental Figure 2). To examine the spectral properties ofMp-phy, we expressed the N-terminal photosensory module ofMp-phy protein with phytochromobilin (PFB) chromophore inEscherichia coli. The recombinant Mp-phy protein exhibiteda typical R/FR photoreversibility (Supplemental Figures 3A and3B), much the same as is observed in the phytochromes in Ara-bidopsis (Mukougawa et al., 2006), suggesting that Mp-PHYencodes a functional and canonical phytochrome.
Mp-phy Protein Is Light-Stable in Vivo
We next examined whether Mp-phy more closely resembledtypical type I or type II phytochromes in termsof light stability. Ten-day-old wild-type plants grown under continuouswhite light weretransferred to the dark for 48 h and then irradiated with either R orFR for different time periods before harvesting. Immunoblotanalysis revealed that the abundance of Mp-phy was slightlyincreased during a prolonged incubation in the dark (Figure 1A).The level of Mp-phy was not changed within 8 h after irradiationwith R or 24 h after FR, though irradiation with R for 24 h causeda slight reduction ofMp-phy (Figures 1B and 1C). Given that phyAin Arabidopsis, rice, and maize show strong downregulation by Rwithin 2 to 4 h (Reddy and Sharma, 1998; Sharrock and Clack,2002; Xie et al., 2014), these results indicate that Mp-phy is light-stable in vivo, which is comparable to the type II phytochromes inArabidopsis and grasses.
Nuclear Translocation of Mp-phy Is Induced by Both Rand FR
In Arabidopsis, phyA is rapidly translocated from the cytosol intothenucleusundereitherRorFR,while thenuclear accumulationofphyB is observed only under R (Kircher et al., 1999; Yamaguchiet al., 1999). To investigate the subcellular localization of Mp-phy,we generated transgenic plants expressing Mp-phy as a fusionprotein with the fluorescent reporter Citrine at the C terminusunder the control of the cauliflower mosaic virus 35S (35S) pro-moter. One-day-old gemmalings grown under continuous whitelight were transferred to the dark for 6 d and then irradiated witheitherRorFR for10minbeforeobservation.Thephy-Citrine fusionprotein in thedark-adapted gemmalingswasmainly located at thecell periphery, although we have been unable to determinewhether this signal was derived from the cytosol or plasmamembrane. Upon the exposure to either R or FR, phy-Citrineaccumulatedwithin10min inasingle largeorganelleof thecell thatwas presumed to be the nucleus (Figure 2A). A mutated Mp-phycontaining an amino acid substitution of the conserved Tyr-241residue to His (Mp-phyY241H) was reported to be constitutivelyactive in vivo (Nishihama et al., 2015), similar to phyBY276H, thecorresponding mutant protein in Arabidopsis (Su and Lagarias,2007; Nishihama et al., 2015). The recombinant Mp-phyY241H
protein completely lacked R/FR photoreversibility, as was ob-servedwithphyBY276H (Supplemental Figures3Cand3D).Wealsogenerated transgenic plants expressing PHYY241H-Citrine underthe control of the 35S promoter and found that the phyY241H-
Citrine fusion protein localized in the nucleus without any expo-sure to light (Supplemental Figure 4). These results suggest thatthe active Pfr form of Mp-phy is translocated into the nucleus.To confirm the light-dependent nuclear translocation of Mp-
phy, we also performed subcellular fractionation experiments.Ten-day-old wild-type plants grown under continuous white lightwere transferred to the dark for 48 h and then irradiatedwith eitherR or FR for 60 min before harvesting. Consistent with the fluo-rescence imaging, immunoblot analysis revealed that the amountof Mp-phy protein in the nucleus-enriched fraction increasedwithin 10 min in response to either R or FR (Figures 2B and 2C).Although light induced rapidnuclear import ofMp-phy, subcellularfractionation results showed that the majority of Mp-phy was stillretained in the cytosol (Figures 2B and 2C). Furthermore, a bandwith lower mobility was also observed in the nucleus-enrichedfraction under FR, suggesting that the Mp-phy protein in thenucleus undergoes some type of posttranslational modificationunder FR (Figure 2C). Contrary to the results obtained with thefluorescence reporter assay, subcellular fractionation revealeda small amount of Mp-phy in the nucleus before light irradiation(Figures 2B and 2C). A similar observation was reported forAt-phyA in dark-grown seedlings, which was presumed to begenerated by brief light exposure before seedling germination
Figure 1. Light Stability of Mp-phy Protein.
Protein accumulation levels of Mp-phy were examined by immunoblotanalyses with anti-Mp-PHY antibodies in wild-type plants. Plants weregrown under continuous white light for 10 d and transferred to either of thefollowing light conditions for the indicated time periods: (A) darkness; (B)darkness for 48 h followedby irradiationwithR (30mmol photonsm22 s21); (C)darkness for 48 h followed by irradiation with FR (30 mmol photons m22 s21).Immunoblotting with anti-Histone H3 is shown as a loading control.
1408 The Plant Cell
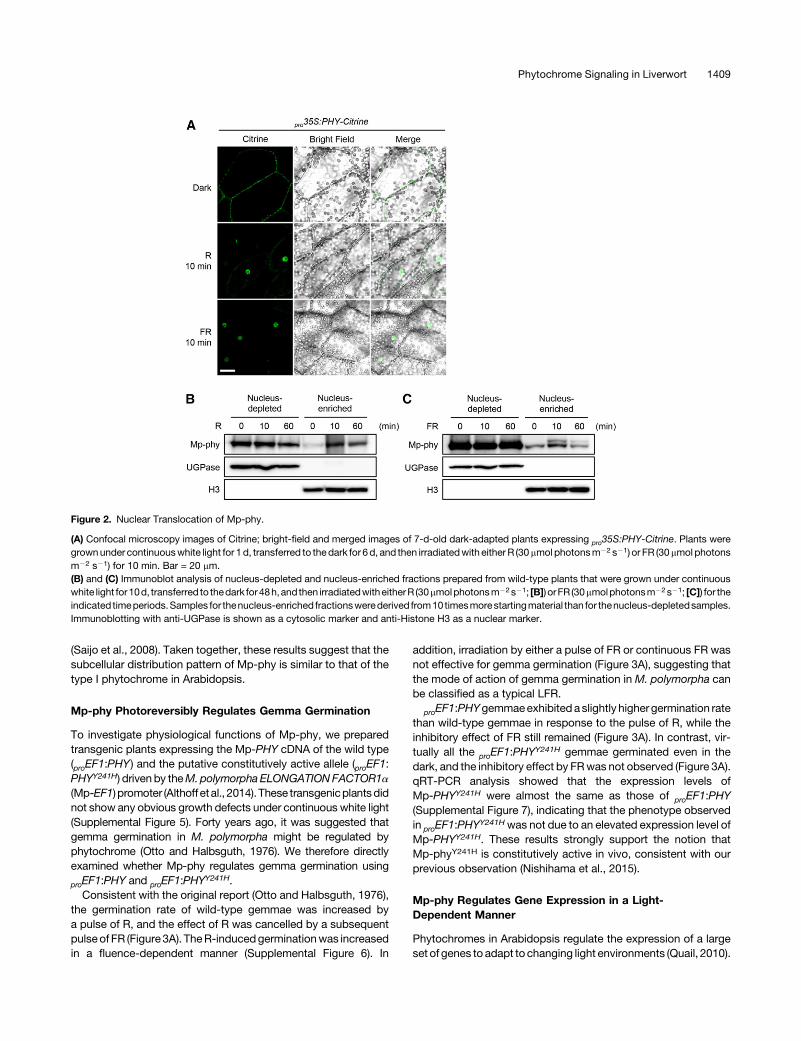
(Saijo et al., 2008). Taken together, these results suggest that thesubcellular distribution pattern of Mp-phy is similar to that of thetype I phytochrome in Arabidopsis.
Mp-phy Photoreversibly Regulates Gemma Germination
To investigate physiological functions of Mp-phy, we preparedtransgenic plants expressing the Mp-PHY cDNA of the wild type(proEF1:PHY) and the putative constitutively active allele (proEF1:PHYY241H) driven by theM.polymorpha ELONGATIONFACTOR1a(Mp-EF1) promoter (Althoff etal., 2014). These transgenicplantsdidnot show any obvious growth defects under continuous white light(Supplemental Figure 5). Forty years ago, it was suggested thatgemma germination in M. polymorpha might be regulated byphytochrome (Otto and Halbsguth, 1976). We therefore directlyexamined whether Mp-phy regulates gemma germination using
proEF1:PHY and proEF1:PHYY241H.
Consistent with the original report (Otto and Halbsguth, 1976),the germination rate of wild-type gemmae was increased bya pulse of R, and the effect of R was cancelled by a subsequentpulse of FR (Figure 3A). TheR-inducedgerminationwas increasedin a fluence-dependent manner (Supplemental Figure 6). In
addition, irradiation by either a pulse of FR or continuous FR wasnot effective for gemma germination (Figure 3A), suggesting thatthe mode of action of gemma germination in M. polymorpha canbe classified as a typical LFR.
proEF1:PHYgemmaeexhibited aslightly higher germination ratethan wild-type gemmae in response to the pulse of R, while theinhibitory effect of FR still remained (Figure 3A). In contrast, vir-tually all the proEF1:PHY
Y241H gemmae germinated even in thedark, and the inhibitory effect by FRwas not observed (Figure 3A).qRT-PCR analysis showed that the expression levels ofMp-PHYY241H were almost the same as those of proEF1:PHY(Supplemental Figure 7), indicating that the phenotype observedin proEF1:PHY
Y241Hwas not due to an elevated expression level ofMp-PHYY241H. These results strongly support the notion thatMp-phyY241H is constitutively active in vivo, consistent with ourprevious observation (Nishihama et al., 2015).
Mp-phy Regulates Gene Expression in a Light-Dependent Manner
Phytochromes in Arabidopsis regulate the expression of a largeset of genes to adapt to changing light environments (Quail, 2010).
Figure 2. Nuclear Translocation of Mp-phy.
(A) Confocal microscopy images of Citrine; bright-field and merged images of 7-d-old dark-adapted plants expressing pro35S:PHY-Citrine. Plants weregrownunder continuouswhite light for 1d, transferred to thedark for 6d, and then irradiatedwith either R (30mmol photonsm22 s21) or FR (30mmol photonsm22 s21) for 10 min. Bar = 20 mm.(B) and (C) Immunoblot analysis of nucleus-depleted and nucleus-enriched fractions prepared from wild-type plants that were grown under continuouswhite light for10d, transferred to thedark for48h,and then irradiatedwitheitherR (30mmolphotonsm22s21; [B]) orFR (30mmolphotonsm22s21; [C]) for theindicated timeperiods.Samples for thenucleus-enriched fractionswerederived from10timesmorestartingmaterial than for thenucleus-depletedsamples.Immunoblotting with anti-UGPase is shown as a cytosolic marker and anti-Histone H3 as a nuclear marker.
Phytochrome Signaling in Liverwort 1409
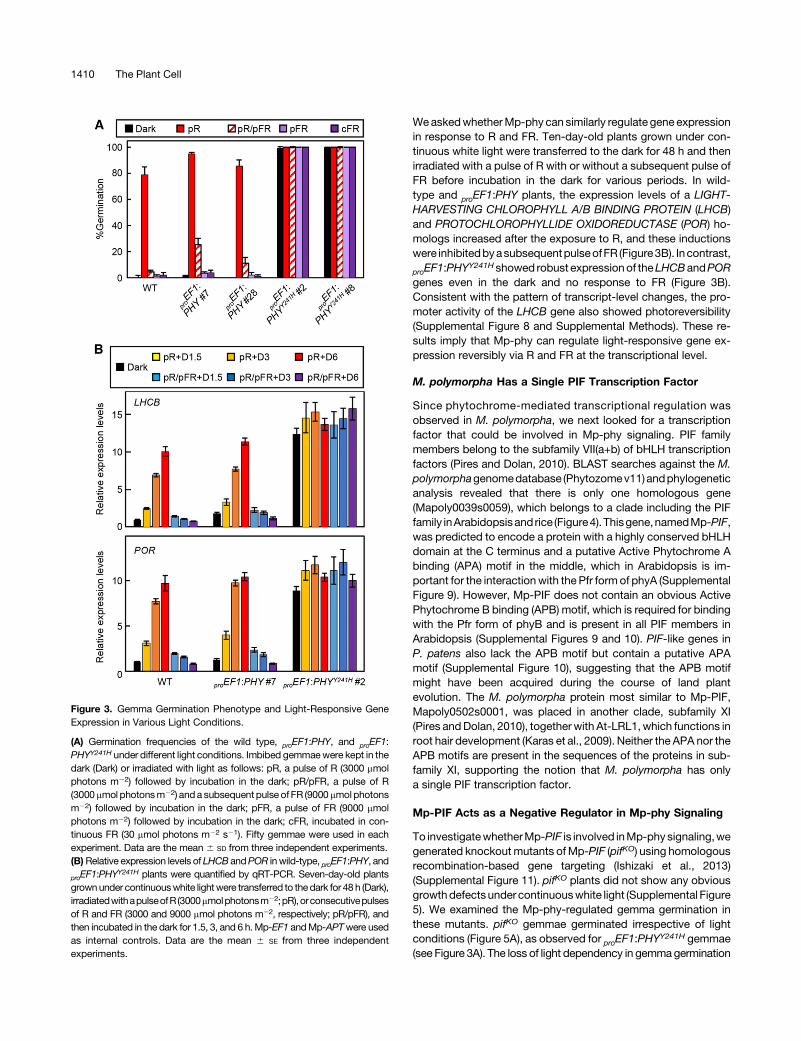
WeaskedwhetherMp-phy can similarly regulate gene expressionin response to R and FR. Ten-day-old plants grown under con-tinuous white light were transferred to the dark for 48 h and thenirradiated with a pulse of R with or without a subsequent pulse ofFR before incubation in the dark for various periods. In wild-type and proEF1:PHY plants, the expression levels of a LIGHT-HARVESTING CHLOROPHYLL A/B BINDING PROTEIN (LHCB)and PROTOCHLOROPHYLLIDE OXIDOREDUCTASE (POR) ho-mologs increased after the exposure to R, and these inductionswere inhibitedbyasubsequentpulseofFR (Figure3B). Incontrast,
proEF1:PHYY241Hshowed robust expressionof theLHCBandPOR
genes even in the dark and no response to FR (Figure 3B).Consistent with the pattern of transcript-level changes, the pro-moter activity of the LHCB gene also showed photoreversibility(Supplemental Figure 8 and Supplemental Methods). These re-sults imply that Mp-phy can regulate light-responsive gene ex-pression reversibly via R and FR at the transcriptional level.
M. polymorpha Has a Single PIF Transcription Factor
Since phytochrome-mediated transcriptional regulation wasobserved in M. polymorpha, we next looked for a transcriptionfactor that could be involved in Mp-phy signaling. PIF familymembers belong to the subfamily VII(a+b) of bHLH transcriptionfactors (Pires and Dolan, 2010). BLAST searches against the M.polymorphagenomedatabase (Phytozomev11) andphylogeneticanalysis revealed that there is only one homologous gene(Mapoly0039s0059), which belongs to a clade including the PIFfamily inArabidopsisand rice (Figure4).Thisgene,namedMp-PIF,was predicted to encode a protein with a highly conserved bHLHdomain at the C terminus and a putative Active Phytochrome Abinding (APA) motif in the middle, which in Arabidopsis is im-portant for the interactionwith the Pfr form of phyA (SupplementalFigure 9). However, Mp-PIF does not contain an obvious ActivePhytochrome B binding (APB) motif, which is required for bindingwith the Pfr form of phyB and is present in all PIF members inArabidopsis (Supplemental Figures 9 and 10). PIF-like genes inP. patens also lack the APB motif but contain a putative APAmotif (Supplemental Figure 10), suggesting that the APB motifmight have been acquired during the course of land plantevolution. The M. polymorpha protein most similar to Mp-PIF,Mapoly0502s0001, was placed in another clade, subfamily XI(Pires andDolan, 2010), together with At-LRL1, which functions inroot hair development (Karas et al., 2009). Neither the APA nor theAPB motifs are present in the sequences of the proteins in sub-family XI, supporting the notion that M. polymorpha has onlya single PIF transcription factor.
Mp-PIF Acts as a Negative Regulator in Mp-phy Signaling
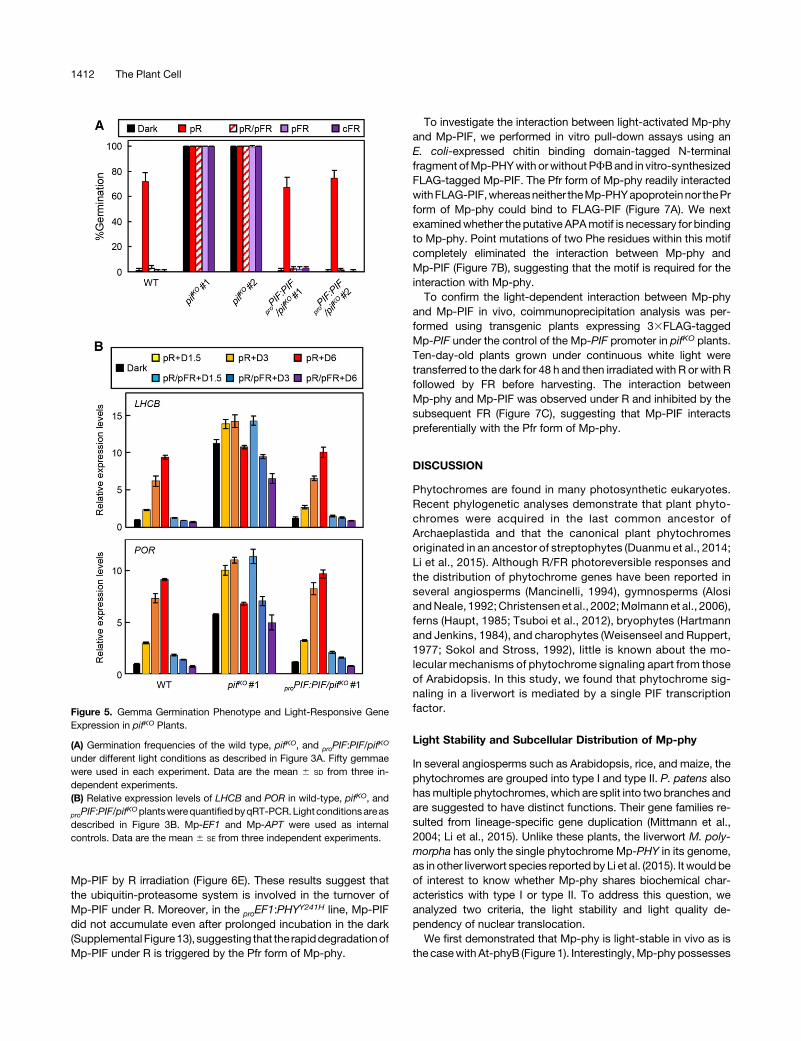
To investigatewhetherMp-PIF is involved inMp-phy signaling,wegenerated knockout mutants ofMp-PIF (pifKO) using homologousrecombination-based gene targeting (Ishizaki et al., 2013)(Supplemental Figure 11). pifKO plants did not show any obviousgrowthdefectsunder continuouswhite light (Supplemental Figure5). We examined the Mp-phy-regulated gemma germination inthese mutants. pifKO gemmae germinated irrespective of lightconditions (Figure 5A), as observed for proEF1:PHY
Y241H gemmae(see Figure 3A). The loss of light dependency in gemmagermination
Figure 3. Gemma Germination Phenotype and Light-Responsive GeneExpression in Various Light Conditions.
(A) Germination frequencies of the wild type, proEF1:PHY, and proEF1:PHYY241H under different light conditions. Imbibed gemmaewere kept in thedark (Dark) or irradiated with light as follows: pR, a pulse of R (3000 mmolphotons m22) followed by incubation in the dark; pR/pFR, a pulse of R(3000mmol photonsm22) and a subsequent pulse ofFR (9000mmolphotonsm22) followed by incubation in the dark; pFR, a pulse of FR (9000 mmolphotons m22) followed by incubation in the dark; cFR, incubated in con-tinuous FR (30 mmol photons m22 s21). Fifty gemmae were used in eachexperiment. Data are the mean 6 SD from three independent experiments.(B)Relative expression levels of LHCB andPOR inwild-type, proEF1:PHY, and
proEF1:PHYY241H plants were quantified by qRT-PCR. Seven-day-old plants
grownunder continuouswhite lightwere transferred to thedark for 48 h (Dark),irradiatedwithapulseofR (3000mmolphotonsm22;pR),orconsecutivepulsesof R and FR (3000 and 9000 mmol photons m22, respectively; pR/pFR), andthen incubated in the dark for 1.5, 3, and 6 h.Mp-EF1 andMp-APTwere usedas internal controls. Data are the mean 6 SE from three independentexperiments.
1410 The Plant Cell

in pifKO was recovered by introducing a genomic fragment of theMp-PIF locus (Figure 5A), clearly showing that Mp-PIF is the generesponsible for the altered light responsiveness of the gemmagermination in pifKO.
We then investigated the role of Mp-PIF in the Mp-phy-regulated expression of LHCB and POR. In pifKO plants, the ex-pression levels of LHCB and PORwere constitutively higher thaninwild-type plants under all the light conditions tested (Figure 5B),although the expression levels gradually decreased during in-cubation in the dark. The complemented lines showed normalexpressionpatternsofLHCBandPORas seen inwild-typeplants.Overall, these results suggest that Mp-PIF negatively regulatesgemmagermination and the expression of light-responsive genesin the Mp-phy signaling pathway.
Mp-phy Regulates Mp-PIF Protein Abundance in aLight-Dependent Manner
Since Mp-PIF appears to be involved in transcriptional regulationin Mp-phy signaling, we examined the molecular mechanism for
light-dependent regulationofgeneexpression. InArabidopsis, thePfr form of phytochromes interacts with PIFs and targets PIFs forproteasomal degradation (Al-Sady et al., 2006; Shen et al., 2007,2008). We therefore examined the light-dependent interactionbetween Mp-phy and Mp-PIF and the degradation of Mp-PIF.Ten-day-old plants grown under continuous white light weretransferred to the dark for 48 h and then irradiated with either R orFR.Mp-PIFwasalmostundetectable incontinuouswhite light, butMp-PIF accumulated within 24 h after the transition to the dark(Figure 6A). Additionally, a pulse of FR before the transition to thedark accelerated the accumulation of Mp-PIF in the dark (Figure6B), probably reflecting the faster FR-mediated photoconversionof Mp-phy to the Pr form compared with its relatively slow con-version in the dark. Mp-PIF that had accumulated in the dark-adapted thalli was rapidly degraded within 30 min by R but not byFR (Figures 6C and 6D). Transcript levels of Mp-PIF were notreduced under R (Supplemental Figure 12), suggesting that therapid reduction of Mp-PIF levels is probably due to post-translational regulation. Furthermore, treatment with MG132, aninhibitor of the 26S proteasome, reduced the degradation of
Figure 4. Phylogeny of the PIF Gene from M. polymorpha and bHLH Transcription Factors from Arabidopsis, Rice, P. patens, Chlorella vulgaris, andChlamydomonas reinhardtii.
An unrooted maximum likelihood phylogenetic tree was constructed by an alignment of sequences corresponding to amino acid residues 520 to 597 ofMp-PIF. The previously reportedC. vulgaris andC. reinhardtii bHLH sequences (Pires and Dolan, 2010) were used as the outgroup. Bootstrap proportionsare indicated at the nodes. Scale bar indicates the average number of substitutions per site. At, Arabidopsis; Os,O. sativa; Pp,P. patens; Mp orMapoly,M. polymorpha; Cv, C. vulgaris; Cr, C. reinhardtii.
Phytochrome Signaling in Liverwort 1411
Mp-PIF by R irradiation (Figure 6E). These results suggest thatthe ubiquitin-proteasome system is involved in the turnover ofMp-PIF under R. Moreover, in the proEF1:PHY
Y241H line, Mp-PIFdid not accumulate even after prolonged incubation in the dark(Supplemental Figure13), suggesting that the rapiddegradationofMp-PIF under R is triggered by the Pfr form of Mp-phy.
To investigate the interaction between light-activated Mp-phyand Mp-PIF, we performed in vitro pull-down assays using anE. coli-expressed chitin binding domain-tagged N-terminalfragment ofMp-PHYwith orwithout PFBand in vitro-synthesizedFLAG-tagged Mp-PIF. The Pfr form of Mp-phy readily interactedwithFLAG-PIF,whereasneither theMp-PHYapoproteinnor thePrform of Mp-phy could bind to FLAG-PIF (Figure 7A). We nextexaminedwhether the putative APAmotif is necessary for bindingto Mp-phy. Point mutations of two Phe residues within this motifcompletely eliminated the interaction between Mp-phy andMp-PIF (Figure 7B), suggesting that the motif is required for theinteraction with Mp-phy.To confirm the light-dependent interaction between Mp-phy
and Mp-PIF in vivo, coimmunoprecipitation analysis was per-formed using transgenic plants expressing 33FLAG-taggedMp-PIF under the control of the Mp-PIF promoter in pifKO plants.Ten-day-old plants grown under continuous white light weretransferred to the dark for 48 h and then irradiated with R or with Rfollowed by FR before harvesting. The interaction betweenMp-phy and Mp-PIF was observed under R and inhibited by thesubsequent FR (Figure 7C), suggesting that Mp-PIF interactspreferentially with the Pfr form of Mp-phy.
DISCUSSION
Phytochromes are found in many photosynthetic eukaryotes.Recent phylogenetic analyses demonstrate that plant phyto-chromes were acquired in the last common ancestor ofArchaeplastida and that the canonical plant phytochromesoriginated in an ancestor of streptophytes (Duanmu et al., 2014;Li et al., 2015). Although R/FR photoreversible responses andthe distribution of phytochrome genes have been reported inseveral angiosperms (Mancinelli, 1994), gymnosperms (AlosiandNeale, 1992;Christensen et al., 2002;Mølmannet al., 2006),ferns (Haupt, 1985; Tsuboi et al., 2012), bryophytes (Hartmannand Jenkins, 1984), and charophytes (Weisenseel and Ruppert,1977; Sokol and Stross, 1992), little is known about the mo-lecular mechanisms of phytochrome signaling apart from thoseof Arabidopsis. In this study, we found that phytochrome sig-naling in a liverwort is mediated by a single PIF transcriptionfactor.
Light Stability and Subcellular Distribution of Mp-phy
In several angiosperms such as Arabidopsis, rice, and maize, thephytochromes are grouped into type I and type II. P. patens alsohasmultiple phytochromes, which are split into two branches andare suggested to have distinct functions. Their gene families re-sulted from lineage-specific gene duplication (Mittmann et al.,2004; Li et al., 2015). Unlike these plants, the liverwort M. poly-morpha has only the single phytochrome Mp-PHY in its genome,as in other liverwort species reported by Li et al. (2015). It would beof interest to know whether Mp-phy shares biochemical char-acteristics with type I or type II. To address this question, weanalyzed two criteria, the light stability and light quality de-pendency of nuclear translocation.We first demonstrated that Mp-phy is light-stable in vivo as is
the casewith At-phyB (Figure 1). Interestingly,Mp-phy possesses
Figure 5. Gemma Germination Phenotype and Light-Responsive GeneExpression in pifKO Plants.
(A) Germination frequencies of the wild type, pifKO, and proPIF:PIF/pifKO
under different light conditions as described in Figure 3A. Fifty gemmaewere used in each experiment. Data are the mean 6 SD from three in-dependent experiments.(B) Relative expression levels of LHCB and POR in wild-type, pifKO, and
proPIF:PIF/pifKOplantswerequantifiedbyqRT-PCR.Light conditionsareas
described in Figure 3B. Mp-EF1 and Mp-APT were used as internalcontrols. Data are the mean 6 SE from three independent experiments.
1412 The Plant Cell

a Lys residue corresponding to Lys-206 of At-phyA, which isthe main ubiquitination site for its proteasomal degradation(Rattanapisit et al., 2016). The type II-like photostability ofMp-physuggests that the mechanism for rapid degradation via theubiquitination of Lys-206 of At-phyA could have been acquiredduring the course of functional differentiation of phytochromesfollowing gene duplication.We also demonstrated that Mp-phy rapidly translocates into
the nucleus in response to either R or FR (Figure 2). AlthoughAt-phyA translocates into the nucleus under both R and FR, itaccumulates only under FR due to its instability under R (Okaet al., 2012). However, Mp-phy is comparatively stable (Figure1B), which may therefore permit its nuclear accumulation underR as well as FR. FHY1 and FHL are known to play an impor-tant role in the nuclear accumulation of At-phyA under FR(Hiltbrunner et al., 2006; Genoud et al., 2008; Rausenbergeret al., 2011). Recently, a functional homologof FHY1 inP. patenswas shown to be similarly involved in the regulation of theFR-induced nuclear translocation of phytochromes (Possartand Hiltbrunner, 2013). These findings suggest that the M.polymorpha FHY1 homolog might regulate the nuclear trans-location of Mp-phy under FR. Taken together, Mp-phy pos-sesses combined properties of type I and type II phytochromes.It has been reported that the phytochrome in the charophyteMesotaenium caldariorum does not fit either the type I or type IIclassifications (Morand et al., 1993). Our results also supportthe hypothesis that the phytochrome in the early diver-gent lineage of streptophytes represents a distinct class ofphytochrome.
Mp-phy-Mediated Responses
Mp-phy regulated gemma germination and light-responsive geneexpression in an R-dependent manner, and the inductive effect ofR was clearly cancelled by a subsequent pulse of FR (Figure 3),indicating that Mp-phy is responsible for the LFR, as are type IIphytochromes in other land plants. Interestingly, neither a pulse ofFR nor continuous FR was effective for gemma germination andthe induction of LHCB expression, although the nuclear accu-mulation of Mp-phy was observed under FR (Figures 2 and 3),suggesting that these responses are not simply regulated by thesubcellular distribution of Mp-phy. In Arabidopsis, the nucleus-localized phyA regulates VLFR and FR-HIR under FR (Shinomuraet al., 1996, 2000). FR-induced nuclear accumulation of phyto-chromeandHIR-like responsesareobserved inP.patens (Possartand Hiltbrunner, 2013). In this study, we could not elucidate thesignificance of the FR-induced nuclear accumulation of Mp-phy.In our previousobservation, however,we found that FR is requiredfor the growth phase transition ofM. polymorpha (Chiyoda et al.,2008; Kubota et al., 2014). The nucleus-localized Mp-phy under
Figure 6. R-Dependent Degradation of Mp-PIF.
Protein accumulation levels of Mp-PIF were examined by anti-Mp-PIFimmunoblot analyses using nucleus-enriched fractions fromwild-type andpifKO plants.(A) to (D) Plants were grown under continuous white light for 10 d andtransferred to either of the following light conditions: (A) darkness for theindicated periods (h); (B) darkness for the indicated periods (h) with andwithout preirradiation of a FRpulse (9000mmol photonsm22; pFR); (C)and (D)darkness for 48 h followed by irradiation with R (30 mmol photonsm22 s21;[C]) or FR (30 mmol photons m22 s21; [D]) for the indicated periods (min).
(E)Plantswere grown under continuouswhite light for 10 d and transferredto thedark for 48 h (D) and then irradiatedwithR (30mmol photonsm22 s21)for 30 min (R). Plants were treated with 50 mM MG132 or DMSO (solventcontrol) for 30 min before irradiation with R. Immunoblotting with anti-Histone H3 is shown as a loading control.
Phytochrome Signaling in Liverwort 1413

FR might be involved in the regulation of the growth phasetransition in M. polymorpha.
We detected a slower migrating band of the Mp-phy protein inthe nucleus under FR, but not under R (Figures 2B and 2C). It hasbeen reported that nucleus-localized At-phyA is phosphorylated
specifically under FR and that this phosphorylation may enhancethe association with COP1 (Saijo et al., 2008), although thefunctional importance of this phosphorylation still remains un-known. Mp-phy might undergo a similar regulation via post-translational modification under FR as observed in At-phyA.
Phytochrome-PIF-Mediated Transcriptional Regulation forR Signaling
In Arabidopsis, PIFs have been shown to play an essential role inphytochrome signaling (Leivar and Monte, 2014). However, re-ports onPIF functions in other plant species are very limited. Here,we demonstrated that the only PIF transcription factor in M.polymorpha, Mp-PIF, functions in phytochrome signaling and isinvolved in the negative regulation of the LFR, affecting gemmagermination probably via light-dependent gene expression (Fig-ures 4 and 5). At-PIFs accumulate in the dark and are degradedupon irradiation with R. In contrast to Arabidopsis, Os-PIL1 hasbeen shownnot to interactwithOs-phyBandhasbeen suggestedto be light-stable in rice (Todaka et al., 2012). Mp-PIF accu-mulated in the dark andwas degraded selectively in response toR possibly via the interaction with the Pfr form of Mp-phy(Figures 6 and 7), suggesting that Mp-PIF is inhibited by light-activated Mp-phy through a molecular mechanism that is alsofound in Arabidopsis. These findings indicate that liverworts, anearly divergent lineage of land plants, had already acquired thetranscriptional regulation system mediated by the phytochrome-bHLH interaction and that the light-dependent degradation ofthe bHLH would be the underlying mechanism of R signaling inland plants. In Arabidopsis, PIFs undergo phosphorylation be-fore their degradation (Al-Sady et al., 2006; Shen et al., 2007,2008). The stimulus-induced phosphorylation and subsequentdegradationof substratesbySCFcomplexeshasbeenobservedin many eukaryotes (Skaar et al., 2013). Further analysis willclarify whether Mp-PIF undergoes phosphorylation before itsdegradation.Our findings highlight the similarities and the differences in PIF-
mediated phytochrome signaling in M. polymorpha and Arabi-dopsis (Figure 8). Under R, Mp-phy translocates into the nucleus(Figure 2) and interacts with Mp-PIF via the APA motif (Figure 7).Mp-PIF is rapidly degraded via the 26S proteasome, probablyfollowing the interactionwith thePfr formofMp-phy (Figures6and7; Supplemental Figure 13). In Arabidopsis, although R inducesnuclear translocationof bothphyAandphyB, phyBpredominantlylocalizes in the nucleus because light-labile phyA is rapidly de-graded in both the cytoplasm and nucleus (Toledo-Ortiz et al.,2010; Oka et al., 2012). phyB interacts with PIFs via the APBmotifand triggers the phosphorylation and proteasomal degradation ofPIFs (Khanna et al., 2004; Al-Sady et al., 2006; Shen et al., 2007,2008; Lorrain et al., 2008). These findings suggest that a commonstrategy of interaction and degradation is utilized under R in M.polymorpha and Arabidopsis, although it is mediated by the APAmotif of Mp-PIF in M. polymorpha. However, the signalingmechanism under FR is different in these two species (Figure 8B).In M. polymorpha, while Mp-phy translocates into the nucleus(Figure 2), Mp-PIF is not degraded under FR (Figure 6). Thesefindings suggest that a very low ratio of the Pfr form of Mp-phyunder FR is not sufficient to induce the degradation of Mp-PIF;
Figure 7. R-Dependent Interaction of Mp-PIF with Mp-phy.
(A) and (B) Pull-down assays of Mp-phy and Mp-PIF. A CBD-taggedN-terminal fragment of Mp-PHY with or without PFB was expressed inE. coli and purified. FLAG-PIF and FLAG-PIFF297A,F300A(mAPA) weresynthesized by in vitro transcription/translation. FLAG-PIF was incubatedwith PHY(N612)-CBD apoprotein (Apo) or the Pfr and Pr forms ofphy(N612)-CBD with PFB (A), and FLAG-PIF and FLAG-PIFF297A,F300A
were incubated with the Pfr and Pr forms of phy(N612)-CBDwith PFB (B).The proteins bound to the chitin resin were probed with anti-Mp-PHY oranti-FLAG antibodies.(C) Coimmunoprecipitation of Mp-phy with Mp-PIF. pifKO and proPIF:PIF-33FLAG/pifKO lines were grown under continuous white light for 10 d andtransferred to the dark for 48 h. Dark-adapted plants were irradiatedwith R(9000 mmol photons m22) or irradiated with R followed by FR (10,800 mmolphotons m22; R/FR). The proteins in the nucleus-enriched fractions wereimmunoprecipitated with an anti-FLAG antibody, and the immunoprecipitateswere probed with anti-Mp-PHY or anti-FLAG antibodies.
1414 The Plant Cell

therefore, the physiological response and gene expression do notoccur (Figure5).Bycontrast, inArabidopsis, anextremely lowratioof thePfr formofphyAcan induce thedegradationofPIF1andPIF3under FR (Park et al., 2004; Al-Sady et al., 2006; Shen et al., 2008).This strong light sensitivity of PIF degradation is considered to beresponsible for seed germination under VLFR condition (Oh et al.,2004;Shenetal.,2008).GiventhatFR-inducedVLFRisnotobservedin M. polymorpha (Figures 3 and 5), high light sensitivity of PIFdegradation may be required for the VLFR. PIF4 and PIF5 are notdegradedunderFR,consistentwith the fact that theydonothave theAPA motif (Lorrain et al., 2008; Leivar and Monte, 2014). PIF4 andPIF5 regulate the shade avoidance response under a low R-to-FRratio (Lorrain et al., 2008). The acquisition of two types of phyto-chromes and functionally diverged PIFsmay have allowed complexresponses to various R to FR ratios during land plant evolution.
Light regulation of phytochrome subcellular localization hasbeen reported in the prasinophyte green algaMicromonas pusilla,
which suggests that light-dependent nuclear translocation ofphytochrome predates the divergence of streptophytes andprasinophytes (Duanmu et al., 2014). However, the signalingmechanisms might be different. Phytochromes in prasinophytescontain an intact histidine kinase domain with light-dependent ki-nase activity, which couldmediate phytochrome signaling (Duanmuet al., 2014). In contrast, all streptophytes examined so far havephytochromes that lack a His residue critical for phosphorelay,suggesting the establishment of distinct signaling mechanisms instreptophytes. PIF-mediated transcriptional regulation could rep-resent one such mechanism, and it may have contributed to theacquisition of diversity in phytochrome signaling. Our results showthat the PIF-mediated regulatory mechanism of R signaling hadalready been established in a liverwort and perhaps in the commonancestor of land plants. It would be of interest to find out whetheracquisition of the phytochrome-PIF system contributed to adap-tation to a terrestrial environment.
Figure 8. A Model of Phytochrome Signaling through PIFs in M. polymorpha and Arabidopsis.
Model of phytochrome signaling under R (A) and FR (B). Schematic illustration forM. polymorpha (left) and Arabidopsis (right). Blue and red circles indicatethe APA motif and the APB motif, respectively. Dotted lines indicate degradation. Dashed lines indicate the nucleus.
Phytochrome Signaling in Liverwort 1415

METHODS
Plant Materials and Growth Conditions
Female andmaleMarchantia polymorpha accessions, Takaragaike-2 (Tak-2)and Takaragaike-1 (Tak-1), respectively (Ishizaki et al., 2008), were culturedaseptically on half-strength Gamborg’s B5 medium (Gamborg et al., 1968)containing 1%sucrose and 1.2%agar under 50 to 60 mmol photonsm22 s21
continuous white light with a cold cathode fluorescent lamp (OPT-40C-N-L;Optrom) at 22°C. F1 spores were obtained by crossing Tak-2 and Tak-1.
Light Sources
Rwasgivenbya redLEDilluminator (MIL-R18,SANYOElectric;peakemissionat 657nm,half-bandwidthof 12nm). FRwasgivenbya far-redLED illuminator(MIL-IF18, SANYO Electric; peak emission at 734 nm, half-bandwidth of13 nm). The light intensity was measured by an LI-250A light meter (LI-CORBiosciences)equippedwithanSKR110red/far-redsensor (Skye instruments).
Cloning of Mp-PHY
Partial phytochrome gene fragments were obtained by sequencing theM.polymorpha EST clone rlwb02n17, which showed significant similarity tothe 39-half of the phytochrome coding sequence in Marchantia paleacea(Suzuki et al., 2001) (GenBank accession number BAB39687). The se-quence of the 59-terminal end was obtained by 59RACE. For 59RACE, totalRNA was isolated from;100 mg ofM. polymorpha thallus tissue using anRNeasy plant mini kit (Qiagen). The first-strand cDNA was synthesizedusing SuperScript III (Thermo Fisher Scientific) and the gene-specificprimer 59-ACCTCCATGTTCAGCT-39. The first-strand cDNA was tailedwith poly dC by terminal deoxynucleotidyl transferase (TaKaRa). 59RACE-PCR of dC-tailed cDNA was performed using the nested gene-specificprimer 59-GCAGGCCGAATACCTGCATG-39 and abridged anchor primer(Thermo Fisher Scientific). Another round of nested PCR was performedwith the nestedgene-specificprimer 59-AGGAATTCACAGGCTGAACG-39and abridged universal amplification primer (Thermo Fisher Scientific).Genomic DNA fragments corresponding to Mp-PHY cDNA were isolatedby screening a PAC genomic library using the primer pair 59-GCAGT-TACGAAGGATTTTGG-39 and 59-TTCTCTGGATGTGATCATGC-39, asdescribed previously (Okada et al., 2000). After SpeI digestion of the PACclone, pMM24-13G6, 7-kb fragments were subcloned and sequenced.
Phylogenetic Analysis
For alignments of amino acid sequences, we used the Muscle program(Edgar, 2004) implemented in Geneious software (version 8.1.3; Biomatters;http://www.geneious.com/) with default parameters. For phylogeneticanalyses, we used the bHLH domain of PIFs and other bHLH transcriptionfactors. The alignment is available as Supplemental Data Set 1. Thephylogenetic treewas constructed using thePhyMLprogramversion 3.2.0(Guindon et al., 2010) using the JTT model and four categories of ratesubstitution. Tree topology, branch length, and substitution rates wereoptimized, and the tree topologywas searched using the nearest-neighborinterchange and subtree pruning and regrafting methods. Bootstrapproportions were computed from 1008 trials. The Chlorella vulgaris andChlamydomonas reinhardtii bHLH sequences reported by Pires andDolan(2010) were used as the outgroup.
Generation of Transgenic Lines
Toobtain the overexpression lines ofMp-PHY, the coding sequence (CDS)ofMp-PHYwasamplifiedbyRT-PCR fromwild-typemRNAwith theprimerpair 59-CACCATGTCGACCACCAAGGTCAC-39 and 59-TTATTTCA-CACTGCCAGCAT-39 and cloned into pENTR/D-TOPO vector (Thermo
Fisher Scientific). The cloned sequence was then transferred to thepMpGWB103vector (Ishizaki etal., 2015) togeneratepMH01.Toobtain theoverexpression linesofMp-PHYY241H, theplasmid carryingMp-PHYcDNAwas mutagenized using a Phusion site-directed mutagenesis kit (NewEngland Biolabs) with the primer pair 59-CACACAAATTCCATGAGGAT-39and 59-CCATAACCCGGTCGTAACCT-39. The cloned sequence was thentransferred to the pMpGWB103 vector to generate pMH03. To obtain theoverexpression lines of phy-Citrine and phyY241H-Citrine fusion proteinsunder the control of the 35S promoter, the CDSs of Mp-PHY andMp-PHYY241Hwithout thestopcodonwereamplifiedbyRT-PCR fromwild-typecDNAwith theprimerpair 59-CACCATGTCGACCACCAAGGTCAC-39and 59-TTTCACACTGCCAGCATCAT-39 and cloned into pENTR/D-TOPOvector. The cloned sequences were then transferred to the pMpGWB106vector (Ishizaki et al., 2015) to generate pMH06 and pMH07. pMH01,pMH03, pMH06, and pMH07 were introduced into F1 sporelings usingAgrobacterium tumefaciens C58C1 GV2260 as previously described(Ishizaki et al., 2008).
To obtain the knockout lines of Mp-PIF, the 59- and 39-homologousarms were amplified from the wild-type genomic DNA by PCR withthe primer pairs 59-CTAAGGTAGCGATTAATGTAGGCGTACTCG-TACTC-39/59-CCGGGCAAGCTTTTAATCTCTTCGAAGCTCATGTG-39and 59-AACACTAGTGGCGCGAACACCTCGAAAGAGAC-3 9/59-TTATCCCTAGGCGCGTGCAAGGAGTCTCAAGTTGG-39, respectively.The amplified 59- and 39-homologous arms were cloned into the PacI andAscI sites, respectively, of thepJHY-TMp1 vector (Ishizaki et al., 2013)withan In-Fusion HD cloning kit (Clontech) to generate pIN005. pIN005 wasintroduced into F1 sporelings. Screening of targeted lines was performedas described previously (Ishizaki et al., 2013). Gene-specific primers usedin the screening are diagrammed in Supplemental Figure 11. To obtain thecomplementation lines of pifKO, the promoter and CDS of Mp-PIF wereamplified from wild-type genomic DNA by PCR with the primer pairs59-CACCTTACGAGGCAGGAGCTC-39/59-GGGAACGAGGTGACT-CATGG-39 and 59-TGCGAGTCTTTGGTGCTAGAG-39/59-TTTGGCG-CGCCTGATCATATTCTGTACTTGG-39, respectively. The amplifiedpromoter fragment was cloned into pENTR/D-TOPO vector. The PmeI-AscIsites of the resulting plasmid carrying the promoter fragment were used forcloning the amplified CDS after digestion with PmeI and AscI. The clonedsequence containing the promoter and the CDS was then transferred to thepMpGWB301 vector (Ishizaki et al., 2015) to generate pIN032. To obtain the33FLAG-tagged Mp-PIF-expressing lines, the CDS of the Mp-PIF withoutthe stop codon was amplified from the wild-type genomic DNA by PCRwith the primer pair 59-CACCTTACGAGGCAGGAGCTC-39 and 59-CATGCTGCTCGGAAGATGA-39 and cloned into pENTR/D-TOPO vector.The cloned sequence was then transferred to the pMpGWB309 vector(Ishizaki et al., 2015) to generate pIN055. pIN032 and pIN055 were in-troduced into the pifKO lines using regenerating thalli as previously de-scribed (Kubota et al., 2013).
Gemma Germination Assay
Gemmae were plated on half-strength Gamborg’s B5 medium containing1% sucrose and 1.2% agar under green light and then irradiated with FR(30 mmol photons m22 s21) for 15 min to inactivate Mp-phy. Gemmaeimbibed in the dark for 1 d were kept under various light conditions andscored for germination in 6 d.
RNA Extraction and RT-PCR
Frozen plants were ground into a fine powder in liquid nitrogen by shakingfor 1minwith ametal cone in acollection tubeusingaMulti-BeadsShocker(Yasui Kikai). Total RNA was extracted from whole plants using TRIzolreagent (Thermo Fisher Scientific) and treated with RNase-Free DNase(Qiagen) following the manufacturers’ instructions. Reverse transcriptionwas performed with;1 mg of RNAwith an oligo(dT) primer using ReverTra
1416 The Plant Cell

Ace (Toyobo) following the manufacturer’s instructions. QuantitativeRT-PCR was performed with a CFX96 real-time PCR detection system(Bio-Rad) using SYBR Green I Nucleic Acid Gel Stain (Lonza) to monitordouble-stranded DNA synthesis. Mp-EF1 and Mp-APT were used as in-ternal controls for normalization of the PCR (Saint-Marcoux et al., 2015).The following thermal cycling profile was used for all PCRs: 95°C for30 s, 40 cycles of 95°C for 5 s, and 60°C for 30 s. Primer pairs used foreach gene were as follows: for Mp-PHY, 59-GGGCTGGGTAGGAAT-CAAGG-39 and 59-CCCTGAACTCAAAATGAACAACG-39; for Mp-PIF,59-CAGCCGATGAGTATGGATGC-39 and 59-AGATGATGGAGC-GAATGCTG-39; for LHCB, 59-GGGATTTCGTTCAGTGGATGG-39and 59-GCTCGCTCCAATTCCGTTC-39; for POR, 59-GTCTGATGAAGC-GAGCAACC-39 and 59-GACGGAACTCTAAGCGAAACC-39; for Mp-EF1,59-AAGCCGTCGAAAAGAAGGAG-39 and 59-TTCAGGATCGTCCGT-TATCC-39; for Mp-APT, 59-CGTCGTTGTTGGAATTGAAGC-39 and 59-TTCTTGGGCTTTCGTTGAGG-39. All samples were amplified in triplicate,and the relative expression levels of the target genes were calculated asdescribed (Vandesompele et al., 2002).
Production of Anti-Mp-PHY and Anti-Mp-PIF Antibodies
For the production of the Mp-PHY antibody, we used an N-terminalfragment of Mp-PHY spanning amino acids 1 to 612 fused to the chitinbinding domain (CBD) at the C terminus, and after transformation ofEscherichia coli strain ER2566, the fusion protein was purified followinga previously described protocol (Mukougawa et al., 2006). For the pro-duction of the Mp-PIF antibody, a nonconserved fragment of the Mp-PIFprotein including amino acids 1 to 257 with a 63His affinity tag at the Nterminus was expressed in E. coli strain Rosetta2(DE3). The E. coli cellsexpressing 63His-PIF(N257) were grown at 37°C to log phase and theninduced with 1 mM IPTG for 24 h at 15°C. Cells were collected and re-suspended in lysis buffer (20mMTris-HCl, pH8.0, 150mMNaCl, 10% [v/v]glycerol, 1 mM DTT, and 0.1 mg/mL lysozyme). The cells were lysed bysonicationandcentrifugedat16,000g for15minat4°C.The insolublepelletwas resuspended in PBS containing 8 M urea and purified by affinitychromatography using Ni-NTA agarose (Qiagen). The recombinant pro-teins were used to raise rabbit polyclonal antibodies (KIWA LaboratoryAnimals).
Protein Extraction and Immunoblot Analysis
Frozenplantswereground intoafinepowder in liquidnitrogenwithamortarand pestle andmixedwith equal volumes of a lysis buffer (50mMTris-HCl,pH 8.0, 150 mMNaCl, 1 mM DTT, 1% [v/v] Triton X-100, 1 mM PMSF, 13Complete Protease Inhibitor [Roche]). The homogenates were centrifugedat 16,000g for 15 min at 4°C. The supernatants were collected, mixed with23 Laemmli sample buffer (100 mM Tris-HCl, pH 6.8, 4% [w/v] SDS, 10%[v/v] 2-mercaptoethanol, and 20% [v/v] glycerol), and boiled at 95°C for5 min. Samples were separated by SDS-PAGE on a 6% acrylamide gel forMp-phy and 15% for Histone H3 and transferred onto polyvinylidenefluoride membranes (Bio-Rad Laboratories). For primary antibodies,polyclonal anti-Mp-PHY (see above) and anti-Histone H3 (Active Motif)were diluted 1:2000 and 1:5000, respectively. For the secondary antibody,ECL Rabbit IgG, HRP-linked whole Ab (GE Healthcare) was diluted1:10,000. Blots were visualized with ECL Prime reagent (GE Healthcare)and ImageQuant LAS 4010 (GE Healthcare).
Subcellular Fractionation
The following procedure was performed under green light. Plants (0.3 g)were harvested and ground into a fine powder in liquid nitrogen andmixedwith 1mLof lysis buffer (20mMTris-HCl, pH8.0, 25% [v/v] glycerol, 10mMMgCl2, 250 mM sucrose, 5 mM DTT, 1 mM PMSF, 20 mM MG132, 13Complete EDTA-Free Protease Inhibitor). The homogenate was filtered
through a 70-mm cell strainer (BD Biosciences), and the flow-through wascentrifuged at 1500g for 10 min at 4°C. The supernatant consisting of thenucleus-depleted fractionwascollectedandmixedwith23Laemmli bufferand boiled at 95°C for 5min. The pellet waswashed two timeswith 1mL ofwash buffer (20 mM Tris-HCl, pH 8.0, 25% [v/v] glycerol, 2.5 mM MgCl2,0.2% [v/v] Triton X-100, 5 mM DTT, 1 mM PMSF, 20 mM MG132, 13Complete EDTA-Free Protease Inhibitor). The final pellet was mixed with50 mL of extraction buffer (20 mM Tris-HCl, pH 8.0, 5 mM EDTA, 150 mMNaCl, 1% [v/v] Triton X-100, 0.5% [w/v] sodium deoxycholate, 0.1% [w/v]SDS, 5 mMDTT, 1 mMPMSF, 20 mMMG132, and 13Complete ProteaseInhibitor) and incubated on ice for 10 min. The extract consisting of thenucleus-enriched fractionwasmixedwith 23 Laemmli buffer and boiled at95°C for 5min. Each fractionwas centrifuged at 16,000g for 15min at roomtemperature. The supernatants were subjected to immunoblot analysis.For primary antibodies, anti-Mp-PHY and anti-Histone H3 were used asdescribed above. Anti-Mp-PIF was diluted 1:2000.
In Vitro Pull-Down Assay
For bait proteins, an N-terminal fragment of Mp-PHY fused to the CBD atthe C terminus with or without PFB was expressed in E. coli as describedabove. The recombinant protein was incubated with chitin resin (NewEngland Biolabs) at 4°C for 1 h andwashed three times with binding buffer(20mMTris-HCl, pH8.0, 150mMNaCl, 0.1% [v/v] TritonX-100, and0.01%[w/v] BSA). The protein concentrations of affinity-bound Mp-PHY apo-protein and holoprotein were estimated by immunoblot analysis. For preyproteins, FLAG-tagged Mp-PIF and Mp-PIFF297A,F300A were synthesizedby an in vitro transcription and wheat germ cell-free translation system(BioSieg) according to the manufacturer’s instructions. The reactionmixture was centrifuged at 15,000g for 10 min, and the supernatant wasused for the pull-down assays after estimation of the product yield byimmunoblot analysis. The bait phy(N612)-CBD holoprotein was irradiatedwithapulseofR (9000mmolphotonsm22;Pfr) or apulseofR followedbyFR(10,800 mmol photons m22; Pr) before use. Ten microliters of the affinity-boundPHY(N612)-CBDapoprotein or holoprotein (Pfr andPr) and10mL ofthe in vitro-translated FLAG-PIF or FLAG-PIFF297A,F300A were mixed with80 mL of binding buffer and incubated at 4°C for 1 h in the dark. Afterwashing the resin three timeswith 1mL of binding buffer under green light,the bound proteins were subjected to immunoblot analysis with anti-Mp-PHY antibody (used as described above) and anti-FLAG M2 antibody(1:2000 dilution).
Coimmunoprecipitation Assay
The following procedure was performed under green light. Plants (2.0 g)were harvested and ground into a fine powder in liquid nitrogen andmixedwith 6 mL of lysis buffer as described in the subcellular fractionationprocedures. After subcellular fractionation as described above, the pelletcontaining nuclei was resuspended with 1 mL of extraction buffer (20 mMTris-HCl, pH 8.0, 5 mM EDTA, 150 mM NaCl, 1% [v/v] Triton X-100, 0.5%[w/v] sodium deoxycholate, 5 mM DTT, 1 mM PMSF, 50 mM MG132, and13CompleteProtease Inhibitor) and incubatedon ice for10min.The lysatewas centrifuged at 16,000g for 15 min at 4°C, and the supernatant wassubjected to immunoprecipitation.Coimmunoprecipitationwasperformedusing the mMACS DYKDDDDK isolation kit (Miltenyi Biotec) according tothe manufacturer’s instructions. The immunoprecipitates were subjectedto immunoblot analysiswith anti-Mp-PHYandanti-FLAGM2antibodies asdescribed above.
Fluorescence Microscopy
Gemmaeweregrownundercontinuouswhite light for 1d, transferred to thedark for 6 d, and then irradiatedwith eitherRor FR for 10min. Fluorescencederived fromCitrine was detected with an Olympus FV1000 confocal laser
Phytochrome Signaling in Liverwort 1417

scanning microscope equipped with a GaAsP detector using a 515-nmlaser for excitation, and a detection window in the range of 535 to 565 nm.
Accession Numbers
The sequences of the Mp-PHY and Mp-PIF genes are available in DDBJunder the following accession numbers: Mp-PHY (LC093264) andMp-PIF(LC093265). The accession numbers of the analyzed proteins in Figure 4are as follows: PIF1 (AEC06979), PIF3 (AEE28458), PIF4 (AEC10198), PIF5(AEE79871), PIF6 (AEE80307), PIF7 (AED97445), PIF8 (ABG25060), SPT(ABH04569), ALC (ABI49493), bHLH23 (AEE85545), bHLH56 (AEE85546),bHLH119 (AEE85547), bHLH127 (AEE85548), and LRL1 (AEC07551) fromArabidopsis; Os01g0286100 (BAF04689), Os03g0639300 (ABF97814),Os03g0782500 (ABF99196), Os05g0139100 (BAS92179), Os07g0143200(BAF20790), Os12g0610200 (ABA99363), andOs02g0795800 (XP_015627343)from rice; and Pp001G155100 (Phpat.001G155100), Pp002G038300(Phpat.002G038300), Pp014G091800 (Phpat.014G091800), Pp017G085300(Phpat.017G085300), and Pp017G062100 (Phpat.017G062100) fromPhyscomitrella patens.
Supplemental Data
Supplemental Figure 1. Alignment of amino acid sequences ofphytochromes from M. polymorpha and M. paleacea.
Supplemental Figure 2. Alignment of amino acid sequences ofphytochromes from M. polymorpha and Arabidopsis.
Supplemental Figure 3. Spectral properties of recombinant Mp-phyand Mp-phyY241H.
Supplemental Figure 4. Light-independent nuclear localization ofphyY241H-Citrine.
Supplemental Figure 5. Phenotypes of wild-type, proEF1:PHY, proEF1:PHYY241H, and pifKO plants.
Supplemental Figure 6. Fluence-response relationship for gemmagermination.
Supplemental Figure 7. Transcript levels of Mp-PHY in transgeniclines.
Supplemental Figure 8. Photoreversible promoter activity of theLHCB gene.
Supplemental Figure 9. Alignment of amino acid sequences ofMp-PIF and PIFs from Arabidopsis.
Supplemental Figure 10. Sequence alignments of the APA and APBmotifs in PIFs.
Supplemental Figure 11. Generation of pifKO plants by homologousrecombination.
Supplemental Figure 12. Transcript levels of Mp-PIF during thetransition from darkness to R.
Supplemental Figure 13. Levels of Mp-PIF protein in transgenic lines.
Supplemental Data Set 1. Text file of the alignment used for thephylogenetic analysis in Figure 4.
Supplemental Methods.
Supplemental References.
ACKNOWLEDGMENTS
We thank Tokitaka Oyama and Tomoaki Muranaka (Kyoto University) forassistanceof luciferasebioluminescenceassayandJamesA.Hejna (KyotoUniversity) for critical reading of the manuscript. This work was supported
by KAKENHI Grant-in-Aids for Scientific Research on Innovative Area(Nos. 23120516 and 25113009 to T.K.), for Scientific Research (B) (No.26291059 to T.K.), for Scientific Research (C) (No. 24570048 to R.N.),for Young Scientists (B) (No. 22770035 to K. Ishizaki), and ResearchFellowships for Young Scientists (24-7049 to K. Inoue) from the JapanSociety for the Promotion of Science.
AUTHOR CONTRIBUTIONS
K. Inoue, R.N., H.K., M.H., R.M., K. Ishizaki, and T.K. designed research.M.N. and Y.T. contributed reagents. K. Inoue, H.K., M.H., and R.M.performed research. K. Inoue, R.N., H.K., M.H., R.M., K. Ishizaki, andT.K. analyzed data. K. Inoue, R.N., K. Ishizaki, and T.K. wrote the article.
Received December 29, 2015; revised May 18, 2016; accepted May 30,2016; published June 1, 2016.
REFERENCES
Alosi, M.C., and Neale, D.B. (1992). Light- and phytochrome-mediatedgene expression in Douglas-fir seedlings. Physiol. Plant. 86: 71–76.
Al-Sady, B., Ni, W., Kircher, S., Schäfer, E., and Quail, P.H. (2006).Photoactivated phytochrome induces rapid PIF3 phosphorylationprior to proteasome-mediated degradation. Mol. Cell 23: 439–446.
Althoff, F., Kopischke, S., Zobell, O., Ide, K., Ishizaki, K., Kohchi,T., and Zachgo, S. (2014). Comparison of the MpEF1a and CaMV35promoters for application in Marchantia polymorpha overexpressionstudies. Transgenic Res. 23: 235–244.
Casal, J.J., Sánchez, R.A., and Botto, J.F. (1998). Modes of action ofphytochromes. J. Exp. Bot. 49: 127–138.
Chen, M., Tao, Y., Lim, J., Shaw, A., and Chory, J. (2005). Regulationof phytochrome B nuclear localization through light-dependentunmasking of nuclear-localization signals. Curr. Biol. 15: 637–642.
Chiyoda, S., Ishizaki, K., Kataoka, H., Yamato, K.T., and Kohchi, T.(2008). Direct transformation of the liverwort Marchantia poly-morpha L. by particle bombardment using immature thalli de-veloping from spores. Plant Cell Rep. 27: 1467–1473.
Christensen, S., LaVerne, E., Boyd, G., and Silverthorne, J. (2002).Ginkgo biloba retains functions of both type I and type II floweringplant phytochrome. Plant Cell Physiol. 43: 768–777.
Christie, J.M., Blackwood, L., Petersen, J., and Sullivan, S. (2015).Plant flavoprotein photoreceptors. Plant Cell Physiol. 56: 401–413.
De Greef, J., Butler, W.L., and Roth, T.F. (1971). Control of senes-cence in marchantia by phytochrome. Plant Physiol. 48: 407–412.
Duanmu, D., et al. (2014). Marine algae and land plants share con-served phytochrome signaling systems. Proc. Natl. Acad. Sci. USA111: 15827–15832.
Edgar, R.C. (2004). MUSCLE: multiple sequence alignment with highaccuracy and high throughput. Nucleic Acids Res. 32: 1792–1797.
Eklund, D.M., et al. (2015). Auxin produced by the indole-3-pyruvicacid pathway regulates development and gemmae dormancy in theliverwort Marchantia polymorpha. Plant Cell 27: 1650–1669.
Flores-Sandoval, E., Dierschke, T., Fisher, T.J., and Bowman, J.L.(2016). Efficient and inducible use of artificial microRNAs inMarchantia polymorpha. Plant Cell Physiol. 57: 281–290.
Fredericq, H., and De Greef, J. (1966). Red (R), far-red (FR) photo-reversible control of growth and chlorophyll content in light-grownthalli of Marchantia polymorpha L. Naturwissenschaften 53: 337.
Gamborg, O.L., Miller, R.A., and Ojima, K. (1968). Nutrient require-ments of suspension cultures of soybean root cells. Exp. Cell Res.50: 151–158.
1418 The Plant Cell

Genoud, T., Schweizer, F., Tscheuschler, A., Debrieux, D., Casal,J.J., Schäfer, E., Hiltbrunner, A., and Fankhauser, C. (2008).FHY1 mediates nuclear import of the light-activated phytochrome Aphotoreceptor. PLoS Genet. 4: e1000143.
Guindon, S., Dufayard, J.F., Lefort, V., Anisimova, M., Hordijk, W.,and Gascuel, O. (2010). New algorithms and methods to estimatemaximum-likelihood phylogenies: assessing the performance ofPhyML 3.0. Syst. Biol. 59: 307–321.
Hartmann, E., and Jenkins, G.I. (1984). Photomorphogenesis of Mossesand Liverworts. In The Experimental Biology of Bryophytes, A.F. Dyerand and J.G. Duckett, eds (London: Academic Press), pp. 203–228.
Haupt, W. (1985). Effects of nutrients and light pretreatment onphytochrome-mediated fern-spore germination. Planta 164: 63–68.
Hiltbrunner, A., Tscheuschler, A., Viczián, A., Kunkel, T., Kircher,S., and Schäfer, E. (2006). FHY1 and FHL act together to mediatenuclear accumulation of the phytochrome A photoreceptor. PlantCell Physiol. 47: 1023–1034.
Huang, X., Ouyang, X., and Deng, X.W. (2014). Beyond repression ofphotomorphogenesis: role switching of COP/DET/FUS in light sig-naling. Curr. Opin. Plant Biol. 21: 96–103.
Huq, E., Al-Sady, B., and Quail, P.H. (2003). Nuclear translocation ofthe photoreceptor phytochrome B is necessary for its biologicalfunction in seedling photomorphogenesis. Plant J. 35: 660–664.
Ishizaki, K., Chiyoda, S., Yamato, K.T., and Kohchi, T. (2008).Agrobacterium-mediated transformation of the haploid liverwortMarchantia polymorpha L., an emerging model for plant biology.Plant Cell Physiol. 49: 1084–1091.
Ishizaki, K., Nishihama, R., Yamato, K.T., and Kohchi, T. (2016).Molecular genetic tools and techniques for Marchantia polymorpharesearch. Plant Cell Physiol. 57: 262–270.
Ishizaki, K., Johzuka-Hisatomi, Y., Ishida, S., Iida, S., and Kohchi,T. (2013). Homologous recombination-mediated gene targeting inthe liverwort Marchantia polymorpha L. Sci. Rep. 3: 1532.
Ishizaki, K., Nishihama, R., Ueda, M., Inoue, K., Ishida, S.,Nishimura, Y., Shikanai, T., and Kohchi, T. (2015). Developmentof gateway binary vector series with four different selection markersfor the liverwort Marchantia polymorpha. PLoS One 10: e0138876.
Ito, S., Song, Y.H., and Imaizumi, T. (2012). LOV domain-containingF-box proteins: light-dependent protein degradation modules inArabidopsis. Mol. Plant 5: 573–582.
Jaedicke, K., Lichtenthäler, A.L., Meyberg, R., Zeidler, M., and Hughes,J. (2012). A phytochrome-phototropin light signaling complex at theplasma membrane. Proc. Natl. Acad. Sci. USA 109: 12231–12236.
Jenkins, G.I. (2014). The UV-B photoreceptor UVR8: from structure tophysiology. Plant Cell 26: 21–37.
Kami, C., Lorrain, S., Hornitschek, P., and Fankhauser, C. (2010). Light-regulated plant growth and development. Curr. Top. Dev. Biol. 91: 29–66.
Karas, B., Amyot, L., Johansen, C., Sato, S., Tabata, S.,Kawaguchi, M., and Szczyglowski, K. (2009). Conservation oflotus and Arabidopsis basic helix-loop-helix proteins reveals newplayers in root hair development. Plant Physiol. 151: 1175–1185.
Kato, H., Ishizaki, K., Kouno, M., Shirakawa, M., Bowman, J.L.,Nishihama, R., and Kohchi, T. (2015). Auxin-mediated transcrip-tional system with a minimal set of components is critical for mor-phogenesis through the life cycle in Marchantia polymorpha. PLoSGenet. 11: e1005084.
Kebrom, T.H., Burson, B.L., and Finlayson, S.A. (2006). Phyto-chrome B represses Teosinte Branched1 expression and inducessorghum axillary bud outgrowth in response to light signals. PlantPhysiol. 140: 1109–1117.
Kebrom, T.H., Brutnell, T.P., and Finlayson, S.A. (2010). Suppres-sion of sorghum axillary bud outgrowth by shade, phyB and de-foliation signalling pathways. Plant Cell Environ. 33: 48–58.
Khanna, R., Huq, E., Kikis, E.A., Al-Sady, B., Lanzatella, C., andQuail, P.H. (2004). A novel molecular recognition motif necessaryfor targeting photoactivated phytochrome signaling to specific ba-sic helix-loop-helix transcription factors. Plant Cell 16: 3033–3044.
Kircher, S., Kozma-Bognar, L., Kim, L., Adam, E., Harter, K.,Schafer, E., and Nagy, F. (1999). Light quality-dependent nuclearimport of the plant photoreceptors phytochrome A and B. Plant Cell11: 1445–1456.
Klose, C., Viczián, A., Kircher, S., Schäfer, E., and Nagy, F. (2015).Molecular mechanisms for mediating light-dependent nucleo/cy-toplasmic partitioning of phytochrome photoreceptors. New Phytol.206: 965–971.
Kubota, A., Ishizaki, K., Hosaka, M., and Kohchi, T. (2013). EfficientAgrobacterium-mediated transformation of the liverwort Marchantiapolymorpha using regenerating thalli. Biosci. Biotechnol. Biochem.77: 167–172.
Kubota, A., Kita, S., Ishizaki, K., Nishihama, R., Yamato, K.T., andKohchi, T. (2014). Co-option of a photoperiodic growth-phasetransition system during land plant evolution. Nat. Commun. 5:3668.
Leivar, P., and Monte, E. (2014). PIFs: systems integrators in plantdevelopment. Plant Cell 26: 56–78.
Li, F.W., Melkonian, M., Rothfels, C.J., Villarreal, J.C., Stevenson,D.W., Graham, S.W., Wong, G.K., Pryer, K.M., and Mathews, S.(2015). Phytochrome diversity in green plants and the origin of ca-nonical plant phytochromes. Nat. Commun. 6: 7852.
Lorrain, S., Allen, T., Duek, P.D., Whitelam, G.C., and Fankhauser,C. (2008). Phytochrome-mediated inhibition of shade avoidanceinvolves degradation of growth-promoting bHLH transcription fac-tors. Plant J. 53: 312–323.
Lu, X.D., Zhou, C.M., Xu, P.B., Luo, Q., Lian, H.L., and Yang, H.Q.(2015). Red-light-dependent interaction of phyB with SPA1 pro-motes COP1-SPA1 dissociation and photomorphogenic developmentin Arabidopsis. Mol. Plant 8: 467–478.
Mancinelli, A.L. (1994). The physiology of phytochrome action. InPhotomorphogenesis in Plants, R.E. Kendrick and and G.H.M.Kronenberg, eds (Dordrecht, The Netherlands: Kluwer AcademicPublishers), pp. 211–269.
Mathews, S. (2006). Phytochrome-mediated development in landplants: red light sensing evolves to meet the challenges of changinglight environments. Mol. Ecol. 15: 3483–3503.
Mittmann, F., Brücker, G., Zeidler, M., Repp, A., Abts, T.,Hartmann, E., and Hughes, J. (2004). Targeted knockout inPhyscomitrella reveals direct actions of phytochrome in the cyto-plasm. Proc. Natl. Acad. Sci. USA 101: 13939–13944.
Mølmann, J.A., Junttila, O., Johnsen, O., and Olsen, J.E. (2006).Effects of red, far-red and blue light in maintaining growth in lat-itudinal populations of Norway spruce (Picea abies). Plant Cell En-viron. 29: 166–172.
Morand, L.Z., Kidd, D.G., and Lagarias, J.C. (1993). Phytochromelevels in the green alga Mesotaenium caldariorum are light regu-lated. Plant Physiol. 101: 97–104.
Mukougawa, K., Kanamoto, H., Kobayashi, T., Yokota, A., andKohchi, T. (2006). Metabolic engineering to produce phytochromeswith phytochromobilin, phycocyanobilin, or phycoerythrobilinchromophore in Escherichia coli. FEBS Lett. 580: 1333–1338.
Nagatani, A., Reed, J.W., and Chory, J. (1993). Isolation and initialcharacterization of Arabidopsis mutants that are deficient in phy-tochrome A. Plant Physiol. 102: 269–277.
Ni, M., Tepperman, J.M., and Quail, P.H. (1998). PIF3, a phyto-chrome-interacting factor necessary for normal photoinduced sig-nal transduction, is a novel basic helix-loop-helix protein. Cell 95:657–667.
Phytochrome Signaling in Liverwort 1419

Ninnemann, H., and Halbsguth, W. (1965). Rolle des phytochromsbeim etiolement von Marchantia polymorpha. Naturwissenschaften52: 110–111.
Nishihama, R., Ishida, S., Urawa, H., Kamei, Y., and Kohchi, T.(2016). Conditional gene expression/deletion systems for Marchantiapolymorpha using its own heat-shock promoter and Cre/loxP-mediated site-specific recombination. Plant Cell Physiol. 57: 271–280.
Nishihama, R., Ishizaki, K., Hosaka, M., Matsuda, Y., Kubota, A.,and Kohchi, T. (2015). Phytochrome-mediated regulation of celldivision and growth during regeneration and sporeling developmentin the liverwort Marchantia polymorpha. J. Plant Res. 128: 407–421.
Oh, E., Kim, J., Park, E., Kim, J.I., Kang, C., and Choi, G. (2004).PIL5, a phytochrome-interacting basic helix-loop-helix protein, isa key negative regulator of seed germination in Arabidopsis thaliana.Plant Cell 16: 3045–3058.
Oka, Y., Ono, Y., Toledo-Ortiz, G., Kokaji, K., Matsui, M.,Mochizuki, N., and Nagatani, A. (2012). Arabidopsis phyto-chrome a is modularly structured to integrate the multiple featuresthat are required for a highly sensitized phytochrome. Plant Cell 24:2949–2962.
Okada, S., et al. (2000). Construction of male and female PAC ge-nomic libraries suitable for identification of Y-chromosome-specificclones from the liverwort, Marchantia polymorpha. Plant J. 24: 421–428.
Otto, K.-R., and Halbsguth, W. (1976). Die förderung der bildung vonprimärrhizoiden an brutkörpern von Marchantia polymorpha L.durch Licht und IES. Z. Pflanzenphysiol. 80: 197–205.
Park, E., Kim, J., Lee, Y., Shin, J., Oh, E., Chung, W.I., Liu, J.R., andChoi, G. (2004). Degradation of phytochrome interacting factor 3 inphytochrome-mediated light signaling. Plant Cell Physiol. 45: 968–975.
Parks, B.M., and Quail, P.H. (1993). hy8, a new class of arabidopsislong hypocotyl mutants deficient in functional phytochrome A. PlantCell 5: 39–48.
Pfeiffer, A., Nagel, M.K., Popp, C., Wüst, F., Bindics, J., Viczián, A.,Hiltbrunner, A., Nagy, F., Kunkel, T., and Schäfer, E. (2012). In-teraction with plant transcription factors can mediate nuclear importof phytochrome B. Proc. Natl. Acad. Sci. USA 109: 5892–5897.
Pires, N., and Dolan, L. (2010). Origin and diversification of basic-helix-loop-helix proteins in plants. Mol. Biol. Evol. 27: 862–874.
Possart, A., and Hiltbrunner, A. (2013). An evolutionarily conservedsignaling mechanism mediates far-red light responses in landplants. Plant Cell 25: 102–114.
Qiu, Y.L., et al. (2006). The deepest divergences in land plants in-ferred from phylogenomic evidence. Proc. Natl. Acad. Sci. USA 103:15511–15516.
Quail, P.H. (2010). Phytochromes. Curr. Biol. 20: R504–R507.Rattanapisit, K., Cho, M.H., and Bhoo, S.H. (2016). Lysine 206 in
Arabidopsis phytochrome A is the major site for ubiquitin-dependent protein degradation. J. Biochem. 159: 161–169.
Rausenberger, J., Tscheuschler, A., Nordmeier, W., Wüst, F.,Timmer, J., Schäfer, E., Fleck, C., and Hiltbrunner, A. (2011).Photoconversion and nuclear trafficking cycles determine phyto-chrome A’s response profile to far-red light. Cell 146: 813–825.
Reddy, R.K., and Sharma, R. (1998). Spatial distribution and tem-poral regulation of phytochromes A and B levels in maize seedlings.Plant Physiol. Biochem. 36: 737–745.
Rösler, J., Jaedicke, K., and Zeidler, M. (2010). Cytoplasmic phy-tochrome action. Plant Cell Physiol. 51: 1248–1254.
Saijo, Y., Zhu, D., Li, J., Rubio, V., Zhou, Z., Shen, Y., Hoecker, U.,Wang, H., and Deng, X.W. (2008). Arabidopsis COP1/SPA1 com-plex and FHY1/FHY3 associate with distinct phosphorylated forms
of phytochrome A in balancing light signaling. Mol. Cell 31: 607–613.
Saint-Marcoux, D., Proust, H., Dolan, L., and Langdale, J.A. (2015).Identification of reference genes for real-time quantitative PCR ex-periments in the liverwort Marchantia polymorpha. PLoS One 10:e0118678.
Sawers, R.J., Linley, P.J., Farmer, P.R., Hanley, N.P., Costich, D.E.,Terry, M.J., and Brutnell, T.P. (2002). Elongated mesocotyl1,a phytochrome-deficient mutant of maize. Plant Physiol. 130: 155–163.
Sharrock, R.A., and Clack, T. (2002). Patterns of expression andnormalized levels of the five Arabidopsis phytochromes. PlantPhysiol. 130: 442–456.
Sheerin, D.J., Menon, C., zur Oven-Krockhaus, S., Enderle, B.,Zhu, L., Johnen, P., Schleifenbaum, F., Stierhof, Y.D., Huq, E.,and Hiltbrunner, A. (2015). Light-activated phytochrome A and Binteract with members of the SPA family to promote photomor-phogenesis in Arabidopsis by reorganizing the COP1/SPA complex.Plant Cell 27: 189–201.
Shen, H., Zhu, L., Castillon, A., Majee, M., Downie, B., and Huq, E.(2008). Light-induced phosphorylation and degradation of thenegative regulator PHYTOCHROME-INTERACTING FACTOR1 fromArabidopsis depend upon its direct physical interactions with pho-toactivated phytochromes. Plant Cell 20: 1586–1602.
Shen, Y., Khanna, R., Carle, C.M., and Quail, P.H. (2007). Phyto-chrome induces rapid PIF5 phosphorylation and degradation inresponse to red-light activation. Plant Physiol. 145: 1043–1051.
Shinomura, T., Uchida, K., and Furuya, M. (2000). Elementary pro-cesses of photoperception by phytochrome A for high-irradianceresponse of hypocotyl elongation in Arabidopsis. Plant Physiol. 122:147–156.
Shinomura, T., Nagatani, A., Hanzawa, H., Kubota, M., Watanabe,M., and Furuya, M. (1996). Action spectra for phytochrome A- andB-specific photoinduction of seed germination in Arabidopsisthaliana. Proc. Natl. Acad. Sci. USA 93: 8129–8133.
Skaar, J.R., Pagan, J.K., and Pagano, M. (2013). Mechanisms andfunction of substrate recruitment by F-box proteins. Nat. Rev. Mol.Cell Biol. 14: 369–381.
Sokol, R.C., and Stross, R.G. (1992). Phytochrome-mediated ger-mination of very sensitive oospores. Plant Physiol. 100: 1132–1136.
Su, Y.S., and Lagarias, J.C. (2007). Light-independent phytochromesignaling mediated by dominant GAF domain tyrosine mutants of Ara-bidopsis phytochromes in transgenic plants. Plant Cell 19: 2124–2139.
Sugano, S.S., Shirakawa, M., Takagi, J., Matsuda, Y., Shimada, T.,Hara-Nishimura, I., and Kohchi, T. (2014). CRISPR/Cas9-mediatedtargeted mutagenesis in the liverwort Marchantia polymorphaL. Plant Cell Physiol. 55: 475–481.
Suzuki, T., Takio, S., Yamamoto, I., and Satoh, T. (2001). Charac-terization of cDNA of the liverwort phytochrome gene, and phyto-chrome involvement in the light-dependent and light-independentprotochlorophyllide oxidoreductase gene expression in Marchantiapaleacea var. diptera. Plant Cell Physiol. 42: 576–582.
Takano, M., Inagaki, N., Xie, X., Kiyota, S., Baba-Kasai, A.,Tanabata, T., and Shinomura, T. (2009). Phytochromes are thesole photoreceptors for perceiving red/far-red light in rice. Proc.Natl. Acad. Sci. USA 106: 14705–14710.
Takano, M., Inagaki, N., Xie, X., Yuzurihara, N., Hihara, F.,Ishizuka, T., Yano, M., Nishimura, M., Miyao, A., Hirochika, H.,and Shinomura, T. (2005). Distinct and cooperative functions ofphytochromes A, B, and C in the control of deetiolation and flow-ering in rice. Plant Cell 17: 3311–3325.
Todaka, D., et al. (2012). Rice phytochrome-interacting factor-likeprotein OsPIL1 functions as a key regulator of internode elongation
1420 The Plant Cell

and induces a morphological response to drought stress. Proc.Natl. Acad. Sci. USA 109: 15947–15952.
Toledo-Ortiz, G., Kiryu, Y., Kobayashi, J., Oka, Y., Kim, Y., Nam,H.G., Mochizuki, N., and Nagatani, A. (2010). Subcellular sites ofthe signal transduction and degradation of phytochrome A. PlantCell Physiol. 51: 1648–1660.
Tsuboi, H., Nakamura, S., Schäfer, E., and Wada, M. (2012). Redlight-induced phytochrome relocation into the nucleus in Adiantumcapillus-veneris. Mol. Plant 5: 611–618.
Vandesompele, J., De Preter, K., Pattyn, F., Poppe, B., Van Roy, N.,De Paepe, A., and Speleman, F. (2002). Accurate normalization ofreal-time quantitative RT-PCR data by geometric averaging ofmultiple internal control genes. Genome Biol. 3: H0034.
Weisenseel, M.H., and Ruppert, H.K. (1977). Phytochrome and cal-cium ions are involved in light-induced membrane depolarization inNitella. Planta 137: 225–229.
Whitelam, G.C., Johnson, E., Peng, J., Carol, P., Anderson, M.L., Cowl,J.S., and Harberd, N.P. (1993). Phytochrome A null mutants of Arabi-dopsis display a wild-type phenotype in white light. Plant Cell 5: 757–768.
Xie, X., Kagawa, T., and Takano, M. (2014). The phytochromeB/phytochrome C heterodimer is necessary for phytochromeC-mediated responses in rice seedlings. PLoS One 9: e97264.
Yamaguchi, R., Nakamura, M., Mochizuki, N., Kay, S.A., andNagatani, A. (1999). Light-dependent translocation of a phyto-chrome B-GFP fusion protein to the nucleus in transgenic Arabi-dopsis. J. Cell Biol. 145: 437–445.
Phytochrome Signaling in Liverwort 1421

DOI 10.1105/tpc.15.01063; originally published online June 1, 2016; 2016;28;1406-1421Plant Cell
Yasuomi Tada, Kimitsune Ishizaki and Takayuki KohchiKeisuke Inoue, Ryuichi Nishihama, Hideo Kataoka, Masashi Hosaka, Ryo Manabe, Mika Nomoto,
Marchantia polymorphaLiverwort Phytochrome Signaling Is Mediated by PHYTOCHROME INTERACTING FACTOR in the
This information is current as of June 5, 2018
Supplemental Data /content/suppl/2016/06/01/tpc.15.01063.DC1.html
References /content/28/6/1406.full.html#ref-list-1
This article cites 91 articles, 33 of which can be accessed free at:
Permissions https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw1532298X
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists


















