荒野心情 荒野心情 墾丁南八解後記 - sow.org.tw · 墾丁南八解後記 選在 3 1 2 5年最後一個假日到南台灣墾丁國 家公享受溫暖的冬陽,這是台南分會第
附錄次
墾丁國家公園墾丁地區大冠鷲(Spilornis cheela hoya)繁殖及棲地利用之調查研究
A Study of the breeding biology and habitat
using of Formosan Crested Serpenteagle
內政部營建署墾丁國家公園管理處委託研究報告
中華民國九十四年十二月
I
PG9403-0384 094-301020100G1-001
墾丁國家公園墾丁地區大冠鷲(Spilornis cheela hoya)繁殖及棲地利用之調查研究
A Study of the breeding biology and habitat
using of Formosan Crested Serpenteagle
受委託者:周大慶
研究助理:潘明雄、胡景程、楊靜櫻、陳宏信、
朱慧敏、簡伶育、張育誠、楊信得、
施月英、陶天麟
內政部營建署墾丁國家公園管理處委託研究報告
中華民國九十四年十二月
目次
目次
目次 ................................................................................................................... I
表次 ................................................................................................................ III
圖次 .................................................................................................................. V
附錄次 ........................................................................................................... VII
摘 要 ...........................................................................................................IX
第一章 前 言 ................................................................................................. 1
第二章 目 的 ................................................................................................. 5
第三章 方 法 ................................................................................................. 7
第一節 找尋巢位及繁殖行為觀察.............................................................. 7
第二節 育雛食物種類及食繭食餘收集...................................................... 7
第三節 棲地偏好 .......................................................................................... 7
一、巢位特徵測量 ................................................................................... 8
二、微棲地特徵測量 ............................................................................... 8
三、巨棲地特徵測量 ............................................................................... 9
第四節 繫放 .................................................................................................. 9
第五節 無線電定位 .................................................................................... 10
第四章 結 果 ............................................................................................... 11
第一節 巢位尋找 ........................................................................................ 11
第二節 育雛食物種類及頻度.................................................................... 14
第三節 棲地偏好 ........................................................................................ 15
一、微棲地特徵 ..................................................................................... 15
I
二、巨棲地特徵 ..................................................................................... 18
第四節 無線電定位 .................................................................................... 26
第五章 討 論 ............................................................................................... 31
第六章 建 議 ............................................................................................... 33
附 錄 ......................................................................................................... 35
附錄一、服務企劃書評審會議紀錄 ......................................................... 35
附錄二、期中簡報審查會議紀錄 ............................................................. 43
附錄三、期末簡報審查會議記錄 ............................................................. 51
參考書目 ......................................................................................................... 59
表次
表次
表 4-1 2005 年墾丁國家公園墾丁及社頂地區猛禽巢位調查摘要 ......... 12
表 4-2 2005 年大冠鷲S19 和S25 巢位育雛生物種類和頻度................... 14
表 4-3 2005 年大冠鷲利用巢樹和逢機取樣樹木特徵比較 ..................... 16
表 4-4 2005 年大冠鷲巢尺寸 ..................................................................... 17
表 4-5 大冠鷲巢位特徵 .............................................................................. 17
表 4-6 2005 年繁殖大冠鷲巢樹林下莖幹數量及草本層和灌叢覆蓋度 . 18
表 4-7 2005 年大冠鷲巢樹所在地形 ......................................................... 18
表 4-8 1995 年至 2005 年間所發現之大冠鷲巢樹所在地形百分比 ....... 19
表 4-9 1995-2005 巢樹所在坡度數量統計 ................................................ 19
表 4-10 2005 年大冠鷲利用巢樹和重要地景距離 ................................. 19
表 4-11 研究區域各種植被及土地類型面積及百分比 ............................ 20
表 4-12 2005 年五個繁殖巢區植被土地類型面積及比例 ....................... 25
表 4-13 2005 年捕捉成鳥之外型量値 ....................................................... 26
表 4-14 2005 年成鳥繫放項目一覽表 ....................................................... 27
表 4-15 2005 年 2 月至 11 月無線電定位個體天數一覽表 ..................... 27
III
圖次
圖次
圖 4-1 2005 年墾丁地區猛禽巢位及配對行為分布地點 ......................... 13
圖 4-2 1995 年至 2005 年墾丁地區大冠鷲利用巢樹種類次數 ............... 15
圖 4-3 研究區位主要土地植被型態和分布 .............................................. 21
圖 4-4 063 個體定位範圍 ........................................................................... 28
圖 4-5 083 個體定位範圍 ........................................................................... 29
圖 4-6 120 個體間定位範圍 ....................................................................... 29
圖 4-7 138 個體定位範圍 ........................................................................... 30
圖 4-8 258 個體定位範圍 ........................................................................... 30
V
附錄次
附錄次
附錄一、服務企劃書評審會議紀錄............................................................. 35
附錄二、期中簡報審查會議紀錄................................................................. 43
附錄三、期末簡報審查會議記錄................................................................. 51
VII
摘要
摘 要
關鍵詞:社頂、大冠鷲、無線電定位
一、研究緣起
猛禽位居食物鏈高層,可做為陸域生態環境的指標物種,用來監測生態系的狀
態。屬於國內二級保育類野生動物的大冠鷲,有關其於墾丁國家公園甚或台灣地區之
生態研究與調查如族群結構、棲地需求、自然史等的基礎資料甚少,有必要加以了解
其對生態系的角色與貢獻,藉由其基礎繁殖生物學上之研究,可以彌補這方面工作之
不足。
二、研究方法及過程
研究方法包括徒步地毯式搜尋研究區域內 1241公頃森林內的大冠鷲巢位和觀察
育雛行為,離巢前的幼鳥則進行繫放追蹤其存活率、擴散等資料。幼鳥離巢後測量巢
位半徑 17.84 公尺內之微棲地特徵,和半徑 250 公尺範圍內土地類型比例及植被類
型。此外亦以誘餌誘捕成鳥,進行無線電定位及追蹤,以了解其棲地利用方式。
三、重要發現
本年度至 11 月中調查期間共發現 5 個大冠鷲巢位,和 11 個鳳頭蒼鷹巢位。此
外,有 5 個 2004 年舊巢區雖未發現築巢,但有觀察到空中對爪、林下交尾、求偶叫
聲及乞食等繁殖行為。5 個大冠鷲巢位中有 3 個巢繁殖失敗,分別受到盜伐者干擾、
獼猴騷擾、颱風吹落巢位等因素影響。本年度觀察大冠鷲育雛行為發現遞食食物種類
之頻度以蜈蚣和蛇類較高;上午 10 點至下午 1 點時段的育雛給食頻率較高。連續森
林內部山溝地形是大冠鷲偏好的營巢地點,山溝中高大的相思樹和茄苳是本區大冠鷲
最常選擇的巢樹種類,巢樹或巢位如未經天候或自然因素破壞或遭受干擾,具有重複
使用的現象。巢位半徑 250 公尺內森林類型有三種分別為次生林、和高位珊瑚礁森林
和海岸林,占全部面積 96.1%。無線電定位的資料顯示,前一年出生的大冠鷲在次年
的梅雨季節開始前往外擴散,擴散的壓力可能來自食物不足和繁殖成鳥的驅趕。現有
的夜棲地定位資料顯示每隻個體有幾處使用頻率較高的地點,但經常在強風天候轉變
IX
夜間棲息地點,而多數個體白天實際活動的範圍相當小和移動的次數則相當低。
四、主要建議事項
由於大冠鷲需要相當廣闊的樹林得以將巢安置於遠離林緣的區域,但是研究區
森林經常遭到盜伐和人為騷擾,因此建議在國家公園內現存的樹林應該小心被保護,
不要再任其切割破碎,特別是位於溪谷和珊瑚礁週遭的遮蔽東北季風的地形。大冠鷲
所選擇繁殖的地點和樹種資料顯示,其偏好環境主要在溪溝和山谷地形的高大植株
上,因此對於上述生物和環境的保護及管理有其重要性。
建議一 森林環境的保護和管理:立即可行建議
主辦機關:內政部營建署墾丁國家公園管理處
協辦機關:內政部警政署墾丁國家公園警察隊
期程與做法:有急迫性,應該列為例行工作。墾管處商請警察隊便衣每天不定期
巡邏各處森林至少兩次,並請兩單位定期檢討工作狀況。
建議二 獼猴數量管理的規劃:中長期建議
主辦機關:內政部營建署墾丁國家公園管理處
協辦機關:彌猴研究相關學者或院校
期程與做法:建議於兩年內實施為期二至三年之短期委託專案研究獼猴族群和活
動範圍變化以及和遊客互動狀況,長期提出前瞻性的經營管理按部
就班做法。
根據過往研究資料顯示,本研究區大冠鷲的繁殖成功率相當高,超過 90%,但
今年所發現的五個巢位中有一個大冠鷲和兩個鳳頭蒼鷹的巢被獼猴騷擾而棄巢,這個
情形過去 3 年沒有發現過,可能是保育獼猴後,其族群數目逐漸增加,和遊客的接觸
也隨之提高,建議未來可將獼猴數量的管理納入經營重點。
建議三 牛隻和鹿隻管理的籌畫:立即可行建議
主辦機關:內政部營建署墾丁國家公園管理處
協辦機關:內政部警政署墾丁國家公園警察隊
期程與做法:短期(一年內)請懇管處檢討梅花鹿復育的階段性任務可否朝向展
示教育可行性,並提出加強非法放牧牛隻管理辦法;中期(兩年內)
摘要
委託專案研究梅花鹿和放牧對森林小苗更新的影響,並提出管理策
略。
非法放牧的牛隻和復育的梅花鹿可能對森林地面水源和植被更新產生長遠影
響,進而影響食物鏈結構。
XI
摘要
ABSTRACT
keywords:Sheding, Spilornis cheela hoya, radio-telemetry
Five Crested Serpent Eagle nests and 11 Crested Goshawk (Accipiter trivirgatus) nests were located by intensive foot survey during the breeding season of 2005 throughout the entire study area. Besides, five breeding pairs showed aerial claws hooking, courtship calling, and food begging behavior were also located near the breeding area of last year, but no breeding nests were found in these areas. Two Crested Serpent Eagle nests of this year were abandoned by illegal logging and Formosan macaques (Macaca cyclopis) harassment. Based on observations of 39 prey items brought to two intensively monitored nest, centipedes (>56 %) and snakes (>28 %) were the most frequently observed prey items, and food usually delivered to nests around noon. The Crested Serpent eagles need relatively larger forest patches so they can place their nests away from the edges. Specifically, the forests that support taller trees in the stream valleys of Kenting should be protected and not fragmented any further. The mean proportion of forest cover within 250 m of all nests is 96.1%, and only the forest types of secondary forest, elevated coral reef forest, and costal forest appeared. Serpent eagles seemed to have a high tendency of using Acacia confusa and Bischofia javanica to place their nests. If nests or nest trees are not disturbed or ruined by monsoon or typhoon, reusing of the placement is probable. Radio-telemetry shows juvenile eagles of last year start to disperse before the rainy season of this year probably owing to the stress of insufficient food or expelling by local breeding pair. According to the information of radio telemetry, the banded eagle usually use several perching places at night steadily but switch to seldom visited and less windy spots to over night when a typhoon or strong winter monsoon hits.
XIII
第一章 前言
第一章 前 言
墾丁國家公園區內地形複雜多變,加上植被類型差異大,孕育了各種適合猛禽
棲息的環境,雖然部份區域經過人為開墾,造成棲地破碎影響,仍有居留及過境猛禽
族群相當數量的分佈。根據研究資料,目前墾丁地區的猛禽有 26 種日行性猛禽紀錄,
包括 6 種留鳥猛禽及 20 種遷徙性猛禽,其中留鳥猛禽以大冠鷲較常見(蔡 1996;劉
1991)。
猛禽位居食物鏈高層,可做為陸域生態環境的指標物種,用來監測生態系的狀
態。屬於國內二級保育類野生動物的大冠鷲,有關其於墾丁國家公園甚或台灣地區之
生態研究與調查如族群結構、棲地需求、自然史等的基礎資料甚少,有必要加以了解
其對生態系的角色與貢獻,藉由其基礎繁殖生物學上之研究,可以彌補這方面工作之
不足。
台灣地區的大冠鷲(Spilornis cheela hoya Swinhoe)隸屬於鷹形目(Falconiformes)
鷲鷹科(Accipitridae)蛇鵰屬(Spilornis),為 1866 年由史溫侯(Swinhoe)所命名(陳
&于 1984)。蛇鵰屬是猛禽家族中相當複雜的 1 個屬,目前該屬多數種類主要分布北
緯 35 度至南緯 9 度之間的印度、中國長江以南到馬來半島、中南半島、菲律賓群島、
西印尼群島、婆羅洲之間的區域,分布相當廣泛而且普遍(許 1995)。雖然眾多的地
方變異造成本屬各種及其各亞種間分類關係混淆和一再修正(Ferguson-lees 2001;
Howard and Moore 1991; del Hoyo 1994; Stepanyan 2001),但目前台灣地區的大冠
鷲被認為地方亞種。
大冠鷲雌雄外型近似,但個體體型及羽色變異頗大,成鳥體長約 70 公分 (陳&
于 1984)。成鳥體色多偏暗,但肩部、腹及脇部點綴白色斑點,尾部及翼緣有相當醒
目的白色帶狀,冠羽短而密集,使得頭頂部位看起來較扁平,頭看起來較大,冠羽警
戒時常豎起,眼先無毛,翅屬寬長型,尾長中等,腳無毛,白色型亞成鳥逐年換羽之
後外型類似成鳥,但仍和成鳥略有差異,通常羽緣較淡,身上多條紋,隨著年齡增長,
尾及翼下白色橫帶轉寬大明顯,體背及胸部羽毛顏色轉深,成熟之後則無明顯差異
(Ferguson-lees and Christie 2001;Hoyo et al. 1994;Severinghaus and Blackshaw 1976;
王等 1991;顏等 1996)。
1
森林範圍和海拔高度大致框架大冠鷲活動範圍,其主要海拔分布由海平面至
1500 公尺(翟 1977;顏 1999),2000 公尺亦相當平常(張 1985),有時可以到達
3000 公尺(周 2001;鄭等 1983),在尼泊爾曾經於 10 月份記錄到高度在 3,350 公尺
的高度出沒(Ferguson-lees and Christie 2001);其水平分布範圍與中低海拔森林範圍
約略吻合,出沒地點之棲地環境相當多樣化,不論乾燥或潮濕、封閉或開放、低地或
山區的原生雨林、次生林到人造林(顏 1996; 張 1985)。
Ueta 和 Minton(1996)在日本的石垣島和西表島曾對另一亞種的大冠鷲
(Spilornis cheela perplexus Swann)進行調查,發現白天他們喜歡在林緣停棲,棲息點
構造以樹木為主,棲息地環境多為潮濕草生地和樹林。於墾丁國家公園的研究顯示,
森林面積較小的環境,大冠鷲可能出現頻率較低,這個研究可能暗示森林區塊的大小
對大冠鷲的分布有相當程度的影響(劉 1991)。
大冠鷲狩獵方式通常安靜地躲在樹上尋找樹叢或地上活動的獵物,發現獵物後
迅速由樹上落到地面以爪制服,然後攜到隱密的樹上進食,不曾發現類似黑鳶等猛禽
在空中進食的行為。其食物種類主要以爬蟲類如青蛙、蛇、蜥蜴為主,有時亦捕時小
型嚙齒動物、生病或受傷的鳥,或節肢動物等(Morioka 1995;Naoroji and Monga
1983;黃 1997;蕭與黃 1996),少數觀察指出大冠鷲亦曾捕食螃蟹及鰻魚(Chou
2003; Ferguson-lees and Christie 2001)。由於食性可能有地方上的差異,本研究為瞭
解墾丁地區大冠鷲食物組成是否有特殊組成或偏好,將針對繁殖期間育雛食物組成,
和覓食地點食繭和食餘進行調查分析。
林等 (1995)曾對台北縣觀音山地區針對 1 巢於繁殖期間進行食性記錄,發現親
鳥供給食物的高峰為 10:00-13:00,多數育雛記錄發生在晴天,86﹪的食物供應記錄
由父鳥所為。黃(1998、2004)也曾對陽明山地區大冠鷲巢進行繁殖季節期間之育雛食
物種類進行觀察。
其獵食環境亦相當多樣化,由人為活動的村莊附近,到茶園、果園等各式農耕
地,以及原生及次生林地等(Ferguson-lees and Christie 2001),顯示這種猛禽對環境
2
第一章 前言
的適應性較其他種類來的佔優勢。
有關大冠鷲的繁殖研究大多侷限在繁殖季期間的巢邊育雛行為觀察(Morioka
1995;Naoroji and Monga 1983;林等 2000;黃 1998;黃 2004)。在印度 Naoroji 和 Monga
(1983)曾對南古加拉特(Gujarat)地區的森林中之大冠鷲另一亞種(Spilornis cheela
melanotis Jerdon)進行巢樹種類、巢區林相、育雛期間獵物種類和行為進行觀察。在
日本 Morioka(1995)彙整日本地區過往有關琉球亞種大冠鷲型態、行為和生殖的觀察。
在台灣,黃(1998)曾對陽明山地區大冠鷲築巢的環境、幼雛擴散等項目進行觀
察。林等(1995)和林等(2000)曾對觀音山地區大冠鷲育雛期間的食性和習性進行調
查。上述研究均在台灣北部地區進行。這些研究顯示大冠鷲僅育 1 雛,由小樹枝組成
的巢相當小,約 50-60 公分直徑,10-30 公分深,襯以新鮮枝葉,巢離地高約 6-20 公
尺,通常築於森林或開放林地分叉的樹枝上,靠近溝壑或溪谷。
大冠鷲的巢由雌雄共同修築,由雌鳥單獨孵蛋,雄鳥負責供給食物。孵蛋時間
約 35 天,國外多數文獻觀察之幼鳥由孵化至離巢時間約 60-65 天(黃 1998;蕭與黃
1996;蕭 2001),最長記錄達 70 天,但不肯定(黃 1998)。
大冠鷲繁殖季節因緯度及高度而有所不同,分布較北的個體產卵於乾季,在印
度北方的山區由 3 月至 7 月,在平原則由 2 至 5 月,印度南方則早在 12 月就開始。
緬甸地區則在 2 月期間;蛋的記錄在爪哇則由 2 月至 11 月均有發現(Ferguson-lees and
Christie 2001; Hoyo et al. 1994)。台灣地區的記錄顯示大冠鷲 3 月中旬開始抱卵。墾
丁地區由於緯度較低,繁殖季發生的時間可能比較早。求偶期間的飛行展示例如俯
衝、翻滾、展示爪、抖翅、或典型地羽毛球狀飛行(將頭、翅、尾縮起上下起伏)偶
爾可見,但較不規則也不積極。
大冠鷲的領域和社會行為資料非常少,由於平常通常為單獨或成對出現,不論
單隻或彼此經常在任何季節於高空盤旋活動,繁殖季有時候小群或家庭成員一起活
動。一般認為大冠鷲的遷移行為不明顯屬於定棲性,但至少亞成鳥屬於四處遊走型
態,國外有少數地區記錄顯示可能隨季節在不同海拔間局部遷徙的現象(Ferguson-lees
3
and Christie 2001)。國內在陽明山地區的觀察發現幼鳥在離巢後 2 個月仍在距巢位
1600 公尺的範圍內活動(黃 1998),其餘擴散資料不詳。
由於全世界近半數熱帶地區的猛禽面臨棲地喪失的問題(Bildstein et al. 1998)。
許多研究發現,棲地破碎化對棲息於連續棲地內的物種有負面衝擊,破碎化直接減少
連續棲地的面積,進而降低各物種的總族群量(Bender et al. 1998)。猛禽的巢位在連續
且合適的棲地內,相互之間呈現約略相等的距離,這種繁殖密度的空間規律化是被領
域行為限制及調整的緣故,但是當棲地遭遇開發而喪失或者棲地品質下降,猛禽的巢
位選擇可能產生變化,繁殖密度則會下降 (Newton 1979)。由於砍伐森林、開發道路、
利用野地、焚墾農業等各種人為干擾,造成棲地破碎或棲地需求不足的問題,進而影
響猛禽的繁殖成功率,已成為猛禽研究的重要課題(Thiollay 1988)。
有關國內猛禽微棲地和巨棲地利用方面的研究僅陳輝勝(1997)曾於墾丁國家公
園內以鳳頭蒼鷹為對象進行數年之研究,不同於國外研究結果,該研究結果顯示破碎
棲地中的個體反而有較高的營巢密度及繁殖成功率,該作者認為這個現象的主因可能
係台灣鳳頭蒼鷹的島嶼生物特性及棲地破碎化降低掠食者數量所致。
4
第一章 前言
第二章 目 的
根據調查資料,墾丁國家公園的猛禽發現數量以南仁山區北段、滿州、社頂
居多,猛禽集中的地點多是大面積的天然林。但在墾丁及滿州等人為開墾、放牧
等的土地利用環境,仍可發現部分大冠鷲族群在此環境下繁殖。由於棲地破碎對
猛禽繁殖成功率的影響為猛禽研究的重要課題之一,為調查墾丁國家公園內大冠
鷲巢位棲地的利用及分布型態,以了解棲地破碎對大冠鷲族群與分布的影響,進
而作為未來經營管理需求上的依據,本計畫預期目標如下:
一、了解研究區內大冠鷲食物需求。
二、大冠鷲繁殖期間育雛行為觀察
三、棲地分析與族群量(密度)監測之初步資料建立
四、巢位選擇與棲地偏好之初步資料建立
五、藉棲地偏好模式及分布數量,提出未來對大冠鷲保育及經營管理之建議。
5
6
第三章 方法
第三章 方 法
第一節 找尋巢位及繁殖行為觀察
每年 3 月至 5 月期間,有系統的在整個研究區內步行搜索每一棵大樹上的巢。
一但發現一個巢,位置就以 GPS 定位計算大冠鷲平均最近巢間距,並標示在 97 正
射化空照圖上,並記錄巢的使用狀況如孵蛋或育雛等。為了降低對巢的干擾,利用
長竿頂的小鏡子或爬到附近的樹上或珊瑚礁岩上,以 Nikon 8 x 40 雙筒望遠鏡或
Nikon 20 x 60 單筒望遠鏡觀察。並記錄親鳥是否在巢內或附近以及蛋或雛鳥的數
目,至於活動的痕跡如是否有新鮮的綠葉在巢內、巢下或附近是否有新鮮的排遺、
食物的殘渣,只有具有上述徵兆的巢才被視為一個佔用或進行繁殖的巢位。調查過
程中如果有發現鳳頭蒼鷹巢位,也一併記錄其位置及使用狀況。
第二節 育雛食物種類及食繭食餘收集
雛食物的種類藉由在巢位附近搭設偽裝帳棚,由窺孔中以高倍率單筒望遠鏡
觀察,記錄親鳥攜回之食物種類及數量,所有食物包括不能立即鑑定物種以錄影
或拍照方式記錄輔助鑑定,並由圖片估算獵物大小。每個巢位每週至少進行 2 整
天的觀察,觀察時間若天候許可,由清晨 5:00 開始至下午 6:00 結束。幼鳥離巢後,
每個偽裝帳仍繼續觀察至幼鳥離巢後 20 天。此外對於研究區中個體進食地點之食
繭及食餘亦進行收集輔助分析。個體間互動或個體行為,如交配、築巢、餵食、
驅趕、理羽和處理食物等,及發生的時間、食物種類亦加以記錄,並辨識個體身
分。
第三節 棲地偏好
棲地的特色由三個尺度來探討 1.巢位的特徵、2.微棲地特徵、3.巨棲地特徵,
第一項的目的在探討巢位偏好,第二、三項目的在發掘棲地偏好,各項的工作方
法如下:
7
一、巢位特徵測量
巢的測量項目包括下列幾項:直徑【(長+寬) / 2】、巢的底座厚度、巢的深
度、巢的高度(巢緣最高點至巢底座) 。巢樹的測量特徵記錄項目包括:種類、胸
徑和高度。巢的位置包括:巢至主幹的距離、支撐樹枝的數目、支撐樹枝的平均
直徑、巢上方覆蓋度。當幼鳥離巢後才測量大冠鷲實際利用的巢樹和逢機取樣的
大樹的差異。座標逢機取樣的大樹以所知實際巢樹的最小胸徑為標準,當逢機取
樣的巢樹胸徑大於已知巢數最小胸徑才被測量。
使用工具包括測量胸徑的胸徑尺或距離的 10 公尺鋼捲尺和 50 公尺皮尺。測
量樹冠層覆蓋度使用 Paul E Lemmon 廠牌 24 格半球面鏡密度計。
二、微棲地特徵測量
微棲地特徵包含測量巢區植被結構和地形,此處「巢區」的界定以巢樹為中
心,半徑 17.84 公尺(面積約 1000 平方公尺)的範圍。測量項目包含胸徑超過 1 公
分的樹木胸徑值及數量、樹下層灌叢及草本層覆蓋比例、樹冠層覆蓋度、平均樹
冠高和林型。地形特徵包括高度、坡度、坡向、地形、巢至最近永久水源的最近
距離、巢至最近可通行車輛或有舖面的道路距離、巢至最近有遊客或經常有人通
行小徑距離、巢至最近有人居住的房舍距離、巢至最近人工房舍或建物距離。
其中樹冠覆蓋度測量方法以巢樹為中心沿 4 個方位,利用 24 格半球面計於
距樹 9 公尺和 15 公尺處各讀取 1 次樹冠層覆蓋度,每個巢位的覆蓋度由此 8 個讀
數平均。由 4 個方位在距離巢樹 20 公尺的地方利用斜度計測量樹冠層高度,由此
4 個讀數平均得到樹冠高。
巢附近的植被分類為次生林或人造林、常綠闊葉林和硬葉樹林 4 大類。次生
林或人造林又細分為 (1) 再生瓊崖海棠 (Calophyllum inophyllum) 林型、(2) 木麻
黃(Casuarina equisetifolia )人造林型、 (3) 欖仁(Terminalia catapa )林型、 (4) 毛
柿(Diospyros discolor )林型、(5) 混合半落葉再生林型、(6) 其他。常綠闊葉林則
細分為海岸林型和榕楠林型( ficus-machilus forest type)。 硬葉樹林則細分為相思
樹( Acacia confusa )林型和珊瑚礁岩植被型。
8
第三章 方法
坡向利用軍用羅盤測量,支撐巢樹的斜坡坡向分別為北方 (337.5°- 22.4°)、
東北方(22.5°- 67.4°)、東方 (67.5°- 112.4°)、東南方(112.5°- 157.4°)、南方 (157.5°-
202.4°)西南方 (202.5°- 247.4°)、西方 (247.5°- 292.4°)和西北方 (292.5°- 337.4°)。
沿著巢樹的坡向再巢樹上下各 5 公尺和 10 公尺的位置以斜度計讀取 4 個坡度後
平均 。海拔高度以農林航測所經建 1 : 5,000 空照圖上之等高線高度及參考 GPS。
地形粗分為鵝鸞鼻半島東坡和西坡,每個位置再細分為懸崖、平原、山腳、
山坡和山脊。
逢機取樣的樹木以 Microsoft Excel 軟體亂數功能,在樣區所在的經緯度範圍
內隨意選取和實際巢樹等數量的座標,然後以 GPS 至野外導航至選定的座標地
點,選取座標點上或距離座標點最近一棵胸徑大於已知最小巢樹胸徑的樹木進行
測量。
三、巨棲地特徵測量
巨棲地特徵項目包括以巢為中心之半徑 250 公尺、500 公尺、1000 公尺範
圍內森林覆蓋面積,以及林緣長度和林緣長度與面積的比值。
巨棲地特徵的測量或計算主要在 ArcMap 軟體上操作,藉由在 97 正射化影像
空照圖上以 Shape file 檔描繪及標示森林範圍和界線,輸入各巢位座標和半徑,計
算各半徑範圍內森林面積和林緣長度等數值。
第四節 繫放
為便利個體辨識及建立長期族群量監測依據,對於研究範圍內幼鳥進行繫
放,繫放時機在雛鳥約 50 日齡左右爬上樹將幼鳥垂降至地面,測量嘴鋒、跗躓、
後爪、翼展、體重各部份形值並拍攝各部位特徵之後,套上翼標、可顯示活動之
無線電發報器(美製 AVM 216 MHz 波段,重量 25g,3 年電池壽命)和腳環,並抽
血 0.1-0.2 毫升後將雛鳥放回巢位。繫放後 3 天,每天都回來觀察繫放個體狀況。
9
成鳥的繫放在冬季 11 至 2 月期間進行,在有大冠鷲固定獵食的地點進行捕
捉,以弓網及電動假蛇引誘,捕捉到的個體測量嘴鋒、跗躓、後爪、翼展、體重
各部份形值並拍攝各部位特徵之後,套上翼標、無線電發報器和腳環並抽血後原
地釋放。
採血方法為採肱骨靜脈 0.1-0.2 毫升血液量,保存在含無水酒精樣本館內。然
後以 PCR 分生技術鑑定性別,鑑定方法參考 Griffiths 等人方法(1996)。
第五節 無線電定位
無線電追蹤方式為兩人同時於不同地點各持一組美製 AVM 廠牌之 LA12-Q
接收器、方向天線和羅盤,由兩地點對準目標發報器的夾角,進行停棲訊號之三
角定位,以其交點做為動物當時位置。調查人員每次定位完畢後以 Garmin Vista
GPS 定出自身所在位置,並記錄觀測人員所在座標、定出交角時間、目標方位和
磁北的順時針方向夾角、天候、訊號強弱、訊號快慢。
10
第三章 方法
第四章 結 果
第一節 巢位尋找
今年 3 月至 6 月 24 日,在整個研究區內步行搜索猛禽的巢位。將發現之巢
位以 1 位英文字母加上 2 位阿拉伯數字編碼,英文字母用來區別物種,後兩碼則
沿用舊巢編號或如為新巢則另編新碼。巢樹則用兩位英文字母加上兩位阿拉伯數
字編碼,如為舊巢樹則沿用編號,如為新發現之巢樹則另編新碼,當探討不同年
間之相同巢位時則冠上西元年以玆區別。
本年度調查期間共發現 5 個大冠鷲巢位,和 11 個鳳頭蒼鷹巢位。其中 5 個
大冠鷲巢位(巢位代號 S19、S24、S25、S26、 S27)定位資料詳如表 4-1,另外
還發現 11 個共域的鳳頭蒼鷹巢位(TG13~TG23) ,各巢位分布如圖 4-1 所示。此
外,有 5 個 2004 年舊巢區(巢區代號 S16、S18、S20、S22、S23),於 3 至 5 月
調查持續觀察到空中對爪、林間求偶、乞食及低聲呼應等配對行為,但未發現新
巢或者舊巢再利用,6 月份之後的調查則未發現上述求偶或配對行為。
除上述巢位之外,8 月下旬由林試所員工轉送一隻幼鳥給研究人員,該幼鳥
係在研究區邊緣外之墾丁森林公園西側山坡被撿拾,拾獲者於 7 月 22 日發現一個
巢連幼鳥被海棠颱風吹落地,該幼鳥因被豢養超過月餘,且羽毛殘破,無法野放,
目前已轉送屏科大野生動物收容中心照料。
11
表 4-1 2005 年墾丁國家公園墾丁及社頂地區猛禽巢位調查摘要
巢位利用階段 物種 發現日期 巢樹編號 樹種 巢位編號 X座標 Y座標 海拔
築巢 孵化 育雛 離巢
大冠鷲 TS05 相思樹 S24 231445 2427349 100m 03_29
大冠鷲 TS10 相思樹 S19 232368 2428864 149 m 03_26
大冠鷲 TS17 相思樹 S25 233349 2424537 97m 05_25
大冠鷲 TS18 澀葉榕 S26 232057 2430227 215m06_07
大冠鷲 TS19 茄苳 S27 231817 2429385 189m 06_19
鳳頭蒼鷹 TG13 茄苳 G013 232555 2429235 - 03_17
鳳頭蒼鷹 TG14 毛柿 G014 232472 2428563 136m 03_26
鳳頭蒼鷹 TG15 茄苳 G015 232218 2428514 103m 03_26
鳳頭蒼鷹 TG16 毛柿 G016 232626 2430372 - 03_27
鳳頭蒼鷹 TG17 相思樹 G012 231700 2427638 159m 04_12
鳳頭蒼鷹 TG18 相思樹 G018 230975 2428087 - 04_13
鳳頭蒼鷹 TG19 山芒果 G005 232910 2429115 90m 04_14
鳳頭蒼鷹 TG20 欖仁 G020 229722 2429384 183m 04_18
鳳頭蒼鷹 TG21 木麻黃 G021 230042 2428085 85m 05_19
鳳頭蒼鷹 TG22 血桐 G022 233258 2430420 77m 05_21
鳳頭蒼鷹 TG23 毛柿 G023 233201 2429916 138m 05_21
資料來源:本研究
4 個今年繁殖之大冠鷲巢位中 S24 和 S26 兩個巢分別於 5 月上旬及 6 月中旬
發現棄巢,其中 S24 的巢樹已連續利用 3 年,在孵蛋期間因山老鼠在距巢樹 15m
附近非法盜伐數棵月橘,伐樹地點較巢位高且可直視巢內,研判干擾過大而棄巢。
棄卵目前則轉交特有生物研究保育中心採樣及收藏,棄巢後 1 個月內仍觀察到大
冠鷲雌、雄鳥在巢附近 100 公尺內活動,或在棄巢上方低飛,棄巢後一周觀察到
兩次雄鳥在交尾前特有之拉長低鳴,但未發現構築新巢行為。
代號 S26 的大冠鷲巢,築於茄苳上,發現後兩週再回去檢視,發現親鳥在巢
附近徘徊,但已不再孵蛋,距巢樹 10 公尺周圍幾棵毛柿樹下有大量經獼猴啃食後
的殘果,巢位傾斜鬆散,推測該巢可能受到獼猴騷擾而遭棄巢。
代號 S19 的巢位為一連續第 2 年利用的巢位,位於梅花鹿復育區第二區內的
相思樹上, 3 月 26 日發現修補舊巢的親鳥於巢樹飛起,5 月 6 日回去檢視已開始
孵蛋,5 月 20 日發現幼鳥已孵出,由體型判斷孵出約 5~6 天,之後陸續觀察 16
天次,共觀察 117 小時 11 分鐘。
S25 巢位發現於 5 月 25 日,發現時小鳥剛孵出 2 至 3 天,頸部稍能抬起,之
後受到人力限制僅觀察 5 天次,計 38 小時 58 分鐘。本巢位於鵝鸞鼻里蕃仔寮旁
12
第三章 方法
的熱帶海岸林內,本區同樣受到非法盜伐樹木的影響極為嚴重,但巢樹四周有小
礁岩屏障,藤蔓交錯,位置隱密,不易被發現。
S27 因 6 月 21 日發現雌鳥向雄鳥乞食的聲音才發現,由於十分靠近研究站而
被忽略,之後 3 天每天都可發現乞食聲,和雌鳥由林中飛出接過雄鳥食物。進入
林中搜尋發現巢樹所在位置藤蔓極為茂密,隱約可見巢位,巢樹種類亦無法靠近
辨識,暫不接近唯恐勉強進入牽動攀爬至樹頂的藤蔓造成棄巢。7 月 10 日觀察到
幼鳥在巢位附近山溝中活動後始測量巢樹。
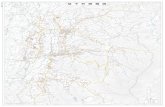
圖 4-1 2005 年墾丁地區猛禽巢位及配對行為分布地點
製圖:本研究
13
第二節 育雛食物種類及頻度
本年度截至 7 月 9 日止,育雛食物的觀察均是由 S19 和 S25 兩巢位附近搭設
的偽裝帳棚窺孔中以高倍率單筒望遠鏡觀察,共 41 筆觀察記錄,除了有 2 筆蛇類
種類無法確定,其餘獵物種類均可辨識。雖曾在巢下搜尋但並未發現任何食餘,
幼鳥吃不完的食物幾乎由親鳥吞食,或者攜出巢外附近巢樹放置一段時間後吞
食,沒有發現丟棄的食物或食餘。曾在每個巢位下方、附近大冠鷲親鳥固定棲息
的樹下或者其他有大量大冠鷲排遺的地點找尋食繭,但均無所獲,顯示大冠鷲對
無脊椎基丁質、甲殼類甲殼和蛇類骨骼等食物的消化能力相當高。
本年度由親鳥攜回的食物種類依次數多寡,分別為節肢動物的大型蜈蚣 23
筆;蛇類 12 筆;甲殼類的黃灰澤蟹和圓形圓盤蟹各 1 筆;兩生類的蟾蜍和澤蛙各
1 筆;蜥蜴類的攀木蜥蜴 1 筆;和哺乳類的赤腹松鼠 1 筆。其中兩筆螃蟹記錄則
均未發現餵食行為(詳見表 4-2)。其中 S19 巢和 S25 巢之平均遞食率分別為 0.24
隻次/小時和 0.31 隻次/小時,食物組成同樣以蜈蚣所佔隻次比例比例最高,蛇類
次之。
表 4-2 2005 年大冠鷲 S19 和 S25 巢位育雛生物種類和頻度
類別 食物 學名 小計 節肢動物 蜈蚣 Scolopendra subspinipes mutilans 22 蛇類 茶斑蛇 Psammodynastes pulverulentus 2
赤尾鮐 Trimeresurus stejnegeri 7 青蛇 Eurypholis major 2 龜殼花 Trimeresurus mucrosquamatus 1
甲殼類 黃灰澤蟹 Geothelphusa albogilva 1 圓形圓盤蟹 Discoplax rotundum 1
兩生類 黑眶蟾蜍 Bufo melanostictus 1 澤蛙 Rana limnocharis B 1
哺乳類 赤腹松鼠 Callosciurus erythraeus 1 蜥蜴類 攀木蜥蜴 Japalura swinhonis 1
總計 40 資料來源:本研究
14
第三章 方法
第三節 棲地偏好
棲地的特色由兩個尺度來探討,一個是繁殖所需的巢樹樹種、巢位所在之位
置、地形等微棲地特徵;另一為領域層次的需求,如活動範圍內地景的組成,或
者和重要地景之間的距離等的巨棲地特徵。
一、微棲地特徵
(一)巢樹的種類
本年度所發現之巢位有三個巢位於相思樹上,一巢位於澀葉榕上,一巢
位於茄苳樹上。綜合本年度和過往的記錄顯示,大冠鷲築巢偏好選擇豆科的
相思樹(41.9%)和大戟科的茄苳(38.7%)(圖 4-2),其餘物種(19.4%)僅
利用 1~2 次。
圖 4-2 1995 年至 2005 年墾丁地區大冠鷲利用巢樹種類次數
0
2
4
6
8
10
12
14
茄苳 相思樹 赤楠 血桐 無患子 澀葉榕 蟲屎
種類
利用
次數
製圖:本研究
15
(二)巢樹的特徵
本年度大冠鷲所選擇之 5 棵巢樹平均樹高 14.02 公尺,胸徑 41.66 公分,
樹冠高度為 12.53 公尺,樹冠層平均覆蓋度為 75.14%,經和逢機取樣的樹比
較之後,發現本年度的巢樹尺寸特徵和逢機選取之巢樹並無明顯差異(表
4-3),此結果和往年研究結論不同。
表 4-3 2005 年大冠鷲利用巢樹和逢機取樣樹木特徵比較
巢樹 逢機取樣 變數 平均值 標準差 標準差 n n P 平均值
平均樹冠高(m) 12.53 3.33 5 12.64 2.46 5 0.89平均覆蓋(%) 75.14 11.92 5 71.04 12.28 5 0.61
樹高(m) 14.02 2.53 5 13.12 2.24 5 0.47胸徑(cm) 41.66 11.55 5 38.68 4.84 5 0.64
資料來源:本研究
(三)巢位的再利用
S24 巢所位在的 TS05 這棵巢樹,在 2000 年至 2005 年之 6 年期間,共
使用 5 次,沒有使用的 1 年為 2001 年,係移到在旁邊距離 28 公尺的一棵蟲
屎上,所有的巢位均位於藤蔓交織的樹幹上,本年度若不是山老鼠騷擾應該
應該可以順利使用。
這棵 TS05 的巢樹,於 5 次利用期間,分別築了 4 個巢,其中 2003 年
和 2004 年都使用同一個巢,2000 年、2002 年、和 2004 年沒有使用前一年
巢位的原因分別為繁殖季結束後巢位被落山風破壞(2000 年);藤蔓過度茂
密蓋住巢位(2002 年、2004 年);以及藤蔓生長旺盛將巢位托離支撐的樹幹。
而 2001 年蟲屎樹上的巢沒有再利用的原因為支撐巢的樹幹被颱風吹斷。
另外 S19 所位居的巢樹 TS10 也是本年度重複利用的巢樹,本巢雖然重
複利用,但因巢位同樣受到生長過度旺盛的藤蔓將巢位托離支撐的樹幹,雖
然躲過去年的杜鵑颱風,但後來被海棠颱風吹落。
上述現象顯示大冠鷲對於巢樹的利用,可能具有相當大的執著性,本區
16
第三章 方法
過往其他連續使用的巢位或巢樹亦具有類似的情形。推測如果不是好的巢樹
有限,就是為了減少在築巢上的能量投資,選擇一個好的營巢地點可以增加
其繁殖成功率。此外,在森林中要選擇一個少有天敵(猴子、人類等)騷擾
的地點,而且地形上必須要能避風,所選擇的樹種要高大穩固支撐力夠並使
巢位遠離地面,最好樹幹上纏繞藤蔓,以增加巢的穩定性以躲避落山風和颱
風,如果巢的附近有許多枝葉屏蔽則更佳,綜合種種條件並不容易。
(四)巢的特徵
本年度無法接近的巢位為 S26,其餘 4 個可接近之巢位經測量之巢的平
均長寬尺寸為 58.13cm,平均的厚度為 38.25cm,深度則為 11.0cm(表 4-4)。
至於巢離地的高度為 10.2m,距主幹距離為 0.38m,支撐巢的樹幹數目為 2.2
根,支撐巢的的樹枝截面積總和平均為 300.5cm(表 4-5)。
表 4-4 2005 年大冠鷲巢尺寸
變數 平均值 標準偏差 n 直徑(cm) 58.13 1.75 4 巢厚(cm) 38.25 9.07 4 巢深(cm) 11.00 4.08 4
資料來源:本研究
表 4-5 大冠鷲巢位特徵
變數 平均值 標準差 n 巢離地高度(m) 10.2 1.92 5 巢至主幹距離(m) 0.38 0.58 5
支撐樹幹數 2.2 1.30 4 支撐枝截面積總和(cm2) 300.5 67.75 4 資料來源:本研究
17
(五)巢樹下層植被
經統計本年度所利用之五棵巢樹之下層植被,胸徑大於 1 公分之莖
幹總數平均為 116.3 根,約每 10 平方公尺一根,莖幹平均截面積總和為
18153.2 公分,林下草本層之覆蓋度為 16.4%(表 4-6)。
表 4-6 2005 年繁殖大冠鷲巢樹林下莖幹數量及草本層和灌叢覆蓋度
變數 平均值 標準差 N 莖幹數 116.3 21.5 5
莖幹截面積總和(cm2) 18153.2 5722.4 林下覆蓋度(%) 16.4 4.4 5
資料來源:本研究
二、巨棲地特徵
(一)巢樹所在的地形特徵
本年度所發現之大冠鷲利用巢樹,有兩棵位於鵝鑾鼻半島西側,3 棵位
於東北季風和大多數颱風路線衝風面之半島東側(表 4-7)。若彙整 1995 年
至 2005 年間所發現之大冠鷲巢位所在地形,發現迎東北季風的鵝鸞鼻半島
東坡和西坡所發現的巢樹累計數量相當,但位於山溝的的比例高於山坡或坡
腳,無位於懸崖、平原、山脊的巢位(表 4-8)。可見微地形蔽風效應的重要
性可能不亞於大尺度的地形效應。此外根據彙整 1995 年至 2005 年 10 年間
本區所發現之巢樹(表 4-9),以南向坡生長的巢樹數量最多,可能為南向坡
較能提供落山風和來自東方和南方颱風的遮蔽。由於許多巢位和幼鳥於颱風
後折損,可見季風和颱風可能為大冠鷲巢位選擇的重要因子,由地形上的巢
樹分布差異,亦可見微地形上的避風效應的相當重要。
表 4-7 2005 年大冠鷲巢樹所在地形
巢樹編號 在半島上的位置 地形 巢樹所在坡向 T005 西側 山溝 SE T012 東側 山溝 S
西側 山腳 T017 SW T018 東側 山坡 SE T019 東側 山溝 S
資料來源:本研究
18
第三章 方法
表 4-8 1995 年至 2005 年間所發現之大冠鷲巢樹所在地形百分比
在半島上的位置 山坡 山溝 百分比 西 2 6 42.11% 東 3 8 57.89%
百分比 26.32% 73.68% 資料來源:本研究
表 4-9 1995-2005 巢樹所在坡度數量統計
坡向 北 東北 東 東南 南 西南 西 西北 次數 1 1 1 2 6 3 2 3
資料來源:本研究
(二)2005 年大冠鷲巢樹和不同地景或人造建物距離
表 4-10 為利用 GIS 系統求得之各巢樹距離最近人工建物、最近永久水
源、無鋪面的道路或小徑、有鋪設柏油或混凝土的道路以及低破碎化森林之
林緣距離分別為 442.3m、353.1m、160.9m、 690.8m、 273.9m,這些項目
距離中除無鋪面道路之外,均較曾經發現之最近巢間距為大(表 4-10)。距
離最近人工建物、有鋪面道路以及林緣的資訊雖然需要進一步的推論分析,
但約略可以顯示大冠鷲對人為擾動的容忍程度可能小於對同類的競爭。距離
永久水源的的距離顯示社頂地區地勢較高,永久地面水源缺乏,但飛行能力
或許可以彌補這方面資源不足,但可能得面對其他競爭者的壓力。距離林緣
較遠顯示大冠鷲築巢偏好於森林內部遠離外界干擾區域,因此連續森林面積
可能較破碎環境更有利大冠鷲繁殖利用。
表 4-10 2005 年大冠鷲利用巢樹和重要地景距離
變數 平均值 標準差 n 距建物(m) 442.3 151.8 5 距水源(m) 353.1 254.9 5 無鋪道路(m) 160.9 121.6 5 鋪面道路(m) 690.8 526.9 5 距林緣(m) 273.9 110.8 5
資料來源:本研究
19
(三)研究區植被分布比例
藉由墾丁國家公園社頂地區發展生態旅遊規劃一案之幫助,得以將研究
區現今多樣化之植被組成,依其不同植相類型如高位珊瑚礁原始林、毛柿林
區、農耕地、次生林、草生地、海岸灌叢、珊瑚礁林投優勢海岸、沙灘、崩
塌地及泥岩惡地、相思樹及木麻黃造林地、檳榔園及竹園、水塘等(表 4-11),
以正射化 97 空照數位圖為底圖,利用 ArcGIS 8.3 製作各種土地植被分布圖
層(圖 4-3),各區和大冠鷲利用之狀況說明如下:
表 4-11 研究區域各種植被及土地類型面積及百分比
類別 面積(公頃) 百分比 次生林 1001.5 43.5% 椰子園 7.3 0.3%
公共設施用地 84.6 3.7% 檳榔園及竹林 8.3 0.4%
海岸林 57.5 2.5% 造林地 48.4 2.1% 農耕地 31.2 1.4% 裸地 55.3 2.4% 草生地 450.5 19.6% 珊瑚礁 85.6 3.7% 海岸灌叢 323.2 14.0%
高位珊瑚礁森林 95.7 4.2% 毛柿林 22.2 1.0% 水塘 1.4 0.1% 沙灘 30.6 1.3% 總計 2303.3 100.0%
資料來源:本研究
20
第三章 方法
圖 4-3 研究區位主要土地植被型態和分布
製圖:本研究
1.高位珊瑚礁原始林
位於墾丁國家公園範圍內之高位珊瑚礁森林是台灣面積最大,且保存
最完整之高位珊瑚礁森林生態系,目前保留較完整之部分為林業試驗所管
轄之試驗林地,目前已設立為”墾丁高位珊瑚礁自然保留區”。其植被屬熱
帶季風林群系,為台灣少見之高位珊瑚礁植物社會,主要可區分為下列四
個生育地型,皮孫木型分布於凹陷谷地,茄苳-台灣膠木型分布於隆起台
地,鐵色、紅柴、樹青、月橘型分布於隆起岩塊,蟲屎、血桐、土楠型分
布於早期人為干擾盛行之平坦谷地。此一森林是台灣唯一一處以柿樹科植
物為最優勢的植物社會,其植物組成與台灣島其它地區明顯不同。
研究期間曾利用陪同林試所工作人員巡山機會,於高位珊瑚礁保留區
圍籬外和梅花鹿復育區研究站東側的高位珊瑚礁原始林步行搜索大冠鷲巢
位,由 1995 年至今,本區僅於 6 月 7 日發現編號 S26 之大冠鷲巢位,但之
後根據獼猴排遺和巢樹旁滿地毛柿落果推測被獼猴干擾而棄巢。推測本區
較成熟完整之林相可能有較多大冠鷲天敵,而且多數藤蔓不易爬到森林高
21
處,因此可供大冠鷲利用之條件環境不多。
2.毛柿林區
位於梅花鹿復育區東北側之毛柿造林地隸屬林務局屏東林區管理處
恆春事業區地第 36 林班地,自民國 9 年起陸續造林,共有造林面積 140
公頃,為現今全台最大之毛柿造林地。毛柿漿果碩大,在果實剛形成時會
吸引獼猴、松鼠啃食和食果性鳥類啄食,果實成熟尚未掉落前白鼻心也會
被毛柿成熟的香氣吸引,掉落地面後為梅花鹿、山猪、及食蛇龜等動物的
食物之一。本區曾發現三巢鳳頭蒼鷹,但從未發現過大冠鷲巢位。
3.農耕地
過去原住民早期耕地的開闢以燒墾、輪耕方式為主,選擇坡地種植旱
稻、甘藷或芋頭,漢民族遷居此地後開始引入水稻田栽植技術,除低窪地
外,多採用梯田方式種植。目前除少數仍有耕作外,農耕地大多已陸續廢
耕成為次生林地,數木多不高大也不茂密,無法被猛禽築巢利用,但常可
見大冠鷲棲息在這些林冠不鬱密區域的樹梢覓食。現存農耕地主要散佈在
聚落週遭和毛柿林區東南側。
4.次生林
次生林為本研究區域內面積最大的植被類型,現存之次生林主要為國
家公園成立之後廢耕之旱田、水稻田、瓊麻園等農地演替而來,另有一部
分為林地燒墾闢地之後漸次復原。墾丁國家公園成立後,徵收了本區大部
分之私有土地,改闢為社頂自然公園及梅花鹿復育區,經造林後林相才慢
慢的經由少數留存於開墾不易之礁岩上或溪溝中之森林自然演替恢復成現
所見之次生林。就現階段演替過程而言,馬纓丹及芒草等植物為此區次生
林演替過程中最早進入之種類,緊接著為構樹、血桐、相思樹、番石榴、
土樟、銀合歡、蟲屎、過山香等樹種,地被及灌木層則以弓果黍、決明、
長穗木等占優勢,亦有為數不少之兩耳草、賽葵、土牛膝等植物。
過往調查顯示,在這個區域內所見到之猛禽巢位,顯然位於較週遭植
22
第三章 方法
被高大之相思樹或茄苳之上,過往燒伐的歷史使得陡峭地形和溪谷區域的
樹木得以保留,而陡峭和溪溝地形使得東北季風無法長驅直入,因此成為
猛禽營巢偏好利用的地方。溪谷林冠鬱密的樹林,往往因林下缺乏陽光而
植被較為稀疏,加上梅花鹿利用,地表往往較為光禿,有時可成為大冠鷲
的獵場。
5.草生地
草生地的形成大多是農田廢耕、長期的過度放牧、或刻意的種植牧草
(畜產試驗所牧草區)所形成。區域內之廢耕農地多以長穗木為優勢種,混
雜龍爪茅、含羞草、野棉花等植物;而長期放牧所形成之草生地除蠅翼草、
鍊莢豆、含羞草之外,則以地下莖強勁耐旱之竹節草分佈最多最廣,伴生
植物有台灣灰毛豆、早田氏爵床、雞角刺、圓葉土丁桂、白茅等草本植物。
6.海岸灌叢及芒草
主要分布於東側之迎風坡面,受冬季強力東北季風的影響,樹形及分
枝多呈現俯臥狀,除零星分佈之台灣海棗外,以林投或芒草之分布為主。
這區的環境看似貧瘠,但在風吹沙、刣牛溪、甚至龍坑附近的礁岩地形,
冬末春初最乾旱的季節,偶爾可見到大冠鷲在此捕食陸蟹和大蝗。因為植
被低矮,缺乏隱蔽,並非大冠鷲主要的棲息環境。
7.珊瑚礁海岸
珊瑚礁石的形成,主要由死亡的珊瑚蟲骨骼、以及部分石灰藻類、有
孔蟲及貝殼等逐漸堆積而成,經過地形抬昇而露出海面成為珊瑚礁海岸。
珊瑚礁石塊表面僅有少量土沙堆積,加上鹽霧影響,可以生存植物種類極
少,以水芫花及乾溝飄拂草為代表。
8.沙灘
本區沙灘之沉積物以石英砂或珊瑚砂為主,故沙灘呈現白黃色。在東
海岸發達的礁岩海岸中,夾雜著少許的沙岸,其中以風吹砂為最著名。植
23
物之種類以馬鞍藤、海埔姜、濱刺草等為主。
9.裸地、崩塌地及泥岩惡地
由於天然崩塌或人為開墾後經雨水嚴重沖蝕所造成的泥岩裸地,主要
分布於刣牛溪支流交會處附近區域,形成本區之特殊景觀。崩塌地上僅見
少數台灣海棗存活,可見台灣海棗為崩塌地演替之先期樹種。
10.相思樹及木麻黃造林地
台灣之造林事業,由日人入據初期(1900 年代)即開始,最初從事樟
樹造林,以供應製樟原料;後亦推廣保安造林、熱帶樹種引進造林等。
相思樹為一般經濟造林之主要樹種,不論在二次大戰前或後,木材的
需求量都非常的大,故造林的樹種以培育短期可以伐採之速生樹種為主。
目前在社頂以西和以南區域的樹林以相思樹為主要樹種,為畜試所過去 20
年來牛羊畜養面積漸減之後分區造林形成。雖然相思林在本區的種植面積
相當大,但造林區域很少進入溪谷,所以相思樹雖然數量多,似乎並非左
右大冠鷲選擇本樹種的原因。
枝條造型特殊的木麻黃則為因應每年的颱風肆虐及盛行的東北季
風,而成為海岸防風造林之最佳樹種,被廣泛種植於恆春半島。但在社頂
至墾丁和社頂至船帆石區內,木麻黃的數量不多,由墾丁至社頂的縣道旁
沿路分布生長,由於壽命問題,許多早期種植之木麻黃逐漸死去。本計畫
和過往研究曾發現鳳頭蒼鷹利用木麻黃,大冠鷲則無。
11.檳榔園及竹林
位於梅花鹿復育區最東側和大坪頂邊坡的帶狀檳榔園及竹林,是早期
在山上開墾及居住的原住民或及漢人所栽植,面積雖然不小,但因樹形構
造簡單,不為大冠鷲和鳳頭蒼鷹棲築巢利用。但調查期間於稀疏的林下,
偶爾可以發現由地面受驚嚇飛起的大冠鷲個體。
24
第三章 方法
12.水塘
早期因人為的開挖或是耕作地的低窪處,廢棄後在雨季或非雨季時形
成臨時水塘,水塘經時間的累積而形成一小型之生態系,吸引了許多的生
物。常見冬季乾旱季節,溪流幾乎完全乾涸,此時大冠鷲常出現在水塘週
遭地面捕食黃灰澤蟹。
(四)巢區土地植被類型分布比例
參考往年研究區域大冠鷲最近巢間距約在 350 公尺左右,因此巨棲地特
徵之植被及土地類型測量僅以巢為中心之半徑 250 公尺為範圍,並分析各半
徑範圍內植被型態比例。至於林緣長度因受到林相過於破碎和無明顯交界,
現地勘查和 GIS 圖層難以謀合,做圖和計算將有誤差,本年度暫不處理此問
題。
綜合各巢區植被,發現土地植被類型僅有前述 12 類土地植被類型中之
7 類,所占面積多寡依序為次生林、高位珊瑚礁森林、裸地、海岸林、草生
地、珊瑚礁岩、水塘等。森林類型有次生林、和高位珊瑚礁森林和海岸林,
占全部面積九成以上,其餘為鑲嵌在森林中小面積之草生地、裸地、水塘、
珊瑚礁岩等植被貧乏區域,所佔比例極低,可見巢區環境選擇對於森林的依
賴性相當高(詳見表 4-12)。
表 4-12 2005 年五個繁殖巢區植被土地類型面積及比例
類型/巢區
S24 S13 S25 S26 S27 小計 百分比次生林 19.366 18.667 15.207 0 7.938 61.178 62.4%
高位珊瑚森林 0 0 0 19.622 11.023 30.645 31.2% 裸地 0.248 1.000 0.694 0 1.003 2.522 2.6% 海岸林 0 0 2.477 0 0 2.477 2.5% 草生地 0.008 0 1.164 0 0 1.172 1.2% 珊瑚礁岩 0 0 0.093 0 0 0.093 0.1% 水塘 0 0 0 0 0.002 0.065 0.1% 總計 19.622 19.622 19.622 19.622 19.622 98.110 100.0%
單位:公頃 資料來源:本研究
25
(五)平均密度
研究區總面積 2303.3 公頃,區域內森林總面積(1240.9 頃),除以穩定
觀察到的大冠鷲配對數目(今年 5 巢和 5 對繁殖行為展示;2004 年 8 巢和 3
對繁殖行為展示),若以 10 個繁殖對推估,則平均營巢密度小於 230 公頃/
對,若扣掉非森林區域為 124 公頃/對,這其中還包括品質不良的森林如造林
地、檳榔園、農耕地等,如果再考慮森林邊緣不適區域,則可用區域將更少。
即使情況如上所述,本研究區仍有高於國外現有許多文獻提及之營巢密度
(Ferguson-lees and Christie 2001;Morioka 1995),因此如果能夠藉由後續繫
放工作,收集研究區所有個體數量、活動範圍等資料,將可以獲得本物種棲
地結構和需求等詳細訊息。
第四節 無線電定位
本年度 2 月至 11 月共追蹤 6 隻繫放個體,其中 3 隻為 2004 年 6 月繫放的幼
鳥,1 隻為今年 2 月,2 隻為 11 月期間繫放之成體。本年度捕捉成鳥外部量值,
詳如表 4-13,繫掛之翼標、腳環、無線電發報器詳如表 4-14。
無線電定位之個體依據個別揹負之發報器之發射頻率給予編號,由 2 月份開
始,代號 063、083、120,3 隻鳥夜間夜棲地定位分別記錄 14、25、109 夜;白天
由清晨至終昏各定位 6、8、19 天數,代號 020 無線電發報器失效,僅有零星目擊
記錄 13 次。成鳥代號 138 及 258 個體均在 11 月捕捉及野放,日間活動各記錄 6
及 4 天,及夜棲地定位各為 15 及 10 夜。
表 4-13 2005 年捕捉成鳥之外型量値
環號 喙長 全頭長 全頭寬 跗趾長 跗趾徑 後爪長 重量 年齡
KTNP011 33.8 94.4 54.1 103.2 14.6x10.3 28.8 1900g 1-2 molting cycle
KTNP6258 32.1 98.2 42.5 95.0 13.6x10.2 25.0 1850g >3 molting cycle
KTNP137 35.7 100.0 52.2 - 19.5x11.2 28.3 2300g >3 molting cycle註 1:長度單位 mm 資料來源:本研究
26
第三章 方法
表 4-14 2005 年成鳥繫放項目一覽表
繫放日期 野放日期 環號 翼標 發報器
型式 顏色編號 廠牌 型式 頻率(MHz)
20050130 20050207 KTNP011 肩套掛式 白 無 AVM 腰掛 216.120
20051108 20051108 KTNP6258 肩套掛式 黃 無 AVM 腰掛 216.258
20051101 20051102 KTNP137 肩套掛式 黃 B AVM 腰掛 216.137 資料來源:本研究
表 4-15 2005 年 2 月至 11 月無線電定位個體天數一覽表
代號註 1 年齡 出生巢/捕捉地 日活動範圍定位天數 夜棲地定位次數063 幼鳥 S22 6 14 083 幼鳥 S16 8 25
成鳥 020 S18 -註 2 - 120 成鳥 社頂公園 19 109 138 成鳥 社頂公園 6 15 258 成鳥 社頂公園 4 10
註 1:無線電定位個體代號採所揹負無線電發報器頻率後 3 碼 註 2:無線電發報器失效,但仍可目擊 資料來源:本研究
由無線電定位工作發現,代號 063、083 亞成鳥個體由出生後至翌年 3 月活
動和夜棲範圍大致在出生地半徑 0.5 公里至 1 公里半左右範圍內活動(圖 4-4、
4-5),3 月份起白天呈現不規律的遊走活動現象,但夜間有時會回到原來出生地附
近活動。代號 020 雖然無法透過無線電定位知道其位置,但透過調查人員、鳥會
賞鳥者、梅花鹿研究站工作人員等人通報 27 次,發現呈現相似的活動類型。
代號 120 成熟個體,為未配對繁殖之流浪漢,在各個領域間遊走,白天常在
不同的區域覓食,常幾天停留在一個地方,然後換到另一個位置再停留數天,夜
棲位置通常靠近其白天活動地點。遇到風大的天氣如颱風和落山風往往收訊訊號
較弱,推測躲到避風的山谷中,這些定位位置距離定位者或者是梅花鹿工作站的
距離有時不會太遠(由收訊強度和角度寬窄判斷)(圖 4-6)。
代號 138 及 258 個體為最近才捕捉到的個體,經繫放之後野放約 1 個月左右,
這兩隻成體所呈現活動範圍相當小,約在 1 平方公里左右的範圍內,由換羽狀況
判斷,這兩隻個體個體均換羽至少 3 輪,以非正式發表之動物園內圈養經驗,大
冠鷲初級飛羽換過 1 輪至少 3 至 4 年,視個體狀況而有差異,野外個體也許因羽
27
毛折損而換的更快或者營養不穩定而較慢,但約略在 3 年左右,因此推斷這兩隻
成體年齡應該都在 10 歲以上,也因此可能較了解環境和有獵捕經驗,因此活動範
圍較未擴散幼體(063、083)或帶有第 1 輪飛羽的個體 120 來的小,至於這是否
是一般性的趨勢尚需更多資料補充(圖 4-7、圖 4-8)。
冬季定位發現大冠鷲喜歡停留在山溝或溪流還有殘存地面水或潮濕土壤的
地點活動,因此許多個體定位發現活動地點密切重疊,例如出生地接近的 063 和
083 在有地面水的區域定位範圍重疊較多(圖 4-7),這個季節在水源週遭還常能
發現的獵物以黃灰澤蟹為主,有時一個地點可以看到 10~20 個被大冠鷲攝食後的
黃灰澤蟹的甲殼。曾經在大圓山腳之屏鵝公路路肩觀察 S22 亞成鳥(過往調查)
在一個上午進食 9 隻黃灰澤蟹,和追逐攀木蜥蜴。
所有個體訊號曾在梅雨季節來臨前分別消失 1 至 2 個月,曾至恆春、港口和
滿州測蒐,發現 083 停留在出火附近的山頭,停留了 2 週。研判應該是研究區域
久旱不雨,梅雨降雨線還未抵達恆春,而滿州附近山頭卻經常烏雲密佈,因此吸
引大冠鷲離開平常棲息範圍前往覓食。但也有可能是受到繁殖個體驅趕。但在 6
月初,除 083 外,所有追蹤個體在梅雨季節期間都回到原來測蒐的地點。
圖 4-4 063 個體定位範圍
製圖:本研究
28
第三章 方法
圖 4-5 083 個體定位範圍
製圖:本研究
圖 4-6 120 個體間定位範圍
製圖:本研究
29
圖 4-7 138 個體定位範圍
製圖:本研究
圖 4-8 258 個體定位範圍
製圖:本研究
30
第三章 方法
第五章 討 論
由巢位找尋的經驗發現,由於大冠鷲的巢位相當隱密,尋找必須仰賴地毯式
搜索,將研究區內的大樹一棵一棵樹尋找,才比較不會有遺漏的情形,過往研究
(Chou et al., 2002)和本研究過往調查顯示大冠鷲的繁殖成功率相當高,約在九
成左右,颱風天候、人為干擾和獼猴騷擾為目前所知之失敗原因。至於離巢後幼
鳥的存活率根據本年度和過往調查,推斷幼鳥第一年的存活率可能不到 6 成左
右,颱風可能是造成多數已離巢之幼鳥死亡的主因。
由繁殖巢邊觀察和野外搜尋發現,雖然蛇類和節肢動物的蜈蚣為大冠鷲主要
的食物種類,但亦不排除兩棲類、蜥蜴類、哺乳類等生物,在食物不足時亦會以
螃蟹為食。
許多研究人員(Crocker-Bedford and Chany 1988, Kennedy 1988, Curnutt and
Robertson 1994) 證明幾種猛禽營巢需要固定的環境特色,或選擇特殊的地點築
巢。由大冠鷲巢樹的觀察可以發現墾丁地區族群築巢偏好選擇相思樹和茄苳,其
餘植物利用較少,由其餘物種利用較少,似乎可以反映前述現象。雖然本年度大
冠鷲巢樹經統計檢定結果不像本區過往研究顯示巢樹和周圍逢機取樣樹木有明顯
差異,可能是年間差異或取樣數的關係。
Cody (1985) 認為鳥類會選擇特定的地點以得到最大的繁殖成功率,其中至
少有 3 個因子被認為會影響鳥類的巢位選擇(1) 巢位被捕食,(2) 環境對蛋和幼雛
的壓迫(Cody 1985),(3) 親鳥的可及性 (Bednarz and Dinsmore 1982)。如果這些現
象可以套用在大冠鷲身上,則高大的樹有利於躲避敵害;穩固的築巢位置可以減
少強風的破壞和蛋及幼雛的脫水效應;而較稀疏的枝幹則有利於親鳥出入穿越。
墾丁地區最重要的環境壓力因子可能是颱風和冬天的東北季風。這個地區的
植被大多受到東北季風的調控,較不抗風的熱帶雨林僅分布於墾丁鵝鸞鼻半島的
西南坡以及山谷或珊瑚礁峽谷中。而較抗風的植被如硬葉樹森林、灌叢、草生地
則分布在露出的山脊和北向坡(Su 1985)。並且墾丁地區鳥巢曝露的方向也可能受
31
季風的影響。例如, Yuang (1996)研究指出墾丁地區小雲雀(Alauda gulgula) 巢的
開口在 3 月份朝向西和南方,此時冬北季風仍然吹拂。可是 4、5 月份築巢的小雲
雀,其巢的開口就沒有很強的方向性。多數大冠鷲的巢多位於向西、南、和西南
的山坡上。 這種分布型式和繁殖季初期未減的東北季風有關。多數鷲巢深入每一
塊森林內部的部分原因可能是這樣的安置可以庇護巢免於風的影響。
Brown and Amadon (1968) 的報告顯示蒼鷹的巢靠近溪流或池塘,本研究多
數鷲巢位於溪谷中,可能和較高的溼度及地形上的庇護有關,如此一來也可以改
善極端環境和風的影響。
Newton (1979) 認為缺乏巢位可能限制幾種猛禽的族群繁殖。由過往觀察資
料顯示本研究區域的大冠鷲繁殖族群年間變動很大,例如最近 3 年資料顯示 2003
年的五對,2004 年的 8 對,今年的 5 對,許多去年舊巢遭天候毀損的巢區,多數
今年沒有在繁殖區附近在發現築巢或繁殖,這個觀察似乎也反映了前述現象。本
研究顯示大冠鷲對於巢樹的利用,可能具有相當大的執著性,本區過往其他連續
使用的巢位或巢樹亦具有類似的情形。推測如果不是好的巢樹有限,就是為了減
少在築巢上的能量投資,選擇一個好的營巢地點可以增加其繁殖成功率。此外,
在森林中要選擇一個少有天敵(猴子、人類等)騷擾的地點,而且地形上必須要
能避風,所選擇的樹種要高大穩固支撐力夠並使巢位遠離地面,最好樹幹上纏繞
藤蔓,以增加巢的穩定性以躲避落山風和颱風,如果巢的附近有許多枝葉屏蔽則
更佳,綜合種種條件並不容易。
大冠鷲狩獵環境相當多樣化,由人為活動的村莊附近,到茶園、果園等各式
農耕地,以及原生及次生林地等(Ferguson-lees and Christie 2001),顯示這種猛禽
對環境的適應性較其他種類來的佔優勢。因此其族群被認為係鷲鷹科中最廣布的
種類。在環境理想的情況下其活動範圍可以小到 5-6 平方公里(Ferguson-lees and
Christie 2001),巢位間隔 1.5 至 2.0 公里(Morioka 1995)。然而本研究顯示其巢位
選擇尺度的領域需求可以比上述密度還高,最近巢間距約在 350 公尺左右。較近
的巢間距有可能的解釋為此處食物豐度較高,或者棲地破碎造成巢區被壓縮,或
者是行為上的適應,也有可能是覓食區和巢區分離的,不論是上述哪些原因,均
32
第三章 方法
需要更多的資料收集才能佐證。
第六章 建 議
建議一 森林環境的保護和管理:立即可行建議
主辦機關:內政部營建署墾丁國家公園管理處
協辦機關:內政部警政署墾丁國家公園警察隊
期程與做法:有急迫性,應該列為例行工作。墾管處商請警察隊便衣每週不定期
巡邏各處森林至少兩次,並請兩單位定期檢討工作狀況。
由於大冠鷲需要相當大的樹林得以將巢安置於遠離林緣的區域,但是遭到盜伐
騷擾,因此建議在國家公園內現存的樹林應該小心被保護,特別是位於溪谷和珊瑚礁
週遭的區域,不要再任其切割破碎,因為這些區域是其遮蔽東北季風的重要棲地。大
冠鷲所選擇繁殖的地點和樹種資料顯示,其偏好環境主要在溪溝和山谷地形的高大植
株上,因此對於上述環境的保護和管理有其重要性。
建議二 獼猴數量管理的規劃:中長期建議
主辦機關:內政部營建署墾丁國家公園管理處
協辦機關:彌猴研究相關學者或院校
期程與做法:建議於兩年內實施為期二至三年之短期委託專案研究獼猴族群和活
動範圍變化以及和遊客互動狀況,長期提出前瞻性的經營管理按部就班做
法。
根據過往研究資料顯示,本研究區大冠鷲的繁殖成功率相當高,超過 90%,但
今年所發現的五個巢位中有一個大冠鷲和兩個鳳頭蒼鷹的巢被獼猴騷擾而棄巢,這個
情形過去 3 年沒有發現過,可能是保育獼猴後,其族群數目逐漸增加,和遊客的接觸
也隨之提高,建議未來可將獼猴數量的管理納入經營方向。
建議三 牛隻和鹿隻管理的籌畫:立即可行建議
主辦機關:內政部營建署墾丁國家公園管理處
協辦機關:內政部警政署墾丁國家公園警察隊
期程與做法:短期(一年內)請懇管處檢討梅花鹿復育的階段性任務可否朝向展
33
示教育可行性,並提出加強非法放牧牛隻管理辦法;中期(兩年內)委託
專案研究梅花鹿和放牧對森林小苗更新的影響,並提出管理策略。
非法放牧的牛隻和復育的梅花鹿可能對森林地面水源和植被更新產生長遠影
響,進而影響食物鏈結構。
34
附錄一
附 錄
附錄一、服務企劃書評審會議紀錄
開會日期:中華民國 94 年 2 月 21 日上午 10 時 0 分 記錄:蔡乙榮
開會地點:墾丁國家公園管理處大型會議室
主持(召集)人:李副處長登志
評審委員:
高雄醫學大學 程委員建中
樹德科技大學 羅委員柳墀
墾丁國家公園管理處 林委員欽旭
墾丁國家公園管理處 馬委員協群
墾丁國家公園管理處 林委員文敏
出列席單位及人員:
墾丁國家公園管理處 許書國 陳榮祥
周大慶
35
主席致辭:首先感謝程老師及羅老師前來協助本處委託研究計畫案評審,評審會議開
始。
業務單位報告:略
參選單位簡報:詳如服務企劃書
會議討論:
一、從計畫名稱來看,這個計畫是一個長期監測計畫嗎?(羅委員柳墀)
回應:預定進行 3年,94 年委託案是 3年計畫的第 1年。(蔡乙榮技士)
二、既然是一個長期監測計畫,是否已有選定可以看出差異的指標來監測其中的變
化。(羅委員柳墀)
回應:預訂以大冠鷲的營巢數目或繁殖配對數做為直接的監測指標,因為繁殖配對
數可以反映出棲地環境的狀況,雖然其中可能還有其他的影響因素,但是,
繁殖配對數或營巢數目還是一個可以採用監測環境變化的指標。(周大慶)
三、選擇以繁殖配對數做為監測指標可能需要注意幾個問題:(一)大冠鷲 1 季是
否只繁殖 1次?有些猛禽是可能會在繁殖失敗後,在同一季再重新築巢進行繁
殖。(二)有些在食物營養充足的環境下,也有可能繁殖 2 次。大冠鷲是否有
可能 1季繁殖 2次?無論如何,如果是以繁殖配對數做為監測指標應該是可行
的,但是,若以營巢數做為監測指標的話,應該是比較不準確的。(羅委員柳
墀)
回應:以上的 2個問題在研究過程中都會加以注意,但是就目前所發現的大冠鷲繁
殖紀錄中,繁殖成功率幾乎多在 9成以上,除非牠的小鳥被人捉了,否則在
這個區域中,大冠鷲幾乎沒有天敵,目前所發現的巢位都是築在沒有猴子活
動的地方,不過若是遇到巢壞了是不是會再築 1個新巢,這一點在研究中會
加以注意。至於會不會因為食物充足而 1季繁殖 2次,這一點不太可能,因
為牠的雛鳥需要很長的時間才會離巢,而且在離巢後親鳥還會繼續的照顧一
段很長的時間,至少還會提供食物 1至 2個月。就目前還在進行無線電追踪
的幾隻幼鳥,已經有 8個月的時間,牠們還都在巢區附近活動,所以應該不
會 1季繁殖 2次。(周大慶)
四、其實生物現象有很多被認為不可能的事情,但是還是會發生,在國外就有一些
例子是這樣。若以 12 個巢位提出這樣的看法,還是會被懷疑,若是 120 個巢,
36
附錄一
那可能就會被相信。在研究中若懷疑森林的完整性或破碎化會影響到猛禽的繁
殖成功率,那應該是要選擇 2個條件不一樣的棲地來進行比較,社頂地區是一
個破碎化的環境,是否要再選擇像萬里得山那樣森林完整的棲地來進行比較。
如果研究假設是認為森林的完整性會有利於大冠鷲的繁殖成功率,那選擇以萬
里得山一帶來做為比較會比鵝鑾鼻半島還要來得好。(羅委員柳墀)
回應:今年的調查將會擴展到大尖石山及社頂以北 2處森林比較完整的地區,這些
地方沒有道路而且人為干擾比較低,萬里得山一帶也是預訂要調查的地區,
這個建議將會列入參考,若可能的話,也會經過修正納入計畫中。(周大慶)
五、森林很完整未必就有利於大型猛禽的繁殖,因為會影響到牠的活動靈活性,而
且食物量充足,並不等於其可用性就高。再提出問題,大冠鷲是否會使用舊巢
繁殖?如果會使用舊巢的話,那應該找巢位的速度會更快。大冠鷲築巢使用相
同的枝椏位置的機率高不高?大約有幾個%?若是今年在這個地方築巢,明年
又在這附近 1百公尺左右築巢,大約有幾%?目前有把握的繁殖配對數大約有
幾個?就目前的資料來看,是否還有需要將萬里得山的資料拿來與鵝鑾鼻半島
的比較?(羅委員柳墀)
回應:大冠鷲會使用舊巢,但是舊巢被颱風破壞的機率相當高,到目前為止 16 棵
巢樹 22 個巢,只有 3個巢是重複使用的,使用相同枝椏位置築巢的機率不
高,重複機率大約不到 1成,不同年在同一個地區築巢的機率很高,大約有
5至 6成,但是個體的辨識上有困難很難確定,有一個築巢地區 3個巢很靠
近,最近巢間距都在 5百公尺以內,也許有可能是那個區域很適合築巢,但
是各巢的活動及覓食是各自分開的,這需要更多巢的資料才能加以印證。目
前有把握在鵝鑾鼻半島上的繁殖配對數大約有 10 至 15 對,而萬里得山則是
一直想納入比較的區域,但是在今年的計畫中可能還沒有辦法做到。(周大
慶)
六、在巢位的尋找上,在歐洲因為樹木會落葉,所以巢位很好找,在台灣則都是常
綠樹木,應該可以配合猛禽的活動頻度高低去找比較容易。(羅委員柳墀)
回應:在墾丁沒辦法這樣尋找,因為大冠鷲在離巢很遠的地方就進入樹冠層在林內
潛行,所以都是一棵樹一棵樹去找巢。(周大慶)
37
七、既然是以這種方式尋求,應該可以一併記錄其他的巢,在德國就有選定 1個區
域由 1個人進行全面的調查,並記錄下所調查到的一切,相信這會是管理處經
營管理上相當有用的資料。(羅委員柳墀)
回應:其實只要發現的巢都有記錄,像:鳳頭蒼鷹、松鼠、樹鵲及黑枕藍鶲等,但
是,如:竹雞及小彎嘴晝眉等在灌木叢中的巢,就不容易發現了。(周大慶)
八、在調查中既然發現大冠鷲的巢和鳳頭蒼鷹的巢可以這麼接近,相信牠們的關係
是很密切的,是否有去加以瞭解?(羅委員柳墀)
回應:對於猛禽的種內或種間關係其實也是想探討的題目,在國外也有很多的報
告,一般而言,在種間除非有特殊的食物上的關係或競爭,否則種間是會比
種內更加靠近,就大冠鷲與鳳頭蒼鷹來看,目前並未發現有捕食上的關係,
所以種間的巢間距要比種內的巢間距小,甚至只有 70 公尺左右,就平均值
來看,大冠鷲與大冠鷲之間或鳳頭蒼鷹與鳳頭蒼鷹之間,都要比大冠鷲與鳳
頭蒼鷹之間來得遠。(周大慶)
九、其實無論種內關係或種間關係都沒有那麼簡單,在國外就有許多有趣的例子,
巢位的選擇可能與避風條件有關,也可能與食物有關,既然野外觀察都已經做
到如此程度,建議也能一併瞭解大冠鷲與鳳頭蒼鷹之間的關係,一定會很有
趣。(羅委員柳墀)
回應:鳳頭蒼鷹的巢都有一併觀察,但是沒把握在這麼短的時間內,就能夠瞭解二
者之間的關係。(周大慶)
十、再來是巢材的部份,希望在研究的時候要特別注意人類使用物品被做為巢材的
情形,尤其是塑膠繩或尼龍繩,在德國研究時曾發現蒼鷹的幼鳥在巢上練習跳
躍時被繩子跘住的情形,在這項研究中若發現巢材使用很多塑膠繩的話,就要
特別注意。另外就是巢位的選擇是否是在避風的位置?在比較寒冷的地區,發
現巢位大多與太陽有關,大多選在東南向。在量測巢材高度的時候,要注意剛
築好在孵蛋的巢與幼鳥離開後的巢,巢材的高度是不一樣的,剛築好的巢會比
較高,幼鳥在練習跳躍時巢會變得比較平,有些巢材也會掉落,這一點必須要
交待清楚。(羅委員柳墀)
回應:巢材的使用情形會加以注意,目前在社頂至鵝鑾鼻所發現的巢位都是在避風
處,至於巢材高度的量測部份會註明量測時期。(周大慶)
38
附錄一
十一、在這資料中提到墾丁的猛禽紀錄是 26 種,但在猛禽手冊簡報時提到有 27
種,到底那一個才是正確的?。(羅委員柳墀)
回應:93 年秋季發現的新紀錄種黃爪隼尚未發表,並未列入猛禽紀錄種數計算。(蔡
乙榮技士)
十二、大冠鷲的研究黃光瀛也做過,不知道有什麼不一樣?若要研究食性的話,那
是不是要蒐集食繭。(羅委員柳墀)
回應:黃光瀛的研究主要是在食性的方面,就目前的觀察大冠鷲的食物幾乎完全消
化,所以沒有辦法由蒐集食繭去瞭解食物的種類,都是要從觀察幼鳥的食物
提供去瞭解,目前已經掌握到幾處牠們經常會去進食的地方,以搭設偽裝帳
的方式可以觀察到很清楚的覓食情形,在冬季大冠鷲主要是在 2條溪流的河
谷覓食,可能是因為比較避風的關係。(周大慶)
十三、在棲地植被的部份,可以由衛星影像圖片或航照圖進行判識。在進行研究時
是否有注意到領域行為?透過一些領域宣示的行為可以標出 1對繁殖對所
利用的領域空間有多大?所選擇的是怎樣的植被環境與組成。(羅委員柳墀)
回應:本研究已使用衛星影像圖片。在觀察中確實可以看到領域宣示的行為,一般
當有其他的個體進入時,使用該棲地的繁殖對會出現尾羽上蹺的宣示行為,
進而由上方接近壓迫,或有對爪的行為。對於使用棲地的植被很想去進行詳
細研究,可是在短短的時間之內及受限於人力,實在沒有辦法同時進行那麼
多的調查分析研究。坦白講,領域是研究猛禽的人最想去瞭解的部份,但是
在這個地方有太多的因素無法確實知道,以幾個巢位靠的那麼近來看,牠們
一升空活動半徑都是幾公里以上,再加上社頂到鵝鑾鼻一帶是屬於被開墾干
擾過的環境,適合的巢樹可能只剩在溪谷而且避風的地方才有,在這個地
方,這樣的環境限制可能就是牠們選擇巢位的主要影響因素。(周大慶)
十四、單以巢位的環境恐怕無法解釋領域空間的選擇,若沒有辦法將領域標示出來
的話,很難去說明利用棲地選擇的條件,以這項計畫來看,研究經費只有
46 萬似乎少了一點。(羅委員柳墀)
回應:在國外也有採用找出最近巢間距的方式,來測量林緣長度及分析其中植被組
成等資料,在這個地方目前實在想不出有什麼因子可以用來解釋領域的問
39
題,有關領域的部份雖然很想多做一些,在受限於人力不足及時間太短的因
素下真得有很大的困難,也許明後年條件允計的話,再多做一些。(周大慶)
十五、剛剛一直在計算將要增加的野外工作就有不少,算一算可能就要增加二三倍
的工作量,但是這些記錄都很重要,也就是數據都是屬於國家的,未來都是
很有用的,以這樣的經費來看,實在有點為研究者抱屈,不過年輕人還是要
撐下去,要撐 3年。前面提到其他的研究大多只是巢邊觀察,這項研究到目
前為止看來也是屬於巢邊觀察。就繁殖研究來看,可能不單單只是巢位的選
擇及棲地環境的因子而已,如分子生物學上的領域及行為互動等也可能是很
重要的因素,另外棲地的利用及是否有共享的因素在其中,或是競爭等,這
可能又要增加工作量,不過也許第一年是準備的資料蒐集,第二年以後人力
與經費較充足,而且比較熟練及資料比較完整的時候,後期再去進行。(程
委員建中)
回應:目前的想法是競爭的因素不外是築巢的範圍及食物上的利用,當然可能還有
其他的因素,不過目前想得到的還是這 2個大的部份,如果要把競爭納入的
話,目前曾觀察到大冠鷲的巢後來是鳳頭蒼鷹在使用,原因是什麼不清楚。
(周大慶)
回應:這種現象在國外也有類似的情形,蒼鷹的巢後來是燕隼在使用,競爭的時候,
體型大的不一定就會佔上風,因為小型的反而比較靈活而且速度快,當然並
不一定是體型的因素或速度的快慢,也可能是其他因素,在猛禽之間常會有
這樣的現象發生。(羅委員柳墀)
十六、資源的競爭,可以包括空間與時間,也可以分為生物性的與非生物性的。在
研究方法中有提到採血樣,將血樣保存在含無水酒精樣本館內,然後以 PCR
分生技術鑑定性別。如果是為了要永久保存血樣在樣本館中供作日後的研究
比對,那是很好的,若不是的話,建議採用羽毛就可以運用 PCR 分生技術鑑
定性別,而不要採用侵入性的方法。(程委員建中)
回應:血樣是保存在樣本管中而不是樣本館,會採用血樣進行性別鑑定是因為配合
的李壽先老師是使用血樣進行鑑定,若羽毛也可以有效鑑定性別,將會一併
考慮。(周大慶)
十七、在統計分析方面都只是一般的基本分析,並未採用比較特別的分析方法,有
40
附錄一
些分析方法是可以用來分析棲地,像棲地型態等,這方面的分析工作或許可
以找一些人幫忙。以上是一些建議,對於前面羅委員所提出的我都贊成。不
過�