
NGHIÊN CU ĐẶC ĐI ÆM HÌNH THÁI VÀ PHÂN T ð MT S TAXON …
80
BỘ GIÁO DỤC VÀ ĐÀO TẠO VIỆN HÀN LÂM KHOA HỌC VÀ CÔNG NGHỆ VIỆT NAM HỌC VIỆN KHOA HỌC VÀ CÔNG NGHỆ ----------------------------- Học viên: Nguyễn Thị Hiên NGHIÊN CỨU ĐẶC ĐIỂM HÌNH THÁI VÀ PHÂN TỬ MỘT SỐ TAXON THUỘC CHI SÂM (PANAX L.) Ở VÙNG TÂY NGHỆ AN LUẬN VĂN THẠC SĨ SINH HỌC Hà Nội, 2020
Transcript of NGHIÊN CU ĐẶC ĐI ÆM HÌNH THÁI VÀ PHÂN T ð MT S TAXON …

BỘ GIÁO DỤC
VÀ ĐÀO TẠO
VIỆN HÀN LÂM KHOA HỌC
VÀ CÔNG NGHỆ VIỆT NAM
HỌC VIỆN KHOA HỌC VÀ CÔNG NGHỆ
-----------------------------
Học viên: Nguyễn Thị Hiên
NGHIÊN CỨU ĐẶC ĐIỂM HÌNH THÁI VÀ PHÂN TỬ
MỘT SỐ TAXON THUỘC CHI SÂM (PANAX L.)
Ở VÙNG TÂY NGHỆ AN
LUẬN VĂN THẠC SĨ SINH HỌC
Hà Nội, 2020

BỘ GIÁO DỤC
VÀ ĐÀO TẠO
VIỆN HÀN LÂM KHOA HỌC
VÀ CÔNG NGHỆ VIỆT NAM
HỌC VIỆN KHOA HỌC VÀ CÔNG NGHỆ
-----------------------------
Học viên: Nguyễn Thị Hiên
NGHIÊN CỨU ĐẶC ĐIỂM HÌNH THÁI VÀ PHÂN TỬ
MỘT SỐ TAXON THUỘC CHI SÂM (PANAX L.) Ở
VÙNG TÂY NGHỆ AN
Chuyên ngành: Thực vật
Mã số: 8 42 01 11
LUẬN VĂN THẠC SĨ: SINH HỌC
NGƢỜI HƢỚNG DẪN KHOA HỌC:
Hƣớng dẫn 1: PGS.TS Phan Kế Long
Hƣớng dẫn 2: TS. Nguyễn Thị Thanh Hƣơng
Hà Nội 2020

LỜI CAM ĐOAN
Tôi xin cam đoan các số liệu và kết quả trong luận văn này là trung
thực và chƣa đƣợc tác giả khác công bố trong bất kỳ công trình nào. Tất cả
các nguồn thông tin trích dẫn trong luận văn đã đƣợc liệt kê trong tài liệu
tham khảo. Nếu có gì sai sót tôi xin chịu hoàn toàn trách nhiệm./.
Hà Nội, ngày tháng năm 2020
Tác giả
Nguyễn Thị Hiên

LỜI CẢM ƠN
Tôi xin cảm ơn Học viện Khoa học và Công nghệ, các thầy cô giáo
Khoa Sinh thái và Tài nguyên sinh vật đã giảng dạy và giúp đỡ tôi trong suốt
quá trình học tập, nghiên cứu.
Đặc biệt, tôi xin gửi lời cảm ơn sâu sắc đến thầy hƣớng dẫn số 1 -
PGS.TS. Phan Kế Long – ngƣời trực tiếp hƣớng dẫn khoa học, cùng cô giáo
hƣớng dẫn số 2 – TS. Nguyễn Thị Thanh Hƣơng đã luôn dành nhiều thời
gian, công sức, tâm huyết và lòng nhiệt tình hƣớng dẫn tôi trong suốt quá
trình thực hiện nghiên cứu và hoàn thành luận văn.
Tôi xin trân trọng cảm ơn Quỹ học bổng Nagao tại Việt Nam, Nhiệm
vụ Điều tra cơ bản mã số UQĐTCB.01/18-19 và Đề tài TN 17-/C04, thuộc
Chƣơng trình Tây Nguyên 2016-2020 đã hỗ trợ về kinh phí giúp tôi hoàn
thiện đề tài khoa học của mình.
Tôi xin trân trọng cám ơn Ban giám hiệu trƣờng THPT Ngô Sĩ Liên –
thành phố Bắc Giang, tỉnh Bắc Giang, các thầy cô giáo trong tổ bộ môn Hóa –
Sinh cùng toàn thể các thầy cô giáo công tác trong trƣờng đã tạo điều kiện hết
mức về thời gian cũng nhƣ công việc ở trƣờng giúp tôi thoàn thành quá trình
học tập và nghiên cứu của mình.
Cuối cùng tôi xin trân trọng cảm ơn gia đình, bạn bè luôn là điểm tựa
vững chắc về vật chất, tinh thần, luôn cổ vũ động viên tôi trong suốt quá trình
học tập và nghiên cứu.
Một lần nữa tôi xin chân thành cám ơn!
Hà nội, ngày tháng năm 2020
Tác giả
Nguyễn Thị Hiên

DANH MỤC CÁC KÝ HIỆU VÀ CHỮ VIẾT TẮT
CR : Cực kì nguy cấp
DNA : Deoxyribonucleic acid
EW : Tuyệt chủng ngoài tự nhiên
ITS : Internal transcribed spacer
IUCN : International Union for Conservation of Nature and
Natural Resources
matK : Maturase K
ML : Maximum Likelihood
MP : Maximum Parsimony
NCBI : National Center for Biotechnology Information
NJ : Neighbor – Joining
PCR : Polymerase Chain Reaction
RAMS : Random amplified microsatellite
RAPD : Randomly Amplified Polymosphic DNA
rbcL : Ribulose bisphosphate carboxylase large chain
RFLP : Restriction Fragment Length Polymorphism
SCN : Sau công nguyên
SSR : Simple Sequence Repeats
TCN : Trƣớc công nguyên
UPGMA : Unweighted pair group method with arithmetic mean

DANH MỤC CÁC BẢNG
Bảng 2.1. Trình tự cặp mồi nhân bản vùng ITS-rDNA bằng kỹ thuật PCR với
cặp mồi theo Phan Kế Long và cs. (2014) ...................................... 16
Bảng 2.2. Các thành phần phản ứng PCR ....................................................... 20
Bảng 2.3. Danh sách trình tự sử dụng trong nghiên cứu thu thập trên Genbank
......................................................................................................... 23
Bảng 3.1. So sánh đặc điểm hình thái giữa Tam thất hoang (P. stipuleanatus)
và Sâm vũ diệp (P. bipinnatifidus) (Nguyễn Tập, 2007) ................ 38
Bảng 3.2. Thành phần nucleotide (%) của các mẫu nghiên cứu ..................... 46
Bảng 3.3. Khoảng cách di truyền (%) giữa các mẫu nghiên cứu (bên trên) và
số lƣợng nucleotide sai khác (bên dƣới) giữa các mẫu nghiên cứu 50

DANH MỤC CÁC ẢNH, HÌNH VẼ
Hình 2.1. Sơ đồ các bƣớc thí nghiệm .............................................................. 17
Hình 2.2. Máy đo NanoDrop one .................................................................... 19
Hình 2.3. Chu trình phản ứng PCR ................................................................. 20
Hình 2.4. Sơ đồ phản ứng QIAquick ............................................................. 21
Hình 3.1. Panax notoginseng - Tam thất (lá mẫu TH1). ................................ 26
Hình 3.2. Panax notoginseng - Tam thất (củ mẫu TH1). .............................. 27
Hình 3.3. Mẫu Panax thu ở khu vực Phu Xai Lai Leng. ............................... 29
Hình 3.4. Panax stipuleanatus - Tam thất hoang (mẫu TB1: lá chét nguyên). 31
Hình 3.5. Panax stipuleanatus - Tam thất hoang (mẫu TB2: lá chét xẻ thùy
sâu). ................................................................................................. 31
Hình 3.6. Panax stipuleanatus - Tam thất hoang (mẫu TB3: lá chét xẻ thùy
sâu) .................................................................................................. 32
Hình 3.7. Panax stipuleanatus - Tam thất hoang (mẫu TB4: lá chét xẻ thùy
nông) ............................................................................................... 33
Hình 3.8. Panax stipuleanatus -Tam thất hoang (mẫu lá ML1) ..................... 34
Hình 3.9. Panax stipuleanatus-Tam thất hoang (mẫu củ ML1) ..................... 35
Hình 3.10. Mẫu (K.M.Feng 13694) isotype (P. stipuleanatus) hiện đang lƣu
trữ tại phòng tiêu bản Viện Thực vật Bắc Kinh (PE0025805) ....... 36
Hình 3.11. So sánh trình tự vùng gen ITS-rDNA của các mẫu Panax TB1-
TB4 và ML1 với trình tự vùng gen của loài P. stipuleanatus ........ 37
Hình 3.12. Lá loài Panax zingiberensis -Tam thất gừng (mẫu lá ML2).. ...... 40
Hình 3.13. Củ của loài Panax zingiberensis -Tam thất gừng (mẫu củ ML2)..... 41
Hình 3.14. So sánh trình tự vùng gen ITS-rDNA của mẫu ML2 với các loài
trong cùng nhóm: P. zingiberensis và P. Wangianus. .................... 43
Hình 3.15. Sắc ký đồ trình tự vùng gen ITS-rDNA của mẫu Panax TB1 ...... 46
Hình 3.16. Trình tự vùng gen ITS-rDNA của các mẫu nghiên cứu................ 49

Hình 3.17. Mối quan hệ họ hàng của các mẫu nghiên cứu với các loài/thứ
trong chi Panax L. trên cơ sở phân tích trình tự vùng gen ITS-
rDNA theo phƣơng pháp Maximum Likelihood (ML). Mô hình tối
ƣu GTR+G+I. Eleutherococcus senticosus là loài ngoài nhóm. Giá
trị ở gốc là giá trị bootstrap >50% .................................................. 51

MỤC LỤC
LỜI CAM ĐOAN
LỜI CẢM ƠN
MỞ ĐẦU .......................................................................................................... 1
CHƢƠNG 1. TỔNG QUAN TÀI LIỆU ........................................................ 4
1.1. GIỚI THIỆU VỀ CHI SÂM PANAX L. ................................................. 4
1.2. TỔNG QUAN CÁC NGHIÊN CỨU VỀ CHI PANAX L. TRÊN THẾ
GIỚI ................................................................................................................ 4
1.3. TỔNG QUAN CÁC NGHIÊN CỨU VỀ CHI PANAX L Ở VIỆT NAM
........................................................................................................................ 8
CHƢƠNG 2. VẬT LIỆU VÀ PHƢƠNG PHÁP NGHIÊN CỨU ............. 14
2.1. VẬT LIỆU, TRANG THIẾT BỊ VÀ DỤNG CỤ NGHIÊN CỨU ....... 14
2.1.1. Vật liệu ............................................................................................ 14
2.1.2. Địa điểm nghiên cứu ....................................................................... 14
2.1.3. Các hóa chất dùng trong nghiên cứu ............................................. 14
2.1.4. Thi t ị và dụng cụ nghiên cứu ....................................................... 15
2.2. PHƢƠNG PHÁP NGHIÊN CỨU ........................................................ 17
2.2.1. Sơ đồ nghiên cứu ............................................................................ 17
2.2.2. Phương pháp thu thập mẫu............................................................. 17
2.2.3. Phương pháp định loại hình thái .................................................... 18
2.2.4. Phương pháp nghiên cứu sinh học phân tử .................................... 18
2.2.4.1. Phƣơng pháp tách chiết DNA tổng số ...................................... 18
2.2.4.2. Phƣơng pháp PCR (Polymerase Chain Reaction) .................... 19
2.2.4.3. Tinh sạch sản phẩm PCR .......................................................... 21
2.2.4.4. Phƣơng pháp phân tích số liệu .................................................. 22

CHƢƠNG 3. KẾT QUẢ VÀ THẢO LUẬN ............................................... 25
3.1. KẾT QUẢ NGHIÊN CỨU ................................................................... 25
3.1.1. Đặc điểm nhận bi t chi Panax ........................................................ 25
3.1.2. Panax notoginseng (Burkill) F.H.Chen ex C.H. Chow - Tam thất
................................................................................................................... 25
3.1.3. Panax stipuleanatus Tsai et Feng- Tam thất hoang ....................... 28
3.1.4. Panax zingiberensis C.Y. Wu & K.M. Feng-Tam thất gừng .......... 39
3.1.5. Đặc điểm hình thái chung của chi Panax ở vùng Tây Nghệ An ..... 43
3.1.6. Mối quan hệ họ hàng của các mẫu nghiên cứu .............................. 45
3.2. THẢO LUẬN ....................................................................................... 52
KẾT LUẬN VÀ KIẾN NGHỊ ...................................................................... 54
1. KẾT LUẬN ................................................................................................. 54
2. KIẾN NGHỊ ................................................................................................ 54
TÀI LIỆU THAM KHẢO ............................................................................ 55
PHỤ LỤC

1
MỞ ĐẦU
1. LÝ DO CHỌN ĐỀ TÀI
Việt Nam nằm trong vùng khí hậu nhiệt đới gió mùa nóng và ẩm nên
có nguồn tài nguyên thực vật phong phú và đa dạng. Thêm vào đó, cộng đồng
các dân tộc Việt Nam từ lâu đời cũng đã có nhiều kinh nghiệm sử dụng các
loại cây cỏ làm thuốc trong đó có Sâm.
Nhân sâm đƣợc dùng trong y học phƣơng Đông hàng ngàn năm nay.
Ngƣời Trung Quốc đã sử dụng nhân sâm từ rất lâu đời nhƣ một thứ dƣợc liệu vô
cùng trân quý, chỉ có các vua chúa và các quan lớn mới có loại thảo dƣợc này.
Tác dụng của nhân sâm đƣợc đề cập trong các trƣớc tác của Shi You (khoảng
những năm 48-33 TCN) hay Shanghan Lun (khoảng những năm 200 SCN).
Chi Nhân sâm (Panax L.) là một chi nhỏ trong họ Ngũ gia bì
(Araliaceae) gồm 15 loài, tất cả đều có giá trị làm thuốc, một số loài thuộc chi
Panax L. (Araliaceae) đƣợc sử dụng có giá trị cao nhƣ Nhân sâm (P.
ginseng), Tam thất (P. notoginseng). Ở Việt Nam, có một số loài thuộc chi
này nhƣ Sâm vũ diệp (P. bipinnatifidus), Tam thất hoang (P. stipuleanatus)
và Sâm ngọc linh (P. vietnamensis) [1,2,3].
Do có giá trị sử dụng và giá trị kinh tế cao, các loài nhân sâm này đã
bị tìm kiếm, khai thác đến mức kiệt quệ trong tự nhiên. Sâm vũ diệp và Tam
thất hoang đã đƣợc xếp hạng cực kỳ nguy cấp (CR), còn Sâm ngọc linh ngày
nay đã đƣợc coi là tuyệt chủng ngoài tự nhiên (EW). Cả ba loài này đã đƣợc
đƣa vào Sách Đỏ Việt Nam (2007), Phụ lục IA - các loài nghiêm cấm khai
thác và sử dụng vì mục đích thƣơng mại của Nghị định 06/2019/NĐ-CP
ngày 22/01/2019, Nghị định 32/2006/NĐ-CP ngày 30/03/2006 về quản lý
thực vật rừng, động vật rừng nguy cấp, quý hiếm. Và đây cũng là ba trong số
17 loài thực vật thuộc Danh mục loài nguy cấp qúy hiếm đƣợc ƣu tiên bảo
vệ theo Nghị định số 160/2013/NĐ-CP ngày 12/11/2013 của Thủ tƣớng
Chính phủ [4].
Vùng Tây Nghệ An có diện tích rất lớn với 1,3 triệu ha với 09 đỉnh núi
thuộc Bắc Trƣờng Sơn cao trên 1000m so với mặt biển nhƣ Pù Hon 1447m,

2
Pù Mát 1814m, Phu Xai Lai Leng 2711m, Pù Hút 2452m, Pù Lon 1561m, Pu
Đen Đin 1540m, Rào Cỏ 2286m, Pù Huống 1200m và Chốp Cháp 1705m có
khả năng phù hợp với điều kiện sinh trƣởng và phát triển của các loài cây
thuốc thuộc chi Sâm (Panax L.). Trên cơ sở nghiên cứu bƣớc đầu và thông tin
từ ngƣời dân địa phƣơng cho biết đã gặp một số cây sâm ở các khe núi Ca
Nọi, Na Khang và Phú Khả thuộc khu vực núi Phu Xai Lai Leng có độ cao
2711m thuộc xã Na Ngoi, huyện Kỳ Sơn, tỉnh Nghệ An nên nhóm nghiên
cứu đã lựa chọn điều tra đại diện các loài cây thuộc chi Sâm (Panax L.) cho
vùng Tây Nghệ An tại khu vực đỉnh núi Phu Xai Lai Leng.
Thêm vào đó, hiện nay Công ty TNHH TH true MILK đã đầu tƣ vƣờn
dƣợc liệu tại các xã Na Ngoi và Mƣờng Lống, Kỳ Sơn, Nghệ An trong đó có
khoảng hơn 1000m2 trồng Sâm. Theo thông tin ghi nhận đƣợc, các loài Sâm
trồng ở đây đƣợc thu thập từ khu vực Tây Nghệ An và nhập từ địa phƣơng
khác về trong đó có cả Sâm ngọc linh đƣợc mang về từ Nam Trà My vì vậy
việc khảo sát, định loại các loài thuộc chi Sâm tại đây có thể mang lại thêm
thông tin về đa dạng loài Sâm tại vùng Tây Nghệ An. Hơn nữa, các loài Sâm
đều sinh trƣởng tốt tại vƣờn ƣơm này, vì vậy đây có thể là mô hình bảo tồn
chuyển vị cho các loài thuộc chi Sâm ở Tây Nghệ An
Nhằm cung cấp những dẫn liệu và cơ sở khoa học phục vụ công tác bảo
tồn, phát triển cũng nhƣ công tác định danh, phân biệt một số loài Sâm ở Tây
Nghệ An, luận văn đã lựa chọn đề tài “Nghiên cứu đặc điểm hình thái và
phân tử một số taxon thuộc chi Sâm (Panax L.) ở vùng Tây Nghệ An”.
2. MỤC TIÊU ĐỀ TÀI
- Mô tả đặc điểm hình thái và sinh học phân tử các loài thuộc chi Sâm
(Panax L.) thu thập ở khu vực Phu Xai Lai Leng, Kỳ Sơn, Nghệ An
- Nghiên cứu một số đặc điểm hình thái và sinh học phân tử của một số
loài thuộc chi Sâm (Panax L.) đƣợc trồng tại Công ty TNHH TH true MILK
Na Ngoi và Mƣờng Lống, Kỳ Sơn, Nghệ An.
3. ĐỐI TƢỢNG VÀ PHẠM VI NGHIÊN CỨU
- Đối tƣợng nghiên cứu: Các loài cây thuộc chi Sâm (Panax L.) thu

3
thập ở vùng Tây Nghệ An gồm các mẫu thu ở khu vực Phu Xai Lai Leng và
vƣờn giống của Công ty TNHH Thtrue MILK ở xã Na Ngoi và Mƣờng Lống,
Kỳ Sơn, Nghệ An.
- Phạm vi nghiên cứu: Vùng Tây Nghệ An
4. Ý NGHĨA KHOA HỌC VÀ THỰC TIỄN CỦA ĐỀ TÀI NGHIÊN CỨU
* Ý nghĩa khoa học
- Góp phần bổ sung hoàn chỉnh vốn kiến thức về hình thái và phân loại
chi Sâm (Panax L.) ở Việt Nam
* Ý nghĩa thực tiễn
- Kết quả của nghiên cứu là cơ sở khoa học để nhận biết bảo tồn các
loài thuộc chi Sâm (Panax L.) tại vùng Tây Nghệ An.
- Thông tin về thành phần loài, phân bố và hiện trạng các loài thuộc chi
Sâm (Panax L.) ở vùng Tây Nghệ An có ý nghĩa lớn đối với việc phát triển
kinh tế vùng, mang lại lợi ích cho ngƣời dân địa phƣơng.
- Kết quả của đề tài là cơ sở khoa học phục vụ cho các ngành ứng dụng
và sản xuất nhƣ Nông - Lâm nghiệp, Dƣợc học, Tài nguyên thực vật, Đa dạng
sinh học và trong công tác đào tạo.
5. ĐIỂM MỚI CỦA LUẬN VĂN
- Mô tả chi tiết đặc điểm hình thái 3 loài thuộc chi Panax L. thu đƣợc ở
vùng Tây Nghệ An
- Sử dụng phƣơng pháp sinh học phân tử hỗ trợ định danh chuẩn các
loài thuộc chi Panax thu đƣợc ở vùng Tây Nghệ An.
- Đã phân tích, xác định Tam thất hoang và Sâm vũ diệp ở Việt Nam là
cùng một loài P. stipuleanatus H.T. Tsai et K.M. Feng.
- Xây dựng bộ tƣ liệu ảnh minh họa đặc điểm hình thái, sinh học phân
tử của các taxon thuộc chi Panax ở vùng Tây Nghệ An: 14 ảnh chụp từ các
mẫu thu trực tiếp trên thực địa, ảnh tham khảo và ảnh chụp từ mẫu trong
phòng tiêu bản.

4
CHƢƠNG 1. TỔNG QUAN TÀI LIỆU
1.1. GIỚI THIỆU VỀ CHI SÂM PANAX L.
Chi Nhân sâm (Panax L.) là một chi nhỏ trong họ Ngũ gia bì
(Araliaceae). Chi này trên thế giới có 8 loài phân bố ở vùng Đông Á,
Hymalaia, Inđôchina, Bắc Châu Mỹ [5]. Sự phân bố của chi Panax L. trên thế
giới cho thấy chúng chỉ xuất hiện ở Bắc bán cầu, kéo dài từ vùng rừng núi
giáp bờ biển phía Đông của Bắc Mỹ bao gồm Bắc Hoa Kỳ và Tây-Nam
Canada (có 2 loài P. quinquefolius và P. trifoliatus) [6, 7, 8]. Vùng Đông Bắc
Á (gồm Viễn Đông Nga, Đông Bắc Trung Quốc, bán đảo Triều Tiên và Nhật
Bản) có 2 loài là P. ginseng và P. japonica. Trung tâm phân bố của chi Panax
L. có thể từ vùng Tây - Nam của Trung Quốc xuống phía Bắc Việt Nam. Thực
chất khu vực này gồm 2 tỉnh biên giới kề nhau là Vân Nam (Trung Quốc) và Lào
Cai (Việt Nam), ở đây đang có tới 7 loài và thứ mọc hoàn toàn tự nhiên, 2 loài
trồng là P. notoginseng (nhập từ Bắc Mỹ) và P. pseudoginseng. Đây có thể coi là
trung tâm phân bố của chi Sâm (Panax L.) trên thế giới. Ở Bắc Mỹ hiện có 3 loài
(P. notoginseng; P. quinquefolius và P. trifoliatus). Giới hạn cuối cùng về phía
Nam của chi Panax L. là loài Sâm Việt Nam (P. vietnamensis) ở Miền Trung
của Việt Nam, tại 14015
’ vĩ độ Bắc.
1.2. TỔNG QUAN CÁC NGHIÊN CỨU VỀ CHI PANAX L. TRÊN THẾ GIỚI
Chi Panax L. đƣợc Carl Linnaeus mô tả lần đầu tiên từ năm 1753 với
loài chuẩn là P. quinquefolius phân bố ở Bắc Mỹ [8]. Sang thế kỷ 19, một số
loài khác đƣợc công bố nhƣ Giả nhân sâm - P. pseudoginseng Wall, ở Nepal;
Nhân sâm - P. ginseng Meyer ở Bắc Triều Tiên, Đông Bắc Trung Quốc và
Viễn Đông Nga; Sâm Nhật P. japonicus Meyer ở Nhật Bản và Trung Quốc;
Sâm vũ diệp - P. bipinnatifidus Seem, ở Đông Bắc Ắn Độ, Nepal và Trung
Quốc (Wen J., 2000) [9]. Nhƣ vậy, đến cuối thế kỷ 19, chi Panax L. có 6 loài.
Năm 1975, Trung Quốc đã bổ sung thêm một loài mới là P. stipuleanatus H.
T. Tsai & K. M. Feng, đồng thời nâng cấp các taxon thứ của P.
pseudoginseng Wall, của các tác giả trƣớc đó thành bậc loài hoàn chỉnh
(Nocerino E. et al., 2000) [10]. Năm 1985[11], một loài mới khác đƣợc tìm

5
thấy ở Việt Nam là Sâm ngọc linh - P. vietnamensis Ha et Grushv. đã nâng
tổng số loài thuộc chi Panax trên thế giới lên 12 loài và 1 thứ. Trong đó Bắc
Mỹ có 2 loài (P. quinquefolius và P. trifolius). Các loài còn lại phân bố ở
châu Á, bao gồm 6 loài ở Trung Quốc, 3 loài có ở Nhật Bản và có 5 loài có ở
Việt Nam (kể cả 2 loài cây trồng nhập nội).
Theo Linnaeus (1753) [8] điểm khác biệt của chi Panax L. so với các
chi khác trong họ Araliaceae là bầu 2 ô, hoa mẫu 5, xếp van hay xếp lợp.
Trong giai đoạn này, số lƣợng loài đã biết còn ít, nên tác giả chỉ giới hạn ở
các đặc điểm của hoa: hoa vừa xếp lợp vừa xếp van. Đây là đặc điểm chính
của chi để phân biệt với các chi khác trong họ Araliaceae. Tuy nhiên, với cách
sắp xếp này thì đặc điểm của chi Panax bao gồm một số chi khác thuộc họ
Araliaceae, đặc biệt là chi Nothopanax.
Theo quan điểm trên, De Candolle (1830) [12] đã chuyển một số loài
thuộc chi Nothopanax vào chi Panax. Kể từ đây, vị trí phân loại và mối quan
hệ phát sinh của chi Panax không có sự đồng nhất về quan điểm giữa các tác
giả. Decaisne và Planchon (1854) [13] cho rằng đặc điểm chính của chi
Panax là đài xếp van, nên chuyển 2 loài P. quinquefolius và P. trifolius sang
chi Aralia, đồng thời hợp nhất hoá các chi Polyscias, Clerodendron,
Pseudopanax và Maralia vào chi Panax. Nhƣ vậy, với cách sắp xếp nhƣ trên,
các tác giả đã cho rằng đặc điểm hình thái của chi Panax bên cạnh đài xếp
van còn có đặc điểm thân gỗ và thân thảo. Đồng ý với quan điểm trên, các tác
giả Bentham và Hooker (1867) [14], Clarke (1879) [15] tiếp tục sát nhập chi
Nothopanax vào chi Panax, đồng thời chuyển chi Panax vào tông (tribe)
Panaceae. Đến thời điểm này, số lƣợng loài thuộc chi Panax lên đến 25 loài,
phân bố chủ yếu ở vùng nhiệt đới châu Á, châu Phi, Newzeland và Australia.
Năm 1868, Seemann [16] đã xem xét lại vị trí phân loại chi Panax và
cho rằng những mô tả của Linnaeus về đặc điểm hình thái đặc trƣng của chi
Panax là không rõ ràng. Theo tác giả, đặc điểm chính để phân biệt chi Panax
so với các chi khác trong họ Araliaceae là đài 5, xếp lợp, bầu 2 ô. Với đặc
điểm này thì 2 loài P. quinquefolius và P. trifolius thuộc về chi Panax và có
mối quan hệ gần với chi Aralia. Đối chiếu với đặc điểm của các chi khác

6
thuộc họ Araliaceae, tác giả ghi nhận có một đặc điểm chỉ ra sự khác biệt giữa
chi Panax so với chi khác là thân dạng củ. Qua đó, tác giả chuyển các loài có
thân dạng củ trƣớc đây ở các chi khác vào chi Panax, đồng thời chuyển một
số loài không có thân dạng củ ra khỏi chi Panax. Nhƣ vậy, chi Panax có 3 đặc
điểm quan trọng làm cơ sở để phân biệt với các chi khác trong họ Araliaceae
là: thân dạng củ, đài 5, xếp lợp, bầu 2 ô. Với quan điểm trên, tác giả đã
chuyển 61 loài ra khỏi chi Panax, và chi này chỉ còn lại các loài: P. trifolius
L., P. quinquefolius L., P. ginseng Mey., P. pseudoginseng Wall., P.
japonicus Mey. và P. bipinnatifidus Seem. Với loài P. fructicosus có đặc
điểm là đài xếp van, Seemann xếp vào chi Nothopanax và đồng thời lấy loài
này là type cho chi Nothopanax. Có lẽ đây là quan điểm đƣợc nhiều nhà
nghiên cứu về chi Panax cho là phù hợp hơn cả. Để khẳng định lại quan điểm
của Seeman (1868) [16], Harm (1897) [17] đã nghiên cứu tất cả các đặc điểm
của các loài Sâm phân bố trên thể giới nhƣ: cơ quan sinh sản, cơ quan sinh
dƣỡng và cho rằng chi Panax có nguồn gốc từ chi Aralia, điểm khác biệt ở
đây là thân dạng củ. Britton và Brown (1913) [18] tiếp tục xem xét vị trí phân
loại chi Panax và đồng tình với quan điểm của Seeman (1868) và Harm
(1897) [17], chỉ ra loài P. quinquefolius có đặc điểm đại diện cho chi: thân
dạng củ, hoa mẫu 5 xếp lợp, bầu 2 ô, và loài đƣợc mô tả lần đầu tiên, nên lấy
loài này làm type danh pháp cho chi Panax.
Hoo (1961) [19] khi nghiên cứu về hệ thống học, mối quan hệ phát sinh
và phân bố của taxon trong họ Araliaceae đã cho rằng chi Panax có nguồn
gốc phát sinh từ Acanthopanax. Vì chi này có đặc điểm hoa mẫu 5, lá có lông
cứng mọc trên gân lá, thân thảo hay thân dạng cây bụi nhỏ. Tác giả cho rằng
đặc điểm thân thảo là đặc điểm tƣơng đồng quan trọng nhất, các loài trong chi
Aralia hầu nhƣ không có đặc điểm này. Quan điểm này của Hoo hầu nhƣ
không nhận đƣợc sự đồng tình của các nhà nghiên cứu về chi Panax.
Năm 1970, Hara [20] xem xét đặc điểm của chi Panax dựa trên mẫu vật
của các loài thuộc chi Panax ở châu Á thu đƣợc ngoài tự nhiên cũng nhƣ dựa
trên mẫu lƣu giữ ở các Bảo tàng thực vật. Qua đó, tác giả thống nhất với các
quan điểm nghiên cứu trƣớc đó về đặc điểm của chi Panax: thân dạng củ, hoa
mẫu 5 xếp van hay xếp lợp, bầu 2 ô và có mối quan hệ gần với chi Aralia.

7
Tuy nhiên, theo tác giả các loài trong chi Panax có đặc điểm hình thái biến
đổi rất lớn so với các bậc phân loại khác trong họ Araliaceae. Do đó, việc
định loại các loài trong chi Panax là việc rất khó khăn nếu chỉ dựa vào một
đặc điểm hình thái nào đó. Theo tác giả, để phân biệt các loài thuộc chi
Panax, trƣớc tiên xếp loài vào 1 trong 3 chuẩn hình thái của thân rễ (rhizome
type): thân rễ có dạng thân củ (carot type); thân rễ có đốt kéo dài; và thân rễ
có đốt ngắn nhƣ đốt trúc. Tiếp theo đó là các đặc điểm hình thái lá, đài, tràng,
số lƣợng chỉ nhụy, quả... Cách phân loại dựa vào hình thái thân rễ trƣớc rồi mới
đến các đặc điểm cơ quan sinh dƣỡng và sinh sản đã đƣợc Xiang và Lowry
(2007) [5] áp dụng để xây dựng khóa phân loại chi Panax ở Trung Quốc.
Wen và Zimmer (1996) [21] dựa vào đặc điểm hình thái, hình thái giải
phẫu, hạt phấn và sinh học phân tử cho rằng chi Panax có mối quan hệ gần
gũi với chi Aralia. Đến thời điểm hiện nay, các hệ thống quan điểm dựa vào
hình thái, giải phẫu và sinh học phân tử đều thống nhất và cho rằng chi Panax
có mối quan hệ gần với chi Aralia, điểm khác khác biệt chủ yếu dễ dàng nhận
thấy so với chi Aralia là thân dạng củ, lá kép hình chân vịt, noãn 2-3.
Sau năm 1970, vị trí phân loại và hệ thống phát sinh của chi Panax hầu
nhƣ không thay đổi. Tuy nhiên do đặc điểm hình thái có biến đổi rất lớn, ngay
trong một loài ở các vùng phân bố khác nhau thì có đặc điểm hình thái cũng
khác nhau (Hara,1970) [20], đồng thời ngày càng có nhiều phát hiện mới nên từ
đó đến nay, việc ghi nhận số lƣợng loài trong chi Panax có sự thay đổi theo thời
gian và tùy vào quan điểm của các nhà nghiên cứu. Ở nửa đầu của thập niên
1970, quan niệm về thành phần loài Sâm châu Á của các nhà nghiên cứu cũng
rất khác nhau và có sự bổ sung nhiều taxon mới. Gần đây, quan hệ phát sinh
giữa các taxon thuộc chi Panax đƣợc xây dựng không chỉ dựa vào đặc điểm hình
thái mà còn sử dụng trình tự các DNA mã vạch - hoặc gọi là chỉ thị DNA), tuy
vậy vẫn chƣa có sự thống nhất giữa các nhóm tác giả. Có thể nhận thấy sự khác
nhau giữa các quan điểm phân loại về số lƣợng loài, sự sắp xếp và phân chia các
loài và dƣới loài trong chi Panax qua các thời kỳ có thể do các nguyên nhân sau:
tính đa dạng về hình thái trong chi Panax, quan điểm về hệ thống phát sinh dựa
vào đặc điểm hình thái rất đa dạng; việc sử dụng các DNA barcode khác nhau để
hỗ trợ cho nghiên cứu phân loại; các phát hiện mới về taxon.

8
Các taxon thuộc chi Panax đã biết đều là các đối tƣợng có giá trị sử
dụng cao và đƣợc xem là “thần dƣợc cuộc sống” (Florence, 1992) [22], do
vậy đã có hàng loạt các công trình nghiên cứu trên thế giới và cũng nhƣ ở
Việt Nam đƣợc tiến hành nhằm thúc đẩy sản xuất, khai thác, sử dụng hiệu
quả. Nhiều nhất là các nghiên cứu về thành phần hóa dƣợc và dƣợc lý, tiếp
đến là nghiên cứu về nhân giống, canh tác, chế biến, hệ thống học thực vật, đa
dạng di truyền, bảo tồn đa dạng nguồn gen... Do đó có khá nhiều công trình
nghiên cứu đƣợc công bố trên các tạp chí, sách chuyên khảo và các thông tin
trên các phƣơng tiện đại chúng liên quan đến Sâm. Nói cách khác, các loài
Sâm là đối tƣợng thực vật có một sự cuốn hút lớn đối với những ngƣời làm
công tác nghiên cứu trong nhiều lĩnh vực trên toàn thế giới. Những công trình
nghiên cứu về sâm vẫn đang đƣợc triển khai cho dù những phát hiện đột phá
về mặt thành phần loài gần đây không nhiều.
1.3. TỔNG QUAN CÁC NGHIÊN CỨU VỀ CHI PANAX L Ở VIỆT NAM
Mặc dù là những loài cây thuốc có giá trị nhƣng các công trình nghiên
cứu về thực vật ở Việt Nam và Đông Dƣơng từ thời Pháp thuộc vẫn chƣa ghi
nhận phân bố của các loài thuộc chi Panax ở Việt Nam. Đến năm 1964, Viện
Dƣợc liệu đã thu đƣợc một số mẫu của loài cây thuốc có tên là “Phan xiết” do
đồng bào ngƣời Dao và Mông ở Sa Pa giới thiệu. Tháng 7 năm 1964 nhà thực
vật học ngƣời Trung Quốc Ngô Chính Dật đã giám định các tiêu bản thuộc
chi Panax L. lƣu giữ tại Viện Dƣợc liệu, trong đó mẫu số 911 và 348 thu ở xã
Tả Phin huyện Sa Pa tỉnh Lào Cai ngày 14/6/1964 đƣợc xác định có tên khoa
học là P. bipinnatifidus Seem. Đến năm 1969, loài này mới đƣợc Grushvitzky
và cộng sự [23] công bố trong một loạt các công trình nghiên cứu về họ
Araliaceae ở miền Bắc Việt Nam [24,25,26]. Từ đó, Sâm vũ diệp đã đƣợc ghi
nhận trong tất cả các tài liệu về cây thuốc và hệ thực vật ở Việt Nam.
Viguier (1908 – 1923) là công trình nghiên cứu đầu tiên ở Việt Nam
chỉ mô tả đặc điểm và phân bố của chi Panax. Nhƣng không trích dẫn tài liệu
tham khảo, dẫn đến gây khó khăn cho ngƣời nghiên cứu tiếp theo và chƣa xác
định đƣợc các loài có phân bố ở Việt Nam. [27]
Phạm Hoàng Hộ (1970) [28] là ngƣời đầu tiên ghi nhận thứ Sâm nhật

9
(P. schinseng Nees var. japonicum Mak.) có ở rừng dày ẩm, núi Langbian
(Lâm Đồng) trong bộ “Cây cỏ Việt Nam”. Ông mô tả ngắn gọn về đặc điểm
hình thái, sinh thái, phân bố và hình vẽ của thứ này.
Năm 1973, Võ Văn Chi & cs trong cuốn “Cây cỏ thƣờng thấy ở Việt
Nam” mô tả Panax pseudoginseng Nees – Tam thất kim bất [29].
Năm 1993, Phạm Hoàng Hộ [30] đã mô tả 3 loài thuộc chi Panax L. có
ở Việt Nam là Sâm hai lần chẻ - P. bipinnatifidus Seem, có ở Hoàng Liên
Sơn, Sâm nhật - P. japonica (Nees) Meyer có ở Langbian, Kon Tum và Giả
nhân sâm - P. pseudoginseng Wall, có ở Sa Pa. Trong lần tái bản vào năm
2000, ngoài hai loài Sâm hai lần chẻ và Giả nhân sâm, Phạm Hoàng Hộ còn
bổ sung thêm một loài Sâm ngọc linh - P. vietnamensis Ha et Grushv. có ở
Gia Lai - Kon Tum. Còn loài Sâm nhật ông ghi có ở Langbian và cả ở Kon
Tum. Nhƣ vậy theo Phạm Hoàng Hộ, trên địa bản tỉnh Kon Tum hoặc Gia Lai
- Kon Tum cũ, có cả hai loài Sâm nhật và Sâm việt nam. Còn đối với Sâm
nhật ở Kon Tum, ông không ghi rõ nguồn mẫu xác định và nơi phân bố cụ
thể, điều này gây trở ngại cho việc các định loài Sâm nhật kể trên có thực sự
phân bố tự nhiên ở Việt Nam hay không. Trong khi đó, loài Sâm nhật do
Phạm Hoàng Hộ đã từng ghi nhận ở Langbian cũng chính là Sâm việt nam
hay Sâm ngọc linh đƣợc Hà Thị Dụng và Grushvitzky [31] (1996) công bố là
loài mới cho khoa học và đặt tên là P. vietnamensis Ha et Grushv.
Năm 1993, Trần Đình Lý trong cuốn “1900 loài cây có ích ở Việt
Nam”, mô tả rất ngắn gọn về phân bố và tác dụng của P. vietnamensis Ha et
Grushv., P. bipinnatifidus Seem. và P. Pseudoginseng Wall. [32].
Năm 1995, Đỗ Tất Lợi trong cuốn “Những cây thuốc và vị thuốc Việt
nam” đã mô tả rất chi tiết về lịch sử ra đời, hình thái, thành phần hóa học, tác
dụng dƣợc lý, công dụng và liều dùng của hai loài là P. vietnamensis Ha et
Grushv và P. pseudoginseng Wall.[33]
Năm 1997, Võ Văn Chi [34] trong cuốn “ Từ điển cây thuốc Việt Nam”
mô tả về P. pseudoginseng Wall. Loài của Nam Trung quốc và Bắc Việt
Nam. Ở nƣớc ta cây mọc hoang và đƣợc trồng nhiều ở Hà Giang, Lào Cai,

10
Cao Bằng tại các vùng núi cao lạnh 1200 – 1500 m; Tam thất hoang P.
bipinnatifidus Seem – Vũ diệp tam thất. Loài của Nam Trung Quốc và Bắc
Việt Nam, ở độ cao 1900 – 2400 m trong rừng ẩm, cây mọc hoang ở vùng núi
cao lạnh của tỉnh Lào Cai. Cũng đƣợc trồng và mọc tốt nhƣ Tam thất; Sâm
ngọc linh P. vietnamensis Ha et Grushv là loài đặc hữu của Việt Nam mọc tập
trung trong 13 xã thuộc 3 huyện miền núi Ngọc linh, thuộc hai tỉnh Kon tum
và Quảng Nam. Cả ba loài này tác giả đều mô tả đầy đủ về hình thái, bộ phận
sử dụng, nơi sống và thu hái, thành phần hóa học cũng nhƣ công dụng.
Năm 1997, Lê Trần Đức trong cuốn “Cây thuốc Việt Nam trồng hái
chế biến trị bệnh ban đầu” đã mô tả rất chi tiết về đặc điểm hình thái, điều
kiện sống, kĩ thuật trồng và chăm sóc, trị sâu bệnh, cách thu hái và chế
biến, công dụng của P. pseudoginseng Wall. Phần cuối mô tả tác giả có
ghi phụ chú cho biết P. vietnamensis Ha et Grushv và củ Tam thất mọc
hoang – P. bipinnatifidus Seem cũng đƣợc dùng với hiệu quả gần nhƣ P.
pseudoginseng Wall. [35]
Năm 2000, Phạm Hoàng Hộ [36] trong cuốn “Cây cỏ Việt Nam” đã
chỉnh sửa thành P. bipinnatifidus Seem. Sâm hai lần kép đƣợc tìm thấy ở núi
Hoàng Liên Sơn. Điểm khác biệt là trong quyển 2 xuất bản năm 2000 Phạm
Hoàng Hộ bổ sung thêm đặc điểm hình thái, sinh thái, phân bố và vẽ hình của
loài P. vietnamensis Dung & Grushv. Sâm việt tìm thấy ở Gia Lai, Kon Tum.
Nhìn chung các công trình của Phạm Hoàng Hộ còn mang tính chất thông kê
đặc điểm hình thái, sinh thái, phân bố và vẽ hình chƣa mô tả đặc điểm chi,
chƣa chỉ ra mẫu nghiên cứu và mẫu chuẩn cũng nhƣ bảo tàng lƣu giữ mẫu.
Tuy nhiên đây đƣợc coi là tài liệu phân loại thực vật về chi Panax ở Việt Nam
có giá trị nhất tới thời điểm này.
Năm 2003, Lê Trọng Cúc & cs [37] trong “Danh lục các loài thực vật
Việt Nam”, tập II mô tả về 3 loài: P. bipinnatifidum Seem. 1868 ( CCVN, 2:
640), - Aralia bipinnatifida C. B. Clarke, 1879, - P. pseudoginseng var.
bipinnatifidus (Seem) H. L. Li, 1942 (FLPF, 54:187), - Vũ diệp tam thất, sâm
vũ diệp, Trúc tiết nhân Sâm, tam thất lá xẻ, Tam thất hoang, Sâm hai lần xẻ,
Hoàng liên thất, phân bố ở Lai Châu (Tà Phỉn), Lào Cai (Sa Pa), còn có ở Ấn

11
Độ, Trung Quốc; P. pseudoginseng Wall. 1829 (CCVN, 2: 641) đƣợc trồng ở
Lào Cai (Sa Pa), Hà Giang (Phố Bằng, So Phin), Cao Bằng (Tắc Tẻ, Thông
Nông), Vĩnh Phúc ( Tam Đảo), còn có ở Nêpal, Trung Quốc; P. stipuleanatus,
Tsai & Feng, 1975 – Tam thất hoang, Tam thất rừng, Bình biên tam thất, Sâm
tam thất, Phan xiết ( H’Mông) phân bố ở Lào Cai (Sa Pa: Núi Hàm Rồng, xã
Tà Phỉn, xã Bản Khoang, còn có ở Trung Quốc (Vân Nam); P. vietnamensis
Ha & Grushv. 1985 (“vietnamensis”) – Sâm ngọc linh phân bố ở Quảng Nam
(Trà My, Trà Linh, Trà Cang), Kon Tum (Đac Glây, Đác Choong, Ngọc Linh,
Đác Tô), Gia Lai (Long Hi), Lâm Đồng (Lang Bian). Tác giả mô tả ngắn
ngọn về hình thái, phân bố, sinh thái và công dụng.
Năm 2005, Nguyễn Tập [6] sau khi nghiên cứu và hệ thống lại các loài
thuộc chi Panax đã kết luận chi Panax ở Việt Nam có 5 loài, trong đó 3 loài
mọc tự nhiên là Sâm vũ diệp - P. bipinnatifidus Seem., Tam thất hoang - P.
stipuleanatus H.T. Tsai et K.M. Feng và Sâm việt nam - P. vietnamensis Ha
et Grushv. Cùng với hai loài nhập trồng là Nhân sâm - P. ginseng và Tam thất
P. notoginseng. Đồng thời cũng chính thức mô tả loài Tam thất hoang - P.
stipuleanatus H.T. Tsai et K.M. Feng. Từ đó đến nay, các tài liệu nhƣ: “Các
loài cây thuốc cần bảo tồn”, “Danh lục đỏ cây thuốc Việt Nam”, “Danh lục
thực vật Việt Nam” .v.v. đều đề cập đến loài Tam thất hoang - P.
stipuleanatus H.T. Tsai et KM. Feng kể trên. Tác giả mô tả đầy đủ chi tiết về
các loài Sâm này.
Năm 2007 “Sách đỏ Việt Nam” [38] mô tả về 3 loài thuộc chi Panax
gồm: P. bipinnatifidus (Seem). 1868 – Sâm vũ diệp (tên khác: Ngật đáp thất,
Tam thất hoang, Tam thất lá xẻ, Trúc tiết nhân sâm, Vũ diệp tam thất) phân
bố ở Lai Châu (Tả Phìn), Lào Cai (Sa Pa, Bát Xát, Than Uyên, núi Hoàng
Liên Sơn), còn có ở Ấn Độ, Nepal, Trung Quốc; P. stipuleanatus H.T. Tsai et
K.M. Feng. 1975 phân bố ở Lào Cai (Sa Pa, Bát Xát, núi Hoàng Liên Sơn),
ngoài ra còn có ở Trung Quốc (Vân Nam); P. vietnamensis Ha & Grushv.
1985 phân bố ở Quảng Nam (Trà My), Kon Tum (Đăk Tô, Đắk Glei, núi
Ngọc Linh, Gia Lai, Lâm Đồng, còn có ở Trung Quốc. Cả ba loài này Sách đỏ
đều ghi lại đặc điểm nhận dạng, sinh học và sinh thái, phân bố, giá trị, tình

12
trạng, phân hạng, biện pháp bảo vệ kèm ảnh minh họa.
Năm 2013, Phan Kế Long & cs. [39] đã ghi nhận bổ sung cho khu hệ
thực vật Việt Nam thứ P. vietnamensis var. fuscidiscus K. Komatsu, S. Zhu &
S.Q. Cai vốn đƣợc phát hiện trƣớc đó ở Vân Nam, Trung Quốc. Nhóm nghiên
cứu đã phát hiện một số điểm phân bố của thứ loài này tại Mƣờng Tè, Tam
Đƣờng và Sìn Hồ tỉnh Lai Châu nên đƣợc đặt tên là Sâm lai châu. Tác giả mô
tả đầy đủ chi tiết về đặc điểm hình thái, phân bố, công dụng.
Năm 2014, Phan Kế Long & cs. [40] tiếp tục nghiên cứu đặc điểm hình
thái và DNA của 7 mẫu cây không có cơ quan sinh sản thuộc chi Panax thu
thập đƣợc ở núi Phu Xai Lai Leng (Kỳ Sơn, Nghệ An). Kết quả cho thấy các
mẫu nghiên cứu có mối quan hệ gần gũi với loài Tam thất hoang - P.
stipuleanatus H.T. Tsai et K.M. Feng (dựa trên trình tự vùng gen ITS-rDNA).
Các mẫu có đặc điểm chung là lá từ không xẻ thùy đến xẻ thùy 1 lần (thùy
nông hoặc sâu); cuống lá có lá kèm.
Đến năm 2016, Trần Ngọc Lân và cộng sự [41] đã nghiên cứu đặc điểm
hình thái, thành phần hóa học và chỉ thị DNA của các mẫu đƣợc gọi là "Sâm
puxailaileng" cũng thu thập ở địa điểm trên. Nghiên cứu đã cho thấy các mẫu
này có mối quan hệ gần gũi với Sâm việt nam (P. vietnamensis), khác xa với
Tam thất hoang (P. stipuleanatus) và Sâm vũ diệp (P. bipinnatifidus) và kết
luận "Sâm phu xai lai leng" thuộc loài P. vietnamensis Ha et Grushv.. Các
mẫu này đƣợc tác giả cho rằng có đặc điểm đặc trƣng phân biệt với "Tam thất
hoang" ở "thân rễ mang các đốt mọc cùng 1 hƣớng, khi sấy khô thƣờng dẻo
và chuyển màu đen" trong khi loài "Tam thất hoang" có "thân rễ có các đốt
thƣờng mọc so le, khi sấy khô thƣờng cứng và chuyển màu trắng". Tuy nhiên,
những đặc điểm quan trọng khác lại chƣa đƣợc mô tả chi tiết nhƣ: có hay
không có lá kèm, hình dạng và màu sắc đĩa tuyến mật, hình dạng và màu sắc
quả chín... Ngoài ra, các tác giả đã không đề cập tới nơi lƣu giữ các tiêu bản
nghiên cứu. Sự không thống nhất của các kết quả nghiên cứu về đặc điểm
hình thái và trình tự vùng ITS-rDNA trên các cá thể thuộc chi Panax cùng thu
thập ở vùng núi Phu Xai Lai Leng ở trên có thể do tại đây có sự tồn tại của
nhiều hơn 1 loài Panax. Cả hai nhóm nghiên cứu đều cho rằng cần có những

13
nghiên cứu sâu hơn về đặc điểm hình thái và chỉ thị phân tử của các loài để có
đƣợc những kết quả chính xác hơn.
Nông Văn Duy & cs (2016) [42] khi nghiên cứu đặc điểm hình thái và
chỉ thị phân tử vùng gen ITS1-5.8S-ITS2, 18S rRNA và matK của quần thể
"Sâm langbiang" phân bố ở Cao nguyên Lâm Viên (núi Lang Biang, Lâm
Đồng) đã công bố đây là thứ mới của loài Sâm việt nam và đặt tên là P.
vietnamensis var. langbianensis N.V. Duy. V.T. Tran & L.N. Trieu.
Từ những nghiên cứu trên cho thấy chi Nhân sâm (Panax L.) ở Việt
Nam gồm 03 loài và 02 thứ ở Việt Nam mọc hoang dã bao gồm Sâm vũ diệp
(P. bipinnatifidus), Tam thất hoang (P. stipuleanatus), Sâm việt nam hay còn
đƣợc gọi là Sâm ngọc linh (P. vietnamensis var. vietnamensis), Sâm lai châu
(P. vietnamensis var. fuscidiscus) và Sâm langbian (P. vietnamensis var.
langbianensis).

14
CHƢƠNG 2. VẬT LIỆU VÀ PHƢƠNG PHÁP NGHIÊN CỨU
2.1. VẬT LIỆU, TRANG THIẾT BỊ VÀ DỤNG CỤ NGHIÊN CỨU
2.1.1. Vật liệu
Mẫu vật nghiên cứu bao gồm:
- 04 mẫu Sâm tự nhiên ở khu vực núi Phu Xai Lai Leng (Ký hiệu
Panax TB1 đến Panax TB4) trong đó có 02 mẫu có hoa non, lá, thân, rễ củ
(Panax TB1 và Panax TB2).
- 03 mẫu Sâm tại vƣờn ƣơm: 01 mẫu Sâm (Ký hiệu Panax TH1)
gồm lá và củ đƣợc thu tại vƣờn ƣơm ở xã Na Ngoi, Kỳ Sơn, Nghệ An; 02
mẫu sâm gồm lá và củ (Ký hiệu Panax ML1 và Panax ML2) đƣợc thu tại
vƣờn ƣơm ở xã Mƣờng Lống, Kỳ Sơn, Nghệ An thuộc sở hữu của Công ty
TNHH TH true MILK.
2.1.2. Địa điểm nghiên cứu
Phòng Thực vật học, Viện Sinh thái và Tài nguyên sinh vật và
phòng Phân loại học thực nghiệm và Đa dạng nguồn gen, Bảo tàng Thiên
nhiên Việt Nam.
2.1.3. Các hóa chất dùng trong nghiên cứu
* Các hóa chất dùng trong nghiên cứu bao gồm:
- Taq PCR mastermix 2x (QIAgen, Mỹ).
- BigDye terminator v3.1 (Applied Biosystems, Mỹ).
- Dung dịch TBE 10x (10mM Tris, 0.9 mM axir boric, 0.01 mM
EDTA) của hãng Invitrogen, Mỹ.
- Thuốc nhuộm an toàn (Malaysia).
- Kít tách chiết DNA (Trung Quốc).
- Kít tinh sạch sản phẩm PCR: QIAgen Quick Gel Extraction Kit
(QIAgen, Mỹ).
- Agarose (Invitrogen, Mỹ).

15
- Sephadex G50 (Sigma, Mỹ).
- Các hóa chất tách chiết và tinh sạch DNA (NaCl, CTAB, EDTA, Tris-
HCl, β-Mercaptoethanol, Isopropanol, Sodium acetate 3M, ethanol 100%,
70%, Rnase (10mg/ml)
- Hóa chất điện di agarose (agarose, đệm TAE, dye ...).
* Hóa chất cho phản ứng PCR:
- Taq DNA polymerase
- MgCl2
- Mastermix 2X bao gồm:
+ dNTP
+ MgCl2
+ Taq DNA polymerase
2.1.4. Thi t ị và dụng cụ nghiên cứu
* Các thiết bị chính sử dụng trong thao tác thực hiện thí nghiệm:
- Tủ lạnh thƣờng và tủ lạnh sâu.
- Bể ổn nhiệt và lò vi sóng.
- Tủ khử trùng, máy sấy chân không.
- Máy khuấy từ, máy trộn.
- Máy soi cực tím.
- Máy khử trùng Mac - 250Nex (Sanyo, Nhật Bản).
- Máy định lƣợng DNA NanoDrop One (Thermo scientific – Mỹ).
- Cân phân tích (Precisa, Thụy Sĩ).
- Máy ổn nhiệt (Memmert, Đức).
- Máy trộn rung (Vortex - Fluka, Mỹ).
- Máy ly tâm 5415D (Eppendorf, Đức).

16
- Máy ly tâm lạnh (Hitachi, Nhật).
- Máy PCR Mastercycler (Eppendorf, Đức).
- Máy điện di agarose gel (BioRad, Mỹ).
- Máy soi gel (UVP, Mỹ).
- Pipette, đầu côn, ống eppendorf.
* Các cặp mồi dùng trong nghiên cứu:
Bảng 2.1. Trình tự cặp mồi nhân bản vùng ITS-rDNA bằng kỹ thuật PCR với
cặp mồi theo Phan Kế Long và cs. (2014)
Ký hiệu
cặp mồi
Trình tự nucleotide (5’– 3’) Kích thƣớc
Lý thuy t
(bp)
Nhiệt độ
bắt cặp
(Tm) C
PaITS-F 5’- CAC TGA ACC TTA TCA TTT
AG AG -3’
700 49-50
PaITS-R 5’- CTT ATT GAT ATG CTT AAA
CTC AG -3'
* Phần mềm xử lý số liệu:
- ClustalW [43 ]
- GeneDoc 2.5 [44]
- MEGA X [45]

17
2.2. PHƢƠNG PHÁP NGHIÊN CỨU
2.2.1. Sơ đồ nghiên cứu
Hình 2.1. Sơ đồ các bƣớc thí nghiệm
2.2.2. Phƣơng pháp thu thập mẫu
Thu thập mẫu cây trƣởng thành trong các chuyến thực địa ở khu vực
núi Phu Xai Lai Leng với cơ quan sinh sản để làm tiêu bản, chụp ảnh, ghi
chép, gắn etiket cho mẫu, mô tả …
Đối với cây non và cây trong vƣờn ƣơm chỉ thu mẫu lá và ghi nhận đặc
điểm hình thái lá, củ; thu 01 lá chét và bảo quản trong túi nylon có chứa
silicagel. Giữ mẫu ở nhiệt độ phòng hoặc -20oC cho đến khi phân tích.
Thiết kế mồi Tách chiết
PCR
Đọc kết quả trình tự
gen
Cặp mồi DNA Tổng số
Phân tích số liệu, so sánh với
các loài thuộc chi Panax và
loài ngoài nhóm
Vùng gen ITS-
rDNA
Cặp mồi
Khai thác trình tự trong NCBI Mẫu lá sâm
Giải trình tự

18
2.2.3. Phƣơng pháp định loại hình thái
Chụp ảnh, mô tả mẫu vật thu thập đƣợc
Phân loại mẫu vật sử dụng phƣơng pháp so sánh hình thái dựa vào các
tài liệu có liên quan, đặc biệt là các tài liệu định loại ở Việt Nam, các nƣớc
lân cận Việt Nam và tài liệu mô tả gốc của các chi.
2.2.4. Phƣơng pháp nghiên cứu sinh học phân tử
2.2.4.1. Phương pháp tách chi t DNA tổng số
DNA tổng số của các mẫu sâm đƣợc tách chiết bằng Dneasy Plant Mini
Kit theo qui trình sau:
- Lấy mẫu lá bảo quản lạnh sâu (-20◦C) đem nghiền trong nitrogen lỏng
(-196◦C) thành dạng bột mịn. Lấy 100mg bột cho vào ống eppendorf 1,5ml.
- Thêm 400μl đệm AP1 và 4ul Rnase A. Trộn đều bằng vortex và ủ ở
65◦C trong 10 phút. Đảo trộn 2 – 3 lần trong quá trình ủ.
- Thêm 130μl đệm AP2. Trộn đều và ủ trong đá 5 phút.
- Hút dịch ly giải vào cột QIAshredder Mini. Ly tâm tốc độ 14000rpm
trong 2 phút.
- Chuyển dịch qua cột vào ống eppendorf 1,5ml mới nhƣng không xáo
trộn. Thêm 1,5 lần thể tích đệm AP3/E và trộn đều bằng pipet.
- Chuyển 650μl hỗn dịch vào cột Dneasy Mini. Ly tâm tốc độ
8000rpm trong 1 phút. Loại bỏ dịch qua cột. Lặp lại bƣớc này với phần
hỗn dịch còn lại.
- Đặt cột vào ống thu mới. Thêm 500μl đệm AW và ly tâm 8000rpm
trong 1 phút. Loại bỏ dịch qua cột.
- Thêm 500μl đệm AW. Ly tâm 14000rpm trong 2 phút. Loại bỏ dịch
qua cột.
- Chuyển cột sang ống eppendorf 1,5ml có ghi nhãn. Thêm 100μl đệm
AE để hoà tan. Ủ ở nhiệt độ phòng 5 phút. Ly tâm 6000rpm trong 1 phút. Lặp
lại bƣớc này, thu đƣợc dịch chiết chứa DNA tổng số, bảo quản ở -20◦C.

19
- Kiểm tra nồng độ DNA tổng số bằng máy định lƣợng DNA
NanoDrop One (Thermo scientific – Mỹ) (Hình 2.2).
Hình 2.2. Máy đo NanoDrop one
2.2.4.2. Phương pháp PCR (Polymerase Chain Reaction)
Ti n hành:
- Sau khi đo nồng độ DNA khuôn, tính và tạo các nồng độ DNA của
các mẫu nhƣ nhau để khi đƣa vào mỗi ống phản ứng thể tích DNA khuôn
đƣợc giống nhau, và đạt nồng độ 10ng trong tổng thể tích là 25 l/ 1 phản ứng.
- Tính lƣợng thể tích các thành phần khác trong phản ứng, trộn đều các
thành phần rồi chia vào các ống đã có khuôn DNA.

20
- Đặt các ống vào máy PCR, khởi động, cài đặt chu trình và cho máy chạy.
Phản ứng PCR đƣợc tiến hành với các thành phần nhƣ Bảng 2.2
Bảng 2.2. Các thành phần phản ứng PCR
TT Thành phần Nồng độ Thể tích (μl)
1 Taq PCR Mastermix 2x 12,5
2 Mồi xuôi 20 mM 1,0
3 Mồi ngƣợc 20 mM 1,0
4 DNA tổng số 10 ng 1,0
5 Nƣớc cất khử ion - 9,5
Tổng 25,0
Phản ứng PCR đƣợc thực hiện với chu trình nhiệt nhƣ sau: (Hình 2.3)
Hình 2.3. Chu trình phản ứng PCR
96oC 96
oC
2
phút
30
giây
50oC
25
giây
72oC 72
oC
40giây 5 phút
35 chu
kì
4oC

21
2.2.4.3. Tinh sạch sản phẩm PCR
Hình 2.4. Sơ đồ phản ứng QIAquick
Các bƣớc tiến hành:
Sử dụng Dneasy plant mini kit (Qiagen, Đức) để tinh sạch sản phẩm
PCR. Các bƣớc tinh sạch sản phẩm:
Bước 1: Chạy điện di sản phẩm PCR trên gel agarose 1%.

22
Bước 2: Cắt đoạn gel chứa băng DNA của từng mẫu, cho vào
eppendorf 1,5 ml.
Bước 3: Cân đoạn gel từng mẫu đã cắt. Bổ sung 3 lần thể tích dung dịch
QG theo trọng lƣợng từng mẫu (1g gel tƣơng đƣơng 1000 μ dung dịch QG).
Bước 4: Ủ ở 50 0C trong 10 phút cho tới khi gel tan hoàn toàn. Để làm tan
gel tốt. lắc đều eppendorf vài lần trong quá trình ủ, mỗi lần cách nhau 2 - 3 phút.
Bước 5: Khi gel đã tan hoàn toàn, kiểm tra màu của dung dịch mẫu.
Màu của dung dịch tốt là màu vàng của QG ban đầu.
Bước 6: Hút toàn bộ dung dịch cho vào cột MinElute đặt trong ống thu
dịch ly tâm của Kit và ly tâm 13.000 rpm trong 1 phút.
Bước 7: Chuyển cột sang ống thu dịch ly tâm khác, cho 700 μl dung
dịch PE vào cột, để mẫu ở nhiệt độ phòng khoảng 5 phút, ly tâm 13.000 rpm
trong 1 phút.
Bước 8: Chuyển cột sang ống thu dịch ly tâm khác và ly tâm với
13.000 rpm trong 1 phút.
Bước 9: Chuyển cột sang eppendorf 1,5 ml đã ghi kí hiệu mẫu, cho 10
μl EB vào cột và ly tâm 13.000 rpm trong 1 phút.
Bước 10: Bỏ cột, bảo quản sản phẩm PCR trong ống eppendorf ở -20 0C.
Bước 11: Sản phẩm PCR sau khi tinh sạch đƣợc gửi đi giải trình tự ở
Công ty Macrogen (Hàn Quốc).
2.2.4.4. Phương pháp phân tích số liệu
- Sử dụng chƣơng trình BLAST để tìm kiếm các trình tự tƣơng đồng đã
đƣợc các tác giả khác công bố trên ngân hàng DNA (Genbank).
- Các trình tự DNA đƣợc so sánh, phân tích với các loài thuộc chi
Panax và loài Eleutherococcus senticosus là loài ngoài nhóm (Bảng 2.3) sử
dụng các phần mềm ClustalW [46], GeneDoc 2.5 [47], phần mềm MEGA X
[48] đƣợc dùng để phân tích tiến hóa phân tử và xây dựng cây phát sinh
chủng loại theo phƣơng pháp Maximum Likelihood (ML) với giá trị

23
bootstrap dùng để đánh giá độ tin cậy của cây phát sinh chủng loại đƣợc
tính với 1.000 lần lấy mẫu thử (resampling) [45, 46].
Bảng 2.3. Danh sách trình tự sử dụng trong nghiên cứu thu thập trên Genbank
TT Loài/thứ ITS
1 P. ginseng U41680
U41681
2 P. quinquefolius U41688
U41689
3 P. japonicus Nhật Bản (JP) U41702
AF263373
4 P. japonicus var. major U41683
5 P. japonicus var. bipinnatifidus U41679
6 P. japonicus var. angustifolius AY271915
7 P. japonicus var. bipinnatifidus HQ112421
8 P. notoginseng U41685
9 P. zingiberensis U41699
10 P. vietnamensis var. vietnamensis
Trà Linh, Quảng Nam KJ418193
11 P. vietnamensis var. langbianensis
Langbian, Lâm Đồng KX768322
12 P. vietnamensis var. fuscidiscus
Mƣờng Tè, Lai Châu, Việt Nam (MT)
KJ418191

24
TT Loài/thứ ITS
13 P. vietnamensis var. fuscidiscus
Tam Đƣờng, Lai Châu, Việt Nam (TĐ) KJ418187
14 P. vietnamensis var. fuscidiscus
Sìn Hồ, Lai Châu, Việt Nam (SH) KJ418184
15 P. pseudoginseng U41693
U41694
16 P. assamicus AY233322
17 P. trifolius U41698
18 P. wangianus U41690
19 P. sinesis AY271920
20 P. omeiensis U41692
U41686
21 P. shangianus AY233328
22 P. stipuleanatus
HQ112444
HQ112441
AY271921
AY271922
23 P. stipuleanatus
Tam Đƣờng, Lai Châu, Việt Nam (TĐ) KJ418196
24 Eleutherococcus senticosus AB570259

25
CHƢƠNG 3. KẾT QUẢ VÀ THẢO LUẬN
3.1. KẾT QUẢ NGHIÊN CỨU
3.1.1. Đặc điểm nhận bi t chi Panax
Trên cơ sở kết hợp giữa phân loại về hình thái và đặc điểm trình tự
vùng gen ITS-rDNA, đề tài đã xác định đƣợc 03 loài thuộc chi Panax ở khu
vực Tây Nghệ An.
Khóa định loại các loài thuộc chi Panax ở vùng Tây Nghệ An
1a. Gốc cuống lá có phần phụ dạng gai.………… 1. Panax notoginseng
1b. Gốc cuống lá không có phần phụ
2a. Lá chét có cuống dài 0,5-0,8 cm………… 2. Panax stipuleanatus
2b. Lá chét không cuống hay gần nhƣ không cuống…3. Panax zingiberensis
3.1.2. Panax notoginseng (Burkill) F.H.Chen ex C.H. Chow -
Tam thất
F.H.Chen ex C.H. Chow, 1975. Acta Phytotax. Sin. 13(2):41; Xiang
Q. et al. 2007. Fl. China, 13: 490.
- Aralia quinquefolius var. notoginseng Burk. 1920
Cây thân thảo, cao 20-60 cm. Thân rễ nạc, rễ củ 1 hoặc nhiều hơn, màu
trắng đục, hình con quay, chiều dài từ 5-12cm, đƣờng kính khoảng 1-5cm. Lá
3-6, mọc vòng ở đỉnh thân, kép chân vịt, gốc cuống không có gai hay có phần
phụ dạng gai. Lá chét 3-7 hình trứng ngƣợc, trứng ngƣợc-thuôn, cỡ 3,5-13 x
1,5-7 cm, dạng màng, hai mặt có lông thƣa trên gân, gốc xiên, mép có răng
nhỏ, đỉnh nhọn. Chƣa thấy đƣợc hoa (Hình 3.1, Hình 3.2). Theo tài liệu
nghiên cứu, loài này có hoa màu lục vàng nhạt với 5 lá đài, cánh hoa 5, nhị 5
và bầu dƣới, 2 ô. Quả mọng hình cầu dẹt, khi chín có mầu đỏ, hạt hình cầu
mầu trắng [34].

26
Hình 3.1. Panax notoginseng (Burkill) F.H.Chen ex C.H. Chow - Tam thất
(lá mẫu TH1). Ảnh: Vũ Đình Duy

27
Hình 3.2. Panax notoginseng (Burkill) F.H.Chen ex C.H. Chow - Tam thất
(củ mẫu TH1). Ảnh: Vũ Đình Duy

28
Loc. class.: China: Yunnan: mountains near Mengzi, forests, 5000 ft.
Typus: A. Henry 11407 (K).
Sinh học và sinh thái: Mùa hoa tháng 7-8, mùa quả tháng 9-10. Cây
mọc trong rừng ở độ cao 1200-1800m.
Phân bố: Việt Nam. Còn có ở Trung Quốc.
Mẫu nghiên cứu: TH1
Địa điểm thu mẫu: Vƣờn ƣơm Công ty TNHH TH true MILK, Na
Ngoi, Kỳ Sơn, Nghệ An.
Nơi lưu trữ mẫu: Bảo tàng Thiên nhiên Việt Nam
Giá trị sử dụng: Cây làm thuốc.
Đặc điểm sinh học phân tử:
Mẫu Panax TH1 có trình tự vùng gen ITS-rDNA dài 588bp và tƣơng
đồng với P. notoginseng U41685 (Phụ lục 1).
Kết hợp giữa kết quả nghiên cứu hình thái và sinh học phân tử, đề tài
có thể khẳng định mẫu TH1 thuộc loài P. notoginseng Seem.
3.1.3. Panax stipuleanatus Tsai et Feng- Tam thất hoang
Tsai & Feng, 1975. Acta Phytotax. Sin. 13(2): 44, pl. 7, f. 4, 6; N. T.
Bân, 2003. Checkl. Pl. Spec. Vietn. 2: 1076; Xiang Q. et al. 2007. Fl. China,
13: 490
Cây thân thảo, sống nhiều năm, cao khoảng 50-70 cm, với thân rễ mập
giống nhƣ rễ tre, dài khoảng 7-12 cm, đƣờng kính 1-2,8 cm (Hình 3.3B, Hình
3.3). Thân khí sinh, mọc thẳng, nhẵn, đặc, cao 35-56, đƣờng kính 0,3-0,5 cm.
Lá mọc vòng ở đỉnh thân, xòe ra. Lá kép chân vịt, có 5 (-7) lá chét, dạng
màng, gân mặt trên lá có lông dạng móc; đôi lá chét dƣới cùng thƣờng nhỏ
nhất, hình thuôn rộng với cuống nhỏ dài khoảng 0,5-0,8 cm, gốc gần tròn,
chóp tròn, mép có răng cƣa hoặc nguyên. Chƣa thấy đƣợc hoa, quả và hạt nên
việc xác định tên các mẫu Sâm Panax thu ở khu vực Phu Xai Lai Leng là P.
stipuleanatus Tsai & Feng chƣa đủ tin cậy (Hình 3.3). Theo tài liệu nghiên

29
cứu, loài này có hoa màu vàng xanh. Cụm hoa tán mọc đơn độc ở đỉnh thân
giữa các lá, dài khoảng 12-16 cm với 5 lá đài nhỏ, cánh hoa 5, nhị 5 và bầu 2
ô. Quả mọng, gần hình cầu dẹt, đƣờng kính 0,6-1,2 cm, khi chín có màu đỏ.
Hạt 1 hoặc 2, màu xám trắng [38].
Ghi chú: Trong số các mẫu vật nghiên cứu thu đƣợc ở khu vực Phu Xai
Lai Leng có hai dạng khác nhau về lá: dạng có lá chét nguyên Hình 3.1C,
Hình 3.1) và dạng có lá chét xẻ (Hình 3.3C, Hình 3.3).
Hình 3.3. Mẫu Panax thu ở khu vực Phu Xai Lai Leng. A. Cây đủ rễ và lá; B.
Rễ củ; C. Lá; D. Hạt phấn; E. Cuống hoa; G. Hoa. Ảnh: Phạm Văn Thế.
Loc. class.: China: Yunnan: Maguan, forests in valleys, 1100-1700 m,
7 Dec. 1947. Typus: K. M. Feng 1364 (KUN)

30
Sinh học và sinh thái: Mùa hoa tháng 4-5, quả tháng 5-9 (10). Nhân
giống tự nhiên chủ yếu bằng hạt. Quả chín thƣờng bị chim ăn, hạt rơi xuống
đất lại bị loại sóc nhỏ ăn nhân hạt. Cá biệt trong tự nhiên, quả chín tồn tại đến
tháng 9 hoặc tháng 10. Cây có thể tái sinh tự nhiên bằng hạt, quả chín rụng
ngay xuống đất, xung quanh gốc cây mẹ, nếu không bị tác động, hạt sẽ nảy
mầm vào tháng 3 năm sau [33]. Phần thân mang lá lụi hàng năm vào mùa
đông, chồi mới mọc lên từ đầu thân rễ, vào đầu mùa xuân năm sau. Khi
thân rễ bị gãy, phần đầu mầm còn lại có khả năng tiếp tục tái sinh. Cây đặc
biệt ƣa ẩm và ƣa bóng, mọc rải rác trên đất có nhiều mùn, dƣới tán 10 rừng
kín thƣờng xanh ẩm, hoặc rừng có xen lẫn với sặt gai, ở độ cao từ 1600-
2300m [47].
Phân bố: Ở nƣớc ta, Tam thất hoang phân bố ở ở Lào Cai, Tây Nghệ
An và Hà Giang. Trên thế giới Tam thất hoang đƣợc tìm thấy ở tỉnh Vân Nam
- Trung Quốc.
Mẫu nghiên cứu: TB1-TB4 và ML1 ở Tây Nghệ An.
Địa điểm thu mẫu: Khu vực Phu Xai Lai Leng (TB1-TB4) và tại Vƣờn
ƣơm Công ty TNHH TH True Milk, Mƣờng Lống, Kỳ Sơn, Nghệ An
Nơi lưu mẫu: Bảo tàng Thiên nhiên Việt Nam.
Giá trị sử dụng: Cây làm thuốc [33].

31
Hình 3.4. Panax stipuleanatus Tsai et Feng - Tam thất hoang
(mẫu TB1: lá chét nguyên). Ảnh: Phạm Văn Thế
Hình 3.5. Panax stipuleanatus Tsai et Feng - Tam thất hoang
(mẫu TB2: lá chét xẻ thùy sâu). Ảnh: Phạm Văn Thế

32
Hình 3.6. Panax stipuleanatus Tsai et Feng - Tam thất hoang
(mẫu TB3: lá chét xẻ thùy sâu)

33
Hình 3.7. Panax stipuleanatus Tsai et Feng - Tam thất hoang
(mẫu TB4: lá chét xẻ thùy nông)

34
Mẫu ML1 thu ở vƣờn ƣơm của Công ty TNHH TH true MILK ở xã
Mƣờng Lống, Kỳ Sơn, Nghệ An có đặc điểm hình thái giống với mẫu TB1
thu ở khu vực Phu Xai Lai Leng nhƣ: Cây thảo, cao khoảng 50-70 cm. Thân
rễ mập giống nhƣ rễ tre. Thân trên mặt đất đơn, mọc thẳng, nhẵn, đặc, cỡ 35-
56 × 0,3-0,5 cm. Lá mọc vòng ở đỉnh thân, xòe ra. Lá kép chân vịt, có 5 lá
chét, dạng màng, gân mặt trên lá có lông dạng móc; đôi lá chét dƣới cùng
thƣờng nhỏ nhất, hình thuôn rộng với cuống nhỏ dài khoảng 0,5-0,7 cm, gốc
gần tròn, chóp tròn, mép có răng cƣa. Chƣa thấy hoa (Hình 3.8).
Hình 3.8. Panax stipuleanatus Tsai et Feng-Tam thất hoang (mẫu lá ML1).
Ảnh: Vũ Đình Duy

35
Hình 3.9. Panax stipuleanatus Tsai et Feng-Tam thất hoang (mẫu củ ML1).
Ảnh: Vũ Đình Duy
Khi đối chiếu với mẫu isotype của (P. stipuleanatus Tsai & Feng)
đƣợc lƣu giữ tại Viện Thực vật Bắc Kinh, Trung Quốc (Hình 3.10) cho thấy
hình thái lá của P. stipuleanatus Tsai & Feng còn có dạng xẻ thùy tƣơng tự
với các mẫu thu ở khu vực Phu Xai Lai Leng. Vì vậy trên cơ sở hình thái có
thể định danh tên khoa học của các mẫu Sâm Panax spp. ở khu vực Phu Xai
Lai Leng chỉ là Panax aff. stipuleanatus Tsai & Feng.

36
Hình 3.10. Mẫu (K.M.Feng 13694) isotype (P. stipuleanatus) hiện đang lƣu
trữ tại phòng tiêu bản Viện Thực vật Bắc Kinh (PE0025805)
Đặc điểm sinh học phân tử:
Các mẫu Panax TB1-TB4 và ML1 có chiều dài vùng gen ITS-rDNA

37
587bp và tƣơng đồng với loài Tam thất hoang P. stipuleanatus thu thập ở
Tam Đƣờng, Lai Châu, Việt Nam (KJ418196). Các mẫu P. stipuleanatus ở
Việt Nam có trình tự vùng gen ITS-rDNA khác với P. stipuleanatus
(AY271922) 01 nucleotide ở vị trí 414 (T>A); khác với các mẫu P.
stipuleanatus HQ112441 và HQ112444 01 nucleotide ở vị trí 139 (->C); Các
mẫu này khác với các mẫu P. stipuleanatus thu thập trên Genbank 01
nucleotide ở vị trí 469 (T>C) (Hình 3.11).
Hình 3.11. So sánh trình tự vùng gen ITS-rDNA của các mẫu Panax TB1-
TB4 và ML1 với trình tự vùng gen của loài P. stipuleanatus
Kết hợp giữa kết quả nghiên cứu hình thái và sinh học phân tử, có
thể khẳng định các mẫu TB1-TB4 và ML1 thuộc loài P. stipuleanatus Tsai
et Feng.

38
Hiện nay, Tam thất hoang (P. stipuleanatus) và Sâm vũ diệp (P.
bipinnatifidus) ở Việt Nam đƣợc phân biệt với nhau bởi hình thái lá chét của
cây trƣởng thành (lá chét hình mác thuôn, nhọn 2 đầu với lá chét xẻ thùy sâu,
hình lông chim). Các đặc điểm còn lại đều gần giống nhau, trong đó đặc điểm
hình thái cụm hoa, hoa, quả đều giống nhau (Bảng 3.1).
Bảng 3.1. So sánh đặc điểm hình thái giữa Tam thất hoang (P. stipuleanatus)
và Sâm vũ diệp (P. bipinnatifidus) (Nguyễn Tập, 2007)
Đặc điểm Tam thất hoang Sâm vũ diệp
Chiều cao 0,25 – 0,75 m 0,3 - 0,7 m
Thân rễ Mập, nằm ngang và thƣờng nổi
trên mặt đất; đƣờng kính 1,5 -
3cm
Mập, nằm ngang và thƣờng
nổi trên mặt đất; đƣờng kính
1,5 - 3,5cm
Phần thân
mang lá
1 - 3 tùy theo số đầu nhánh của
thân rễ; đƣờng kính thân từ 0,3
– 0,6 cm
1 - 3 tùy theo số đầu nhánh
của thân rễ; đƣờng kính thân
từ 0,3 – 0,6 cm
Lá Kép chân vịt gồm 1-3 cái, mọc
vòng ở ngọn; có cuống dài 5 -
10 cm; 5 lá chét; có cuống
ngắn, hình thuôn hay mác
thuôn, nhọn 2 đầu, mép có răng
cƣa; ở cây non có thể gặp lá xẻ
thùy nông, mép thùy nông khía
răng cƣa
Kép chân vịt, mọc vòng ở
ngọn; thƣờng gồm 3 cái; 3 –
5 lá chét xẻ thùy sâu, hình
lông chim, mép khía răng
cƣa
Cụm hoa Tán đơn, mọc ở ngọn; cuống
cụm hoa 5 – 10 cm; mang từ 30
– 80 hoa; cuống hoa mảnh dài 1
– 1,5 cm.
Tán đơn, mọc ở ngọn; cuống
cụm hoa 5 – 10 cm; mang từ
20 – 90 hoa; cuống hoa mảnh
dài 1 – 1,5 cm.

39
Đặc điểm Tam thất hoang Sâm vũ diệp
Hoa Hoa màu vàng xanh, 5 lá đài
nhỏ, 5 cánh hoa; 5 nhị; đầu vòi
nhụy chẻ đôi. Bầu 2 ô
Hoa màu vàng xanh, 5 lá đài
nhỏ, 5 cánh hoa; 5 nhị; đầu
vòi nhụy chẻ đôi. Bầu 2 ô
Quả Hình cầu đến hình cầu dẹt;
đƣờng kính 0,6 – 1,2cm; khi
chín có màu đỏ. Hạt: 2. Hạt gần
hình cầu hoặc gần giống hạt
đậu; màu trắng xám; vỏ cứng,
có rốn hạt
Hình cầu đến hình cầu dẹt;
đƣờng kính 0,6 – 1,2cm; khi
chín có màu đỏ. Hạt: 2. Hạt
gần hình cầu hoặc gần giống
hạt đậu; màu trắng xám; vỏ
cứng, có rốn hạt
Mùa hoa Tháng 3 – 4 Tháng 3 – 4
Mùa quả Tháng 4 – 8 (10) Tháng 4 – 8 (9)
Độ cao 1600 – 2400 m a.s.l. 1600 – 2400 m a.s.l.
Nông Văn Duy et al. (2016) [42] đã không phát hiện/chỉ ra sự sai khác
ở các vùng gen matK, ITS1-5.8S-ITS2 và 18S rDNA của các mẫu P.
stipuleanatus và P. bipinnatifidus thu thập ở Hồ Thầu, Tam Đƣờng, Lai Châu.
Zhou et al. (2018) [48] sau khi phân tích 129 mẫu Panax spp. thuộc 20 địa
điểm trên cơ sở nghiên cứu hình thái và phân tích 07 vùng gen đã kết luận P.
stipuleanatus có 2 dạng lá xẻ và không xẻ. Thêm vào đó, Trần Công Luận và
cs. (2009) [49] đã xác định có sự tƣơng đồng về thành phần hóa học của P.
stipuleanatus và P. bipinnatifidus thu thập ở Trạm nghiên cứu dƣợc liệu Sa
Pa, Lào Cai. Trên cơ sở các nghiên cứu trên, chúng tôi cho rằng Tam thất
hoang và Sâm vũ diệp thu đƣợc ở Tây Nghệ An có thể thuộc cùng một loài.
Điều này mở ra hƣớng nghiên cứu tiếp theo cho đề tài.
3.1.4. Panax zingiberensis C.Y. Wu & K.M. Feng-Tam thất gừng
Acta Phytotax. Sin. 13(2): 42, pl. 6, f. 5–6. 1975
C.Y. Wu & K.M. Feng, 1975. Acta Phytotax. Sin. 13(2): 42; Xiang Q.

40
et al. 2007. Fl. China, 13: 490
Thân thảo, cao 20-60 cm. Thân rễ nạc, dạng củ mọc ngang, dài khoảng
7-12cm, đƣờng kính 1-3cm, bò lan nhƣ rễ gừng. Lá 3-7, mọc vòng ở đỉnh
thân, kép chân vịt, gốc cuống lá không có gai hay có phần phụ dạng gai nhỏ.
Lá chét 3-5, không cuống hay gần nhƣ không cuống, hình bầu dục đến trứng
ngƣợc thuôn, cỡ 6-18x 3-6 cm, dạng màng, hai mặt đều có lông trên gân, gốc
hình nêm, mép có răng cƣa nhỏ, đỉnh có mũi hay mũi nhọn dài. Chƣa thấy
đƣợc hoa (Hình 3.12, 3.13). Theo tài liệu nghiên cứu, cụm hoa ở gốc, nằm ở
bên lá, cuống hoa dài 6-8 cm, ở phía cuối có một lá bắc và lá bắc con dạng
màng. Tràng hoa màu trắng, họng vàng. Bầu nhẵn, chia 3 ô. [38]
Hình 3.12. Lá loài Panax zingiberensis C.Y. Wu & K.M. Feng -Tam thất
gừng (mẫu lá ML2). Ảnh: Vũ Đình Duy.

41
Hình 3.13. Củ của loài Panax zingiberensis C.Y. Wu & K.M. Feng -Tam thất
gừng (mẫu củ ML2). Ảnh: Vũ Đình Duy.
Loc. class.: China: Yunnan: Maguan, broad-leaved evergreen forests,

42
1971. Typus: Z. R. Yang 7302 (KUN).
Sinh học và sinh thái: Mùa hoa tháng 7-8, mùa quả tháng 8-10. Mọc
trong rừng thƣờng xanh lá rộng.
Phân bố: Lào Cai (Sa Pa), Nghệ An, Còn có ở Trung Quốc.
Mẫu nghiên cứu: ML2
Địa điểm thu mẫu: Vƣờn ƣơm Công ty TNHH TH true MILK, Mƣờng
Lống, Kỳ Sơn, Nghệ An.
Nơi lưu mẫu: Bảo tàng Thiên nhiên Việt Nam.
Đặc điểm sinh học phân tử:
Mẫu Panax ML2 có trình tự vùng gen ITS-rDNA 587bp và tƣơng đồng
với loài P. zingiberensis. Mẫu Panax ML2 khác với P. wangianus ở 06
nucleotide ở vị trí 27 (C>T), 57 (C>T), 96 (T>C), 103 (C>A), 179 (C>T) và
526 (A>G). (Hình 3.14).

43
Hình 3.14. So sánh trình tự vùng gen ITS-rDNA của mẫu ML2 với các loài
trong cùng nhóm: P. zingiberensis và P. Wangianus.
Kết hợp giữa kết quả nghiên cứu hình thái và sinh học phân tử có thể
khẳng định mẫu ML2 thuộc loài P. zingiberensis Wu et Feng.
3.1.5. Đặc điểm hình thái chung của chi Panax ở vùng Tây Nghệ An
Panax L., Sp. Pl. 1058. 1753; Gen. Pl. 481. 1754, Proparte; ed. 6. 554.
1764; Jussieu, Gen. Pl. 218. 1789, Proparte; DC. Prodr. 4: 252. 1830; Walp.
Rep. 5: 925. 1846; Decne. et Planch, in Rev. Hortic. 105. 1854; Miquel in
Ann. Mus. Bot. Lugd.-Bat. 1: 14; 1863; Maxim. Diagn. Plant. Nov. 1: 264.
1866; Benth. et Hook. F. Gen. Pl.l: 938. 1867; Seem, in Joum. Bot. 6: 62.
1868; Graham in Joum. Arnold Arbor. 47: 132-136. 1966.; Grushv. I.V.,
Svortsona, N.T; Ha, T.D.; Hara. H. in Joum. Jap. Bot. 45:197-212. 1970;

44
Hoo, G., Tseng, c. J., Fl. China 54: 179-188, 1978; p. H. Ho, Fl. Vietnam 2:
515-516. 1999; Xiang Q. & p.p. Lowry n, Fl. China 13:489-491.2007;
Lectotype: p. quinquefolius Linn. sp. Pl. ed. 1513. 1753 (Graham 1966).
- Cây thảo, sống nhiều năm, cao đến khoảng lm.
- Thân rễ dày, mập, thƣờng nằm ngang gồm nhiều đốt ngắn, mỗi đốt có
1 vết sẹo lõm do thân khí sinh lụi hàng năm để lại, đôi khi ở một số cây phần
cuối thân rễ có một hoặc vài củ dạng con quay gần hình cầu đƣờng kính có
thể đến 5cm; riêng ở loài P. notoginseng thân rễ có dạng củ mọc hƣớng thẳng
hình thoi hoặc hình trụ; rễ nhỏ mọc xung quanh thân rễ. Thân trên mặt đất
thƣờng là 1 hoặc có thể 2-4 thân, hƣớng thẳng đứng, không phân nhánh, nhẵn,
gốc có vảy mỏng, lõi thân xốp.
- Lá mọc vòng ở đỉnh thân, thƣờng 3-5 lá; gốc cuống lá có thể có lá
kèm dạng mũi mác hoặc không; lá kép chân vịt có 5 (ít khi 3 hoặc 6- 7) lá
chét, hai lá chét ngoài cùng thƣờng có kích thƣớc nhỏ hơn; lá chét nguyên
hoặc có răng cƣa hoặc xẻ thùy lông chim; phiến lá hình trứng, trứng ngƣợc,
elip hoặc thuôn, mỏng, hai mặt màu xanh, mũi lá có đuôi nhọn, gốc lá hình
nêm, hẹp dài hoặc lệch, gân lá có lông cứng ở hai mặt.
- Cụm hoa thƣờng mang một tán đơn độc ở đỉnh; cuống hoa dài hơn
cuống lá; cụm hoa gồm từ 40 - 140 hoa; cúống hoa nhỏ đƣợc bao phủ bởi
nhiều mấu nhỏ dạng gai thịt dài 0,04 - 0,08 mm. Hoa lƣỡng tính, tỏa tròn,
đƣờng kính 2,0-3,5 mm, đài có 5 răng thấp dạng tam giác, nhẵn; cánh hoa 5,
màu xanh hoặc xanh mép hơi trắng, nhẵn; nhị 5, chỉ nhị mảnh, dài bằng hoặc
dài vƣợt quá cánh hoa, bao phấn 2 ô, đính lƣng; đĩa hoa lồi đến phẳng hoặc
lõm, màu xanh hơi ngả vàng đến xanh có vòng tím hoặc tím hoàn toàn; bầu 2
ô hoặc 1 ô, mỗi ô 1 noãn, vòi nhụy hợp hoặc xẻ 2.
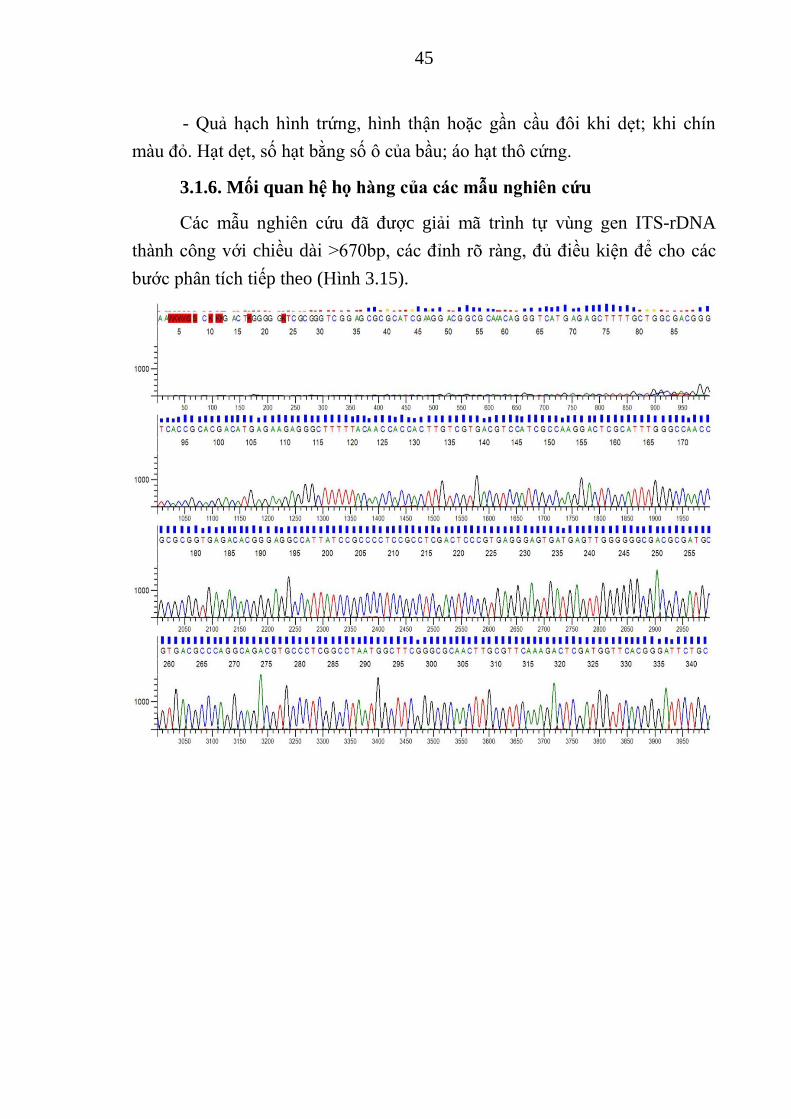
45
- Quả hạch hình trứng, hình thận hoặc gần cầu đôi khi dẹt; khi chín
màu đỏ. Hạt dẹt, số hạt bằng số ô của bầu; áo hạt thô cứng.
3.1.6. Mối quan hệ họ hàng của các mẫu nghiên cứu
Các mẫu nghiên cứu đã đƣợc giải mã trình tự vùng gen ITS-rDNA
thành công với chiều dài >670bp, các đỉnh rõ ràng, đủ điều kiện để cho các
bƣớc phân tích tiếp theo (Hình 3.15).

46
Hình 3.15. Sắc ký đồ trình tự vùng gen ITS-rDNA của mẫu Panax TB1
Chiều dài vùng gen ITS-rDNA sau khi chỉnh sửa, loại bỏ mồi, dao
động từ 587 (TB1-TB4 và ML1) đến 588bp (ML2 và TH1) (Bảng 3.2, Hình
3.14). Các mẫu P. stipuleanatus TB1-TB4 và ML1 giống hệt nhau. Khoảng
cách di truyền giữa các mẫu này với mẫu P. zingiberensis ML2 là 4,3%
(tƣơng ứng với 25 nucleotide sai khác), với mẫu P. notoginseng TH1 là 4,8%
(tƣơng ứng với 28 nucleotide sai khác). Khoảng cách di truyền giữa mẫu ML2
và TH1 là 2,9% (tƣơng ứng với 17 nucleotide sai khác) (Bảng 3.2).
Bảng 3.2. Thành phần nucleotide (%) của các mẫu nghiên cứu
Mẫu A C G T Chiều dài
TB1 21,9 31,3 28,6 18,1 587
TB2 21,9 31,3 28,6 18,1 587
TB3 21,9 31,3 28,6 18,1 587
TB4 21,9 31,3 28,6 18,1 587

47
ML1 21,9 31,3 28,6 18,1 587
ML2 21,9 30,6 28,9 18,5 588
TH1 21,8 30,4 29,1 18,7 588
Trung ình 21,9 31,1 28,7 18,2 587,3

48

49
Hình 3.16. Trình tự vùng gen ITS-rDNA của các mẫu nghiên cứu

50
Bảng 3.3. Khoảng cách di truyền (%) giữa các mẫu nghiên cứu (bên trên) và
số lƣợng nucleotide sai khác (bên dƣới) giữa các mẫu nghiên cứu
TB1 TB2 TB3 TB4 ML1 ML2 TH1
TB1 - 0 0 0 0 4,3 4,8
TB2 0 - 0 0 0 4,3 4,8
TB3 0 0 - 0 0 4,3 4,8
TB4 0 0 0 - 0 4,3 4,8
ML1 0 0 0 0 - 4,3 4,8
ML2 25 25 25 25 25 - 2,9
TH1 28 28 28 28 28 17 -
Trong mối quan hệ họ hàng với các loài trong chi Panax L. (Hình
3.15), các mẫu P. stipuleanatus TB1-TB4 và ML1 nằm trong nhóm P.
stipuleanatus và có mối quan hệ gần gũi nhất với P. stipuleanatus KJ418196
thu ở Tam Đƣờng, Lai Châu; Mẫu P. zingiberensis ML2 có quan hệ gần gũi
với P. zingiberensis U41699; Mẫu P. notoginseng TH1 có quan hệ gần gũi
với P. notoginseng U41685.

51
Hình 3.17. Mối quan hệ họ hàng của các mẫu nghiên cứu với các loài/thứ
trong chi Panax L. trên cơ sở phân tích trình tự vùng gen ITS-rDNA theo
phƣơng pháp Maximum Likelihood (ML). Mô hình tối ƣu GTR+G+I.
Eleutherococcus senticosus là loài ngoài nhóm. Giá trị ở gốc là giá trị
bootstrap >50%

52
3.2. THẢO LUẬN
Vùng Tây Nghệ An có diện tích rất lớn với 1,3 triệu ha với 09 đỉnh núi
thuộc Bắc Trƣờng Sơn cao trên 1000 m so với mặt biển nhƣ Pù Hon 1447 m,
Pù Mát 1814 m, Phu Xai Lai Leng 2711 m, Pù Hút 2452 m, Pù Lon 1561 m,
Pu Đen Đin 1540 m, Rào Cỏ 2286 m, Pù Huống 1200 m và Chốp Cháp 1705
m có khả năng phù hợp với điều kiện sinh trƣởng và phát triển của các loài
cây thuốc thuộc chi Sâm (Panax L.). Theo nghiên cứu của Phan Ke Long et
al., 2014 [40] và Trần Ngọc Lân và cs., 2016 [41] cho thấy tại vùng núi Phu
Xai Lai Leng có thể tồn tại 02 loài thuộc chi Panax. Trong nghiên cứu này,
chúng tôi thu thập đƣợc các mẫu ngoài tự nhiên có hình thái lá chét giống với
mô tả của Tam thất hoang và Sâm vũ diệp. Tuy nhiên, kết quả nghiên cứu
hình thái thấy có sự khác biệt 3 dạng lá (lá không xẻ, lá xẻ thùy nông và lá xẻ
thùy sâu), tiếp tục phân tích bằng kỹ thuật sinh học phân tử trên có sở phân
tích trình tự vùng gene ITS-rDNA đã xác định các mẫu này đều cùng là loài
Tam thất hoang P. stipuleanatus.
Mẫu vật ở vƣờn ƣơm của Công ty TNHH TH true MILK ở xã Na Ngoi
và Mƣờng Lống, Kỳ Sơn, Nghệ An khá đa dạng, nguồn gốc cây trồng ở đây
đƣợc du nhập từ các tỉnh lân cận về trồng và một số cây do ngƣời dân địa
phƣơng thu thập trong vùng. Trên cơ sở phân loại hình thái kết hợp phân tích
trình tự vùng gen ITS-rDNA, chúng tôi đã xác định đƣợc các loài trồng ở đây
bao gồm Tam thất P. notoginseng, Tam thất hoang P. stipuleanatus và P.
zingiberensis. Trong đó, loài P. zingiberensis đã đƣợc ghi nhận ở Việt Nam
[38] tuy nhiên chƣa có mẫu nghiên cứu vì vậy cần tiếp tục điều tra nghiên cứu
để xác định nơi phân bố, điều kiện sống, tình trạng bảo tồn và thu thập tiêu
bản của loài này ở khu vực Tây Nghệ An phục vụ công tác nghiên cứu, bảo
tồn, xác định giá trị dƣợc liệu và phát triển bền vững.
Ở Việt Nam, các loài thuộc chi Panax L. đều ở tình trạng Cực kỳ nguy
cấp (CR) và đƣợc bảo vệ tại Nghị định 06/2019/NĐ-CP ngày 22/01/2019
(nhóm IA). Nguyễn Tập (2007) [50], Sâm ngọc linh đã đƣợc nghiên cứu khá
kỹ lƣỡng trong đó đã phát hiện ra hàm lƣợng “ocotillol saponin” nhƣ
majonoside-R2 rất cao (5,5% trong thân rễ khô) có tác dụng an thần cao mà

53
P. ginseng không có. Hơn nữa, majonoside-R2 có khả năng kháng Epstein-
Barr virus early antigen (EBV-EA) là tác nhân gây bệnh ung thƣ vòm họng
(Yamasaki, 1999) [51]. Dựa trên giá trị của Sâm ngọc linh đã đƣợc chứng
minh, Trên cơ sở thành công của Công ty TNHH TH true MILK trong bảo
tồn và phát triển một số loài thuộc chi Panax L. ở huyện Kỳ Sơn, Nghệ An
cho thấy tiềm năng phát triển nguồn dƣợc liệu quý hiếm này tại vùng Tây
Nghệ An. Hơn nữa, có thể nghiên cứu chuyển vị Sâm ngọc linh P.
vietnamensis var. vietnamensis tại đây nhằm phát triển kinh tế địa phƣơng,
xóa đói, giảm nghèo.

54
KẾT LUẬN VÀ KIẾN NGHỊ
1. KẾT LUẬN
Qua quá trình nghiên cứu chi Panax ở vùng Tây Nghệ An đã thu đƣợc
kết quả sau:
1. Đã phân tích tổng hợp đƣợc đặc điểm hình thái các loài Sâm (Panax
L. ) ở khu vực Phu Xai Lai Leng bao gồm P. stipuleanatus Tsai et Feng, P.
zingiberensis Wu et Feng và P. notoginseng Seem.
2. Các mẫu Panax TB1-TB4 thu đƣợc ở núi Phu Xai Lai Leng và mẫu
ML1 thu tại vƣờn ƣơm xã Mƣờng Lống, Kỳ Sơn, Nghệ An có chiều dài vùng
gen ITS-rDNA 587bp và tƣơng đồng với loài Tam thất hoang P. stipuleanatus
thu thập ở Tam Đƣờng, Lai Châu, Việt Nam (KJ418196). Các mẫu P.
stipuleanatus ở Việt Nam có trình tự vùng gen ITS-rDNA khác với
P.stipuleanatus (AY271922) 01 nucleotide ở vị trí 414 (T>A); khác với các
mẫu P. stipuleanatus HQ112441 và HQ112444 01 nucleotide ở vị trí 139 (-
>C); Các mẫu này khác với các mẫu P. stipuleanatus thu thập trên Genbank
01 nucleotide ở vị trí 469 (T>C).
Mẫu Panax TH1 thu tại xã Na Ngoi, Kỳ Sơn, Nghệ An có trình tự vùng
gen ITS-rDNA dài 588bp và tƣơng đồng với P. notoginseng U41698.
Mẫu Panax ML2 thu tại vƣờn ƣơm xã Mƣờng Lống, Kỳ Sơn, Nghệ An
có trình tự vùng gen ITS-rDNA 587bp và tƣơng đồng với loài P.
zingiberensis. Mẫu Panax ML2 khác với P. wangianus ở 06 nucleotide ở vị
trí 27 (C>T), 57 (C>T), 96 (T>C), 103 (C>A), 179 (C>T) và 526 (A>G).
2. KIẾN NGHỊ
Tiếp tục điều tra nghiên cứu hình thái học, sinh học phân tử để có thêm
thông tin về chi Panax giúp khai thác, bảo vệ và sử dụng nguồn tài nguyên
thiên nhiên ở vùng Tây Nghệ An.

55
TÀI LIỆU THAM KHẢO
1. Nguyễn Bá, 2007, Thực vật học, NXB Giáo dục, Hà Nội.
2. Nguyễn Tiến Bân (chủ biên) và nhiều ngƣời khác, 1996, Sách đỏ
Việt nam, phần (II), NXB Khoa học và kỹ thuật, Hà Nội, tr. 29-31, 204-208.
3. Lê Thanh Sơn và Nguyễn Tập, 2006, Những đặc điểm sinh thái cơ
bản của Sâm ngọc linh, Tạp chí Dược liệu, 11(4), tr. 145-147.
4. Chính phủ nƣớc CHXHCNVN và dự án của Quỹ Môi trƣờng toàn
cầu VIE/91/G31, 1995, K hoạch hành động đa dạng sinh học của Việt nam.
5. Xiang Q., P. P. Lowry, 2007, Flora China, 13, pp.1-548. Science
Press & Missouri Botanical Garden Press, Beijing & St. Louis.
6. Nguyễn Tập, 2005, Các loài thuộc chi Panax L. ở Việt Nam, Tạp chí
Dược liệu, 10(3), tr. 71 – 76.
7. Linnaeus C., 1735, Systema naturae, pp. 8-13, Leyden.
8. Linnaeus C., 1753, Species plantarum, Laurentius Salvius, pp.133-
134, 202, 273, 1058.
9. Wen J., 2000, Species diversity, Nomenclature, phylogeny,
Biogeography and Classificational Ginseng genus (Panax L., Araliaceae),
Proceeding of the International Ginseng Workshop “Utilza of Biotechno,
genetic and cultural approaches for North American and Asian Ginseng
improvement”, Zamir K. Punja, pp. 29-45
10. Nocerino E. et al., 2000, Fitoterapia, 71, pp. 1-5.
11. Ha T.D. & Grushvitsky I.V., 1985, A new species of the genus
Panax (Araliaceae) from Vietnam, Bot. J. (Leningrad), tr. 519-522.
12. Candolle, A. P. de, 1830, Prodromus systematis naturalis regni
vegetabilis, Araliaceae, Vol. 4, Treuttel & Wurtz, Paris, pp.251-266.
13. Decaisne, J. & E. Planchon, 1854, Esquisse d’une monographie des
Araliacées. Rev. Hort. 4, pp.104-109.
14. Bentham G. and J. D. Hooker, 1867, Genera plantarum, Araliaceae,

56
L. Reeve & Co., London, Vol I, pp. 931-947.
15. Clarke C. B., 1879, The Flora of British India, London, vol 2.
pp.808.
16. Seemann, 1868, Journal of Botany, vol 6, pp.54.
17. Harms H, 1897, Zur Kenntnis der Gattungen Aralia und Panax,
Botanische Jahrbucher fur Systematik Pflarzengeschichte, 23, pp.1–23.
18. Britton, N. L. and A. Brown, 1913, An illustrated flora of the
northern United States, Canada and the British pos-sessions, New York:
Charles Scribner’s Sons, vol2, 2nd ed.
19. Hoo, G., 1961, The Systematics, relationship and distribu-tion of the
Araliaceae of China. Bulletin of Amoi Uni-versity (NaturalSciences), 8(1),
pp. 1–11.
20. Hara H., 1970, On the Asiatic species of genus Panax, J. Jap. Bot.,
45, pp. 197-222.
21. Wen J., Zimmer E. A., 1996, Phylogeny and Biogeography of Panax
(Araliaceae): Inference from ITS sequences of nuclear ribosomal DNA, Mol.
Phylogen., 6, pp. 166-167.
22. Florence c., Lee, 1992, FacTsabout Ginseng, the elixir of life,
Hollym, pp.44-46.
23. Grushvitzky, I. V., Tikhomirov, V. N., Aksenov, E. S. & Shibakina,
G. V, 1969, Succulent fruit with carpophore inspecies of the genus
Stilbocarpa Decne et Planch. (Araliaceae), Bull. Moscow Soc. Natur., Biol,
74, pp. 64–76.
24. Due N. M., Nham N. T. et al.,1993, Saponin from Vieetnamese
Ginseng, Panax vietnamensis Ha et Grushv, collected in central Vietnam, I,
Chemical phrm. Bull., 42, pp. 2010-2014.
25. Due N. M., Kasai R. et al., 1994, Saponin from Vieetnamese
Ginseng, Panax vietnamensis Ha et Grushv, collected in central Vietnam, II,
Chemical pharm. Bull., 42, pp. 115-122.

57
26. Due N. M., Kasai R. et al.,1997, Saponin composition of Vietnamese
Ginseng and its significance from phamacognostical points of view,
Proceeding of the First Indochina Conference on Pharmaceutical Sciences,
Pharmacy in Harmony, Bangkok, Thailan, pp. 273-283.
27. Viguier R., 1908-1923, Flore Générale De L’Indo-Chine, Part 2(5),
pp. 1162-1163.
28. Phạm Hoàng Hộ, 1970, Cây cỏ Miền nam Việt Nam, Nxb Bộ Văn
hóa giáo dục và thanh niên trung tâm học liệu, 1, tr.989.
29. Võ Văn Chi, & cs, 1973, Cây cỏ thƣờng thấy ở Việt Nam, Nxb Khoa
học kĩ thuật, Hà Nội, 3, tr. 109.
30. Phạm Hoàng Hộ, 1993, Cây cỏ Miền Nam Việt Nam, Nxb Trẻ, Hà
Nội, 2(2), tr. 640-641.
31. Grushvitzky, I.V., Skvortsova, N.T., Ha Thi Dung & Arnautov,
N.N., 1996, Fam. Araliaceae Juss. - Ngu gia bi. Vascular plants Synopsis of
Flora, 2, pp.16-42.
32. Trần Đình Lý & cs, 1993, 1900 loài cây có ích ở Việt Nam, Nxb Thế
giới, tr. 32-33.
33. Đỗ Tất Lợi, 1995, Những cây thuốc và vị thuốc Việt Nam, Nxb
Khoa học và kĩ thuật, Hà Nội, tr. 376-379, 1004-1007.
34. Võ Văn Chi 1997, Từ điển cây thuốc Việt nam, Nxb Y học, Hà Nội,
tr. 19-21, 64.
35. Lê Trần Đức, Cây thuốc Việt Nam trồng hái chế biến trị bệnh ban
đầu, Nxb Nông nghiệp, Hà Nội, tr. 1013-1026.
36. Phạm Hoàng Hộ, 2000, Cây cỏ Việt Nam, Nxb Trẻ, Hà Nội, 2, tr.
515-516.
37. Lê Trọng Cúc & cs, 2003, Danh lục các loài thực vật Việt Nam, Nxb
Nông Nghiệp, Hà Nội, tr.640.
38. Bộ Khoa học và Công nghệ, Viện Khoa học và Công nghệ, 2007,
Sách đỏ Việt Nam, Phần II thực vật, Nxb Khoa học tự nhiên và Công nghệ,

58
Hà Nội, tr. 85-89.
39. Phan Ke Long, Le Thanh Son, Phan Ke Loc, Vu Dinh Duy & Pham
Van The, 2013, Lai chau ginseng Panax vietnamensis var. fuscidiscus K.
Komatsu, S. Zhu & S. Q. Cai I. Morphology, Ecology, Distribution and
Conservation Status. In Proc. 2nd VAST-KAST Workshop on Biodiversity
and Bio-Active Compounds, tr. 65-73.
40. Phan Ke Long, Tran Thi Viet Thanh, Nguyen Thien Tao, Phan Ke
Loc, Nguyen Tu Lenh, Nguyen Tien Lam, Dang Xuan Minh, 2014,
Morphological and moleculer characteristics of Panax sp. (Araliaceae) from
Phu Xai Lai Leng Mountain, Nghe An province, Vietnam, Journal of Biology,
36(4), tr. 494-499.
41. Trần Ngọc Lân & cs, 2016, Kết quả nghiên cứu về loài Sâm
Phuxailaileng ở vùng núi cao tỉnh Nghệ An, tạp chí Khoa Học và Công Nghệ
Nghệ An, số 12/2016. Tr7-11.
42. Nong Van Duy, Le Ngoc Trieu, Nguyen Duy Chinh, Van Tien Tran
(2016). A new variety of Panax (Araliaceae) from Lam Vien Plateau,
Vietnam and its molecular envidence. Phytotaxa 277 (1), pp. 47-58
43. Thompson, J.D., Higgins, D.G. and Gibson, T.J. CLUSTAL W,
1994, improving the sensitivity of progressive multiple sequence alignment
throught sequence weighting, positions-specific gap penalties and weight
matrice choice, Nucleic Acids Research, 22, pp. 4673-4680.
44. Nicholas K.B., Nicholas H.B.J., 1997, GeneDoc Analysis and
Visualization of Genetic Variation, EMBNEW, pp 4-14
45. Tamura K., Dudley J., Nei M., Kumar S., 2007, MEGA4: Molecular
Evolutionary Genetics Analysis (MEGA) sofware version 4.0”, Molecular
Biology and Evolution 10.1093/molbev/msm092.
46. http://en.wikipedia.org/wiki/Maximum_parsimony_(phylogenetics).
47. Komatsu K., Tohda C. & Zhu S., 2005, Ginseng drugs – Molecular
and Chemical characteristics and possibility as antidementia drugs, Current

59
Topics Nutraceutical Res, 3, tr. 47-64.
48. Zhou M., Gong X. & Pan Y., 2018, Panax species identification with
the assitance of DNA data, Genetic Resources and Crop Evolution, 65, pp.
1839-1856
49. Trần Công Luận, Lƣu Thảo Nguyên, Nguyễn Tập, 2009, Nghiên cứu
thành phần hóa học của hai loài Sâm vũ diệp (P. bipinnatifidus Seem.)và Tam
thất hoang(P. stipuleanatus Tsai et Feng), Tạp chí Dƣợc liệu, 14(1), tr. 17-23.
50. Nguyễn Tập, 2007, Cẩm nang cây thuốc cần bảo vệ ở Việt Nam,
IUCN, tr.161-170.
51. Konoshima T., Takasaki M., Ichiishi E., Murakami T., Tokuda H.,
Nishino H., Duc N.M., Kasai R., Yamasaki K., 1999, Cancer
chemopreventive activity of majonoside-R2 from Vietnamese ginseng, Panax
vietnamensis, Cancer Letters, 147(2), pp.11-16.

PHỤ LỤC
Phụ lục 1: So sánh trình tự vùng gen ITS-rDNA của các mẫu nghiên cứu với
các loài trong chi Panax.L (bảng)





Phụ lục 2: Thực nghiệm trong phòng thí nghiệm của Bảo tàng Thiên nhiên
Việt Nam

Hà Nội, ngày tháng năm 2020
Xác nhận của cán ộ hƣớng dẫn
Học viên cao học
Phan K Long
Nguyễn Thị Thanh Hƣơng
Nguyễn Thị Hiên

BẢNG TRA CỨU TÊN TIẾNG VIỆT
Tên Việt Nam Trang xuất hiện
Sâm vũ diệp 3, 13, 16, 17, 18, 45, 58, 60
Tam thất hoang
3, 14, 16, 17, 28, 34, 36, 37, 38, 39, 40, 44, 45, 58,
60
Sâm ngọc linh 3, 18, 58, 59
Tam thất 3, 16, 25, 31, 32, 33, 58
Tam thất gừng 39
Sâm nhật bản 13, 15, 28
Sâm lai châu 18
Sâm langbian 18
Sâm việt nam 15, 17, 18
Sâm puxailaileng 17

BẢNG TRA CỨU TÊN KHOA HỌC
Tên latinh Trang xuất hiện
Aralia bipinnatifida 15
Aralia quinquefolia
var. notoginseng 31
Aralia. 11
Araliaceae 1, 7, 8, 9, 10, 11, 13
Eleutherococcus
senticosus 28, 57
Eleutherococcus
senticosus 30
P. bipinnatifidum 15
P. japonica 7, 13, 15
P. trifolius 7, 9, 29
Panax 1, 2, 4, 5, 6, 7, 10, 11, 12, 13, 17, 19, 28, 34, 35, 43,
44, 46, 49, 50, 52, 56, 57, 58, 59, 60
Panax ginseng 3, 1, 7, 16, 28, 57, 59
Panax vietnamensis 8, 13, 14, 15, 16, 17
Panax. japonicus var.
major 28
Panax. notoginseng 3, 7, 16, 28, 29, 31, 32, 33, 34, 50, 52, 56, 57, 58
Panax. pseudoginseng
var. bipinnatifidus 15
Panax. stipuleanatus 3, 6, 9, 15, 16, 17, 29, 30, 25, 34, 35, 37, 38, 39, 40,
41, 42, 43, 44, 45, 46, 50, 52, 56, 57, 58, 60
Panax. vietnamensis
var. fuscidiscus 16, 18, 29, 57
Panax. vietnamensis
var. langbianensis 17, 18, 29, 57
Panax. vietnamensis
var. vietnamensis 29, 57,59
Panax. zingiberensis 29, 39, 47, 48, 49, 50, 52, 56, 57, 58, 60
Panax.assamicus 29, 57
Panax.bipinnatifidus 3, 8, 10, 12, 13, 14, 15, 16, 17, 18, 45, 46, 57
Panax.japonicus 28, 57
Panax.japonicus
var.angustifolius 28, 57
Panax.japonicus
var.bipinnatifidus 28, 29, 57
Panax.major 57
Panax.omeiensis 29, 57
Panax.pseudoginseng 8, 10, 13, 14, 15, 29, 57,
Panax.pseudoginseng
var.elegantior 57

Panax.quinquefolius 7, 9, 10, 28, 57
Panax.shangianus 29, 57
Panax.sinensis 29, 57
Panax.wangianus 29, 49, 57





![T¥ rÔ? Ñ ä>Y¾Y¶Ø Yk³ Ë]äNR æm È`pû iØt4èÃZ9: èY¾Y Ø Yk³ Ë]äNR æm È`pû iØt4èÃZ9: è Áö¤Âk W>³*](https://static.fdocument.pub/doc/165x107/60492622dd3de204c645b5a9/t-r-yy-yk-nr-m-p-it4fz9-yy-yk-nr.jpg)












