Myofibroblast proliferation and heterogeneity are ... · RESEARCH ARTICLE SUMMARY SKIN REPAIR...
10
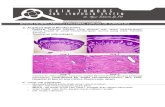
RESEARCH ARTICLE SUMMARY ◥ SKIN REPAIR Myofibroblast proliferation and heterogeneity are supported by macrophages during skin repair Brett A. Shook*, Renee R. Wasko, Guillermo C. Rivera-Gonzalez, Emilio Salazar-Gatzimas, Francesc López-Giráldez, Biraja C. Dash, Andrés R. Muñoz-Rojas, Krystal D. Aultman, Rachel K. Zwick, Vivian Lei, Jack L. Arbiser, Kathryn Miller-Jensen, Damon A. Clark, Henry C. Hsia, Valerie Horsley* INTRODUCTION: Fibroblasts produce extra- cellular matrix (ECM) molecules that regulate tissue strength and resilience. Imbalanced ECM maintenance leads to tissue dysfunction. Al- though multiple populations of fibroblasts support uninjured skin function, the extent of fibroblast diversity and the presence of func- tionally distinct subsets in adult fibrotic skin are poorly defined. The inability to find a single molecular marker that identifies all activated “myofibroblasts” during wound healing sug- gests the existence of multiple subsets of ECM- producing myofibroblasts. Furthermore, little is known about the cellular and molecular mechanisms that promote the expansion of individual myofibroblast subsets and support cellular heterogeneity. Although macrophages influence myofibroblast numbers and ECM de- position after injury, subpopulations of wound bed macrophages have only recently been de- fined, and specific interactions with fibroblasts have not been explored. RATIONALE: Variation in healing and scarring rates in multiple tissues suggests that fibro- blast diversity exists. To develop therapies target- ing fibrotic responses, functionally distinct subsets of myofibroblasts and the mechanisms that support individual populations must be uncovered. We used a comprehensive, hierar- chical fluorescence-activated cell sorting strat- egy to define myofibroblast subsets in skin wound beds from adult mice. This strategy revealed distinct subsets of wound bed myofi- broblasts, including an abundant population that contains the cell surface marker profile of adipocyte precursor cells (APs). We examined myofibroblast subsets in different cutaneous fibrotic contexts and explored mechanisms that selectively promote the expansion of APs. RESULTS: Genetic lineage tracing and flow cytometry revealed distinct subsets of wound bed myofibroblasts that express smooth muscle actin and collagen. The most abundant pop- ulations were CD26-expressing APs and a sub- set with high cell surface levels of CD29 (CD29 High ). Although transcriptomic analysis revealed that each myofibroblast subset has a distinct gene expression profile, functional analyses suggest that myofibroblast subsets make both overlap- ping and distinct contribu- tions to repair. APs were significantly reduced and CD29 High cells were more abundant in wound beds from aged mice and skin from mice that underwent bleomycin-induced fibrosis, suggesting that the fibrotic environment influences myofibroblast composition. Injury and repair-related changes in AP transcription implicated macrophage signaling in the modulation of AP gene ex- pression. Genetic ablation and cell transplan- tations of different myeloid cells revealed that macrophages expressing macrophage galac- tose N-acetylgalactosamine–specific lectin 2 ( Mgl2/CD301b) directly stimulate proliferation in a subset of APs and not in other myofibroblast subsets. By combining in vitro cytokine stimula- tion with in vivo signaling pathway inhibition, we identified multiple CD301b + macrophage– secreted factors (platelet-derived growth factor C and insulin-like growth factor 1) that selectively stimulate AP proliferation, thus supporting the heterogeneity of wound bed myofibroblasts. CONCLUSION: We identified multiple popu- lations of skin myofibroblasts and observed that the composition of myofibroblasts is dependent upon the fibrotic environment. Distinct inter- actions allow CD301b + macrophage–derived sig- naling to selectively activate the proliferation of APs and not other myofibroblasts. These results have potential for the development of therapies that target multiple cellular populations or sig- naling pathways under conditions associated with excessive or deficient ECM deposition, such as wound healing and fibrosis. ▪ RESEARCH Shook et al., Science 362, 909 (2018) 23 November 2018 1 of 1 The list of author affiliations is available in the full article online. *Corresponding author. Email: [email protected] (B.A.S.); [email protected] (V.H.) Cite this article as B. A. Shook et al., Science 362, eaar2971 (2018). DOI: 10.1126/science.aar2971 CD29 High Adipocyte precursor Adipocyte CD29 Low Aging Young adult Uninjured skin Selective activation of adipocyte precursor proliferation Scab Wound Activated fibroblast (myofibroblast) Fibroblast CD301b + Mφ Regulation of myofibroblast diversity in skin wounds. After injury, multiple subsets of fibroblasts become activated myofibroblasts that contribute to tissue repair and scar formation. Wound bed macrophages expressing CD301b selectively activate proliferation in APs and not other myofibroblasts.With age, impaired healing is associated with a reduction in CD301b + macrophages (Mf) and APs. These findings identify distinct cellular and molecular interactions that support myofibroblast heterogeneity. ON OUR WEBSITE ◥ Read the full article at http://dx.doi. org/10.1126/ science.aar2971 .................................................. on September 12, 2020 http://science.sciencemag.org/ Downloaded from
Transcript of Myofibroblast proliferation and heterogeneity are ... · RESEARCH ARTICLE SUMMARY SKIN REPAIR...

RESEARCH ARTICLE SUMMARY◥
SKIN REPAIR
Myofibroblast proliferation andheterogeneity are supported bymacrophages during skin repairBrett A. Shook*, Renee R. Wasko, Guillermo C. Rivera-Gonzalez, Emilio Salazar-Gatzimas,Francesc López-Giráldez, Biraja C. Dash, Andrés R. Muñoz-Rojas, Krystal D. Aultman,Rachel K. Zwick, Vivian Lei, Jack L. Arbiser, Kathryn Miller-Jensen, Damon A. Clark,Henry C. Hsia, Valerie Horsley*
INTRODUCTION: Fibroblasts produce extra-cellular matrix (ECM) molecules that regulatetissue strength and resilience. Imbalanced ECMmaintenance leads to tissue dysfunction. Al-though multiple populations of fibroblastssupport uninjured skin function, the extent offibroblast diversity and the presence of func-tionally distinct subsets in adult fibrotic skinare poorly defined. The inability to find a singlemolecular marker that identifies all activated“myofibroblasts” during wound healing sug-gests the existence ofmultiple subsets of ECM-producing myofibroblasts. Furthermore, littleis known about the cellular and molecularmechanisms that promote the expansion of
individual myofibroblast subsets and supportcellular heterogeneity. Although macrophagesinfluence myofibroblast numbers and ECM de-position after injury, subpopulations of woundbed macrophages have only recently been de-fined, and specific interactions with fibroblastshave not been explored.
RATIONALE: Variation inhealing and scarringrates in multiple tissues suggests that fibro-blastdiversity exists. Todevelop therapies target-ing fibrotic responses, functionally distinctsubsets ofmyofibroblasts and themechanismsthat support individual populations must beuncovered. We used a comprehensive, hierar-
chical fluorescence-activated cell sorting strat-egy to define myofibroblast subsets in skinwound beds from adult mice. This strategyrevealed distinct subsets of wound bed myofi-broblasts, including an abundant populationthat contains the cell surface marker profile ofadipocyte precursor cells (APs). We examinedmyofibroblast subsets in different cutaneousfibrotic contexts and explored mechanismsthat selectively promote the expansion of APs.
RESULTS: Genetic lineage tracing and flowcytometry revealed distinct subsets of woundbed myofibroblasts that express smooth muscleactin and collagen. The most abundant pop-ulations were CD26-expressing APs and a sub-setwithhigh cell surface levels of CD29 (CD29High).Although transcriptomic analysis revealed thateach myofibroblast subset has a distinct geneexpression profile, functional analyses suggestthat myofibroblast subsets make both overlap-
ping anddistinct contribu-tions to repair. APs weresignificantly reduced andCD29High cells were moreabundant in wound bedsfrom aged mice and skinfrommice that underwent
bleomycin-induced fibrosis, suggesting that thefibrotic environment influences myofibroblastcomposition. Injury and repair-related changesin AP transcription implicated macrophagesignaling in the modulation of AP gene ex-pression. Genetic ablation and cell transplan-tations of different myeloid cells revealed thatmacrophages expressing macrophage galac-tose N-acetylgalactosamine–specific lectin 2(Mgl2/CD301b) directly stimulate proliferationin a subset ofAPs andnot in othermyofibroblastsubsets. By combining in vitro cytokine stimula-tion with in vivo signaling pathway inhibition,we identified multiple CD301b+ macrophage–secreted factors (platelet-derived growth factorCand insulin-like growth factor 1) that selectivelystimulate AP proliferation, thus supporting theheterogeneity of wound bed myofibroblasts.
CONCLUSION: We identified multiple popu-lations of skinmyofibroblasts and observed thatthe composition of myofibroblasts is dependentupon the fibrotic environment. Distinct inter-actions allow CD301b+macrophage–derived sig-naling to selectively activate the proliferation ofAPs and not other myofibroblasts. These resultshave potential for the development of therapiesthat target multiple cellular populations or sig-naling pathways under conditions associatedwith excessive or deficient ECM deposition,such as wound healing and fibrosis.▪
RESEARCH
Shook et al., Science 362, 909 (2018) 23 November 2018 1 of 1
The list of author affiliations is available in the full article online.*Corresponding author. Email: [email protected] (B.A.S.);[email protected] (V.H.)Cite this article as B. A. Shook et al., Science 362, eaar2971(2018). DOI: 10.1126/science.aar2971
CD29High
Adipocyte precursor
Adipocyte
CD29Low
Agin
g
Youn
g ad
ult
Unin
jure
d sk
inSelective activation of adipocyte precursor proliferation
Scab
Wound
Activated fibroblast
(myofibroblast)
Fibroblast
CD301b+ Mφ
Regulation of myofibroblast diversity in skin wounds. After injury, multiple subsets offibroblasts become activated myofibroblasts that contribute to tissue repair and scarformation. Wound bed macrophages expressing CD301b selectively activate proliferation inAPs and not other myofibroblasts. With age, impaired healing is associated with a reduction inCD301b+ macrophages (Mf) and APs. These findings identify distinct cellular and molecularinteractions that support myofibroblast heterogeneity.
ON OUR WEBSITE◥
Read the full articleat http://dx.doi.org/10.1126/science.aar2971..................................................
on Septem
ber 12, 2020
http://science.sciencemag.org/
Dow
nloaded from

RESEARCH ARTICLE◥
SKIN REPAIR
Myofibroblast proliferation andheterogeneity are supported bymacrophages during skin repairBrett A. Shook1*, Renee R.Wasko1, Guillermo C. Rivera-Gonzalez1, Emilio Salazar-Gatzimas2,Francesc López-Giráldez3, Biraja C. Dash4, Andrés R. Muñoz-Rojas5, Krystal D. Aultman1,Rachel K. Zwick1, Vivian Lei1, Jack L. Arbiser6, KathrynMiller-Jensen1,5, Damon A. Clark2,Henry C. Hsia4, Valerie Horsley1,7*
During tissue repair, myofibroblasts produce extracellular matrix (ECM) molecules for tissueresilience and strength. Altered ECM deposition can lead to tissue dysfunction and disease.Identification of distinct myofibroblast subsets is necessary to develop treatments for thesedisorders.We analyzed profibrotic cells duringmouse skin wound healing, fibrosis, and agingand identified distinct subpopulations of myofibroblasts, including adipocyte precursors(APs). Multiple mouse models and transplantation assays demonstrate that proliferation ofAPs but not other myofibroblasts is activated by CD301b-expressing macrophages throughinsulin-like growth factor 1 and platelet-derived growth factor C.With age, wound bed APsand differential gene expression between myofibroblast subsets are reduced. Our findingsidentify multiple fibrotic cell populations and suggest that the environment dictatesfunctional myofibroblast heterogeneity, which is driven by fibroblast-immune interactionsafter wounding.
Tissues sustain resilience and strength throughthe maintenance of extracellular matrix(ECM) molecules by mesenchymal cells.Under disease states, profibrotic conditionslead to excessive and disordered ECM de-
position that impairs tissue function (1, 2). Addi-tionally, dysregulated ECM is associated withaged skin and age-related defective wound heal-ing (3–8). Variability in rates of wound healing,scarring, and fibrosismay result from functionally
distinct mesenchymal cells (9, 10). Thus, identi-fying distinct mesenchymal cell populations thatcontribute to fibrosis and the mechanisms thatdrive cellular diversity has substantial implica-tions for disease treatment (2, 11–13).Prior experiments in mice have demonstrated
that embryonic mesenchymal precursors express-ing Engrailed (En1) orDelta-like homolog 1 (Dlk1/Pref1) generate skin fibroblast and adipocytelineages (14–16). During skin repair after injury,myofibroblasts expressing alpha smooth muscleactin (SMA), Pdgfra, Sca1, Itga8, CD34, andDpp4(CD26) migrate, proliferate, and deposit ECM(17–19). Myofibroblasts do not form lipid-filledadipocytes within regenerated tissue after a stan-dard small skin injury (14, 15, 20, 21) but can formadipocytes that regenerate hair follicles in largewounds (20). How environmental conditions al-ter functional cellular diversity and the contribu-tion ofmesenchymal subsets to tissue fibrosis arenot well understood.In this study, we uncover unappreciated het-
erogeneitywithinwound bedmyofibroblasts thatis dependent on the tissue environment. In par-ticular, we show that the predominant popula-tion of myofibroblasts is adipocyte precursor cells
RESEARCH
Shook et al., Science 362, eaar2971 (2018) 23 November 2018 1 of 8
1Molecular, Cellular and Developmental Biology, YaleUniversity, New Haven, CT 06511, USA. 2InterdepartmentalNeuroscience Program, Yale University, New Haven, CT06511, USA. 3Yale Center for Genome Analysis, Yale Schoolof Medicine, New Haven, CT 06510, USA. 4Department ofSurgery (Plastic), Yale School of Medicine, New Haven, CT06510, USA. 5Department of Biomedical Engineering, YaleUniversity, New Haven, CT 06511, USA. 6Department ofDermatology, Atlanta Veterans Administration Health Center,Emory University, Atlanta, GA 30322, USA. 7Department ofDermatology, Yale University, New Haven, CT 06511, USA.*Corresponding author. Email: [email protected] (B.A.S.);[email protected] (V.H.)
A C
B
1acS
102
103
104
105
102 104 102 10443D
C
102
103
104
105
Per
cent
of c
ells
CD
34+
CD
29H
igh
AP
CD
29Lo
w
CD34+AP
SMA
D
E
F
102 1040
20406080
100
5d WB
5d WB
Collagen I
% o
f Max
Singlets;Sytox-;Lin-(CD45-;CD31-)
102 104
5d WB
43D
C 102
103104105
102 104
NW
Non-wounded
CD29LowCD29High
CD29LowCD29High
CD29
Per
cent
SM
A+
Rel
ativ
e ge
ne e
xpre
ssio
nR
elat
ive
gene
exp
ress
ion
Col IActa2 (SMA)
Col IActa2 (SMA)
102 104
CD90P
erce
nt C
D90
+
0
50
100
0
50
100
Per
cent
Col
I+
APCD34+
0
50
100102 104
CD29
102
103
104
105
102 104 102 104
CD26
Singlets;Sytox-;Lin-
102
103
104
105
Per
cent
of c
ells
Dlk
1 Cre
ER
trac
edE
n1C
re tr
aced
0
50
1000
50
100
Wound bed myofibroblastmolecular markers
EmbryonicLineage tracing model
SMA, Sca1, Pdgfra, Itga8Dlk1CreERDriskell et al., 2013
SMA, Sca1, Pdgfra, CD26,CD34, Itga8
En1CreRinkevich et al., 2015
Mesenchymal cell lineage tracing
dwat
pc
0
4
8
12
05
10154080
012345
0.0
0.5
1.0
1.5
(rel
ativ
e to
NW
cell
popu
latio
n)(r
elat
ive
to N
W s
kin)
NW
5d W
B7d
WB
2w W
B
020406080
100
% o
f Lin
- Cel
ls
epidermisscab
woundbed
papillaryand reticular
dermis
Fig. 1. Dermal mesenchymal cell heterogeneity changes after injury. (A) Molecular markers ofwound bed myofibroblasts identified by using genetic lineage tracing (14, 15). (B) FACS analysisand quantification of CD34 andCD29 subsets of SCA1+;CD26High lineage-traced cells in nonwoundedskin (n = 4 samples). (C) FACS plots and quantification of cellular subsets in nonwounded skin(n = 8 samples) and 5-day (n = 19 samples), 7-day (n = 4 samples), and 14-day (n = 7 samples)wound beds. (D and E) Real-time quantitative polymerase chain reaction analysis of SMA(Acta2) and Col I (Col I) in mesenchymal subsets isolated from nonwounded skin (D) or 5-daywound beds (E). (F) Representative flow cytometry histograms and quantification of SMA, CD90,and Col I in mesenchymal subsets (n = 3 samples). Error bars indicate the mean ± SEM. NW,nonwounded;WB, wound bed; pc, panniculus carnosus; dwat, dermal white adipose tissue.
on Septem
ber 12, 2020
http://science.sciencemag.org/
Dow
nloaded from

(APs) derived from En1 lineage–traced fibroblasts(14, 15, 21, 22), which contribute to tissue repairand ECM production and modulation. We showthat, in wound beds of agedmice, APs aremarked-ly reduced and wound bed myofibroblast sub-populations become more homogeneous in theirgene expression profiles and localization. Ourdata indicate that CD301b+ macrophage–derivedplatelet-derived growth factor C (PDGFC) andinsulin-like growth factor (IGF) signaling con-tributes to myofibroblast heterogeneity by selec-tively promoting the proliferation of wound bedAPs and not other myofibroblast subsets. Thesefindings define major subsets of wound bed myo-fibroblasts and identify immune andmolecularinteractions that promote functional cellular het-erogeneity under distinct fibrotic conditions.
Mesenchymal cell heterogeneity underfibrotic conditions
Myofibroblasts within wound beds express PDGFreceptor a (PDGFRa), CD34, and SCA1 and de-rive from embryonic precursors that expressEn1 or Dlk1/Pref1 (14–16) (14, 15, 23, 24) (Fig. 1A).Because PDGFRa, CD34, and SCA1 define APs(SCA1+;CD34+;CD29+) (25, 26), we sought to deter-mine whether APs were derived from En1- andDlk1-expressing precursors. We confirmed thatEn1Cre;Rosa26-LSL-tdTomato– and Dlk1CreER;mT/mG–traced cells expressed CD26 and SCA1(14, 15) (Fig. 1B). Ninety-six percent of tdTomato+;SCA1+;CD26+ cells in En1Cre;tdTomato and 97%of GFP+;SCA1+;CD26+ cells in Dlk1CreER;mTmGnonwounded skin express APmarkers CD29 andCD34 (Fig. 1B and fig. S1).Flow cytometry analysis of immune and en-
dothelial lineage–negative cells (Lin−) isolatedfrom uninjured dermis and 5-day wound bedsrevealed four populations of cells: CD29+;CD34+,CD34+, CD29High (cells with high surface levelsof CD29), and CD29Low cells (Fig. 1C). We defineCD29+;CD34+ cells as APs because 90% retainSCA1+ expression after injury and have adipo-genic potential in vitro and in vivo (26–28) (fig.S2A). Although AP and CD29Low cells were themost abundant populations in nonwounded skin,wound beds contained increased proportions ofCD29High cells (Fig. 1C).To determine which populations were myofi-
broblasts, we analyzed the expression of SMAand collagen I (Col I). Within wound beds, eachcell population up-regulated SMAandCol ImRNAexpression compared with that in cell popula-tions from uninjured skin (Fig. 1, D and E); how-ever, flow cytometry revealed that only APs andCD29High cells were enriched for SMA, Col I,and the fibroblast marker CD90 in wound beds(Fig. 1F).To further analyze the fibrotic nature of these
cell populations, we examined the expressionof profibrotic proteins SCA1, CD9, CD26, andPDGFRa (14–16, 22, 29) (figs. S1B and S2, A to D).A greater percentage of APs and CD29High andCD29Low cells expressed CD9 after injury (fig. S2D),and CD9+ APs have decreased in vitro adipogenicpotential compared with CD9− APs (fig. S2, Eand F). These data suggest that fibrotic cells are
heterogeneous and distinct between uninjuredand injured skin. Further, at least two major pop-ulations of fibrotic mesenchymal cells exist inskin wounds: APs and CD29High cells.We next examined CD29 and CD34 popula-
tions in bleomycin-induced fibrosis. CD29High
cells increased more robustly and fewer APswere observed after bleomycin treatment than inthe context of wound healing (fig. S2, G and H).Colocalization with other profibrotic markerswas not markedly changed (fig. S2, B to D). Thus,the profibrotic cellular composition in bleomycin-
treated skin is distinct from that in wound heal-ing, suggesting that distinct strategies are requiredto treat tissue fibrosis under different pathologicalconditions.Because SMA andCD9 expression increased in
multiple populations of mesenchymal cells with-in skin wounds, we sought to design a compre-hensive hierarchical marker panel to delineatemesenchymal heterogeneity in skin wounds byusing six fibrotic markers (Fig. 2, A and B, andfig. S1). Lin− cells were subdivided on the basisof PDGFRa and SCA1 (14, 15). Whereas PDGF
Shook et al., Science 362, eaar2971 (2018) 23 November 2018 2 of 8
0
20
40
60 *
****
Per
cent
Col
I+P
erce
nt S
MA
+
-
Hig
h
Low
+-
Hig
h
Low
+
NW 5d NW 5d NW 5d NW 5d
-
Hig
h
Low
+-
Hig
h
Low
+
NW 5d NW 5d NW 5d NW 5d
-
Hig
h
Low
+-
Hig
h
Low
+
NW 5d NW 5d NW 5d NW 5d
020406080
100
020406080
100
0
50
100
020406080
100
020406080
100
***
***
**
** **
****
****
********
****
020406080
100
******
*****
D
Singlets; Sytox-; Lin-
1acS
102103104105
102 104
A C
B
Non-wounded
5d wound bed
2 GFP+;Sca1-
102103104105
102 104
3 GFP-;Sca1-
102103104105
0
0
102 104
1 GFP+;Sca1+
102103104105
102 104
43D
C
Pdgfra-GFP
43D
C
102103104105
102 104
1 GFP+;Sca1+
CD29
1
102103104105
102 104
Pdgfra-GFP
Singlets; Sytox-; Lin-
1acS
1
23
CD29
43D
C
43D
C
CD29CD29
CD29LowAP CD29High
Per
cent
of L
in-C
ells
CD26:CD9:
E
048
123060
**
****
01020
4060
DAPICD29CD45 CD26
F
dwat
pc
epidermis scab
papillaryand reticular
dermis
Enriched in CD29High cells (CD26Low;CD29High)
Enriched with APs (CD26High;CD29+)2. Signal LUTs
Wound midline
normalized distance - superficial to deep
Sig
nal I
nten
sity
Rat
io(c
d26)
/(cd
26+
cd29
)Distribution of signal
normalized distance - midline to edge
1. Bisect wound bed at midlineand subtract CD45+ pixels
4. Graphic representation 3. Average
signal intensity
mask of CD29 and CD26
0 0.2 0.4 0.6 0.8 10.40.60.8
0 0.2 0.4 0.6 0.8 10.50.60.7
064128191255 AU
CD26 signal intensityCD29 signal intensity
Fig. 2. Skin wounds contain multiple myofibroblast subsets. (A and B) FACS plots detailing thegating strategy to definemesenchymal subpopulations. (C to E) Quantification of the relative abundancesof prevalent profibrotic subsets (n = 6 samples) (C) and colocalization with SMA (D) and Col I (E) innonwounded skin and 5-daywound bedmesenchymal subsets (n=3 samples). (F) Pipeline for processingimmunostained tissue sections to infer the locations of APs (CD29+;CD26High) and CD29High cells inday 5 wound beds.Yellow lines delineate wound edges. Scale bar, 250 mm. Error bars indicate the mean ±SEM. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001. AU, arbitrary units; LUT, look-up table.
RESEARCH | RESEARCH ARTICLEon S
eptember 12, 2020
http://science.sciencem
ag.org/D
ownloaded from

signaling is central to fibrosis (2, 21, 30–32), weincluded PDGFRa− cells in our analysis becausePDGFRa− profibrotic cells may also contribute torepair (14, 33). We further subdivided popula-tions on the basis of CD29 and CD34 expressionand then by the presence of CD26 (high or low)and CD9 (CD9+ or CD9−). This analysis revealedthat 54% of Lin− cells in nonwounded skin con-tained surface markers that prospectively iden-tify APs: PDGFRa+;SCA1+;CD29+;CD34+ (Fig. 2Aand figs. S2A and S3A). Within the AP pool,66%were CD26High;CD9− cells (Fig. 2, A to C, andfig. S4, A and B). The only other nonimmune ornonvascular populations that contribute morethan 3% of total cells in nonwounded skin werePDGFRa−;SCA1−;CD29High (CD29High) cells (~7%)and PDGFRa−; SCA1−;CD29Low (CD29Low) cells(~15%) (Fig. 2, A to C, and figs. S2A and S3A).In 5-day wound beds, the relative abundance
of APs decreased as PDGFRa+;CD29High cellsand CD29Low cells became more abundant (Fig.2, A to C, and figs. S3 and S4, A and B). BecauseCD34+ cells were mostly negative for SMA (Fig.1F) and not abundant during repair, we excludedthem from further analysis. CD29High andCD29Low
cells remained predominantly CD26Low;CD9+ andCD26Low;CD9−, respectively (Fig. 2, A to C, andfig. S4, A and B). Forty-six percent of APs wereCD26High;CD9+ cells in 5-day wound beds, com-pared with 24% in nonwounded skin (Fig. 2, A toC, and fig. S4, A and B). The shift from CD9− toCD9+ APs persisted 14 days after injury (fig. S3E).Immunofluorescent staining of fluorescence-activated cell sorting (FACS)–isolated cells andwound beds showed greater intensity of CD26staining in APs than in other cells, confirming
our flow cytometry results (fig. S4, C and D). SMAand Col I were expressed by APs and CD29High
cells in wound beds regardless of the cells’ CD26or CD9 expression; however, few CD29Low cellsexpressed SMA, Col I, CD90, or ER-TR7 (Figs. 1,D to F, and 2, D and E, and fig. S4, C to H).To explore the spatial organization of myofi-
broblast subsets, we regionally dissected woundbeds for flow cytometry and examined the sig-nal intensity of CD26 and CD29 in nonimmune(CD45−) cells in tissue sections. Although APsare found throughout the wound bed, CD29High
cells were biased toward the most superficialregion of the outer wound bed edge (Fig. 2F andfig. S5). CD29High cells were more abundant inthe upper dermis of nonwounded skin, whichcontributes to the superficial regenerating der-mis (15). Human skin has a composition of me-senchymal cells similar to that of mouse skin.However, greater percentages of these popula-tions were CD26High and CD9+ cells (fig. S6), in-dicating that human skin may be more biasedtoward fibrotic responses.To determine whether repair-related myofi-
broblast heterogeneity arises from conversionbetween cellular subsets, we performed geneticlineage tracing by using inducible Cre-lox mouselines that label profibrotic cells: Dlk1CreER;mT/mG (postnatal labeling) and PdgfraCreER;mT/mG (adult labeling) (fig. S7A) (15). In uninjuredskin, several mesenchymal populations were la-beled in Dlk1CreER mice, whereas PdgfraCreERmice predominantly labeled APs (~94%) (fig. S7, Band C). PdgfraCreER lineage–traced cells in 5-daywound beds contributed to APs, SCA1−;CD29Low
cells, and a rare population of SCA1+;CD29High
cells (fig. S7, C to F). SCA1+;CD29High cells areCD9+ during repair and similar in size to APs(fig. S7, E and F), suggesting that they are pro-fibrotic. Two weeks after injury, PdgfraCreERlineage–traced cells comprised ~80% APs, ~10%SCA1+;CD29High cells, and 7% SCA1−;CD29Low
cells (fig. S7D). These data suggest that APs con-tribute to multiple myofibroblast subpopulations;however, they do not contribute to the expan-sion of SCA1−;CD29High cells. Yet, because non-AP,CD29+ cells are labeled in PdgfraCreER mice, wecannot rule out the possibility that the prolifera-tion of CD29+ cells (fig. S12) also contributes tomyofibroblast heterogeneity after injury.
Myofibroblast subsets have distinctgene expression profiles
The comparison of transcriptional profiles ofCD29Low and CD29High cells and APs that wereeither CD9 positive or negative by RNA sequenc-ing (RNA-seq) (n = 2 profiles for each population)confirmed significant diversity amongwound bedmyofibroblasts (Fig. 3, A and B). Although tran-scriptomic analysis revealed that CD9+ and CD9−
APs were similar, each mesenchymal subset ex-pressed distinct mRNAs (figs. S8 and S9 andtable S1). For instance, CD29High cells had ele-vated expression of Pdgfrb, CD146, NG2, andother perivascular cell or pericyte markers (34)compared with APs and CD29Low cells and hadelevated Acta2 expression in nonwounded skin(Fig. 1D).Although transcriptomeswere distinct between
cellular subsets, Ingenuity Pathway Analysis(IPA) predicted common active biofunctionsand similar upstream activators of gene expression
Shook et al., Science 362, eaar2971 (2018) 23 November 2018 3 of 8
AP - CD9+
AP - CD9-
B
C
D E F
A
-20 0 20
50
-5-1
0P
C2
(10.
3%)
PC1 (77.2%)
CD29High
CD29Low
Extracellularmolecules
Ccl2, Ccl7, Ccl11, Csf1, Cxcl1, Cxcl2, Cxcl10,Cxcl12, Cxcl13, Cxcl14, Figf, Gas6, Il6, Il33,Nov, Ogn, Sfrp4, Timp1, Vasn, Wisp2, Wnt2
Ccl3l3, Ccl6, Gdf15, Hbegf, Inhba, Il1A, Il1B,Il1rn, Pdgfa, Pf4 (Cxcl4), Spp1 (Opn), Tgfb2,Tnf, Vegfc, Wnt5A
Receptors Adora2b, Cxcr7*, Dpp4 (Cd26), Il6st*, Il11ra*,Il13ra1*, Lpar1*, Osmr1*, Pdgfra*, Sfrp2,Sfrp4, Tgfbr2, Tgfbr3
Ccr2, Endra, Endrb, Fgfr4*, Il2rg, Il10ra,Itgam, Itgb2*, Pdgfrb
Adamts1, Adamts2, Adamts4, Adamts5,Col5a2, Col14a1, Fbln1, Fbn1, Has1, Loxl1,Mmp2, Mmp3, Mmp23, Mmp27, Plod2, Podn,Timp3
ECMcomponents/
regulators
Adamtsl2, Col6a3, Col7a1, Col12a1, Mmp13,Tnc
Enriched in APs vs CD29High Enriched in CD29High vs APs
148
1100
695
504
1305
939
AP - CD9
-
CD29High
CD29Lo
w
CD29High
AP - CD9+
AP - CD9-
0
500
1000
1500
# Significant differentiallyexpressed genes at alpha 0.05
* Predicted upstream activators of unique gene expression profiles
LOX activity
CD
29H
igh
AP
-CD
9-
AP
-CD
9+
AP
CD
29H
igh
0200400600800
Mig
ratio
n di
stan
ce (μ
m)
AP
CD
29H
*
0
500
1000
1500
2000
2500
Rel
ativ
e flu
ores
cenc
e
Hyd
roxy
prol
ine
(μg/
106 c
ells
)
0
1
2
3
4
wound explantmigration assay
bisectedwound bed
3d migrationdistance
CD45 CD26 CD29**
**
Collagen production Collagen crosslinking
Fig. 3. Myofibroblast subsets can distinctively regulate repair. (A) Transcriptomic principal components analysis of myofibroblast subsets. (B) Table ofgenes with statistically significant differences in expression between cellular subsets. (C) Wound healing–related genes enriched in APs (CD9− and CD9+
AP populations) or CD29High cells. (D and E) Quantification of hydroxyproline content (n = 7 samples) (D) and lysyl oxidase (LOX) activity (n = 4 samples;*P = 0.0416) (E) in cells from day 5 wounds. (F) Migration distance of APs (CD26High) (asterisks) and CD29High cells (arrowheads) from cultured wound beds(n ≥ 250 cells from 3 wound beds). Scale bar, 10 mm. Error bars indicate the mean ± SEM.
RESEARCH | RESEARCH ARTICLEon S
eptember 12, 2020
http://science.sciencem
ag.org/D
ownloaded from

profiles (figs. S10 and S11 and table S2), suggestingsome functional redundancy among myofibro-blasts. However, many genes differentially ex-pressed between APs and CD29High cells havebeen implicated in wound healing (Fig. 3C andfigs. S8 and S9). Additionally, each myofibro-blast population was enriched for different ECMcomponents and modifiers (Fig. 3C and figs. S8and S9). Both CD9+ and CD9−APs were enrichedfor many cytokine genes (Ccl2, Cxcl1, Cxcl10, andCxcl12) and ECM components (Col5a2, Fbln1,Fbn1, Has1, and Loxl1) that promote rapid ECMdeposition (35–37). The enrichment of genes in-volved in repair and fibrosis changed amongmyofibroblast populations between day 5 andday 14 of repair (fig. S8B), indicating that myo-fibroblast subsets can distinctively influenceboth the proliferative and maturation phases oftissue repair.To determine whether myofibroblast subpop-
ulations were functionally distinct, we examinedcollagen production, collagen cross-linking, andthe migration of APs and CD29High cells. We didnot observe differences in collagen productionor cellular migration; however, we detected anincreased ability of APs to cross-link collagencompared with CD29High cells, consistent withelevated Lox expression in APs (Fig. 3, C to F,and fig. S8A).Because myofibroblast numbers increase after
injury (Fig. 1C and fig. S3), we examined in vivoproliferation within the different mesenchymalsubsets (APs and CD29High and CD29Low cells)during tissue repair. Proliferation increased inAPs and CD29High and CD29Low cells after injury,and CD26Low cells were more proliferative thanCD26High cells within each cellular subset (fig.S12, A to D). Taken together, these data demon-strate that the dermis contains tremendous het-erogeneity within profibrotic cells, which havedistinct functions during tissue repair.
Myofibroblast composition and geneexpression are altered during aging
Age-related defects in repair are associated withreduced myofibroblasts and dysfunctional ECMdeposition (3–6) (fig. S13, A and B). To determinewhether mesenchymal populations were alteredwith age, we analyzed 5-day wound beds in youngand aged mice. The relative abundance of APsdecreased and CD29High cells increased in woundbeds from agedmice (Fig. 4, A to D), with reducedpercentages of CD9+ cells in all mesenchymalpopulations (Fig. 4C), suggesting that fibrotic cellsare lost or unstimulated with age.Analysis of transcriptional changes in myofi-
broblasts during aging by RNA-seq (n = 2 pro-files) revealed fewer differentially expressed genesbetween myofibroblast subsets (fig. S14, A to C)because of age-related down-regulation of manygenes within individual populations (fig. S14D).Comparing the transcriptomes of myofibroblastpopulations in young versus aged mice revealedage-related changes in gene expression of extra-cellular molecules (Fig. 4E) and increased expres-sion ofmultiplemetalloproteases inmyofibroblasts,consistent with the ability of aged fibroblasts to
break down ECM faster than young fibroblastsand impair healing (4, 6).
APs become fibrotic after injury
To identifymolecularmechanisms regulating APmyofibroblasts during repair, we isolated APs
from uninjured skin and 5-day wound beds andperformed RNA-seq (n = 2 profiles). Injury andrepair up-regulated Acta2 (SMA) and severalsecreted factors implicated in tissue repair (fig.S15, A to C). Several adipogenic genes and in vitroadipogenic potential were reduced in wound bed
Shook et al., Science 362, eaar2971 (2018) 23 November 2018 4 of 8
B
D
C
A
AP - CD9+ Extracellular Molecules
Adamts1, Ccl6, Cgref1, Clec3b, Col4a1, Col4a2,Col8a1, Col12a1, Cpz, Crlf1, Nov, Pamr1, Thbs2,Vcan
CD29High Extracellular Molecules
Mmp3, Mmp12, Thbs4
102
103
104
105
102 104
Young
43D
C
CD29
Singlets;Sytox-;Lin-
102
103
104
105
102 104
Aged
CD29LowAP CD29High
Per
cent
of L
in- C
ells
CD26:- +-+CD9:
Hig
h
Low
Hig
h
Low
***
****
Youn
gA
ged
Youn
gA
ged
Youn
gA
ged
Youn
gA
ged
- +-+
Hig
h
Low
Hig
h
Low
Youn
gA
ged
Youn
gA
ged
Youn
gA
ged
Youn
gA
ged
- +-+
Hig
h
Low
Hig
h
Low
Youn
gA
ged
Youn
gA
ged
Youn
gA
ged
Youn
gA
ged
****
- +-+
Hig
h
Low
Hig
h
LowCD26:
CD9:
CD29LowCD29HighAP
Youn
gA
ged
AP - CD9- Extracellular Molecules
Adamts1, Ccl6, Cgref1, Clec3b, Col4a1, Dmkn,Eln, Fgl2, Fbln5, Igfbp5, Itgbl1, Mmp12, Nov,Opn, Pamr1, Plxdc2, Vcan
CD29Low Extracellular Molecules
Apod, Ccl3l3, Ccl4, Csf1, Cxcl3, Il1A, Il1rn, Il23a,Il36g, Mmp9, Mmp12, Mmp13, Nov, Prelp,Thbs4, Tnf, Tpsab1/Tpsb2,
Genes Differentially Expressed With Age
Enriched in Young Enriched in Aged
E
**
05
10152025
0
10
20
30
010203040 ****
DAPICD45 CD29CD26
dwatpc
epidermis
scab papillaryand reticular
dermis
Enriched in CD29High cells (CD26Low;CD29High)
Enriched with APs (CD26High;CD29+)
2. Signal LUTs
mask of CD29and CD26
064128191255 AU
CD26 signal intensityCD29 signal intensity
0 0.2 0.4 0.6 0.8 1normalized distance - superficial to deep
0.40.6
Sig
nal I
nten
sity
Rat
io(c
d26)
/(cd
26+
cd29
)
Distribution of signal
0 0.2 0.4 0.6 0.8 1normalized distance - midline to edge
0.40.50.6
1. Bisect wound bed at midlineand subtract CD45+ pixels
4. Graphic representation
3. Average signal intensity
Wound midline
Fig. 4. Myofibroblast composition and gene expression is altered during aging. (A) FACSplots for 5-day wounds from young and aged mice. (B) Quantification of the relative abundance ofprevalent profibrotic subsets in 5-day wounds (n = 4 samples). (C) Pie charts depicting CD9 andCD26 colocalization. (D) Pipeline for processing immunostained wound bed sections to infer APand CD29High cell locations in day 5 wound beds from aged mice. Yellow lines delineate wound edges.Scale bar, 250 mm. (E) Genes with altered differential expression with age. Black and red textindicates enrichment in young and aged mice, respectively. Error bars indicate the mean ± SEM.**P < 0.01, ***P < 0.001, ****P < 0.0001.
RESEARCH | RESEARCH ARTICLEon S
eptember 12, 2020
http://science.sciencem
ag.org/D
ownloaded from

APs (fig. S15, D and E). Thus, APs displayed dra-matic alterations within the wound environmentthat limit their adipogenic potential, promotemyofibroblast gene expression, and may explainthe myofibroblast origin of adipocytes in largewounds (20).
Macrophage signaling selectivelyactivates proliferation of wound bed APs
Because delayed healing in aged mice is associ-ated with decreased APs and APs rapidly increasefrom days 3 to 7 after injury, when new dermal
tissue is generated, we investigated potential sig-naling pathways that could affect AP numbersduring repair. IPA predicted that injury-relatedchanges in AP gene expression could result frommonocyte-macrophage–derived ligands (fig. S15F).Macrophage ablation reduces wound bed myofi-broblast numbers, impairsmyofibroblast function,and impairs wound healing (38–41); however,the underlying mechanisms are ill defined. Toexamine the contribution ofmonocytes andmac-rophages to myofibroblast heterogeneity, weablated macrophages by using LysMCre;iDTR
mice (38, 41–43) (Fig. 5A). Ablating monocytesandmacrophages reduced all AP subsets (fig. S16)and diminished AP proliferation in wounds with-out significantly changing CD29High or CD29Low
populations (Fig. 5B and fig. S16A). Additionally,pharmacological reduction of macrophages de-creased the percentage of dividing APs from 25to 9% in controls (fig. S17), indicating that themyeloid lineage in 5-day wound beds selectivelyactivates the proliferation of APs and not othermyofibroblasts.
CD301b+ macrophages activate APproliferation during wound healing
During the mid-phase of wound healing (days 3to 7), the myeloid lineage is composed predom-inantly of monocyte and macrophage subsets(41, 44, 45) (fig. S18A). We have previously shownthat wound bed macrophages expressing mac-rophage galactose-type C-type lectin 2 (Mgl2/CD301b) contribute to repair by promoting pro-liferation and fibroblast repopulation and thatCD301b+ macrophages are ablated in Mgl2DTR
mice (41, 46, 47). Whereas ablating all macro-phages in LysMCre;iDTR mice decreased theproliferation of all subsets of APs, ablating CD301b-expressing macrophages reduced the prolifer-ation of CD26Low APs in 5-day wounds, with nochange in CD29High or CD29Low cell proliferation(Fig. 5C). This suggests that diversity in woundbed macrophages (48) allows the proliferationof different AP subsets to be differentially reg-ulated, thus promoting myofibroblast heteroge-neity. ReducedAPproliferation indiphtheria toxin(DT)–treatedMgl2DTR mice resulted in a ~50%reduction in 5-ethynyl-2′-deoxyuridine–positive(EdU+) APs in 7-day wound beds that was ob-served only when newly generated CD301b+
macrophages were ablated during the prolifera-tive phase of repair (Fig. 5D). Consistent with thismodel, cell transplantation of CD301b+ macro-phages, and not that of other immune cells, in-creasedAPproliferation (from 18 to 28%),whereasCD29High and CD29Low cells were unaltered (Fig. 5,E and F). Further, cultured CD301b+ macrophagesdoubled AP proliferation in vitro (Fig. 5G), dem-onstrating direct signaling between CD301b+
macrophages and APs.To identify signaling molecules that activate
AP proliferation during repair, we compared thetranscriptomes of CD301b+ macrophages withthose of F4/80− immune cells isolated from day5 wounds (Fig. 6A and fig. S18, B and C) (n = 2profiles per group).We identified ligands enrichedin CD301b+ macrophages that bind to receptorson APs (Fig. 6B and fig. S18D) and validated theseresults by quantifying protein secretion (fig.S18E). Cultured APs were treated with candidatemolecules, and only PDGFC and IGF1 inducedproliferation (Fig. 6C). To determine whetherPDGFC and IGF1 signaling pathways contrib-ute to AP proliferation in vivo, we administeredligand-neutralizing antibodies or receptor antag-onists after injury (Fig. 6D). Local injection ofPDGFC- or IGF1-neutralizing antibodies in vivoreduced AP proliferation; however, no change inthe proliferation of other cells was detectable.
Shook et al., Science 362, eaar2971 (2018) 23 November 2018 5 of 8
Fig. 5. CD301b+ macrophages selectively stimulate AP proliferation during wound healing.(A) Quantification of wound bed macrophage depletion (n ≥ 3 samples; P = 0.004). (B) Quantificationof myofibroblast proliferation in wound beds (n = 4 samples). (C) Quantification of cell proliferationin wound beds of CD301b+ macrophage–depleted mice (Mgl2DTR) (n = 3 samples). (D) Quantificationof EdU-incorporating APs in mice receiving DTon days 2, 4, and 6 after injury (left) (n = 3 samples,P = 0.0469) and DTon days 2 and 3 or 3, 4, and 6 after injury (right) (n = 3 samples, P = 0.0116).Mice were given two injections of EdU per day from days 3 through 7 after injury. (E) FACS plotsof immune cell populations isolated for transplants. (F and G) Quantification of EdU-incorporatingcells after injection of select immune cell subsets in vivo (n = 5 samples, P = 0.0146) (F) or Transwellcoculture (n = 6 samples, P = 0.001) (G). Error bars indicate the mean ± SEM. *P < 0.05,**P < 0.01, ***P < 0.001. IV DT, intravenous DT; Mo/Mf, monocyte-macrophage.
RESEARCH | RESEARCH ARTICLEon S
eptember 12, 2020
http://science.sciencem
ag.org/D
ownloaded from

Additionally, the inhibition of PDGFRa and IGF1receptor (IGF1R) or downstream phosphatidyl-inositol 3-kinase (PI 3-kinase) signaling selec-tively reduced AP proliferation (Fig. 6D). Wedid not observe spatial biasing of CD301b+ ma-crophages in wound beds (Fig. 6E), and theexpression of wound healing–associated geneschanged minimally in myofibroblasts from 5-daywound beds ofMgl2DTRmice relative to controls(fig. S19). These data suggest that the distinctgene expression profile of each myofibroblastsubset results from interactions with other tis-sue resident cells, such as keratinocytes (49).As a result, the delayed re-epithelialization andrevascularization observed in Mgl2DTR mice(41) may result from CD301b+ macrophages in-teracting with keratinocytes and endothelialcells. IGF1 can stimulate repair, potentially bypromoting the migration and proliferation ofkeratinocytes and fibroblasts (35, 50–53), yetthe contribution of PDGFC to healing has notpreviously been explored. To examine the con-tribution of PDGFC signaling to wound healing,we locally injected a PDGFC-neutralizing anti-body at the periphery of wound beds and exam-ined skin repair.We did not observe gross changesin re-revascularization and myofibroblast repop-ulation in 5-day wound beds compared withcontrols; however, we observed a slight decreasein re-epithelialization (fig. S20). These data demon-strate that multiple ligands produced by CD301b+
macrophages activate the proliferation of APs,and not other myofibroblast subsets, duringwound healing.Numbers of CD301b+macrophages increase in
wounds as AP abundance increases (41). How-ever, wound beds from aged mice contain fewerCD301b+ cells than those from young controls,and human keloid scars, which have been shownto contain many CD26+ fibroblasts (54), areenriched with CD301+ cells (fig. S21). Thus, theinteraction between CD301b+ macrophages andmesenchymal cells may provide a therapeutic tar-get for fibrosis-related diseases.
Discussion
Dermal cells, including fibroblasts and adipo-cytes, support epidermal functions and integrity(11, 12). Although a common embryonic precur-sor for SMA+ wound bed myofibroblasts exists(14, 15, 20, 33, 55, 56), the diversity of mesenchy-mal cells during adult tissue repair is ill defined.In this study, we discovered that after injury, skinwound beds contain tremendous mesenchymalheterogeneity, similar to what is observed duringlung fibrosis (57). We identified two major classesof SMA+ and Col I+ myofibroblasts that arisefrom different cellular origins: cells with a cellsurface marker profile of APs, and CD29High
cells. During tissue repair, a greater percentageof APs express profibrotic cell surface proteinsCD26High and CD9, with reduced adipogenic po-tential. Spatially, these two myofibroblast popu-lations are distinct, with APs evenly distributedwithin wounds and CD29High cells biased towardsuperficial, outer regions of wound beds. RNA-seq and functional analysis of these myofibro-
blast subsets revealed that each subset has distincttranscriptomes with some functional overlap.With age, the abundance of APs decreases and
CD29High cells are more prevalent, as differentialgene expression between myofibroblast subsetsis reduced. Althoughmyofibroblasts are dynamicafter injury in mouse skin, human dermal fibro-blasts express profibrotic cell surface proteins inuninjured skin, possibly resulting in stronger fi-brotic biasing in humans. These studies illuminatedistinct functional subsets of fibrotic cells, pro-viding a stepping stone to develop therapeutic
strategies that promote efficient wound healingand treat fibrosis.
Regulation of functionalmyofibroblast diversity
In this study, we have shown that CD26High myo-fibroblasts are largely CD34+;CD29+ APs thatfunction asmyofibroblasts in regeneratingmouseskin. Although previous reports did not observethe same degree of CD34+ and CD29+ colocaliza-tion on myofibroblasts (14, 16), these differenceslikely result from changes in fibroblast surface
Shook et al., Science 362, eaar2971 (2018) 23 November 2018 6 of 8
ASytox- Singlets
CD45- CD45+, CD11b+
F4/80- F4/80+, CD206+
CD301b- CD301b+
CD301b+ Macrophage
Factor
AP Receptor
Ccr5
Igf1rPdgfra
Cxcr2Ccl8
Ccr1 Ccl6
Igf1Pdgfc
Cxcl4
RNA sequencing10 1000
1010
00
CD301b+ macrophage expression (FPKM)
F4/
80- i
mm
une
cell
expr
essi
on (
FP
KM
)
518 genes upregulated
613
gene
s up
regu
late
d B
Per
cent
EdU
+ A
Ps
******
***
C
E
0.5%
FBS
+BS
A
10%
FBS
+BS
AC
CL6
10n
gC
CL6
100
ngC
CL8
10n
gC
CL8
100
ngC
XCL4
20n
gC
XCL4
100
ngIG
F1 1
0ng
PDG
FC 1
5ng
0
5
10
15
20
250 3 5
Days Post Wounding
Wound Sacrifice4hr16hr
EdUnAb or
AntagonistD
Per
cent
EdU
+
In vitro stimulation
CD29HighAP CD29Low AP
DM
SO
Cre
nola
nib
Lins
itini
bW
ortm
anni
n0
1
2
3
4
5
IgG
αPD
GF
CαI
GF
10
2
4
6
8
IgG
αPD
GF
CαI
GF
10
5
10
15
IgG
αPD
GF
CαI
GF
10
1
2
3
4
5*
****
***
Neutralizing AntibodyReceptor
Antagonists
CD301b1. Bisect wound bed at midlineand generate intensities (LUT)
0 0.2 0.4 0.6 0.8 1normalized distance - superficial to deep
0
50Distribution of CD301b in Wound Beds
0 0.2 0.4 0.6 0.8 1normalized distance - midline to edge
0
50
CD
301b
Inte
nsity
(A
U)
2. Average signal
intensity
064128191255 AU
Woundmidline
Fig. 6. CD301b+ macrophage–derived ligands activate AP proliferation. (A) Cell populationsisolated from 5-day wound beds for RNA-seq (left) and FPKM (fragments per kilobase per million)scatter-plotting (right). (B) Table of ligands enriched in CD301b+ macrophages that bind to receptorson APs. (C) Quantification of AP proliferation after the administration of ligands. Fetal bovineserum (FBS) at 10% is a positive control (n = 5 samples, ***P < 0.001). BSA, bovine serum albumin.(D) Quantification of in vivo cellular proliferation after the administration of PDGFC (n = 6 samples,*P = 0.0337)– and IGF1 (n = 6 samples, *P = 0.0436)–neutralizing antibodies (nAbs) or antagonistsagainst PDGFRa (crenolanib) (n = 6 samples, ***P = 0.0001), IGFR1 (linsitinib) (n = 4 samples,*P = 0.01017), or PI 3-kinase (wortmannin) (n = 4 samples, **P = 0.0028). DMSO, dimethylsulfoxide. (E) Pipeline for processing immunostained wound bed sections to infer the distributionof CD301b+ macrophages in day 5 wounds (n = 6 samples). Yellow lines delineate wound edges.Scale bar, 250 mm. Error bars indicate the mean ± SEM.
RESEARCH | RESEARCH ARTICLEon S
eptember 12, 2020
http://science.sciencem
ag.org/D
ownloaded from

marker expression associated with different agesand hair follicle stages (29). Our data reveal thatbiased proliferation and plasticity of fibroblastsubsets promote myofibroblast heterogeneity inskin wounds. Our lineage tracing data suggestthat a combination of proliferation and plasticitysupports fibroblast heterogeneity within regen-erating skin.Althoughmultiple signals likely influence myo-
fibroblast heterogeneity, our study highlights theimportance of myofibroblast-macrophage inter-actions and, particularly, of PDGFC and IGF1 inpromoting myofibroblast heterogeneity and re-pair. These data correspond with the function ofmacrophages in tissue fibrosis (1, 58), the abilityof exogenousPDGFC to rescuedelayed skinwoundhealing in diabetic mice (59), and the promotionof fibroblast proliferation and repair by IGF1(35, 50–53). In various tissues, macrophages ex-press Pdgfc and Igf1 after injury or under path-ological conditions (60–64). PDGF signaling andIGF signaling cooperate synergistically to pro-mote fibroblast proliferation and enhance woundhealing without increasing scarring (65–67). Treat-ments aimedat fine-tuning thenumber of CD301b+
macrophages could be of tremendous clinicalvalue, as reduction in CD301b+macrophages andCD26-expressing fibroblasts is associated withaging and defective wound healing and keloidscars contain excessive ECM, CD26-expressingfibroblasts, andCD301+ cells. Further understand-ing of howmyofibroblast subsets function and areinfluenced by the microenvironment during fi-brosis and pathologies with irregular ECMhomeo-stasis will allow optimization of treatments forthese encumbering diseases.
REFERENCES AND NOTES
1. T. A. Wynn, K. M. Vannella, Macrophages in tissue repair,regeneration, and fibrosis. Immunity 44, 450–462 (2016).doi: 10.1016/j.immuni.2016.02.015; pmid: 26982353
2. S. Ebmeier, V. Horsley, Origin of fibrosing cells in systemicsclerosis. Curr. Opin. Rheumatol. 27, 555–562 (2015).doi: 10.1097/BOR.0000000000000217; pmid: 26352735
3. A. Gosain, L. A. DiPietro, Aging and wound healing. World J.Surg. 28, 321–326 (2004). doi: 10.1007/s00268-003-7397-6;pmid: 14961191
4. C. B. Ballas, J. M. Davidson, Delayed wound healing in agedrats is associated with increased collagen gel remodeling andcontraction by skin fibroblasts, not with differences inapoptotic or myofibroblast cell populations. Wound RepairRegen. 9, 223–237 (2001). doi: 10.1046/j.1524-475x.2001.00223.x; pmid: 11472619
5. T. Fujiwara et al., Age-associated intracellular superoxidedismutase deficiency potentiates dermal fibroblast dysfunctionduring wound healing. Exp. Dermatol. (2017). doi: 10.1111/exd.13404; pmid: 28677217
6. N. Vedrenne, B. Coulomb, A. Danigo, F. Bonté, A. Desmoulière,The complex dialogue between (myo)fibroblasts and theextracellular matrix during skin repair processes and ageing.Pathol. Biol. (Paris) 60, 20–27 (2012). doi: 10.1016/j.patbio.2011.10.002; pmid: 22099331
7. E. C. Naylor, R. E. B. Watson, M. J. Sherratt, Molecular aspectsof skin ageing. Maturitas 69, 249–256 (2011). doi: 10.1016/j.maturitas.2011.04.011; pmid: 21612880
8. E. Kohl, J. Steinbauer, M. Landthaler, R. M. Szeimies, Skinageing. J. Eur. Acad. Dermatol. Venereol. 25, 873–884 (2011).doi: 10.1111/j.1468-3083.2010.03963.x; pmid: 21261751
9. T. Tabib, C. Morse, T. Wang, W. Chen, R. Lafyatis, SFRP2/DPP4and FMO1/LSP1 define major fibroblast populations inhuman skin. J. Invest. Dermatol. 138, 802–810 (2018).doi: 10.1016/j.jid.2017.09.045; pmid: 29080679
10. C. Philippeos et al., Spatial and single-cell transcriptionalprofiling identifies functionally distinct human dermal fibroblast
subpopulations. J. Invest. Dermatol. 138, 811–825 (2018).doi: 10.1016/j.jid.2018.01.016; pmid: 29391249
11. R. R. Driskell, F. M. Watt, Understanding fibroblastheterogeneity in the skin. Trends Cell Biol. 25, 92–99 (2015).doi: 10.1016/j.tcb.2014.10.001; pmid: 25455110
12. J. M. Sorrell, A. I. Caplan, Fibroblast heterogeneity: More thanskin deep. J. Cell Sci. 117, 667–675 (2004). pmid: 14754903
13. R. G. Marangoni et al., Myofibroblasts in murine cutaneousfibrosis originate from adiponectin-positive intradermalprogenitors. Arthritis Rheumatol. 67, 1062–1073 (2015).doi: 10.1002/art.38990; pmid: 25504959
14. Y. Rinkevich et al., Identification and isolation of a dermallineage with intrinsic fibrogenic potential. Science 348, aaa2151(2015). doi: 10.1126/science.aaa2151; pmid: 25883361
15. R. R. Driskell et al., Distinct fibroblast lineages determine dermalarchitecture in skin development and repair. Nature 504, 277–281(2013). doi: 10.1038/nature12783; pmid: 24336287
16. D. Jiang et al., Two succeeding fibroblastic lineages drivedermal development and the transition from regeneration toscarring. Nat. Cell Biol. 20, 422–431 (2018). doi: 10.1038/s41556-018-0073-8; pmid: 29593327
17. I. A. Darby, B. Laverdet, F. Bonté, A. Desmoulière, Fibroblastsand myofibroblasts in wound healing. Clin. Cosmet. Investig.Dermatol. 7, 301–311 (2014). pmid: 25395868
18. B. Hinz et al., Recent developments in myofibroblast biology:Paradigms for connective tissue remodeling. Am. J. Pathol.180, 1340–1355 (2012). doi: 10.1016/j.ajpath.2012.02.004;pmid: 22387320
19. T. A. Wynn, T. R. Ramalingam, Mechanisms of fibrosis:Therapeutic translation for fibrotic disease. Nat. Med. 18,1028–1040 (2012). doi: 10.1038/nm.2807; pmid: 22772564
20. M. V. Plikus et al., Regeneration of fat cells frommyofibroblasts during wound healing. Science 355, 748–752(2017). doi: 10.1126/science.aai8792; pmid: 28059714
21. T. Iwayama et al., PDGFRa signaling drives adipose tissuefibrosis by targeting progenitor cell plasticity. Genes Dev. 29,1106–1119 (2015). doi: 10.1101/gad.260554.115; pmid: 26019175
22. G. Marcelin et al., A PDGFRa-mediated switch toward CD9high
adipocyte progenitors controls obesity-induced adiposetissue fibrosis. Cell Metab. 25, 673–685 (2017). doi: 10.1016/j.cmet.2017.01.010; pmid: 28215843
23. R. Higashiyama et al., Differential contribution of dermal residentand bone marrow-derived cells to collagen production duringwound healing and fibrogenesis in mice. J. Invest. Dermatol. 131,529–536 (2011). doi: 10.1038/jid.2010.314; pmid: 20962852
24. O. Kanisicak et al., Genetic lineage tracing defines myofibroblastorigin and function in the injured heart. Nat. Commun. 7, 12260(2016). doi: 10.1038/ncomms12260; pmid: 27447449
25. R. Berry, M. S. Rodeheffer, Characterization of the adipocytecellular lineage in vivo. Nat. Cell Biol. 15, 302–308 (2013).doi: 10.1038/ncb2696; pmid: 23434825
26. G. C. Rivera-Gonzalez et al., Skin adipocyte stem cell self-renewal is regulated by a PDGFA/AKT-signaling axis. Cell StemCell 19, 738–751 (2016). pmid: 27746098
27. C. D. Church, R. Berry, M. S. Rodeheffer, Isolation and study ofadipocyte precursors. Methods Enzymol. 537, 31–46 (2014).doi: 10.1016/B978-0-12-411619-1.00003-3; pmid: 24480340
28. M. S. Rodeheffer, K. Birsoy, J. M. Friedman, Identification ofwhite adipocyte progenitor cells in vivo. Cell 135, 240–249(2008). doi: 10.1016/j.cell.2008.09.036; pmid: 18835024
29. B. M. Lichtenberger, M. Mastrogiannaki, F. M. Watt, Epidermalb-catenin activation remodels the dermis via paracrinesignalling to distinct fibroblast lineages. Nat. Commun. 7,10537 (2016). doi: 10.1038/ncomms10537; pmid: 26837596
30. T. Ostendorf, F. Eitner, J. Floege, The PDGF family in renalfibrosis. Pediatr. Nephrol. 27, 1041–1050 (2012). doi: 10.1007/s00467-011-1892-z; pmid: 21597969
31. L. J. Reigstad, J. E. Varhaug, J. R. Lillehaug, Structural andfunctional specificities of PDGF-C and PDGF-D, the novelmembers of the platelet-derived growth factors family. FEBS J.272, 5723–5741 (2005). doi: 10.1111/j.1742-4658.2005.04989.x; pmid: 16279938
32. J. C. Bonner, Regulation of PDGF and its receptors in fibroticdiseases. Cytokine Growth Factor Rev. 15, 255–273 (2004).doi: 10.1016/j.cytogfr.2004.03.006; pmid: 15207816
33. S. Dulauroy, S. E. Di Carlo, F. Langa, G. Eberl, L. Peduto,Lineage tracing and genetic ablation of ADAM12+ perivascularcells identify a major source of profibrotic cells during acutetissue injury. Nat. Med. 18, 1262–1270 (2012). doi: 10.1038/nm.2848; pmid: 22842476
34. S. Paquet-Fifield et al., A role for pericytes as microenvironmentalregulators of human skin tissue regeneration. J. Clin. Invest. 119,2795–2806 (2009). pmid: 19652362
35. S. Werner, R. Grose, Regulation of wound healing by growthfactors and cytokines. Physiol. Rev. 83, 835–870 (2003).doi: 10.1152/physrev.2003.83.3.835; pmid: 12843410
36. J. Ding, E. E. Tredget, The role of chemokines in fibrotic woundhealing. Adv. Wound Care 4, 673–686 (2015). doi: 10.1089/wound.2014.0550; pmid: 26543681
37. M. Xue, C. J. Jackson, Extracellular matrix reorganizationduring wound healing and its impact on abnormal scarring.Adv. Wound Care 4, 119–136 (2015). doi: 10.1089/wound.2013.0485; pmid: 25785236
38. I. Goren et al., A transgenic mouse model of induciblemacrophage depletion: Effects of diphtheria toxin-drivenlysozyme M-specific cell lineage ablation on woundinflammatory, angiogenic, and contractive processes.Am. J. Pathol. 175, 132–147 (2009). doi: 10.2353/ajpath.2009.081002; pmid: 19528348
39. T. Lucas et al., Differential roles of macrophages in diversephases of skin repair. J. Immunol. 184, 3964–3977 (2010).doi: 10.4049/jimmunol.0903356; pmid: 20176743
40. R. Mirza, L. A. DiPietro, T. J. Koh, Selective and specificmacrophage ablation is detrimental to wound healing in mice.Am. J. Pathol. 175, 2454–2462 (2009). doi: 10.2353/ajpath.2009.090248; pmid: 19850888
41. B. Shook, E. Xiao, Y. Kumamoto, A. Iwasaki, V. Horsley, CD301b+macrophages are essential for effective skin wound healing.J. Invest. Dermatol. 136, 1885–1891 (2016). doi: 10.1016/j.jid.2016.05.107; pmid: 27287183
42. B. E. Clausen, C. Burkhardt, W. Reith, R. Renkawitz, I. Förster,Conditional gene targeting in macrophages and granulocytesusing LysMcre mice. Transgenic Res. 8, 265–277 (1999).doi: 10.1023/A:1008942828960; pmid: 10621974
43. T. Buch et al., A Cre-inducible diphtheria toxin receptormediates cell lineage ablation after toxin administration.Nat. Methods 2, 419–426 (2005). doi: 10.1038/nmeth762;pmid: 15908920
44. J. M. Daley, S. K. Brancato, A. A. Thomay, J. S. Reichner,J. E. Albina, The phenotype of murine wound macrophages.J. Leukocyte Biol. 87, 59–67 (2010). doi: 10.1189/jlb.0409236;pmid: 20052800
45. R. Mirza, T. J. Koh, Dysregulation of monocyte/macrophagephenotype in wounds of diabetic mice. Cytokine 56, 256–264(2011). doi: 10.1016/j.cyto.2011.06.016; pmid: 21803601
46. Y. Kumamoto et al., CD301b+ dermal dendritic cells driveT helper 2 cell-mediated immunity. Immunity 39, 733–743 (2013).doi: 10.1016/j.immuni.2013.08.029; pmid: 24076051
47. B. Yang et al., IL-27 facilitates skin wound healing through inductionof epidermal proliferation and host defense. J. Invest. Dermatol. 137,1166–1175 (2017). doi: 10.1016/j.jid.2017.01.010; pmid: 28132857
48. M. S. Hu et al., Delivery of monocyte lineage cells in abiomimetic scaffold enhances tissue repair. JCI Insight 2,e96260 (2017). doi: 10.1172/jci.insight.96260; pmid: 28978794
49. S. Werner, T. Krieg, H. Smola, Keratinocyte-fibroblast interactionsin wound healing. J. Invest. Dermatol. 127, 998–1008 (2007).doi: 10.1038/sj.jid.5700786; pmid: 17435785
50. Y. Ando, P. J. Jensen, Epidermal growth factor and insulin-likegrowth factor I enhance keratinocyte migration. J. Invest.Dermatol. 100, 633–639 (1993). doi: 10.1111/1523-1747.ep12472297; pmid: 8491986
51. I. Haase, R. Evans, R. Pofahl, F. M. Watt, Regulation ofkeratinocyte shape, migration and wound epithelializationby IGF-1- and EGF-dependent signalling pathways. J. Cell Sci.116, 3227–3238 (2003). pmid: 12829742
52. J. G. Simmons, J. B. Pucilowska, T. O. Keku, P. K. Lund, IGF-Iand TGF-beta1 have distinct effects on phenotype andproliferation of intestinal fibroblasts. Am. J. Physiol.Gastrointest. Liver Physiol. 283, G809–G818 (2002).doi: 10.1152/ajpgi.00057.2002; pmid: 12181198
53. S. M. Rudman, M. P. Philpott, G. A. Thomas, T. Kealey, Therole of IGF-I in human skin and its appendages: Morphogenas well as mitogen? J. Invest. Dermatol. 109, 770–777 (1997).doi: 10.1111/1523-1747.ep12340934; pmid: 9406819
54. Y. Xin et al., Expansion of CD26 positive fibroblast populationpromotes keloid progression. Exp. Cell Res. 356, 104–113(2017). pmid: 28454879
55. H. Suga et al., Tracking the elusive fibrocyte: Identification andcharacterization of collagen-producing hematopoietic lineagecells during murine wound healing. Stem Cells 32, 1347–1360(2014). doi: 10.1002/stem.1648; pmid: 24446236
56. B. Hinz, Formation and function of the myofibroblast duringtissue repair. J. Invest. Dermatol. 127, 526–537 (2007).doi: 10.1038/sj.jid.5700613; pmid: 17299435
57. J. R. Rock et al., Multiple stromal populations contribute topulmonary fibrosis without evidence for epithelial to
Shook et al., Science 362, eaar2971 (2018) 23 November 2018 7 of 8
RESEARCH | RESEARCH ARTICLEon S
eptember 12, 2020
http://science.sciencem
ag.org/D
ownloaded from

mesenchymal transition. Proc. Natl. Acad. Sci. U.S.A.108, E1475–E1483 (2011). doi: 10.1073/pnas.1117988108;pmid: 22123957
58. K. M. Vannella, T. A. Wynn, Mechanisms of organ injury andrepair by macrophages. Annu. Rev. Physiol. 79, 593–617(2017). doi: 10.1146/annurev-physiol-022516-034356;pmid: 27959618
59. D. G. Gilbertson et al., Platelet-derived growth factor C(PDGF-C), a novel growth factor that binds to PDGF a andb receptor. J. Biol. Chem. 276, 27406–27414 (2001).doi: 10.1074/jbc.M101056200; pmid: 11297552
60. A. Triantafyllopoulou et al., Proliferative lesions andmetalloproteinase activity in murine lupus nephritis mediatedby type I interferons and macrophages. Proc. Natl. Acad. Sci. U.S.A.107, 3012–3017 (2010). doi: 10.1073/pnas.0914902107;pmid: 20133703
61. H. R. Chang, H. J. Kim, X. Xu, A. W. Ferrante Jr., Macrophageand adipocyte IGF1 maintain adipose tissue homeostasisduring metabolic stresses. Obesity (Silver Spring) 24, 172–183(2016). doi: 10.1002/oby.21354; pmid: 26663512
62. P. Ramachandran et al., Differential Ly-6C expression identifiesthe recruited macrophage phenotype, which orchestrates theregression of murine liver fibrosis. Proc. Natl. Acad. Sci. U.S.A.109, E3186–E3195 (2012). doi: 10.1073/pnas.1119964109;pmid: 23100531
63. J. Tonkin et al., Monocyte/macrophage-derived IGF-1orchestrates murine skeletal muscle regeneration and
modulates autocrine polarization. Mol. Ther. 23, 1189–1200(2015). doi: 10.1038/mt.2015.66; pmid: 25896247
64. D. Pohlers, R. Huber, B. Ukena, R. W. Kinne, Expression ofplatelet-derived growth factors C and D in the synovialmembrane of patients with rheumatoid arthritis andosteoarthritis. Arthritis Rheum. 54, 788–794 (2006).doi: 10.1002/art.21670; pmid: 16508943
65. S. E. Lynch, J. C. Nixon, R. B. Colvin, H. N. Antoniades, Role ofplatelet-derived growth factor in wound healing: Synergisticeffects with other growth factors. Proc. Natl. Acad. Sci. U.S.A.84, 7696–7700 (1987). doi: 10.1073/pnas.84.21.7696;pmid: 3499612
66. S. E. Lynch, R. B. Colvin, H. N. Antoniades, Growth factors inwound healing. Single and synergistic effects on partialthickness porcine skin wounds. J. Clin. Invest. 84, 640–646(1989). doi: 10.1172/JCI114210; pmid: 2788174
67. S. E. Lynch et al., The effects of short-term application of acombination of platelet-derived and insulin-like growth factorson periodontal wound healing. J. Periodontol. 62, 458–467(1991). doi: 10.1902/jop.1991.62.7.458; pmid: 1920013
ACKNOWLEDGMENTS
We thank members of the Horsley lab, R. Atit, and A. MacLeodfor critical reading of the manuscript. We thank B. Hogan andC. Barkauskas for sharing PdgfraCreER mice and C. Wolfrum forsharing Dlk1CreER mice before the initial mouse line publications andR. Atit for En1Cre;Rosa26-LSL-tdTomato mice. Funding: This work was
supported in part by NIH grants to V.H. from NIAMS (AR060295 andAR069550) and NIA through pilot project grants from the ClaudeD. Pepper Older Americans Independence Center at Yale (NIAP30AG21342) awarded to V.H. This research was supported in part by aNew Investigator award to H.C.H. from the Yale Department of SurgeryOhse Fund. B.A.S. is a New York Stem Cell Foundation–DruckenmillerFellow. This research was supported by the New York Stem CellFoundation. Author contributions: B.A.S. conceived, designed, andoversaw the experiments with suggestions from V.H., J.L.A., K.M.-J.,D.A.C., andH.C.H., R.R.W, G.C.R.-G., and A.R.M.-R. performed experimentsand analyzed the data. B.C.D., E.S.-G., K.D.A., R.K.Z., and V.L. assistedwith collection and analysis of data. F.L.-G. analyzed the RNA-seq data.B.A.S. and V.H. wrote the manuscript. Competing interests: Theauthors have no competing interests. Data and materials availability:RNA-seq data are available at the Gene Expression Omnibus (GEO)under accession numbers GSE105788, GSE105789, and GSE105790.
SUPPLEMENTARY MATERIALS
www.sciencemag.org/content/362/6417/eaar2971/suppl/DC1Materials and MethodsFigs. S1 to S21Tables S1 to S3References (68–78)
23 October 2017; resubmitted 20 July 2018Accepted 4 October 201810.1126/science.aar2971
Shook et al., Science 362, eaar2971 (2018) 23 November 2018 8 of 8
RESEARCH | RESEARCH ARTICLEon S
eptember 12, 2020
http://science.sciencem
ag.org/D
ownloaded from

repairMyofibroblast proliferation and heterogeneity are supported by macrophages during skin
Damon A. Clark, Henry C. Hsia and Valerie HorsleyDash, Andrés R. Muñoz-Rojas, Krystal D. Aultman, Rachel K. Zwick, Vivian Lei, Jack L. Arbiser, Kathryn Miller-Jensen, Brett A. Shook, Renee R. Wasko, Guillermo C. Rivera-Gonzalez, Emilio Salazar-Gatzimas, Francesc López-Giráldez, Biraja C.
DOI: 10.1126/science.aar2971 (6417), eaar2971.362Science
, this issue p. eaar2971; see also p. 891Sciencetissue repair and aging, which may have therapeutic implications for chronic wounds and fibrotic disease.composition and gene expression changed during aging. Thus, macrophage-fibroblast interactions are important duringCD301b-expressing macrophages selectively activated proliferation of APs, but not other myofibroblasts. Myofibroblast myofibroblasts, including cells identified as adipocyte precursors (APs). In cellular ablation mouse models,fibrosis, and aging (see the Perspective by Willenborg and Eming). They identified distinct subpopulations of
examined cells during mouse skin wound healing,et al.much ECM is deposited, fibrosis and scarring results. Shook Fibroblasts deposit extracellular matrix (ECM) molecules to regulate tissue strength and function. However, if too
Myofibroblast diversity with injury and aging
ARTICLE TOOLS http://science.sciencemag.org/content/362/6417/eaar2971
MATERIALSSUPPLEMENTARY http://science.sciencemag.org/content/suppl/2018/11/19/362.6417.eaar2971.DC1
CONTENTRELATED
http://stke.sciencemag.org/content/sigtrans/13/648/eaay8690.fullhttp://stke.sciencemag.org/content/sigtrans/12/564/eaao3469.fullhttp://stm.sciencemag.org/content/scitransmed/10/432/eaai8524.fullhttp://stm.sciencemag.org/content/scitransmed/9/371/eaaf8611.fullhttp://stm.sciencemag.org/content/scitransmed/9/415/eaal2774.fullhttp://stm.sciencemag.org/content/scitransmed/10/451/eaap8798.fullhttp://science.sciencemag.org/content/sci/362/6417/891.full
REFERENCES
http://science.sciencemag.org/content/362/6417/eaar2971#BIBLThis article cites 78 articles, 15 of which you can access for free
PERMISSIONS http://www.sciencemag.org/help/reprints-and-permissions
Terms of ServiceUse of this article is subject to the
is a registered trademark of AAAS.ScienceScience, 1200 New York Avenue NW, Washington, DC 20005. The title (print ISSN 0036-8075; online ISSN 1095-9203) is published by the American Association for the Advancement ofScience
Science. No claim to original U.S. Government WorksCopyright © 2018 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of
on Septem
ber 12, 2020
http://science.sciencemag.org/
Dow
nloaded from