
medfazna encimatika - bio.ijs.sibio.ijs.si/~krizaj/group/Predavanja 2010/Encimi, ki delujejo na...
34
1 Slika iz: Lin et al., (1998) Science 279. Encimi, ki delujejo na agregiranih substratih (npr. lipidnih membranah). medfazna encimatika klasična encimatika membranska površina
Transcript of medfazna encimatika - bio.ijs.sibio.ijs.si/~krizaj/group/Predavanja 2010/Encimi, ki delujejo na...

1
Slika iz: Lin et al., (1998) Science 279.
Encimi, ki delujejo na agregiranih substratih (npr. lipidnih membranah).
medfazna encimatika
klasična encimatika
membranska površina

2
• Dostopajo do substrata na fazni meji med vodnim in lipidnim okoljem
• Evolucija v smeri produktivne interakcije s površino agregatov
⇒ učinkovita vezava substrata v aktivno mesto
• Encimi lipidnega metabolizma:
- PI-3-kinaze, gliceridne lipaze, bakterijska sfingomielinaza, fosfolipaze A1 in A2,
PI-fosfolipaza C
• Sekretorne PLA2 – paradigma medfazne encimatike
• Sekretorne fosfolipaze A2 (sPLA2)
• Substrati – značilnosti fosfolipidnih agregatov
• Zakaj je medfazna kinetika zapletena?
• Kako jo uporabljamo v praksi?

3
PLA2
POPC
• Cepijo sredinsko (sn-2) estrsko vez v molekulah glicerofosfolipidov
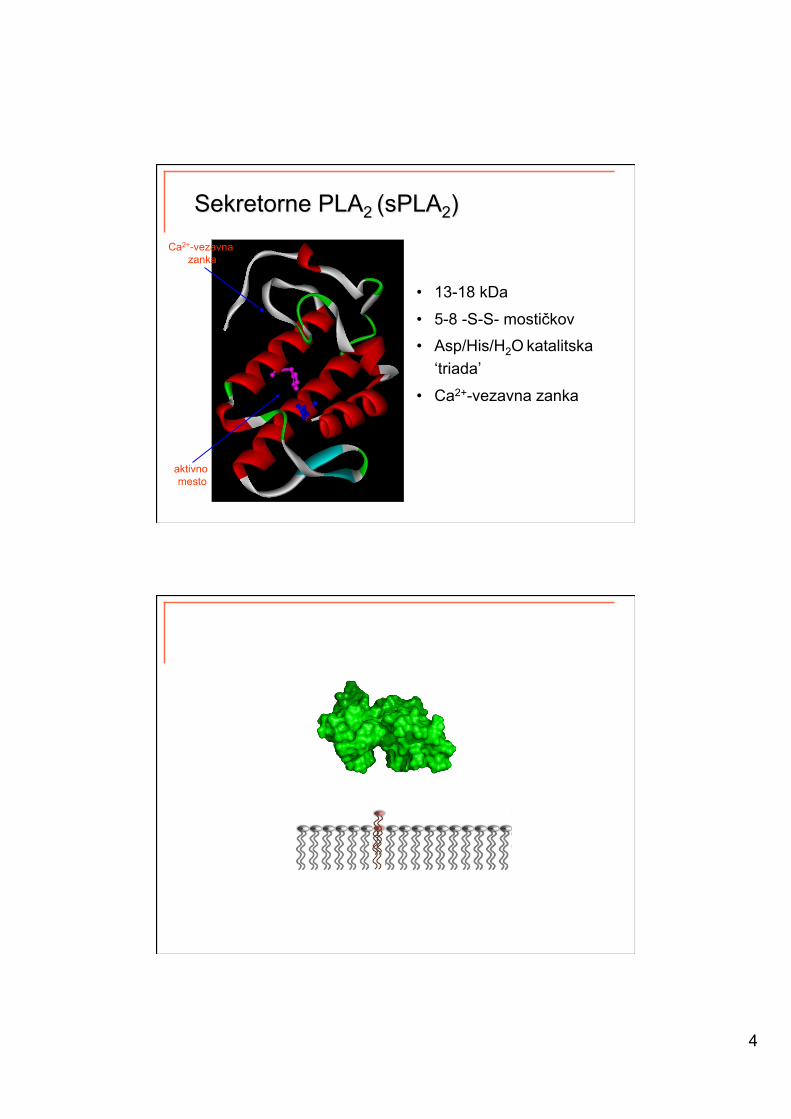
4
• 13-18 kDa
• 5-8 -S-S- mostičkov
• Asp/His/H2O katalitska ‘triada’
• Ca2+-vezavna zanka
Ca2+-vezavna zanka
aktivno mesto
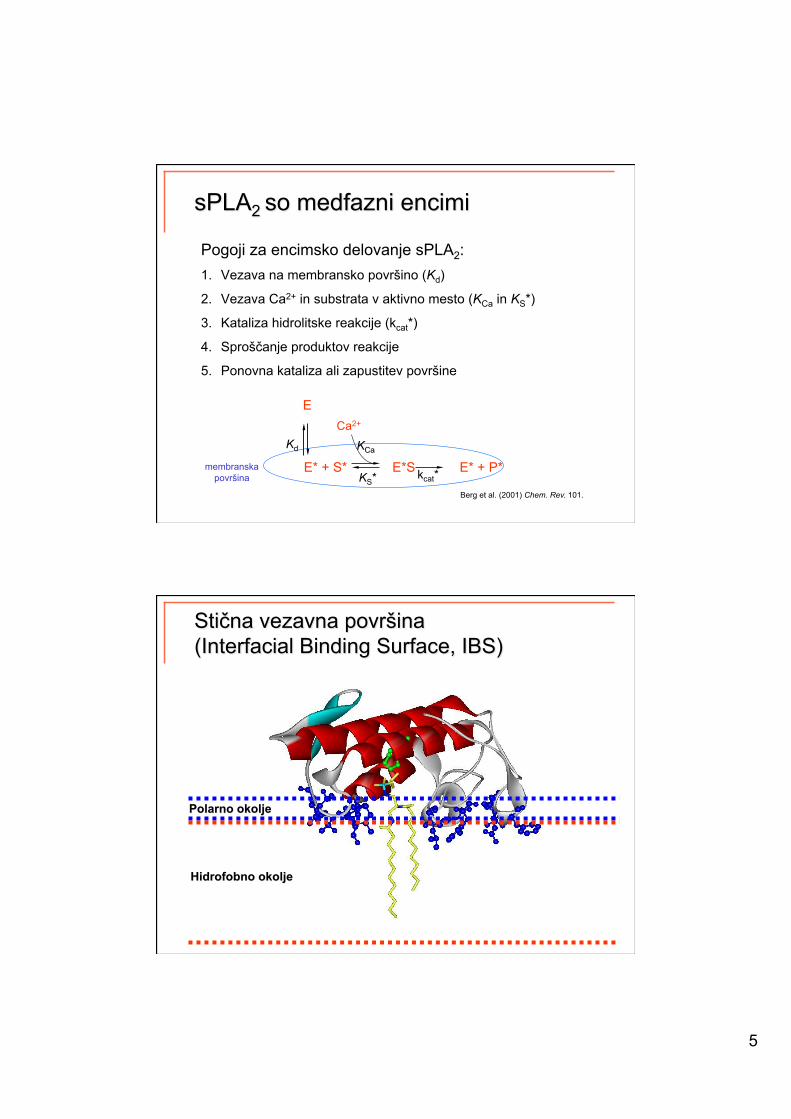
5
Pogoji za encimsko delovanje sPLA2: 1. Vezava na membransko površino (Kd)
2. Vezava Ca2+ in substrata v aktivno mesto (KCa in KS*)
3. Kataliza hidrolitske reakcije (kcat*)
4. Sproščanje produktov reakcije
5. Ponovna kataliza ali zapustitev površine
E* + S* E*S E* + P* KS* kcat*
Kd
E
Berg et al. (2001) Chem. Rev. 101.
membranska površina
Ca2+

6
F124
N119 R118
V31 F24
L19
T20
L2
L3
K69
R72
T70 S67
H48
Interakcije z membrano: – centralni obroč hidrofobnih
in aromatskih ostankov – polarni, bazični, kisli
ostanki na robovih
Petan et al., (2005) Biochemistry, 44.
Ca2+-vezavna zanka
Kalcij je nujen za vezavo fosfolipidne molekule v aktivno mesto in katalitsko reakcijo, ni pa potreben za vezavo sPLA2 na membransko površino.
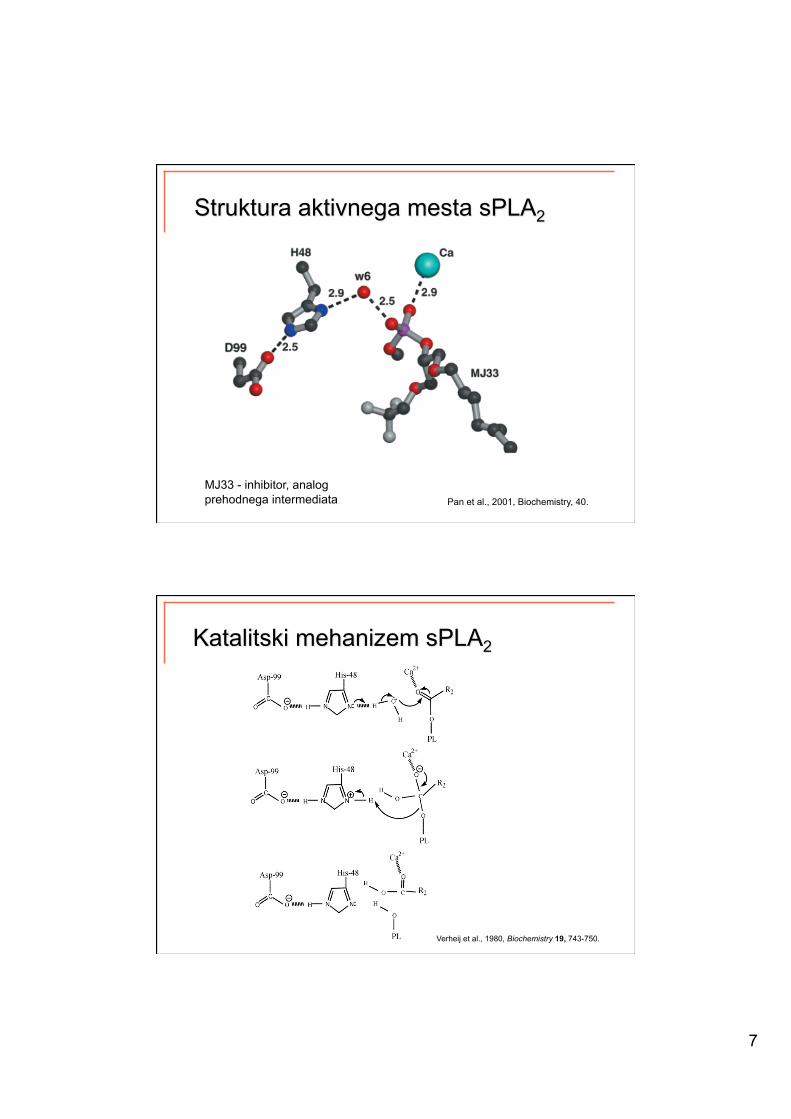
7
Pan et al., 2001, Biochemistry, 40.
MJ33 - inhibitor, analog prehodnega intermediata
Verheij et al., 1980, Biochemistry 19, 743-750.

8
• “Specifičnost” ali afiniteta za vezavo na membrano
• fizikalno-kemijske lastnosti celotne membranske površine in zgradba IBS-a
• večina sPLA2 se zelo dobro veže na negativno nabite (PG, PS, PM), a veliko slabše na zwitterionske (elektronevtralne) membranske površine (PC, PE)
• Substratna specifičnost aktivnega mesta
• struktura fosfolipidne molekule in njeno prileganje v aktivno mesto • večina sPLA2 nima izražene specifičnosti do posameznih fosfolipidov

9
• Bistvo razumevanja medfazne kinetike se skriva v poznavanju narave agregiranih substratov
• Že najbolj enostavni eksperimentalni modeli za sPLA2 (npr. suspenzija fosfatidilholinskih liposomov v vodi) so zelo kompleksni za kinetsko analizo
• Amfipatske molekule => specifična organizacija/agregacija molekul v vodnem okolju
• Na strukturo agregatov vplivajo: – Dolžina in (ne)nasičenost verig CH – Struktura polarne glave – Metoda disperzije – Koncentracija soli – Temperatura – Prisotnost drugih amfipatskih molekul

10
< 4 CH2-skupin < 10 CH2-skupin
• SUV, LUV, GUV – ”Small/Large/Giant unilamellar vesicles” (unilamelarni vezikli)
• MLV – multilamelarni vezikli
> 10 CH2-skupin

11
16:0-18:1 PG
16:0-18:1 PC
16:0-18:1 PS
1-palmitoil-2-oleoil-sn-glicero-3-fosfoglicerol
1-palmitoil-2-oleoil-sn-glicero-3-fosfoholin
1-palmitoil-2-oleoil-sn-glicero-3-fosfoserin
(−)
(+/−)
(−)
12
• MLV potiskamo skozi polikarbonatne filtre z izbranim premerom por (0,015 do 12 µm, tipično 100 nm)
LipoFast Extruder, Avestin, Canada
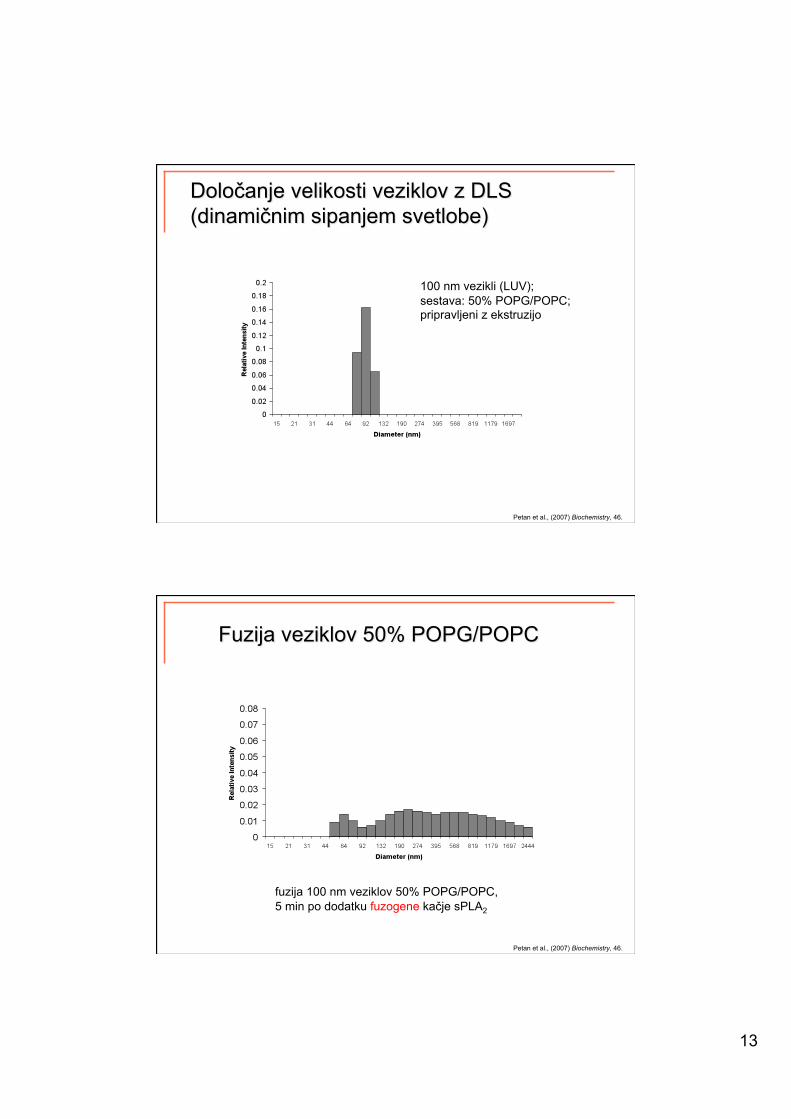
13
100 nm vezikli (LUV); sestava: 50% POPG/POPC; pripravljeni z ekstruzijo
Petan et al., (2007) Biochemistry, 46.
fuzija 100 nm veziklov 50% POPG/POPC, 5 min po dodatku fuzogene kačje sPLA2
Petan et al., (2007) Biochemistry, 46.

14

15
Omejitev dostopnosti substrata na 2D: 1. Encim se mora vezati na površino v odvisnosti od dostopne
vezavne površine, ki jo določa:
- celotna koncentracija substrata
- število, velikost, vrsta (struktura) in dinamika agregatov
2. Aktivnost je odvisna od površinske koncentracije substrata
3. Specifična struktura (fizikalno-kemijske lastnosti) agregata vpliva na vse aspekte medfazne kinetike
Primer delovanja sPLA2 na veziklih PC - “time scrambling in hopping mode”
t1=0 t3>0
=> v danem trenutku vsaka molekula encima “vidi” drugačno okolje!
t2=0 t1>0

16
• Tudi S in P se lahko izmenjujejo med agregati
Počasna izmenjava molekul: fosfolipidi v veziklih ~ nekaj ur, dni
kataliza sPLA2~ nekaj ms
Hitra izmenjava molekul: detergent v micelah ~ nekaj ms
fosfolipidi v mešanih micelah ~ sekunde
MERITVE ODRAŽAJO HITROST KATALITSKE
REAKCIJE
MERITVE ODRAŽAJO HITROST PREHAJANJA S IN P
Hitrosti prehajanja E, S in P med agregati vplivajo na makroskopsko stanje sistema, ki ga eksperimentalno zasledujemo:

17
• Substrat se lahko nahaja v konformaciji, ki pospeši ali inhibira vezavo
• Sosednje molekule lahko sterično ovirajo vezavo E* in S*
• Fazne razlike/prehodi in lateralne domene, substratov, produktov, inhibitorjev…
• Poškodbe površine
• Kopičenje produktov vpliva na strukturo, afiniteto vezave, aktivnost
• Dinamika izmenjave S, P in E
• Procesivnost katalitske reakcije
• Zasedenost vezavnih mest (“crowding”)
=> možne so številne kinetske poti
E* + S* E*S E* + P* KS* k*cat
Kd
E E + S ES E + P
K’s Ksd Kd
kcat KS

18
• Vsaka molekula encima mora videti enako! • Povprečje mikroskopskih dogodkov = makroskopsko stanje
sistema
1. “Scooting”– drseča/procesivna kataliza, ni izmenjave E, S ali P med agregati
2. “Quasi-scooting”– zelo hitra izmenjava E, S, P med agregati (hitrejša od katalitske reakcije)
1 2 3
4 n
n >> 1000 “scooting mode”
E* izvaja procesivno katalizo do popolne hidrolize zunanje plasti vezikla
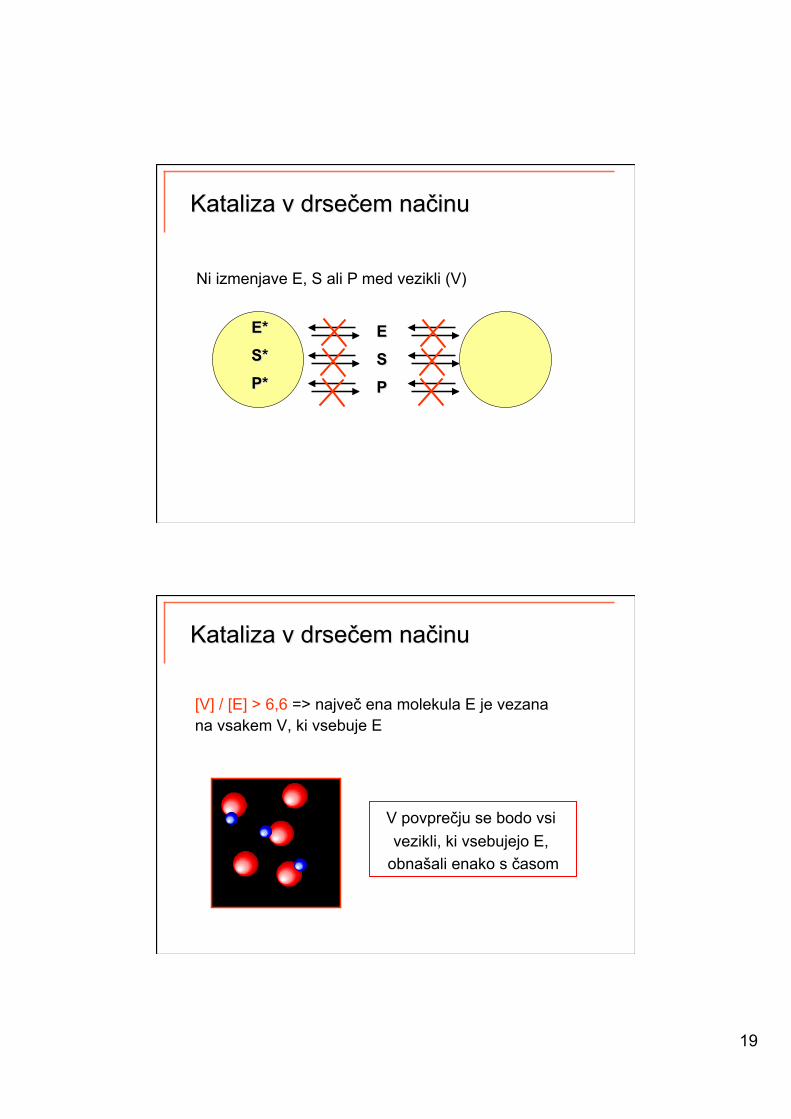
19
Ni izmenjave E, S ali P med vezikli (V)
[V] / [E] > 6,6 => največ ena molekula E je vezana na vsakem V, ki vsebuje E
V povprečju se bodo vsi vezikli, ki vsebujejo E,
obnašali enako s časom
20
⇒ merjenje nastajanja produkta v časovni enoti bo vsota produktov, ki nastajajo na vsakem veziklu, ki vsebuje encim.
⇒ lahko povežemo mikroskopske dogodke na posameznem veziklu z makroskopskim obnašanjem sistema
• pH-stat metoda
• vezikli DMPM (SUV) – Kd ~ 0.1 pM – vezava E*V je res
ireverzibilna!
=> hidrolizira se le del veziklov
DTPM – nehidrolizabilni dietrski fosfolipidi
DMPM

21
• [E] / [V] > 5 => razgradi se le 60% vseh PL molekul, t.j. vsi PL na zunanji strani vseh veziklov
• Ni sproščanja vsebine veziklov
• Reakcija se ustavi, ko ni več PL na zunanji strani E-vsebujočih veziklov
(dokaz: “navidezna aktivacija sPLA2” s polikationskim peptidom polimiksinom B (PxB))
PxB omogoča prehajanje DMPG med vezikli, ne pa tudi DMPA
• Začetno hitrost v0 in konstanto prvega reda ki dobimo z nelinearno regresijo iz krivulje Pt/Pmax (t)
• Ns – število molekul S v zunanjem sloju veziklov dobimo iz: Pmax = [E]celotnaNs
• kcat*, KM*, KP* so še vedno neznanke!
Pt kit = Pmax – ln 1 – ( ) +
Nski
v0 – 1 ( ) Pt
Pmax ( )
kcat* Nski =
KM*(1 + 1/KP*)
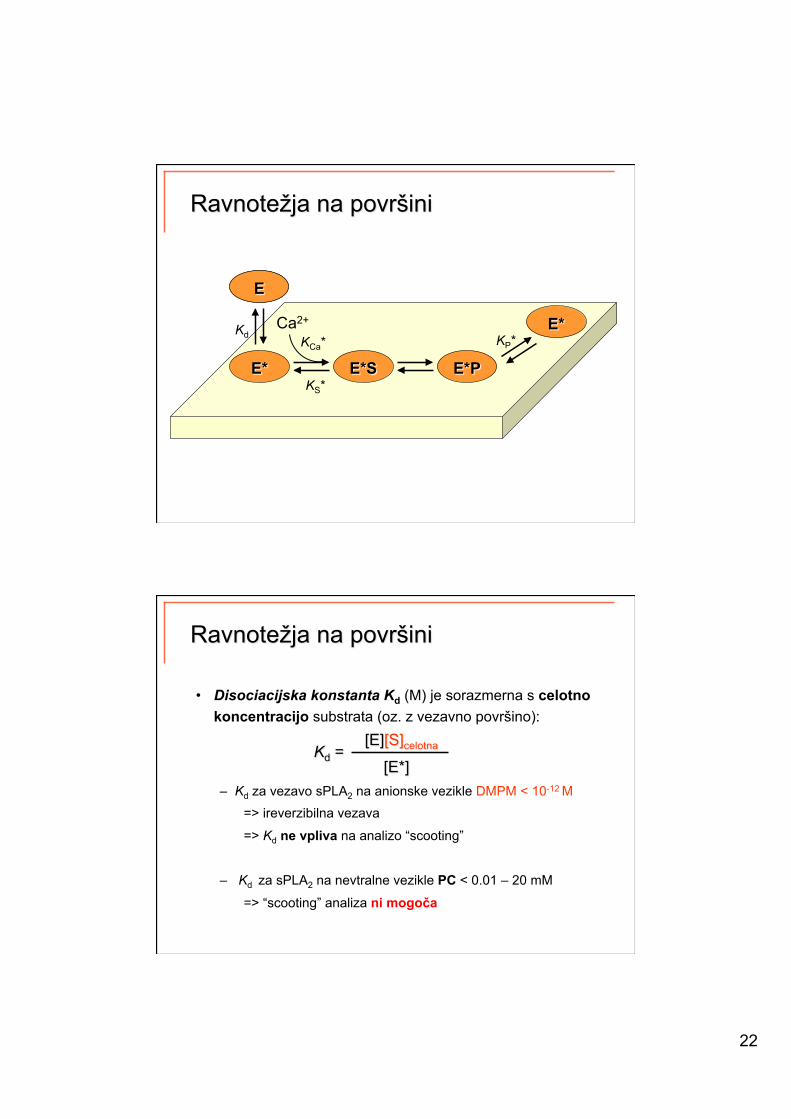
22
KCa* KP*
KS*
Ca2+ Kd
• Disociacijska konstanta Kd (M) je sorazmerna s celotno koncentracijo substrata (oz. z vezavno površino):
– Kd za vezavo sPLA2 na anionske vezikle DMPM < 10-12 M => ireverzibilna vezava
=> Kd ne vpliva na analizo “scooting”
– Kd za sPLA2 na nevtralne vezikle PC < 0.01 – 20 mM
=> “scooting” analiza ni mogoča

23
• 2D konstante ravnotežij za vezavo ligandov:
E* + L* E*L (L = S, P, I)
konstante KS*, KP*, KI* (molski odstotek)
– Določimo jih lahko s spreminjanjem površinskega odstotka liganda (XL*):
XL* = KL* => [E*] = [E*L]
Ko je molski odstotek liganda enak KL* velja, da polovica molekul encima vsebuje vezan ligand.
• Nevtralni razredčevalec (NR) = 2D inertno topilo – ne veže se v aktivno mesto, a je podoben substratu! – tvori vezavno površino za encim – npr. za ppPLA2: 2-heksadecil-glicero-3-PC
• Idealen NR omogoča spreminjanje XL od 0 do 1 in s tem določanje KCa*, KS*, KP*, KI* – metoda temelji na tem, da se E*L bolj počasi inaktivira (alkilacija
His-48 s p-BPB) kot E*
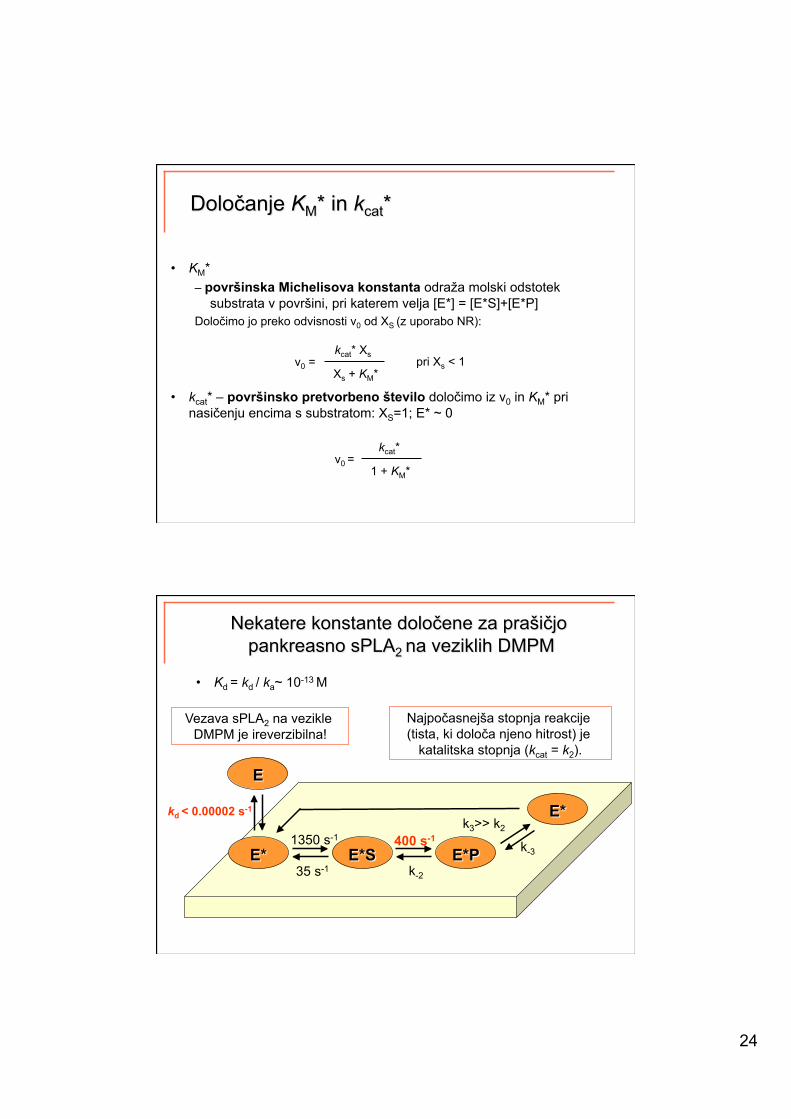
24
• KM* – površinska Michelisova konstanta odraža molski odstotek
substrata v površini, pri katerem velja [E*] = [E*S]+[E*P] Določimo jo preko odvisnosti v0 od XS (z uporabo NR):
• kcat* – površinsko pretvorbeno število določimo iz v0 in KM* pri nasičenju encima s substratom: XS=1; E* ~ 0
kcat* v0 =
1 + KM*
kcat* Xs v0 =
Xs + KM* pri Xs < 1
• Kd = kd / ka~ 10-13 M
kd < 0.00002 s-1
k-3
k3>> k2
35 s-1
1350 s-1
k-2
400 s-1
Vezava sPLA2 na vezikle DMPM je ireverzibilna!
Najpočasnejša stopnja reakcije (tista, ki določa njeno hitrost) je
katalitska stopnja (kcat = k2).

25
“Scooting” analiza je le eksperimentalni “trik”, ki omogoča
kinetsko analizo sPLA2 le pod posebnimi pogoji.

26
Beers et al. (2002) JBC 277.
antibakterijsko delovanje sPLA2 iz skupine IIA
• Afiniteta za vezavo na membrane – Kd
• Vpliv Kd na encimsko aktivnost – metoda FABP
• Vpliv Kd na afiniteto vezave Ca2+ – appKCa

27
• Težavno pri visokih afinitetah (PM, PG)
• Produkti vplivajo na Kd
• nehidrolizabilni fosfolipidi
• odsotnost Ca2+
• encimsko neaktivne mutante
• Tehnike temeljijo na: • fizično ločevanje E in E*
• spremembe spektralnih lastnosti (Trp, FRET)
• kinetska ali ravnotežna analiza s SPR
nehidrolizabilni dietrski fosfolipidi
Kd S
ni hidrolize
Ca2+
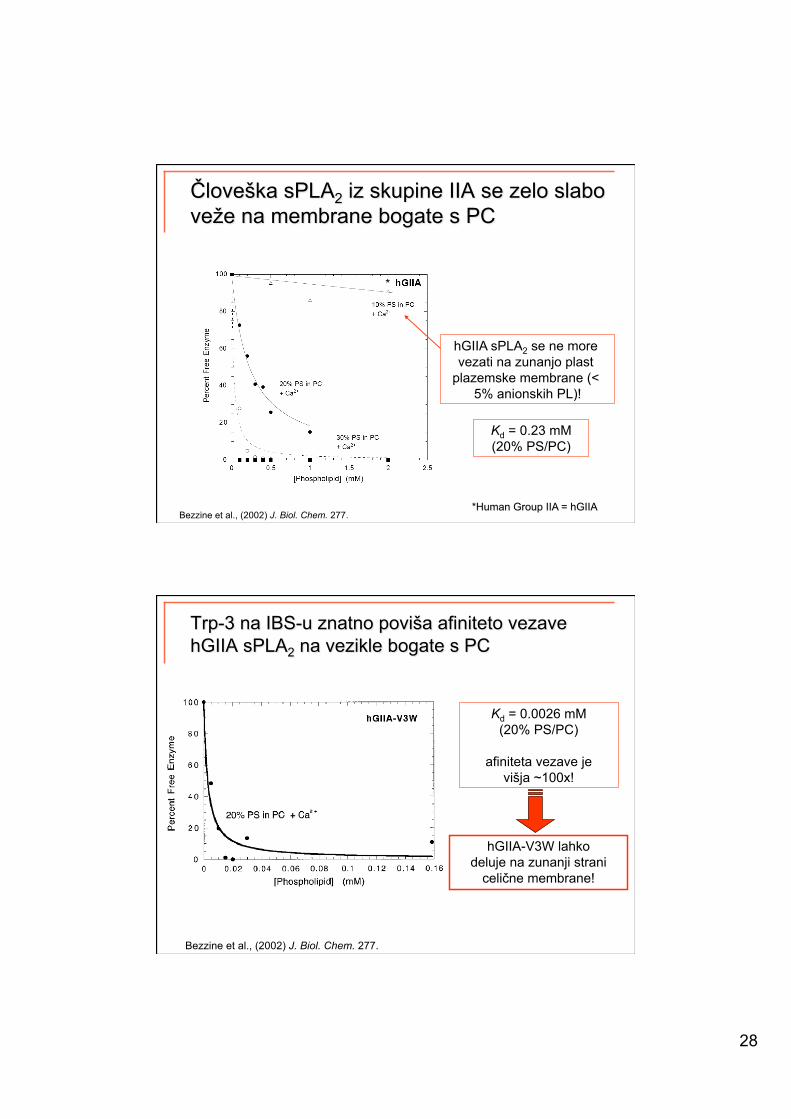
28
Bezzine et al., (2002) J. Biol. Chem. 277.
hGIIA sPLA2 se ne more vezati na zunanjo plast
plazemske membrane (< 5% anionskih PL)!
*
Kd = 0.23 mM (20% PS/PC)
Bezzine et al., (2002) J. Biol. Chem. 277.
Kd = 0.0026 mM (20% PS/PC)
afiniteta vezave je višja ~100x!
hGIIA-V3W lahko deluje na zunanji strani
celične membrane!

29
• Začetna hitrost hidrolize (v0)
• Prednost metode: substrat so lahko poljubni
glicerofosfolipidi s poljubnimi strukturami agregatov
• Zelo občutljiva metoda (0,01-5 pmol sPLA2)
11-dansyl-undecanoic acid - DAUDA FABP
MK
LUV
PLA2
30
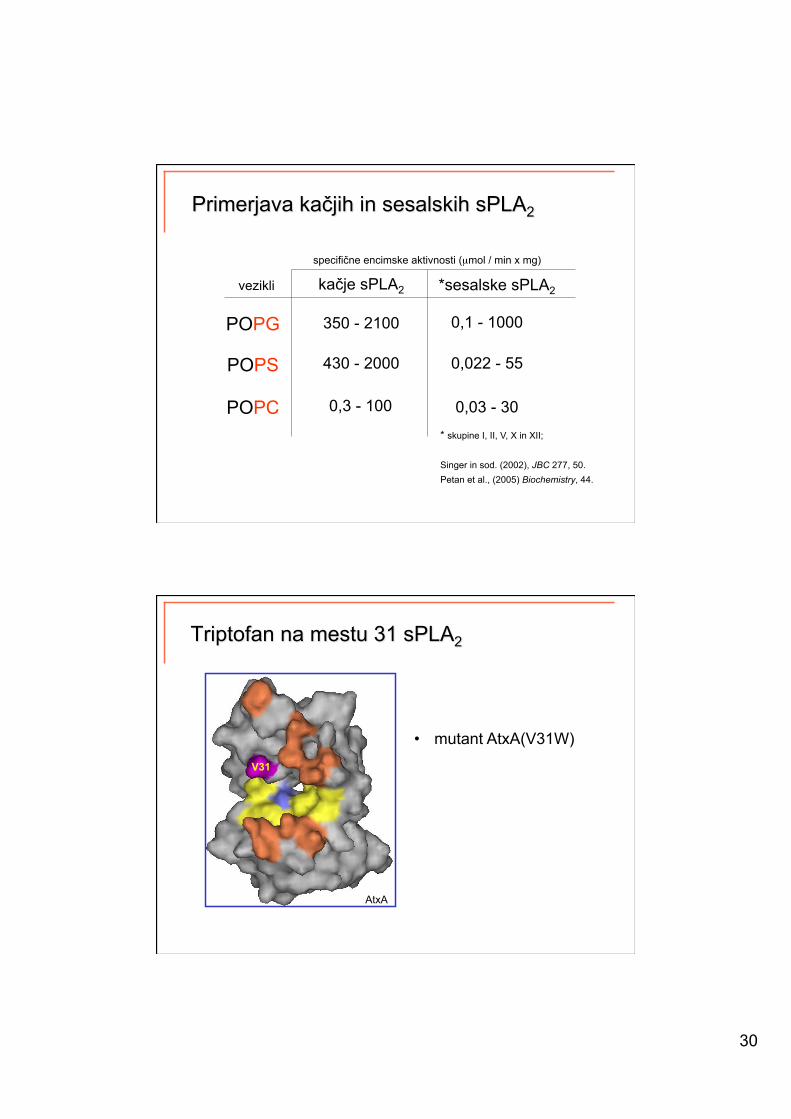
kačje sPLA2 *sesalske sPLA2
specifične encimske aktivnosti (µmol / min x mg)
POPG
POPS
POPC
vezikli
* skupine I, II, V, X in XII;
Singer in sod. (2002), JBC 277, 50. Petan et al., (2005) Biochemistry, 44.
350 - 2100 0,1 - 1000
430 - 2000
0,3 - 100
0,022 - 55
0,03 - 30
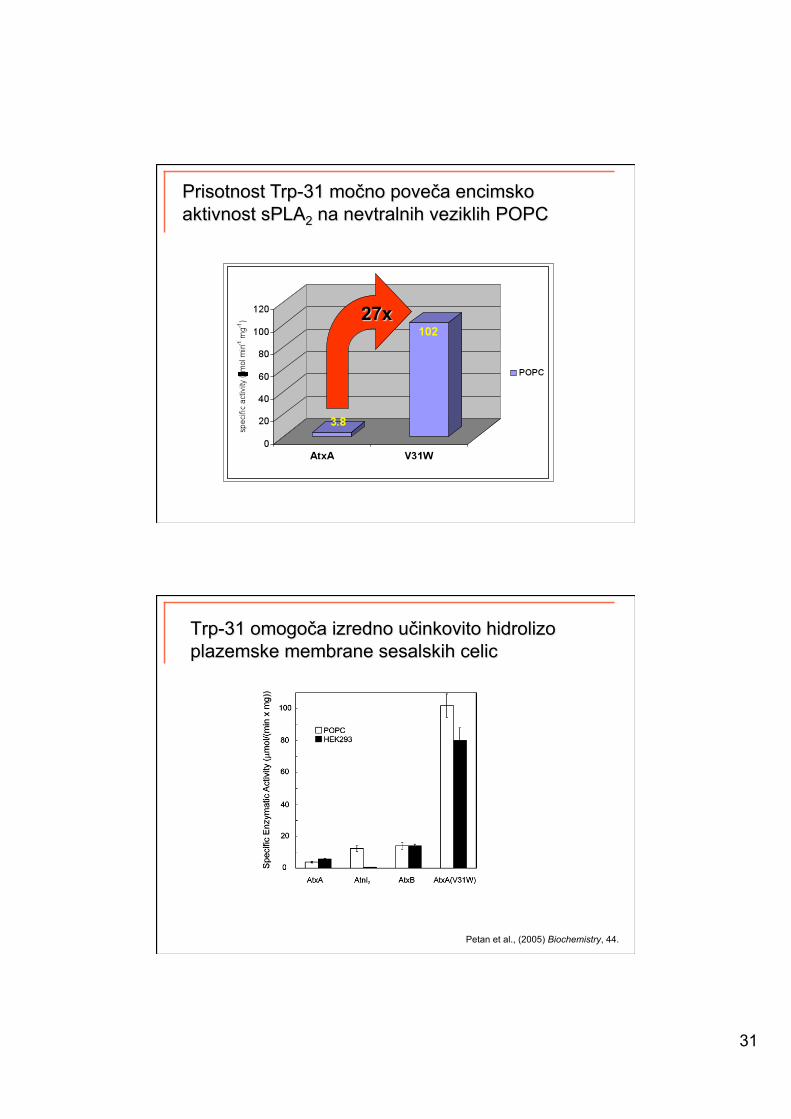
AtxA
• mutant AtxA(V31W)
31
Petan et al., (2005) Biochemistry, 44.

32
• Meritve v0 v odvisnosti od [Ca2+] z metodo FABP
• Vezikli poljubne sestave
• Določimo navidezno KCa (appKCa)
vmax [Ca2+] v0 =
[Ca2+] + appKCa
Ca2+
S
Ca2+
Kd sPLA2
KCa
KS*
appKCa = f (Kd, KCa, [V])
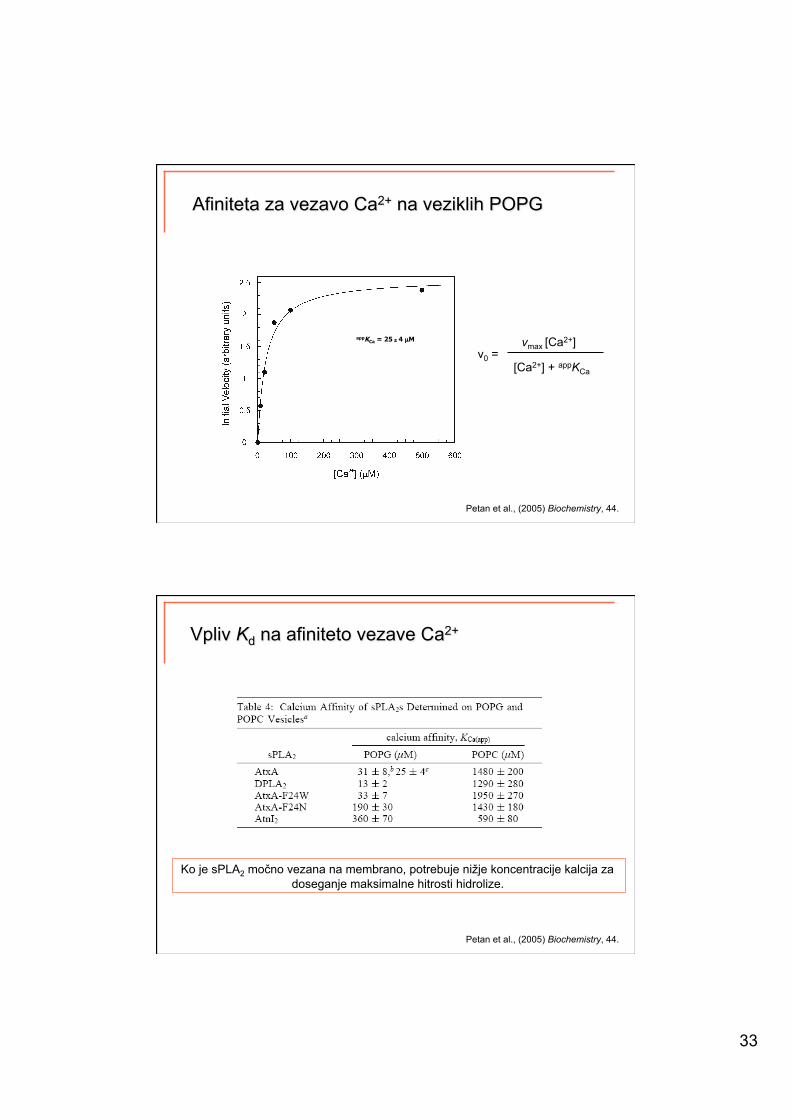
33
Petan et al., (2005) Biochemistry, 44.
appKCa = 25 ± 4 µM vmax [Ca2+] v0 =
[Ca2+] + appKCa
Petan et al., (2005) Biochemistry, 44.
Ko je sPLA2 močno vezana na membrano, potrebuje nižje koncentracije kalcija za doseganje maksimalne hitrosti hidrolize.

34

![[] dE =E [ ] - IJSbio.ijs.si/brigita/faculty/pdf/encimatika/P11 Pretvorba...presentacija razreda II lizosomska pot E-64 klorokin šibke baze (-) (-) antigenska presentacija razreda](https://static.fdocument.pub/doc/165x107/5fee9ee89ebc3679ce43a9c8/-de-e-pretvorba-presentacija-razreda-ii-lizosomska-pot-e-64-klorokin.jpg)
















