
Mécanismes de survenue des anomalies chromosomiques de ... · blocage de la fourche de...
73
Mécanismes de survenue des anomalies chromosomiques de structure JM DUPONT Laboratoire de Cytogénétique Hôpitaux Universitaires Paris Centre - Site Cochin Université Paris Descartes - Faculté de Médecine Paris
Transcript of Mécanismes de survenue des anomalies chromosomiques de ... · blocage de la fourche de...

Mécanismes de survenue des anomalies chromosomiques de structure
JM DUPONTLaboratoire de CytogénétiqueHôpitaux Universitaires Paris Centre - Site CochinUniversité Paris Descartes - Faculté de MédecineParis

Anomalies de structure Fréquence = 0,5% équilibrées 0,06% déséquilibrées

Anomalies de structure
Cassure double brin‣Pathologique
Fréquence = 0,5% équilibrées 0,06% déséquilibrées

Anomalies de structure
Cassure double brin‣Pathologique‣Physiologique
Fréquence = 0,5% équilibrées 0,06% déséquilibrées

CDB physiologiques
• CDB méiotiques déclenchées par Spo11
• Première étape de la recombinaison méiotique
• CDB déclenché par le complexe RAG
• Recombinaison V(D)J

Cassures double-brin (CDB) pathologiques
• ≈ 10 CDB / j / cell
• Stress réplicatif
• O2- provenant du métabolisme oxydatif -> radicaux hydroxyl (≈ 100/h/cell)
• Radiations ionisantes
• rayons γ, rayons X (≈ 300 Millions/personne/heure)
• Erreur de processus enzymatiques normaux
• Défaut de la Topoisomérase II (pas de ligation), action du complexe RAG en dehors de la région V(D)J
• Stress mécanique

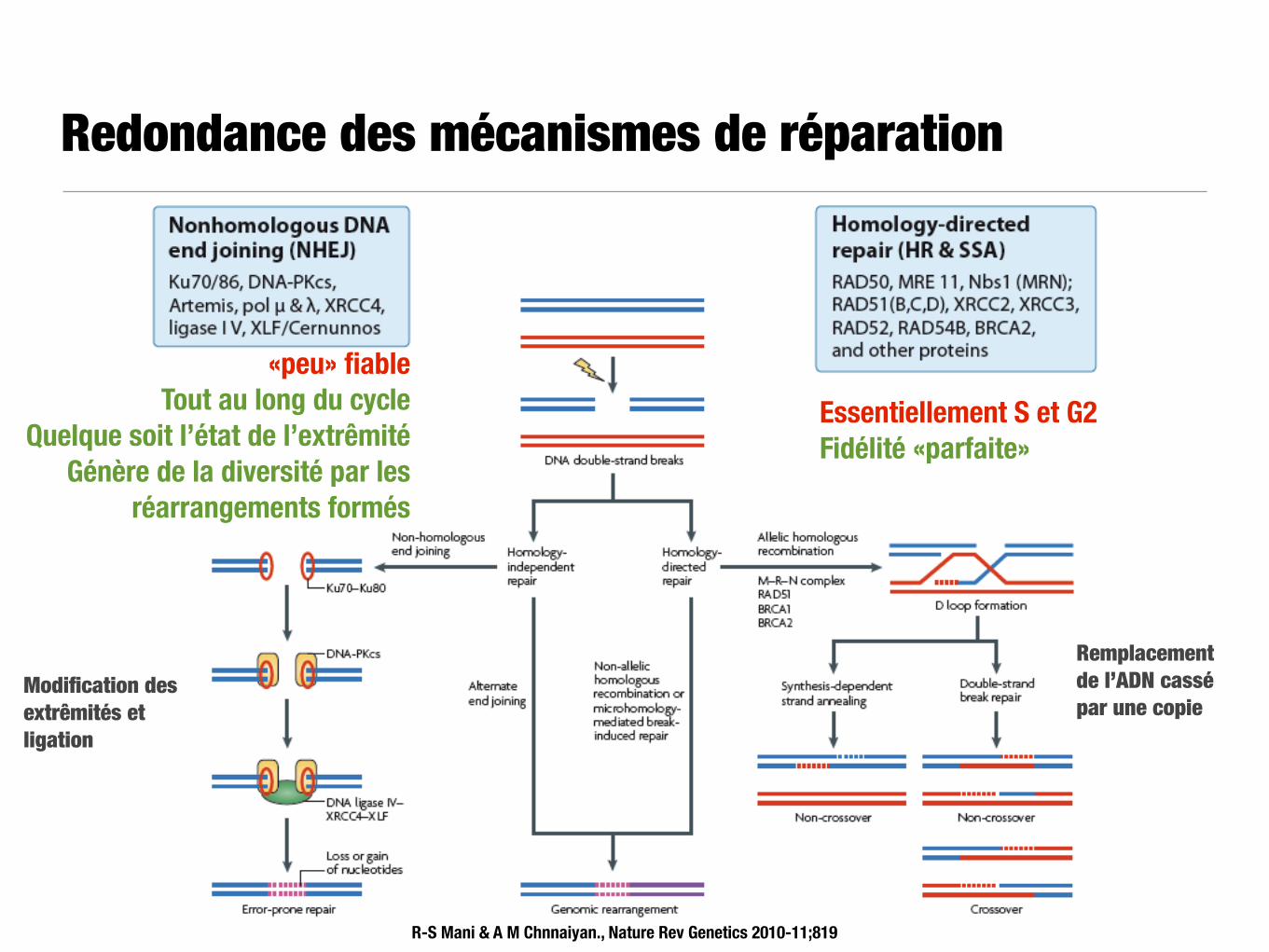
Redondance des mécanismes de réparation
R-S Mani & A M Chnnaiyan., Nature Rev Genetics 2010-11;819
Modification des extrêmités et ligation
«peu» fiable Tout au long du cycle
Quelque soit l’état de l’extrêmité Génère de la diversité par les
réarrangements formés
Redondance des mécanismes de réparation
R-S Mani & A M Chnnaiyan., Nature Rev Genetics 2010-11;819
Remplacement de l’ADN cassé par une copie
Essentiellement S et G2 Fidélité «parfaite»
Modification des extrêmités et ligation
«peu» fiable Tout au long du cycle
Quelque soit l’état de l’extrêmité Génère de la diversité par les
réarrangements formés

Réparation par fusion des extrémités• Action itérative et aléatoire de plusieurs
enzymes pour obtenir des extrémités complémentaires
• polymérases : Pol µ and λ• nucléases : artemis:DNA-PKcs
MR Lieber, Annu Rev Biochem, 2010-79;181

Réparation par fusion des extrémités• Action itérative et aléatoire de plusieurs
enzymes pour obtenir des extrémités complémentaires
• polymérases : Pol µ and λ• nucléases : artemis:DNA-PKcs
• Ligation finale
• DNA ligase IV ±XRCC4±XLF• Pas d’homologie étendue
nécessaire
MR Lieber, Annu Rev Biochem, 2010-79;181

Réparation par fusion des extrémités• Action itérative et aléatoire de plusieurs
enzymes pour obtenir des extrémités complémentaires
• polymérases : Pol µ and λ• nucléases : artemis:DNA-PKcs
• Ligation finale
• DNA ligase IV ±XRCC4±XLF• Pas d’homologie étendue
nécessaire
• «Cicatrice» possible au point de jonction
• Résultats variables pour un même point de cassure -> très flexible
• Diversité (réarrangement V(D)J)MR Lieber, Annu Rev Biochem, 2010-79;181

Réparation par recombinaison homologue
• Remplacement de la région lésée par une copie homologue
PJ Hastings et al., Nat Rev Genetics, 2009-10;551
Préparation des extrémités
Copie
Réparation
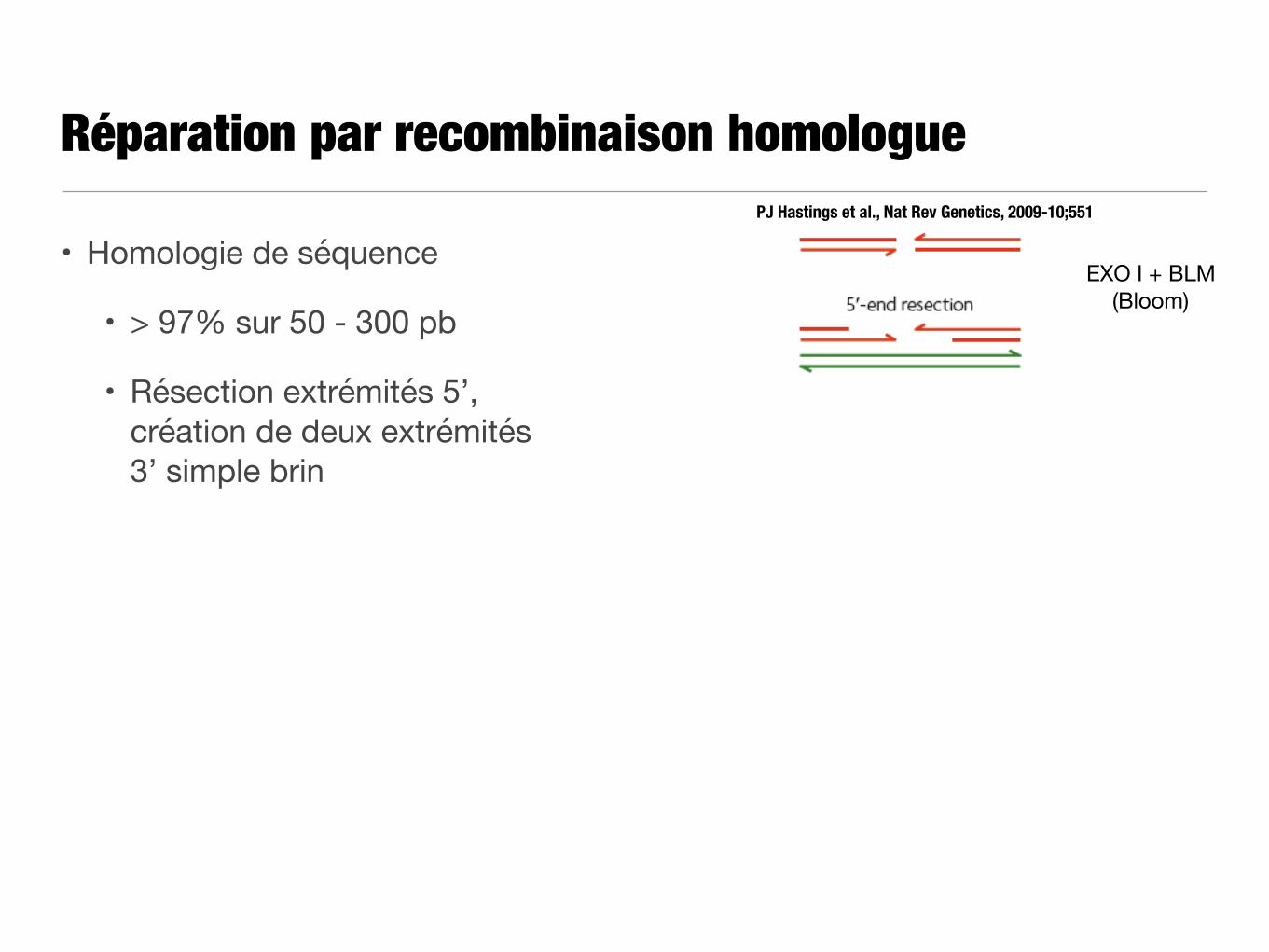
Réparation par recombinaison homologue
• Homologie de séquence
• > 97% sur 50 - 300 pb
• Résection extrémités 5’, création de deux extrémités 3’ simple brin
PJ Hastings et al., Nat Rev Genetics, 2009-10;551
EXO I + BLM (Bloom)

T Sugiyama et N Kantake, J Mpl Biol 2009-390;45
Réparation par recombinaison homologue
• Homologie de séquence
• > 97% sur 50 - 300 pb
• Stabilisation de l’ADN simple brin
• RPA, RAD52
PJ Hastings et al., Nat Rev Genetics, 2009-10;551
EXO I + BLM (Bloom)

Réparation par recombinaison homologueT Sugiyama et N Kantake, J Mpl Biol 2009-390;45
RAD51 + BRCA2 + cohesines
• Homologie de séquence
• Stabilisation de l’ADN simple brin
• Formation d’un nucléofilament
• RAD51-ssDNA
• Formation d’une synapse -> Recherche d’homologie
ME Moynahan, M Jasin, Nat Rev Mol Cell Biol, 2010-11;196
ATP

Réparation par recombinaison homologueT Sugiyama et N Kantake, J Mpl Biol 2009-390;45
RAD51 + BRCA2 + cohesines
DSBR Double Strand Break Repair
SDSA Synthesis-Dependant strand
annealing
• Homologie de séquence
• Stabilisation de l’ADN simple brin
• Formation d’un nucléofilament
• Recherche d’homologie
• Etape clé = invasion du brin homologue (D-loop) : RAD51

Réparation par recombinaison homologue : Crossing over / Conversion génique
• Homologie de séquence
• Etape clé = invasion du brin homologue (D-loop)
• Résolution
• DSBR (Holliday junction)
• Crossing over
• Conversion génique
PJ Hastings et al., Nat Rev Genetics, 2009-10;551
Endonucléase
Double Holliday Junction

Choix du mécanisme de réparation
• Compétition entre HR et NHEJ
• HR : S et G2
• NHEJ : Tout le cycle mais essentiellement G0 et G1
M Shrivastav et al., Cell Res, 2008-18;134

Choix du mécanisme de réparation
• Compétition entre HR et NHEJ
• HR : S et G2
• NHEJ : Tout le cycle mais essentiellement G0 et G1
• Eléments de régulation
• Partenaire disponible pour HR ++
• Régulation de la formation d’ADN simple brin par CDK (Cycline Dependant Kinases)
• Protéines participant au «choix» :
• BRCA1 favorise HR
• ATM favorise NHEJDaley J, Sung P. Cell 2013, Mar;49(5):840-1.
M Shrivastav et al., Cell Res, 2008-18;134

Et si il n’y a pas de deuxième extrémité ?
• Mécanismes de réparation normal
• Capture télomérique
• Synthèse d’un néo-télomère
Cassure télomérique

Et si il n’y a pas de deuxième extrémité ?
• Mécanismes de réparation pathologique
• Fusion des chromatides soeur -> dicentrique
• NHEJ
PJ Hastings et al., Nat Rev Genetics, 2009-10;551
Cassure télomérique

Et si il n’y a pas de deuxième extrémité ?
• Mécanismes de réparation pathologique
• Fusion des chromatides soeur -> dicentrique
• NHEJ
• Auto appariement sur des séquences répétées
Cassure télomériqueHermetz KE et al.. 2014. PLoS Genet 10:e1004139.

Et si il n’y a pas de deuxième extrémité ?
• Mécanismes de réparation pathologique
• Fusion des chromatides soeur -> dicentrique
• NHEJ
• Auto appariement sur des séquences répétées
• Formation d’un anneau
Cassure télomérique

Cassures par Erreurs de réplication
D Branzei et M Foiani, Nat Rev Mol Cell Biol, 2010-11;208
J Atkinson et P McGlynn, Nucleic Acids Res, 2009-37;3475
Arrêt de la fourche de réplication

Cassures par Erreurs de réplication
D Branzei et M Foiani, Nat Rev Mol Cell Biol, 2010-11;208
J Atkinson et P McGlynn, Nucleic Acids Res, 2009-37;3475
Arrêt de la fourche de réplication

Cassures par Erreurs de réplication
D Branzei et M Foiani, Nat Rev Mol Cell Biol, 2010-11;208
J Atkinson et P McGlynn, Nucleic Acids Res, 2009-37;3475
Arrêt de la fourche de réplication
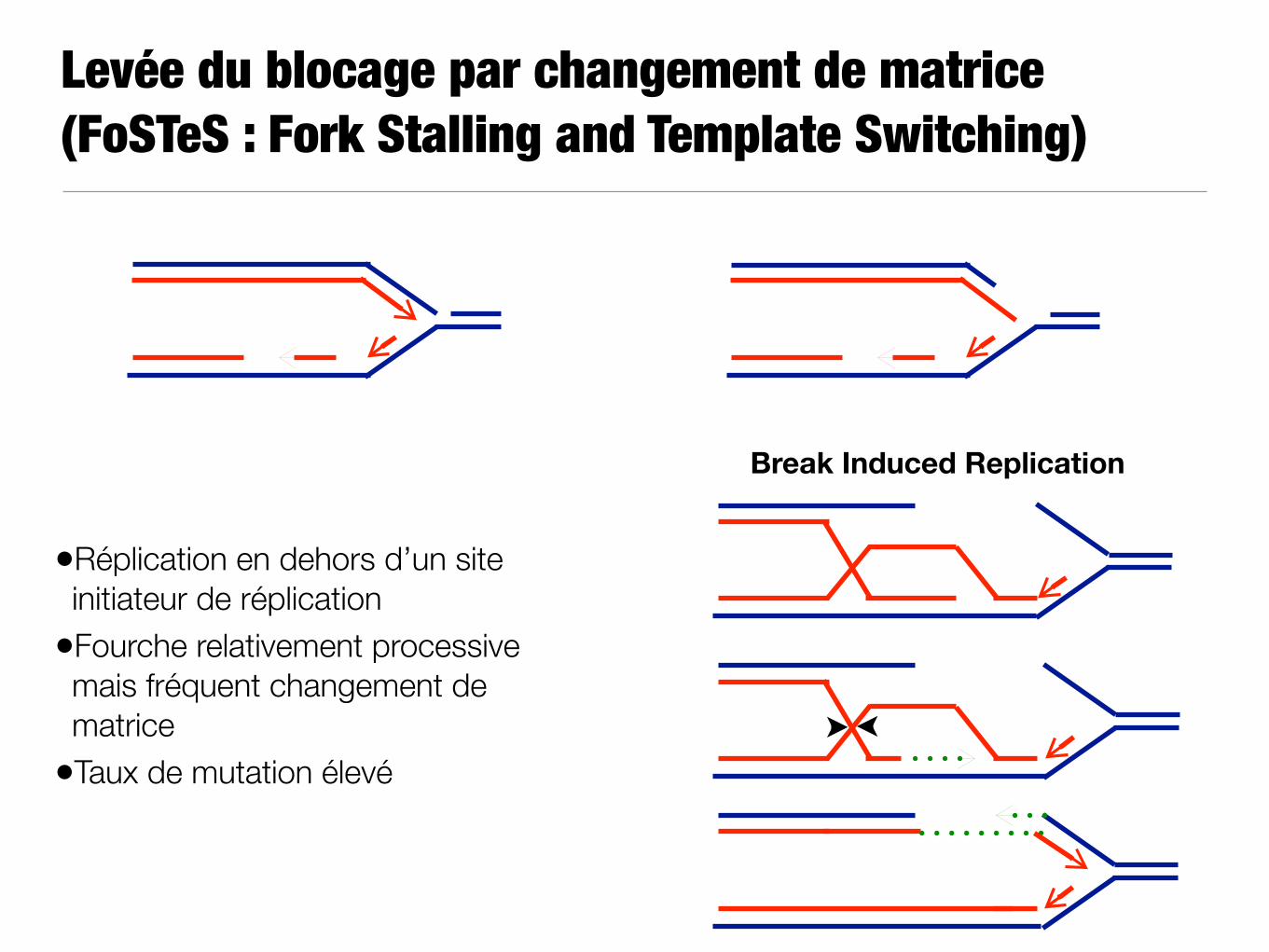
Levée du blocage par changement de matrice (FoSTeS : Fork Stalling and Template Switching)
Break Induced Replication
•Réplication en dehors d’un site initiateur de réplication
•Fourche relativement processive mais fréquent changement de matrice
•Taux de mutation élevé

Les anomalies de structure résultent d’un défaut de réplication ou de réparation de cassures !
• CNVs = Copy Number Variant
• Réarrangements Inter or intra chromosomiques

Clonage des points de cassure par séquençage haut débit
Weckselblatt B, Rudd MK. 2015. Human structural variation: mechanisms of chromosome rearrangements. Trends Genet

Les CNV résultent d’un défaut de recombinaison ou de réplication !
Modified from W Gu et al., PathoGenetics, 2008-I:4
COMPLEX
DeletionDuplicationTriplication

CNV récurrents : NAHR 2 fois plus de délétions que de duplications !
M Sasaki et al., Nat Rev Mol Cell Biol, 2010-11:182
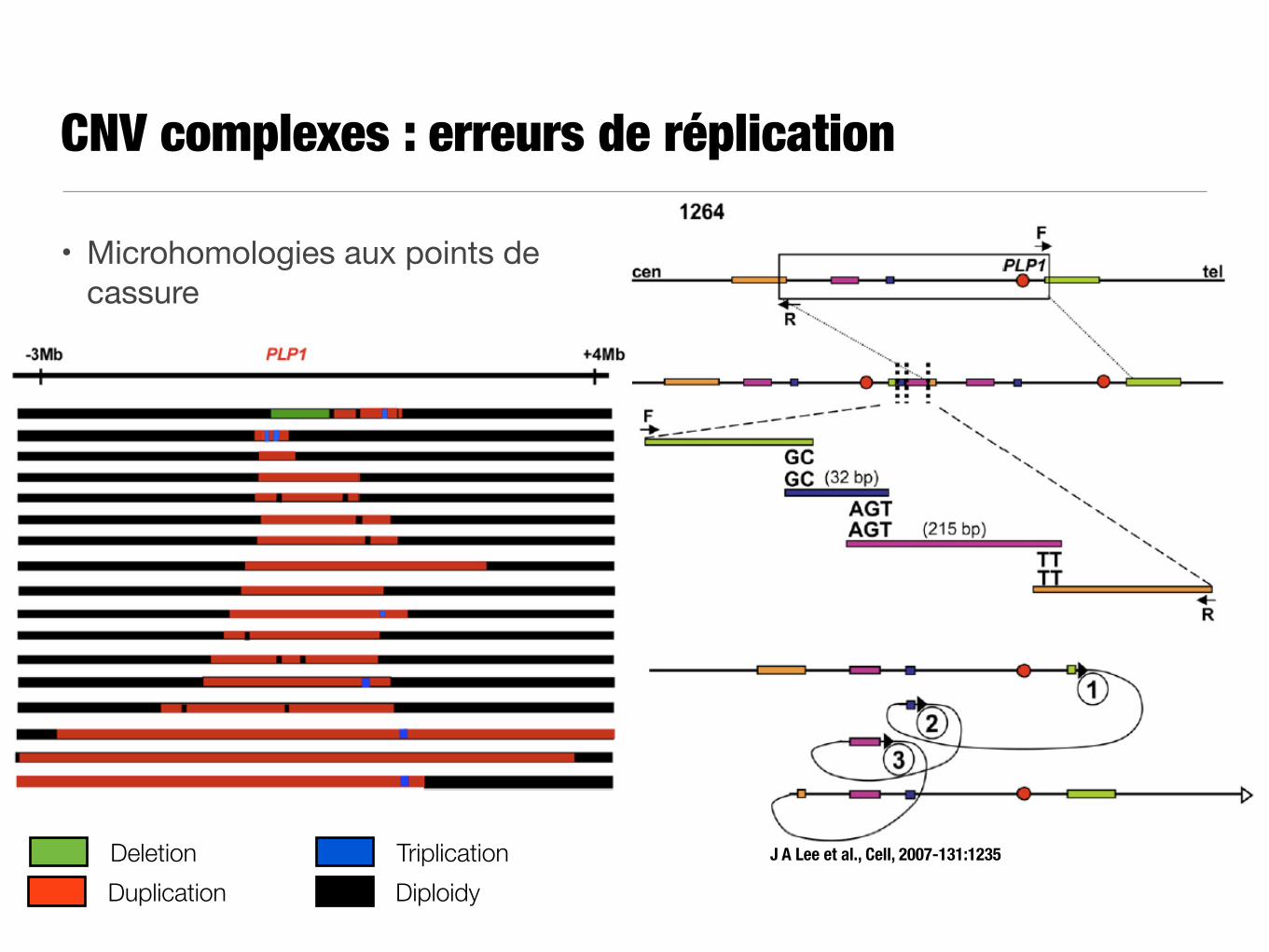
CNV complexes : erreurs de réplication
• Microhomologies aux points de cassure
J A Lee et al., Cell, 2007-131:1235
DuplicationDeletion Triplication
Diploidy

CNV complexes : FoSTeS / MMBIR
P J Hastings et al., Nat Review Genetics, 2009-10:551J A Lee et al., Cell, 2007-131:1235
• FoSTeS (Fork Stalling and Template Switching): blocage de la fourche de réplication sans cassure d’ADN
• MMBIR (Microhomology Mediated BreakInduced Replication): blocage de la fourche en raison d’une cassure d’ADN

CNV complexes : association de différentes erreurs de réplication / réparation
• Sauvetage de réplication
• Template switching
• NHEJ
• MMBIR
B Weckselblatt and M K Rudd., Trends in Genetics, 2015-31;587-599

Réarrangements inter or intra chromosomiques : NHEJ
Translocations réciproques ≈ 30% microhomology
CNVs ≈ 70% microhomology
Constitutional Cancer
Control individuals 1000 genome project
C. Chiang, J. C. Jacobsen, C. Ernst, C. Hanscom, et al., Nat Genet 44, 390-7 (2012).

Facteurs favorisant les remaniements
R-S Mani & A M Chnnaiyan., Nature Rev Genetics 2010-11;819

Facteurs favorisant les remaniements
• Proximité des molécules d’ADN
• Fréquence des cassures double brin
• Erreurs de réparation

Facteurs favorisant les remaniements
• Proximité des molécules d’ADN
• Territoires chromosomiques : taille, contenu en gènes, recouvrement
• Fréquence des cassures double brin
• Erreurs de réparation
K J Meaburn et al., Semin Cancer Biol 2007-17;80

Facteurs favorisant les remaniements
• Proximité des molécules d’ADN
• Territoires chromosomiques : taille, contenu en gènes, recouvrement
• Zones de transcription (Transcription factories), différenciation cellulaire
• Fréquence des cassures double brin
• Erreurs de réparation
E Gaffe, PLoS Genet, 2010-6;e1001011MR Branco and A Pombo, PLoS Biol, 2006-4;e138

Facteurs favorisant les remaniements
• Proximité des molécules d’ADN
• Territoires chromosomiques : taille, contenu en gènes, recouvrement
• Zones de transcription (Transcription factories), différenciation cellulaire
• Chronologie de réplication
• Translocations majoritairement entre séquences se répliquant en même temps
• Fréquence des cassures double brin
• Erreurs de réparation
E Gaffe, PLoS Genet, 2010-6;e1001011

Facteurs favorisant les remaniements
Triplex DNA Hairpins Quadruplex DNA
Cruciform DNA Left-handed Z-DNA
L Visser et al, Hum Mol Genet 2009-18;3579
• Proximité des molécules d’ADN
• Fréquence des cassures double brin
• Stress réplicatif
• Non-B DNA
• Erreurs de réparation

Facteurs favorisant les remaniements
• Proximité des molécules d’ADN
• Fréquence des cassures double brin
• Stress réplicatif
• Non-B DNA
• Séquences répétées
• Erreurs de réparation

Facteurs favorisant les remaniements
• Proximité des molécules d’ADN
• Fréquence des cassures double brin
• Stress réplicatif
• Non-B DNA
• Séquences répétées
• Erreurs de réparation
I Voineagu et al., PNAS 2008-105;9936

Cas de la t(11;22)(q23;q11)
• Palindromes en 11q 23 et 22q11
• ADN cruciforme
• Cassure double brin
• Réplication ? Autre ?
• Réparation par NHEJ
• Homologie qq nucléotides
• petite délétion au point de cassure

Facteurs favorisant les remaniements
• Proximité des molécules d’ADN
• Fréquence des cassures double brin
• Erreurs de réparation
• Structure du génome favorise les erreurs de recombinaison

Erreurs de recombinaison : NAHR (Non Allelic Homologous Recombination)
J A Bailey and E E Eichler, Nat Review Genet, 2006-7:552
• Favorisées par la présence de séquences répétées : LCR (Low Copy Repeats) ou duplicons
• ≈ 13% du génome
• > 95 - 97% homologie
• 50 - 300 pb
Duplicons inter chromosomiques
Duplicons intra chromosomiques

Mécanismes de protection contre les NAHR
• Inhibition des cassures au niveau des séquences répétées
• Homologie de séquence ≈ parfaite pour autoriser la recombinaison

Mécanismes de protection contre les NAHR
M Sasaki et al., Nat Review Mol Cell Biol 2010-11;182
• Inhibition des cassures au niveau des séquences répétées
• Homologie de séquence ≈ parfaite pour autoriser la recombinaison
• Utilisation préférentielle de l’homologue

Mécanismes de protection contre les NAHR
M Sasaki et al., Nat Review Mol Cell Biol 2010-11;182
• Inhibition des cassures au niveau des séquences répétées
• Homologie de séquence ≈ parfaite pour autoriser la recombinaison
• Utilisation préférentielle de l’homologue
• Contrôle de la qualité de l’homologie (RAD51 - ssDNA)

Facteurs favorisant les remaniements
• Proximité des molécules d’ADN
• Fréquence des cassures double brin
• Erreurs de réparation
• Structure du génome favorise les erreurs de recombinaison
• Rôle du stress cellulaire

Le stress cellulaire peut entraîner un déficit en Cohésines• 2 rôles principaux
• Cohésion des chromatides soeurs
• Organisation du génome
C H Haering, 2003K Nasmyth, An Rev Genet, 2009

Le stress cellulaire peut entraîner un déficit en Cohésines
S Covo et al., PLoS Genetic 2010-e1001006
• Rôle des cohésines
• Maintiennent les chromatides soeurs à proximité
• Favorisent la recherche d’homologie sur la chromatide soeur
• Stress facilite la recombinaison entre homologue au lieu d’une recombinaison entre chromatides sœurs

Stress cellulaire et défaut de recombinaison
• Mutation des protéines de réparation,
• Stress cellulaire
• Réduction RAD51ME Moynahan, M Jasin, Nat Rev Mol Cell Biol, 2010-11;196
ATP

MMBIR versus BIR : rôle de RAD51
T Sugiyama et N Kantake, J Mpl Biol 2009-390;45
MMBIRBIR
• MMBIR versus BIR
• BIR : Homologie étendue (> 50 pb)
• MMBIR : Microhomologie de quelques nucléotides suffit
• BIR nécessite RAD51 pour l’invasion du brin homologue
• MMBIR : mécanisme alternatif impliquant RAD52
• Le stress cellulaire réduit la quantité de RAD51 disponible

Conclusion
Cas sure

Conclusion
HR
NHEJBIR
Compétition Protéines régulatrices : BRCA1
Cycle cellulaire
Cicatrice au point de cassure
Cas sure
Restauration fidèle
Position dans le noyau
Structure du génomeEnvironnement cellulaire

Conclusion
HR
NHEJBIR
NAHR
Compétition Protéines régulatrices : BRCA1
Anomalies récurrentes mais pas seulement (ALU, LINE …)
Cycle cellulaire
Cicatrice au point de cassure
LCR
Cas sure
Restauration fidèle
Position dans le noyau
Structure du génomeEnvironnement cellulaire

Conclusion
HR
NHEJBIR
NAHR
FoSTeS / MMBIR
Compétition Protéines régulatrices : BRCA1
Anomalies récurrentes mais pas seulement (ALU, LINE …)
Cycle cellulaire
Stress cellulaire
Anomalies complexes
Cicatrice au point de cassure
LCR
Cas sure
Restauration fidèle
Position dans le noyau
Structure du génomeEnvironnement cellulaire

Mais comment expliquer çà ??
Stephens, P. J. et al. Cell 144, 27-40 (2011).

Hypothèse d’un évènement initial unique = chromothripsis
• Concernerait 2 à 3% des cancers
• jusqu’à 25% des cancers osseux
• Réparation par les mécanismes habituels (MMBIR / NHEJ)
• Très nombreux réarrangements créés en même temps

Accumulation d’anomalies ou chromothripsis ?
• Les cassures ne sont pas réparties aléatoirement
• Clustering sur un chromosome (ou une partie)
• Mais partenaires peuvent être distants
• Il n’y a pas de duplications, soit 1 copie, soit deux copies
Stephens, P. J. et al. Cell 144, 27-40 (2011).

Mécanisme : cassures multiples réparées par NHEJ
• Soit radiation ionisante / radicaux libres
• sur un chromosome en cours de condensation -> clustering des anomalies…
• Retard de migration en mitose
• Cassures lors de la cytokinèse (micro noyaux)
Kloosterman WP et al. Hum Mol Genet 2011, May 15;20(10):1916-24

Mais identification de situations incompatibles
1. Liu P, et al. Cell 2011, Sep 16;146(6):889-903.
• Patients porteurs de remaniements complexes
• duplications
• NHEJ possible si réplication préalable du chromosome
• triplications
• non explicables par NHEJ

Le phénomène de chromothripsis est-il spécifique aux cellules cancéreuses ?
• Patients avec caryotypes apparemment équilibrés et phénotype anormal
• 20% : CCR méconnus
Chiang C et al.. Nat Genet 2012;44(4):390-7.

Un nouveau concept : la survenue de réarrangements multiples suite à un évènement unique
• Cassures multiples • Erreurs multiples de réparation
Maher CA, Wilson RK. Cell 2012, Jan 20;148(1-2):29-32.
Chromothripsis Chromoanasynthesis
Soit

Un nouveau concept : la survenue de réarrangements multiples suite à un évènement unique• Chromothripsis
• NHEJ
Zhang CZ, Leibowitz ML, Pellman D. Genes Dev 2013, 27(23):2513-30.
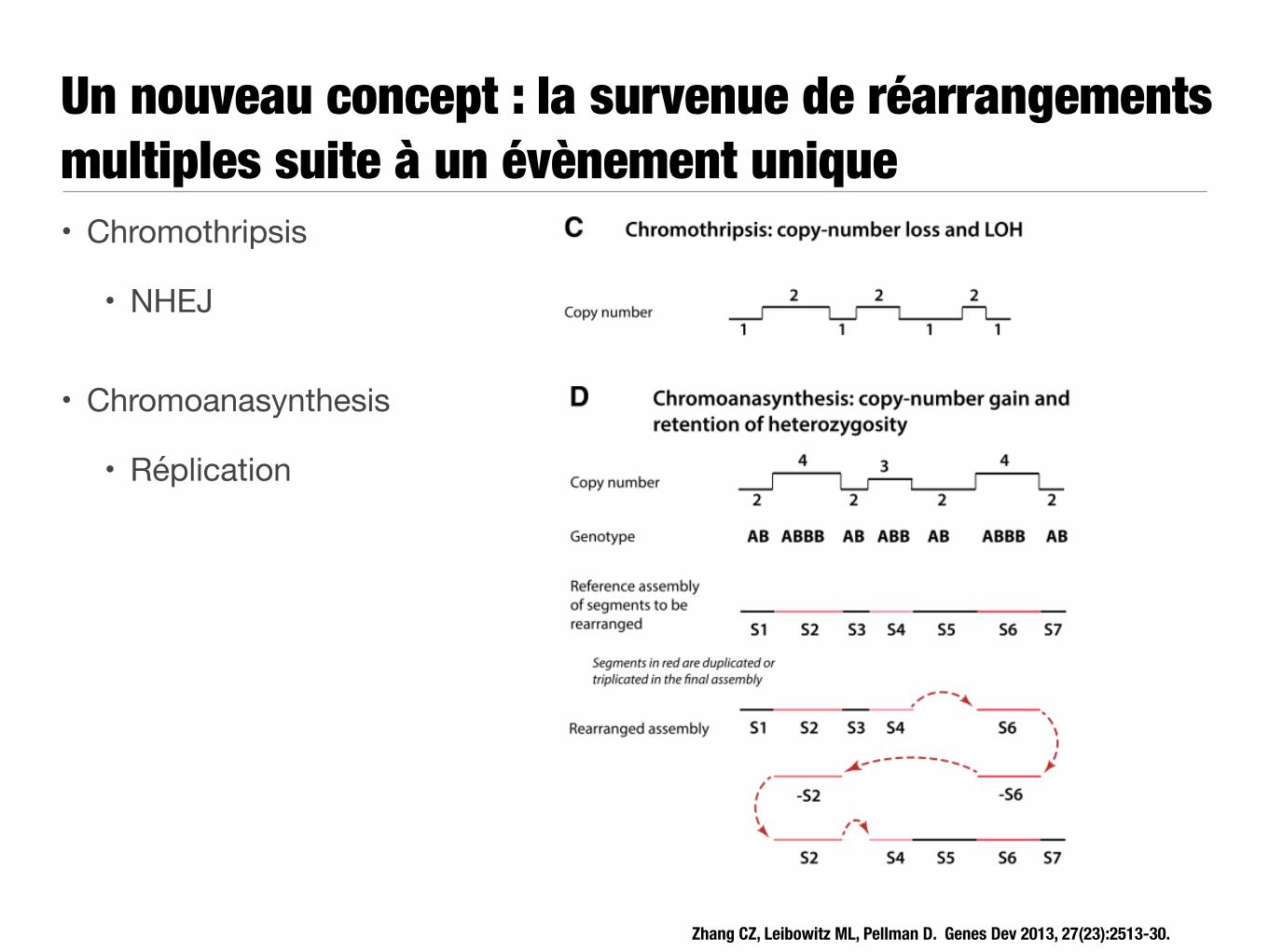
Un nouveau concept : la survenue de réarrangements multiples suite à un évènement unique• Chromothripsis
• NHEJ
• Chromoanasynthesis
• Réplication
Zhang CZ, Leibowitz ML, Pellman D. Genes Dev 2013, 27(23):2513-30.

Un nouveau concept : la survenue de réarrangements multiples suite à un évènement unique• Chromothripsis
• NHEJ
• Chromoanasynthesis
• Réplication
• Chromoplexy
• NHEJ
• Translocation induced chromothripsis
Zhang CZ, Leibowitz ML, Pellman D. Genes Dev 2013, 27(23):2513-30.

Une catastrophe initiale localisée
• Radiation ionisante sur des chromosomes condensés
Kloosterman WP et al. Hum Mol Genet 2011, May 15;20(10):1916-24
Chromothripsis

Une catastrophe initiale localisée
• Radiation ionisante sur des chromosomes condensés
• Erreur de réparation
• FoSTeS / MMBIR
• Stress cellulaire
Chromoanasynthesis

Dérapage des mécanismes de réparation ?
Chiang C et al.. Nat Genet 2012;44(4):390-7.
• Cas de l’insertion d’un transgène
• génération de remaniements complexes au sein du transgène et aux points d’insertion
• «Chromoanasynthesis»
• secondaire à une cassure unique
• dérapage des mécanismes de réparation

Une catastrophe initiale localisée
• Radiation ionisante sur des chromosomes condensés
• Erreur de réparation
• FoSTeS / MMBIR
• Stress cellulaire
• Micronuclei
• Cassures dans le matériel isolé
• Réintégration lors d’un cycle ultérieur
Zhang CZ, Leibowitz ML, Pellman D. Genes Dev 2013, 27(23):2513-30.

Conséquences du phénomène de chromoanagenesis
• Dans tous les cas remaniement en profondeur du génome
• En général mort cellulaire
• Déséquilibre d’expression génique (➘, fusion, ➚ (chromosomes minutes))
• avantage prolifératif et cancérisation• Explique les cancers sans instabilité génomique
• Remaniement complexe constitutionnel dans les formes «mineures»
• Rôle évolutif
• Spéciation ?


















