
,B-Galactosidase in Escherichia coli - Journal of Bacteriology

AIDS RESEARCH AND HUMAN RETROVIRUSESVolume 10, Number 6,1994Mary Ann Liebert, Inc., Publishers
Linear Epitopes of HIV-1, Presented as Hybrids withEscherichia Coli ß-Galactosidase or Synthetic Peptides
MARIA G. ISAGULIANTS,12 LIDIA L. SUKHANOVA,3 MICHAEL LEVI,4 ALKEKSEJ PH. BOBKOV,iTATJANA I. KALININA,1 ULLA RUDÉN,2 VALERIJ D. SMIRNOV,i and BRITTA WAHREN^
ABSTRACT
HIV-1B cell epitopes from gp41, the T cell epitope of p34pol, and a cluster of B and T epitopes from pl7«a« were
selected. The epitopes were presented as synthetic peptides and as either N- or C-terminal insertions into ß-galactosidase. Hybrids were efficiently expressed in E. coli and easily pu rifled when epitopes were inserted at theß-galactosidase C terminus. Sera from HIV-1-infected individuals reacted in peptide- and hybrid protein-basedenzyme-linked immunosorbent assays (ELISAs) mostly with the immunodominant site of gp41. The second siteof gp41 and also sites from pl7 and p34 appeared to be immunorecessive. A few of the HIV-1-positive sera exhib-ited several immunorecessive reactivities. HIV-1-positive sera from the former Soviet Union and Cuba had reac-tivities similar to those of American, African, and west European sera. Some sera could not be evaluated as
specifically HIV-1 seropositive because of their broad reactivities with a multitude of peptides and proteins, un-
related to HIV-1. Extensive tests were performed to define unspecific reactivities by absorption, blocking, andsandwich ELISAs. The application of the hybrid protein assay substantially improved the specificity of theELISA tests. Thus, hybrid protein-based ELISAs appeared to be more suitable than peptide-based ELISAs, es-
pecially for the evaluation of immunorecessive reactivities.
INTRODUCTION
1^ valuation of the antigenicity of short peptide sequences" "^within HIV-1 proteins is useful for the study of their biolog-ical and immunological significance. Screening of the HIV-1+sera for antibodies to the sequences of the HIV-1 proteins,thought so far to evoke low/rare (immunorecessive) antibody re-
sponse, is of interest. B cell epitopes predicted by hydrophilicityand ß turn locations are often immunorecessive. Eleven epitopeswere thus located in gpl60,'-3 but only 5 were actually markedby significant immune responses. Antibodies to the other sixwere present in only 1-3% of the HIV-1+ sera.4-5 Much evidencehas accumulated indicating that these predicted epitopes, al-though they might be silent in the native protein, can induce neu-
tralizing antibodies or contain T cell epitopes,58 the B epitopesproximate to or overlapping the latter (see Ref. 9, and referencestherein). Silence may be due either to poor recognition and/orpoor presentation to the host immune system, leading to the lowlevel or absence of antibody production.10 We attempted to de-
velop a procedure for the evaluation of such anti-epitope anti-bodies, keeping in mind their possible predictive value in AIDSprogression.
In the present article, peptides belonging to structurally and/orfunctionally significant regions of HIV-1, but supposedly evok-ing a low (immunorecessive) antibody response, were selectedand the presence of anti-peptide antibodies in panels of HIV-1+sera of different origin was studied by means of two indirect en-
zyme-linked immunosorbent assays (ELISAs). The peptides in-clude amino acids (aa) 738-748 of gp41 (RD) or its longer (aa732-746) variant (RD'), aa 940-949 of p34í>°' (P), and aa 99-115(EA) and 105-115 (EE) of pllgag. The epitopes originate fromconserved regions of HIV-1. The sequences are represented byconsensus sequences and are common to several strains of HIV-1 (HXB2, BRU, RF, SF2, and others), with some switches to thehomologous amino acids. ' '
Epitope RD is known to influence noticeably the host immuneresponses, lowering the natural peripheral blood mononuclearcell (PBMC) killer activity and changing lymphokine produc-
1D. I. Ivanovsky Institute of Virology, Academy of Medical Sciences, Gamaleja Str. 16, Moscow, 123098, Russia.2Department of Virology, Swedish Institute for Infectious Disease Control, 105 21 Stockholm, Sweden,institute of Viral Preparations, Academy of Medical Sciences, I-Dubrovskaja Str. 15, Moscow, 109088, Russia."Department of Clinical Virology, Karolinska Institute, 105 21 Stockholm, Sweden.
655

656 ISAGULIANTSETAL.
tion.12'13 Antibodies to that gp41 region neutralize various HIV-1 isolates, but not HIV-2, acting on the early stages of virus ab-sorbtion and penetration.
P, predicted as an epitope because of its location in a ß turn,stimulates the in vitro proliferation of T cells in 48% of HIV-1-infected individuals,5 indicating the presence of a T epitope.
EA and EE were selected from the cluster of neighboring Band T epitopes at the C terminal of pllgag (aa 78-132 ofpl7).14-16 Antibodies to aa 86-115 of pl7 were once reported tohave HIV-1-neutralizing activity.16 EA (EE) might also containa T cell epitope belonging to HLA-A2-restricted CD8 cytotoxicT lymphocytes (CTLs).17
The two indirect ELISA procedures used in this study includeone, based on recombinant proteins carrying HIV-1 epitopes,and another utilizing synthetic peptides, which represent thesame or overlapping HIV-1 determinants. As a convenient refer-ence the immunodominant epitope aa 593-604 of gp41 (LG),successfully used to screen sera for HIV-1,4,18,19 was chosen.
half sites Bam HI
partially filled In Incomplète
half altes Xhol
Incomplete partially tilled In
GG AT [_CCT A g|
c cG AGC
TCG AGTC
protruding ends of the vector
Protruding ends of DNA-duplexes, coding for the
HIV-1 antlgenlc determinants (
and plaamld vectors, prepared for their
cloning ( |_| ).
FIG. 2. Structure of the protruding ends of the DNA duplexes,coding for the HIV-1 epitopes, and the pCSG197-derived plas-mid vector, prepared for their cloning in monomeric form, aswas proposed by Khudjakov et al.23 In the pEX3-derived vectorXhol is exchanged for Sa/GI.
MATERIALS AND METHODS
OligonucleotidesOligonucleotides (19 to 55-mers), coding for the selected
HIV-1 epitopes, were synthesized manually using the H-phos-phonate approach20ab as described in Ref. 21. The oligonu-cleotide (oligo) sequences, composed of codons preferred byEscherichia coli,22 together with the amino acid epitopes en-coded by them, are given in Fig. 1. The DNA duplex cohesiveend structures, permitting cloning of their monomers in plasmid
A (+) 5' CCA CTA GGC ATT TOG GGT TGC TCT GGA AAA CTC ATT TGC OC
( ) 3' GT GAT CCG TAA ACC CCA ACG AGA CCT TTT GAG TAA ACG CGA GC
(+) 5' CCT GGA GAG CGT GAT CGC GAC CGT TCC ATT CGT CTT GC
{-) 3' GA CCT CTC GCA CTA GCG CTG GCA AGG TAA GCA GAA CGA GC
C (+) 5' CCG GAA GAG GAA CAG AAT AAA TCT AAG AAA AAG GC(-) 3' GC CTT CTC CTT GTC TTA TTT AGA TTC TTT TTC CGA GC
D (+> 5' CCG GAA GCA CTC GAC AAG ATC GAA GAG GAA CAG AAT AAA TCT AAG AAA AAG GC
(-) 3 GC CTT CGT GAG CTG TTC TAG CTT CTC CTT GTC TTA TTT AGA TTC TTT TTC CGAGC
E (+) 5' CCA GTT TAC TAT CGT GAC TCC CGT AAT CCT CTT GC
(-) 3' GT CAA ATG ATA GCA CTG AGG GCA TTA GGA GAA CGA GC
FIG. 1. Primary structure of DNA duplexes, coding for theHIV-1 epitopes from gp41 aa 593-604 (duplex A) and aa738-748 (B), from pl7 aa 105-115 (c) and aa 99-115 (D), andfrom p34 aa 940-949 (E). The peptide structure of the epitopes isshown above. Roman numerals designate the oligos synthesized.
vectors, were designed as in Fig. 2. Deprotected oligos were pu-rified by polyacrylamide gel electrophoresis (PAGE)-7 M urea
electrophoresis.Construction of recombinant plasmids
The DNA duplexes (Fig. 1) were cloned in the gene ofchimera "cro(l-7 aa)-preS of HBV-/ac/Z gene product" as a
preS substitute via pSCG19724 BamHl and Xhol restriction andpartial filling of the cohesive ends by the Klenow fragment ofDNA pol I and dATP, dGTP, and dTTP. Cloning into the pEX3polylinker25 was done accordingly, using BamHl and Sa/GI(Table 1). Vector DNA was purified by electrophoresis in 2%agarose NuSieve GTG (FMC, Philadelphia, PA). The DNA frag-ments were squeezed from agarose by repeated freeze-thawingfollowed by centrifugation, and sedimented from the supernatantwith ethanol. The ligation of 0.3-0.5 p.g of DNA duplex and0.1-0.2 p.g of vector was performed for 16 hr at 4°C. The compe-tent E. coli K-12 AHlAtrp, lacZ~am cells, carrying a copy of a
defective X prophage XN^, IN^ 53cI857AHl,26 were preparedvia the Hanahan method27 and transformed with the ligationmixture as in Ref. 28.
The recombinant clones were selected by colony blot hy-bridization. Plasmid DNA was isolated using an alkaline lysisprocedure28 and submitted to restriction enzyme mapping. ThepEX3-based recombinant plasmids were digested with restric-tion enzyme pairs: EcoRV/BamHl and EcoRV/Pstl (the Pstl sitewas next to Sa/GI,25 and was lost after ligation). The Southernblots of resulting gels were hybridized with predenatured 32P-la-beled DNA duplexes. The difference in hybridization patternsshows the insertions localized between the BamHl and Pstl sitesof the pEX3 polylinker. The pCSX analysis (where X = LG, RD,EA or P) was done accordingly, using PstllBaml and PstVXhol.The DNA of clones used for expression was sequenced by theSänger method as modified for plasmids.29
Hybridization with oligonucleotide probesThe terminal 32P label was introduced into preannealed DNA
duplexes by the Klenow fragment of DNA pol I and the four

EVALUATION OF IMMUNORECESSIVE REACTIVITIES 657
a-32P-dNTPs. The colony and Southern blotting were done on
Hybond N and Hybond N+, using the Amersham (ArlingtonHeights, IL) protocols. The hybridization conditions were as de-scribed by Church and Gilbert.30
Expression assayThe expression from the phage X PR promoter was induced by
raising the cell incubation temperature from 30 to 42°C. Afterthe 4- to 6-hr induction period, cells were harvested and dis-rupted by sonication. The lysates were analyzed by indirectELISA with HIV-1+ control sera and by 12.5% sodium dodecylsulfate (SDS)-PAGE.31 The gels were blotted onto nitrocellulosemembranes (0.45 mm; Schleicher & Schuell, Keene, NH) andhybridized with HIV-1-positive sera selected for having anti-gp41 reactivity, or with rabbit sera against synthetic peptidesfrom HIV-1, as in Ref. 32.
Pure ß-galactosidase (ß-Gal) (Promega, Madison, WI) pro-vided a concentration standard for the evaluation of hybridyields. Serial dilutions of ß-Gal were made and run in the ß-Gal-specific indirect ELISA with anti-ß-Gal monoclonal antibody(MAb; Promega). Dilutions in the range of 0.1 to 10 u.g/ml gavea linear concentration/absorption value dependence. The celllysate dilutions were prepared to give optical density (OD) val-ues in the range appropriate for reliable ß-Gal content quantita-tion. The overall protein content was estimated by the method ofBradford, using a protein assay dye reagent provided by Bio-Rad(Richmond, CA).
Inclusion body purification was done as in Ref. 28; proteinswere collected by centrifugation at 50,000g, as in Ref. 33, andused directly to coat ELISA plates. The construction and evalua-tion of antigenicity of the ß-Gal-gp41 hybrid are described inRef. 34.
Study populationThe ELISA study using recombinant proteins was performed
on a panel containing serum samples of 56 HIV-1-infected pa-tients from the former Soviet Union and 12 from Cuba. TheELISA study, based on peptides, was performed on a panel con-
taining serum samples from 19 U.S., 10 African (Ivory Coast),and 36 west European (including 7 Swedish) HIV-1-seroposi-tive individuals.
ELISA using recombinant proteins and Escherichia colicell lysates
Plates (Linbro, Cat. No. 76381-04 [Flow Laboratories,McLean, VA] or Cat. No. 3590-[Costar, Cambridge, MA]) were
coated overnight with recombinant proteins (2-5 pg/ml) or
pCSX/E. coli cell lysates (10-20 p.g/ml) in 0.05 M sodium car-
bonate buffer, pH 9.6, at 20°C. Positive and negative controlantigens were prepared in the same way and coated on the same
plate in equal amounts, as judged by ß-Gal and total protein con-
tent. The HIV-1+ sera were diluted 1:100, anti-HIV-1 IgG and(Du Pont [Wilmington, DE] HIV-1 strong positive control) seri-ally with phosphate-buffered saline (PBS) containing E. coli celllysate (1-3 mg/ml) and 0.05% Tween 20. Anti-peptide rabbitsera were diluted serially with PBS, containing 1% fish skingelatin. The indirect ELISA was conducted as in Ref. 28. Optical
density values twice that of the same serum on ß-Gal-coatedplates indicated positive HIV-1 status.
PeptidesThe peptide sequences are given in Table 2. Peptides LG and
P were also obtained as hybrids (LGTT and PIT) with T cell epi-tope aa 831-843 (TT)35 from tetanus toxoid, linked to the C ter-minus for further use in immunization. Longer peptides were
supposed to be more effective in plate coating. Peptides were
synthesized on 30 mg of resin (Fmoc, 0.3-0.5 mmol/g)(NovaSyn PR 500; Novabiochem, Switzerland) in polypropy-lene bags (160 mesh/in.; McMaster-Carr, Chicago, IL). Penta-fluorophenyl esters of Fmoc-protected amino acids were fromNovabiochem. The synthesis protocol36 was applied with hy-droxybenztriazole as a condensation catalyst. The cleavage anddeprotection were done with scavengers, recommended byApplied Biosystems (Foster City, CA). The peptides were ana-
lyzed by reversed-phase high-performance liquid chromatogra-phy (HPLC) on a Pep-S5 column (Pharmacia-LKB, Bromma,Sweden) using a linear gradient of 0-60% CH3CN in H20, con-
taining 0.1 % trifluoroacetic acid. Because HPLC showed pep-tides to be from 50 (LGTT) to 95% (P) pure, they were used forplate coating directly.
Peptides. Peptide EE conjugated to bovine serum albumin(BSA), RD', and SVVEEIVEEEG (MAL) from Plasmodiumfalciparum were kind gifts of lu. A. Semiletov and T. V. Firsova(D. I. Ivanovsky Institute of Virology, Moscow), M. Jansson andM. Sällberg (National Bacteriological Laboratory, Stockholm,Sweden), and N. Ahlborg (Immunology Institute, StockholmUniversity, Sweden).
Peptide ELISA
Micro-ELISA plates (MaxiSorb F96; Nunc, Roskilde,Denmark) were coated with 5-10 p:g of peptide/ml at 20°Covernight and at 4°C for 5 days and stored at 4°C until use. TheELISA was done as in Ref. 19; sample dilutions were made inPBS, containing 0.5% (w/v) BSA, 2% (v/v) whole goat serum,0.05% Tween 20, and 0.01% merthiolate. The dilution buffercomponents were supposed to take care of 94.5-95.3% of non-
specific binding.37 Values above the mean OD + 3SD of HIV-1-controls were considered positive for the sera exhibiting specificbinding.
Specificity assayELISAs on plates coated with peptides of non-HIV-1 origin,
such as TT or SVVEEIVEEEG from P. falciparum, or BSA,were performed to discover unspecific reactivities. Absorptionvalues in the non-HIV-1 peptide ELISA exceeding the cutoff ofthe negative control +7SD were indicative of high unspecificbinding serum activity.
The free peptide competition assay38 was carried out to estab-lish the specific character of the serum binding of the ß-Gal hy-brids and peptides. Up to 10 p.g/ml of the peptides, used as anti-gens or representing the epitope inserted in the hybrid, was used.A decrease in OD >50% was indicative of specific blocking.Blocking in the negative control was done accordingly.
Additional confirmatory tests for the sera with indefiniteHIV-1 status in the hybrid protein assay included indirect ELISA

658 ISAGULIANTS ET AL.
Table 1. Recombinant plasmids, determining the Synthesis of Hybrid Proteins on the Basis of ß-GALAcrosiDASE withInsertion of HIV-1 Antigenic Determinants (AD)
PLASMID, CARRYINGAD GENE
ENCODED HIV-1AD ( a.a. )
STRUCTURE OF THE HYBRID PROTEIN
pCSLG
pCSRD
pCSEE
pCSEA
pCSP
pEXLG
pEXRD
pEXEA
pEXP
593-604 gp41
738-748 gp41
105-115 p17
99-115 p17
940-949 p34
593-604 gp41
737-749 gp41
99-115 p17
940-949 p34
cro(1-7) AD
cro(1-7)
cro(1-7) AD
cro(1-7)
cro(1-7)
cro(1-7)
laclZ-gene product
AD laclZ-gene product
laclZ-gene product
AD laclZ-gene product
cro(1-7) AD laclZ-gene product
cro(1-7) laclZ-gene product AD
cro(1-7) laclZ-gene product AD
laclZ-gene product AD
laclZ-gene product AD
with dilutions of sera, and double-sandwich ELISA using anti-ß-Gal MAb (2 p:g/ml)-coated plates. Sera shown to be positive byone of the specificity tests were considered to be positive.
RESULTS
Construction of recombinant ß-galactosidases carryingHIV-1 antigenic determinants
The recombinant plasmids were constructed to encode for ß-galactosidase (ß-Gal) hybrids with HIV-1 epitopes inserted inthe 3' end of lacZ or the 5' end of cro-lacIZ genes. Plasmidsalong with the hybrids they encode are depicted in Table 1.
For conveying the detailed epitope recognition study the re-
combinant plasmids had to determine the efficient synthesis ofthe hybrids. The efficiency of transcription/translation is not af-fected if the coding DNA sequence is inserted into the 3' end ofthe chimeric gene, as it is in the pEX vector series.25'33.39 Hybridsof this type were shown to preserve the beneficial properties ofß-Gal, that is, the ability to expose inserted sequences and ease ofpurification thanks to the inclusion body formation (see, e.g.,Refs. 33 and 39). Indeed, recombinant E. coli K-12-AHlAtrp/pEXX, where X = LG, RD, EA, or P (Table 1), on am-
plification and thermoinduction expressed hybrid proteins withdual ß-Gal-HIV-1 epitope antigenic properties, as proved byELISA. The yields of the hybrids according to quantitativeELISA amounted of 10-15% of total cell protein content. Theyield of the ß-Gal hybrids with epitopes inserted at the N termi-nus was less than 1 %. The low level ofhybrid production may bedue to hybrid instability, caused by the disruption of the ß-Galcompact tertiary structure, as described in Ref. 33. But since for a
small insertion this disruption should not be too drastic, it wasmore probably caused by the changes in the mRNA 5'-end sec-
ondary structure, which might have involved the determinants oftranslation initiation or led to mRNA instability.40 The low levelof synthesis precludes detailed characterization of that type ofhybrid and their use for serum analysis.
Characterization of the recombinant ß-galactosidasescarrying HTV-1 epitopes
The ß-Gal hybrids with N-terminal LG, EA, and P insertionsand all hybrids with the C-terminal insertions reacted in indirectELISA with strong HIV-1+control sera. The OD values obtainedusing ß-Gal hybrids as antigens were 2 to 15 times higher thanthe ones obtained using carrier ß-Gal (Fig. 3). The reactivity was
50-100% blocked by the peptides, representing epitopes in-

EVALUATION OF IMMUNORECESSIVE REACTIVITIES
Table 2. Peptide Sequences, Locations, and Antigenicity, Determined by Peptide ELISA
Name
Peptide
Location ofHIV-1 part
(aa)Complete amino acid
sequence
Reactivity(%) ofHIV-1-positive
sera
Mean ODofpositivesamples
659
MeanODofHTV-
negativesera (+3 SD)
JB-4 aa 594-613 of gp41,HXB2 strain'9
LG aa 593-604 of gp414.5LGTT» aa 593-604 of gp41RD' aa 732-746 of gp41,
HXB2 strainEEC aa 105-115 of pl7EA aa99-115ofpl7P aa 940-949 of p343PTT»
GIWGCSGKLICTTAVPWNAS 100
LGIWGCSGKLIC 97
LGIWGCSGKLIC 97QY1KANSKF1GITEL0
GIEEEGGERDRDRSI 7.5
EEEQNKSKKKA 5
EALDKIEEEQNKSKKKA 7.5
VYYRDSRNPL 12.5VYYRDSRNPL
QY1KANSKFIGITEL
0.95(0.43-1.50)
0.68(0.42-0.94)
0.99(0.29-1.50)
0.26(0.24-0.29)
0.27(0.26-0.28)
0.47(0.39-0.53)
0.19(0.11-0.48)
0.15(0.04-0.29)
0.13(0.08-0.17)
0.08(0.06-0.11)
0.11(0.08-0.16)
0.16(0.09-0.22)
0.23(0.15-0.30)
0.05(0.03-0.08)
aUsed as a hybrid with 15-aa-long peptide sequences, representing a T cell epitope from tetanus toxoid QYIKANSKFIGITEL (TT).:bNon-HTV-1 part of the peptides is printed in cursive.cUsed for coating plates as a conjugate with BSA.
serted in (5-Gal. HIV-1- sera did not have any reactivity againstHIV-1 epitope-carrying hybrids (Fig. 3).
The strong reactivity of HIV-1+ sera to cro-ß-Gal-LG was
also demonstrated by immunoblot (Fig. 4). A major band withmobility characteristic of a 140-kDa protein and HIV-1 im-munospecificity appeared in immunoblots of the thermoinducedE. coli/pEXLG cell lysate with HIV-1+ serum (lane 3), whereasonly minor bands, caused by anti-E coli reactivities of humanserum, were present in lane 1, containing£. co///pEX3 thermoin-duced cell lysate. We have been unable so far to find HIV-1+ sera
having a comparably strong reactivity to hybrids carrying EA, P,orRD.
The HIV-1 immunospecificity of the hybrid cro-ß-gal-P was
also demonstrated by Western blot with rabbit serum against therelevant synthetic peptide. The rabbit serum against peptide Precognized a 140-kDa protein already in the lysate of nonin-duced E. coli/pEXP cells. The protein was the main product inthe lysate of induced cells. Immunoblotting uncovered partialdegradation of the hybrid on overnight thermoinduction. It didnot appear if the induction period was less than 6 hr.
Screening of the HIV-1-positive serum panel, usinghybrid ß-galactosidases with HIV-1 epitopes inserted atthe C terminus
The panel of HIV-1+ sera from Cuba and the former SovietUnion was subjected to indirect ELISA, using hybrid ß-Gal-coated plates. The results are summarized in Fig. 5. None of thesera exhibited any nonspecific reactions that could have maskedthe recognition of HIV-1 epitopes.
The majority of the positive sera (53 of 56 Soviet, 12 of 12Cuban) reacted with the hybrid cro-ß-Gal-LG. It was recognizedby all but one of the HIV-1+ sera reactive with the cro-ß-Gal-
gp41 hybrid (Fig. 5A). A number of HIV-1+ sera reacted withhybrids containing the following insertions: aa 738-748 of gp41(RD) (3.5%; 2 of 56 Soviet sera), aa 940-949 of p34 (P) (5.9%; 3Soviet, 1 Cuban serum), and aa 99-115 of pl7 (EA) (4.4%; 2Soviet, 1 Cuban serum) (Fig. 5B).
Representativity ofantibodies to selected HIV-1 epitopesstudied by the peptide-based indirect ELISA
The peptides, representing the same epitopes, are depicted inTable 2. A preliminary check for the occurrence of side reactivi-ties with the non-HIV parts of the peptide preparations (such as
TT) and BSA showed that none of the HIV-1- sera reacted withthem, whereas 38% of the HIV-1+ sera did. Nonspecific reac-
tions were exhibited by 2 of 19 American, 9 of 10 African, 7 of29 west European, and 7 of 7 Swedish sera. The value of the ab-sorption varied from 0.3 to 1.5, whereas the mean value of nega-tive sera +3SD never exceeded 0.3. Plates coated with BSA andpeptides of non-HIV origin, such as SVVEEIVEEEG from P.falciparum, as well as uncoated ELISA plates, were tested withthe same result. The reactivity could not be inhibited by the addi-tion of the free peptide to the sera at the dilution stage, which in-dicated the nonspecific character of the binding.
Blocking the plates prior to incubation with sera loweredsome, but not all, nonspecific reactions. The absorption values inELISAs using plates coated with peptides of non-HIV-1 origin,and blocked with 5% BSA, still reached 0.8-0.9 in three sera. In33% of the total sera it still exceeded the cutoff value (the mean
OD of the negative panel +3SD and, in 11 %, +7SD).Most of the sera (97%) reacted with the immunodominant
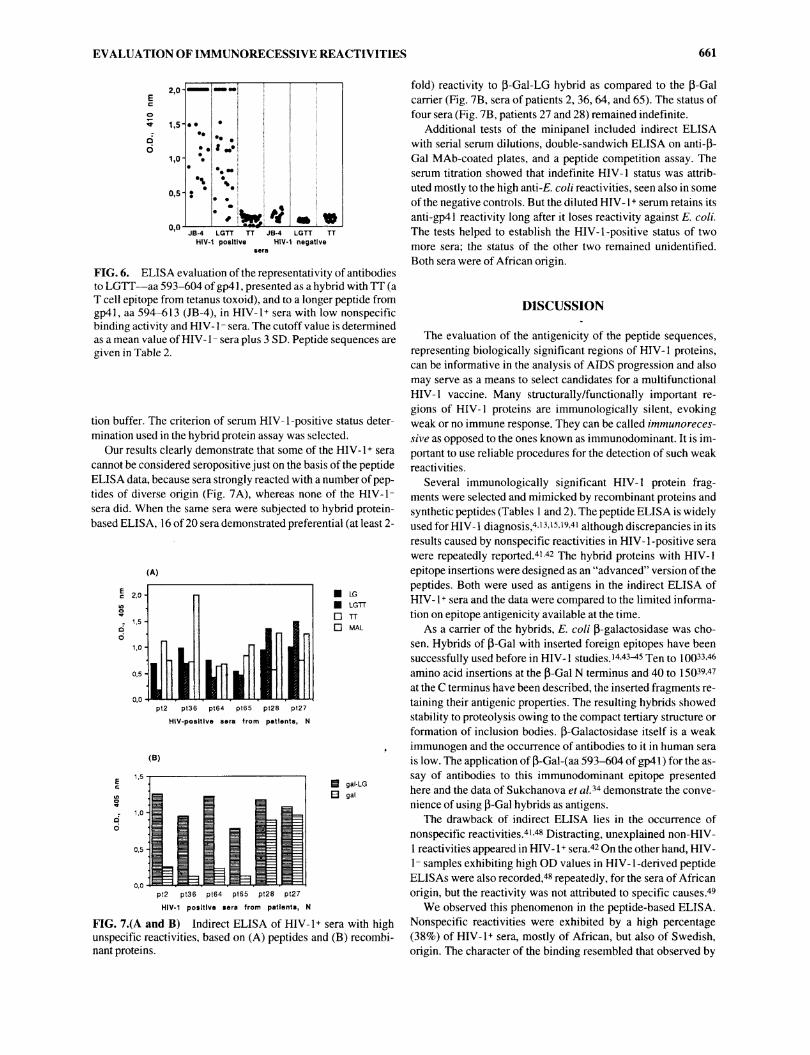
epitope LG, represented also by LGTT (Table 2), as shown inFig. 6. Addition of the next eight C-terminal amino acids plus

660 ISAGULIANTS ET AL.
HIV-1 positive sera, N HIV-1 negative sera, N 12 3 4
ÜL
LG-b-gal
P-b-galü b-gal
o 1,5
0,5
3 0,2E2 EA-b-gal ¿ b-gal o °'1
(d)
FIG. 3. Evaluation of the antigenicity of hybrids of ß-galac-tosidase with HIV-1 epitope insertion at the C terminus: aa
593-604 (LG-b-gal) (a) and aa 738-748 (RD-b-gal) (b) of gp41,aa 99-115 ofpl7s"g (EA-b-gal) (c), and aa 940-949 of p34H (P-b-gal) (d) via indirect ELISA with HIV-1+ (left) and HIV-U(right) sera. The results of the HIV-1+ and HIV-1- serumELISAs with carrier protein-ß-galactosidase (b-gal) are givenas negative controls.
FIG. 4. Immunoblotting of the E. coli/pEXLG lysates with anHIV-1+ serum: E. coli/pEXLG, clone 15, before thermoinduc-tion (lane 2) and after 4 hr of thermoinduction (lane 3); E.coli/pEXLG, clone 13, after thermoinduction (lane 4). Thelysate of the thermoinduced E. coli/pEX3 cells is given as a neg-ative control (lane 1 ). The arrow shows the position of ß-galac-tosidase hybrid.
aÖ
(a)
GP41 b-gal
peptide cyclization led to 1TJ0% recognition by HIV-1+ sera, as
was previously described.19The side reactivity masked weak interactions of anti-HIV-1
antibodies with the epitope sequences. Therefore the sera were
divided into a group exhibiting nonspecific reactivity (25 indi-viduals, 38%) and into those that did not (40 individuals, 62%).Only the latter group was subjected to the study of weak interac-tions. From 5 to 12.5% of HIV-1+ sera were found to possess re-activities with P, EE, EA, or RD; both the occurrence and anti-body titers were low (Table 2).
Application ofhybrid ß-galactosidase for the assay ofsera with high nonspecific reactivities
To make a direct comparison between two variants of ELISA,a group of 20 HIV-1+ sera from a mixed U.S.-African-Europeanpanel, exhibiting nonspecific reactivities, was selected. Thismini panel was subjected to analysis by both methods under con-
ditions used for peptide ELISA. Prior to the incubation with hu-man sera, antigen-coated plates were blocked with serum dilu-
(b)E °'4C
3 0,3
i °-2
0,1
0,0
m VA mEA RD b-gal
FIG. 5. ELISA evaluation of antibodies to HIV-1 epitopes,presented as insertions in the C terminus of ß-Gal, in the sera ofHIV-1-infected patients. The cutoff value is determined byELISA of HIV-1 + sera with E. coli ß-Gal (b-gal) and amounts to0.22. (a) ELISA data obtained using a hybrid of ß-Gal with theinsertion of aa 593-604 of gp41 (LG) and a hybrid of ß-galac-tosidase with gp41 (GP41 ); (b) ELISA data obtained using a hy-brid of ß-Gal with the insertions of aa 738-748 of gp41 (RD), aa940-949 of p34 (P), and aa 99-115 of pl7 (EA).
EVALUATION OF IMMUNORECESSIVE REACTIVITIES 661
• 1,5HoÓ
0,5
0,0
•*
*««
fed 4JB-4 LGTT TT JB-4 LGTT TT
HIV-1 positive HIV-i negative6era
FIG. 6. ELISA evaluation of the representativity of antibodiesto LGTT—aa 593-604 ofgp41, presented as a hybrid with TT (aT cell epitope from tetanus toxoid), and to a longer peptide fromgp41, aa 594-613 (JB-4), in HIV-1+ sera with low nonspecificbinding activity and HIV-1- sera. The cutoff value is determinedas a mean value of HIV-1- sera plus 3 SD. Peptide sequences are
given in Table 2.
tion buffer. The criterion of serum HIV-1-positive status deter-mination used in the hybrid protein assay was selected.
Our results clearly demonstrate that some of the HIV-1+ seracannot be considered seropositive just on the basis of the peptideELISA data, because sera strongly reacted with a number of pep-tides of diverse origin (Fig. 7A), whereas none of the HIV-Usera did. When the same sera were subjected to hybrid protein-based ELISA, 16 of 20 sera demonstrated preferential (at least 2-
(A)
pt2 pt36 pt64 pt65 pt28 pt27HIV-posltlve sera from patients, N
(B)
m gal-LGH gal
pl2 pt36 pt64 pl65 p128 pt27HIV-1 positive sera from patients, N
FIG. 7.(A and B) Indirect ELISA of HIV-1+ sera with highunspecific reactivities, based on (A) peptides and (B) recombi-nant proteins.
fold) reactivity to ß-Gal-LG hybrid as compared to the ß-Galcarrier (Fig. 7B, sera of patients 2, 36, 64, and 65). The status offour sera (Fig. 7B, patients 27 and 28) remained indefinite.
Additional tests of the minipanel included indirect ELISAwith serial serum dilutions, double-sandwich ELISA on anti-ß-Gal MAb-coated plates, and a peptide competition assay. Theserum titration showed that indefinite HIV-1 status was attrib-uted mostly to the high anti-E. coli reactivities, seen also in some
of the negative controls. But the diluted HIV-1+ serum retains itsanti-gp41 reactivity long after it loses reactivity against E. coli.The tests helped to establish the HIV-1 -positive status of twomore sera; the status of the other two remained unidentified.Both sera were of African origin.
DISCUSSION
The evaluation of the antigenicity of the peptide sequences,representing biologically significant regions of HIV-1 proteins,can be informative in the analysis of AIDS progression and alsomay serve as a means to select candidates for a multifunctionalHIV-1 vaccine. Many structurally/functionally important re-
gions of HIV-1 proteins are immunologically silent, evokingweak or no immune response. They can be called immunoreces-sive as opposed to the ones known as immunodominant. It is im-portant to use reliable procedures for the detection of such weakreactivities.
Several immunologically significant HIV-1 protein frag-ments were selected and mimicked by recombinant proteins andsynthetic peptides (Tables 1 and 2). The peptide ELISA is widelyused for HIV-1 diagnosis,4'13'15'19'41 although discrepancies in itsresults caused by nonspecific reactivities in HIV-1-positive sera
were repeatedly reported.41-42 The hybrid proteins with HIV-1epitope insertions were designed as an "advanced" version of thepeptides. Both were used as antigens in the indirect ELISA ofHIV-1 + sera and the data were compared to the limited informa-tion on epitope antigenicity available at the time.
As a carrier of the hybrids, E. coli ß-galactosidase was cho-sen. Hybrids of ß-Gal with inserted foreign epitopes have beensuccessfully used before in HIV-1 studies.1443^5 Ten to 10033-46amino acid insertions at the ß-Gal N terminus and 40 to 15039-47at the C terminus have been described, the inserted fragments re-
taining their antigenic properties. The resulting hybrids showedstability to proteolysis owing to the compact tertiary structure or
formation of inclusion bodies. ß-Galactosidase itself is a weakimmunogen and the occurrence of antibodies to it in human sera
is low. The application of ß-Gal-(aa 593-604 of gp41 ) for the as-
say of antibodies to this immunodominant epitope presentedhere and the data of Sukchanova et al?4 demonstrate the conve-
nience of using ß-Gal hybrids as antigens.The drawback of indirect ELISA lies in the occurrence of
nonspecific reactivities.41.48 Distracting, unexplained non-HIV-1 reactivities appeared in HIV-1 + sera.42 On the other hand, HIV-1- samples exhibiting high OD values in HIV-1-derived peptideELISAs were also recorded,48 repeatedly, for the sera of Africanorigin, but the reactivity was not attributed to specific causes.49
We observed this phenomenon in the peptide-based ELISA.Nonspecific reactivities were exhibited by a high percentage(38%) of HIV-1+ sera, mostly of African, but also of Swedish,origin. The character of the binding resembled that observed by

662 ISAGULIANTS ET AL.
Calfruny et al. in human anti-nuclear antibody-positive sera,where it was attributed to the high concentration of immunecomplexes.50 The side reactivity masked any possible weak in-teractions of the anti-HIV-1 antibodies with the epitopes. But itdid not affect so much the results of the indirect hybrid protein-based ELISA. The application of the latter diminished the num-
ber of sera with indefinite status from 38% seen in peptide-basedELISAs to 6%, and, after confirmatory tests, to 3%.
One of the reasons for this may lie in the efficiency of the platecoating. The optimal conditions are known to depend on theproperties of the peptides (see, e.g., Ref. 51). Therefore in thepeptide-based ELISA, control of the plate-coating efficiency isessential. Hybrid proteins in general are better coating agentsthan peptides, leaving no free space on the ELISA plate surfacefor nonspecific plastic-protein interactions.
The choice of negative controls is also important. The ELISAof the HIV-1 -seropositive panel with the protein carrier ß-Gal re-veals the presence of any side reactions in the sera from the verybeginning. ß-Galactosidase is a better negative control than anyof the peptides ofnon-HIV-1 origin, because its properties are al-most identical to the ones of the positive antigen (hybrid ß-Gal),whereas the properties of the peptides used as positive and nega-tive antigens may vary.
With the exception of the sera exhibiting nonspecific reactivi-ties, both variants of indirect ELISA gave the same recognitionfrequencies for immunodominant, as well as immunorecessive,HIV-1 epitopes, showing that both peptides and hybrid proteinswith these peptide sequences inserted successfully mimick lin-ear HIV-1 epitopes. Hybrid proteins appear to be more conve-
nient antigens in the study of weak immune interactions, makingit less sensitive to interference by nonspecific reactivities.
Our data on the reactivity of the immunodominant epitopeLG, obtained both in peptide and hybrid protein ELISAs, matchthose ofGnann et al.,4 except for the total lack of anti-gp41 reac-
tivity in two sera originating from the former USSR. Their HIV-1-positive status was confirmed by indirect assay with recombi-nant ß-Gal-p24 protein. Typically this lack of reactivity is due to
incipient seroconversion and was shown to give place abun-dantly to reactivity with gp41 and other viral antigens.41 The ex-istence of HIV-1-positive sera negative for gp41 (2 of 68; Fig.5A) shows the necessity for using more than one antigen in theevaluation of serum HIV-1-related status.
The occurrence of antibodies to the sequences comprising Tcell or B cell epitopes, recognized by neutralizing antibodies,was low (5-12.5%) and independent of the origin of the serum.
The findings on the occurrence of antibodies to immunoreces-sive HIV-1 epitopes adds to the limited data available from ear-
lier studies.5'13 Analysis of the data from both panels revealedthat the immunorecessive reactivities were attributed to the samesera with the exception of anti-P antibodies, which occurredmore often. This is remarkable, because the presence of antibod-ies to certain HIV-1 epitopes was reported to be of prognosticvalue for disease progression. Thus the decline of anti-p24 (re-viewed in Ref. 41), pl7,52 and gpl20 V3 loop53 antibodies wasobserved in parallel with the progression of severe AIDS symp-toms. As no single peptide-specific antibody has so far been re-
ported to be a definite indicator ofpatient status or prognosis, it isnecessary to screen sera for the coincident presence (or absence)of antibodies to certain epitopes along with a survey of AIDSprogression.
ACKNOWLEDGMENTS
This work was supported in part by a grant from the SwedishInstitute, Stockholm, Sweden. We are grateful to Prof. S. Brittonfor constant support, and to Dr. M. M. Garaev, Dr. V.-A.Sundqvist, and Dr. E. V. Kazenova for useful discussions. Wethank Dr. Yu. A. Semiletov and T. V. Firsova for the gift of two
peptides, Dr. R. P. Pavlutchenkova for obtaining the hyperim-mune rabbit serum against one of the peptides, Dr. S. Cox forhelp in manuscript preparation, and Dr. S. A. Arakelov for shar-ing his experience on HIV-1 ELISA.
REFERENCES
1. Modrow S, Hahn BH, Shaw GM, Gallo RC, Wong-Staal F, andWolf H: Computer-assisted analysis of envelope protein sequencesof seven human immunodeficiency virus isolates: Prediction of anti-genic epitopes in conserved and variable regions. J Virol1987;61:570-578.
2. Steinberg MJE, Barton GJ, Zvelebil MJ, Cookson J, and CoatesARM: Prediction of antigenic determinants and secondary struc-tures of the major AIDS virus proteins. FEBS Lett1987;218:231-237.
3. Krchnak V, Mach O, and Maly A: Computer prediction of potentialdeterminants from protein amino acid sequence. Anal. Biochem1987;165:200-207.
4. Gnann IW, Schwimmbeck PL, Nelson JA, Truax AB, and OldstoneMBA: Diagnosis of AIDS by using a 12-amino acid peptide repre-senting an immunodominant epitope of the HIV. J Infect Dis1987;2:261-267.
5. Schrier RD, Gnann JW, Landes R, Lockshin C, Richman D,McCutchan A, Kennedy C, Oldstone MBA, and Nelson JA: T cellrecognition of HIV synthetic peptides in a natural infection. JImmunol 1989;142:116-1176.
6. Ho DD, Kaplan JC, Raskauskas IE, and Gurney ME: Second con-
served domain of gpl20 is important for HIV infectivity and anti-body neutralization. Science 1988;239:1021-1023.
7. Ho DD, Samgadharan MG, Hirsch MS, Schooley RI, Rota TR,Kennedy RC, Chanh TC, and Sato L: HIV neutralizing antibodiesrecognize several conserved domains on the envelope glycopro-teins. J Virol 1987;61:2024-2028.
8. Cease KB, Margalit H, Cernette JC, Putney SD, Robey WG, QuangC, Streicher HZ, Fischinger PJ, Gallo RC, De Lisi C, and BerzofskyJA: Helper T-cell antigenic site identification in AIDS virus gpl20envelope protein and induction of immunity in mice to the nativeprotein using a 16-residue synthetic peptide. Proc Nati Acad SeiUSA 1987;84:4249^t253.
9t Graham S, Wang ECY, Jenkins O, and Borysiewicz LK: Analysis ofthe human T-cell response to picomaviruses: Identification ofT-cellepitopes close to B-cell epitopes in poliovirus. J Virol1993;67:1627-1637.
10. Miedema F, Tersmette M, and van Lier RAW: AIDS pathogenesis:A dynamic interaction between HIV and the immune system.Immunol Today 1990; 11:293-297.
11. Myers G, Rabson A, Josephs SF, Smith TF, Berzofsky JA, andWong-Staal F (Eds.): Human Retroviruses and AIDS. Los AlamosNational Laboratory, Los Alamos, New Mexico, 1989.
12. Mertins S, Cauda R, Tubarello M, Ortona L, Kennedy R, Rubin M,Pizzo P, Chahn T, and Kanda P: HIV gp41 synthetic peptides affectthe natural immune response. Vlth Int. Conf. on AIDS, SanFrancisco, California, 1990;3:160.
13. Cauda R, Tumbarello M, Ortona L, Kennedy RC, Chanh TC, TyringS, and Kanda P: HIV gp41 synthetic peptides influence normal pro-

EVALUATION OF IMMUNORECESSIVE REACTIVITIES 663
duction of cytokines. Vlth Int. Conf. on AIDS, San Francisco,California 1990:3:143.
14. Mathiesen T, Broliden P-A, Rosen J, and Wahren B: Mapping ofIgG subclasses and T-cell epitopes on HIV proteins by syntheticpeptides. Immunology 1989;67:453-459.
15. Kchaitov RM, Sidorovitch IG, Pavlikov SP, Nikolaeva IA,Ivastchenko ME, Andreev SM, and Skljarov LJ: The use of syn-thetic peptides in the diagnostics of the HIV infection. Immunologia(USSR) 1990;2:15-20 [in Russian].
16. Sarin PS, Sun DK, Thornton AH, Naylor PH, and Goldstein AL:Neutralization of HTLV-III/LAV replication by antiserum to thy-mosine a 1. Science 1986;232:1135-1137.
17. Achour A, Picard O, Zagury D, Sarin PS, Gallo RC, Naylor PH, andGoldstein AC: HGP-30, a synthetic analogue of HIV pi 7, is a targetfor cytotoxic lymphocytes in HIV infected individuals. Proc NatiAcad Sei USA 1990;87:7045-7049.
18. Wang JJG, Steel S, Wisniewolsky R, and Wang CJ: Detection of an-
tibodies to human T-lymphotropic virus type III by using a syntheticpeptide of 21 amino acid residues corresponding to a highly anti-genic segment of gp41 envelope protein. Proc Nati Acad Sei USA1986;83:6159-6163.
19. Broliden PA, Rudén U, Ouattara AS, de Thé G, S0lver E, Trojnar J,and Wahren B: Specific synthetic peptides for detection of and dis-crimination between HIV-1 and HIV-2 infection. AIDS1991;4:952-958.
20a. Froehler BC, Ng PC, and Matteucci MD: Synthesis of DNA via de-oxynucleoside H-phosphonate intermediates. Nucleic Acids Res1986;14:5399-5407.
20b. Garegg PJ, Redberg T, Stawinski J, and Strömberg R: NucleosideH-phosphonates. III. Chemical synthesis of oligodeoxyribonu-cleotides by the hydrogen phosphonate approach. Tetrahedron Lett1986;27:4051-4054.
21. Isaguliants MG, Lutchin SV, and Smirnov VD: Chemical synthesisand enzymatic assembly of DNA, coding for the epitopes of humanimmunodeficiency virus type I. Khimia Prirodnikh Soedinenij(USSR) 1990;3:393-399. [in Russian]
22. Wong JT-F and Cedergren R: Natural selection versus primitivegene structure as determinant of codon usage. Eur J Biochem1986;159:175-180.
23. Khudjakov JE, Samoshin VV, Kalinina TI, and Smirnov VD: Theconstruction of repeated sequences with the use of syntheticoligonucleotides. Materials of the conference, New Trends inBiotechnology, Puschino-na-Oke, 1988, 108. [in Russian]
24. Khudjakov JE, Kalinina TI, Nepljueva VS, and Smirnov VD: Thecorrelation between translation initiation efficiency and secondarystructure of mRNA in the hybrid gene cro-lacLZ. MolecularnayaBiología (USSR) 1987;21:1504-1512. [in Russian]
25. Stanley KK and Luzio JP: Construction of a new family of high effi-ciency bacterial exptression vectors: Identification of cDNA clonescoding for human liver proteins. EMBO J 1984;3:1429-1434.
26. Bernard H-U, Remaut E, Hershfield MV, Das HK, Helinski DK,Yanofsky C, and Franklin N: Construction of plasmid cloning vehi-cles that promote gene expression from the bacteriophage P pro-moter. Gene 1979;5:59-76.
27. Hanahan D: Studies of transformation of E. coli with plasmids. JMol Biol 1983;166:557-580.
28. Maniatis T, Fritsch EF, and Sambrook J: Molecular Cloning: ALaboratory Manual, 2nd ed. Cold Spring Harbor Laboratory Press,Cold Spring Harbor, New York, 1989.
29. Korneluk RG, Quan F, and Gravel R: Rapid and reliable deoxy se-
quencing of double-stranded DNA. Gene 1985;40:317-323.30. Church GM and Gilbert W: Genomic sequencing. Proc Nati Acad
Sei USA 1984;81:1991-1995.31. Laemmli UK: Cleavage of the structural proteins during the assem-
bly of the head of bacteriophage T4. Nature (London) 1970;227:680-685.
32. Davis LG, Dibner MD, and Baltley F: Basic Methods in MolecularBiology. Elsevier, New York, 1986.
33. Stanley K: Solubilization and immune detection of ß-galactosidasehybrid proteins carrying foreign antigenic determinants. NucleicAcids Res 1983;11:4077^1092.
34. Sukchanova LL, Goltsov VA, Zvonarev AJ, Sazonov AE, KarasevaEV, Alatortseva GI, Amiantova II, and Zajtsev IZ: Synthesis of theantigenically active HIV-1 polypeptides in bacterial systems. In:Genetic and Cellular Engineering in the Solution of FundamentalProblems of Biotechnology, Vol. 1. Semenova VF (Ed). Tartu,USSR, 1989, pp. 113-119. [in Russian]
35. Demotz S, Lanzavecchia A, Eisel U, Niemann H, Widmann C, andCorradin G: Delineation of several DR-restricted tetanus toxin T cellepitopes. J Immunol 1989;142:394-402.
36. Sällberg M, Rudén U, Magnius LO, Norrby E, and Wahren B: Rapid"tea-bag" peptide synthesis using 9-Fmoc protected amino acids ap-plied for antigenic mapping of viral proteins. Immunol Lett1991;30:59-68.
37. Vogt RF, Phillips DL, Henderson-LO, Whitfield W, and SpiertoFW: Quantitative differences among various proteins as blockingagents for ELISA microtiter plates. J Immunol Methods1987;101:43-50.
38. Steward MW, Stanley CM, Dimarchi R, Mulcahy G, and Doel TR:High-affinity antibodies induced by immunization with a syntheticpeptide is associated with protection of cattle against foot-and-mouth disease. Immunology 1991;72:99-103.
39. Broekhuijsen MP, van Rijn JMM, Blom AJM, Powels PH, Enger-valk BE, Brown F, and Francis MJ: Fusion proteins with multiplecopies of the major antigenic determinant of foot-and-mouth diseasevirus protect both the natural host and laboratory animals. J GenVirol 1987;68:3137-3143.
40. Hargrove JL and Schmidt FH: The role of mRNA and protein stabil-ity in gene expression. FASEB J 1989;3:2360-2370.
41. Lindhardt BO: Serological investigations of HIV infection. DanishMed Bull 1991;38:22-36.
42. Chrystie I, Palmer S, Voller A, and Banatvala J: False positive serol-ogy and HIV infection. Lancet 1993;341:441^142.
43. Aepinus C, Voll R, Broker M, and Fleckenstein B: A rev/beta-galac-tosidase fusion protein binds in vitro transcripts spanning the rev-re-
sponsive element of HIV-1. FEBS Lett 1990;263:217-221.44. Marcus-Secura CJ, Woerner AM, Klutch M, and Quinnan GV Jr:
Reactivity of an HIV gag gene polypeptide expressed in E. coli withsera from AIDS patients and monoclonal antibodies to Gag.Biochim Biophys Acta 1988;949:213-223.
45. Mikhailov MV, Vasilyev AL, Nikolaeva IA, Arsenyan SG, andSverdlov ED: Antigenic determinants synthesized in a library ofrandomly cloned fragments of the HIV-1 genome. FEBS Lett1990;259:318-320.
46. Winther MD, Allen G, Bonford RH, and Brown F: Bacteriallyexpressed antigenic peptide from FMDV capsid elicits variableimmunologie responces in animals. J Immunol 1986;136:1835-1840.
47. Scholl B-C, von Hintzenstern J, Borisch B, Traupe B, et al.:Procaryotic expression of immunogenic polypeptides of the largephosphoprotein (pi50) of human cytomegalovirus. J. Gen Virol1988;69:1195-1204.
48. Martin PW, Burger DR, Caouette S, Goldstein AS, and Peetoom F:Importance of confirmatory test after strongly positive HTLV-IIIscreening test. N Engl J Med 1985;314:1577.
49. Lauritzen E and Lindhardt BO: Antibodies against HIV detected byimmunoblotting. In: Handbook ofImmunology ofProteins, Vol. II.Bjerrum O and Heegaard N (Eds.). CRC Press, Boca Raton, Florida,1988, pp. 117-129.
50. Calfruny WA, Heruth DP, Jaqua MJ, and Plagemann PGW:Immunoglobulins that bind to uncoated ELISA plate surfaces:Appearance in mice during infection with lactate-dehydrogenase-el-

664 ISAGULIANTS ET AL.
53. Jansson M, Wahren B, Scarlatti G, Principi N, Lombardi V,Livadiotti S, Elia L, Plebani A, Wigzell H, and Rossi P: Patterns ofIgG subclasses reactivity to HIV-1 envelope peptides in childrenborn to HIV-1 infected mothers. AIDS 1992;6:365-371.
evating virus and in human anti-nuclear antibody positive sera. JMed Virol 1986;19:175-186.
51. Geetligs HI, Weijer WJ, Bloemhoff W, Welling GW, and Wester S:The influence of pH and ionic strength on the coating of peptides ofherpes simplex virus type 1 in an enzyme-linked immunosorbent as-
say. J Immunol Methods 1988; 106:239-244.52. Lange JM, de Wolf F, Krone WJA, Danner SA, Coutinho RA, and
Goudsmit J : Decline of antibodies to outer core protein p 17 is an ear-
lier serological marker of disease progression in HIV infection thananti-p24 decline. AIDS 1987; 1:155-159.
Address reprint requests to:Britta Wahren
Department of VirologySwedish Institute for Infectious Disease Control
105 21 Stockholm, Sweden

This article has been cited by:
1. BERNHARD ECKER, SABINE VOLLENHOFER, TINA BARES, THOMAS SCHALKHAMMER, MANFREDSCHINKINGER, FRITZ PITTNER. 1996. Overexpression and Purification of a Recombinant Chimeric HIV Type 2/HIVType 1 Envelope Peptide and Application in an Accelerated Immunobased HIV Type 1/2 Antibody Detection System (AIBS): ANew Rapid Serological Screening Assay. AIDS Research and Human Retroviruses 12:12, 1081-1091. [Abstract] [Full Text PDF][Full Text PDF with Links]
2. IAN J. LAUDER, HSIANG JU LIN, EDWARD B. SIWAK, F. BLAINE HOLLINGER. 1996. Kernel Density Analysis ofVariable and Conserved Regions of the Envelope Proteins of Human Immunodeficiency Virus Type 1 and Associated Epitopes.AIDS Research and Human Retroviruses 12:2, 91-97. [Abstract] [Full Text PDF] [Full Text PDF with Links]














![[JP Morgan] All You Ever Wanted to Know About Corporate Hybrids but Were Afraid to Ask](https://static.fdocument.pub/doc/165x107/55205d904979590a3f8b467e/jp-morgan-all-you-ever-wanted-to-know-about-corporate-hybrids-but-were-afraid-to-ask.jpg)



