LA MEMBRANA CELLULARE: IPOTESI DI DANIELLI E …users.unimi.it/monticy/dida/03.00 Membrane...
61
LA MEMBRANA CELLULARE: IPOTESI DI DANIELLI E DAVSON (1935) poro polare molecola lipidica molecola proteica
Transcript of LA MEMBRANA CELLULARE: IPOTESI DI DANIELLI E …users.unimi.it/monticy/dida/03.00 Membrane...

LA MEMBRANA CELLULARE: IPOTESI DI DANIELLI E DAVSON (1935)
poro polare
molecola lipidicamolecola proteica

LA MEMBRANA CELLULARE: MODELLO A MOSAICO FLUIDO

I MOVIMENTI DEI FOSFOLIPIDI NEL DOPPIO STRATO

PROTEINE DI MEMBRANA E TRASPORTI
diffusione trasporto attivotrasporto facilitato
proteina canale proteine trasportatrici
energia

UOVO DI ARBACIA
concentrazioni di Na e K intra- ed extracellulari
[Na+], [K +]; unità arbitrarie
acqua di mare
[K +] i
[K +]e
[Na+] i
[Na+]e

GLOBULO ROSSO O ERITROCITA
EMOGLOBINA
Emoglobina: pigmento respiratorio
vaso sanguignoglobulo rosso

LA MEMBRANA CELLULARE COME BARRIERA
membrana
FASE 1composizione 1
FASE 2composizione 2
prodotti di rifiuto
materiale ossidabile

Liquidointerstiziale
Vasisanguigni
cellula
LE BARRIERE DA SUPERARE

LIQUIDO INTERSTIZIALE
•pO2, pCO2
•concentrazione sostanze nutritizie•concentrazione prodotti di rifiuto•pH•pressione osmotica•concentrazione degli anioni•concentrazione dei cationi•volume dell’ambiente interno
Variano solo entrolimiti ristrettissimicompatibili con lavita.
“OMEOSTASI”

OSMOSI
membranaH2O H2O + saccarosio
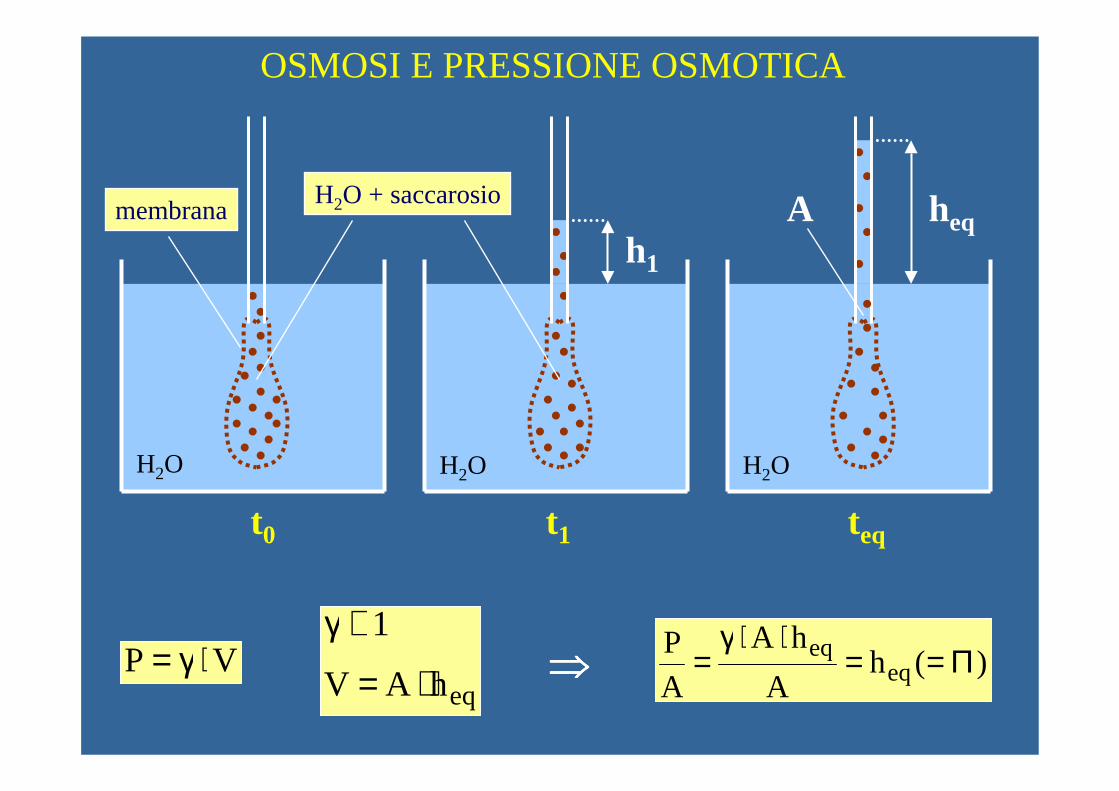
OSMOSI E PRESSIONE OSMOTICA
t0 t1 teq
h1
heq
H2O H2O H2O
H2O + saccarosio A
VP ⋅γ=eqhAV
1
⋅=
≅γ)(h
A
hA
AP
eqeq Π==
⋅⋅γ=⇒⇒⇒⇒
membrana
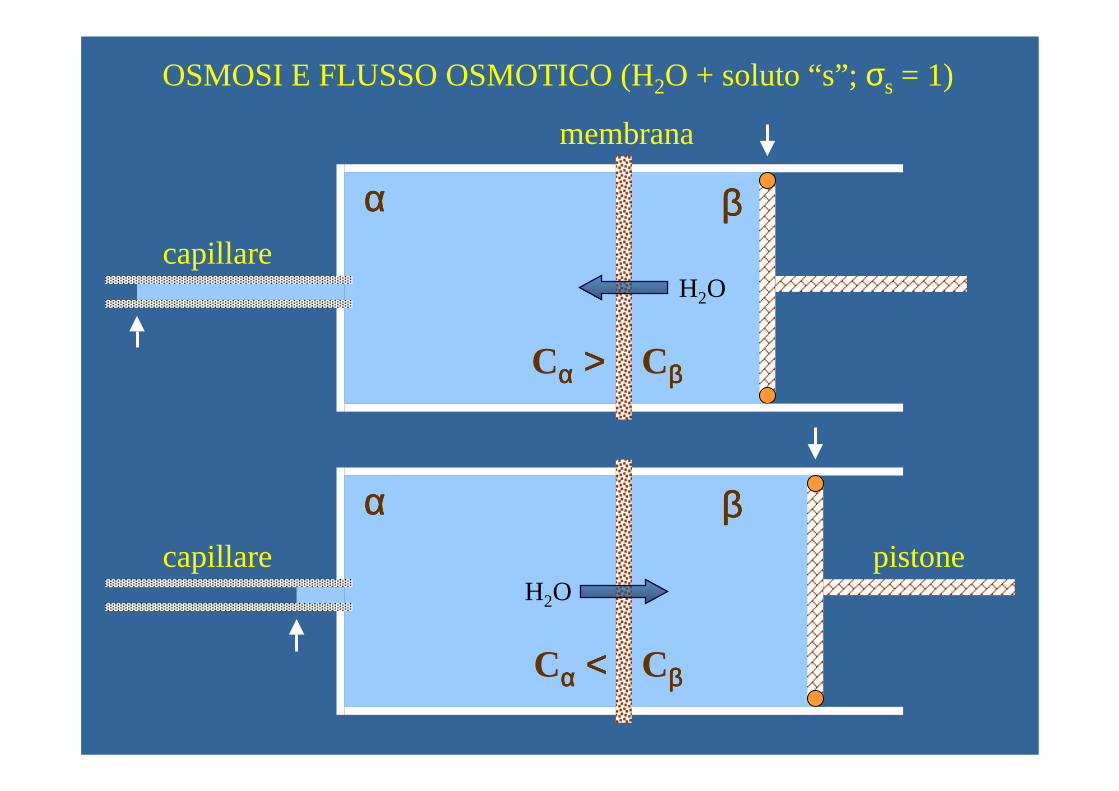
OSMOSI E FLUSSO OSMOTICO (H2O + soluto “s”; σs = 1)
membrana
capillare
capillare pistone
Cαααα >>>> Cββββ
Cαααα <<<< Cββββ
αααα ββββ
αααα ββββ
H2O
H2O

P V = n R T
nP = R T
Vn
c = V
= R T c
⋅ ⋅ ⋅
⋅ ⋅
Π ⋅ ⋅
LEGGE DI VAN’T HOFF
P: pressione,
V: volume,
n: numero di moli,
R: costante dei gas,
T: temperatura assoluta,
c: concentrazione,
Π: pressione osmotica.

OSMOLARITÀ
k = 1, 2……m
ck = c1, c2……cm
H2O
∑∑∑∑====
ΦΦΦΦ====m
kkkk cnOsmolarità
1
n = 1; saccarosio, glucosio etc.n = 2; NaCl, KCl, HCl etc.n = 3; MgCl2, Na2SO4 etc.
specie presenti
concentrazioni

a

OSMOLARITÀ DELLA SOLUZIONE FISIOLOGICA
H2O + NaCl
[NaCl] = 0,9% = 9 g/L
∑∑∑∑====
ΦΦΦΦ====m
kkkk cnOsmolarità
1
mOsm 45,28615493,02cnOsm NaClNaClNaClSF =⋅⋅=⋅Φ⋅=
mM 154]L[mol 154,0 ]mol[g 44,58
]L[g 9c
44,58PM
93,0
2n
1-1-
1-
NaCl
NaCl
NaCl
NaCl
=⋅=⋅⋅=
=
=Φ
=

METODI DI DETERMINAZIONE DELLA PRESSIONE OSMOTICA
1. metodo diretto,2. plasmolisi,3. abbassamento del punto di congelamento,4. tensione di vapore,5. composizione chimica.

ATTIVITÀ E COEFFICIENTI OSMOTICI
0,77930,77180,10
0,82200,81910,05
0,92830,92740,005
NaCl
f
KCl
fM⋅L-1
da R. Plonsey, D. G. Fleming “Bioelectric phenomena”
2
2i i
i
= R T a
a = f c
-log(f) = 0,509 z µ
µ = forza ionica della soluzione
1µ = c z
2
Π ⋅ ⋅⋅
⋅ ⋅
⋅ ⋅∑

DIFFUSIONE
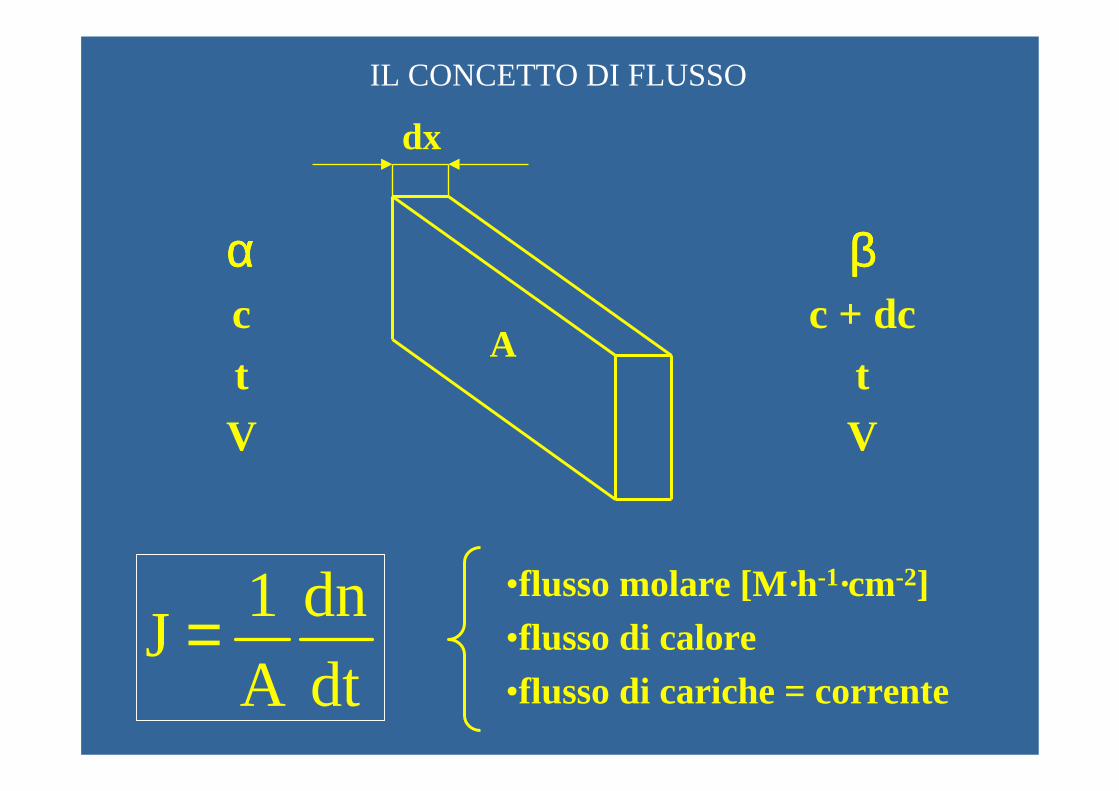
dx
A
ααααctV
ββββc + dc
tV
IL CONCETTO DI FLUSSO
1 dnJ
A dt=
•flusso molare [M·h-1·cm-2]•flusso di calore•flusso di cariche = corrente

dx
A
ααααc
ββββc + dc
DIFFUSIONE – I legge di Fick
1 dnJ P c - P (c dc) -P dc
A dt= = ⋅ ⋅ + = ⋅
1 dn-
dcJ P dc
xA dt-D
d= ⋅ ⋅==
I legge di Fick
se dx↑↑↑↑ ⇒⇒⇒⇒ interazioni ↑↑↑↑ ⇒⇒⇒⇒ P ↓↓↓↓
DP
dx=
[P] = cm⋅s-1 [D] = cm2⋅s-1
Adolf Eugen Fickfisiologo tedesco, 1829 – 1901

DIFFUSIONE ATTRAVERSO LA MEMBRANA CELLULARE
α βc = c c∆ −
membrana cellulare
fase lipidica
fase αacquosa
fase βacquosa
∆x
cα
cβ
?
concentrazione del soluto nella fase lipidicaB =
concentrazione del soluto nella fase acquosa
α, βc = concentrazioni soluto nella membrana alle superfici limite∗
⇒⇒⇒⇒α α
β β
c = B c
c = B c
∗
∗
⋅
⋅ α β α βc = c c = B (c c )∗ ∗ ∗∆ − ⋅ −→→→→

αc∗
membrana cellulare
fase lipidica
fase αacquosa
fase βacquosa
α β α β
c D A BJ = -D A = - (c c ) = -k A (c c )
x x
∗∆ ⋅ ⋅⋅ ⋅ ⋅ − ⋅ ⋅ −∆ ∆
∆x
cα
cββc∗
D Bk = (costante di permeabilità)
x
⋅∆
cα, cβmisurabili
MISURA DEL COEFFICIENTE DI PERMEABILITÀ

EFFETTI DEL COEFFICIENTE DI RIPARTIZIONE
r < 1
r·(cα - cβ) < (cα – cβ)
r1 > 1
r1·(cα - cβ) > (cα – cβ)

ce
ci
FLUSSI UNIDIREZIONALI
e i e i e i i eJ = -k A (c c ) = -k A c k A c = J J→ →⋅ ⋅ − ⋅ ⋅ + ⋅ ⋅ +
m
ci
ce
Je→iJi→e
m
ce
ci
ce
ci
m m
= +
0

RAPPORTO SUPERFICIE/VOLUME
ie i
ii
ie i
ie i
dn=k A (c -c )
dtn
=cV1 dn A
=k (c -c )V dt Vdc A
=k (c -c )dt V
⋅ ⋅
⋅ ⋅ ⋅
⋅ ⋅
ce
ci = ni/V
V
A
la variazione di concentrazione all’interno della cellula dipende dal
rapporto superficie/volume⇒⇒⇒⇒
r = 1 cm A/V = 3 cm-1
r = 50 µm A/V = 600 cm-1⇒⇒⇒⇒ equilibrio raggiunto 200 volte prima

Lo scoiattolo artico assume la forma sfericaper ridurre “S/V” e quindi minimizzare ilflusso (la perdita) di calore.
Hannah Carey – Wisconsin University: in The Scientist 17(20): 27, 2003
tcorporea = 36÷÷÷÷37 °°°°C →→→→ tamb + 1÷÷÷÷2 °°°°C
fcardiaca = 200÷÷÷÷300/min →→→→ 3÷÷÷÷5/min
letargo

PERMEABILITÀ AGLI ZUCCHERI
Molecola glucidica:
1. grande importanza metabolica,
2. poco adatta ad attraversare le membrane lipidiche per
a) struttura,
b) numerosi gruppi –OH.
Esperimento:
eritrociti umani
cglucosio, e↑↑↑↑ →→→→ Pglucosio↓↓↓↓ ⇒⇒⇒⇒ trasporto legato al grado di saturazione del trasportatore
cglucosio, e↑↑↑↑
cglucosio, i
Jglucosio
cglucosio, e
Jglucosio
⇒⇒⇒⇒
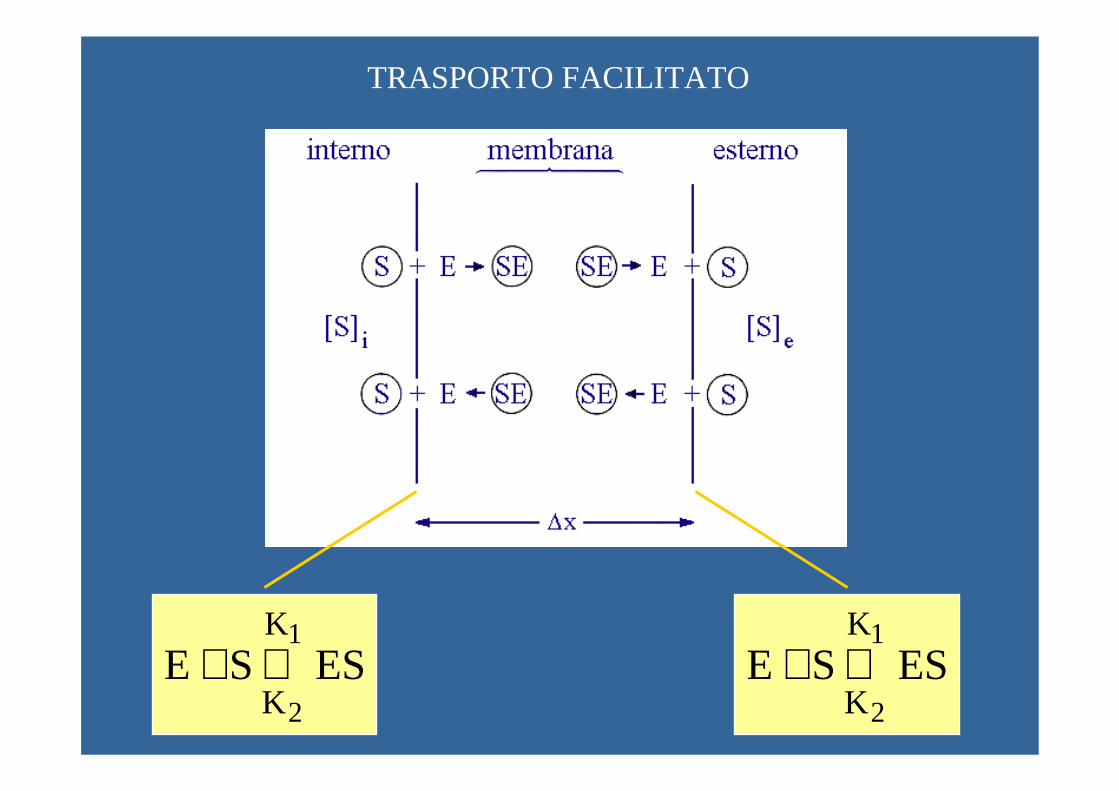
TRASPORTO FACILITATO
ES SE1
2
K
K⇔+ ES SE
1
2
K
K⇔+

ce
ci ↑↑↑↑
Je→i
Ji→e
Je→i
Ji→e
variabili nel tempo ⇒⇒⇒⇒
ce
ci
tempo
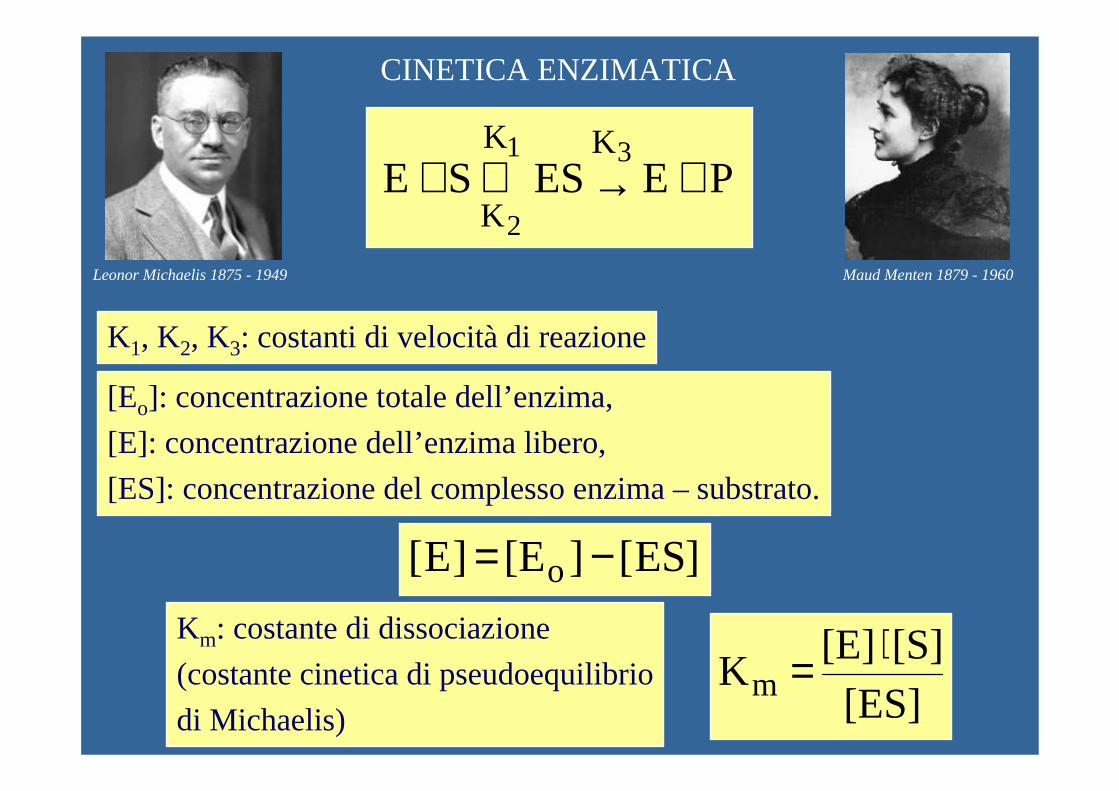
CINETICA ENZIMATICA
PE ES SE31
2
KK
K+→⇔+
]ES[][E ]E[ o −=
K1, K2, K3: costanti di velocità di reazione
[Eo]: concentrazione totale dell’enzima,
[E]: concentrazione dell’enzima libero,
[ES]: concentrazione del complesso enzima – substrato.
[ES][S][E]
Km⋅=
Km: costante di dissociazione
(costante cinetica di pseudoequilibrio
di Michaelis)
Leonor Michaelis 1875 - 1949 Maud Menten 1879 - 1960
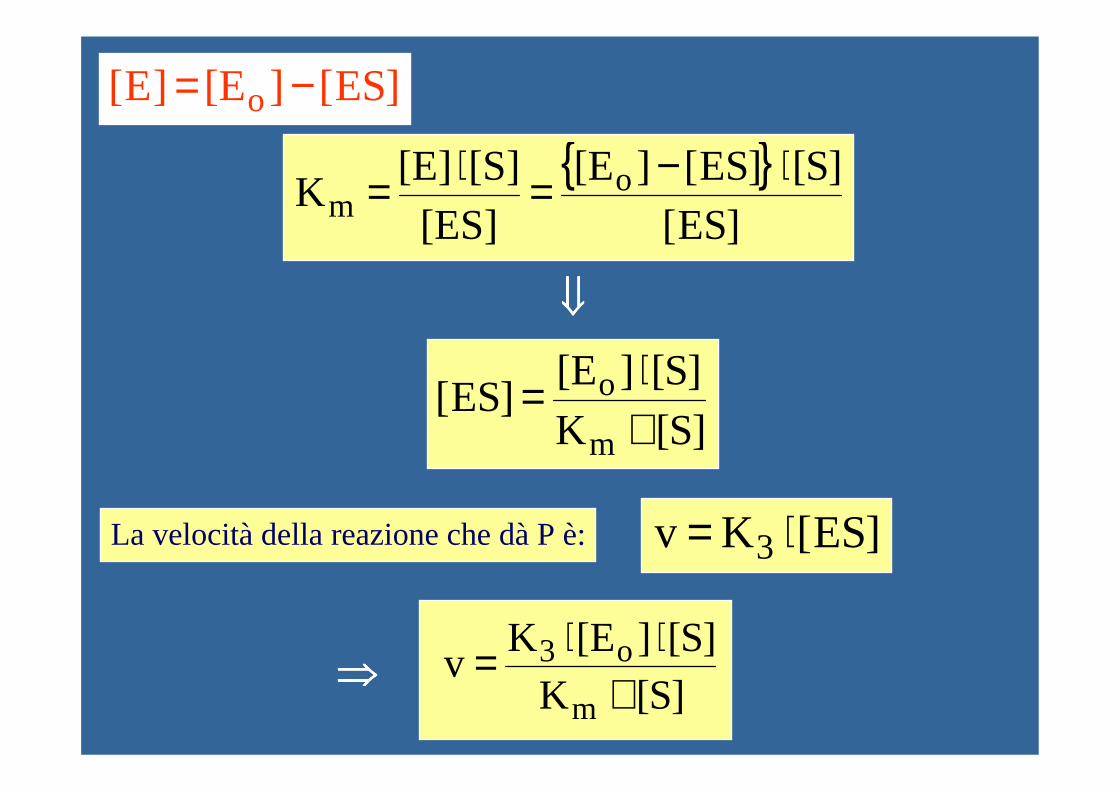
{ }]ES[
]S[]ES[][E
[ES][S][E]
K om
⋅−=⋅=
]S[K]S[][E
]ES[m
o
+⋅=
]ES[K v 3 ⋅=
]ES[][E ]E[ o −=
⇓⇓⇓⇓
La velocità della reazione che dà P è:
]S[K]S[][EK
vm
o3
+⋅⋅=⇒⇒⇒⇒

][EK V o3max ⋅=
]S[K]S[V
vm
max
+⋅=
Quando l’enzima è saturo: [ES] = [Eo]
]ES[K v 3 ⋅= ⇒⇒⇒⇒
]S[K]S[][EK
vm
o3
+⋅⋅= ⇒⇒⇒⇒
equazione di Michaelis-Menten
la velocità di una reazione enzimatica dipende solo da [S]
⇓⇓⇓⇓
v = f([S])
Vmax determinata sperimentalmente: ([S], v) ⇒ (calcolo) Km
Maud Menten 1879 - 1960

[S]
v
Vmax
]S[K]S[V
vm
max
+⋅=
maxmax
m
V1
]S[1
VK
v1 +⋅=
linearizzazione
⇓⇓⇓⇓
EQUAZIONE DI MICHAELIS-MENTEN
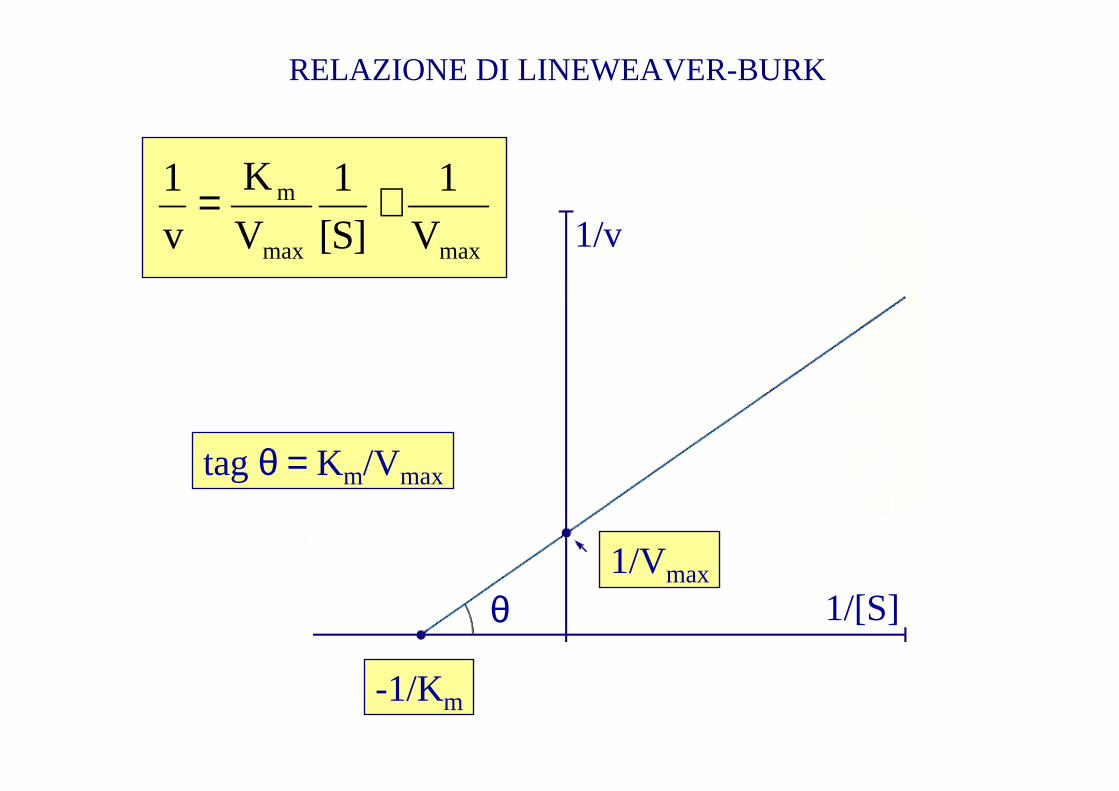
maxmax
m
V
1
]S[
1
V
K
v
1 +=
-1/Km
1/[S]
1/v
θ
tagθ = Km/Vmax
1/Vmax
RELAZIONE DI LINEWEAVER-BURK
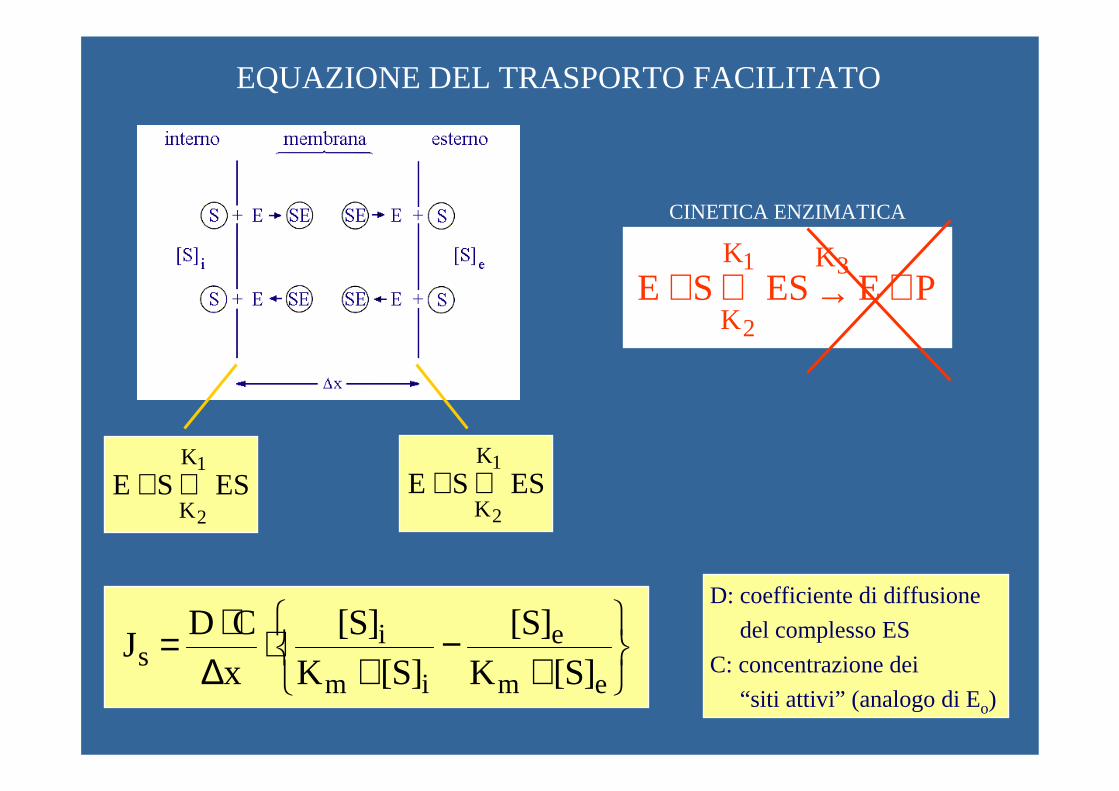
EQUAZIONE DEL TRASPORTO FACILITATO
PE ES SE31
2
KK
K+→⇔+
ES SE1
2
K
K⇔+ ES SE
1
2
K
K⇔+
CINETICA ENZIMATICA
+−
+⋅
∆⋅=
em
e
im
is ]S[K
]S[]S[K
]S[xCD
J
D: coefficiente di diffusione
del complesso ES
C: concentrazione dei
“siti attivi” (analogo di Eo)

x[ES]
D Js ∆∆⋅=
im
ie)(i s ]S[K
]S[Cx
D J
+⋅⋅
∆=→
em
ei)(e s ]S[K
]S[Cx
D- J
+⋅⋅
∆=→
{ }{ } { }emim
eimi)(e se)(i ss ]S[K]S[K
]S[]S[KxCD
JJ J+⋅+
−⋅∆⋅=+= →→
A B
BA [ES][ES] [ES] −=∆
im
ioA ]S[K
]S[][E ]ES[
+⋅=
em
eoB ]S[K
]S[][E ]ES[
+⋅=CINETICA ENZIMATICA
⇓⇓⇓⇓

+−
+⋅=
em
e
im
imaxs ]S[K
]S[]S[K
]S[J J
+−
+⋅
∆⋅=
em
e
im
is ]S[K
]S[]S[K
]S[xCD
J]ES[K v 3 ⋅=
Jmax
quando [ES] = [Eo] (enzima saturo)
v = Vmax
Jsv
P = D/∆xK3
TRASPORTO FACILITATOCINETICA ENZIMATICA
xCD
Jmax ∆⋅=

im
imaxs ]S[K
]S[J J
+⋅=
V = ∞t = 0, [S]e = 0
t ↑, [S]e = 0
Js
t = 0, [S]it ↑, [S]i ↓
⇓⇓⇓⇓
+−
+⋅=
em
e
im
imaxs ]S[K
]S[]S[K
]S[J J
]S[K]S[V
vm
max
+⋅=
eq. di Michaelis-Menten
equilibrio (Js = 0) solo quando [S]i = 0
⇓⇓⇓⇓
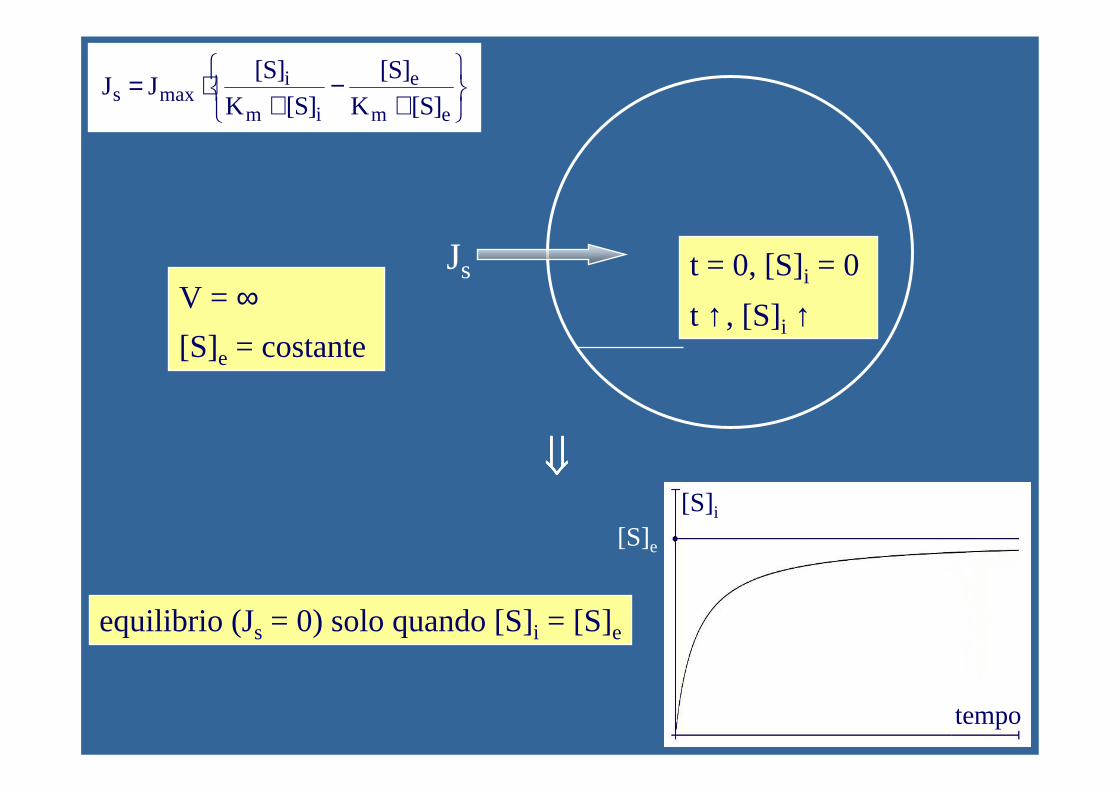
V = ∞[S]e = costante
Js t = 0, [S]i = 0
t ↑, [S]i ↑
⇓⇓⇓⇓
+−
+⋅=
em
e
im
imaxs ]S[K
]S[]S[K
]S[J J
equilibrio (Js = 0) solo quando [S]i = [S]e
tempo
[S]i[S]e

INIBITORI
competitivi non competitivi florentina (aglicone di florizina)
dinitrofluorobenzene
florizina composti a base di Hg vari narcotici
competitivi
⇓⇓⇓⇓Js ↓ o Js = 0
non competitivi
⇓⇓⇓⇓Js varia per interazioni
competizione fra molecole (S e inibitore) per il trasportatore
⇓
numero di siti attivi per S ↓ → Km varia

AUMENTO DI Js
l’inibitore può favorire il distacco
ES → E + S⇓⇓⇓⇓
Js ↑
• eritrociti umani: aumenta la fuoruscita di glucosio in presenza di mannosio
n. trasportatori ↑ (?)
• insulina → Jglucosio↑
affinità glucosio-trasportatore ↑ (?)

TRASPORTO FACILITATO - CONCLUSIONI
soluto trasportato barriera sede del trasporto K membrane lipidiche naturali membrane lipidiche artificiali glucosio membrana placentare barriera ematoencefalica membrana del globulo rosso Fe, vitamina B12 epitelio intestinale membrana del globulo rosso Ca orletto a spazzola delle cellule
• formazione e scissione del legame fra S ed E determinate da particolari caratteristiche:
chimiche, stereochimiche, fisiche;
• specifico (ma vi sono casi di trasportatori per più soluti);
effettiva traslocazione del complesso (se liposolubile); es. valinomicina + K+
• natura
trasportatore fisso → formazione di canale selettivo

αααα ββββ
STATO DI EQUILIBRIO (esempio idraulico)
∆∆∆∆h = hαααα - hββββ
∆∆∆∆h = 0

αααα ββββ
STATO STAZIONARIO (esempio idraulico)
∆∆∆∆h = hαααα - hββββ = cost
P

∆∆∆∆P = Pαααα – Pββββ
faseαααα faseββββ
'OH2
J
m

faseαααα faseββββ
''OH2
J
cαααα > cββββ
∆∆∆∆P = 0
m

∆∆∆∆P = Pαααα – Pββββ
faseαααα faseββββ
cαααα > cββββm
'OH 2
J
''OH 2
J
''OH
'OHOH 222
J-JJ =
filtrazione
osmosi

faseαααα faseββββ
''OH2
J
cαααα > cββββ
∆∆∆∆P = 0
m
sJ

faseαααα faseββββ
'OH2
J
cαααα (A+B-) = cββββm
+AJ
-BJ
∆∆∆∆P = Pαααα – Pββββ

faseαααα faseββββ
''OH2
J
cαααα (A+B-) > cββββ
∆∆∆∆P = 0
m
+AJ
-BJ

faseαααα faseββββ
cββββ
∆∆∆∆P = 0
m
+AJ
-BJ
cαααα (A+B-) =
++++ −−−−∆∆∆∆V

faseαααα faseββββ
cβ1β1β1β1
cβ2β2β2β2
….cββββn
m
i+AJ
i-BJ
cα1α1α1α1 ≠cα2α2α2α2 ≠….
cααααn ≠
++++ −−−−∆∆∆∆V
∆∆∆∆P

∆∆∆∆P
αααα ββββ
cαααα1……cααααm
M
cββββ1……cββββm
∆∆∆∆E
IL POTENZIALE ELETTROCHIMICO
j i
ii T,p,n
Gµ
n≠
∂= ∂ % ( ) ( )0
i i i i iµ µ T VP RTln c z FE= + + +% %

( )i αi βi i i i∆µ µ -µ V∆P RT∆ln c z F∆E= = + +% % %
LA DIFFERENZA DI POTENZIALE ELETTROCHIMICO

αi αii
i βi i βi
RT c A cE - ln ln
z F c z c= =
( )i i i∆µ RT∆ln c z F∆E= +%
IL POTENZIALE DI EQUILIBRIO

LA LEGGE DI NERNST
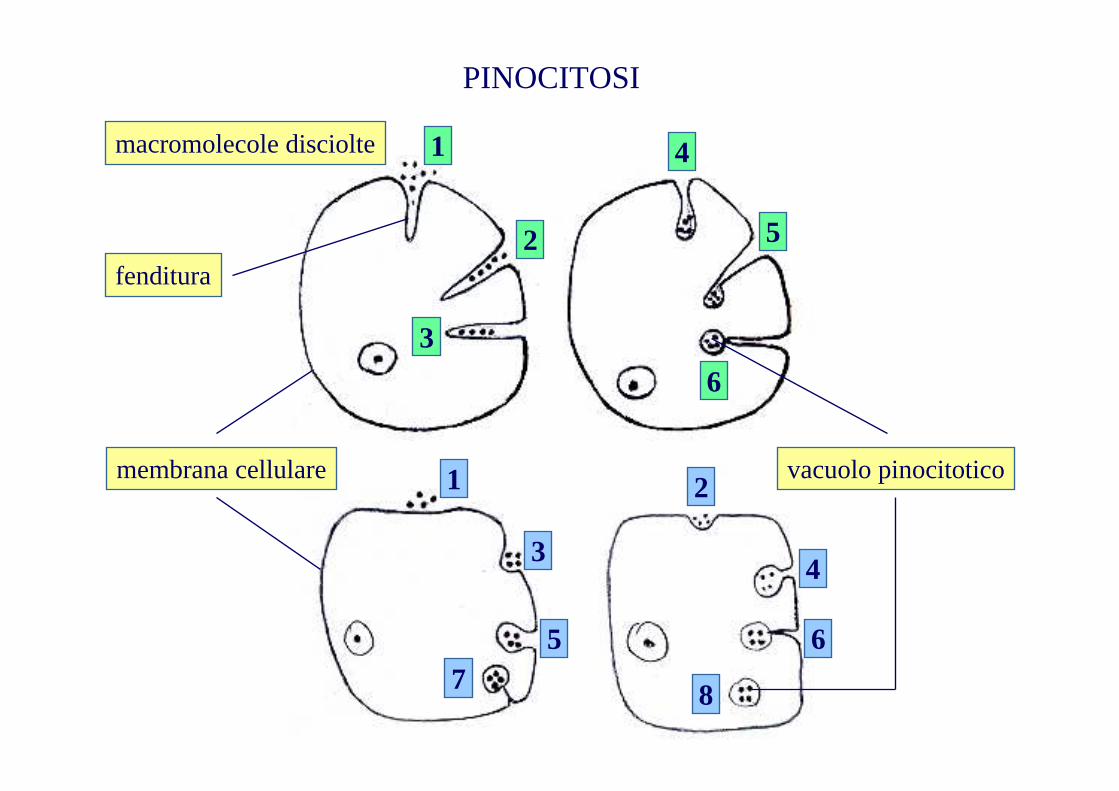
PINOCITOSI
vacuolo pinocitoticomembrana cellulare
fenditura
macromolecole disciolte 1
2
3
4
5
6
1 2
34
5 67 8

FAGOCITOSI
particella solida o batterio
nucleo membrana cellulare
1 23
4
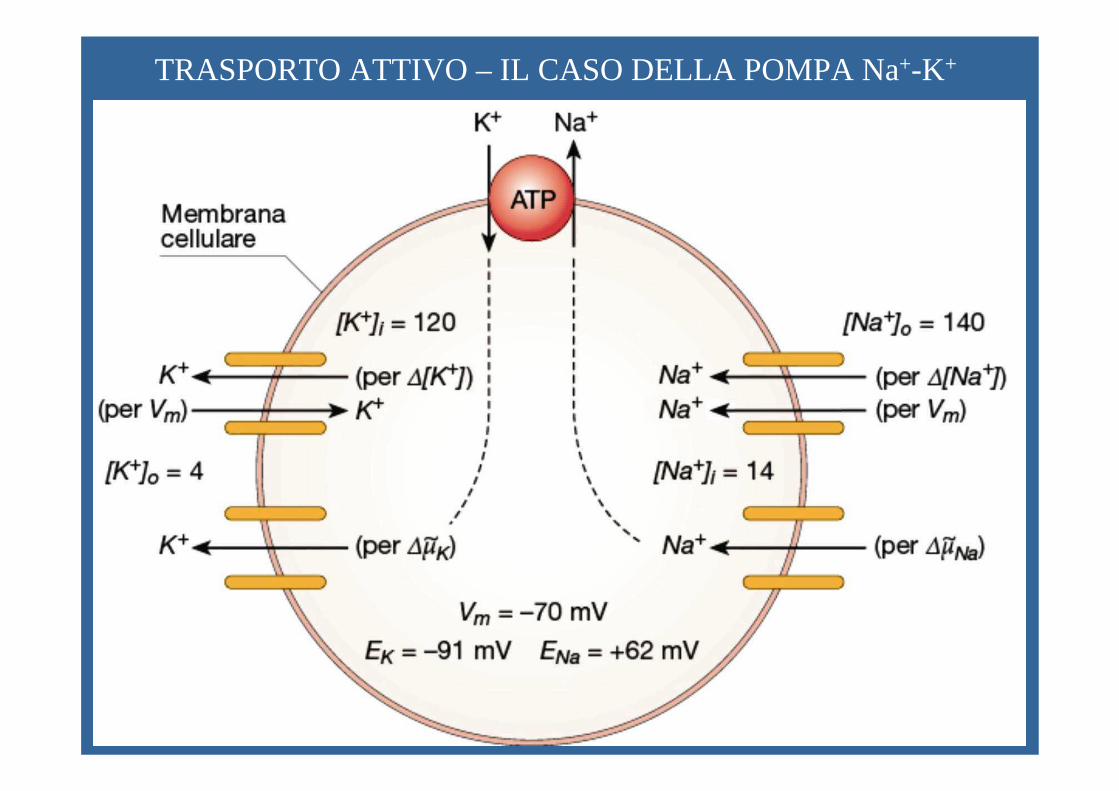
TRASPORTO ATTIVO – IL CASO DELLA POMPA Na+-K+

“Things should be made as simple as
possiblebut not any simpler.”
…Albert Einstein (1879 – 1955)