
La chromatine - tagc.univ-mrs.frtagc.univ-mrs.fr/puthier/images/download/3_chromatine.pdf · La...
27
La chromatine
Transcript of La chromatine - tagc.univ-mrs.frtagc.univ-mrs.fr/puthier/images/download/3_chromatine.pdf · La...

La chromatine

La chromatine
Dans le noyau des cellules eucaryotes l'ADN génomique est fortement compacté.
La chromatine correspond à l'association d'ADN et de protéines structurales
Hétérochromatine structure dont l'état de condensation n'est pas altéré au cours du cycle cellulaire.
Euchromatine structure décondensée à l'interphase (G1/S/G2).
L'hétérochromatine est localisée principalement en région péri-nucléaire et nucléolaire
L'hétérochromatine constitutive
L'hétérochromatine facultative
L'euchromatine est plutôt répartie à l'intérieur du nucléoplasme

L'hétérochromatine constitutive:
Hétérochromatine constitutive
Correspond à des régions quasi dénuées de gènes
Formée principalement des télomères et centromères et certaines régions de certains chromosomes (1,9,13,14,15,Y)
Régions riches en séquences répétées (séquences « satellites »)
(ex: séquences « α-satellites » au niveau des centrosomes)
Très polymorphe
Elle est stable et donc retrouvée à toutes les étapes de développement ou de la différenciation chez tous les individus
Coloré par les bandes C (« C banding », sulfate de baryium)

Hétérochromatine constitutive
‘rDNA’ is a region containing a cluster of repeat units for the ribosomal RNA genes, which specify a type of non-coding RNA

L'hétérochromatine facultative
Correspond à des régions transcriptionnellement inactives (mais contenant des gènes)
Son état est réversible
Son état « inactif » est fonction du développement, de l'état de différenciation, du stimuli,...
Elle n'est pas particulièrement enrichie en séquences satellite
Cas particulier du corpuscule de Barr
Inactivation de l'X (ou lyonisation) se déroule dans les stades précoces de l'embryogenèse (blastocyste).
Toute la descendance d'une cellule porte le même chromosome X inactivé
Mosaïcisme

Hétérochromatine
L'hétérochromatine
Est répliquée tardivement (lié à la forte compaction)
Est méthylée sur les cytosines (5-Méthylcytosine)
Hétérochromatine constitutive
Très méthylée
Hétérochromatine facultative
Méthylée au niveau des ilots CpG (un C suivi d'un G, p correspond à phosphate)
CpG retrouvés dans les régions régulatrices des gènes
→

Organisation de la chromatine

Figure 4: Chromatin fibers purified from chicken erythrocytes. Each nucleosome (~12-15 nm) is well resolved, along with the linker DNA between the nucleosomes. Given the resolution, other components, if present, such as a transcribing RNA polymerase or transcription factor complexes, should be resolvable
Probing Nanometer Structures with Atomic Force Microscopy. Zhifeng Shao. News Physiol Sci 14, 1999;

Une structure dynamique
Chromatine: des contraintes antinomiques
Nécessité de compacter l'ADN (~ 2 mètres !)
Intervention de protéines spécialisées
Nécessité d'être « modelable » car l'ADN doit être accessible rapidement
Pour induire la réplication
Pour induire la transcription
Pour la réparation
Nécessite la mise en place d'une machinerie spécifique

Histones et nucléosomes
Les histones sont des petites protéines (11-22 kDa) basiques (riches en arginine et lysine) très conservées au cours de l'évolution . On en a décrit 5 à ce jour.
les histones H2A, H2B, H3 et H4 peuvent s'associer pour former un octamère (nucléosome)
(H2A,H2B,H3,H4) x 2
Premier niveau de compaction de l'ADN
~ 11nm
Un octamère est entouré d'une boucle d'ADN
de 146pb
L'histone H1 permet la compaction des
nucléosomes
Rigidifie la structure hélicoïdale (30 nm)
ainsi obtenue
2ème degré de condensation de l'ADN

Les histones
Les histones contiennent:
Une région centrale (appelée « histone-fold ») contenant trois hélices α séparées par 2 boucles
Permet des interactions entre histones
Des extrémités amino- et carboxy-terminales.
Les extrémités amino-terminales (dans une moindre mesure carboxyterminales) sont les cibles de nombreuses modifications post-traductionnelles.
N-ter C-ter

Modifications post-traductionnelles des histones
Les modifications les mieux décrites sont:
L’acétylation → ajout d'un groupement acétyle
Fonction carbonyle, fonction méthyle
La méthylation → ajout d'un groupement méthyle
La phosphorylation → ajout d'un phosphate
L’ubiquitinylation → ajout d'ubiquitine
Autres modifications:
telles que la sumoylation, la glycosylation, la biotinylation, l'ADP-ribosylation,...

La réaction d'acétylation
L'acétylation se produit sur un groupement aminé
Soit à l'extrémité N-terminale d'une protéine
Soit A l'extrémité de la chaîne latérale d'une lysine.

Modifications alternatives

Résidus sujets à modification

Nomenclature des modifications

Modificateurs
"Variations sur le thème du code histone". Dominique Ray-Gallet, Annabelle Gérard, Sophie Polo, Geneviève Almouzni. MEDECINE/SCIENCES 2005 ; 21 : 384-9

Enzymes intervenant dans la méthylation

Code génétique des histones
L'acétylation de plusieurs lysines des histones H3 et H4 est impliquée dans la régulation de l'expression génique
Les fonctions amine des lysines sont chargées positivement à pH neutre
Permet l'interaction avec l'ADN
La neutralisation de ces fonctions par les HAT (acétylation) déstabilise la liaison à l'ADN)
ADN plus facilement accessible

Code génétique des histones
La distribution des modifications d'histone permet de réguler la dynamique chromatinienne
Hétérochromatine
H3K9me2, H3K9me3
Régions transcriptionnellement actives et TSS
H3K4me3, H3K4me2, H3K36me, H3K4me1, H3ac, H4ac
Chromosome X inactif
H3K27me3, H3K9me2, H4K20me, H2AK119ub1
Chromosome condensés durant la mitose et la méiose
Phosphorylation de H3 (sérine et Thréonine)
H3S10ph, H3S28ph, H3Thr11ph

Modifications post-traductionelles
Chromatin disruption and modification. AP Wolffe and JJ Hayes. Nucleic Acids Research, Vol 27, Issue 3 711-720
Les parties amino-terminales des histones H2B, H3 et H4 et la partie carboxy-terminale de H2A sont présentées à l'extérieur du nucléosome.
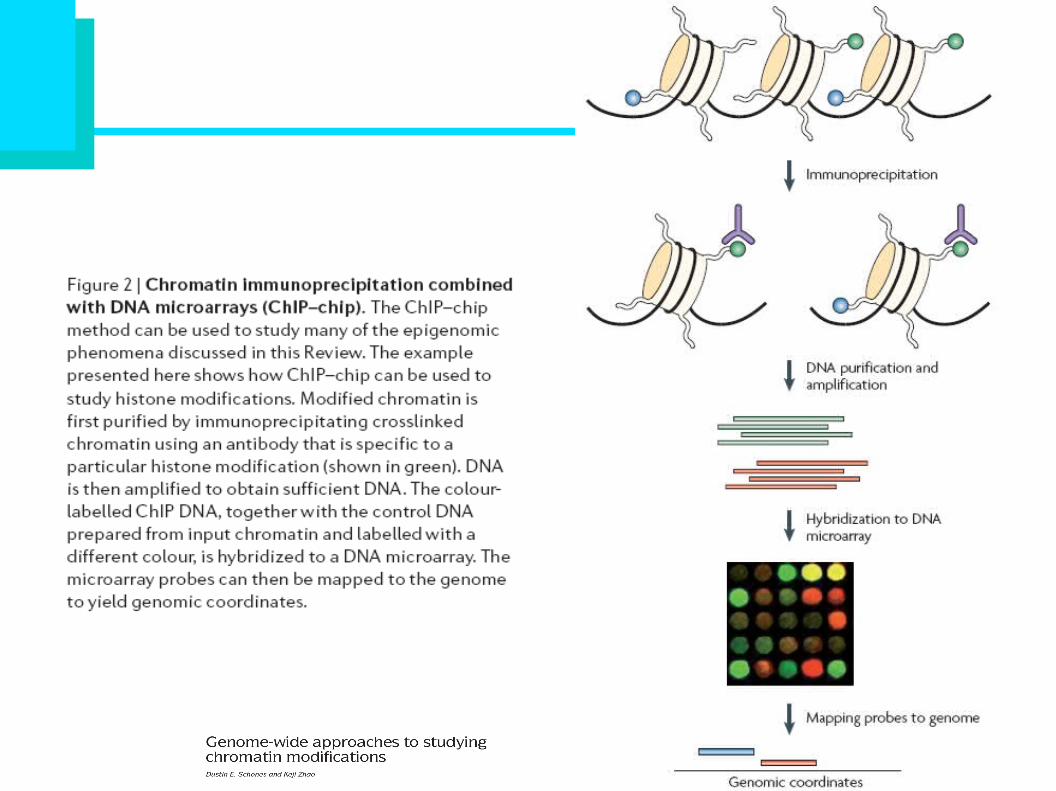

Dynamic Remodeling of individual nucleosomes

sphéroplaste: Cellules, provenant de bactéries et de levures qui ont partiellement perdu leur paroi donc leur forme caractéristique, et sont devenues rondes.
Micrococcal Nuclease (S7 Nuclease) is a relatively nonspecific nuclease that digests single-stranded and double-stranded nucleic acids.

Dynamic Remodeling of individual nucleosomes

(A) Average nucleosome profiles of all genes in the yeast genome from −600 bp to +1,000 bp with respect to the transcription start site (TSS). Nucleosome positions are shown as gray ovals below the profile. The intensity of the filled oval reflects the average probability score of the nucleosomes.
Dynamic Remodeling of individual nucleosomes

(E) The genes in the yeast genome were sorted in descending order according to their transcription rates [16], and the average promoter nucleosome profiles for the top 500 genes (orange) and the bottom 500 genes (black) are plotted.
Dynamic Remodeling of individual nucleosomes


















