El cloroplasto: un organelo clave en la vida y en el aprovechamiento ...

B3
Bio
logi
a
B2
LE
ZIO
NE
30 μm140 μm
20 μm 20 μm
01
01 La cellula: sguardo d’insieme
LE DIMENSIONI DELLE CELLULE
La cellula è l’unità base dei viventi.
Esistono due tipi di cellule: le cellule procariotiche e le cellule eucariotiche. La cellula procarioti-
ca è più piccola e ha una struttura più semplice rispetto a quella eucariotica, tuttavia le funzioni
fondamentali di una singola cellula procariotica di un batterio sono simili a quelli che governa-
no i circa 60 000 miliardi di cellule eucariotiche del corpo umano.
L’osservazione della struttura cellulare richiede strumenti opportuni perché le cellule, in genere,
sono molto piccole.
0,1 nm 1 nm
Atomi
Piccole molecole
Lipidi
Proteina
Fago T2 (virus)
Cloroplasto(organello)
La maggior parte dei batteri
Gran parte delle cellulevegetali e animali
Uovo di pesce
Colibrì
Uomo
Balenottera azzurra
Sequoia della California
10 nm 100 nm 1 μm 10 μm 100 μm 1 mm 1 cm 0,1 m 1 m 10 m 100 m 1 km
Microscopio elettronico
Microscopio ottico
Occhio nudo
IL MICROSCOPIO
Il più piccolo oggetto che normalmente un essere umano riesce a distinguere misura circa
0,2 mm (200 μm) . I microscopi accrescono il potere di risoluzione in modo tale che riusciamo a
osservare le cellule e le loro strutture interne.
I microscopi sono fondamental-
mente di due tipi:
risoluzione di circa 0,2 μm);
Vediamo ora alcuni esempi di come
appaiono le cellule osservate con le
tecniche di microscopia ottica.
Nel microscopio ottico
si distinguono due viti:
quella macrometrica,
che serve per allontanare
o avvicinare il tubo
portalenti al vetrino, per la
messa a fuoco, e quella
micrometrica, consente
movimenti più fini per
regolare la messa a fuoco.
La distanza che deve
separare due oggetti
perché l’occhio li
percepisca come distinti
prende il nome di
risoluzione; due oggetti
più vicini sembrano
un’unica macchia.
Le strutture interne delle
cellule sono difficili da
distinguere sotto la luce
visibile, perciò spesso
le cellule subiscono un
trattamento chimico che
ne colora i componenti
con tinte diverse per far
risaltare certe strutture.
micron cubi: un micron è pari a un millesimo di millimetro, cioè 1 μm = 10–3 mm = 10–6 m
Le cellule sono minuscole: il volume di una cellula varia da 1 μm3 a 1000 μm3 . Non
mancano le eccezioni: le uova degli uccelli sono enormi, e alcuni tipi di alghe unicel-
lulari possono essere visti a occhio nudo.
Le dimensioni ridotte dipendono dalla necessità di mantenere un adeguato rappor-to tra superficie e volume. Il volume infatti determina la quantità di attività chimica
svolta dalla cellula nell’unità di tempo, mentre la superficie determina la quantità di
sostanze che la cellula può scambiare con l’esterno (cioè i nutrienti che vengono pre-
levati dall’ambiente e i prodotti di scarto che sono riversati all’esterno).
In una scala logaritmica, ogni unità è 10 volte più grande della precedente.
Il diametro della maggior parte delle cellule è compreso nell'intervallo 1-100 μm
Questa scala logaritmica mostra le dimensioni relative delle
molecole, delle cellule e degli organismi pluricellulari.
Molte cellule sono più piccole di 200 μm e quindi invisibili per l’occhio umano.
Oculare
Lenti dell’oculare
Lenti dell’obiettivo
Lenti del condensatore
Fonte di luce
Nella microscopia in campo chiaro, la luce
passa direttamente attraverso queste cellule
umane. A meno che non siano presenti pig-
menti naturali, vi è poco contrasto e i detta-
gli non vengono distinti.
Nella microscopia a fluorescenza, una so-
stanza naturale all’interno delle cellule o un
colorante fluorescente che si lega a uno spe-
cifico componente cellulare viene eccitato da
un raggio di luce e viene osservata la luce fluo-
rescente emessa a una lunghezza d’onda supe-
riore proveniente direttamente dal colorante.
Nella microscopia a contrasto di fase, il
contrasto dell’immagine viene aumentato
enfatizzando le differenze di indice di rifra-
zione (la capacità di deviare la luce), esaltan-
do così le regioni chiare e scure della cellula.
La microscopia confocale si basa sull’emis-
sione in fluorescenza ma sfrutta un sistema
di focalizzazione sia della luce eccitante sia di
quella emessa. Il risultato è un’immagine bi-
dimensionale più nitida di quella che si può
ottenere con la microscopia a fluorescenza
standard.
Per potenziare
l’osservazione delle
cellule al microscopio
ottico o elettronico sono
state sviluppate numerose
tecniche.

B5
Bio
logi
a
B4
LE
ZIO
NE
200 nm
02
02 Le cellule procariotiche
I biologi classificano tutti gli esseri viventi in tre domini: gli archei, i batteri e gli eucarioti.
Gli archei e i batteri sono costituiti da cellule procariotiche (di dimensioni comprese tra 0,25
e 4 μm), che non possiedono compartimenti interni delimitati da membrane. Probabilmente
le prime cellule avevano un’organizzazione simile a quella dei moderni procarioti, che ancora
oggi costituiscono la maggior parte degli organismi viventi. Le cellule che costituiscono que-
sti organismi condividono alcune caratteristiche comuni: sono circondate da una membrana,
contengono citoplasma, almeno una molecola di DNA e un certo numero di ribosomi, organuli
deputati alla sintesi delle proteine.
La molecola di DNA
contenuta nel nucleoide
è costituita da un numero
di coppie di basi (bp) che
spazia da 3 a 5 000 000.
Le cellule procariotiche so-
no delimitate da una mem-brana plasmatica semiper-
meabile, composta da un
doppio strato fosfolipidico.
All’interno della membrana plasmatica vi è il citoplasma, materiale semifluido entro cui av-
vengono le reazioni cellulari. Il citoplasma è composto da due parti: il citosol, che è la parte più
fluida, e le particelle insolubili sospese in esso.
Una particolare zona del citoplasma chiamata nucleoide, contiene il DNA, che si trova sotto
forma di un’unica molecola circolare, alla quale sono associate proteine, anche se non in modo
altamente organizzato come negli eucarioti.
Nel citoplasma sono sempre presenti i ribosomi liberi del diametro di 25 nm, in cui ha luogo
la sintesi proteica. I ribosomi sono costituiti da RNA e proteine.
Molti procarioti possiedono una parete cellulare, costituita
da peptidoglicani, situata esternamente alla membrana pla-
smatica. Essa dà sostegno alla cellula e ne determina la for-
ma. A seconda del tipo di parete cellulare (più o meno spes-
sa), i batteri si dividono in Gram positivi e Gram negativi.
Attorno alla parete cellulare, alcuni batteri presentano
uno strato di muco composto per lo più da polisaccaridi,
denominato capsula. La capsula può avere diverse funzio-
ni: proteggere i batteri procarioti che infettano gli animali
dall’attacco del sistema immunitario, favorire l’adesione ad
altre cellule o a un substrato; prevenire l’essiccamento della
cellula.
Nel corso dell’evoluzione, alcuni procarioti hanno sviluppato strutture specializzate. Nei
cianobatteri, per esempio, sono presenti speciali membrane, capaci di svolgere la fotosintesi.
Numerosi procarioti, inoltre, si muovono grazie ad appendici denominate flagelli, costi-
tuite da proteine in grado di contrarsi. Il flagello ha l’aspetto di un cavatappi e ruota sul
proprio asse come un’elica. Altri procarioti invece hanno strutture dette pili, che hanno una
struttura simile ai flagelli ma sono molto numerosi e più piccoli e permettono alla cellula
procariotica di aderire ad altri organismi, per lo scambio di materiale genetico o per ricava-
re protezione e cibo. I pili sessuali intervengono nella coniugazione batterica.
Capsula
Batterio Pseudomonas aeruginora
Ribosomi
Nucleoidi
Membrana plasmatica
Flagello
Membrana plasmaticaPeptidoglicanoMembrana esterna(assente in alcuni batteri)
Parete cellulare

B7
Bio
logi
a
B6
LE
ZIO
NE
1,5 μm
0,8 μm 25 nm
0,5 μm
30 nm0,1 μm
03
03 Le cellule eucariotiche
Le cellule eucariotiche possono essere animali o vegetali. Alcuni organuli sono presenti in entrambi i
tipi di cellula, mentre altri sono tipici dell’una o dell’altra. Nelle fotografie ottenute al microscopio elet-
tronico, molti organuli delle cellule vegetali appaiono quasi identici per forma a quelli osservati nelle
cellule animali. Le strutture cellulari specifiche delle cellule vegetali comprendono la parete cellulare e
i cloroplasti. Le cellule animali contengono invece i centrioli, che non si trovano nelle cellule vegetali.
CELLULA ANIMALE
I ribosomi sono i siti dove vengono sintetizzate le proteine.
Sono costituiti da RNA ribosomiale (rRNA) e oltre 50 pro-
teine. Si possono trovare liberi nel citoplasma o attaccati al
reticolo endoplasmatico.
La parete delle cellule vegetali è una struttura semirigida che si trova
all’esterno della membrana plasmatica. Ha tre importanti funzioni:
fornisce sostegno alla cellula; fa da barriera contro le infezioni fungi-
ne; contribuisce a dare forma alla pianta. Al microscopio, possiamo
riconoscere nella parete cellulare delle cellule vegetali i plasmodesmi,
piccoli canali che permettono a due cellule vegetali vicine di scam-
biarsi varie sostanze (proteine, ioni, acqua, macromolecole e RNA).I perossisomi sono piccoli organuli (dal
diametro di 0,2-1,7 mm) con un interno
granulare che contiene speciali enzimi;
grazie ai quali si possono demolire con
reazioni di ossidazione i perossidi tossici
(come l’acqua ossigenata o perossido di
idrogeno H2O2).
I mitocondri sono
le centrali energe-
tiche della cellula.
Un citoscheletro composto
di microtubuli, filamenti
intermedi e microfilamenti
sostiene la cellula ed è coin-
volto nel movimento della
cellula e dei suoi organuli.
Il nucleo è il luogo dove si
trova la maggior parte del
DNA cellulare che, con le
proteine a esso associate,
costituisce la cromatina.
I centrioli sono
coinvolti nella di-
visione nucleare.
La membrana plasmatica
separa la cellula dall’ambien-
te e regola il flusso di mate-
riali dentro e fuori di essa.
Il reticolo endoplasma-tico ruvido è il sito dove
si svolge gran parte della
sintesi proteica.
MitocondrioCitoscheletro
Nucleolo
NucleoReticolo endoplasmatico ruvido
Ribosomi
Reticolo endoplasmatico liscio
Membrana plasmastica
Esterno della cellula
Interno della cellula
Apparato di Golgi
Ribosomi (legati al RE ruvido)
Centrioli
Perossisomi
CELLULA VEGETALE
0,5 μm
25 nm
0,75 μm
0,75 μm
0,5 μm
1 μm
I vacuoli sono presenti soprattutto nel-
le cellule vegetali e nei protisti e con-
tengono una soluzione acquosa con
molte sostanze disciolte. Sono circon-
dati da una singola membrana e hanno
varie funzioni:
degli scarti della cellula;
-
cupano oltre il 90% del volume della
cellula e sono i responsabili del turgo-re della cellula stessa;
-
tali del fiore o dei frutti, che servono
ad attirare gli animali che favorisco-
no l’impollinazione o la dispersione
del seme;
-
te certi vacuoli contengono enzimi
che idrolizzano le proteine del seme
scindendole in monomeri utilizzabi-
li per l’alimentazione e lo sviluppo.
Le cellule animali possono avere va-
cuoli, ma di dimensioni ridotte. Le
cellule eucariotiche (sia vegetali che
animali) possono inoltre contenere ve-
scicole, con struttura simile ai vacuoli,
dimensioni molto inferiori e funzione
di trasporto.
Sui ribosomi vengono sin-
tetizzate le proteine.
L’apparato di Golgi ela-
bora e impacchetta le
proteine.
Proteine e altre molecole
tossiche vengono modi-
ficate chimicamente nel
reticolo endoplasmatico
liscio.
I cloroplasti utilizzano l’e-
nergia solare per produrre
zuccheri attraverso la foto-
sintesi.
Parete cellulare
Ribosomi liberi
Nucleolo
Vacuolo
Nucleo
Reticolo endoplasmatico ruvido
Reticolo endoplasmatico liscio
Perossisomi
Mitocondrio
Apparato di Golgi
PlasmodesmiMembrana plasmatica
Cloroplasti

B9
Bio
logi
a
B8
LE
ZIO
NE
04
04 Gli organuli cellulari
Le cellule eucariotiche sono tipiche dei membri del dominio degli eucarioti, che comprende i
protisti, i vegetali, i funghi e gli animali. Le dimensioni delle cellule eucariotiche sono fino a 10
volte più grandi di quelle delle cellule procariotiche.
Le cellule eucariotiche sono caratterizzate dalla presenza di compartimenti delimitati da mem-
brane, gli organuli, nei quali si verificano determinate reazioni chimiche in base agli enzimi
presenti.
Ciascun organulo svolge, nel proprio tipo particolare di cellula, un ruolo specifico definito dalle
reazioni chimiche che è capace di compiere. Consideriamo ora gli organuli uno per uno.
IL NUCLEO
IL RETICOLO ENDOPLASMATICO
L’osservazione al microscopio elettronico di una cellula eucariotica rivela una rete di membrane
interconnesse che si dirama per tutto il citoplasma della cellula eucariotica formando tubuli e sac-
chetti appiattiti. L’insieme di queste membrane prende il nome di reticolo endoplasmatico (RE).
In base all’aspetto, si distinguono il reticolo endoplasmatico ruvido (RER) e il reticolo endo-plasmatico liscio (REL).
Lo spazio interno del
RE, che è separato e
distinto dal citoplasma
circostante, si chiama
lume.
Nelle cellule eucariotiche
è presente un vasto
sistema di membrane
interne (endomembrane)
che delimitano comparti
separati del citoplasma.
I due comparti principali
di questo sistema sono il
reticolo endoplasmatico e
l’apparato di Golgi.
Gli organuli cellulari
Organulo Struttura Funzioni Presenza nelle cellule
Dimensioni
Nucleo racchiuso da una doppia
membrana
duplica il DNA, controlla l’attività
cellulare e, nel nucleolo, assembla i
ribosomi
animali e
vegetali
diametro circa
5 μm (nucleolo
1-3 μm)
Ribosomi formati da RNA e
proteine
sintetizzano le proteine animali e
vegetali
diametro circa
20 nm
Reticolo
endoplasmatico
ruvido
sistema di membrane
singole
sintetizza membrane e proteine animali e
vegetali
molto variabili
Reticolo
endoplasmatico
liscio
sistema di membrane
singole
sintetizza i lipidi, trasforma sostanze
tossiche, idrolizza il glicogeno e
accumula ioni calcio
animali e
vegetali
molto variabili
Apparato di
Golgi
sistema di membrane
singole
elabora e smista le proteine;
nelle cellule vegetali sintetizza i
polisaccaridi per la parete
animali e
vegetali
lunghezza circa
1 μm
Lisosomi racchiusi da una singola
membrana
digeriscono le macromolecole
idrolizzandole
solo animali diametro 0,1-1
μm
Perossisomi racchiusi da una singola
membrana
raccolgono e decompongono i
perossidi
animali e
vegetali
diametro 0,2-
1,7 μm
Vacuoli racchiusi da una singola
membrana
accumulano prodotti di scarto; nelle
cellule vegetali mantengono il turgore
e contengono pigmenti
animali e
vegetali
molto variabili
Mitocondri racchiusi da una doppia
membrana
producono energia tramite la
respirazione cellulare
animali e
vegetali
lunghezza
2-8 μm
Cloroplasti racchiusi da una doppia
membrana
producono energia tramite la
fotosintesi
solo vegetali lunghezza
2-10 μm
Altri plastidi
(cromoplasti e
leucoplasti)
racchiusi da una doppia
membrana
contengono pigmenti e accumulano
amido e grassi
solo vegetali diametro circa
2-10 μm
1 μm
Il nucleo è il luogo in cui avviene la duplicazione del DNA; esso contiene inoltre una zona,
il nucleolo, dove ha inizio il montaggio e la sintesi dei ribosomi a partire dall’RNA riboso-
miale e da specifiche proteine.
Il nucleo è avvolto da una doppia membrana che costituisce l’involucro nucleare. Le due
membrane dell’involucro nucleare sono separate da uno spazio di circa 20 nm e sono per-
forate da circa 3500 pori, che mettono il nucleo in comunicazione con il citoplasma.
0,5 μm
Nucleoplasma
Membrana esternaMembrana interna
L'involucro nucleare è in continuità con il reticolo endoplasmatico
Nucleolo
CromatinaInvolucro nucleare
Poro nucleare
Il reticolo endoplasmatico ruvido è costellato di ribosomi che sono i siti della sintesi proteica.
Essi sono responsabili del suo aspetto granuloso. Le proteine appena sintetizzate dai riboso-
mi entrano nel lume del reticolo e qui vengono modificate dal punto di vista chimico.
Il reticolo endoplasmatico
liscio svolge quattro funzio-
ni importanti:
-
pidi;
-
sformazione di sostanze
tossiche, farmaci e pesti-
cidi;
sede in cui avviene la sin-
tesi del glicogeno;
-
gazzinare ioni calcio.
Nucleo
Ribosomi
Lume
LumeInterno della cellula

B10 B11
BiologiaL E Z I O N E
Bio
logi
a
04
Gli organuli cellulari04 L’APPARATO DI GOLGI
I LISOSOMI
0,5 μm
L’apparato di Golgi elabora e impacchet-
ta le proteine.
Le vescicole contenenti proteine provenienti dal reticolo endopla-
smatico trasferiscono le sostanze all’apparato di Golgi.
L’apparato di Golgi è for-
mato da cisterne appiatti-
te, circondate da singola
membrana e svolge diver-
se funzioni:
-
ticolo endoplasmatico e
le elabora ulteriormen-
te (attaccandovi catene
di oligosaccaridi);
smista le proteine pri-
ma che vengano inviate
alle loro destinazioni
finali;
-
no sintetizzati i poli-
saccaridi per la parete
delle cellule vegetali.
Nucleo Interno della cellula
Esterno della cellulaProteine per uso esterno alla cellula
Proteine per uso interno alla cellula
Membrana plasmatica
Cisterne dell'apparato di Golgi
Reticolo endoplasmatico ruvido
Reticolo endoplasmatico liscio
I MITOCONDRI
I CLOROPLASTII lisosomi sono i siti dove
avviene l’idrolisi del mate-
riale assunto dalla cellula
per fagocitosi. Questi or-
ganuli isolano gli enzimi
digestivi dal citoplasma e
distruggono i batteri che
entrano nell’organismo e
i propri materiali (autofa-gia). Sono presenti solo
nelle cellule animali.
Un lisosoma primario
viene generato dal Golgi.
Il lisosoma si fonde
con un fagosoma.
Le piccole moleco-
le generate dalla di-
gestione diffondo-
no nel citoplasma.Il materiale non digerito
viene rilasciato all’esterno.
Lisosoma primario
Fagosoma
Particelle di cibo assunte per fagocitosi
Lisosoma secondario
Interno della cellula
Membrana plasmatica
Apparato di Golgi
Esterno della cellula
0,6 μm
La membrana interna è la barriera principale tra il citosol e gli enzimi mitocondriali; è molto
estesa e ripiegata a formare delle creste; è impermeabile a tutti gli ioni e alle molecole polari:
per attraversarla, queste sostanze devono utilizzare proteine di trasporto. Le membrane divi-
dono il mitocondrio in due parti: lo spazio intermembrana e la matrice (sostanza gelatinosa
delimitata dalla membrana interna).
Le creste contengono mo-
lecole cruciali per la pro-
duzione di ATP a partire
da altre molecole.
La membrana esterna è liscia e permeabile alla mag-
gior parte delle molecole e di piccoli ioni. Svolge una
funzione protettiva.
Spazio intermembrana
Matrice Creste
Membrana esternaMembrana interna
Matrice Creste
Membrana esterna
Membrana interna
La matrice, sostanza gelatinosa delimitata
dalla membrana interna, contiene riboso-
mi, DNA e diversi enzimi che partecipano
alla respirazione cellulare.
75 μm
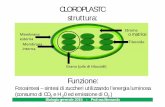
I cloroplasti sono organuli presenti solo nella cellu-
la vegetale: contengono il pigmento verde clorofilla
e sono la sede della fotosintesi.
Secondo la teoria endosimbiontica, come i mitocon-
dri, anche i cloroplasti erano dei procarioti fotosin-
tetici entrati in simbiosi con una cellula più grande:
contengono infatti ribosomi e un proprio DNA cir-
colare. Come i mitocondri, anche i cloroplasti sono
costituiti da una doppia membrana, una esterna e una
interna: in esse si trovano le molecole di clorofilla.
Le membrane interne del cloroplasto hanno l’aspet-
to di una pila di sacchetti discoidali. Queste pile,
dette grana, sono formate da una serie di compar-
timenti circolari piatti (i sacchetti), addossati l’uno
all’altro, che prendono il nome di tilacoidi.
Cloroplasti Cellula della foglia

B13
Bio
logi
a
B12
LE
ZIO
NE
05
05 Citoscheletro, ciglia e flagelli
IL CITOSCHELETRO
Oltre ai numerosi organuli dotati di membrana, il citoplasma eucariotico contiene una serie di fibre
lunghe e sottili che costituiscono il citoscheletro. I componenti essenziali del citoscheletro sono
i microfilamenti, i filamenti intermedi e i microtubuli, tre importanti strutture che mantengono e
rinforzano la forma della cellula e contribuiscono al suo movimento.
Il citoscheletro svolge varie funzioni importanti: sostiene la cellula e ne mantiene la forma; è alla
base del movimento cellulare; tiene in posizione gli organuli all’interno della cellula; alcune fibre
del citoscheletro funzionano da binari o da sostegni per proteine motrici che spostano gli organuli
all’interno della cellula; infine, interagisce con strutture extracellulari contribuendo a mantenere
in sede la cellula.
25 nm
Dimero di tubulina Subunità fibrosa Monomero
di β-tubulina
β α
7 nm
Monomerodi α-tubulina
Monomero di actina
Filamenti intermediMicrofilamenti Microtubuli
8–12 nm
20 μm 10 μm 10 μm
EstremitàEstremità+–EstremitàEstremità
+–
CIGLIA E FLAGELLI
Molte cellule eucariotiche presentano ciglia o flagelli, che utilizzano per muoversi nell’ambiente
circostante. Sia le ciglia che i flagelli eucariotici derivano dal montaggio di speciali microtubuli
e hanno un’identica struttura interna, anche se sono diversi per lunghezza e per tipo di battito. Il
movimento deriva dallo scorrimento l’una sull’altra delle doppiette di microtubuli.
I flagelli eucariotici
sono diversi dai flagelli
procariotici.
Vedremo nelle prossime
lezioni che cos’è l’ATP e
che cosa sono i centrioli.
~50 nm
La sezione trasversale rivela la disposi-
zione «9+2» dei microtubuli, con le nove
coppie di microtubuli fusi…
… e i due microtubuli più interni non fusi.
Da uno dei due microtubuli di ogni doppietta parte un raggio che collega la doppietta con il
centro della struttura. Le nove doppiette microtubulari di ciglia e flagelli sono tenute insie-
me dalla proteina dineina, che ha attività ATPasica.
Nel citoplasma, alla base di ciascun ciglio o flagello eucariotico, c’è un organulo chiamato
corpuscolo basale, nel quale si prolungano le nove doppiette di microtubuli. Quasi uguali ai
corpuscoli basali delle ciglia e dei flagelli sono i centrioli.
Doppietto di microtuboli
«Raggi»
Proteina motrice (dineina)
Proteina di connessione (nexina)
Questa sezione trasversale di singolo ciglio mostra la disposizione dei microtuboli e delle proteine.
Microfilamenti
-
teina actina, spesso interagiscono
con filamenti di altre proteine.
-
lula e guidano il movimento
cellulare, che comprende la
contrazione, lo scorrimento del
citoplasma e la costrizione del
corpo cellulare che prelude alla
divisione cellulare.
-
lamenti di miosina, determina-
no la contrazione muscolare.
Filamenti intermedi
-
teine fibrose orga-
nizzate in strutture
robuste a forma di
fune che stabiliz-
zano la struttura
della cellula e con-
tribuiscono a man-
tenerne la forma.
intermedi aiutano
a tenere insieme
due cellule vicine.
Microtubuli
-
lecole della proteina tubulina, che consiste
di due subunità, α-tubulina e β-tubulina.
in seguito all’aggiunta o alla sottrazione di
dimeri di tubulina.
cromosomi.
movimento delle cellule.
movimento delle vescicole trasportate dal-
le proteine motrici.

B14 B15
LE
ZIO
NE
LE
ZIO
NE
Bio
logi
a06 L’ATP
LA STRUTTURA DELL’ATP
Per la cattura e il trasferimento dell’energia libera necessaria a svolgere lavoro, le cellule uti-
lizzano una particolare molecola: l’adenosintrifosfato o ATP. L’ATP agisce come una sorta
di «moneta» dell’energia. Così come
gli esseri umani guadagnano denaro
con il proprio lavoro e poi lo spen-
dono per comprare ciò di cui hanno
bisogno, così le cellule utilizzano
una parte dell’energia libera prodot-
ta dalle reazioni esoergoniche per
costruire l’ATP, che poi potrà essere
usata per alimentare reazioni endo-
ergoniche.
L’IDROLISI DELL’ATP
L’ATP è un nucleotide prodotto dalle cellule in vari modi
e utilizzato per diversi scopi. La molecola dell’ATP è for-
mata da una base azotata, l’adenina, legata a una molecola
di ribosio che porta attaccata una sequenza di tre gruppi
fosfato, tenuti insieme da legami fosfodiestere.
CH2O–O P
O
O–
PP O
O
O–
O
O
O–
H
H H
OH OH
NH2
H
H
H
N
N
N
NC
C
C
C
C
O
C C
–
Adenina
Ribosio
Adenosina
(adenosinmonofosfato)AMP
(adenosindifosfato)ADP
(adenosintrifosfato)ATP
Gruppi fosfato
Dal punto di vista energetico
l’ATP ha due proprietà che lo
rendono particolarmente utile
alla cellula: quando viene idro-
lizzato libera una quantità consi-
stente di energia; può fosforilare,
cioè donare un gruppo fosfato,
a molte molecole diverse. L’i-
drolisi di uno dei legami fosfo-
diesfere dell’ATP libera energia
(7 kcal/mol), con formazione di
ADP (adenosindifosfato) e uno
ione fosfato (HPO42–, abbreviato
in Pi):
EnergiaEnergia
ATP
ADP
+ Pi
Reazione esoergonica:(libera energia)⁰ Respirazione cellulare ⁰ Catabolismo
Reazione endoergonica:(richiede energia)⁰ Trasporto attivo⁰ Movimenti cellulari⁰ Anabolismo
La sintesi di ATP da ADP e Pi richiede energia. Le rea-
zioni cellulari esoergoniche liberano l’energia neces-
saria per sintetizzare ATP a partire da ADP e fosfato.
L’idrolisi di ATP in ADP e Pi libera energia. L’energia
liberata dall’idrolisi dell’ATP in ADP + Pi è utilizzata
dalla cellula per alimentare le reazioni endoergoniche.
07
07Gli enzimi
LA CATALISI ENZIMATICA
Nelle reazioni chimiche, la soglia energetica da superare perché la reazione avvenga è detta ener-gia di attivazione. Esistono sostanze in grado di abbassare l’energia di attivazione, e accelerare
così la reazione stessa: sono dette catalizzatori.
Un catalizzatore è una sostanza che accelera una reazione chimica senza consumarsi
durante la trasformazione.
I catalizzatori biologici più comuni sono gli enzimi, che sono complesse molecole proteiche
molto specifiche: un enzima catalizza un’unica reazione chimica, riconosce in genere un solo
reagente o gruppo di reagenti, si lega a esso ed esce inalterato dalla reazione che catalizza.
Il nome degli enzimi deriva dalla loro funzione specifica e spesso termina col suffisso -asi. In particolare, esistono sei classi di enzimi:
le ossidoreduttasi, che catalizzano reazioni redox;
le transferasi, che catalizzano reazioni di trasferimento di gruppi tra due substrati;
le idrolasi, che catalizzano reazioni di idrolisi;
le liasi, che catalizzano reazioni di addizione ai doppi legami;
le isomerasi, che catalizzano reazioni di isomerizzazione;
le ligasi, che catalizzano reazioni di sintesi.
L’enzima RNA-polimerasi
catalizza la sintesi
dell’RNA, ma non quella
del DNA. Ogni enzima
infatti è specifico per la
reazione che catalizza.
In una reazione catalizzata
da un enzima, i reagenti
prendono il nome di
substrati. Le molecole
di substrato si legano
a un determinato sito
dell’enzima, chiamato
sito attivo, dove avviene
la catalisi.
Negli enzimi allosterici una molecola si può
legare specificamente
all’enzima (di solito in
posizione diversa dal
sito attivo), inducendo
un cambiamento
conformazionale
dell’enzima, e
una conseguente
variazione dell’attività
dell’enzima, che può
essere sia positiva
(attivazione allosterica),
che negativa (inibizione
allosterica).
Enzima Complesso enzima-substrato Enzima
Sito attivo Prodotto
Le molecole di substra-
to si adattano perfetta-
mente al sito attivo…
La specificità di un enzima deriva dalla pre-
cisa forma tridimensionale del suo sito at-
tivo, al quale si adatta soltanto una ristretta
gamma di substrati (a volte anche solo uno).
Le molecole diverse per forma, gruppi fun-
zionali o proprietà non riescono ad aderire
al sito attivo e legarsi a esso.
La demolizione del complesso enzima-substrato gene-
ra il prodotto. L’enzima è nuovamente disponibile per
catalizzare un’altra reazione.
… mentre molecole di-
verse non vi si adattano.
ATP (modello compatto) ATP (formula di struttura)

B16 B17
BiologiaL E Z I O N E
Bio
logi
a
07
07 Gli enzimi
FATTORI CHE INFLUENZANO GLI ENZIMI
La struttura tridimensionale degli enzimi e le proprietà chimiche delle catene laterali dei loro siti
attivi li rendono fortemente sensibili al pH e alla temperatura.
I COENZIMI
Talvolta, perché si manifesti l’attività di un enzima, è necessaria la partecipazione di molecole
non proteiche:
i coenzimi sono composti organici, in genere di dimensioni relativamente piccole rispetto
all’enzima con cui formano un legame transitorio: uno stesso coenzima può essere utilizzato
anche da più enzimi diversi. Sono i principi attivi delle vitamine;
i cofattori sono ioni inorganici come lo zinco o il ferro;
i gruppi prostetici sono gruppi molecolari legati covalentemente all’enzima.
Il riscaldamento tende ad aumentare la velocità di una reazione enzimatica perché a temperatu-
re più alte aumenta la percentuale di molecole di reagente dotate di energia cinetica sufficiente a
fornire l’energia di attivazione per la reazione. Tuttavia, se la temperatura diventa troppo elevata,
gli enzimi iniziano a disattivarsi perché alcuni dei legami deboli presenti all’interno della loro
molecola si spezzano. Quando poi il calore arriva ad alterare la struttura terziaria, si dice che
l’enzima si denatura e perde la propria funzionalità.
Alcuni enzimi, come
quelli dei batteri
estremofili, agiscono
anche ad alte
temperature.
Velo
cità
di r
eazi
one
1 2 3 4 5 6 7 8 9 10 11pHAcido Basico
12
Ciascun enzima pre-
senta un picco di atti-
vità in corrispondenza
di un particolare valore
di pH. Per esempio, la
pepsina è una protea-
si particolarmente atti-
va nell’ambiente acido
del lo stomaco.
Amilasi salivare
Pepsina Arginasi
Velo
cità
di r
eazi
one
Temperatura
Un enzima è più attivo a una determina-
ta temperatura ottimale; a temperature
superiori, la denaturazione ne riduce o
annulla l’attività.
Velocità massima
Temperatura ottimale
Il coenzima NAD deriva dalla vitamina B2
(nicotinammide adenin dinucleotide).
Grazie alla sua capacità di trasportare ener-
gia ed elettroni, è uno dei principali tra-
sportatori di energia delle reazioni redox,
nonché un intermediario universale degli
scambi energetici cellulari. Si presenta in
due forme chimicamente distinte: la forma
ossidata NAD+ e la forma ridotta NADH.
La reazione di riduzione NAD+ + 2H
NADH + H+ equivale al trasferimento di
due atomi di idrogeno (2H+ + 2e-).
Forma ossidata (NAD+)+
NAD+AH2 BH2
A BNADH
H+
N
N
N
CH2
H HH H
OH OH
O
NH2
OP
N
CH2
H HH H
OH OH
O
H
OP
CONH2
+N
H
CONH2
+
O
O
O–
O–
O
H
+NADH2HH+
H+Ogni freccia nera
ricurva rappresen-
ta una reazione di
ossidazione o di ri-
duzione. La freccia
larga in blu indica il
percorso degli elet-
troni trasferiti.… lasciando
libero un H+.
La struttura ad anel-
lo del NAD+ acqui-
sta 2e- e 1H+…
Due atomi di idro-
geno (2e- + 2H+)
vengono trasferiti.Ossidazione Ossidazione
Ossidazione
Riduzione Riduzione
Riduzione
Forma ridotta
Un altro coenzima
comune che trasferisce
elettroni durante il
metabolismo del glucosio
è il FAD, (flavin adenin dinucleotide).

B19
Bio
logi
a
B18
LE
ZIO
NE
08
08
I carboidrati possono essere
legati alla superficie esterna
delle proteine (formando glico-
proteine) o dei lipidi (forman-
do glicolipidi).
Nelle cellule anima-
li, alcune proteine di
membrana sono asso-
ciate a filamenti della
matrice extracellulare.
Alcune proteine in-
tegrali attraversano
completamente il dop-
pio strato fosfolipidi-
co; altre vi sono inse-
rite solo parzialmente.
Alcune proteine per-
mettono alle cellule di
aderire l’una all’altra.
Le proteine periferiche di
membrana non sono inseri-
te nel doppio strato fosfoli-
pidico e sono prive di regio-
ni idrofobe.
Le molecole di colesterolo disperse nel dop-
pio strato fosfolipidico, fra le code idrofobe
dei fosfolipidi, condizionano la fluidità degli
acidi grassi della membrana (nelle cellule
animali).
Alcune proteine di
membrana interagi-
scono con il citosche-
letro della cellula.
Una proteina intrinseca, o integrale, è «agganciata» all’interno della
membrana grazie alle catene laterali idrofile e idrofobe dei suoi amminoa-
cidi. Le estremità idrofile si protendono nell’ambiente acquoso esterno alla
cellula e sul versante citoplasmatico; le catene idrofobe sono invece rivolte
verso l’interno lipidico della membrana.
Nuove porzioni di membrana possono esse-
re prodotte in punti particolari, per esempio
del reticolo endoplasmatico.
La membrana che si forma viene ricavata da
quella plasmatica mediante endocitosi.
Le proteine di membrana sono inserite
nel RER.
Anche le vescicole che gemmano
dall’apparato di Golgi sono rivestite da
membrana.
Le vescicole possono rimanere all’in-
terno della cellula in forma di organuli,
come accade per i lisosomi.
Oppure esse possono fondersi con la
membrana plasmatica e liberare il loro
contenuto all’esterno della cellula (eso-
citosi). La loro membrana viene così
integrata nella membrana plasmatica.
Le membrane biologiche hanno in comune molte pro-
prietà perché condividono la medesima organizzazione
molecolare. Esse devono la propria struttura fisica e il
proprio funzionamento ai loro costituenti: lipidi, protei-
ne e carboidrati. I componenti principali della membra-
na sono i fosfolipidi che sono organizzati a formare un
doppio strato, con le code polari e idrofobe rivolte verso
l’interno della membrana, le teste polari verso l’interno
della cellula. Il doppio strato fosfolipidico serve da «la-
go» lipidico in cui «galleggiano» varie proteine. Questa
struttura generale va sotto il nome di modello a mosai-co fluido.
La struttura delle membrane biologiche
Le membrane biologiche sono strutture fondamentali per l’attività della cellula: sono altamente
organizzate, permettono alle cellule di interagire tra loro e regolano gli scambi con l’ambiente.
Esterno della cellula
Doppio strato fosfolipidico
Interno della cellula
La struttura delle membrane biologiche è comune a tutte le membrane, sia quelle che delimitano
la cellula sia quelle che rivestono gli organuli e le vescicole.
Negli eucarioti, la membrana plasmatica viene continuamente rinnovata per compensare le per-
dite dovute alla fagocitosi.
Nelle cellule eucariotiche esiste una stretta relazione tra tutte le membrane presenti, che sono
strutture estremamente dinamiche: si formano, passano da un organulo all’altro, si interrompo-
no e si fondono fra loro. All’interno delle cellule eucariotiche, inoltre, intere porzioni di mem-
brane con caratteristiche strutturali diverse si spostano seguendo la propria funzione.
Nucleo
Reticolo endoplasmatico ruvido
Reticolo endoplasmatico liscio
EsocitosiEndocitosi
Apparato di Golgi
Ambiente intracellulare
Lisosoma
Membrana plasmaticaAmbiente extracellulare