
Kosmos Numer 4 (249) Tom 49, 2000
21
K osmos Tom 49, 2000 Numer 4 (249) Strony 537-557 PROBLEMY NAUKBIOLO G^CZNYCH_____________ Polskie Towarzystwo Przyrodników im. Kopernika J anina K aczanowska , D orota W łoga , M auryla K iersnowska , Ewa joachimiak i A ndrzej K aczanowski Zakład Cytofizjologii, Instytut Zoologii, Uniwersytet Warszawski Miecznikowa 1, 02-096 Warszawa ROLA CYTOSZKIELETU W ROZWOJU ORGANIZMÓW JEDNOKOMÓRKOWYCH (NA PRZYKŁADZIE ORZĘSKA TETRAHYMENA I PORÓWNAWCZO DROŻDŻA SCHISOSACCHAROMYCES POMBE) RÓŻNORODNE FUNKCJE CYTOSZKIELETU W REGULACJI ROZWOJU KOMÓREK EUKARIOTYCZNYCH Cytoszkielet wszystkich komórek organi- zmów eukariotycznych jest zbudowany głównie z włókien mikrotubularnych i mikrofilamentów aktynowych wraz z towarzyszącymi im białkami motorycznymi, z cytoszkieletu podbłonowego (złożonego głównie ze spektiyn i sieciowanej aktyny) oraz z tzw. filamentów pośrednich. Tym białkom strukturalnym towarzyszy bardzo wie- le białek regulujących stabilność i przemiany w cytoszkielecie. Cytoszkielet jest nie tylko stru- kturą komórki stanowiącą jej trwałe rusztowa- nie i wyznaczającą jej kształt, ale pełni w niej również inne funkcje. Mechanizm ruchu komó- rek opiera się na funkcjonowaniu cytoszkieletu rzęsek, wici i przemieszczających się po podłożu wypustek cytoplazmatycznych wspartych stru- kturami cytoszkieletu. Cytoszkielet buduje również szlaki komunikacji wewnątrz komórki umożliwiając przemieszczanie się poszczegól- nych organelli komórkowych. Jądra, mitochon- dria, czy też pęcherzyki sekrecyjne mogą się podczepiać do białek motoiycznych, a nastę- pnie wraz z nimi przemieszczać po włóknach mikrotubul lub mikrofilamentów do punktów docelowych, gdzie podlegają zakotwiczaniu. W ten sposób cytoszkielet uczestniczy w endocy- tozie i sekrecji. Cytoszkielet podtrzymuje także terytorialne zróżnicowanie komórki decydując między innymi o rozmieszczeniu różnych rodza- jów białek, a także transkiyptów (mRNA róż- nych białek), które mogą być docelowo tran- sportowane, a następnie zakotwiczane do błony komórkowej i tam dopiero wykorzystywane do syntezy białek (Hazelrigg 1998). Kształt typowej komórki pobranej z organi- zmu Metazoa, np. fibroblastów lub komórek nabłonkowych, może się zmieniać w zależności od tego, czy komórka przylega do podłoża lub do innych komórek, czy też jest zawieszona w środowisku płynnym, a także od tego czy jest to komórka spoczynkowa, czy też dzieli się na dwie komórki potomne. Cytoszkielet zmienia się w zależności od różnicujących sygnałów z otocze- nia, a także od sygnałów wewnętrznych związa- nych z przebiegiem cyklu komórkowego i two- rzeniem polarności komórki. A więc cytoszkielet jest strukturą dynamiczną, podlegającą reorga- nizacji na drodze polimeryzacji i depolimeiyza- cji włókien, zarówno w cyklu podziałów mitoty- cznyeh, jak i w trakcie procesów różnicowania. Cytoszkielet jest nie tylko odbiorcą sygna- łów rozwojowych, ale także pełni funkcje regu- latora w szlakach przekazywania sygnałów ze środowiska do komórki. A oto znamienny przy- kład: komórki linii 3T3 myszy w obecności czyn- ników wzrostu zaczynają przylegać do macierzy pozakomórkowej znajdującej się na dnie szalki i wchodzą w replikację DNA i podziały komór- kowe. Pobudzenie syntezy DNA odbywa się przez aktywację czynnikami wzrostu recepto- rów integrynowych obecnych w błonie komó- rek. Ta aktywacja prowadzi z jednej strony do przylegania komórek do podłoża, a z drugiej sygnał ten jest przekazywany do jądra inicjując syntezę DNA. Otóż, przyleganie do podłoża ko- mórek prowadzi do lokalnej reorganizacji cyto- szkieletu, tworzą się wtedy miejsca przylegania (ang. foci) podtrzymywane spolimeryzowanymi
Transcript of Kosmos Numer 4 (249) Tom 49, 2000

KosmosTom 49, 2000Numer 4 (249)Strony 537-557
PROBLEMY NAUKBIOLO G^CZNYCH_____________Polskie Towarzystwo Przyrodników im. Kopernika
Ja n in a Ka c z a n o w s k a , D o r o ta W ło g a , Ma u r y l a K ie r s n o w s k a , Ew a j o a c h im ia k i A n d r ze j Ka c zan o w sk i
Zakład Cytofizjologii, Instytut Zoologii,Uniwersytet Warszawski Miecznikowa 1, 02-096 Warszawa
ROLA CYTOSZKIELETU W ROZWOJU ORGANIZMÓW JEDNOKOMÓRKOWYCH (NA PRZYKŁADZIE ORZĘSKA TETRAHYMENA I PORÓWNAWCZO DROŻDŻA
SCHISOSACCHAROMYCES POMBE)
RÓŻNORODNE FUNKCJE CYTOSZKIELETU W REGULACJI ROZWOJU KOMÓREKEUKARIOTYCZNYCH
Cytoszkielet wszystkich komórek organizmów eukariotycznych jest zbudowany głównie z włókien mikrotubularnych i mikrofilamentów aktynowych wraz z towarzyszącymi im białkami motorycznymi, z cytoszkieletu podbłonowego (złożonego głównie ze spektiyn i sieciowanej aktyny) oraz z tzw. filamentów pośrednich. Tym białkom strukturalnym towarzyszy bardzo wiele białek regulujących stabilność i przemiany w cytoszkielecie. Cytoszkielet jest nie tylko strukturą komórki stanowiącą jej trwałe rusztowanie i wyznaczającą jej kształt, ale pełni w niej również inne funkcje. Mechanizm ruchu komórek opiera się na funkcjonowaniu cytoszkieletu rzęsek, wici i przemieszczających się po podłożu wypustek cytoplazmatycznych wspartych strukturami cytoszkieletu. Cytoszkielet buduje również szlaki komunikacji wewnątrz komórki umożliwiając przemieszczanie się poszczególnych organelli komórkowych. Jądra, mitochondria, czy też pęcherzyki sekrecyjne mogą się podczepiać do białek motoiycznych, a następnie wraz z nimi przemieszczać po włóknach mikrotubul lub mikrofilamentów do punktów docelowych, gdzie podlegają zakotwiczaniu. W ten sposób cytoszkielet uczestniczy w endocy- tozie i sekrecji. Cytoszkielet podtrzymuje także terytorialne zróżnicowanie komórki decydując między innymi o rozmieszczeniu różnych rodzajów białek, a także transkiyptów (mRNA różnych białek), które mogą być docelowo transportowane, a następnie zakotwiczane do błony komórkowej i tam dopiero wykorzystywane do syntezy białek (H a z e l r i g g 1998).
Kształt typowej komórki pobranej z organizmu Metazoa, np. fibroblastów lub komórek nabłonkowych, może się zmieniać w zależności od tego, czy komórka przylega do podłoża lub do innych komórek, czy też jest zawieszona w środowisku płynnym, a także od tego czy jest to komórka spoczynkowa, czy też dzieli się na dwie komórki potomne. Cytoszkielet zmienia się w zależności od różnicujących sygnałów z otoczenia, a także od sygnałów wewnętrznych związanych z przebiegiem cyklu komórkowego i tworzeniem polarności komórki. A więc cytoszkielet jest strukturą dynamiczną, podlegającą reorganizacji na drodze polimeryzacji i depolimeiyza- cji włókien, zarówno w cyklu podziałów mitotyczny eh, jak i w trakcie procesów różnicowania.
Cytoszkielet jest nie tylko odbiorcą sygnałów rozwojowych, ale także pełni funkcje regulatora w szlakach przekazywania sygnałów ze środowiska do komórki. A oto znamienny przykład: komórki linii 3T3 myszy w obecności czynników wzrostu zaczynają przylegać do macierzy pozakomórkowej znajdującej się na dnie szalki i wchodzą w replikację DNA i podziały komórkowe. Pobudzenie syntezy DNA odbywa się przez aktywację czynnikami wzrostu receptorów integrynowych obecnych w błonie komórek. Ta aktywacja prowadzi z jednej strony do przylegania komórek do podłoża, a z drugiej sygnał ten jest przekazywany do jądra inicjując syntezę DNA. Otóż, przyleganie do podłoża komórek prowadzi do lokalnej reorganizacji cytoszkieletu, tworzą się wtedy miejsca przylegania (ang. foci) podtrzymywane spolimeryzowanymi

538 Janina Kaczanowska i w spółau torzy
wiązkami aktyny, natomiast spada stabilność cytoplazmatycznych mikrotubul. Zatem w obecności czynników wzrostu adhezja komórek do podłoża wraz z reorganizacją cytoszkieletu stanowi sygnał inicjacji syntezy DNA i namnażania się komórek. Jednakże komórki 3T3, nawet przy braku czynników wzrostu, mogą tworzyć miejsca przylegania, jeżeli ich wewnętrzny stabilizujący cytoszkielet mikrotubularny zostanie zdemontowany (przez podanie nokodazolu lub winblastyny). Tej wymuszonej adhezji towarzyszy także wejście komórek w replikację DNA (B e rs h a d s k y i współaut. 1996). Tak więc organizacja cytoszkieletu ma bezpośredni wpływ na proliferację komórek. Inne doświadczenia (SOR- g e r i współaut. 1997) wykazały, że obecność lub odwrotnie depolimeiyzacja struktur mikro- tubularnych wpływają na dalsze losy komórek indukując apoptozę (w przypadku demontażu cytoszkieletu) lub różnicowanie (przy cytoszkie- lecie stabilnym). Te obserwacje wykazują, że zmiany w cytoszkielecie mogą same stanowić wewnętrzne sygnały aktywujące ścieżki przekażnictwa kontrolujące zarówno namnaża- nie się, jak też apoptozę i różnicowanie komórek.
Dobrym modelem do badania udziału cytoszkieletu w rozwoju są wolnożyjące jednokomórkowe organizmy zdolne do procesów płciowych (a więc takie, u któiych można badać te procesy stosując metody genetyki rozwoju), jak komórki drożdży Schisosaccharomyces pombe i orzęsków z rodzaju Tetrahymena. Te organizmy stały się modelowymi obiektami dla badania roli cytoszkieletu w rozwoju. W komórkach spoczynkowych (nie dzielących się) ich cytoszkiele- ty tworzą bardzo określony wzór morfologiczny. Wznowienie cykli komórkowych, a także podjęcie procesów płciowych przez komórki, wiąże się z zasadniczą reorganizacją tego wzoru, to znaczy z reorganizacją cytoszkieletu. A więc są to modele, w których można badać zależność pomiędzy przemianami w cytoszkielecie a realizacją programu rozwoju. Obserwacje tych zmian pozwoliły wyznaczyć rolę cytoszkieletu w przebiegu szeregu procesów rozwoju komórek, w których udział i regulacyjna rola cytoszkieletu była zupełnie nieoczekiwana.
W tym artykule rozważymy rolę cytoszkieletu:
— w różnicowaniu biegunów komórki, a więc omówimy zagadnienie powstawania polar- ności komórki i wyznaczania stref wzrostu w trakcie cyklu komórkowego:
— w regulacji transkrypcji genów kodujących struktury cytoszkieletalne (zjawisko jak dotąd wykryte tylko w komórce Tetrahymena) i
regulacji posttranslacyjnej stabilności struktur mikrotubularnych.
Kolejnym przedstawionym zagadnieniem będzie udział cytoszkieletu w procesie płciowym (koniugacji): utworzeniu połączenia koniugują- cych komórek, indukcji mejozy i regulacji przebiegu dalszych etapów procesu płciowego Tetrahymena. Uwarunkowania wejścia komórek w mejozę u S. pombe są inne i wobec tego te procesy nie będą tutaj omawiane.
W szczepach Tetrahymena thermophila opisano wielokrotnie proces starzenia się klonów wegetatywnych przejawiający się wzrostem liczby komórek bezpłodnych, niezdolnych do tworzenia pronukleusów. Szereg badań wskazuje na to, że bezpłodność ta jest związana ze zmianami starczymi w tworzeniu i funkcjonowaniu cytoszkieletu.
A oto bardziej szczegółowy zarys poszczególnych zagadnień.
Pierwsze pytanie dotyczy roli cytoszkieletu podbłonowego i mikrotubul w wyznaczaniu po- larności komórki i określenie strefy wzrostu komórki. Mikrotubularne organellum komórki eukariotycznej nazwane centrosomem w komórkach zwierząt, komórkach niższych roślin i wielu pierwotniaków, wyznacza biegun przedni komórki. Na elektronogramach centrosom ma postać spolaryzowanej pałeczki złożonej ze struktury wewnętrznej przypominającej tzw. „koło zębate” (ang. cartwheel structure) i dziewięciu koliście rozmieszczonych potrójnych włókienek mikrotubularnych. Centrosom jest otoczony warstwą białek pericentrosomalnych i razem z nimi tworzy centriolę. Komórka w trakcie przygotowania się do podziału odtwarza centriolę potomną usytuowaną pod kątem prostym w stosunku do centrioli macierzystej. Centriola może pełnić różne funkcje w komórce; np. w migrującej komórce ameboidalnej centriola wraz z aparatem Golgiego wyznacza kierunek ruchu i biegun przedni komórki, a w organizmach posiadających rzęski lub wici centriola w kontakcie z błoną komórkową staje się ciałkiem podstawowym. Położenie ciałka podstawowego reguluje tworzenie potomnych ciałek podstawowych w okolicy tej struktury macierzystej i wyznacza miejsca tworzenia wici i rzęsek (K r io u tc h k o v a i O n ish ch en k o 1999).
W komórkach roślin wyższych i innych typach komórek acentriolarnych (np. zmutowanych liniach zwierzęcych, w których brak centrioli) wykazano, że białka pericentrosomalne, a nie struktura centrosomu, są odpowiedzialne za organizację biegunów wrzeciona mitotycznego.
Ale są i inne sposoby wyznaczania biegunów wrzeciona mito tycznego, np. u ssaków komórka

Rola cytoszkieletu w rozwoju organizmów 539
jajowa jest pozbawiona centrosomu lub ma tylko pojedyńczą centriolę. Przy zapłodnieniu plemnik wnosi nie tylko haploidalne jądro, ale także centriolę wyznaczającą oś ciała zygoty (Hable i Kroft 2000). Miejsce wniknięcia plemnika z dwiema centriolami do acentriolarnego jaja nicienia Caenorhabditis elegans wyznacza biegun tylny zygoty, a równocześnie na przeciwległym końcu przednim zachodzi gromadzenie się aktyny. Tworzy się zatem spolaryzowana oś przednio-tylna zarodka i względem tych struktur polarnych odbywa się asymetryczne rozmieszczanie zarówno ziarnistości biegunowych tylnych, jak i asymetryczne zgrupowanie różnych receptorów i podbłonowych kinaz.
Powstają pytania, jak odbywa się wytworzenie biegunów komórki w trakcie podziału komórki o mitozie wewnątrzjądrowej (np. Schi- sosaccharomyces pombe) oraz w komórce także o mitozie wewnątrzjądrowej, ale trwale spolaryzowanej , w której oś ciała wyznaczają powielone spolaryzowane rzędy ciałek podstawowych (np. Tetrahymena).
W trakcie podziału mitotycznego komórki centriolarnej dwie centriole, macierzysta i potomna, stanowią bieguny wrzeciona mitotycznego i wyznaczają dwubiegunową oś tego podziału. W przypadku komórek acentriolarnych, takich jak komórki roślin wyższych, funkcje biegunów wrzeciona mitotycznego spełniają centra mikrotubulogenezy — MTOC (ang. mic- rotubule organizing center). W linii zmutowanych acentriolarnych komórek macierzystych Drosophila hodowanych in vitro wykazano, że specyficzna odmiana białka tubulinowego, a mianowicie y-tubulina, odgrywa zasadniczą rolę w organizacji biegunów wrzeciona. W stadium cyklu komórki, który sprzyja polimeryzacji stabilnych mikrotubul, y-tubulina pojawia się w centrach mikrotubulogenezy tworząc funkcjonalne bieguny mitotyczne. Wynika stąd wniosek, że warunki aktywujące wytworzenie dwóch lub więcej MTOC w komórce mogą stopniowo prowadzić do wytworzenia dwubiegunowej osi komórki, często z wtórną eliminacją nadliczbowych MTOC (Debec i współaut. 1995). Obecnie wiadomo, że wytworzenie dwubiegunowego wrzeciona związane jest także z polimeryzacją mikrotubul w bezpośrednim sąsiedztwie kondensujących chromosomów i wymaga aktywności kinazy fosforylującej histon H3 (kinaza typu Aurora), współdziałających z fosfatazą (typu PP1) (Hsu i współaut. 2000) i innymi kinazami towarzyszącymi funkcjonowaniu wrzeciona mitotycznego (Robinson i Spu- DICH 2000).
W komórkach o mitozie centriolarnej bieguny wrzeciona mitotycznego są źródłem sygnału
wyznaczającego oś komórki i położenie bruzdy cytokinetycznej rozdzielającej komórki potomne. Tworzone na centrosomie mikrotubule są strukturami polarnymi; ich koniec minus (-) jest przy centrosomie, natomiast peryferyczne końce stanowią końce plus (+). Polaryzacja mikrotubul stanowi instrukcję strukturalną dla białek motorycznych i przyległej błony komórkowej. W okresie poprzedzającym podział mito- tyczny komórki, dwubiegunowe wrzeciono mi- totyczne zakotwicza się biegunami do błony komórkowej. Zakotwiczenie to stabilizuje oś wrzeciona mitotycznego, a sygnały wysyłane z biegunów wyznaczają prostopadłe położenie bruzdy cytokinetycznej (Robinson i Spudich 2000). Zbudowanie prawidłowej struktury wrzeciona mitotycznego w stadium metafazy stanowi punkt kontrolny (ang. spindle checkpoint) dla przejścia komórki w stadium anafazy i wytworzenia bruzdy cytokinetycznej (Rudner i Murray 1996). Ten sygnał oznacza równocześnie proteolizę szeregu białek, w tym białka łączącego dwie chromatydy i cyklin stanowiących składnik MPF (ang. mitosis promoting factor), oraz rozpoczęcie demontażu struktur mikrotu- bularnych wrzeciona. Natomiast bardziej zagadkowy jest udział pewnej kinazy białkowej, POLO-kinazy, związanej z białkiem motorycz- nym typu kinezyn (u Drosophila nazwanego białkiem PAVAROTTI), a także innych kinaz białkowych, np. NIMA , Aurora i innych (Fry 1998, Hsu i współaut. 2000). Te kinazy początkowo mają różną lokalizację w komórce, ale w trakcie funkcjonowania wrzeciona przemieszczają się do bruzdy cytokinetycznej. Mutanty zarówno POLO-kinazy, jak i kinezyny (PAVAROTTI) wchodzą w mitozę, osiągają telofazę, ale nie tworzą pierścienia cytokinetycznego (Adams i współaut. 1998).
A więc zarówno w przypadku mitoz centrio- larnych, jak i acentriolarnych wyznaczenie osi wrzeciona mitotycznego i biegunów komórki odgrywa zasadniczą rolę w wyznaczaniu położenia bruzdy cytokinetycznej (Robinson i Spudich 2000). Z kolei, wyznaczanie położenia bruzdy podziałowej może warunkować przebieg dalszych procesów rozwojowych, jak to ma miejsce w bruzdkowaniu zarodka nicienia Caenorhabditis elegans. Prawidłowe wyznaczenie położenia bruzd cytokinetycznych jest pod kontrolą genów programu rozwoju i jest niezbędne dla przeżycia zarodka i prawidłowego przebiegu różnicowania blastomerów (Watts i współaut. 2000).
Zarówno u grzybów (drożdże), jak i u orzę- sków mitoza odbywa się wewnątrz otoczki jądrowej. Jest to tzw. acentriolarna mitoza zamknięta, w której w kontakcie z otoczką jądro

540 Janina Kaczanowska i współau torzy
wą tworzą się bieguny zwane ciałkami polarnymi — SPB (ang. spindle polar body), w przeciwieństwie do mitoz otwartych, w których otoczka jądrowa rozprasza się w stadium tworzenia dwubiegunowego wrzeciona. Powstaje pytanie, jaki jest mechanizm przestrzennej zależności między wytworzeniem osi komórki a wyznaczeniem położenia bruzdy podziałowej w przypadku komórki apolarnej (S. pombe) i komórki stale spolaryzowanej (Tetrahymena). Tetrahymena (Rye. 1) jest pobudliwą, trwale spolaryzo-
Ryc.l. Schemat wzoru rozmieszczenia mikrotubu- larnych struktur kortykalnych Tetrahymena.
Biegun przedni jest miejscem od którego rozpoczyna się przebieg struktur południkowych (włókna mikrotubularne i rzędy ciałek podstawowych rzęsek). Pod biegunem przednim zlokalizowany jest aparat gębowy. Na biegunie tylnym, cytoprokt lokuje się koło podgębowego rzędu No. 1, a 90° na prawo znajdują się otworki Wodniczki tętniącej.
waną komórką (a więc pływającym „jednokomórkowym neuronem”), a morfogeneza podziałowa ma charakter metameryczny. Ponadto, znaczniki (markery) powierzchniowe ciała obu obiektów pozwalają na badanie korelacji wzrostu komórki z przemianami w cytoszkielecie. Powstaje pytanie, czy ciałka podstawowe rzęsek Tetrahymena pełnią funkcję w regulacji polaryzacji ciała i czy odbierają sygnały wyznaczające położenie bruzdy cytokinetycznej.
Genom S. pombe jest już poznany, a badania dotyczące sekwencjonowania genomu T. ther- mophila podjęte są na szeroką skalę. W ciągu ostatniego roku grupa badaczy z USA przedstawiła plan częściowego sekwencjonowania genomu T. thermophila (ang. Tetrahymena genome project). Dalsza realizacja projektu umożliwi poszukiwanie poszczególnych genów w genomie, a także wstępną charakterystykę ich produktów białkowych. Opracowanie metod gene
tyki molekularnej dla T. termophila spowodowało, że orzęsek ten stał się doskonałym obiektem badań procesów regulacji transkrypcji i zmian posttranskrypcyjnych białek cytoszkieletu. Istnieje opracowana metoda pozbawiania komórki Tetrahymena prawie wszystkich rzęsek, to znaczy eliminacji z komórki ogromnej ilości spoli- meryzowanej tubuliny. Stworzyło to realną możliwość zbadania, czy ilość spolimeryzowanej i pula niespolimeryzowanej tubuliny w komórce Tetrahymena mogą stanowić sygnał dla regulacji poziomu transkrypcji tubuliny, i jak są regulowane posttranslacyjne zmiany w polimerach tubuliny.
Komórki T. thermophila w określonych warunkach są zdolne do procesu płciowego (koniugacji). Wytworzenie pary indukuje nie tylko mejozę mikronukleusów w obu partnerach, ale także zmianę cytoszkieletu wewnątrzcytopla- zmatycznego w poszczególnych stadiach koniugacji, tzn. podczas powstawania połączeń koniugacyjnych, tworzenia pronukleusów, ich wymiany i krzyżowej fuzji jąder prowadzącej do powstania jąder zygotycznych (synkarionów) oraz różnicowania jąder postzygotycznych w nowe makronukleusy i mikronukleusy (patrz art. Kościuszko i Przyboś w tym numerze KOSMOSU). We wszystkich tych stadiach pojawiają się przejściowo określone struktury cytoszkiele- talne. A więc losy jąder w takich przejściowo wielojądrowych komórkach koniugantów zależą od wykształcenia i funkcjonowania cytoszkieletu indukowanego czasowym połączeniem się komórek dwóch różnych typów płciowych.
Wraz z wiekiem klonu T. thermophila, mierzonym liczbą podziałów komórkowych odbytych po uprzedniej koniugacji, maleje liczba komórek płodnych, to znaczy zdolnych do wytworzenia przedjądrzy (pronukleusów). Komórki te są żywotne i mogą parować z komórkami innych typów płciowych i zawsze w obu komórkach pary zachodzi mejoza mikronukleusa. Jednakże wszystkie jądra postmejotyczne są następnie resorbowane. Dotychczasowe badania wskazują, że utrata zdolności do wytworzenia przedjądrzy i w konsekwencji odtworzenia w trakcie koniugacji nowych makronukleusów i mikronukleusów łączy się z utratą zdolności do wytworzenia specyficznych struktur cyto- szkieletalnych wokół któregokolwiek z jąder postmejotycznych. A więc istnieją przesłanki aby sądzić, że wraz z wiekiem klonalnym zachodzą zmiany, które uniemożliwiają prawidłową organizację cytoszkieletu koniugacyjnego, co powoduje bezpłodność. Zadaniem tego artykułu jest próba scharakteryzowania udziału cytoszkieletu w omówionych tutaj zjawiskach rozwojowych.

Rola cytoszkieletu w rozwoju organizmów 541
BADANIA NAD POLARNOŚCIĄ — ROLA CENTRÓW ORGANIZACJI MIKROTUBUL W PRZEBIEGUMORFOGENEZY PODZIAŁOWEJ
ODMIENNY WZOR PRZESTRZENNY WZROSTU KOMORKI I
TYPU MORFOGENEZY U TETRAHYMENA I S. POMBE
Jak się odbywa morfogeneza podziałowa Schisosaccharomyces pombe? Komórka, która znajduje się w fazie G1 cyklu komórkowego rośnie wyłącznie na starym (przedpodziałowym) biegunie i jest to jednobiegunowa faza wzrostu (Ryc. 2). Wejściu w fazę s DNA cyklu towarzyszy
aktywacja przez pobudzenie receptora typu Rho (ang. Ras homolog) (F u r g e i współaut. 1998). Małe białko Rho zlokalizowane w błonie komórkowej, po przyłączeniu cząsteczki GTP w miejsce GDP, działa jako GTPaza aktywując PAK-ki- nazy, np. Rho-kinazę. Wykazano, że aktywacja tej kinazy jest niezbędna dla podtrzymania dwubiegunowości komórki w mitozie i koordynacji zmian w błonie komórkowej z cyklem komórkowym (V e r d e i współaut. 1998). Białka te
Ryc. 2. Zmiany w położeniu strefy wzrostu w cyklu komórkowym drożdża Schisosaccharomyces pombe (dalsze objaśnienia w tekście).
kontrolowane genetycznie przejście z jedno- biegunowego wzrostu do wzrostu komórki na obu końcach (faza wzrostu dwubiegunowego). Dobudowa błony komórkowej w regionie drugiego bieguna wymaga uprzedniego przemieszczenia do tego bieguna, wzdłuż osi komórki, F-aktyny i innych białek z cytoplazmy. Po ukończeniu replikacji (stadium G2/M), w otoczce jądra komórkowego drożdża tworzą się dwa biegunowe ciałka polarne (SPBs) (M asu d a 1995). Te nowe ciałka biegunowe w otoczce jądra wiążą y-tubulinę i białka GTBPs (ang. gamma-tubulin binding proteins) (S c h ie b e l 2000). Pomiędzy ciałkami biegunowymi tworzy się wewnątrzjądrowe wrzeciono mitotyczne. Dwubiegunowość komórki jest regulowana przez białka motoryczne wrzeciona mitotyczne- go oraz przez stan polimeryzacji mikrotubul (Saw in i N u r s e 1998, F u r g e i współaut. 1998). We wczesnych stadiach mitozy, przynajmniej trzy białka kortykalne: F-aktyna, białko cyto- szkieletalne łącznikowe KELCH, wiążące ciałka polarne (SPBs) z biegunami komórki, i białko RAL 3, gromadzą się symetrycznie na obu starych biegunach komórki. RAL 3 nie jest wprawdzie białkiem cytoszkieletalnym, ale pośredniczącym (ang. scaffold protein), które wiąże wiele białek cytoszkieletu. Te bieguny regulują oś wrzeciona mitotycznego i zapewniają osiowy wzrost komórki. Procesy lokalizacji białek cyto- szkieletalnych, regulujących biegunowy wzrost komórki drożdża, kontrolowane są przez kilka białek, w tym przez produkt genu sspl, który jest kinazą serynowo/treoninową typu PAK-kinazy (M a tsu sa k a i współaut. 1995). Cechą kinaz PAK (ang. pRas activated kinases) jest ich
w czasie anafazy przemieszczają się do regionu tworzącej się bruzdy. Z podanych tutaj danych wynika, że w S. pombe istnieje mechanizm sprzęgający aktywność SPBs z przemianami w cytoszkielecie, który prowadzi do dwubiegunowego podziału komórki.
Nasze wiadomości o mechanizmach podziału wegetatywnego Tetrahymena są znacznie skromniejsze. W przeciwieństwie do większości innych komórek posiada ona względnie stabilny cytoszkielet nadający komórce stały kształt i stałą polaryzację przekazywaną podczas kolejnych podziałów komórkowych. Oś ta jest wyznaczona południkowym rozmieszczeniem podpowierzchniowych struktur cytoszkieletal- nych, które wraz z systemem błony powierzchniowej i podbłonowych cystern stanowią tzw. korteks orzęska. W obrębie struktur południkowych wyróżnia się rzędy ciałek podstawowych rzęsek z towarzyszącymi im włókienkami kotwiczącymi. Ciałko podstawowe, o strukturze przypominającej centriolę, jest strukturą spolaryzowaną; ma swoją część przednią i tylną oraz stronę lewą i prawą (B e is s o n i J e rk a -D z ia d o s z 1999). Tak więc struktury południkowe składają się z podjednostek spolaryzowanych stanowiących domeny korteksu (ang. basal body domain), które można wyznakować szeregiem przeciwciał (W illia m s i współaut. 1990, F r a n k e l 1999). Ciałko podstawowe odbiera również sygnały zewnętrzne regulujące ruch rzęski, a więc jest strukturą spolaryzowaną nie tylko morfologicznie, ale także fizjologicznie (H e n n e s s e y i K u r u v i l la 1999). Ciałka podstawowe zawierają y-tubulinę i powielają się podobnie do centrioli, a ostatnio wykazano, że inaktywacja funkcji

542 J anina Kaczanowska i w spółau torzy
genu y-tubuliny (u innego o rzęska, Parame- ciuni) zatrzymuje ich powielanie w korteksie (Ruiz i współaut. 1999).
Orzęsek ma wyróżnialny morfologicznie biegun przedni i tylny. Pod biegunem przednim Tetrahymena występuje charakterystyczna struktura aparatu gębowego, złożona z zespołów orzęsionych ciałek podstawowych tworzących błonki. Rytmiczny ruch błonek napędza pokarm do zagłębienia, w którym odbywa się zlokalizowana fagocytoza. Aparat gębowy jest asymetryczny i ma różne krawędzie, lewą i prawą oraz przednią i tylną (Kiersno w ska i G oliń - ska 1996). Położenie gęby jest wyznacznikiem zarówno przodu ciała, jak i strony brzusznej orzęska. W okolicy bieguna tylnego występuje wsparty cytoszkieletem cytoprokt, któiy jest zlokalizowanym miejscem defekacji, czyli usuwania na zewnątrz niestrawionych zawartości wodniczek pokarmowych. Także w okolicy bieguna tylnego występują stałe, wzmocnione cytoszkieletem, otworki Wodniczki tętniącej — CVPs (ang. contractile vacuole pores), przez które usuwany jest nadmiar wody i płynne metabolity komórki. Struktury południkowe oraz struktury gęby, cytoproktu i otworki Wodniczki tętniącej są morfologicznymi znacznikami biegunów komórki (ang. cortical landmarks) i polarności korteksu. Struktury te tworzą (Ryc. 1) określony kortykalny wzór przestrzenny cytoszkieletu (ang. cortical pattern) (Na nney 1966, Frankel 1989).
W okresie międzypodzialowym przyrost powierzchni ciała Tetrahymena zachodzi w środkowej strefie komórki (Ryc. 3). Towarzyszy temu polimeryzacja struktur cytoszkieletalnych oraz proliferacja ciałek podstawowych i rzęsek w
rzędach południkowych. Nowe ciałka podstawowe (jedno lub więcej) powstają z przodu istniejącego, orzęsionego ciałka podstawowego i początkowo są nieorzęsione. Następnie dobu- dowywane są rzęski i struktury włókniste kotwiczące ciałka podstawowe w korteksie i w ten sposób powstają nowe domeny kortykalne (Al len 1969, szczegółowo geometrię tego procesu u Tetrahymena omawia Frankel 1999). Pozycja ciałek podstawowych w rzędach południkowych jest pod kontrolą genetyczną (Nanney 1975). Komórka jest spolaryzowana jako całość, bowiem w mutancie T. thermophila (disorder) ciałka podstawowe i rzęski nie tworzą regularnych rzędów, a mimo to komórka zachowuje nadal charakterystyczny kształt i prawidłowe położenie bruzdy podziałowej (Jerka-D zia- d o sz i współaut. 1995).
Komórka orzęska w trakcie podziału wegetatywnego dzieli się poprzecznie względem osi przodo-tylnej, która zostaje zachowana w komórkach potomnych. Oznacza to, że potomny osobnik przedni (zwany proterem) tworzy się z przedniej części ciała osobnika rodzicielskiego i wytwarza nowy biegun tylny, a osobnik tylny (zwany opistorem) wytwarza nowy biegun przedni i tworzy się z tylnej części ciała osobnika rodzicielskiego. Ponieważ bruzda podziałowa Tetrahymena przebiega poprzecznie w stosunku do rzędów rzęsek i rozdziela tworzący się biegun tylny protera i biegun przedni opistora, to podział ten nie jest podziałem symetrycznym (Rye. 4A), lecz metamerycznym, czyli segmen - talnym (Rye. 4B) (Frankel 1989).
W trakcie podziału Tetrahymena zachodzi częściowa reorganizacja cytoszkieletu. W tylnej połowie komórki tworzy się nowy aparat gębowy
- strefa wzrostu korteksu - zróżnicowana strefa bruzdy
Ryc. 3. Zmiany w położeniu strefy wzrostu i tworzenie strefy bruzdy w trakcie morfogene- zy podziałowej T etrah ym en a (dalsze objaśn ienia w tekście).
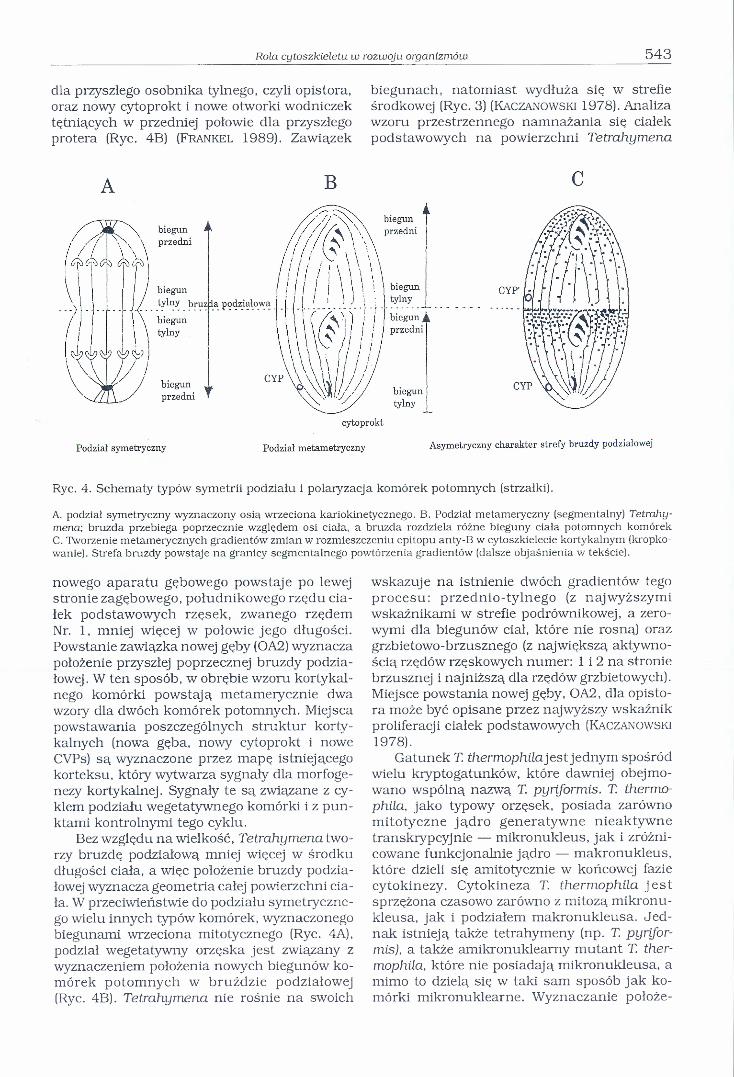
Rola cytoszkieletu w rozwoju organizmów 543
dla przyszłego osobnika tylnego, czyli opistora, oraz nowy cytoprokt i nowe otworki wodniczek tętniących w przedniej połowie dla przyszłego protera (Ryc, 4B) (F r a n k e l 1989). Zawiązek
biegunach, natomiast wydłuża się w strefie środkowej (Ryc. 3) (K a c za n o w sk i 1978). Analiza wzoru przestrzennego namnażania się ciałek podstawowych na powierzchni Tetrahymena
Ryc. 4. Schematy typów symetrii podziału i polaryzacja komórek potomnych (strzałki).
A. podział symetryczny wyznaczony osią wrzeciona kariokinetycznego. B. Podział metameryczny (segmentalny) Tetrahymena; bruzda przebiega poprzecznie względem osi ciała, a bruzda rozdziela różne bieguny ciała potomnych komórek C. Tworzenie metamerycznych gradientów zmian w rozmieszczeniu epitopu anty-B w cytoszkielecie kortykalnym (kropkowanie). Strefa bruzdy powstaje na granicy segmentalnego powtórzenia gradientów (dalsze objaśnienia w tekście).
nowego aparatu gębowego powstaje po lewej stronie zagębowego, południkowego rzędu ciałek podstawowych rzęsek, zwanego rzędem Nr. 1, mniej więcej w połowie jego długości. Powstanie zawiązka nowej gęby (OA2) wyznacza położenie przyszłej poprzecznej bruzdy podziałowej. W ten sposób, w obrębie wzoru kortykal- nego komórki powstają metamerycznie dwa wzory dla dwóch komórek potomnych. Miejsca powstawania poszczególnych struktur korty- kalnych (nowa gęba, nowy cytoprokt i nowe CVPs) są wyznaczone przez mapę istniejącego korteksu, który wytwarza sygnały dla morfoge- nezy kortykalnej. Sygnały te są związane z cyklem podziału wegetatywnego komórki i z punktami kontrolnymi tego cyklu.
Bez względu na wielkość, Tetrahymena tworzy bruzdę podziałową mniej więcej w środku długości ciała, a więc położenie bruzdy podziałowej wyznacza geometria całej powierzchni ciała. W przeciwieństwie do podziału symetrycznego wielu innych typów komórek, wyznaczonego biegunami wrzeciona mitotycznego (Rye. 4A), podział wegetatywny orzęska jest związany z wyznaczeniem położenia nowych biegunów komórek potomnych w bruździe podziałowej (Rye. 4B). Tetrahymena nie rośnie na swoich
wskazuje na istnienie dwóch gradientów tego procesu: przednio-tylnego (z najwyższymi wskaźnikami w strefie podrównikowej, a zerowymi dla biegunów ciał, które nie rosną) oraz grzbietowo-brzusznego (z największą aktywnością rzędów rzęskowych numer: 1 i 2 na stronie brzusznej i najniższą dla rzędów grzbietowych). Miejsce powstania nowej gęby, OA2, dla opistora może być opisane przez najwyższy wskaźnik proliferacji ciałek podstawowych (K a c za n ow sk i
1978).Gatunek T. thermophilajest jednym spośród
wielu kryptogatunków, które dawniej obejmowano wspólną nazwą T. pyriformis. T. thermo- phila, jako typowy orzęsek, posiada zarówno mitotyczne jądro generatywne nieaktywne transkiypcyjnie — mikronukleus, jak i zróżnicowane funkcjonalnie jądro — makronukleus, które dzieli się amitotycznie w końcowej fazie cytokinezy. Cytokineza T. thermophila jest sprzężona czasowo zarówno z mitozą mikronu- kleusa, jak i podziałem makronukleusa. Jednak istnieją także tetrahymeny (np. T. pyriformis), a także amikronuklearny mutant T. thermophila, które nie posiadają mikronukleusa, a mimo to dzielą się w taki sam sposób jak komórki mikronuklearne. Wyznaczanie położę-

544 J anina Kaczanowska i współau torzy
nia bruzdy podziałowej u T. pyriformis, a także w komórkach gatunków mikronuklearnych zachodzi bez udziału struktur wrzeciona mitotycznego. Proces morfogenezy podziałowej w komórce Tetrahymena ma charakter meta- meryczny i jest zdeterminowany wyłącznie przez reorganizację cytoszkieletu kortykalnego.
A więc u orzęsków w trakcie podziału nie tworzy się dwubiegunowa oś ciała komórki pomimo zasadniczego podobieństwa strukturalnego, sposobu powielania oraz budowy centro- somu (ciałek podstawowych rzęsek). Z drugiej strony, w dzielących się komórkach drożdży tworzy się dwubiegunowa oś ciała, pomimo zasadniczo odmiennej struktury i lokalizacji SPB w stosunku do centrosomu. Natomiast we wszystkich tych strukturach mamy pewne cechy wspólne, tzn. występowanie y-tubuliny, powielanie struktury w określonej fazie cyklu i aktywność w organizowaniu polimeryzacji tu- buliny.
Powstaje pytanie, czy i jakie istnieją punkty kontrolne w cyklu podziału orzęska i drożdża, które determinują zależność pomiędzy strukturami o charakterze centrów polimeryzacji mi- krotubul zawierających y-tubulinę a wyznaczeniem i funkcjonowaniem bruzdy cytokinetycznej.
PODOBIEŃSTWA W KONTROLI SPRZĘŻENIA CYKLU
KOMÓRKOWEGO Z FOSFORYLACJA BIAŁEK CYTOSZKIELETU
U S. POMBE I TETRAHYMENA
Aktywacja dwóch servnowo/ treoninowych kinaz białkowych: p34cdc /cyklina B oraz kinazy NIMA (ang. never in mitosis Aspergillus) powoduje jednoczesną fosforylację białek wchodzących w skład wielu struktur komórki drożdży S. pombe. Te kinazy wpływają na profazalną kondensację chromosomów i fosforylują bieguny wrzeciona mitotycznego, kinetochory oraz SPB. Jednocześnie, fosfoiylacje te są wykrywane przeciwciałem MPM-2 w ekstrahowanych cytoszkieletach dzielących się komórek. Następnie, kinaza NIMA, podobnie jak cyklina B, jest degradowana w stadium telofazy (Ye i współaut. 1995).
To samo przeciwciało, MPM-2, wiąże się z wieloma strukturami cytoszkieletu w interfa- zalnej komórce Tetrahymena (K ie r s n o w s k a i G o liń s k a 1996), a fosforylacja poszczególnych struktur cytoszkieletu zmienia się w trakcie cyklu komórkowego (K a c za n o w s k a i współaut. 1999). Co więcej w Tetrahymena wykryto zarówno homolog kinazy p 3 4 cdc2 (G o n d a i współaut. 1999 a, patrz także poniżej), jak i homolog kinazy NIMA (W a n g i współaut. 1998).
W czasie podziału komórki Tetrahymena tworzą się dwa metameryczne gradienty niektó
rych własności korteksu, np. wzrostu rzęsek (F r a n k e l i współaut. 1981). Granicą pomiędzy tymi gradientami jest bruzda (F r a n k e l 1989). W strefie bruzdy zahamowane jest tworzenie nowych ciałek podstawowych, a strefy wzrostu aktywują się w środkowych rejonach powstającego protera i opistora (Fig. 3) (K a c za n o w sk a i współaut. 1993). Znakowanie dzielących się te- trahymen przeciwciałem MPM-2 wykazało, że fosforylacja w południkowych rzędach ciałek podstawowych rzęsek jest najbardziej intensywna w przedniej okolicy komórki oraz bezpośrednio za bruzdą podziałową, a spada stopniowo w dalszych ciałkach podstawowych. Wykazano również różnice w ufosforylowaniu struktur nowej i starej gęby. A więc w czasie podziału Tetrahymena intensywność fosforylacji ciałek podstawowych tworzy wzdłuż południków metameryczne gradienty, a na ich granicy powstaje bruzda podziałowa (K a c za n o w sk a i współaut. 1999).
Podobne dwa gradienty wiązania pewnych epitopów białek cytoszkieletu podbłonowego (W illia m s i współaut. 1987, 1990), rozdzielone strefą bruzdy, powstają w dzielących się Tetrahymena (Rye. 4C) (K a c za n o w sk a i współaut.1999). Ponadto, w mutantach T. thermophila dzielących się na dwa nierówne osobniki potomne wykazano, że przesunięciu bruzdy towarzyszy odpowiednia zmiana w gradientach fosforylacji ciałek podstwowych i epitopów białek podbłonowych (K r z y w ic k a i współaut. 1999).
Te dane wskazują, że zarówno u S. pombe, jak i u Tetrahymena zmiany w intensywności fosforylacji w cytoszkielecie dzielących się komórek, wykrywane przeciwciałem MPM-2, były zawsze wskaźnikiem procesu tworzenia bruzdy podziałowej (LOGARINHO i S u n k e l 1998, B a h le r i współaut. 1998).
WYZNACZANIE POŁOŻENIA I FUNKCJONOWANIA BRUZDY
PODZIAŁOWEJ S. POMBE, POŚREDNIE DANE DOTYCZĄCE
TETRAHYMENA
M a ta i N u r s e (1997) stwierdzili, że położenie bruzdy podziałowej u S. pombe wyznaczane jest przez kortykalne punkty odniesienia, którymi są rosnące bieguny komórki (Ryc. 2). Pierwszym etapem jest aktywacja biegunów wrzeciona mitotycznego, a następnie biegunów komórki. Położenie bruzdy podziałowej w połowie osi ciała u S. pombe kontrolowane jest przez produkty genu Midlp i dwie kinazy seryno- wo/treoninowe: Poml i Pio 1 (tak zwana POLO- kinaza). Produkty fosforylacji cytoszkieletu przez kinazę Plol wykrywane są również przeciwciałem MPM-2 (B A h le r i współaut. 1998). Podobnych białek nie zidentyfikowano jeszcze

Rola cytoszkieletu w rozwoju organizmów 545
u Tetrahymena. Istnieją jednak pośrednie dowody na to, że w trakcie podziału komórki zachodzi aktywacja przynajmniej bieguna przedniego Tetrahymena, skoro gęba OA1 i część apikalna komórki podlega kilkustopniowej reorganizacji. Reorganizacja ta jest związana z cyklem zmian w ufosforylowaniu poszczególnych struktur cytoszkieletu gęby, wykrywanych przeciwciałem MPM-2 (Kaczanowska i współaut. 1999).
Przemieszczanie się w region bruzdy aktyn i aktyno-podobnych białek (np. profiliny, fim- bryny ) oraz białek enzymatycznych, (jak kilku kinaz, np. cdc7 — kinazy i GTPazy, np. Spglp) reguluje powstanie i funkcjonowanie bruzdy podziałowej S. pombe. Zmiany te odbywają się po przejściu metafazowo-anafazowym cyklu, tzn. po uruchomieniu proteolizy i inaktywacji MPF. Wszystkie te przemiany stanowią składowe nowej ścieżki sygnalizacyjnej łączącej cykl jądrowy z przebiegiem cytokinezy (Evangelista i współaut. 1997, Sparks i współaut. 1999, Balasubramanian i współaut. 2000).
Podczas cytokinezy w komórce Tetrahymena, podobnie jak w innych badanych komórkach, tworzy się strukturalny pierścień (Jerka- Dziadosz i współaut. 1995) wyznaczający miejsce podczepienia cytoszkieletu pierścienia kurczliwego (Jerka-Dziadosz 1981). Obecnie zlokalizowano w bruździe dzielącej się Tetrahymena szereg białek pierścienia kurczliwego, np. aktynę (HlRONO i współaut. 1987), profilinę (Edematsu i współaut. 1992) i białko podobne do fimbryny (Watanabe i współaut. 1998). Podobnie w bruździe drożdża zlokalizowano białka zależne od kinazy zależnej od kalmoduliny u drożdża (McCollum współaut. 1995) i podobną kinazę w pierścieniu cytokinetycznym Tetrahymena thermophila (Gonda i współaut. 1999 a, b).
Wszystkie te dane wskazują na podobieństwo molekularne mechanizmów regulujących zarówno przemiany cytoszkieletalne w podziale dwubiegunowym drożdży, jak i w podziale seg- mentalnym orzęsków.
WPŁYW ZMIAN POSTTRANSLACYJNYCH TUBULIN W CYTOSZKIELECIE T. THERMOPHILA NA RÓŻNICOWANIE WŁASNOŚCI STRUKTUR MIKROTUBULARNYCH
s tu k t u r y tu b u lin o w e ja k o źró d ło syg n ału
REGULUJĄCEGO TRANSKRYPCJĘ GENÓW TUBULINOWYCH
Zmiany posttranslacyjne tubuliny
Struktura korteksu Tetrahymena jest bardzo złożona (Allen 1967). Wiele struktur mikro- tubularnych wykazuje różną stabilność w cyklu komórkowym, jak też różną odporność na czynniki depolimeryzujące (Frankel 1999). W komórce T. thermophila wyróżniono co najmniej kilkanaście różnych typów struktur mikrotu- bularnych. Tetrahymena jest zatem znakomitym modelem do badania wpływu zmian post- translacyjnych tubulin na właściwości mikro- tubul oraz zależnej od tubuliny regulacji funkcjonowania genów tubuliny.
Podstawowym budulcem mikrotubul są heterodimery tubulinowe złożone z cząsteczek a- i (3-tubuliny. Występowanie wielu struktur mikrotubularnych o różnorodnych własnościach i stabilności w obrębie jednej cyto plazmy wynika, albo z obecności izoform tubulinowych, albo ich modyfikacji posttranslacyjnych oraz ze współdziałania z białkami MAP, białkami moto- rycznymi z rodziny kinezyn i dynein oraz białkami opiekuńczymi (Luduena 1993, Lee 1993, Skoufias i Scholey 1993, Soares i współaut. 1995, W illiams i Nelsen 1997). W jaki sposób
może być regulowane powstawanie mikrotubul o różnych właściwościach, a w konsekwencji różne funkcje mikrotubul ?
Po pierwsze, jest to związane z występowaniem wielu genów dla a- i/lub (3-tubuliny (Little i Seehaus 1988, Burns 1991), a w rezultacie wielu izotypów tubuliny i heterodimerów tubulinowych. Taka różnorodność powoduje, że dany rodzaj mikrotubul jest budowany tylko z pewnych rodzajów heterodimerów, co może wpływać na późniejsze własności mikrotubul, ich trwałość wiązania białek motorycznych lub białek MAP oraz oddziaływania z innymi białkami itp. Okazuje się, że w niektórych tkankach (np. gonady męskiej Drosophila) ekspresji ulegają tylko niektóre geny tubulinowe i nigdy nie stwierdza się obecności izotypów powstałych na skutek ekspresji pozostałych genów tubuliny (Savage i współaut. 1989, Hoyle i Raff 1990). W ten sposób, w niektórych wyspecjalizowanych tkankach dochodzi do wytworzenia mikrotubul o ściśle określonych właściwościach.
Po drugie, właściwości mikrotubul związane są również ze zmianami posttranslacyjnymi i oddziaływaniem z innymi białkami, np. białkami MAP, kontrolującymi stabilność mikrotubul, białkami opiekuńczymi TCP-1 powodującymi zmiany w strukturze tubulin i różną ich stabilność (Gao i współaut. 1993) oraz czynnikami destabilizacji mikrotubul w trakcie cyklu ko

546 Janina Kaczanowska i współau torzy
mórkowego, zmieniającymi kinetykę polimeryzacji/ depolimeiyzacji heterodimerów tubulino- wych (Soa r e s i współaut. 1995, A n d e r s e n1999). Z kolei, mikro tubule w rzęskach Tetra- hymena są przykładem struktur bardzo stabilnych, które nie ulegają depolimeiyzacji pod wpływem czynników, które depolimeryzują mi- krotubule w komórkach interfazowych i podziałowych.
Komórki Tetrahymena mają tylko jeden gen dla a-tubuliny i dwa geny dla P-tubuliny, kodujące takie samo białko (ostatnie doniesienia wskazują na istnienie trzech genów dla (3-tubu- liny) (G a e r t ig i współaut. 1993, Gu — informacja ustna). Przy tak małej różnorodności heterodimerów powstających z prawie jednolitej puli monomerów tubulinowych, pojedyncza komórka jest w stanie wytworzyć aż około 17 różnych struktur mikrotubularnych. Zatem, dysponująca znacznie mniejszą liczbą izotypów tubuliny Tetrahymena wytwarza mikrotubule o różnych własnościach w liczbie podobnej jak organizmy wielokomórkowe, mające wiele izotypów tubuliny. Ponieważ niewielka liczba genów tubulinowych charakteryzuje organizmy wytwarzające wici i rzęski, być może im mniejsza jest różnorodność heterodimerów, tym bardziej stabilne struktury mikrotubularne mogą być z nich wytworzone w komórce (G a e r t ig i współaut. 1993). Dalsze badania tubulin Tetrahymena wykazały występowanie kilku izomerów a- tubulinowych (G a e r t ig i współaut. 1995). Skoro same izoformy tubulin nie są źródłem zmienności mikrotubul Tetrahymena wydaje się, że w komórce tej główną rolę w różnicowaniu własności poszczególnych izomerów tubuliny mają modyfikacje posttranslacyjne.
Tubuliny w procesie obróbki posttranslacyj- nej mogą ulegać acetylacji, tyrozylacji/detyro- zylacji, poliglicylacji, poliglutamylacji i fosforylacji. Większość miejsc modyfikacji posttransla- cyjnych tubulin znajduje się na C-końcu tego białka. Jak do tej pory żadnej z tych modyfikacji nie udało się przypisać specyficznej funkcji (Hai i Gorovsky — informacja ustna, H ira n o -O h n is i i W a ta n a b e 1989, P e n q u e i współaut. 1991). Wydaje się, że przynajmniej z dwóch powodów Tetrahymena jest odpowiednim obiektem do badań zmian posttranslacyjnych tubulin: (1) małej liczby genów kodujących tubulinę (jeden gen a-tubuliny i dwa, być może trzy geny (3-tu- buliny), (2) łatwości konstruowania mutantów i manipulacji genetycznych przez wprowadzenie do komórki zmutowanego genu i jego podstawianie w miejsce dzikiego. W ten sposób kon- strukt znajduje się na swoim miejscu w genomie i podlega ekspresji takiej, jak gen dziki (G a e r t ig i współaut. 1994).
W ostatnim okresie dokładnie przebadano funkcje jakie spełnia acetylacja lizyny 40 zlokalizowanej na N-końcu a-tubuliny. W tym celu w komórkach Tetrahymena zastąpiono gen dziki konstruktem, w którym kodowana tubulina w miejscu lizyny 40 miała podstawioną argininę (oba aminokwasy zawierają grupę zasadową). Okazało się, że zmodyfikowane białko nie było acetylowane. Tym niemniej komórki tworzące zmutowane białko nie wykazywały żadnych zauważalnych zmian w porównaniu z komórkami kontrolnymi (G a e r t ig i współaut. 1995). Wydaje się więc, że acetylacja lizyny 40 nie ma wpływu na stan mikrotubul. Dlaczego więc aminokwas ten jest konserwowany ewolucyjnie? Nasuwa się kilka odpowiedzi.
1. Być może modyfikacja ta działa synergi- stycznie z innymi. W komórce Tetrahymena wykazano istnienie izoform a-tubuliny i P-tu- buliny związanych ze zmianami posttranslacyj- nymi. Wszystkie izoformy a-tubuliny są acetylowane na lizynie, ale różnią się ładunkiem. Świadczy to o występowaniu jeszcze innych modyfikacji posttranslacyjnych w tubulinie. Być może zmiany we właściwościach mikrotubul spowodowane przez te modyfikacje wraz z acetylacją, doprowadzają do zmian właściwości a-tubuliny, a następnie powodują zmianę w funkcji heterodimeru tubulinowego i całej mi- krotubuli.
2. Być może jednak „sprawność” mikrotubul i całej komórki pod wpływem acetylacji lizyny 40 zmienia się w sposób na razie nieuchwytny do wykazania, ale na tyle istotny, że jest konserwowany ewolucyjnie.
3. Możliwe również, że a-tubulina jest drugorzędnym (przypadkowym) substratem dla modyfikującej ją acetylazy, której główny sub- strat ma miejsce acetylacji o podobnej budowie jak (3-tubulina. Zatem tubulina byłaby acetylo- wana „przy okazji” acetylacji innego białka, którego modyfikacja mogłaby mieć dużo większe znaczenie. Można by zatem mówić o konserwacji ewolucyjnej enzymu modyfikującego tubulinę, a nie samej modyfikacji (G a e r t ig i współaut. 1995).
Udało się również uzyskać mutanty Tetrahymena, w których na C-końcu a- lub p-tubuliny, usunięto miejsca poliglicylacji kwasu glutaminowego. Badania wykazały, że brak poliglicylacji a-tubuliny nie powoduje zmian ruchliwości i przeżywalności komórek. Natomiast w przypadku P-tubuliny efekt mutacji zależy od liczby zmutowanych miejsc. W P-tubulinie na C-końcu występują cztery miejsca poliglicylacji. Zastąpienie trzech miejsc poliglicylacji powoduje spowolnienie wzrostu, zmniejszoną ruchliwość, a także zatrzymanie podziałów. Mutanty bardzo

Rola cytoszkieletu w rozwoju organizmów 547
często wykazują nienormalną wielkość, a także zwiększoną liczbę jąder. Zdarza się również, że komórki nie przechodzą cytokinezy lub też bruzda podziałowa jest położona niesymetrycznie. Takie fenotypy mogą być wynikiem nieprawidłowych oddziaływań mikrotubul z białkami motorycznymi i strukturalnymi związanymi z tubulinami. Natomiast zastąpienie czterech miejsc poliglicylacji (3-tubuliny jest letalne. Ekspresję nieprawidłowych fenotypów komórek zawierających zmutowany gen można jednak odwrócić. Do komórek zawierających zmutowany gen (3-tubuliny wszczepiono gen a-tubuliny, w którym C-koniec zastąpiono C-końcem pochodzącym z dzikiego genu (3-tubuliny. Takie komórki nie wyrażały nieprawidłowego fenotypu (Xia i współaut. 2000, Gaertig — informacja ustna).
Udział tubuliny w regulacji transkrypcji genów tubulin
Jedną z ciekawych własności tubuliny jest jej udział w regulacji ekspresji genów, z czego najlepiej poznanym przykładem jest regulacja ekspresji genów a- i (3-tubuliny. Badania z inhibitorami tubulin na komórkach ssaków wykazały, że przy zastosowaniu środków depolime- ryzujących mikro tub ule obserwowano spadek ilości mRNA tubulin, a przy zastosowaniu środków stabilizujących mikrotubule, wzrost jego ilości (Cle v e lan d i współaut. 1981, 1983). W następnych badaniach stwierdzono, że pod wpływem zmian w poziomie monomeru tubuli- nowego następuje zmiana ilości mRNA tubulin. Wzrost ilości monomerów tubuliny pobudza specyficzną degradację mRNA (3-tubuliny (Ba - churski i współaut. 1994, C le ve land 1989).
Przyżyciowe odrzęsianie Tetrahymena przy pomocy czynników chemicznych (Thompson i współaut. 1974) powoduje odpadnięcie wszystkich rzęsek, z zachowaniem ich ciałek podstawowych. Zachodząca następnie synchroniczna i masowa regeneracja rzęsek pozwala na badanie molekularnych mechanizmów tego procesu.
Wydaje się, że w komórkach Tetrahymena regulacja transkrypcji genów tubulinowych przebiega w nieco inny sposób niż w komórkach ssaków. Depolimeryzacja mikrotubul przy pomocy oiyzaliny lub kolchicyny wywołuje wzrost ilości mRNA dla a-tubuliny, a co za tym idzie wzrost ilości monomeru a-tubulinowego, a więc nie występuje charakterystyczna dla komórek ssaków degradacja mRNA tubulinowego. Co ciekawsze, to samo zjawisko (czyli wzrost ilości monomerów tubuliny) występowało pod wpływem czynników stabilizujących mikrotubule, między innymi taksolu (St a r g e ll i współaut.
1992). Wydaje się więc, że Tetrahymena generalnie reaguje na zakłócenia w cy to szkielecie poprzez wzrost ilości mRNA dla a-tubuliny.
Dalsze badania wykazały, że Tetrahymena posiada dwa niezależnie działające mechanizmy reagowania na zniszczenie mikrotubul (Gu i współaut. 1995). Pierwszy z nich jest uruchamiany, gdy komórka Tetrahymena zostaje od- rzęsiona, a więc zniszczeniu uległy mikrotubule budujące rzęski. W odpowiedzi na odrzęsienie, w komórce zostaje uruchomiona transkrypcja wszystkich genów kodujących tubulinę (gen a- tubulinowy i dwa geny (3-tubuliny). Natomiast w przypadku, gdy nastąpi tylko depolimeiyza- cja mikrotubul cytoplazmatycznych bez odrzę- siania, pobudzana jest ekspresja genu a-tubu- liny i tylko jednego z dwóch genów (3-tubuliny.
W jaki sposób komórka „rozróżnia”, które ze struktur mikrotubularnych uległy zniszczeniu? Po pierwsze, u orzęsków, jak to zbadano w Paramecium caudatum, kanały wapniowe zależne od napięcia występują wyłącznie na rzęskach (M a c h e m e r i O g u r a 1979, patrz art. Fa b - c za k i Fa b c z a k w tym numerze KOSMOSU). Zniszczenie rzęsek może być w jakiś sposób skorelowane z otwarciem tych kanałów, a to mogłoby powodować specyficzny sygnał uruchamiający translację genów tubulinowych. Kanały tego samego typu mogłyby występować powszechnie w rzęskach orzęsków, w tym również u Tetrahymena. Następnie, rzęski zawierają unikalną kinezynę (B er n ste in i współaut. 1994) i dyneinę (Pa s c h a l i współaut. 1992), a w rzęskach Tetrahymena wykazano także istnienie unikalnej kinazy tubulinowej Ca2+ /kalmo- dulino-zależnej (H ir a n o -O h nish i i W ata n a b e 1989) i to decydowałoby o specyfice sygnału transkrypcji genów tubulinowych.
Trudniej jest określić jaki mechanizm mógłby uruchamiać transkrypcję tubulin po zniszczeniu mikrotubul cytoplazmatycznych. Pewne przypuszczenia opierają się na wynikach wcześniejszych doświadczeń nad wpływem czynników depolimeryzujących i taksolu na poziom mRNA tubulin (Sta r g e ll i współaut. 1992). Prawdopodobnie w obu przypadkach, zarówno w przypadku depolimeryzacji mikrotubul pod wpływem oryzaliny, jak i w przypadku ich stabilizacji taksolem, dochodzi do spadku ilości monomerów tubulin: w przypadku depolimeryzacji monomery są wiązane przez czynnik depolimeryzujący, natomiast w przypadku stabilizacji taksolem monomery pozostają związane w polimery mikrotubul.
Pomimo pewnych wątpliwości interpretacyjnych, w Tetrahymena wykazano po raz pierwszy, że cytoszkielet jest składnikiem ścieżki

548 Janina Kaczanowska ł współau torzy
transdukcji sygnału w regulacji transkrypcji genu tubuliny i może uczestniczyć w regulacji funkcji genów. Cytoszkielet nie jest więc jedynie
„wykonawcą” odebranych sygnałów, ale również ich źródłem i ścieżką przekazu (Gu — informacja ustna).
ROLA CYTOSZKIELETU W KONIUGACJI
PRZEBIEG KONIUGACJI
Warunkiem koniecznym do wejścia komórek w koniugację (Ryc. 5A) jest ich okresowe głodzenie (inicjacja) oraz wzajemne oddziaływanie na siebie komórek różnych typów płciowych (kostymulacja), które następnie łączą się przednimi okolicami ciała (przedgębowymi) (Bruns i Brussard 1974). Połączenie koniugacyjne (ang. conjugation junction), które trwa około 10 godzin, pobudza proces mejozy (Ryc. 5A, b). Jedno z czterech jąder postmejotycznych dzieli się mitotycznie (mitoza pregamiczna), zaś trzy pozostałe są przesuwane ku tyłowi komórki, gdzie ulegają degradacji (Ryc. 5A, c-e). W wyniku podziału mitotycznego w każdym z koniu- gantów powstają dwa przedjądrza: stacjonarne i migracyjne (Rye. 5A, f). Po wzajemnej wymianie przedjądrzy migracyjnych (Ryc. 5 A, g-h), dochodzi do ich fuzji z przedjądrzami stacjonarnymi (Ryc. 5 A, i). Powstałe w ten sposób jądro zygotyczne, zwane także synkarionem, przechodzi dwa podziały postzygotyczne (5 A, j-m), po czym jądra potomne podlegają różnicowaniu: dwa z nich, zlokalizowane w przedniej części komórki, stają się zawiązkami nowych ma- kronukleusów, zaś dwa jądra położone w tyle komórki, pozostają mikronukleusami (Ryc. 5A, n-o). W tym czasie stary, przedkoniugacyjny makronukleus przesuwa się ku tyłowi komórki i ulega degradacji (Ryc. 5A, o-p.). W trakcie koniugacji orzęska zachodzą więc wielokrotne podziały mikronukleusa (mejoza, mitoza pregamiczna) oraz postzygotyczne podziały synkario- nu bez cytokinezy. W ten sposób, przejściowo, powstają wielojądrowe komórki. O losach poszczególnych jąder, to znaczy o kolejnych podziałach, różnicowaniu lub resorpcji decydują: położenie jąder w cytoplazmie, gotowość jąder do odpowiedzi na określone sygnały docierające do nich z cytoplazmy w poszczególnych etapach koniugacji oraz specyficzne stuktuiy cytoszkie- letalne. Tetrahymena thermophita jest więc komórką, w której można badać udział cytoszkieletu w kontroli przebiegu koniugacji.
Z badań nad innymi typami komórek (oocyt Drosophila) wiadomo, że wewnątrzcytopla- zmatyczne mikrotubule pełnią dwojaką rolę: są zaangażowane w przemieszczanie się jądra oo-
cytu w cytoplazmie oraz w transport ważnych determinant rozwojowych (mRNA, białek).
Istnienie kompartmentalizacji cytoplazmy w koniugujących tetrahymenach sugerują doświadczenia N an n ey ’a (1953): wymuszone silnym wirowaniem przemieszczenie mikronu- kleusów w koniugantach powodowało zmiany w liczbie tworzących się mikronukleusów i zawiązków makro nuklearny eh.
Zależności między stanem cytoplazmy a losami jąder obserwowano również u Paramecium (M ikam i 1980, G ran dch am p i B e is s o n 1981, Y a n a g i 1987). W komórce wegetatywnej mikro- nukleus nigdy nie przekształca się w zawiązek makronuklearny. Dopiero proces płciowy umożliwia podziały jąder, fuzję przedjądrzy i różnicowanie zawiązków makronuklearny eh. Przeniesienie mikronukleusa z komórki wegetatywnej P. caudatum do koniuganta w okresie II podziału postzygotycznego lub tuż po tym podziale, może zmusić przeszczepiony, „wegetatywny” mikronukleus do różnicowania się w zawiązek makronuklearny z „ominięciem” etapu mejozy i fuzji przedjądrzy (M ikam i i N g 1983). Zatem w przypadku Paramecium, ani fuzja przedjądrzy, ani nawet podziały mejotyczne nie są warunkami niezbędnymi dla procesu różnicowania zawiązków makronuklearnych, konieczny jest natomiast odpowiedni „stan” cytoplazmy i cytoszkieletu.
ZMIANY POWIERZCHNIOWE I CYTOSZKIELETALNE W
REGIONIE POŁĄCZENIA KONIUGACYJNEGO
W okresie kostymulacji dochodzi do zmiany morfologii przedniego (powyżej gęby) bieguna komórki (ang. tip transformation) (W o l f e i G r im es 1979). Staje się on gładki, pozbawiony rzęsek, ciałek podstawowych, mikrotubul korty kalnych i alweoli ( E l l i o t t i T r e m o r 1958, SUGANUMA i współaut. 1984, WOLFE 1982). Błonę komórkową w tym rejonie podściela rozbudowana warstwa gęsto upakowanego materiału fibrylarnego (W o l f e 1985). W wierzchołkach kostymulowanych komórek, pod tą elektronowo gęstą warstwą, występują filamenty zbudowane z białka o masie 64 kDa, tak zwane filamenty fenestrynowe (N e ls e n i współaut. 1994).
Opisanym wyżej lokalnym zmianom korty- kalnym towarzyszy gromadzenie się w szczyto-

Rola cytoszkieletu w rozwoju organizmów 549
wej części komórki transbłonowego białka (ang. transmembrane protein) — glikoproteiny wią
żącej się z konkanawaliną A (Wolfe i współaut. 1986). Konkanawalina A sprzężoną z FITC, zna-
Ryc. 5. Przemiany jądrowe i cytoszkieletalne w trakcie koniugacji T. thermophila.
A. Podczas pierwszego podziału mejotycznego mikrotubule cytoplazmatyczne tworzące gęstą siatkę częściowo zanikają (5A, a, b). Pod koniec mejozy nowe mikrotubule polimeryzują w regionie połączenia koniugacyjnego i wokół dwóch (5A, c) spośród czterech jąder postmejotycznych (tzw. mikrotubule perinuklearne). Mikrotubule perinuklearne utrzymują się wokół jądra, które „podczepia” się do połączenia koniugacyjnego i w ten sposób jest rekrutowane do trzeciego podziału prezygotycznego (5A, d). W podczepieniu tym biorą udział mikrotubule polimeryzujące z regionu połączenia koniugacyjnego i mikrotubule perinuklearne. Nieliczne mikrotubule powstałe wokół otoczki drugiego jądra, szybko zanikają. Wokół pozostałych dwóch jąder postmejotycznych nie obserwuje się mikrotubul perinuklearnych (Gaertig i F leury 1992, T akagi i współpaut. 1991, K iersnowska dane niepublikowane). W wyniku mitozy pregamicznej powstają dwa przedjądrza (stacjonarne i migracyjne). Oba przedjądrza połączone są z sobą i z regionem połączenia koniugacyjnego pasmami mikrotubul (5A, e, f). Podczas wymiany przedjądrzy, mikrotubule otaczające przedjądrza migracyjne oraz mikrotubule odchodzące od połączenia koniugacyjnego tworzą tzw. koszyczek mikrotubularny (5A, g, f). Łączące się przedjądrza otoczone są pasmami mikrotubul (5A, i; tzw. channel-like structure — wg Gaertig a i Fleury 1992) zanikającymi przed pierwszym podziałem postzygotycznym (5A, j). W stadiach postzygo tycznych jądra nie są otoczone mikrotubulami perinuklearnymi (5A, j-p). Mikrotubule cytoplazmatyczne (tworzące siatkę wewnętrzną) stopniowo odtwarzane są w okresie różnicowania zawiązków makronu- klearnych (5A , o, p). Oznaczenia: Mi — mikronukleus, Ma — makronukleus, r — resorbowane jądra postmejotyczne, zMa — zawiązki makronuklearne, sMa — skondensowany przedkoniugacyjny makronukleus: na schemacie w jednym koniugancie (z wyjątkiem 5 A, g, h) przedstawiono wcześniejsze i późniejsze stadium tego samego etapu przemian jądrowych, w rzeczywistości procesy jądrowe zachodzą synchronicznie. B. Rozmieszczenie fenestryny w koniugantach, objaśnienia w tekście.

550 Janina Kaczanowska i współau torzy
kuje nie tylko przedni biegun kostymulowa- nych komórek, ale również, aż do końca parowania, połączenie koniugacyjne.
Kostymulowane komórki łączą się zmienionymi biegunami i dzięki częściowej fuzji błon tworzą pary. W strefie połączenia koniugacyj- nego powstają bardzo liczne pory o średnicy około 0,2 pm ( E l l i o t t i T r e m o r 1958, W o l f e 1982) łączące cytoplazmę obu koniugantów i umożliwiające wymianę makromolekuł (Mc Don a ld 1966). W okresie wymiany przedjądrzy organizacja i wielkość porów ulegają zmianie, umożliwiając migrację przedjądrzy (O r ia s
1986).Znakowanie przeciwciałem antyfenestryno-
wym połączenia koniugacyjnego wykazuje, że filamenty fenestiynowe obrzeżają pory. To znakowanie utrzymuje się przez cały okres parowania komórek, przy czym wzmożona intensywność znakowania tego regionu jest obserwowana po III podziale prezygotycznym i podczas wymiany przedjądrzy (N e ls e n i współaut. 1994).
CYTOSZKIELET MIKROTUBULARNY
Podczas koniugacji Tetrahymena organizacja somatycznych struktur kortykalnych nie ulega zmianie, natomiast w aparacie gębowym dochodzi do zaniku wiązki mikrotubularnej włókien gębowych (ang. deep fibers, DF) oraz resorpcji kilkunastu ciałek podstawowych membranelli gębowych. Nieczynna, wypłycona, z niefunkcjonalnymi membranellami gęba nie ulega dalszym zmianom, aż do czasu ukończenia przemian jądrowych. Dopiero w ekskoniu- gancie dochodzi do resorpcji „starych”, zamrożonych dotychczas struktur gębowych i utworzenia nowego aparatu gębowego na drodze tak zwanej wymiany gęby (ang. oral replacement), po czym komórki mogą podejmować fagocytozę (K ie r s n o w s k a i współaut. 1993).
W wegetatywnych komórkach Tetrahymena, oprócz mikrotubul kortykalnych uporządkowanych w określone struktury, występują również mikrotubule tworzące gęstą sieć we- wnątrzcytoplazmatyczną oraz mikrotubule we- wnątrzjądrowe (w obu jądrach), uczestniczące w ich podziale (La F ou n ta in i D a v id s o n 1980). W koniugujących tetrahymenach obserwuje się zmiany w cytoszkielecie wewnątrzcytopla- zmatycznym. Dotychczasowa, rozbudowana sieć mikrotubul stopniowo zanika, tworzą się natomiast specyficzne, niezależne od wrzecion mitotycznych, przejściowe struktuiy związane tylko z niektórymi mikronukleusami w wieloją- drowym koniugancie. I tak, wokół jednego z czterech jąder postmejotycznych pojawiają się
mikrotubule perinuklearne, jednocześnie w rejonie połączenia koniugacyjnego polimeryzują mikrotubule rozrastające się w głąb cytopla- zmy. Biorą one udział w podczepieniu do połączenia koniugacyjnego jednego z jąder postmejotycznych, leżącego w pobliżu tego połączenia i otoczonego mikrotubulami perinuklearny- mi. Tylko to jądro dzieli się mitotycznie (trzeci podział prezy go tyczny). Jedno z jąder potomnych, przylegające ściśle do połączenia koniugacyjnego, jest pronukleusem migracyjnym, drugie, położone poniżej, jest pronukleusem stacjonarnym. Oba pronukleusy są przez cały czas otoczone materiałem mikrotubularnym, który nie tylko podczepia oba jądra do połączenia koniugacyjnego, ale również łączy je ze sobą (nie jest to pozostałość wrzeciona mitotyczne- go). Szczególnie silnie rozbudowana otoczka mi- krotubularna obejmuje pronukleusy migracyjne tuż przed przejściem do komórki partnera i w samym momencie transferu tego jądra. Mi- krotubularna „tuba” otacza także łączące się pronukleusy i następnie jądro zygotyczne (G a e r t ig i FLEURY 1992). Zanikanie wewnętrznej sieci oraz pojawianie się nowych struktur mikrotubularnych jest przedstawione na Rye. 5A.
Podanie inhibitorów antytubulinowych (no- kodazol, winblastyna) w czasie koniugacji Tetrahymena nie tylko wstrzymuje podziały jąder na skutek depolimeiyzacji wrzecion podziałowych, lecz również uniemożliwia fuzję przedjądrzy (H a m ilto n i współaut. 1988, K a cza n o w sk i i współaut. 1991). Dzięki doświadczeniom z nokodazolem udało się również wykazać, że już przedjądrza mogą różnicować się w zawiązki makronuklearne oraz wskazać na zależność między różnicowaniem zawiązków makronu- klearnych a położeniem jąder postzygo tycznych w cytoplazmie komórki (K a c za n ow sk i i współaut. 1991).
FILAMENTOWE ELEMENTY CYTOSZKIELETU
W skład cytoszkieletu otaczającego przedjądrza oprócz mikrotubul wchodzą również dwa typy filamentów zbudowane odpowiednio z białka 64 kDa, tworzącego filamenty fenestrynowe (N e ls e n i współaut. 1994) oraz białka 49 kDa (T akag i i współaut. 1991). Po III podziale prezygotycznym (powstanie przedjądrzy) obserwuje się wzmożoną polimeryzację filamentów fene- strynowych w rejonie połączenia koniugacyj- nego oraz wokół przedjądrzy migracyjnych i stacjonarnych (Ryc. 5B, a, b). Powstałe filamenty fenestrynowe delikatnymi wiązkami łączą ze sobą przedjądrza, a mocniejszymi podczepiają jądra migracyjne bezpośrednio do połączenia

Rola cytoszkieletu w rozwoju organizmów 551
koniugacyjnego. W trakcie wymiany przedjądrzy pojawia się bardzo silne zagęszczenie fila- mentów fenestrynowych wokół jądra przechodzącego przez połączenie koniugacyjne, tworzących tzw. koszyczek (Rye. 5B, c, d). Po wymianie przedjądrzy, związane z nimi struktury fene- strynowe utrzymują się tylko przez krótki czas. W dalszych przemianach jądrowych nie obserwowano udziału filamentów fenestrynowych (Rye. 5B, e, f). A więc przeciwciało skierowane przeciwko fenestrynie rozpoznaje tylko jądra funkcjonujące jako przedjądrza (C o le i S o e l t e r 1997).
Białko o masie molekularnej około 49 kDa ma podwójną naturę - występuje w mitochon- driach w postaci monomeru — enzymatycznego białka o właściwościach syntazy cytrynianowej (K o jim a i współaut. 1997) oraz w cytoplazmie — w formie spolimeryzowanej, tworząc 14 nm fi- lamenty nie wykazujące aktywności syntazy cytrynianowej. Obie formy, to jest enzymatyczna i filamentowa, są kodowane przez ten sam gen (Num ata i współaut. 1991, N u m ata 1996), a różnice we właściwościach obu form (nieznacznie wyższa masa molekularna i dodatkowy punkt izoelektryczny dla formy filamentowej) są prawdopodobnie związane z modyfikacjami post- translacyjnymi. Warto jednocześnie zaznaczyć, że oczyszczone białko 49 kDa, w określonych warunkach in vitro, może przechodzić z formy spolimeryzowanej w formę monomeryczną (enzym) i odwrotnie.
Filamenty 14 nm biorą udział w morfogene- zie aparatu gębowego (N u m ata i współaut. 1983) oraz w koniugacji (N u m ata i współaut. 1985). W czasie procesu płciowego filamenty te pojawiają się pod koniec drugiego podziału mejotycznego w rejonie połączenia koniugacyjnego. Polimeryzują one z tego regionu i sięgają do około 1/3 w głąb komórki (T ak ag i i współaut. 1991). W czasie III podziału prezygotycznego 14 nm filamenty kolokalizują się z mikrotubulami otaczającymi oba przedjądrza oraz pronukleusy migracyjne przechodzące przez połączenie koniugacyjne. Znakowanie przeciwciałem skierowanym przeciwko 14 nm filamentom zanika po fuzji przedjądrzy.
PRZERWANIE POŁĄCZENIA MIĘDZY KOMÓRKAMI PARY NIE
WSTRZYMUJE PRZEMIAN W CYTOSZKIELECIE
Wczesne koniuganty (tj. około 2,5 godz.) można rozdzielić mechanicznie bez uszkodzenia komórek (K ie r s n o w s k a i współaut. 2000). Okazuje się, że jeśli rozdzielano koniuganty, w których mikronukleus został już pobudzony do podziału mejotycznego (stadium I i II profazy mejotycznej I), to były one zdolne utworzyć pro
nukleusy, które łączyły się ze sobą (cytogamia). Z jądra zygotycznego powstawały nowe makro- nukleusy i mikronukleusy. Mikrotubule i włókna fenestrynowe tworzyły struktury charakterystyczne dla poszczególnych stadiów koniugacji. A więc, kontakt z drugą komórką podczas procesu płciowego jest niezbędny do zainicjowania koniugacji, ale nie jest konieczny do jej kontynuowania. Świadczy to o tym, że raz pobudzone komórki (pobudzenie ścieżki/ścieżek sygnalizacji wewnątrzkomórkowej) mogą realizować cały, złożony program koniugacji oraz, że pobudzone komórki mają potencjalną zdolność do przejścia pełnego programu koniugacji, a obserwowane zmiany w tym programie wynikają z hamowania już pobudzonych szlaków rozwojowych.
WPŁYW INHIBITORA KINAZ BIAŁKOWYCH, 6-DMAP,
NA PRZEBIEG KONIUGACJI
W obecności inhibitora pewnej klasy kinaz serynowo/treoninowych, 6-DMAP, mejoza i mitoza pregamiczna są opóźnione i często przebiegają asynchronicznie w obu komórkach pary. W niektórych koniugantach zachodzi nawet tylko jeden podział mejotyczny. Jednakże wokół żadnego z jąder (bez względu na ich liczbę) nie występują mikrotubule pericentriolarne. Natomiast polimeryzacja mikrotubul w regionie połączenia koniugacyjnego przebiega tak jak w koniugantach kontrolnych. Mikrotubule te rozrastają się w głąb cytoplazmy i niektóre-wiązki mogą dochodzić (a nawet otaczać) do jednego lub dwóch jąder (mogą to być również produkty I-ego podziału mejotycznego) położonych w pobliżu połączenia koniugacyjnego (Kiersnowska i Włoga — obserwacje nie publikowane). Jądra te mogą znaleźć się w połączeniu koniugacyj- nym, ale nie dochodzi do przejścia ich do partnera, ani też do fuzji jąder i rozwoju postzygo- tycznego. Wyniki te świadczą o tym, że cytosz- kielet perinuklearnyjest niezbędny do utworzenia funkcjonalnych pronukleusów, przemieszczenia się ich do komórki partnera, fuzji oraz do różnicowania się zawiązków makronuklear- nych.
Podobne skutki działania 6-DMAP na cyto- szkielet komórki obserwowano w oocytach jeżowców (S t - P ie r r e i D u fr e s n e 1990, D u fr e s n e i współaut. 1991, S t - P ie r r e i współaut. 1994). Przeniesienie zapłodnionych oocytów do środowiska z 6-DMAP powoduje zatrzymanie procesów, w które zaangażowane są mikrotubule: migrację przedjądrzy, lokalizowanie przedjądrza i wreszcie fuzję przedjądrzy. W zapłodnionym jaju poddanym działaniu 6-DMAP, nie obserwowano mikrotubul astralnych plemnika,

552 J anina Kaczanowska i współau torzy
chociaż polimeryzacja mikrotubul kortykal- nych nie była zakłócona. 6-DMAP podany w okresie metafazy powoduje rozpad wrzeciona podziałowego (S t - P ie r r e i D u fr e s n e 1990). Ponadto, w komórkach tych wykryto, że filament pośredni (p 117) jest silnie ufosforylowany w czasie, gdy zachodzi fuzja przedjądrzy, po czym poziom jego fosforylacji znacznie maleje. Pod wpływem 6-DMAP obserwuje się silniejszą
defosforylację filamentu p l l 7 (S t - P ie r r e i współaut. 1994).
Przedstawione dane sugerują, że warunkiem przejścia przez punkt kontrolny początku rozwoju postzygotycznego w koniugantach Tetrahymena jest wytworzenie otoczki cytoszkieletalnej wokół rekrutowanego pregamicznego jądra, co jest skorelowane w tym stadium z aktywacją nieznanej, kluczowej fosforylacji białek.
STARZENIE SIĘ KLONU — W ZRO ST LICZBY KOM OREK BEZPŁODNYCH
Bezpośrednio po przejściu komórek płodnych przez koniugację, ekskoniuganty Tetrahymena nie są dojrzałe do ponownego parowania i dopiero po przejściu 40-60 podziałów od ostatniej koniugacji stają się dojrzałe płciowo. W warunkach naturalnych mechanizm ten wyklucza parowanie komórek siostrzanych, które odpływają od siebie; zapewnia to odpowiedni poziom polimorfizmu genetycznego. Z drugiej strony, komórki, które przeszły dużą liczbę podziałów (200-300) pozostając cały czas zdolnymi do parowania, na ogół posiadają silnie uszkodzony mikronukleus. Stopniowo, wraz z upływem czasu mierzonego liczbą podziałów, wzrasta prawdopodobieństwo pojawienia się w mikronukleusie mutacji i takich uszkodzeń genomu, które powodują aneuploidię mikronu- kleusów. I rzeczywiście, w staiych klonach obserwowane są komórki z bardzo małymi mikro- nukleusami, a nawet komórki całkowicie ami- kronuklearne. Jednakże jedynie komórki ami- kronuklearne są nieżywotne (z wyjątkiem szczepu BI). Przyczyny tego stanu rzeczy są obecnie jeszcze nieznane, ponieważ nie wykryto nawet śladowej ekspresji genów mikronuklear- nych.
Proces koniugacji jest nieuchronnie „godziną prawdy” dla komórek. Wówczas mogą ujawnić się wszystkie defekty mikronukleusa. Jeżeli mimo uszkodzenia genomu mikronukleus jest zdolny utworzyć funkcjonalne pronukleusy, to nawet gdy tylko jeden partner posiada uszkodzony mikronukleus, makronukleusy obu eks- koniugantów, powstałe z zygotycznych jąder, są defektywne. W koniugacji „staiych” komórek obserwuje się przypadki, gdy oba ekskoniuganty danej paiy są nieżywotne. A więc, w mikronukleusie mogą gromadzić się takie defekty jak mutacje letalne o charakterze dominującym, które nie zaburzają przebiegu koniugacji komórek, ale ujawniają się w nowych makronukleu- sach. Stanowi to endogenne zagrożenie dla populacji orzęsków powodując nieuchronne straty w liczebności populacji.
Można więc oczekiwać, że w warunkach naturalnych dobór faworyzuje częste koniugacje. Tak jest w istocie, co wykazano na drodze masowych izolacji nowych szczepów T. thermophila ze śródleśnych zbiorników wodnych. Były one identyfikowane jako T. thermophila przy pomocy przeciwciał skierowanych przeciwko specyficznym antygenom powierzchniowym tego gatunku. Okazało się, że wśród tych szczepów zawsze (bez względu na porę roku) znajdowano linie komórek niedojrzałych do koniugacji, a więc takich, które stosunkowo niedawno przeszły proces płciowy. Częstym koniugacjom sprzyja występowanie u T. thermophila aż siedmiu różnych typów płciowych, które w warunkach naturalnych były reprezentowane w prawie równych proporcjach (D o e r d e r i współprac. 1995, 1996).
Innym przystosowaniem do wzrastającej z wiekiem klonu kumulacji uszkodzeń genomu Tetrahymena jest utrata zdolności do tworzenia przedjądrzy z aneuploidalnych mikronukleu- sów. Dlaczego jednak jest to proces przystosowawczy umożliwiający przeżycie? Otóż, bez względu na liczbę podziałów wegetatywnych komórki komplementarnych typów płciowych pozostają zdolne do parowania, co indukuje proces mej ozy. Jednakże w przypadku aneuploidalnych mikronukleusów, wszystkie jądra postmejotyczne są resorbowane. Koniugant z aneuploidalnym mikronukleusem nie tworzy pronukleusów, jest więc bezpłodny, ale wciąż zachowuje zdolność przyjęcia pronukleusa od swojego partnera. Jeśli komórka z aneuploidalnym jądrem koniuguje z komórką posiadającą nieuszkodzony mikronukleus (tak zwana koniugacja poronna) (O r ia s 1986), bezpłodna komórka pary otrzymuje pronukleus migracyjny od swojego zdrowego partnera (Ryc. 6A). Tylko otrzymanie pronukleusa od partnera umożliwia odtworzenie żywotnej linii, ponieważ amikronu- klearne komórki T. thermophila giną. Płodny partner staje się w ten sposób jednostronnym dawcą mikronukleusa, a komórka niepłodna

Rola cytoszkieletu w rozwoju organizmów 553
Ryc. 6. Schematy koniugacji poronnej: a. dwóch szczepów niepłodnych i b. koniugacja osobnika płodnego z osobnikiem bezpłodnym (dalsze objaśnienia w tekście).
jego biernym biorcą. Proces ten jest więc naturalną „transplantacją” pronukleusa do komórki pozbawionej własnego mikronukleusa. Po tym jednostronnym transferze jądra każda z komórek posiada po jednym haploidalnym jądrze, które początkowo nie dzieli się i nie resorbuje. Przedkoniugacyjne makronukłeusy również nie resorbują się. Nie dzielące się mikronukleusy w każdym partnerze podlegają jednak diploidyza- cji. Oba ekskoniuganty koniugacji poronnej stają się heterokarionami: komórka bezpłodna ma teraz transplantowany diploidalny homo- zygotyezny mikronukleus partnera i własny makronukleus sprzed koniugacji, a płodny partner ma homozygotyczny mikronukleus powstały z diploidyzacji własnego pronukleusa oraz własny makronukleus sprzed koniugacji (Ryc. 6A). Każdy z ekskoniugantów posiadając swój przedkoniugacyjny makronukleus wyraża swój przedkoniugacyjny typ płciowy. Komórki
te mogą natychmiast (jeśli nadal przebywają w środowisku pozbawionym pokarmu) przystąpić do ponownej koniugacji (A l l e n 1967 a), podczas której każdy z partnerów jest zdolny do wytworzenia w pełni funkcjonalnych pronukleusów, a następnie odtworzenia nowego aparatu jądrowego. Warto zauważyć, że w ten sposób w komórce niepłodnej zachodzi całkowite usunięcie jej poprzedniego genomu (ang. genomie exclusion). Utrata płodności z jednej strony i zachowanie przez komórkę bezpłodną zdolności do przyjęcia pronukleusa migracyjnego z drugiej strony „ratuje” płodną komórkę, która miała „pecha” i związała swój los z komórką, która jest defektywna. W przeciwnym przypadku liczne uszkodzenia genomu występujące w jądrze ge- neratywnym jednego z partnerów podlegałyby postzygotycznej ekspresji w makronukleusie obu koniugantów, uśmiercając także tą komórkę rodzicielską, która była płodna.
UDZIAŁ CYTOSZKIELETU W REGULACJI PŁODNOŚCI
Jak opisano wcześniej wewnątrzcytopla- zmatyczny cytoszkielet koniugantów płodnych tworzy specyficzne stuktury związane z: (1) połączeniem koniugacyjnym, (2) jądrem post- mejotycznym, (3) pronukleusami. W komórkach niepłodnych występują tylko struktury polimeryzujące w okolicy połączenia koniuga- cyjnego. Żadne z jąder nie jest otaczane przez mikrotubule perinuklearne (G a e r t ig i F le u r y 1992). Przypomnijmy, że w koniugacji normalnej mikrotubule perinuklearne występują wokół jednego z jąder postmejotycznych (położonego w pobliżu połączenia koniugacyjnego) i właśnie to jądro jest rekrutowane do podziału
pregamicznego. Tak więc, determinacja wyboru czy Tetyrahymena thermophila pozostanie komórką płodną czy bezpłodną wiąże się ze zdolnością do nukleacji cytoszkieletu wokół jądra postmej otycznego.
W czasie koniugacji dwóch linii niepłodnych (Rye. 6B) makronukłeusy w obu koniugujących komórkach wydłużają się w kierunku połączenia koniugacyjnego. Pomiędzy przednią, silnie wyciągniętą częścią makronukleusa a połączeniem koniugacyjnym powstaje cytoszkielet złożony z mikrotubul i filamentów 49 kDa (N u m ata i współaut. 1985). W tym samym czasie wszystkie cztery postmejotyczne mikronukleusy nie

554 J anina Kaczanowska i w spółau torzy
płodnych komórek pozostają w cytoplazmie, niektóre z nich nawet w pobliżu połączenia koniugacyjnego, i dopiero później podlegają re- sorpcji. Aneuploidalne komórki zachowują więc zdolność do nukleacji struktur cytoszkieletal- nych w rejonie połączenia koniugacyjnego, ale nie wokół mikronukleusów. Być może w „starych", aneuploidalnych mikronukleusach zachodzą takie zmiany w otoczce jądrowej, które uniemożliwiają polimeryzację mikrotubul wokół produktów podziału tego jądra oraz ich podczepienie do połączenia koniugacyjnego i są one jedną z przyczyn ich degradacji.
Jedynym warunkiem przywrócenia zdolności do normalnej koniugacji komórki, która jest biorcą obcego mikronukleusa jest wymiana własnego, „złego” mikronukleusa na „dobry” otrzymany od partnera (A l l e n 1967b). Skoro jednak mikronukleus jest transkrypcyjnie „milczący”, to fakt “ten, znany od blisko 30 lat, stanowi nie lada zagadkę i rodzi następujące pytania:
— czy cytoszkielet perinuklearny, który podczas normalnej koniugacji powstaje na otoczce jednego z czterech produktów mejozy warunkuje jego „przeżycie” w cytoplazmie komór
ki, a następnie podział, który prowadzi do wytworzenia funkcjonalnych pronukleusów?
— czy wzrastająca wraz w wiekiem klonal- nym bezpłodność koniugantów jest związana z uszkodzeniami struktury chromatyny?
— czy DNA zawarte w mikronukleusie może spełniać podczas koniugacji jakąś funkcję strukturalną, niezwiązaną z transkrypcją warunkując powstanie odpowiedniego cytoszkieletu perinuklearnego?
— czy u Tetrahymena występuje „czujnik” molekularny wykrywający uszkodzenia DNA (np. odcinki jednoniciowego DNA), który blokuje powstanie na otoczce jednego z czterech produktów mej ozy cytoszkieletu mikrotubularne- go? Czujnik taki mógłby być analogiczny do kompleksów jakie tworzą w komórkach ssaków poli(ADP-rybozylowane) białka i białko p53 w miejscach uszkodzeń DNA, co wstrzymuje podział komórki (E n o ch i N o r b u r y 1995).
Na te pytania nie znamy na razie odpowiedzi. Ale warto zauważyć, że są to pytania dotyczące występowania w czasie koniugacji Tetrahymena „punktu kontrolnego” uruchamiającego rozwój postzygotyczny, a zarazem roli cytoszkieletu w regulacji płodności.
DEVELOPMENTAL ROLE OF THE CYTOSKELETON IN UNICELLULAR ORGANISMS
Sum m ary
The ability of eukaryotic cells: to assemble the spatial pattern of intracellular structures, to perform directed movements, and to participate in reception, transduction and monitoring of external signals are well acknowledged functions of the intracellular cytoskeleton. Recent studies on unicellular organisms like a ciliate protozoan Tetrahymena and a yeast Schisosaccharomyces pombe revealed new and unexpected functions of the intracellular cytoskeletons in development. In this article the role of the cytoskeleton and reassembling in driving mitotic cycle and sexual processes are reviewed mainly focusing on recent advances in studies on Tetrahymena, with the comparative reference to achievements gained on the yeast model. In this aim and scope of this article, the following problems are discussed: 1. Molecular and structural understanding of the role of the cytosceleton in maintaining and clonal propagation of cellular asymmetry and polarity. 2. Recent advances in studies on mechanisms of the positioning and asymetry of the fission-line region during so called asymmetric cytokinesis. 3. Discovery of the internal signalling system monitoring
the expression of the tubulin gene by the integrity of the preexisting microtubular system and genetic control of posttranslatiónal diversification of microtubules in Tetrahymena. 4. Studies on temporal reassembling of the intra- cytoplasmic, (non-cortical) cytoskeleton during cell to cell contact during conjugation of Tetrahymena thermophila and its role in the stimulation of meiosis, cross fertilization and postzygotic nuclear differentation within the common cytoplasmic compartment. 5. It has been well known that the clonal senescence of Tetrahymena thermophila with gradually declining its fertility during conjugation and with the mechanism of the survival of the mates due to mechanism of „genomic exclusion" is an evolutionary strategy of this species to select the unaffected micronuclear genomes. But only recently, the clonal senescence is ascribed to the failure in reassembling of the cytoskeleton in conjugating mates. This discovery may be crucial for search for the role of cytoskeleton in checkpoint control of elimination of the defective and maintaining of the unaffected sets of the micronuclear chromosomes.
LITERATURA
A dams R. R., A lvaro A.,Tavares M., Salzberg A., B elen H. J., G lover D. M., 1998. Pavarotti encodes a kinesin-like protein required to organize the central spindle and contractile ring fo r cytokinesis. Genes & Dev. 12, 1483- 1494.
A llen S. L., 1967 a. Cytogenetics o f genomic exclusion.Genetics 55, 797-822.
Allen S. L., 1967 b. Genomic exclusion in Tetrahymena. A rapid means fo r inducing homozygous diploid lines in Tetrahymena pyriformis. Science 155, 575-577.
Allen R. D., 1967. Fine structure, reconstitution andpossible functions o f components o f the cortex o f Tetrahymena pyriformis. J.Protozool. 14, 553-565.
Allen R. D., 1969. The morphogenesis o f basal bodies and accessory structures o f the cortex o f the ciliated proto- zoanTetrahymenapyriformis. J. Cell Biol. 40, 716-733.
A ndersen S. S. L., 1999. Balanced regulation o f microtubule dynamics during the cell cycle: a contemporary review. BioEssays. 21, 53-60.

Rola cytoszkieletu w rozwoju organizmów 555
B ac h u r sk i C. J., T h e o d o r a k is N. G., C o u ls o n R. M. R., C levelan d D. W., 1994. An amino-terminal tetrapeptide specifies cotranslational degradation o f y-tubulin but not fi-tubulin mRNAs. Mol. Cell Biol. 14, 4076-4086.
BAh le r J., St e e v e r A . B ., W h e a t le y S ., W a n g Y ., P r in g le J. R., G o u ld K. L., M c c o llu m D., 1998. Role o f Polo Kinase and M idlp in Determining the Site o f Cell Division in Fission YEast. J. Cell B io l. 143, 1603-1616.
B alas u b r am a n ian M. K., M c c o llu m D., S u r a n a U., 2000. Tying the knot: linking cytokinesis to the nuclear cycle. J. Cell Sci. 113, 1503-1513.
B eisson J ., J e r k a -D z ia d o s z M ., 1999. Polarities o f the centri- olar structure: Morphogenetic consequences. Biol.Cel91, 367-378.
B er s h a d sk y A., C h a u s o w s k y A., B ec k e r E., L yu b im o v a A., G eig e r B., 1996. Involvement o f microtubules in the control o f adhesion-dependent signal transduction. Curr. Biol. 6, 1279-1289.
B ernste in M., B e ec h P. L., K a t z S. G., R o ze n b a u m J. L., 1994. A new kinesin-like protein (K lpl) localized to a single microtubule o f the Chlamydomonas fagellum. J. Cell Biol. 125, 11313-1326.
B runs P., B r u ssard T. P, 1974. Pair formation inTetrahyme- napynformis, an inducible developmental system. Exp. Cell Res. 188, 337-344.
B urns R. G., 1991. a-, (3-, andy-tubulins: sequence comparison and structural constraints. Cell Motil. Cytoskel. 20, 181-189.
C le v e la n d , D. W., 1989. Autoregulated control o f tubulin synthesis in animal cells. Curr. Opin. Cell Biol. 1, 10-14.
C levelan d D. W., Ł o pa ta M. A., S h e r lin e P., K ir s c h n e r M. W., 1981. Unpolymerized tubulin modulates the level o f tubulin mRNAs. Cell 25, 537-546.
C leveland D. W., P itt e n g e r M. F., F e r a m isc o J. R., 1983. Elevation o f tubulin levels by microinjection supresses new tubulin synthesis. Nature 305, 738-740.
C ole E., S o e l te r T. A., 1997. A mutational analysis of conjugation in Tetrahymena thermophila. 2. Phenotypes affecting middle and late development: third prezygotic nuclear division, pronuclear exchange, pronuclear fu sion and postzygotic development. Devel. Biol. 189, 233-245.
D ebec A ., D etraves C., M o n tm o r y C., G e r a u d , W r ig h t M .,1995. Polar organization o f gamma-tubulin in acentdolar mitotic spindles o f Drosophila melanogaster cells. J.Cell Sci., 108, 2645-2653.
D o e r d e r F. P, G a t e s M. A. Q ., E b e r h a a r d t F. P., A rslan yo lu M., 1995. High frequency o f sex and equal frequencies o f mating types in natural populations o f the ciliate Tetrahymena thermophila. Proc. Natl Acad. Sci. USA92, 8715-8718.
D o e r d e r F. P, A r s lan yo lu M., S a a d Y., K a c z m a r e k M., M e n d o za M., M ita B., 1996. Ecological genetics o f Tetrahymena thermophila: Mating types, i-antigens, multiple alleles and epistasis. J. Eukaryot.. Micr. 43, 95-100.
D ufre sne L., N e a n t J., S t -P ie r r e J., D u b e F., G u e r r ie r P., 1991. Effects o f 6-dimethylaminopurine on microtubules and putative intermediate filaments in sea urchin embryos. J.Cell Sci. 99, 721-730.
E d em atsu H., H iro no M., W atan ab f . Y., 1992. Tetrahymena profilin is localized in the division furrow . J. Biochem. 112, 637-642.
E llio tt A . M ., T r e m o r J., 1958. The fine structure o f the pellicle in the contact area o f conjugating Tetrahymena pyriformis. J. Biophys. Biochem. Cytol. 4, 839-840.
E noch T., N o r b u r y C., 1995. Cellular responses to DNA damage: cell cycle checkpoint, apoptosis and the roles o f p53 and ATM. Trends in Biochem. Sci. 20, 426-430.
Eva n g e lista M., B lu n d e ll K, L o n g tin e M. S., C h o w N., A d am e s J. R., P r in g le M. P., B o o n e C., 1997. Bnilp., a yeastformin linking cdc42p. and the actin cytoskeleton
during polarized morphogenesis. Science 276, 118- 122.
F ran k e l J., 1989. Pattern Formation: Ciliate Studies and Models. Oxford Univ. Press, NY.
F ran k e l J., 2000. Cell biology o f Tetrahymena thermophila. [W:] Methods in Cell Biol 62. D. J., A sai J. D., F o r n e y (red.) Acad Press, str. 28-125.
F ran k e l J . ,N elsen E. M., M a r t e l E., 1981. Development o f the ciliature o f Tetrahymena thermophila II. Spatial subdivision prior to cytokinesis. Devel. Biol. 88, 39-54.
F ry A. M., M era ld i P., N igg E. A., 1998. A centrosomal function fo r the human Nek2 protein kinase, a member of the NIMA family o f cell cycle regulators. EMBO J. 17, 47-481.
F u r g e K. A., W ong K., A r m s tr o n g J., B ala s u b r a m a n ia n M., A lb r ig h t C. F., 1998. Byr 4 and Cdc 16form a two-component GTPase-activating protein fo r the Spgl GTPase that controls septation in fission yeasts. Curr. Biol. 8, 947-954.
G a e r t ig J., F l e u r y A ., 1992. Spatio-temporal reorganization o f intracytoplasmic microtubules is associated with nuclear selection and differentiation during the development process in the ciliate Tetrahymena thermophila. Protoplasma 167, 74-87.
G a e r t ig J., T h a t c h e r T . H., M c g r ath K. E., C a lla h a n R. C., G o r o v sk y M. A., 1993. Perspectives on tubulin isotype function and evolution based on the observations that Tetrahymena thermophila microtubules contain a single a- and (3-tubulin. Cell Motil. Cytoskeleton 25, 243-253.
G a e r t ig J., T h atc h e r T . H., G u L., G o r o v s k y M. ,1994. Electroporation-mediated repleacement of a positively and negatively selectable Q-tubulin gene in Tetrahymena thermophila. Proc. Natl. Acad. Sci. USA 91, 4549- 4553.
G a e r t ig J., C ru z M. A., B o w en J., Gu L., P e n n o c k D. G., G o r o v sk y M., 1995. Acetylation o f lysine 40 in a-tubulin is not essential in Tetrahymena thermophila. J. Cell Biol. 129, 1301-1310.
G a o Y., V ain b e r g I. E., C h o w R. L. C o w an N. J., 1993. Two cofactors and cytoplasmic chaperonins are required fo r the folding o f alfa- and beta- tubulin. Mol.Cell Biol. 13, 2478-2485.
G o n d a K., N ish ib o r i K., O h b a H., W a t a n a b e A., N u m a t a O., 1999a. Molecular cloning o f the gene fo r p85 that regulates the initiation o f cytokinesis in Tetrahymena. Biochem. Biophys. Res. Comm. 264, 112-118.
G o n d a K., Ka t o h M., H an y u K., W a t a n a b e Y. N u m a t a O., 1999b. Ca2+/calmodulin andp85 cooperatively regulate in initiation of cytokinesis in Tetrahymena. J. Cell Sci. 112,3619-3626.
G r an d c h am p P. S., B eisson J., 1981. Positional control o f nuclear differentiation in Paramecium Dev. Biol. 81, 336-341.
Gu L., G a e r t ig J., S ta r g e l l L. A., G o r o v s k y M. A. 1995 Gene-specific signal transduction between microtubules and tubulin genes in Tetrahymena thermophila. Mol.Cell Biol. 15, 5173-5179.
H ab le W. E., K r o p f D . L., 2000. Spermentry induces polarity in fucoid zygotes. Development 127, 493-501.
H a m il t o n E. P., S u h r -J e s s e n P. B., O rias E., 1988. Pronuclear fusion failure: an alternate conjugationcd pathway in Tetrahymena thermophila, induced by vinblastine. Genetics 118, 627-636.
H en n e s se y T . M., K u r u v illa H . G., 1999. Electrophysiology o f Tetrahymena. [W:] Methods Cell Biol. 62. D. J. A sai J. D. F o r n e y J. D. (red.), Acad. Press , str. 363-378.
H ir a n o -O h nish i J., W atan ab e Y., 1989. Ca2+/calmodulin — dependent phosphorylation o f ciliary -tubulin in Tetrahymena. J. Biochem. 105, 858-860.
H iro no M., N a k a m u r a M., T su n e m o to M., Y a s u d a T ., O h b a H., N u m ata O., 1987. Tetrahymena actin: Localization and

556 Janina Kaczanowska i współau torzy
possible biological roles o f actin in Tetrahymena cells. J. Biochem. 102, 537-545.
H o y le H. D., R a f f E . C ., 1990. Two Drosophila beta- tubulin isoforms are not functionally equivalent. J. Cell Biol. I l l , 1009-1026.
Hsu J. Y., S un Z-W., Li X., R e u b e n M., T a t c h e ll K., B ish o p
D. K., G r u sh c o w J. M., B r am e C. J., C a l d w e l l J. A., H u n t D. F., L in R., S m ith M. M., A llis C. D., 2000. Mitotic phosphorylation o f histone H3 is governed by Ipll /aurora Kinase and Glc7/PPl phosphatase in budding yeast and nematodes. Cell 102, 279-291.
J er k a - D zia d o sz M., 1981. Cytoskeleton-related structures in Tetrahymena thermophila microfilament at the apical and division-furrow ring. J. Cell Sci. 51, 241-253.
J er k a -D zia d o sz M., J e n k in s I. M., N e lse n E . M., W ill ia m s N .E., J a e c k e l -W illlams R., F r a n k e l J., 1995. Cellular polarity in ciliates: persistence o f global polarity in a disorganized mutant o f Tetrahymena thermophila that disrupts cytoskeletal organization. Devel. Biol. 169, 644-661.
K ac z a n o w s k a J., B u za n sk a L.,O str o w sk i M., 1993. Relationship between spatial pattern o f basal bodies and membrane skeleton (epiplasm) during the cell cycle of Tetrehymena: cdaA mutant and anti-membrane skeleton immunostaining. J. Euk. Microbiol. 40, 747-754.
K a c z a n o w sk a J ., J o a c h im ia k E ., B u za n s k a L., K r aw c zyń sk a W ., W h e a tle y D. N ., K ac z an o w sk i A., 1999. Molecular subdivision o f the cortex o f dividing Tetrahymena is coupled with the formation o f the fission zone. Devel. Biol. 212, 150-164.
K ac zan o w sk i A., 1978. Gradients o f prolferation o f ciliary basal bodies and the determination o f the position o f the oral primordium in Tetrahymena J. Exp. Zool. 204, 417-430.
K a c zan o w sk i A., R a m e l M., K a c z a n o w s k a J., W h e a tle y D., 1991. Macronuclear differentiation in conjugating pairs of Tetrahymena treated with antitubulin drug nocoda- zole. Exp. Cell Res. 195, 330-337.
K ie r sn o w s k a M., K ac z a n o w sk i A., DE H a l l e r G., 1993. Inhibition o f oral morphogenesis during conjugation o f Tetrahymena thermophila and its resumption after cell separation. Europ. J. Protistol. 29, 359-369.
K ie r sn o w s k a M., G o l in s k a K ., 1996. Patterns o f the phospho- rylated structures in the morphostatic ciliate Tetrahymena thermophila: MPM-2 immunogoldlabelling. Acta Protozool. 35, 297-308.
K ie r sn o w s k a M., K ac z an o w sk i A., M o r g a J., 2000. Macronuclear development in conjugants o f Tetrahymena thermophila, which were artificially separated at meiotic prophase. J. Euk. Micr. 47, 139-147.
K o j im a H ., W a t a n a b e Y ., N u m a t a O., 1997. The dual functions o f Tetrahymena citrate synthase are due to the polymorphism o f its isoforms. J. Biochem. 122, 998-1003.
K rio u tc h k o v a M. K., O n ish c h e n k o G. E., 1999. Structural and functional characteristics o f the centrosome in gametogenesis and early embryogenesis o f animals. Int. Rev. Cytol. 185, 107-151.
K r z yw ic k a A., K ie r sn o w s k a M., W ło g a D., K a c z a n o w s k a J.,1999. Analysis o f the effects o f the cdaKl mutation o f Tetrahymena thermophila on the morphogenesis o f the fission line. Europ. J. Protistol. 35, 342-352.
LA F o u n ta in J. R., D av id so n L. A., 1980. An analysis of spindle ultrastructure during anaphase o f micronuclear division in Tetrahymena Cell Motil. 1, 41-61.
Lee G., 1993. Non- motor microtubule- associated proteins. Curr. Opin. Cell Biol. 5, 88-94.
L ittle M., S e e h a u s T ., 1988. Comparative analysisof tubulin sequences. Comp. Biochem. Physiol. 90B, 655-670.
L o g ar in h o E ., S u n k e l C. E ., 1998. The Drosophila POLO kinase localises to multiple compartments o f the mitotic apparatus and is required fo r the phosphorylation o f MPM2 reactive epitopes. J. Cell Sci. I l l , 163-167.
L u d u e n a R. F., 1993. Are tubulin isotypes functionally significant? Mol. Biol. Cell 4, 445-457.
M a c h e m e r H .,O g u ra A., 1979. Ionic conductances o f membranes in ciliated and deciliated Paramecium. J. Physiol. 296, 49-60.
M a s u d a H., 1995. The formation and functioning o f yeast mitotic spindles. BioEssays 17, 45-51.
M a t a J., N u r se P., 1997. Teal and the microtubular cytos- keleton are importantfor generating global spatial order within the fission yeast cell. Cell 89, 939-949.
M a t s u s a k a T., H ir ata D., Y a n a g id a M., T o d a T., 1995. A novel protein kinase gene ssp l+ is required fo r alteration of growth polarity and actin localization in fission yeast. EMBO J. 14, 3325-3338.
M c C o llu m D., B a la s u b r a m a n ia n MK., P e l c h e r LE., H em m in g - s e n S M ., G o uld KL., 1995. Schisosaccharomycespombe cdc4+ gene encodes a novel EF-hand protein essential fo r cytokinesis. J. Cell Biol. 130, 651-660.
Mc D onald B. B., 1966. The exchange ofRNA and protein during conjugation in Tetrahymena J. Protozool. 13, 277-285.
M ikam i K., 1980. Differentiation o f somatic and germinal nuclei correlated with intracellular localization in Paramecium caudatum exconjugants. Dev. Biol. 80, 46-55.
M ikam i K., N g S. F., 1983. Nuclear differentiation in Paramecium tetraurelia. Exp. Cell Res. 144, 25-30.
N a n n e y D. L., 1953. Nucleoplasmic interactions during conjugation in Tetrahymena. Biol. Bull. 105, 133-148.
N a n n e y D. L., 1966. Cortical integration in Tetrahymena: An exercise in cytogeometry. J. Exp. Zool. 161, 307-317.
N a n n e y D. L., 1975. Patterns o f basal body addition in ciliary rows o f Tetrahymena. J. Cell Biol. 79, 727-736.
N e lsen E. M., W ill ia m s N. E., Yi H., K n aa k J., F r a n k e l J.,1994. „Fenestrin” and conjugation in Tetrahymena thermophila. J. Euk. Microbiol. 41, 483-495.
N u m ata O., 1996. Multifunctional proteins in Tetrahymena 14-nmfilament protein/citrate synthase and translation elongation factor-1 -a. Int. Rev. Cytol. 164, 1- 35.
N u m a t a O., H iro no M., W a t a n a b e Y., 1983. Involvement of Tetrahymena intermediate filament protein, a 49 K protein, in the oral morphogenesis. Exp. Cell Res. 148, 207-220.
N u m a t a O., S ugai T., W a t a n a b e , 1985. Control o f germ cell nuclear behaviour at fertilization by Tetrahymena intermediate filament protein. Nature 314, 192-194.
N u m a t a O., T a k e m a s a T., T ak a g i I., H iro n o M., H ir an o H., C h ib a J., W a t a n a b e Y., 1991. Tetrahymena 14-nmfilament — forming protein has citrate synthase activity. Biochem. Biophys. Res. Comm. 174, 1028-1034.
O r ia s E ., 1986. Ciliate conjugation. [W:] The Molecular Biology o f a Ciliated Protozoa. J. G., G a l l (red.). Acad. Press N.Y., str. 45-84.
P a s c h a l B. M., M ikam i A., P fis t e r K. K., VALLee R. B., 1992. Homology o f the 74kD cytoplasmic dynein subunit with a flagellar dynein polypeptide suggest an intracellular targeting Junction. J. Cell Biol. 118, 1133-1143.
P e n q u e D., Ga e lg o L., R o d r ig u e s -P o u sa d a C., 1991. Multiple a-tubulin isoforms in cillia and cytoskeleton o f Tetrahymena pyrform is generated by post-translational modification. Studies durind reciliation. Eur. J. Biochem. 195, 487-494.
R o b in s o n D. N., S pu d ic h J. A., 2000. Towards a molecular understanding o f cytokinesis. Trends in Cell Biol. 10, 228-237.
R u d n e r A. D., M u r r ay A. W., 1996. The spindle assembly checkpoint. Curr. Opin. Cell Biol. 8, 773-780
Ruiz F., B e isso n J., R o s s ie r J., D u pu is -W ill ia m s P., 1999. Basal body duplication in Paramecium requires y-tubulin. Curr. Biol. 9, 43-46
S aw in K. E., N u rse P., 1998. Regulation o f cell polarity by microtubules in fission yeast. J. Cell Biol. 142, 457- 471.

Rola cytoszkieletu w rozwoju organizmów 557
Savage C ., H a m e n il M ., C u lo t t i J. G., C o u ls o n A ., A lb e r ts o n D. G., C h alfie M., 1989. mec-7 is a fi-tubulin gene required fo r the production o f 15 protofilament microtu- bules in Caernorhabditis elegans. Genes Dev. 3, 870- 881.
Schebel E., 2000. Tubulin complexes binding to the centro- some, regulation and microtubule nucleation. Curr. Opin. Cell Biol. 12, 113-118.
S k o u fias D. A., S c h o le y J. M., 1993. Cytoplasmic microtu- bule-based motor proteins. Curr. Opin. Cell Biol. 5, 95-104.
S o ares H., Cy r n e L., C a s a lo u C., G u e r r e ir o P., R o d r ig u e s - P o u s a d a C., 1995. Will the chaperonin CCT be involved in the functional role o f tubulins in Ciliates? [W :] Protis- tological Actualites. G. B r u g e r o lle ]-P. M ig n o t (red.). Clermont-Ferrand Univ., str. 34- 43.
S parks C. A., M o r ph e w M., M c c o llu m D., 1999. Sid2p., a spindle pole body kinase that regulates the onset o f cytokinesis. J. Cell Biol. 146, 777-790.
S o r g er P. K., D o b le s M ., T o u r n e b ize R ., H ym an A . A ., 1997. Coupling o f cell division and cell death to microtubule dynamics. Curr.Op. Cell Biol. 9, 807 -814.
St a r g e ll L. A., H eru th D. P., G a e r t ig J . ,G o r o v s k yM., 1992. Drugs affecting microtubule dynamics increase a-tubulin mRNA accumulation via transcription in Tetrahymena thermophila. Mol. Cell Biol. 12, 1443-1450.
St -P ierre J., D u fr e sn e L., 1990. Identification and localization o f proteins immunologically related to intermediate proteins in sea urchin eggs and embryo cell. Cell Motil. Cytoskel. 17, 75-86.
St -P ierre J., V in c e n t M., D u fr e sn e L., 1994. Effects of 6-dimethylaminopurine on the length o f the cell cycle and the state o f phosphorylation o f putative intermediate filament proteins in sea urchin embryos. Cell Motil. Cytoskel. 29, 131-140.
S u g an u m a Y., S h im o d e C, Y a m a m o t o H., 1984. Conjugation in Tetrahymena: Formation o f a special junction area for conjugation during the co-stimulation period. J. Electron Microsc. 33, 10-18.
T akag i I., N u m a t a O., W a t a n a b e Y., 1991. Involvement o f 14-nm filament-forming protein and tubulin in gametic pronuclear behavior during conjugation in Tetrahymena. J. Protozool. 38, 345-351.
T h o m pso n G. A., B au g h L. C., W a l k e r L. F., 1974 Non-lethal deciliation o f Tetrahymena by a local anesthetic and its utility as a tool fo r studying cilia regeneration. J. Cell Biol. 61, 253-257.
V erd e F., W ile y D. J., N ur se P., 1998. Fission yeast orb6, a ser/thrprotein kinase related to mammalian rho- kinase and myotonic dystrophy kinase, is requiredfor maintenance o f cell polarity and co-ordinates cell morphogenesis
with the cell cycle. Proc. Natl. Acad. Sci USA 95, 7526- 7531.
W a n g S., N a k a s h im a S., SAKAI H., N u m a t a O. F ujiu K., N o saw a Y ., 1998. Molecular cloning and cell cycle dependent expression o f a novel NIMA (Never in Mitosis A.nidulansj-related protein kinase (TtNRK) in Tetrahymena cells. Biochem J. 334, 193-203.
W a t a n a b e A., K u r as aw a Y., W a t a n a b e Y., N u m a t a O., 1998. A new Tetrahymena actin-binding protein is localized in the division furrow. J. Biochem. 123, 607-6113.
W a t t s J. L., M o r t o n D. G., B e s tm a n J., K e m ph u e s K. J., 2000. The C. elegans par-4 gene encodes a putative serine- threonine kinase required fo r establishing embryonic asymmetry. Development 127, 1467-1475.
W ill ia m s N. E., H o n ts J. E., J a e c k e l -W ill ia m s R. F., 1987. Regional differentiation o f the membrane skeleton in Tetrahymena. J.Cell Sci. 87, 457-463.
W ill ia m s N. E., H o n ts J. E., K a c z a n o w s k a J., 1990. The formation o f basal body domains in the membrane skeleton o f Tetrahymena. Development 109, 935-942.
W ill ia m s N . E., N else n E. M., 1997. HSP70 and HSP90 homologs are associated with tubulin in hetero-oli- gomeric complexes, cilia and the cortex o f Tetrahymena. J. Cell Sci., 110, 1665-1672.
W o lf e J., 1982. The conjugation junction in Tetrahymena: Its structure and development. J. Morphol. 172, 159- 178.
W o lf e J., 1985. Cytoskeletal reorganization and plasma membrane fusion in conjugating Tetrahymena. J. Cell Sci. 73, 69-85.
W o lf e J., G rim e s G. W., 1979. Tip transformation in Tetrahymena: a morphologenetic response to interactions between mating types. J. Protozool. 26, 82-89.
W o lf e J., P ag lia r o L ., F o r tu n e H., 1986. Coordination o f concanavalin-A-receptor distribution and surface differentiation in Tetrahymena. Differentiation 31, 1-9.
Y an a g i A., 1987. Positional control o f fates o f the nuclei produced after meiosis in P. caudatum. Devel. Biol. 122, 535-539.
Y e X . S., X u G., P u R. T., F in c h e r R. R., M c g u ir e S. L., O sm ani A. H. AND O sm an i S. A., 1995. The NIMA protein kinase is hyperphosphorylated and activated downstream o f p34 2/ cyclin B: co-ordination o f two mitosis promoting kinases. EMBO J. 14, 986-994.
X ia L ., Ha i B ., G a o Y ., B u r n e tte D ., T h azh a th R., D u an J., B re M.H., L e villie r s N., G o r o v s k y M. A., G a e r tig J., 2000. Polyglycylation o f tubulin is essential and affects cell motility and division in Tetrahymena thermophila. J. Cell Biol. 149, 1097-1106.


















