
Iodation des protéines par voie enzymatique : 2. Formation d'un composé intermédiaire...
5
European J. Biochem. 5 (1968) 114-118 Iodation des protkines par voie enzymatique 2. Formation d’un composC intermediaire peroxydase-haloghe J. NUNEZ et J. POMMIER Laboratoire de Biochimie GAnArale et Comparee du College de France, Paris (Rep le 26 decembre 1967/1 avrill968) In the thyroid gland the iodination of thyroglobulin is performed by a peroxidase. Hors- radish peroxidase also catalyses, in the presence of an H,O,-generating system, both the iodide oxidation and the iodination of tyrosine residues of a protein. In the present work we have shown that the enzyme forms with the halogen an intermediary compound which is relatively stable and can be isolated. The halogen, probably present in an oxidised form in the complex, is directly transferred to the protein. In a first type of experiment, we have identified an iodinated derivative of the peroxidase after incubation of the enzyme with an H,O,-generating system (glucose + glucose oxidase). Iodine-labelled enzyme, isolated by filtration on Sephadex G-25, contains the halogen partly in reactive form (approximately 20 and partly as mono- and diiodotyrosine. I n the experimental conditions adopted, the enzyme itself can be a protein acceptor of iodine. In a second type of experiment the enzyme is preincubated with the H,O,-generating system and iodide (15 pg) labelled with a first isotope l3II. After a few minutes, thyroglobulin is added to the reaction mixture together with more iodide (100 pg) labelled with SL second isotope lZ5I. After a 20-minute incubation the 1311/1251 isotope ratio is always higher in thyroglobulin than in the excess free iodide. I n the same experiments the iodination heterogentity of thyroglobulin was also confirmed by sucrose gradient ultracentrifugation. The 131I/1Z5I ratio is much higher in the thyroglobulin fractions which sediment rapidly than in those which have a slower sedimentation coefficient. In a third type of experiment the iodinated peroxidase is mixed a t 0” with thyroglobulin, and the reactive iodine contained in the enzyme is directly transferred to the protein. No free iodide can be detected. Such a transfer does not require a H,O,-generating system. All these results show the existence of a enzyme-halogen complex in which the halogen is very probably in an oxidised form. Since the halogen is transferred directly from the complex to the thyroglobulin, an interaction between the enzyme and the tyrosine residues of the protein can be assumed. La glande thyroide contient un systkme per- oxydasique qui, en presence d‘un systkme donateur d’eau oxygBn8e catalyse l’oxydation de l’iodure et permet l’halogbnation de la thyroglobuline [l -41. Diverses peroxydases vBg6tales ou animales cataly- sent la m6me reaction [a-61. Nous avons montrb notamment gue la peroxydase de raifort, enzyme cristallis&e, est un bon modhle de la &action d’ioda- tion d’un accepteur proteique [5]. Dans ce travail, nous avons mis en Bvidence la formation d’un com- plexe intermediaire enzyme-halogkne et un transfert direct de l’halogbne du complexe b l’accepteur proteique. Enzymes. Peroxydase de raifort (EC 1.11.1.7) et glucose oxydase (EC 1.1.3.4). MATBRIEL ET METHODES Produits Les produits utilisBs sont: de la peroxydase de raifort de Rz = 3 (Boehringer), du glucose anhydre (Prolabo), de la glucose oxydase grad A (Boehringer), de la shrumalbumine de pore grad A (Calbiochem), de la thyroglobuline purifiee de mouton [7] et de la pronase (Calbiochem) . Mdthodes de purification Les milieux rBactionnels sont purifihs ou analyses : (a) par percolation sur SephadexG-25 ou 6-200, bquilibre avec un tampon phosphate 0,05 M pH 6; (b) par chromatographie sur papier en solvant
Transcript of Iodation des protéines par voie enzymatique : 2. Formation d'un composé intermédiaire...

European J. Biochem. 5 (1968) 114-118
Iodation des protkines par voie enzymatique
2. Formation d’un composC intermediaire peroxydase-haloghe
J. NUNEZ et J. POMMIER
Laboratoire de Biochimie GAnArale e t Comparee du College de France, Paris
( R e p le 26 decembre 1967/1 avrill968)
In the thyroid gland the iodination of thyroglobulin is performed by a peroxidase. Hors- radish peroxidase also catalyses, in the presence of an H,O,-generating system, both the iodide oxidation and the iodination of tyrosine residues of a protein.
I n the present work we have shown that the enzyme forms with the halogen an intermediary compound which is relatively stable and can be isolated. The halogen, probably present in an oxidised form in the complex, is directly transferred to the protein.
In a first type of experiment, we have identified an iodinated derivative of the peroxidase after incubation of the enzyme with an H,O,-generating system (glucose + glucose oxidase). Iodine-labelled enzyme, isolated by filtration on Sephadex G-25, contains the halogen partly in reactive form (approximately 20 and partly as mono- and diiodotyrosine. I n the experimental conditions adopted, the enzyme itself can be a protein acceptor of iodine.
In a second type of experiment the enzyme is preincubated with the H,O,-generating system and iodide (15 pg) labelled with a first isotope l3II. After a few minutes, thyroglobulin is added to the reaction mixture together with more iodide (100 pg) labelled with SL second isotope lZ5I. After a 20-minute incubation the 1311/1251 isotope ratio is always higher in thyroglobulin than in the excess free iodide. I n the same experiments the iodination heterogentity of thyroglobulin was also confirmed by sucrose gradient ultracentrifugation. The 131I/1Z5I ratio is much higher in the thyroglobulin fractions which sediment rapidly than in those which have a slower sedimentation coefficient.
In a third type of experiment the iodinated peroxidase is mixed a t 0” with thyroglobulin, and the reactive iodine contained in the enzyme is directly transferred to the protein. No free iodide can be detected. Such a transfer does not require a H,O,-generating system.
All these results show the existence of a enzyme-halogen complex in which the halogen is very probably in an oxidised form. Since the halogen is transferred directly from the complex to the thyroglobulin, an interaction between the enzyme and the tyrosine residues of the protein can be assumed.
La glande thyroide contient un systkme per- oxydasique qui, en presence d‘un systkme donateur d’eau oxygBn8e catalyse l’oxydation de l’iodure et permet l’halogbnation de la thyroglobuline [l -41. Diverses peroxydases vBg6tales ou animales cataly- sent la m6me reaction [a-61. Nous avons montrb notamment gue la peroxydase de raifort, enzyme cristallis&e, est un bon modhle de la &action d’ioda- tion d’un accepteur proteique [5]. Dans ce travail, nous avons mis en Bvidence la formation d’un com- plexe intermediaire enzyme-halogkne et un transfert direct de l’halogbne du complexe b l’accepteur proteique.
Enzymes. Peroxydase de raifort (EC 1.11.1.7) et glucose oxydase (EC 1.1.3.4).
MATBRIEL E T METHODES
Produits Les produits utilisBs sont: de la peroxydase de
raifort de Rz = 3 (Boehringer), du glucose anhydre (Prolabo), de la glucose oxydase grad A (Boehringer), de la shrumalbumine de pore grad A (Calbiochem), de la thyroglobuline purifiee de mouton [7] et de la pronase (Calbiochem) .
Mdthodes de purification Les milieux rBactionnels sont purifihs ou analyses :
(a) par percolation sur SephadexG-25 ou 6-200, bquilibre avec un tampon phosphate 0,05 M pH 6; (b) par chromatographie sur papier en solvant
Vo1.5, No.l,1968 J. NUNEZ et J. POMMIER 115
n-butanol-acide acetique-eau (8:2:2, v/v/v). On &pare ainsi les protkines halogknees marquees ob- tenues, qui restent B l’origine, de l’iodure libre en excits et Bventuellement des mono- et diiodotyrosines ; (c) par ultracentrifugation en gradient de saccharose de 100-250 g/l poursuivie pendant 15 h B 39,000 tours/ min dans le rotor XW 39 de la centrifugeuse Spinco L2.
Me‘tthodes de comptage Les comptages ont kti: rkalises au moyen d’un
cristal B scintillation monte sur un spectromAtre B deux Bchelles de comptage (Auto y Packard) rkgl6es pour permettre la mesure simultanee des deux iso- topes, 1311 et lZ5I. Dans les conditions de comptage adoptkes, un faible pourcentage du spectre d’1311 se superpose B celui d’1251. Un calcul simple permet de corriger l’activiti: obtenue pour 1251.
Hydrolyse des protdines ioddes obtenues Elle est realishe par action de la pronase B la con-
centration finale de 0,5g/100ml B 37” pendant 15 heures.
Afesure de l’activite‘ peroxydasique vis-ci-vis du gaiacol
L’activitb peroxydasique des diverses fractions obtenues par ultracentrifugation en gradient de saccharose des milieux rbactionnels des divers essais decrits plus loin est mesuree en prenant le gafacol eomme substrat. A chaque fraction sont ajout6s 0,2 ml de galacol (0’25 mg), 0,7 ml de tampon phos- phate 0,05 M de pH 6 et 50 p1 de solution d’H,O, 50mN. On determine pour chaque fraction l’aug- mentation d’absorbance B 470mp en une minute B temperature ordinaire dans les cuves de 1 ml du spectrophotometre Zeiss.
Conditions d’incubation Dans tous les essais, l’incubation a 6ti: realishe B
37” dans le tampon phosphate 0’05 M pH 6 ; le systeme donateur d’H,O, etait constitue par un mklange de 10 pg de glucose oxydase et de 37,5 pg de glucose.
Les essais permettant de mettre en bvidence, par double marquage, le complexe enzyme-haloghe, ont 6th realisbs de la fagon suivante: 250 pg de peroxy- dase dissous dans 1 ml de tampon phosphate 0,05 M pH 6, sont prAincub6s pendant des temps variant de 3 L 10 min selon les essais, en presence du systbme generateur d’H202, de 15 pg d’IK et d’une dose traceuse d’WNa. A la fin de cette prbincubation, une partie aliquote est prelevite et deposee sur un chromatogramme sur papier; 2 mg de thyroglobuline, 100 pg d’IK et m e dose traceuse d’1251Na sont alors ajoutes simultanbment au milieu de prbincubation ; l’incubation est poursuivie pendant 20 minutes. 8*
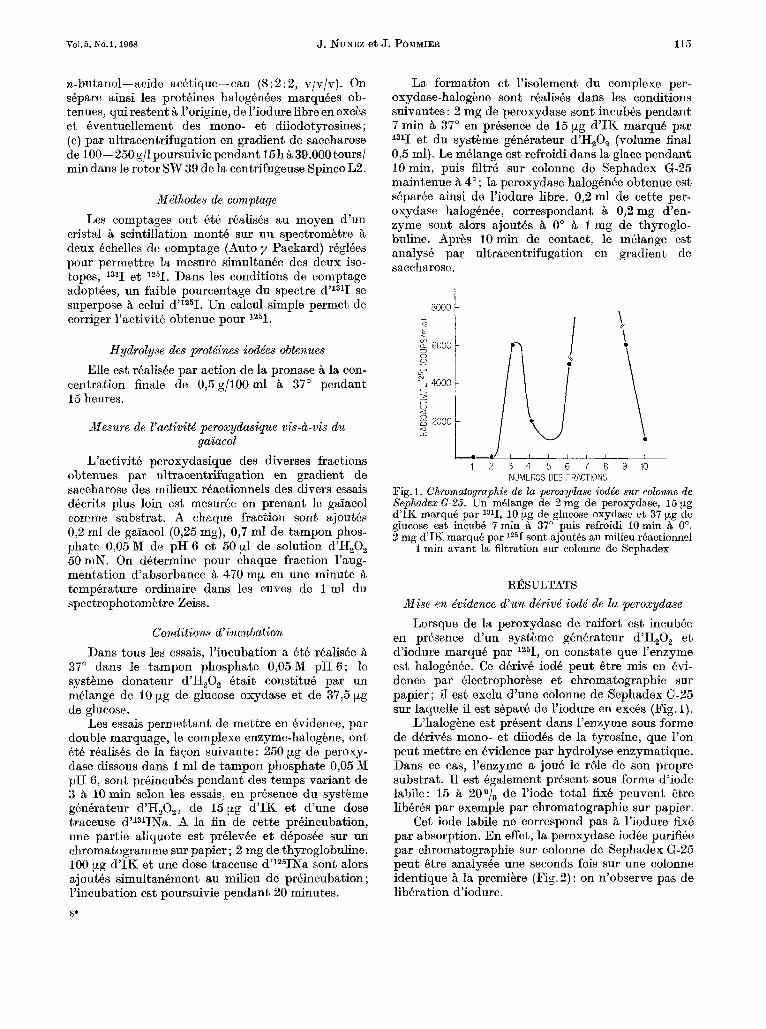
La formation et l’isolement du complexe per- oxydase-haloghe sont realisks dam les conditions suivantes : 2 mg de peroxydase son6 incubks pendant 7 min B 37” en presence de 15 pg d’IK marque par 1311 et du systeme generateur d’HzO, (volume final 0,5 ml). Le melange est refroidi dans la glace pendant lOmin, puis filtri: sur colonne dc Sephadex G-25 maintenue B 4”; la peroxydase halogknke obtenue est skpart?e ainsi de I’iodure libre. 0,2 ml de cette per- oxydase halogenke, correspondant A 0,2 mg d’en- zyme sont alors ajoutes B 0” B 1 mg de thyroglo- buline. Aprks 10min de contact, le melange est analysk par ultracentrifugation en gradient de saccharose.
aooo c \ I
\. L, - J I 1 , , , I I I
1 2 3 4 5 6 7 8 9 10 NUMEROS DES FRACTIONS
Fig. 1. Chroinatographie de la peroxydase iodke sur colonne de Sephadex G-25. Un melange de 2 mg de peroxydase, 15 pg d’IK marque par 1311, i0 pg de glucose oxydase et 37 pg cle glucose est incub6 7 rnin B 37” puis refroidi 10 min B 0”. 2 mg d‘IK marqni! par lz5I sont ajoutks au milieu rriactionnel
1 min avant la filtration sur colonne de Scphadex
R~SULTATS
Mise en Lvidence d’un ddrive‘ iode’ de la peroxydase Lorsque de la peroxydase de raifort est incubbe
en presence d’un systbme genkrateur d’H,O, e t d’iodure marque par 1251, on constate que l’enzyme est halogkn8e. Ce derive iod6 peut &tre mis en kvi- dence par hlectrophorbse et chromatographie sur papier; il est excIu d’une colonne de Sephadex G-25 sur laquelle il est skparb de l’iodure en exc& (Fig. 1).
L’halogene est present dans l’enzyme sous forme de derives mono- et diiodes de la tyrosine, que l’on peut mettre en evidence par hydrolyse enzymatique. Dans ce cas, l’enzyme a jouk le r61e de son propre substrat. I1 est egalement present sous forme d’iode labile: i5 L 2001, de l’iode total fix6 peuvent &re lib&& par exemple par chromatographie sur papier.
Cet iode labile ne correspond pas L l’iodure fix6 par absorption. En effet, la peroxydase iodee purifXe par chromatographie sur colonne de Sephadex G-25 peut &re analysee une seconds fois sur une colonne identique B la premiere (Fig. 2 ) : on n’observe pas de libkration d’iodure.

116 Iodation des proteines par voie enzymatique European J. Biochem.
Ces rbsultats indiquent donc que la peroxydase incubbe en prbsence d’iodure et en absence d’accep- teur proteique oxyde celui-ci, joue le r61e de substrat vis-a-vis de l’halogitne oxyde et renferme une partie de l’halogitne sous forme labile. Par eontre en pre- sence d’accepteur protitique l’enzyme n’est pas halogbnke [5].
A la suite de ces rbsultats, nous avons emis l’hypothitse que la fraction d’iodure labile est fixee sur l’enzyme en un complexe intermediaire, probable- ment sous forme oxydee, pour &re ensuite directe-
Fig. 2. Analyse par chromatographie du pic de peroxydase iodke isolk duns les conditions de la Fig.1. Colonne de
Sephadex G-25 maintenue Q 4”
ment transferee au substrat proteique (P) selon le schema suivant :
+ I + P E C--1 E-I-+ E-I oxydb ---+ E + P-I (complexe intermediaire) .
Pour verifier cette hypothitse, nous avons realis6 l’ex- perience ditcrite dans le paragraphe suivant.
Mise e n hidence d’un complexe intermdddiaire peroxydase-iode pur double murpuuge successi f avec
deux isotopes de l’iode La peroxydase est prbincubee en presence du
systitme gbnbrateur de H,O, et d’iodure marquB par un premier isotope de l’iocle, 1311-. Au bout de 3 B 10 min, selon les essais, de la thyroglobuline, l’ac- cepteur protitique, et un deuxiitme isotope de l’iode, 125I--, sont ajoutbs simultanbment et l’incubation est poursuivie pendant 20 min. Au bout de ce temps le milieu reactionnel est analys6. L’iodure en ex& (marque par 1311 et lZ5I) est s6parb des enzymes et de la thyroglobuline par chromatographie sur Sepha- dex 6-25. La thyroglobuline, exclue de la colonne en m6me temps que la peroxydase et la glucose oxydase, est separbe de ces dernibres par ultracentrifugation en gradient de saccharose. En determinant le rap- port 1311/1251 de l’iodure, de la thyroglobuline et du
milieu ritactionnel total b la fin de la rbaction, on montre que le rapport des deux isotopes est toujours plus &levit dans la thyroglobuline que dans l’iodure ou dans le milieu reactionnel total (Tableau). Ces rbsultats concordent avec l’hypothhse de la formation d’un complexe intermediaire peroxydase-iode oxyde, form6 au cours de la preincubation et B partir duquel l’haloghe est transfitrit directement & l’accepteur protitique .
Tableau. Double marquwge de la thyroglobuline par lZ5I et 1311
250 pg de peroxydase de raifort, 10 pg de glucose oxydase et 37,5 pg de glucose sont pr6incubks pendant 7 minutes en presence de 15 pg d’IK marque par lalI; on introduit alors 2 m g de thyroglobuline et 1OOpg d’IK marque par lz5I; l’incubation est poursuivie pendant 20 minutes. L’iodure libre en exchs est skpar6 par filtration sup Sephadex G-25; la thyroglobuline est isolke par ultracentrifugation en gradient de saccharose. On etablit le rapport 1311/1251 dans l’iodure libre en exchs ou dans le milieu d‘incubation total, dans la thyroglobuline totale c t dans les fractions de thyroglobuline sQparQes par ultracentrifugation. Ces divers rapports sont
calculks par rkfhrence Q ceux de l’iodure ou du milieu d’incubation total, arbitrairement ramen& Q 100
Rapport 131~/ ie51 dsns
les thyroglobulines separees par ultra-
le milieu 1% t h w - centrifugation cn Essai I- total globulinc gradient de saccllarose
Molecules DIol6cules ((rapides @ nlentes s
totmale
- 1 100 - 115 - 2 100 - 3 100 - 136 173 126 4 - 100 110 136 99 5 - 100 159 208 139 6 - 100 119 138 105
- - 117
Analyse de la thyroglobuline doublement marquke obtenue
La thyroglobuline obtenue itu cours des experien- ces de double marquage decrites ci-dessus est analysite par ultracentrifugation en gradient de saccharose (Fig. 3).
On observe la presence de deux pics doublement marques par les deux isotopes. Le premier, de faible coefficient de s6dimentation) correspond B la per- oxydase iod6e. Le second, de coefficient de &dimen- tation bgal b 19 S environ, est celui de la thyro- globuline. Le pic de peroxydase contient relative- ment beaucoup plus de 1311 que celui de thyro- globuline. Ce resultat s’explique aisement par le fait qu’au cours de la prhincubation la peroxydase a joub simultanement le r61e d’enzyme et de substrat.
Le pic de thyroglobuline est aussi doublement marque. On constate que le rapport l3lI/1Z5I est d’autant plus grand que les fractions sbdimentent plus vite (Fig.3 et Tableau).

Vol. 5, No. 1, 1968 J. NUNEZ e t J. POMMIER 117
0 10 20 30 40 NUMEROS DES FRACTIONS
Fig. 3. Analyse par ultracentrifugation de la thyroglobuline obtenue au cours d’une expkrience de double marquage. 250 pg de peroxydase de raifort, 10 pg de glucose oxydase et 37,5 pg de glucose sont preincubes pendant 7 min L 37” en presence de 15 pg d‘IK marque par W. On introduit alors 2 mg de thyroglobuline et I00 pg d’IK marque par lZ51. L’incubation est poursuivie pendant 20 min, l’iodure libre en excb est sbpare par filtration sur Sephadex 6-25. La centrifugation est poursuivie pendant 15 h B 35.000 tours/min en gradient de saccharose (100-250 g/l) dans le rotor SW 39 de la cen- trifugeuse Spinco L2. , Radioactivit6 de l3II; x , radio-
activite de 1251; A , rapport 1311/1251
~ . o o n M o o o a - m ~ ~ ~ . ~ ~ I
10 20 30 40 NUMEROS DES FRACTIONS
1
075 C ._ < 7 a
05
025
Fig. 4. Analyse par ultracentrifugation d’un mClange de per- oxydase halogknke et de thyroglobuline. La solution contenait 0,2 ml de peroxydase halogenbe (isolbe par filtration sur Sephadex G-25 dans les conditions de la Fig. 1) correspondant L 0,2 mg d‘enzyme et 1 mg de thyroglobuline; le melange etait maintenu B 0” pendant 10 min. La centrifugation a B t B poursuivie pendant 15 h B 35.000 tours/min en gradient de saccharose (100-250 g/l) dans le rotor SW 29 de la centri- fugeuse Spinco L 2 . 0 , radioactivite de lZSI; x , augmentation
de l’absorbance B 470 mp
Isolernent du cornplexe intermkddiaire peroxydase- halogthe et transfert ?L f ro id de l’halogkne ir. l’accepteur
protkdique De la peroxydase iodee marquee par 1311 et isolbe
B basse temperature (voir partie exphimentale) est
m6langBe pendant 10 min B 0” B de la thyroglobuline. On constate aprbs ultracentrifugation engradient de saccharose que I0 B 25O/, de l’iode initialement present dans la peroxydase halogBnBe sont transfBr6s sur la thyroglobuline (Fig.4).
On ne met pas en Bvidence par chromatographie sur papier d’iodure libre marque par l31I. I1 s’agit donc rBellement d’un transfert direct d’halogbne de l’enzyme halogBnBe B la thyroglobuline.
Pour verifier ce rBsultat, l’experience est r6pBtBe dans des conditions identiques, mais en ajoutant au mblange refroidi le deuxibme isotope de l’iode lZ5I-. On constate qu’il n’est ni fix6 B la peroxydase, ni incorpore dans la thyroglobuline. Pendant le contact entre la peroxydase iodBe et la prothine acceptrice, il n’intervient donc, dans ces conditions, aucune rBaction d’oxydation de l’iodure libre. La presence d’I3lI au niveau de la thyroglobuline pouvait cepen- dant correspondre B une absorption d’iodoperoxy- dase sur l’accepteur proteique, et non Li un transfert d’halogbne. Pour bcarter cette Bventualith, nous avons v6rifiB par ultracentrifugation en gradient de saccharose qu’aucune activiti: peroxydasique vis-8-vis du gaiacol ne peut Btre mesurite au niveau du pic de la thyroglobuline (Fig. 4).
DISCUSSION Dans la suite de reactions peroxydasiques qui
conduisent It l’iodation de proteines par voie enzy- matique, nous avons pu mettre en evidence un com- plexe intermediaire form6 entre l’enzyme et l’halo- gbne.
L’existence de ce complexe a &ti: etablie en pr8- incubant l’enzyme avec un premier isotope de l’iode, l3II--, puis en l’incubant avec lZ5I- et l’accepteur protkique introduits simultanement. On constate que le rapport 1311/1251 de la protBine iodee obtenue est supBrieur, B la fin de l’incubation, B celui de l’iodure libre en excbs. Ce resultat ne peut &re compris que si une fraction d’1311- forme avec l’enzyme, au cours de la preincubation, un complexe intermediaire relativement stable, dans lequel il n’est pas 6chan- geable avec l’iodure libre en excbs present dans le milieu; d’autre part, cette fraction d’I3lI- doit &re cBd6e directement B l’accepteur protbique lorsque celui-ci est introduit dans le milieu. En effet, si cette fraction d’I3lI- etait libBree dans le milieu avant d’btre incorportte dans la proteine, elle s’hchangerait ou se melangerait instantanement avec l’iodure libre ; les rapports l3lI/lZ5I de la prothine marquee et de l’iodure devraient alors Btre identiques.
Le nombre de sites de l’enzyme Atant forcement rbduit, la mise en Bvidence du complexe intermediaire a &tit rendue possible parce que nous avons utilise de 1’1311 dont l’activiti: specifique etait 10 B 15 fois plus &levee que celle de lZ51 et parce que l’enzyme est partiellement inactivee au cours de la preincubation ;

118 J. NIJNEZ et J. POMMIER: Iodation des protCines par voie enzymatique European J. Biocheni.
en l’absence d’accepteur prothique, l’enzyme est son propre substrat. De ce fait, m e fraction seulement d’131I (environ 20 O / o ) retenu par l’enzyme participe B la formation du complexe. Le reste est substitui: sur les ritsidus de tyrosine de la peroxydase.
Lorsque la thyroglobuline doublement marquee, ainsi obtenue, est analysee par ultracentrifugation en gradient de saccharose, on constate que les frac- tions qui sedimentent le plus rapidement contiennent relativement plus d’1311. Ce resultat confirme l’h6t8ro- genhite d’iodation de la thyroglobuline [8-131; on sait que les fractions les plus rapides sont celles dont le contenu en haloghe est le plus BlevB [ 131. I1 semble done logique que les molitcules les plus halogenees et qui ont donc B t B iodees les premieres soient relative- ment plus riches en 1311. Ce resultat est donc &gale- ment en accord avec l’hypothbse de la formation d’un complexe enzyme-iode oxydB et du transfert direct d’halogene de l’enzyme B l’accepteur proteique.
L’isolement du complexe intermediaire enzyme- halogene a && rendu possible en utilisant des con- ditions precises de concentration en iodure, une durhe d’incubation bien determinee et en operant & basse temperature pendant la purification. Lorsque l’on observe ces diverses prheautions, on montre que 20 Of,,
environ de l’iode renferme dans le complexe enzyme- halogene sont transferbs sur la prothine acceptrice. Lorsque le transfert est realis6 en presence d’iodure marque par lZ5I, on constate que ce dernier isotope n’est pas incorpor6. Ces resultats montrent gue I31I est effectivement oxydb, fix6 B I’enzyme et enfin transfer6 directement de l’enzyme B la thyroglo- buline .
La formation d’un complexe intermbdiaire entre la peroxydase de raifort et un donateur d’hydrogene oxydB a &ti! postuke par Mann d8s 1931 [15]:
E + AH, $ E--AH,+ E-AH. + H.
et les arguments experimentaux indirects ont B t B fournis Bgalement, notamment par Hosoya [IS], puis par Brill et Weinryb [17].
Lorsque le donateur d’hydrogene est l’iodure, le complexe enzyme-substrat oxyde devrait contenir l’haloghe sous forme de I. ou de I+, selon qu’un ou deux islectrons sont mis en jeu.
Enfin, un transfert direct d’iode oxyde implique probablement une interaction entre le complexe en- zyme-haloghe et la protitine acceptrice (P) :
ou E + I- + P + E-I oxyd6 - P + E + P-I . Diverses donnhes qui seront publiQes ulterieure-
ment indiquent en effet que la prothine acceptrice forme egalement un complexe intermediaire avec l’enzyme.
Ce travail a b6nBficik d’une subvention du Centre Na- tional de la Recherche Scientifique (Equipe de Recherche No 18) et d’une aide de la DBlkgation Gbn6rale la Recherche Scientifique et Techniquc (Comiti: d0 Biologie MolBculaire. Contrat No 66.00.268).
BIBLIOGRAPHIE 1. Alexander, N. M., J . Biol. Chem. 234 (1959) 1530. 2. Hosoya, R., Kondo, Y., et Ui,N., J . Biochem.52 (1962) 180. 3. De Groot. L. S.. et Davis. A. M.. Endocrinoloau. 70
Y Y ,
(1962) 492. . 4. Klebanoff. S. J.. Yio. C., et Kessler. D.. Biochim. Bio- , ,
phys. Acta, 58 (1662) 58. 5. Nunez, J., Pommier, J., El Hilali, M., et Roche, J., J .
Labelled Compmnds, 1 (1965) 128. 6. Taurog, A., et Howells, E. M., J . Biol. Chem. 241 (1966)
1329. 7. Nunez, J., Mauchamp, J., et Roche, J., Biochim. Bio-
phys. Acta, 86 (1964) 86. 8. Ingbar, Sh., Askonas, B. A., et Work, T. S., Endocrino-
logy, 64 (1959) 110. 9. Roche, J., Nunez, J., et Gruson, M., Compt. Rend. doc.
Biol. 154 (1960) 2194. 10. Ui, N., Tarutani, O., Kondo, Y., et Tamura, H., Nature,
191 (1961) 1199. 11. Robbins, J., J. Biol. Chem. 238 (1963) 182. 12. Bouchilloux, S., Rolland, &I., Torresani, J., Roques, M.,
et Lissitzky, S., Biochim. Biophys. Acta, 93 (1964) 93. 13. Nunez, J., Mauchamp, J., Pommier, J., Circovic, T., et
Roche, J., Bioehem. Biophys. Res. Commun. 23 (1966) 761.
14. Robbins, J., Salvatore, G., Vecchio, G., et Ui, N., Bio- chim. Biophys. Acta, 127 (1966) 101.
15. Mann, P. J. G., Biochem. J . 25 (1931) 918. 16. Hosoya, T., J. Biochem. 47 (1960) 794. 17. Brill, A. S., et Weinryb, T., Biochemistry, 6 (1967) 358.
J. Nunez et J. Pommier Laboratoire de Biochimie GCnCrale et ComparBe du College de France 15 Place Marcellin Bcrthelot, F-75 Paris-5, France


















