Hum. Reprod.-1998-Hamamah-20-30

of 11
-
Upload
yuanda-khan -
Category
Documents
-
view
229 -
download
0
Transcript of Hum. Reprod.-1998-Hamamah-20-30
-
7/25/2019 Hum. Reprod.-1998-Hamamah-20-30
1/11
Role of the ionic environment and internal pH
on sperm activity
Samir Hamamah
1
3
and Jean-Luc Gatti
2
^nite de Biologie de la Reproduction, Departement de Gynecologie-
Obstetrique, Faculte de Medecine, CHU Bretonneau, 37044 Tours cedex,
and
2
URA INRA -CNRS 1291, INRA -Station de Physiologie de la
Reproduction des Mammiferes Domestiques, 37380 Nouzilly, France
3
To whom correspondence should be addressed at: IVF Center, A. Beclere Hospital,
157 rue Porte de la Trivaux, 92141 Clamart, France
In most species, once formed in the testis, spermatozoa are bathed in a fluid
where they remained immobile and with a very low level of metabolism.
This immotile status is understandable in view of the need to preserve the
sperm energy reserve and to decrease the risk of alteration to membranes,
internal structures and biochemical compounds by endogenous oxidizing
agents produced by mitochondrial activity. This quiescent phase can be of
different lengths and finishes when the semen is released into the external
environment where the spermatozoa become motile and metabolically active.
For invertebrates, and some fish, sexual activity is generally seasonal and
fertilization is external. Spermatozoa, once differentiated in the gonad,
remain there completely quiescent until they are released into the external
medium, which is either fresh water or sea water. Dilution of the testicular
fluid surrounding the spermatozoa allows the initiation of motility and
metabolism. In fact, this seminal fluid has an inhibitory effect on sperm
activity. For birds and mammals (including humans), the situation is much
more complex. In these species, sperm production is almost continuous
although for some of them, seasonal variations occur. When spermatozoa
are released from the Sertoli cells, they are rapidly exported from the testis
to the epididymis where the composition of the surrounding medium is
profoundly modified. For most species, the spermatozoa remain immobile
in the lower part of the epididymis, even though they have gained the
capability to be fully motile as shown by dilution in an adequate medium.
In
vivo,motility is activated when the spermatozoa are mixed with secretions
from the different accessory glands during ejaculation. This paper will
review the role played by environm ental factors , such asions,in the activation
of sperm motility and metabolism of different species of invertebrates and
vertebrates. Special attention is given to changes in sperm internal pH, its
regulation and role in the activation of sperm axonemal movement.
Key words:
human/internal pH/mammal/motility/spermatozoa
Introduction
The activation of sperm motility occurs in response to changes in the external
medium. Among the external factors, ions and particularly internal pH (pHj)
20 Europe an Society for Human Reproduc tion and Embry ology Hum an Reproduction Volume 13 Supplement 4 199H
byguestonSeptember17,2015
http://humrep.oxfordjournals.org/
Downloadedfrom
http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/ -
7/25/2019 Hum. Reprod.-1998-Hamamah-20-30
2/11
Sperm internal pH
seem to play a pivotal role in sperm physiology in invertebrates, lower vertebrates
and, to some extent, in mammals. Over the past 20 years, an increasing amount
of data have been collected on the regulation of pH
{
in a large number of cell
types. These studies have been conducted to determine the mechanisms involved
in this pH, regulation. These mechanisms include: plasma membrane Na
+
/H
+
and K
+
/H
+
exchange, NaCl-HCO
3
co-transport, H
+
ATPases, etc. One of the
reasons for the rapid progress in this area is the fact that somatic cells are large
enough to be impaled without damage by microelectrodes during electro-
physiological studies; sperm cells are too small for such studies to be carried out
easily, although several attempts have been made, with variable results. In these
studies, sperm degradation produced by the microelectrode was difficult to
evaluate. The main progress in our understanding of sperm pHj regulation and
of other internal ions, is the result of analysis, either by the accumulation of
radioactive probes or fluorescent indicators. One of the main drawbacks of these
methods is the difficulty of determining the participation of each of the
several compartments forming the spermatozoa (i.e. acrosomal vesicle, nucleus,
mitochondria, cytoplasm) in the measurements. This leads to an average pH
;
, ion
concentration or changes in concentration of the cell popu lation. This last problem
can be partially resolved by using video-microscopy coupled with computer
image analysis, but this type of approach is time-consuming and is not completely
exempt from the effects of compartmentation.
Sea urchin sperm activation and pHj regulation
Sea urchin semen is the oldest and the best demonstrated example of the tight
relationship between external ionic conditions, pH
i5
motility and metabolic
activation (Lillie, 1919). Sea urchin spermatozoa are immotile and show no
oxygen consumption in the coelomic fluid after the induction of spawning. In
the field, their motility as well as their metabolism are turned on after spawning
in sea water, in which spermatozoa can be active for several hours. Under normal
conditions, sperm activation is accompanied by a release of H
+
ions. Measurement
of sperm pHj was carried out using [
31
P]-nuclear magnetic resonance (NMR), by
the fluorescent probe 9 amino-acridine, and also by the distribution of radioactive
probes (Christen
et al,
1982, 1983a,b; Lee
et al,
1983). These studies have
shown that the dilution of spermatozoa in artificial sea water containing sodium,
produces a pH; increase of 0.4-0.5 pH units. The activation of motility as well
as the pHj increase are inhibited when the external media is acidified (pH < 6.5),
lacks sodium ions or when the K
+
concentration is >30 mM (Christen
et al,
1982, 1983a,b; Lee et al, 1983). Under these inhibitory conditions, addition of
a diffusible base-like NH
4
C1 in the external medium produces an activation of
motility and metabolism by raising the pHj over a critical threshold value of 7.5
(Christen
et al,
1982, 1983; Lee
et al,
1983).
Proton release and, as a result, pHj increase, occurs by the activation of a
plasma membrane sodium-proton exchange. This Na
+
/H
+
exchange is controlled
21
byguestonSeptember17,2015
http://humrep.oxfordjournals.org/
Downloadedfrom
http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/ -
7/25/2019 Hum. Reprod.-1998-Hamamah-20-30
3/11
S.Hamamah and J.-L.Gatti
by the plasma membrane potential and blocked by depolarization when the
external K
+
concentration is >30 mM (Christen et al, 1986). In coelom ic fluid,
a high CO
2
level which decreases the pHj and an external concentration of
potassium of ~30 mM may explain why spermatozoa are not activated, even
though a there is a high NaCl concentration (Christen et al., 1983a; Johnson
et al.,
1983). It is interesting to note that, in the presence of sodium and low
potassium, the pHj is maintained at an almost constant value of 7.2-7.3, within
the external pH (pH
e
) range of 6.9-8.3 while, in the absence of sodium, the pHj
increases linearly from 6.1 to 7.3 and the difference between the pH
e
and the
pHj remains almost constant at ~0.8-1.0 pH units (see Table I in Christen
et al,
1982). This suggests that, in the absence of sodium, the protons are distributed
passively and no other pHj regulatory mechanism is functional.
How does pHj act on motility and metabolism? The activity of the outer arm
dynein-ATPase from sea urchin sperm axonemes shows a strict pH dependence.
This key enzyme for flagellar beating is almost inactive at pH 7.2 and its activity
increases at pH 7.4-8.0 (Christen et al, 1982). However, the action of pH on
regulatory mechanisms involving the cAMP pathway and protein phosphorylation
also seems likely. In inactive sea urchin spermatozoa, the ATP level is high and
oxygen consumption is negligible. The turn-on of the dynein-ATPases by pH
produces ADP and then increases mitochondrial activity (Christen et al, 1982,
1983a,b, 1986). Mitochondrial activity then produces H
+
ions which are exported
from the cell by Na
+
/H
+
exchange, and then a compensatory efflux of Na
+
by
the Na
+
/K
+
-ATPase is necessary to maintain a stable internal ionic environment
(Gatti and Christen, 1985).
Motility and metabolism are not the only steps in sea urchin sperm physiology
in which ions are involved. Changes in pHj and calcium influx also play a pivotal
role in the acrosome reaction and in the responses to egg-released factors (for
review, see Ward and
Kopf
1993).
Fish sperm activation
Teleost fish reproduction has also been studied for a long time because of their
econom ic interest, particularly salmonids, and cyprinids (Stoss, 1983). Both species
have external fertilization and spawning occurs in fresh water, but activation of
sperm motility is dependent upon different external factors: for salmonid fishes it
is external ions (H
+
, K
+
and Ca
2 +
), while for cyprinids, osmotic pressure is the
most important factor. O smotic pressure activation is also found in sea water fishes
although interaction w ith external ions has also been suggested.
Trout sperm activation and pH
t
regulation
Trout spermatozoa are immobile in testicular fluid. Under natural conditions,
trout spermatozoa are released into the fresh water where they are active for
< 1 min. Since the spermatozoa are released at the same time as the eggs at a
byguestonSeptember17,2015
http://humrep.oxfordjournals.org/
Downloadedfrom
http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/ -
7/25/2019 Hum. Reprod.-1998-Hamamah-20-30
4/11
Sperm internal pH
sperm/egg ratio of ~10
9
, almost 100% of eggs are fertilized. Meanwhile, for
artificial insemination, simplified media isosmotic to seminal plasma have been
made and, in these media, the presence of high potassium concentrations
(>30 mM) and low pH (1.0 pH units (pHj
-
7/25/2019 Hum. Reprod.-1998-Hamamah-20-30
5/11
S.Hamamah and J.-L.Gatti
flow cytometry (Marian et al., 1997). This study has shown that the pHj of the
spermatozoa increases slightly by 0.2-0.3 pH units at activation by a Na
+
/H
+
-
dependent mechanism which is amiloride sensitive. This Na
+
/H
+
mechanism is
inactivated by high osmolarity and the change in the pHj is not the trigger of
motility, although the value of the pH; can influence the length of the motility
phase. The decrease in pHj observed at dilution may be partially explain by the
production of protons during the utilization of ATP by the axonemal dynein-
ATPase activity (Perchec et al., 1995).
Bird sperm motility activation and pH
regulation
In birds, spermatozoa formed in the testis are expelled continuously in the
epididymis. The spermatozoa stay in the epididymis for ~1 day. Only a small
percentage of the spermatozoa from the upper part of epididymis are able to
move after dilution, while >70% spermatozoa from the lower part are capable
(Clulow and Jones, 1982). Sperm motility after dilution is dependent upon the
temperature: they are immotile at 30C and motile at 40C (Ashizawa
et al.,
1989a,b). This temperature inhibition can be removed by increasing the pH
e
or
increasing the external Ca
2 +
concentration. Measurement of the pH; using a
9 amino-acridine fluorescent dye shows that pH; is dependent upon the pH
e
(Ashizawa
et al.,
1989a,b). At 30C, external pH and pHj are equilibrated for
pH
e
of 6.0-10.0. At 40C, a difference of 0.2-0.3 pH units (pHj
-
7/25/2019 Hum. Reprod.-1998-Hamamah-20-30
6/11
Sperm internal pH
Table I.
Internal pH (pH
;
) of human spermatozoa under different ionic conditions: sodium
(NaM), and potassium (KM) at different external pH (pH
e
). Values are given as mean SD
(data modified from Hamamah
et al,
1996)
PH
6
7.2
7.4
7.8
8.2
NaM
(140 mM Na
+
)
pH,
6.8 0.2
7.0 0.1
7.3 0.1
7.6 0.1
KM
(140 mM K
+
)
pHi
7.2 0.1
7.3 0.1
7.6 0.1
7.9 0.1
indicators (Florman et al, 1989; Vredenburgh-W ilberg and Parrish, 1995; Brook
et al,
1996; Hamamah
et al,
1996; Cross and Razy-Faulkner, 1997), [
31
P]-NMR
(Smith et al, 1985; Robitaille et al, 1987; Hamam ah et al, 1995) and also by
the distribution of a radioactive amine (Gatti
et al,
1993a; Ham amah, 1996).
It has been proposed that inhibition of motility of the spermatozoa of hamster,
rat, bull and dog in the caudal epididym is is due to an acidic pH, maintained by the
presence of lactate (Babcock, 1983; A cott and Carr, 1984; Carr
et al,
1985; Turner
and Reich, 1985). However, the concentration of this compound
in vivo
is not
sufficient by itselftodecrease the pH;. Meanwhile, the pH of the epididymal cauda
fluid is acidic in most species and this pH
e
acts directly on pHj (Mann, 1964; Gatti
et al, 1993a). Rat spermatozoa are immobile in epididymal cauda fluid and their
motility initiation is accompanied by an intracellular alkalization mediated m ay be
by a sodium-proton exchange mechanism (Wonget al, 1981;Wong andLee,1983)
but calcium may also be an important factor (A rmstrong
et al,
1992).
Babcocket al.(1983) showed that anp jalkalization stimulated both metabolism
and motility of epididymal bull spermatozoa. Protein phosphorylation, dependent
upon changes in bull sperm pH, have also been reported (Carr and Acott, 1989).
pH
e
also acts on the mobility of spermatozoa from the epididymal cauda sperm ato-
zoa ram and boar (Gattiet al, 1993b). In both species, spermatozoa are activated
by dilution in an alkaline medium while, after 1 h, a peak of m otility around
pH
e
7-8 is found.
Recently, we analysed the effect of pH
e
on the human sperm motility in a medium
supporting in-vitro fertilization. We observed that motility parameters, in particular
percentage motility, were slightly affected by alkaline pH values (pH
e
= 8.0) after
a long period of incubation. Optimum pH
e
would be ~7.2 (Table I).
Only a few studies have addressed the question of the mechanism implicated
in the regulation of the sperm pH;. In ram spermatozoa, external potassium has
been shown to act on pHj through a complex mechanism involving calcium
(Babcock and Pfeiffer, 1987; Rigoni
et al,
1987). Analysis of the pH
{
of ram
and boar spermatozoa diluted in different ionic conditions (with or without
sodium and potassium ions) has also shown that the pH
;
is strongly dependent
upon the pH
e
and that it increases linearly with pH
e
(Gatti et al, 1993a). In this
study, the presence or absence of sodium in the dilution media had almost no
effect on the pHj value. The slightly more alkaline pHj of ~0.1-0.2 pH units
25
byguestonSeptember17,2015
http://humrep.oxfordjournals.org/
Downloadedfrom
http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/ -
7/25/2019 Hum. Reprod.-1998-Hamamah-20-30
7/11
S.Hamamah
and
J.-L.Gatti
Table II.
Effect of different external pH (pHe) on the percentage of motile ejaculated washed
spermatozoa as a function of time. Motility was evaluated by computer-assisted sperm analysis
(CASA).Values are given as mean SD
(n
= 6)
Time of incubation
pH
e
30 min 150 min pHj
7.2 60 2 1 61 13 6.8
7.4 53 27 55 23 7.0
7.8 65 1 4 49 22 7.3
8.2 55 3 0 43 16 7.6
pH
;
= internal pH.
which was observed in potassium media may be due to the dissipation of the
plasma membrane potential gradient. The same type of measurement was
conducted with ejaculated human spermatozoa (Hamamah
et al.,
1996) and it
was also found that pHj is highly dependent upon pH
e
: a quasi-linear relationship
existing between pH
e
and pH; in the presence of potassium and sodium (Table II).
The pHj values were also slightly lower in presence of sodium than in presence
of potassium; and probably for the same reason. Recently, a study on the
mechanism of pHj regulation was carried out on mouse cauda epididymal
spermatozoa (Zeng
et al.
1996). After dilution, two regulation mechanisms were
found: one appears to be a NaCl/HCO
3
exchange which seems specific to the
sperm cell, the other mechanism, which allows the cell to recover after
alkalinization and acidification, was ion independent and not precisely defined.
These differing results suggest that the mammalian sperm membrane is
permeable to protons, and that large a change in pH
e
produces changes in pHj
that can not be overridden by the pHj regulatory mechanisms which may exist.
In mam mals, ejaculated spermatozoa, w hile motile, are unable to fertilize. They
must undergo a capacitation step that takes placein vivo in the female genital tract
and which is the prelude to the acrosome reaction in the egg zona pellucida (for
review s, see Yanagimachi,1994;Breitbarte?al.,1997).External calciumisrequired
for both capacitation and the acrosome reaction, but several other factors, e.g. the
pH of the medium , and the presence of glucose, external potassium and bicarbonate,
are also involved. Sperm pH
i5
the Na
+
/K
+
-ATPase, a proton ATPase, and Ca-
ATPase are also cellular mechanisms that have been implicated as factors in this
phenomenon (for reviews, see Fraser, 1993; Kopf
et al.,
1994; Breitbart
et al,
1997).
R ecent studies have also suggested that
the
increase of pH during capacitation
is more likely to be an accompanying phenomenon rather than the trigger of this
physiological step (Uguz
et al.,
1994). Progesterone, a component of follicular
fluid, and some ofitsderivatives are able to induce sperm capacitation and prom ote
the acrosome reaction via an influx of calcium (Hyne et al., 1981; Thomas and
Meizel, 1988; Blackmore et al., 1990). The effect of the addition of progesterone
and human follicular fluid (HFF) on sperm pHj was analysed by Hamamah
et al.
(1996). Addition of progesterone, oestradiol-17(3 or 20% HFF to spermatozoa
incubated did not induce any rapid
pH ;
variation. A slight change in pH (~0.2 units)
26
byguestonSeptember17,2015
http://humrep.oxfordjournals.org/
Downloadedfrom
http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/ -
7/25/2019 Hum. Reprod.-1998-Hamamah-20-30
8/11
Sperm internal pH
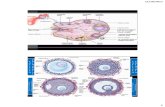
Figure 1. Hypothetical pathway of the sperm internal pH change during the acrosome reaction (AR). See
text for explanation.
occurred with progesterone after 15 min. Florman
et al.
(1989) suggested that
this slow increase in pHj could be interpreted by the recruitment of spermatozoa
undergoing the acrosome reaction . Since the acrosomal vesicle represents an acidic
compartment ofthespermatozoa (Christen
e t al,
1983; Thomas and Meizel, 1988;
Florman
et al.,
1989, 1995). This compartment lowers the overall pH, of the cell.
When this vesicle loses its containment, and hence when its pH equilibrates with
the pH
e
, an increase in sperm pHj occurred (see Figure 1).This is mainly due to the
fact that the acrosome represents a small volume with a large pH gradient (see
1996).
Conclusions
Eukaryotic cells have powerful membranes mechanisms that control the pHj
(exchange of Na
+
/H
+
, K
+
/H
+
, NaCl/HCO
3
etc). Invertebrate and vertebrate
spermatozoa have a different type of mechanism or lack apparent large regulatory
mechanisms to maintain their pHj at a constant value.
In the sea urchin, the only mechanism is a Na
+
/H
+
exchange regulated by the
plasma membrane potential. In trout spermatozoa, pHj is mainly dependent upon
pH
e
.Inthesecells,the plasma membrane potentialisregulated by both the potassium
and proton gradients. It seems that for fish sperm atozoa w ith external fertilization,
and particularly those spawning in fresh water, regulation of the internal milieu
after release is not a priority since the spermatozoa are rapidly altered by the effects
of osmotic pressure, and the egg can be fertilized only for a short period.
7
byguestonSeptember17,2015
http://humrep.oxfordjournals.org/
Downloadedfrom
http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/ -
7/25/2019 Hum. Reprod.-1998-Hamamah-20-30
9/11
S.Hamamah
and
J.-L.Gatti
Mamm alian sperm pHj regulation has not been fully investigated yet. In simple
media, the value of pH
;
is mainly dependent on the value of the pH
e
, suggesting
that the membrane is permeable to protons. No important regulatory mechanisms,
such as N a
+
/H
+
or K
+
/H
+
exchange, have been found. This may be due to the fact
that after ejaculation, mam malian spermatozoa are in a physiological m ilieu where
the pH is well maintained. The different mechanisms described appear to be able
to regulate only to a very sm all degree the different physiological processes before
fertilization or to preserve the sperm pH; during these different steps. Further
research is required to gain a better understanding of sperm ionic regulation, with
a view to obtaining be tter media for short- or long-term preservation and to improve
conditions for the spermatozoa during in-vitro fertilization.
References
Acott, T.S. and Carr, D.W. (1984) Inhibition of bovine spermatozoa by cauda epididymal fluid: II.
Interaction of pH and a quiescence factor. BiolReprod. 30, 926-935.
Armstrong, V.L., Clulow J., Murdoch, R.N. and Jones, R.C. (1994) Intracellular signal transduction
mechanisms of rat epididymal spermatozoa and their relationship to motility and metabolism.
Mol.
Reprod.
Dev.,
38, 77-84.
Ashizawa K., Maeda, S. and Okauchi, K. (1989a) The mecanisms of reversible immobilization of
fowl spermatozoa at body temperature.
J.
Reprod.
Fertil,
86, 271-276.
Ashizawa, K., Suzuki, Y. and Okauchi, K. (1989b) Flagellar movement in demembranated
preparations of ejaculated fowl spermatozoa. J. Reprod.Fe rtil.,
86 ,
263-270.
Ashisawa , K., W hishart, G.J., Na kao, H. and Tsuzuki, Y. (1994) Inhibition of temperature dependent
immobilization of fowl spermatozoa at body temperature by an increased internal pH.
J
Reprod.
Fertil,
101,
593-598.
Babcock, D.F. (1983) Examination of the intracellular ionic environment and of ionophore action
by null point measurement employing the fluoresceine chromophore. J. Biol. Chem.,
258,
6380-6389.
Babcock, D.F., Rufo G.A. and Lardy, H.A. (1983) Potassium-dependent increases in cytosolic pH
stimulate metabolism and motility of mammalian sperm. Proc. Natl. Acad. Sci. USA,
50 ,
1327-1331.
Babcock , D.F. and Pfeiffer, D.R. (1987) Independent elevation of cytosolic C a
+ +
and pH by
voltage dependant and pH sensitive mechanisms.
J. Biol. Chem.,
262, 15041-15047.
Billard, R. (1990) Artificial insemination in fish. In Lamming, G.E. (ed.), Marshall s Physiology
of Reproduction.
Churchill Livingstone, Edinburgh, UK, pp. 870-888.
Billard, R. and Cosson, M.-P. (1992). Some problems related to the assessment of sperm motility.
J. Exp. Zool.,
261, 122-131.
Billard, R., Cosson, J., Perchec, G. and Linhart, O. (1995) Biology of sperm and artificial
reproduction in carp.
Aquaculture,
129, 95-112.
Blackmore, P.F., Beebe, S.J., Danforth, D.R. and Alexander, N. (1990) Progesterone and 17|3-
hydroprogesterone. Novel stimulators of calcium influx in human sperm.
J. Biol. Chem.,
25 ,
1376-1380.
Boitano, S. and Omoto, C. (1991) Membrane hyperpolarization activates trout sperm motility
without an increase in intracellular pH. J. Cell Sci.,
98,
343-349.
Breitbart, H., Rubinstein S. and Lax, Y. (1997) Regulatory mechnisms in acrosomal exocytosis.
Rev.Reprod. 2, 165-174.
Brook, Ph., Lawry, J., Cooke, I.D. and Barratt, Ch.L.R. (1996) Measurement of intracellular pH
in human spermatozoa by flow cytometry with the benzo(Qxanthene dye SNAFL-1: a novel,
single excitation, dual emission, molecular probe.
Mol. Hum.
Reprod.
2,
18-25.
28
byguestonSeptember17,2015
http://humrep.oxfordjournals.org/
Downloadedfrom
http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/ -
7/25/2019 Hum. Reprod.-1998-Hamamah-20-30
10/11
Sperm internal pH
Carr, D.W. and Acott, T.S. (1989) Internal pH regulates bovine sperm motility and protein
phosphorylation. BiolReprod.
41,
907-920.
Carr, D.W., Usselman, M.C. and Acott, T.S. (1985) Effect of pH, lactate, and viscoelastic drag on
sperm motility: a species comparison. BiolReprod.
30 ,
913-925.
Christen, R., Schackmann, R.W. and Shapiro, B.M. (1982) Elevation of the internal pH activates
respiration and motility of sperm of the sea urchin
Strongylocentrotus purpuratus. J. Biol. Chem.,
257, 14881-14890.
Christen, R., Shackmann, R.W., Dalquist, F.W. and Shapiro, B.M. (1983a)
31
P-NMR analysis of
sea urchin sperm activation.
Exp. Cell Res.,
149, 289-294.
Christen, R., Shackmann, R.W. and Shapiro, B.M. (1983b) Metabolism of sea urchin sperm.
Interrelationships between intracellular pH, ATPase activity and mitochondrial respiration.
J.
Biol. Chem.,
258,
5392-5399.
Christen, R., Schackmann, R.W. and Shapiro, B.M. (1986) Ionic regulation of sea urchin sperm
motility, metabolism and fertilizing capacity. J. Physiol. (London),
379,
347-365.
Christen, R., Gatti, J.-L. and Billard, R. (1987) Trout sperm motility. The transient movement of
trout sperm is related to changes in the concentration of ATP following the activation of flagellar
movement.
Eur. J. Biochem.,
166,
667-671.
Clulow, J. and Jones, R.C. (1982) Production, transport, maturation, storage and survival of
spermatozoa in the male Japanese quail,
Coturnix coturnix. J.
Reprod.
Fe rtil.,
64 , 259-266.
Cosson, M.-P, Billard, R. and Letellier, L. (1989) Rise in intracellular Ca
2 +
accompanies the
initiation of trout sperm motility. Cell. Motil. Cytoskel,
14 ,
424-434.
Cross, N.L. and Razy-Faulkner, P. (1997) Control of human sperm intracellular pH by cholesterol
and its relationship to the response of the acrosome to progesterone. Biol.Reprod.
56,
1169-1174.
Dacheux, J.-L., Druart X., Syntin, P. et al. (1995) Secretions and roles of epididymal proteins.
Frontiers Endocrinol,
11,
13-33.
Florman, H.M., Tombes, R.M., First, N.L. and Babcock, D.F. (1989) An adhesion associated
agonist from the zona pellucida activates G protein-promoted elevations of internal C a
+ +
and
pH that mediate mammalian sperm acrosomal exocytosis.
Dev. Biol.,
135,
133-146.
Florman, H.M., Lemos, F.J., Arnoult, C.
et al.
(1995) Exocytotic ion channels in mammalian
sperm. In Fenichel, P. and Parinaud, J. (eds), Human Acrosome Reaction. Vol. 236. INSERM/
John Libbey Eurotext Ltd., Paris, France, pp. 179-189.
Fraser, L.R. (1993) Calcium channels play a pivotal role in the sequence of ionic changes involved
in initiation of mouse sperm acrosomal exocytosis. Mol.Reprod.Dev.,
36,
368-376.
Gatti, J.-L. and Christen, R. (1985) Regulation of internal pH of sea urchin sperm. Role for the
Na/K pump. J. Biol. Chem.,
260,
7599-7602.
Gatti, J.-L. (1987)Role du pH interne et du potentiel de membrane dans Vactivation de la mobilite
et du metabolisme des
spermatozo ides.
These, Universite de Nice.
Gatti, J.-L., Billard, R. and Christen, R. (1990) Ionic regulation of the plasma membrane potential
of rainbow trout
(Salmo gairdneri)
sperm: role in the initiation of motility.
J. Cell. Physiol,
143, 546-544. .
Gatti, J.-L., Chevrier, C , Paquignon, M . and Dache ux, J.L. (1993a) External ionic conditions,
internal pH and motility of ram and boar spermatozoa. J. Reprod.Fertil,
98,
439-449.
Gatti, J.-L., Chevrier, C , Paquignon, M. et al. (1993b) Relation between internal pH (pHj) and
motility of ram and boar spermatozoa. In Baccetti, B. (ed.), Comparative Spermatology 20 Years
After.
Raven Press, New York, USA, pp. 3 69-37 2.
Hamamah, S., Seguin, E, Barthelemy, C.
et al.
(1995) Quantification of energetic patterns of
human spermatozoa during capacitiation process by
31
P magnetic resonance spectroscopy
(
31
P-MR S). In Fenichel,P.and Pa rinaud, J. (eds),
Human Acrosome Reaction.
Vol. 236. INSERM /
John Libbey Eurotext Ltd., Paris, France, pp. 395-396.
Hamamah, S., Magnoux E., Royere, D.
et al.
(1996) Internal pH of human spermatozoa: effect of
ions,
human follicular fluid and progesterone. Mol. Hum. Reprod. 2, 219-224.
Hyne, R.V. and Garbers, D.L. (1981) Requirement of serum factors for capacitation and the
acrosome reaction of guinea pig spermatozoa in buffered medium below pH 7.8.Biol.Reprod.
24,
257-266.
Johnson, C.H., Clapper, D.L., Winkler, M.M.
et al.
(1983) A volatile inhibitor immobilizes sea
uurchin sperm in semen by depressing the intracellular pH. Dev. Biol.,
98,
493-501.
29
byguestonSeptember17,2015
http://humrep.oxfordjournals.org/
Downloadedfrom
http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/ -
7/25/2019 Hum. Reprod.-1998-Hamamah-20-30
11/11
S.Hamamah
and
J.-L.Gatti
Kopf G.S., Kalab, P., Leclerc, P.
et al.
(1994) Signal transduction in mammalian spermatozoa. In
Verhoen, G. and Habenicht, U.-F. (eds),
Molecular and Cellular Endocrinology of the Testis.
Springer-Verlag, Berlin, Germany, pp. 153-183.
Lee, H.C., Johnson, C.H. and Epel, D. (1983) Changes in internal pH associated with initiation of
motility and acrosome reaction of sea urchin sperm.
Dev. Biol.,
95,31-45.
Lillie, F.R. (1919)
Problems of Fertilization.
University of Chicago Press, Chicago, USA.
Mann, T. (1964) The Biochemistry of Semen and ofth Male Reproductive Tract. John Wiley and
Sons, New York, USA.
Marian, T., Krasznai, Z., Balkay, L.
et al.
(1997) Role of extracellular and intracellular pH in Carp
sperm motility and modifications by hyperosmosis of regulation of the Na
+
/H
+
exchanger.
Cytometry,
27, 374-382.
Morisawa, M. (1994) Cell signaling mechanisms for sperm motility. Zool. Sci.,
11,
647-662.
O'Donnell, J.M. (1969) Electronic counting and sizing of mammalian spermatozoa and cytoplasmic
droplets.
J.
Reprod.
Fertil,
19 ,
263-272.
Perchec, G., Jeulin, C , Cosson, J., Andre, F. and Billard, R. (1995) Relationship between sperm
ATP content and motility of carp spermatozoa.
J. Cell Sci.,
108,
747-753.
Perchec-Poupard, G., Gatti, J.-L., Cosson, J.
et al.
(1997) Effects of extracellular environment on
the osmotic signal transduction involved in activation of motility of carp spermatozoa.
J.
Reprod.
Fertil,
110,315-327.
Redondo-M iiller, C , C osson, M.- P, Cosson, J. and Billard, R. (1991) In vitro maturation of the
potential for movement of carp spermatozoa. Mol.Reprod.Dev.,29, 259-270.
Rigoni, F., Dell'Antone, P. and Deana, R. (1987) Evidence for a D pH-driven Ca
2 +
uptake in
EGTA-treated bovine spermatozoa.
Eur. J. Biochem.,
169,
417-422.
Robitaille, P.M., Robitaille, P.A., Martin, PA. and Brown, G.G. (1987) Phosphorus-31 nuclear
magnetic resonance studies of spermatozoa from the boar, ram, goat and bull. Comp. Biochem.
Physiol,
87B, 285-296.
Smith, M.B., Babcock, D.F. and Lardy, H.A. (1985) A
31
P-NMR study of the epididymis and the
epididymal sperm of the bull and hamster.
Biol.
Reprod.
33,
1029-1040.
Stoss J. (1983) Fish gamete preservation and spermatozoan physiology. In Hoar,W.S.,Randall, D.
and Donaldson, E.M. (eds), Fish Physiology. Academic Press, New York, USA, pp. 305-350.
Tanimoto, S. and Morisawa, M. (1988) Roles for potassium and calcium channels in the initiation
of sperm motility in rainbow trout. Dev. Growth Differ,
30,
117-124.
Tanimoto, S., Kudo, Y, Nakasawa, T. and Morisawa, M. (1994) Implication that potassium flux
and increase in intracellular calcium are necessary for the initiation of sperm motility in salmonid
fishes.
Mol.
Reprod.
Dev.,
39,409-414.
Thomas, P. and Meizel, S. (1988) An influx of extracellular calcium is required for initiation of
the human sperm acrosom e reaction induced by hum an follicular fluid. Gamete Res.,20,397-411.
Turner, T.T. and Reich, G.W. (1985) Cauda epididymal sperm motility: comparison among five
species.
Biol.
Reprod.
32, 120-128.
Uguz, C , Vredenburgh, W.L. and Parrish, J.J. (1994) Heparin-induced capacitation but not
intracellular alkalinization of bovine sperm is inhibited by Rp-Adenosine-3,5-cyclic
monophosphorothioate. Biol.Reprod.
51,
1031-1039;
Vredenburgh-Wilberg, W.L. and Parrish J.J. (1995) Intracellular pH of bovine sperm increases
during capacitation.
Mol.
Reprod.
Dev.,
40, 490-502.
Ward, C. and
Kopf
G.S. (1993) Molecular events mediating sperm activation.Dev. B iol,
158,
9-34.
Wong, P.Y.D., Lee, W M . and T sang, A.Y.F. (1981) The effects of extracellular sodium on acid
release and motility initiation in rat caudal epididymal spermatozoa in vitro. Exp. Cell Res.,
131, 97-104.
Yanagimachi, R. (1994) Mammalian fertilization. In Knobil, E., Neill, J.D., Ewing, L.
et al.
(eds),
The Physiology of Reproduction.
Raven Press Ltd, New York, USA, pp. 189-232.
Zeng, Y,Oberdorf J.A. and Florman, H.M. (1996) pH regulation in mouse sperm: identification
of Na
+
-, Cl -, and HCO
3
-dependent and arylminobenzoate-dependent regulatory mechanisms
and characterisation of their roles in sperm capacitation.
Dev. Biol,
173,
510-520.
3
byguestonSeptember17,2015
http://humrep.oxfordjournals.org/
Downloadedfrom
http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/http://humrep.oxfordjournals.org/
![Sophismata Buridani ([Reprod.])](https://static.fdocument.pub/doc/165x107/625d10ffa98da525ef7f60fa/sophismata-buridani-reprod.jpg)