LA MITOSI In biologia, la moltiplicazione equivale alla divisione !

(Laura Condorelli 2018) Pagina 1
Genetica
Struttura DNA
Cromosomi I monomeri del DNA si chiamano
nucleotidi.
–
Le basi azotate sono
classificate in purine (A e G,
costituite da un doppio anello) e
pirimidine (T e C, costituite da un
anello singolo).
.
I legami a idrogeno
.
Il DNA è formato da 2 filamenti avvolti ad elica, tra loro
’ ’
’ ’ à 5’ à ’ 5’ S de
’ à
La doppia elica del DNA si avvolge attorno a strutture
proteiche dette istoni e forma dei nucleosomi.
A loro volta i nucleosomi si avvolgono a spirale formando la
struttura condensata del cromosoma.
Ogni cromosoma
.

(Laura Condorelli 2018) Pagina 2
Nel cariotipo i cromosomi sono ordinati in base
alla grandezza e alla posizione del centromero,
prima compaiono gli autosomi (controllano i
caratteri) e per ultima la
. Ci sono 22
coppie di autosomi e una coppia di cromosomi
sessuali, XX per le femmine, XY per i maschi.
Rispetto al centromero si riconoscono un
braccio più corto (p) e uno più luungo (q)..
La parte finale dei cromosomi prende il nome di telomero. I cromosomi hanno una tipica struttura a bande;
le bande chiare sono ricche di basi AT e contengono pochi geni; le bande scure sono ricche di CG e
contengono molti geni.
I cromosomi sono identici a 2 a 2 per grandezza e posizione del centromero e si dicono omologhi
(co
à . Questi cromosomi, identici per struttura e funzione, si dicono
omologhi.
I cromosomi omologhi sono ereditati uno dal padre e uno dalla madre.
La condizione per cui ogni carattere è controllato da 2 alleli posti sui cromosomi omologhi si dice diploidia
e viene indicata con 2n.
Le cellule della linea germinale (ovociti e spermatozoi) sono necessariamente aploidi n. Cellule uovo e
spermatozoi, infatti, dopo la fecondazione ristabiliscono il corredo cromosomico diploide.
Struttura RNA
I R ’ stituiti da uno zucchero (il ribosio), un
gruppo fosforico e una base azotata. Le basi azotate del RNA sono A,U,C,G. La timina viene cioè sostituita
’U Il RNA, a contrario del DNA, può uscire dal nucleo. Il RNA è a catena singola ed è di 3 tipi
1. m-RNA (messaggero): sta nel nucleo e nel citoplasma, si
chiama anche trascritto ed è la copia del gene, porta il
messaggio dal nucleo al citoplasma
2. r-RNA (ribosomiale): è costituente dei ribosomi (luogo in cui
avviene la sintesi proteica)
3. t-RNA (transfer): è il traduttore, ha una forma a trifoglio,porta
’ ’
aminoacido
anticodone

(Laura Condorelli 2018) Pagina 3
Duplicazione del DNA 1. La duplicazione avviene
subito dopo la divisione cellulare.
Subito dopo che una cellula si è
divisa, infatti, avviene una fase di
crescita in cui vengono sintetizzati
RNA e proteine, poi viene duplicato il
DNA. Ogni cromosoma, infatti, dopo
la divisione cellulare, che sia mitosi o
meiosi, è composto da un solo
E’
ogni cromosoma sia presente nella
cellula in duplice copia. Il processo
avviene in questo modo
1 I ’
Il DNA, a questo punto è diviso nei suoi
due filamenti e viene mantenuto diviso da
altri enzimi (SSBs)
2 Il DNA viene poi ’
DNA polimerasi 3, che è DNA
dipendente (crea un polimero di DNA
utilizzando come stampo il DNA stesso .
’
meccanismo di complementarietà delle
basi azotate.
à
. La doppia elica del DNA è
antiparallela e anche la duplicazione avviene in
modo diverso per le due catene. In particolare:
uno dei due filamenti viene copiato in modo
continuo (leading strand ’ frammenti
di Okazaki (lagging strand). Questo perché
’ III ò
direzi 5’-> ’
’ à ’ H
’
copiata in direzione opposta e necessita dei seguenti enzimi
A. R : R ’ à ’
B. III à ’
C. DNA polimerasi I rimpiazza il primer, sostituendolo con DNA
D. Ligasi unisce i vari frammenti di Okazaki
Al seguente link sono spiegati la struttura e la replicazione del DNA.
https://youtu.be/dKubyIRiN84

(Laura Condorelli 2018) Pagina 4
Sintesi proteica
R – R
e che come la DNA polimerasi sfrutta il meccanismo di complementarietà delle basi
azotate. Non tutto il DNA viene trascritto; si parla di eucromatina (trascritto) ed eterocromatina (non
trascritto).
Rispetto alla DNA polimerasi, però, copia solo il gene ( 1 pezzo di DNA presente su una delle due catene,
sin. locus) e non tutto il DNA. Un gene è definito come un pezzo di DNA che controlla un carattere, ovvero
codifica per una proteina. La RNA polimerasi utilizza U al posto di T. Il meccanismo di trascrizione del gene
viene regolato in ogni cellula da un gene promotore posto a monte del gene da trascrivere. Al gene
promotore si attacca la RNA polimerasi, ma solo se è presente il complesso dei fattori di trascrizione (sono
un complesso di molecole proteiche che si attaccano al DNA. Il primo fattore di trascrizione si lega a
sequenze di basi TATA e si chiama pertanto TATAbox. Questo favorisce il legame degli altri fattori di
trascrizione con il DNA).
Il primo RNA formato si chiama trascritto
primario o pre mRNA e contiene anche la copia
degli introni, che vengono poi tagliati durante il
processo di splicing. Non sempre dallo stesso gene
si forma lo stesso m-RNA, nel senso che non sempre
gli esoni vengono trascritti, oppure in cellule diverse
vengono trascritti esoni diversi, come nel caso
successivo (splicing alternativo). Affinche il m-
RNA sia stabile occorrono altri due processi:
’ cappuccio ’ à 5’ ’
di molteplici basi A (poli A ’ à ’
L’emofilia è una malattia che si verifica in seguito
ad uno splicing anomalo che porta alla non
codificazione del 4^ esone
Al link seguente potete seguire una video lezione
sulla simulazione di laboratorio. Potete svolgere la
simulazione che si trova qui e compilare la seguente
scheda.

(Laura Condorelli 2018) Pagina 5
Ad esempio l’emofilia è
una malattia che si origina
in seguito alla non
trascrizione dell’esone
numero 4 in seguito ad
una delezione della
regione intronica in cui si
attacca lo snRNP
Il trascritto (m–RNA) si porta nel citoplasma,
prende contatto con la sub-unità piccola del
ribosoma, successivamente si lega anche la
sub-unità grande.
Nel ribosoma si riconoscono 2 siti: sito P e sito
A. Nella traduzione interviene anche il t-RNA
(traduttore, transfer). Esso ha una forma a
’
una tripletta complementare al codone, in alto
’
corrispondenza (tra codoni e aminoacidi) è
fissata dal codice genetico.
Nel sito P è esposto il primo codone, corrispondente a metionina per tutti gli esseri viventi.
Il secondo codone è esposto nel sito A.
Arriva il t- R ’ ’
’

(Laura Condorelli 2018) Pagina 6
A questo punto il
ribosoma si sposta
di una posizione,
nel sito P compare
il terzo codone e la
catena
aminoacidica si
allunga.
’
ha legato
’
glicina alla
fenilalanina il
ribosoma si sposta
ancora. Nel sito P
compare il codone
CCU e nel sito A
AAU,
corrispondente
’
asparagina.
Ancora una volta il
ribosoma si sposta,
P ’
codone AAU e nel
sito A compare
GAU.
’
acido aspartico.
Il processo
continua fino a
che nel sito P non
compare una
delle 3 triplette
non senso
(UAA,UAG,
UGA) . E’
segno che la
sintesi proteica è
finita.
Una video animazione è visibile l seguente link
https://youtu.be/fOl7lrNuOnk
La stessa catena di m-RNA viene contemporaneamente letta da più ribosomi (poliribosoma) e si formano più
proteine.
A questo processo partecipano dunque tutti e 3 i tipi di RNA, messaggero, transfer, ribosomiale.

(Laura Condorelli 2018) Pagina 7
Codice genetico Il linguaggio degli acidi nucleici è un linguaggio di 4 basi azotate.
I ’ ’
proteina, formata da varie combinazioni dei 20 aminoacidi.
Il linguaggio delle proteine, quindi, utilizza 20 aminoacidi, quello degli acidi nucleici utilizza 4 basi azotate.
Il codice genetico esprime la corrispondenza tra basi azotate e aminoacidi.
Le combinazioni di 4 basi azotate, prese a 3 a 3, sono 64, molte più dei venti aminoacidi necessari per le
proteine.
Il codice genetico è quindi ridondante (contiene più informazioni di quelle necessarie). Il codice genetico
esprime la corrispondenza tra codoni (triplette di basi azotate poste sul m-RNA) e aminoacidi.
I ’ U U U U A.
’ ò
.
I codoni sono triplette sul m –RNA.
Gli anticodoni sono triplette sul t –RNA.
Il t- R ’ ’

(Laura Condorelli 2018) Pagina 8
Regolazione genica 1) Nei procarioti
I geni sono in qualche modo sottoposti a feedb k ’ :
possiamo avere geni strutturali (sono quelli che codificano per proteine o altro) e geni regolatori. Tra questi
esistono promotori e operatori, che sono siti di legame per proteine regolatrici, le quali si chiamano
repressori.
Operone lac (inducibile): geni per la
scissione da lattosio a glucosio+galattosio
(scissione di un unico legame). Ci sono 3
’
β ).
Promotore: è il sito attacco per RNA
polimerasi II
Operatore: lega il repressore (proteina che
quando è legata al sito operatore
impedisce ala RNA polimerasi di copiare,
perché la blocca).
Regolatore: codifica per il repressore.
In assenza di lattosio (induttore) la
proteina repressore è legata al gene
operatore e quindi la trascrizione dei geni
strutturali non può avvenire.
S ’
induttore, legandosi al repressore e di
fatto, inattivandolo, in modo che la RNA
polimerasi possa copiare i geni strutturali.
Esperimento su PhET
Operone trp (reprimibile): trp è un a.a.
prodotto da 5 geni strutturali. Se trp è
presente la cellula non lo sintetizza. In
questo caso il repressore, per essere
attivo, deve legarsi ad un corepressore
(che è proprio il trp).
2) Negli eucarioti
Hanno maggior quantità di DNA (geni per comunicazione tra cellule, per differenziamento cellulare, per
proteine che uniscono cellule di tessuti). Il DNA viene normalmente suddiviso in eucromatina (viene
trascritto) ed eterocromatina (di norma non trascritto). Le sequenze geniche comprendono gli esoni e le
sequenze correlate (introni, geni promotori, pseudogeni [simili ai geni, ma non codificanti, risultato
’ voluzione], frammenti genici). Il DNA non genico è suddiviso in sequenze presenti in unica copia, ad
esempio gli SNPs (single nucleotide polimorphism, si trovano anche in sequenze codificanti) che
differiscono per un solo nucleotide.

(Laura Condorelli 2018) Pagina 9

(Laura Condorelli 2018) Pagina 10
Sequenze ripetute in tandem (satelliti, ministaelliti, microsatelliti): presenti in più di una copia, sono
piccole e normalmente occupano posizioni specifiche sul cromosoma. Possono essere quindi ricercate ed
utilizzate per esami sul DNA.
satelliti: ’ terocromatina pericentrica, cioè posizionata vicino al centromero.
minisatelliti che servono a stabilizzare DNA nelle vicinanze dei telomeri (servono ad evitare
’ ). Sono costituiti da sequenze ripetute di
esanucleotidi TTAGGC che servono a proteggere i cromosomi. Altri minisatelliti sono le sequenze
altamente variabili VNTR (variable number tandem repeats) che sono altamente polimorfiche e
hanno un core GGGCAGGAXG in sequenze non codificanti, usate nel fingerprinting (per studi
forensi, medici, evoluzionistici
microsatelliti sono numerosi e con un numero di ripetizioni che varia moltissimo (fino a1000x->alto
polimorfismo). Possono essere mono, di (ATATAT) o trinucleotidici (GAT-GAT-GAT), come i
STR (short tandem repeats), che vengono utilizzati come marcatori molecolari ’
(ogni individuo ha un numero variabile di S R ’
altre sequenze poco ripetute (da 10 a 1000x): servono per produrre t-RNA e m-RNA
trasposomi: ’ à -incolla o copia e incolla
Geni interrotti (esoni e introni) ogni esone
codifica per un dominio (parte di proteina). Il
trascritto primario si chiama anche pre-mRNA
subisce il processo di splicing ad opera di
riboproteine nucleari o snurp (SnRNP). La
stabilizzazione del trascritto serve a proteggerlo
da enzimi idrolitici prevede due processi:
Aggiunta cappuccio
guaninico
Aggiunta sequenze
poliA
Più sequenze regolatrici Ogni gene inizia con un promotore e termina con un terminatore (da non
’
come gli operoni, in cui i geni codificatori sono attaccati tra loro, negli eucarioti sono sparsi nel cromosoma
e quindi devono condividere le stesse sequenze di controllo. RNA polimerasi non riconosce il gene
promotore, ma necessita di altre proteine, che formano il complesso di trascrizione. RNA polimerasi è
inoltre di 3 tipi
II. trascrive geni codificanti I. trascrive i geni per r-RNA III. trascrive i geni per t-RNA
Famiglie geniche: solo pochi geni sono presenti in copia singola, sono cioè aploidi. Al contrario, la maggior
parte di essi ha copie diverse (i geni per gli anticorpi sono tantissimi, ad esempio). Le famiglie geniche sono
le copie di uno stesso gene che hanno subìto mutazioni diverse. Quelli che non codificano (perché hanno
perso il promotore oppure manca il sito di riconoscimento per lo splicing) si chiamano pseudogeni.
Meccanismi di regolazione che agiscono prima della trascrizione: sono legati alla struttura condensata del
DNA in nucleosomi (alcune parti del DNA non si despiralizzano [eterocromatina] e quindi non vengono
copiate). Si parla di rimodellamento del DNA per indicare la despiralizzazione delle parti che vengono
trascritte [eucromatina]. Normalmente nelle fenmine il 75% di uno dei due cromosomi X (diploide) è
eterocromatina e si chiama corpo di Barr. Nella sindrome di Klinefelter una delle due X è un corpo di Barr.

(Laura Condorelli 2018) Pagina 11
Nel gatto il fenotipo a tartaruga alcune cellule hanno attivo il cromosoma X che porta il gene rosso (per il
manto del pelo), altre hanno attivo il gene nero.
Geni costitutivi o housekeeping: ’ r enzimi
della glicolisi). La maggior parte dei geni, al contrario, viene attivata o soppressa.
Come già detto, RNA polimerasi non
riconosce il gene promotore (che è
formato da almeno 3 parti: legame della
RNA polimerasi, legame per il fattore di
trascrizione, legame della proteina
regolatrice), ma necessita di altre proteine,
dette fattori di trascrizione. Queste
proteine non sono normalmente legate ai
siti di legame del promotore, ma possono
essere messe a contatto coi siti di legame
specifici se attivati da proteine attivatrici
attaccate a sequenze intensificatori o
enhancer (oppure silenziatori, detti anche
silencer ai quali invece si legano dei
repressori proteici). Un modello prevede
che il DNA si ripieghi su se stesso
portando le proteine specifiche (fattori
generali della trascrizione) a contatto col
promotore (sito attacco RNA polimerasi).
Splicing alternativo: in cui anche alcuni esoni, a seconda del tessuto, vengono tagliati insieme agli introni,
dando luogo a proteine leggermente diverse in tessuti diversi (vedi tropomiosina nel muscolo scheletrico,
tessuto connettivo, fegato e cervello).
Anche il processo di traduzione può essere regolato, così come può essere regolata la longevità della proteina
(prima opportunamente marcata con una piccola proteina chiamata ubiquitina). Molte altre molecole di
ubiquitina si aggiungono e il complesso proteina-poliubiquitina viene inserito in un proteasoma che la
demolisce. HPV fa demolire in questo modo la proteina p53 che serve ad inibire la divisione cellulare e
produce cancro della cervice.
Ciclo cellulare
Tra due successive divisioni cellulari (mitosi o
meiosi, seguite da citodieresi, ovvero la divisione
del citoplasma) la cellula è in interfase. Questa
avviene in 3 distinti periodi. La fase più lunga è la
G1, durante la quale la cellula sintetizza moltissime
molecole, si accresce. Prima della divisione
successiva, la cellula duplica il DNA (fase S). Nella
fase G2 la cellula prepara gli organelli, gli enzimi
che serviranno alla divisione

(Laura Condorelli 2018) Pagina 12
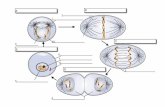
Divisioni cellulari
Mitosi Profase:
Scompare la membrana
nucleare.
I cromosomi si spiralizzano (si
condensano, si avvolgono a
gomitolo). Questo succede
perché la struttura condensata
del cromosoma permette alo
stesso di muoversi. Al
contrario, durante la sintesi
proteica e durante la
duplicazione del DNA, è
necessario che lo stesso sia
despiralizzato.
I centrioli, già duplici e disposti
ai poli opposti della cellula,
emettono le fibre che formano
’
Metafase:
I cromosomi si dispongono
sulla piastra equatoriale.
Le fibre del fuso mitotico
prendono contatto coi
centromeri.
Anafase:
Si separano i cromatidi e vanno
ai poli opposti della cellula.

(Laura Condorelli 2018) Pagina 13
Telofase
I cromosomi si despiralizzano,
ricompare la membrana
nucleare.
Al seguente link vedete una animazione video
https://youtu.be/ofjyw7ARP1c
Meiosi Consiste in 2 divisioni successive senza che ci sia duplicazione del DNA.
.
La meiosi 2 è equazionale e simile alla mitosi.
Profase 1:
Scompare la membrana
nucleare.
I cromosomi si
spiralizzano.
I centrioli, duplici,
emettono le fibre che
’
Metafase 1:
I cromosomi omologhi
si dispongono sulla
piastra equatoriale (vi
sono a questo punto 4
cromatidi affiancati, si
chiama fase delle
tetradi, può avvenire il
crossing-over).
Le fibre del fuso
mitotico prendono
contatto coi centromeri.

(Laura Condorelli 2018) Pagina 14
Anafase 1:
Si separano i
cromosomi omologhi e
vanno ai poli opposti
della cellula.
Telofase1 profase 2
Sono gli stessi processi
della telofase mitotica,
solo che non vengono
portati a termine,
giacchè inizia subito la
profase 2.
. Metafase 2:
I cromatidi si
dispongono sulla piastra
equatoriale.
Le fibre del fuso
mitotico prendono
contatto coi centromeri.
. Anafase 2:
Si separano i cromatidi
e vanno ai poli opposti
della cellula.
Telofase 2
I cromosomi si
despiralizzano,
ricompare la membrana
nucleare. (si veda il
disegno sotto)
Al seguente link vedete una animazione video
https://youtu.be/nMEyeKQClqI

(Laura Condorelli 2018) Pagina 15
Crossing over Avviene durante la metafase 1.
Consiste nello scambio tra
cromatidi interni.
Aumenta la variabilità genetica
delle popolazioni.
.

(Laura Condorelli 2018) Pagina 16
Genetica mendeliana Mendel è un monaco e conduce i suoi esperimenti su piante di pisello.
Si tratta di linee pure, sono cioè individui o
Mendel analizza essenzialmente 2 caratteri : colore del seme e forma del seme.
Si osserva che alcuni caratteri (dominanti) non saltano generazioni, altri (recessivi) saltano generazioni.
I ’
Nella figura sotto, ad esempi ’
I ’
degli alleli dominanti.
Ogni gene controlla un carattere ed occupa una
posizione specifica del cromosoma (e ovviamente
del suo omologo), tanto più che si usa il termine
locus (dal latino-luogo, posizione) come sinonimo
del gene.
Per comprendere le leggi di Mendel è utili imparare ad utilizzare il quadrato di Punnett: consiste in uno
schema; genotipi maschile e femminile sono posti nel riquadro in alto a destra, in alto e a sinistra della
tabella sono invece i gameti maschile e femminile rispettivamente e dentro lo schema vi sono i genotipi degli
zigoti, ovvero dei figli della coppia presa in esame

(Laura Condorelli 2018) Pagina 17
Prima e seconda legge di Mendel Prima legge di Mendel: legge dell’uniformità degli ibridi Incrociando tra loro 2 individui appartenenti a 2 linee pure (geni omozigoti, ovvero alleli uguali) e che
differiscono per un carattere (il colore del seme, giallo o verde), si osserva che gli ibridi di prima
generazione sono tutti uguali tra loro e manifestano il carattere dominante giallo. I due genitori hanno il
seguente genotipo
genotipoGG fenotipo: seme giallo
genotipo vv fenotipo: seme verde
Attraverso la meiosi i
gameti si separano, perciò
il corredo cromosomico
sarà n. Il corredo
cromosomico diploide 2n
viene ristabilito dopo la
fecondazione.
Col quadrato di Punnett lo schema sopra è
evidenziato in modo più semplice.
I figli saranno tutti uguali tra loro, i rapporti
genotipici saranno 4/4 Gv e il fenotipo sarà per tutti
giallo (carattere dominante).
Seconda legge di Mendel: legge della segregazione In seconda generazione ricompare il
carattere che sembrava scomparso in F1 e
che è detto recessivo, in proporzione
fenotipica 3.1.
Nel quadrato di Punnet è importante scrivere
ai margini della colonna la sigla dei gameti
aploidi.
Rapporti genotipici: 1(GG):2(Gv):1(vv)
Rapporti fenotipici: 3G:1v

(Laura Condorelli 2018) Pagina 18
Dominanza incompleta Si riscontra ad esempio nel colore de fiori bocca di
leone, che possono essere rossi, bainchi e rosa. Gli
alleli sono due: B= bianco; R= rosso.
P ’
’
’ à
’ à
’ à
In F1 avremo quindi 4/4 di individui rosa.
Rapport genotipici: 4/4 RB
Rapport fenotipici: 4/4 rosa
Incrociando due individui della F1 osserviamo che i
rapporti genotipici sono sempre quelli della 2^ legge
di Mendel
Rapporti genotipici: 1(RR):2(RB):1(RR)
I rapporti fenotipici ancora una volta saranno uguali
a quelli genotipici.
Rapporti fenotipici: 1(rosso):2(rosa):1(bianco)
Codominanza Si riscontra ad esempio nei gruppi sanguigni, che
possono essere:A-B-0-AB. Sono dovuti a 3 tipi di
alleli (A, B, 0), che codificano per un antigene posto
sulla superificie del globulo ro ’
à ’ ’ B à ’
B ’ 0 à ’
una specie di serratura posta sulla superficie del
globulo rosso. Essa, però, è anche una molecola in
grado di scatenare la risposta immunitaria, cioè la
produzione di anticorpi, che avranno forma
’
entrare nel globulo rosso e ucciderlo.
P ’ ’
’ ’ B che codifica
’ B
Chi possiede il gruppo sanguigno AB ha sia
’ ’ B
E ’ ò
’ - ’ B ò
’ -B. chi ha tutti e due gli
antigeni non avrà anticorpi, mentre chi non ha
antigeni avrà entrambi gli anticorpi.

(Laura Condorelli 2018) Pagina 19
genotipo fenotipo
(gruppo
sanguigno)
antigeni anticorpi dona a riceve da
AA/A0 A A Anti B A, AB A,0
BB/B0 B B Anti A B,AB B,0
AB AB A,B nessuno AB tutti
00 0 nessuno Anti A, Anti B tutti 0
genotipo fenotipo
(fattore Rh)
antigeni anticorpi dona a riceve da
Rh+Rh+/Rh+Rh- Rh+ Rh+ nessuno Rh+ tutti
Rh-Rh- Rh- nessuno Anti Rh+ tutti Rh-
Terza legge di Mendel
legge della segregazione indipendente
Incrociando 2 individui appartenenti a 2 linee pure e che differiscono per 2 caratteri, si osserva che in F2 essi
segregano indipendentemente e compaiono nuove combinazioni di caratteri in proporzioni fenotipiche
9:3:3:1.
N.B. questo vale solo se i geni stanno su cromosomi diversi, ovvero se sono geni indipendenti, altrimenti
segregano come nella seconda legge di Mendel, a meno che non si verifichi il crossing over.

(Laura Condorelli 2018) Pagina 20
In realtà durante la F1 le cose vanno in modo
abbastanza normale. Incrociando due individui,
appartenenti a due linee pure e che differiscono per
due caratteri (che sono indipendenti tra loro, ovvero
stanno su due cromosomi differenti), In F1 gli ibridi
di prima generazione manifesteranno tutti entrambi i
caratteri dominanti.
Il genotipo di entrambi i genitori è questo:
Alla meiosi si separano i cromosomi omologhi,
pertanto il maschio produce solo gameti GL e la
femmina solo vr.
Andando avanti nella seconda generazione le cose si complicano. Bisogna considerare, infatti, che alla
I ’
non è assolutamente detto che tutti i cromosomi paterni si disp ’
Anzi, è molto probabile che ciò non avvenga. Nel disegno sotto, ad esempio, si sono considerate 3 coppie di
à ’ afase
I è casuale e tutte le sei combinazioni di gameti hanno la stessa probabilità di formarsi: Nel quadrato di
Pnnett, pertanto, vanno prese in considerazioni tutte le probabilità.
Con due soli geni e due paia di cromosomi
omologhi le possibili combinazioni sono 4.
Esempio di applicazione di 3^ legge (incrocio tra
due individui della F1 con g . Ogni
individuo potrà produrre gameti GL,Gr,vL,vr.
Ognuno di questi gameti avrà la stessa probabilità
di formarsi. A seguito della fecondazione nella
seconda generazione avremo dunque i seguenti
rapporti fenotipici:
9(GL):3(Gr):3(vL):1(vr)

(Laura Condorelli 2018) Pagina 21

(Laura Condorelli 2018) Pagina 22
Test- cross o reincrocio Serve per stabilire se un individuo con fenotipo dominante
(di cui pertanto non conosciamo il genotipo) abbia genotipo
omozigote o eterozigote. Tutti sanno che se vedo una
persona con gli occhi azzurri, allora so che è ’
omozigote recessivo (perché il carattere recessivo si
manifesta solo allo stato omozigote), ma se ne vedo una
con gli occhi scuri non posso sapere se è omo o eterozigote.
Questo esperimento (usato in laboratorio e non per gli
uomini), consiste ’ ’
S ’
.
Se invece è eterozigote metà degli ibridi sono recessivi,
come risulta dagli schemi a fianco.
Ereditarietà legata al sesso Gli esperimenti di Morgan su Drosophyla
hanno permesso di capire che in molte
specie il cromosoma Y non porta geni, se
non quelli del sesso.
Per i geni portati dal cromosoma X,
pertanto, le femmine sono diploidi, i
maschi aploidi (emi .
Per la specie umana le malattie principali
’
Geni associati Sono geni che stanno sullo stesso cromosoma,
pertanto non segregano indipendentemente alla
meiosi.
Possono ricombinare solo nel caso di crossing over
: : : :0:0: :
.
La frequenza di ricombinazione viene utilizzata per
costruire le mappe cromosomiche, poiché è indice
della distanza tra geni.
Link ad un video su genetica https://youtu.be/DS2aZGrsl2Y
Esercizi Flashquiz; biologia
A. Acidi nucleici; B. duplicazione codice; C. trascrizione-traduzione- D. mutazioni 2018; E. genetica mendeliana; F. genetica completo
Video- lezioni (NON per miei alunni) 1. Acidi nucleici 2. Duplicazione DNA-codice genetico 3. Trascizione-traduzione 4. Mitosi e meiosi 5. Mutazioni 6. Genetica mendeliana 1^ e 2^ legge 7. Genetica mendeliana 3^ legge
Credits: alunna Alyn Marin classe 2G, alunno Pietro Monti, classe 2F, Besta (A.S. 2014-15)