
Formulario de autorización de publicación en el ...
69
Formulario de autorización de publicación en el Repositorio Institucional 1. Identificación del documento y autor Nombre del autor: Mariana Izaguirre Beltrán Correo electrónico: [email protected] Facultad: Facultad de Medicina Veterinaria y Zootecnia Región Veracruz Programa académico: Maestría en Ciencia Animal Título del documento: Evaluación de la actividad antibacteriana del plasma sanguíneo del cocodrilo de pantano (Crocodylus moreletii) sobre cepas de Staphylococcus aureus. Tipo de documento Tesis de licenciatura: Tesis de maestría: ( ) Tesis doctoral: Otro: ( ) ( X ) ( ) Parte de un libro ( ) Objeto de aprendizaje: ( ) Disertación ( ) Otro ( ) Temas del trabajo recepcional: (palabras clave de 5 términos): Resistencia bacteriana, Staphylococcus aureus, Crocodylus moreletii, plasma sanguíneo 2. Autorización de la publicación de la versión electrónica del documento En consonancia con lo dispuesto por El Abogado General de nuestra Universidad, en el Oficio AG/884/2106, en mi carácter de Maestra, doy mi autorización a la Dirección General de Bibliotecas para que este trabajo recepcional sea publicado en el repositorio institucional de la Universidad. 3. Tipo formato del documento: PDF: ( X ) Otro, especifique: Tesis Fecha de entrega:28/10/2020 MVZ Mariana Izaguirre Beltrán Nombre y firma
Transcript of Formulario de autorización de publicación en el ...
Formulario de autorización de publicación en el Repositorio Institucional
1. Identificación del documento y autor
Nombre del autor: Mariana Izaguirre Beltrán
Correo electrónico: [email protected]
Facultad:
Facultad de Medicina Veterinaria y Zootecnia Región Veracruz
Programa académico:
Maestría en Ciencia Animal
Título del documento:
Evaluación de la actividad antibacteriana del plasma sanguíneo del cocodrilo de pantano (Crocodylus
moreletii) sobre cepas de Staphylococcus aureus.
Tipo de documento
Tesis de licenciatura:
Tesis de maestría:
( ) Tesis doctoral:
Otro:
( )
( X ) ( )
Parte de un libro ( ) Objeto de aprendizaje: ( )
Disertación ( ) Otro ( )
Temas del trabajo recepcional: (palabras clave de 5 términos):
Resistencia bacteriana, Staphylococcus aureus, Crocodylus moreletii, plasma sanguíneo
2. Autorización de la publicación de la versión electrónica del documento
En consonancia con lo dispuesto por El Abogado General de nuestra Universidad, en el Oficio AG/884/2106, en mi
carácter de Maestra, doy mi autorización a la Dirección General de Bibliotecas para que este trabajo recepcional
sea publicado en el repositorio institucional de la Universidad.
3. Tipo formato del documento:
PDF: ( X ) Otro,
especifique:
Tesis
Fecha de entrega:28/10/2020
MVZ Mariana Izaguirre Beltrán
Nombre y firma

I
UNIVERSIDAD VERACRUZANA
FACULTAD DE MEDICINA VETERINARIA Y ZOOTECNIA
EVALUACIÓN DE LA ACTIVIDAD ANTIBACTERIANA DEL PLASMA SANGUÍNEO
DEL COCODRILO DE PANTANO (CROCODYLUS MORELETII) SOBRE CEPAS DE
STAPHYLOCOCCUS AUREUS
TESIS
COMO REQUISIO PARCIAL PARA OBTENER EL GRADO DE:
Maestra en Ciencia Animal
PRESENTA:
MVZ. Mariana Izaguirre Beltrán
Directora: Dra. Carolina Barrientos Salcedo
Co-Director: Dr. Alfonso Alexander Aguilera
Asesora: Dra. Concepción del Carmen Ahuja Aguirre
VERACRUZ, VER. DICIEMBRE DE 2020

1

2
DEDICATORIAS
A mi familia, por ser las personas que siempre me impulsan a continuar y nunca
rendirme. Por tenerme paciencia y comprenderme en esos momentos de estrés
constante, mil gracias.

3
INSTITUCIONES DONDE SE DESARROLLÓ EL TRABAJO DE TESIS
Facultad de Medicina Veterinaria y Zootecnia, Universidad Veracruzana.
Laboratorio de Química Médica y Quimiogenómica, Facultad de Bioanálisis,
Universidad Veracruzana
DIRECTORES DE TESIS
Directora: Dra. Carolina Barrientos Salcedo
Co-Director: Alfonso Alexander Aguilera
Asesora: Concepción del Carmen Ahuja Aguirre

4
INSTITUCIÓN BECARIA
Durante el curso de los dos años del programa de Maestría en Ciencia Animal (1 de
Agosto de 2018 – 31 de julio de 2020), la autora de esta tesis contó con el apoyo en
forma de BECA DE MANUTENCIÓN por parte del CONSEJO NACIONAL DE CIENCIA Y
TECNOLOGÍA (CONACyT), con número de becaria (CVU/Becario): 922695

5
RECONOCIMIENTOS
A la Facultad de Medicina Veterinaria y Zootecnia de la Universidad
Veracruzana, por ser la institución que me brindó la oportunidad de la realización del
posgrado.
Al Consejo Nacional de Ciencia y Tecnología por el apoyo financiero durante el
periodo en el que se llevó a cabo el proyecto.
Al Laboratorio de Química Médica y Quimiogenómica y al Cepario
Microbiológico de la Facultad de Bioanálisis de la Universidad Veracruzana, por
brindar las facilidades para que se llevara a cabo todo el trabajo del laboratorio y el
análisis de datos.
A la Dra. Carolina Barrientos Salcedo, por su todo lo que me enseñó y gran apoyo
durante el desarrollo del estudio.
A la Dra. Concepción del Carmen Ahuja Aguirre, por su asesoría, apoyo y por
escucharme siempre que lo necesité durante el tiempo de la maestría.
Al Dr. Alfonso Alexander Aguilera, por todos sus comentarios y críticas constructivas
durante el desarrollo del proyecto.
A las Dras. Patricia Cervantes Acosta y Lorena López De Buen, por todos los
comentarios y ayuda que me dieron en cada uno de los exámenes tutoriales.
Al Rancho El Colibrí de la Antigua, UMA Cacahuatal, en particular al MCA. Sergio
Alberto Viveros Peredo, por las facilidades otorgadas para los animales que se
utilizaron para este estudio.
A la Universidad Nacional de Quilmes (UNQ), Argentina y al equipo del
Laboratorio de Microbiología Molecular, por la oportunidad de hacer una estancia
de investigación en sus instalaciones y por toda la paciencia y los conocimientos
transmitidos.
Al Programa Iberoamericano de Ciencia y Tecnología para el Desarrollo, por el
apoyo económico otorgado durante la estancia de investigación en la UNQ, en Buenos
Aires, Argentina.

6

7
CONTENIDO
INTRODUCCIÓN .............................................................................................. 14
1. ANTECEDENTES .......................................................................................... 16
1.1. Staphylococcus aureus: características y mecanismos de resistencia ............... 16
1.1.1. Estructura antigénica ............................................................................. 17
1.1.2. Enzimas y toxinas de Staphylococcus aureus ............................................. 18
1.1.3. Staphylococcus aureus en la salud humana ............................................... 20
1.2. Resistencia bacteriana ......................................................................... 21
1.3. Generalidades del sistema inmunológico ...................................................... 23
1.3.1. Sistema inmunológico innato (SII) ........................................................... 24
1.3.2. Sistema inmunológico adaptativo (SIA) .................................................... 24
1.4. Sistema inmune de los reptiles ................................................................... 26
1.4.1. Sistema inmune innato (SII) en reptiles ................................................... 26
1.4.1.1. Lisozimas ........................................................................................... 27
1.4.1.2. Proteínas antimicrobianas .................................................................... 27
1.4.1.3. Sistema del complemento .................................................................... 27
1.4.1.4. Leucocitos no específicos ..................................................................... 28
1.4.2. Respuesta inflamatoria ........................................................................... 29
1.5. Actividad de los componentes sanguíneos de los cocodrilianos sobre células de
mamíferos ...................................................................................................... 29
1.6. Actividad antimicrobiana de los componentes sanguíneos de cocodrilianos ....... 30
HIPÓTESIS ..................................................................................................... 32
OBJETIVO GENERAL ........................................................................................ 32
OBJETIVOS ESPECÍFICOS ................................................................................ 32

8
2. MATERIALES Y MÉTODOS ............................................................................. 33
2.1. Características del estudio ......................................................................... 33
2.2. Obtención del plasma sanguíneo de Crocodylus moreletii ............................... 33
2.2.1. Procesamiento de las muestras sanguíneas ............................................... 34
2.2.2. Cuantificación de proteínas del plasma sanguíneo de cocodrilo y humano ..... 35
2.2.3. Electroforesis de proteínas en geles de poliacrilamida en presencia de sds .... 36
2.3. Conteo y evaluación de reactividad de linfocitos por ensayo MTT .................... 39
2.4. Aislamiento y confirmación de las cepas de Staphylococcus aureus ................. 41
2.4.1. Caracterización morfológica .................................................................... 45
2.4.2 Prueba de catalasa en lámina ................................................................... 45
2.4.3. Prueba de coagulasa .............................................................................. 46
2.4.4. Prueba de susceptibilidad y resistencia antimicrobiana ............................... 47
2.5. Evaluación de la actividad antibacteriana por el método de microdilución en caldo
.................................................................................................................... 49
2.6. Análisis estadístico .................................................................................... 50
3. RESULTADOS .............................................................................................. 51
3.1. Ensayo de microdilución en caldo ............................................................... 51
3.2. Ensayo MTT ............................................................................................. 53
4. DISCUSIÓN ................................................................................................ 55
5. CONCLUSIONES .......................................................................................... 58
REFERENCIAS ................................................................................................. 59

9
LISTA DE CUADROS
CUADRO 1. Cuantificación de proteínas de referencia BSA……………………………. 35
CUADRO 2. Valores de las densidades ópticas obtenidas con cada
concentración de plasma de cocodrilo para la cepa de referencia de S. aureus
ATCC 1556…………………………………………………………………………………………………………
50
CUADRO 3. Valores de las densidades ópticas obtenidas con cada
concentración de plasma de cocodrilo para la cepa de referencia del aislado
clínico humano de S. aureus………………………………………………………………………………
51

10
LISTA DE FIGURAS
FIGURA 1. Ejemplares de C. moreletii sacados de los estanques para la
toma de muestra sanguínea……………………………………………………………………….
33
FIGURA 2. Toma de muestra sanguínea de C. moreletii…………………………… 33
FIGURA 3. Plasma sanguíneo de C. moreletii alicuotado…………………………… 34
FIGURA 4. Cuantificación de proteínas de referencia (albúmina sérica
bovina)…………………………………………………………………………………...................
35
FIGURA 5. Electroforesis en gel de poliacrilamida para lectura de
proteínas en plasma de C. moreletii con marcador de peso
molecular…………………………………………………………………………………………………….
37
FIGURA 6. Adición del Ficoll® Paque Plus GE, EE.UU a los tubos con
sangre heparinizada……………………………………………………………………………………
39
FIGURA 7. Linfocitos humanos observados por medio de microscopio
óptico y cámara de Neubauer…………………………………………………………………….
40
FIGURA 8. A) Preparando cajas de Petri de agar sal y manitol; B) Tubos
de cultivo con agar BHI; C) Cajas de cultivo con agar sal y manitol y agar
BHI……………………………………………………………………………………………………………..
42
FIGURA 9. Observación de cepas de Staphylococcus aureus 24 h en
estufa de cultivo………………………………………………………………………………………….
40
FIGURA 10. A) Siembra de S. aureus en tubos de cultivo para su
almacenamiento; B) Cepa de S. aureus en tubo de cultivo con aceite
mineral esterilizado…………………………………………………………………………………….
43
FIGURA 11. A) Staphylococcus aureus en agar sal y manitol; las colonias
doradas (flecha roja) representan la fermentación del manitol. B) S.
aureus en agar sangre; se observa la hemólisis beta en forma de halos
claros alrededor de las colonias………………………………………………………………….
43
FIGURA 12. Morfología colonial de Staphylococcus aureus……………………… 44
FIGURA 13. Realizando prueba de catalasa………………………………………………. 45
FIGURA 14. Formación de la fibrina (flecha roja) en fibrinógeno en
plasma humano………………………………………………………………………………………….
45
FIGURA 15. Halos de inhibición (mm) en cultivo de S. aureus con discos
de novobiocina (28 mm), sulfonamidas/trimetoprim (17 mm) y
vancomicina (19 mm) el halo gris que se observa indica la inhibición
bacteriana que provoca cada antibiótico……………………………………………………
47

11
FIGURA 16. Halos de inhibición (mm) en cultivo de S. aureus con discos
de penicilina (0 mm) y cefoxitín (0 mm); el halo gris que se observa
indica la inhibición bacteriana que provoca cada antibiótico……………………
47
FIGURA 17. Placa con las diferentes diluciones de plasma de cocodrilo…. 49
FIGURA 18. Inhibición bacteriana de la cepa de referencia ATCC 1556 de
S. aureus con plasma de cocodrilo, esto con respecto al control del
antibiótico Trimetoprim/Sulfametoxazol (SXT). No paramétrico, Kruskal-
Wallis test. (P < 0.05)………………………………………………………………………………..
51
FIGURA 19. Inhibición bacteriana del aislado clínico humano de S. aureus
20090127 con plasma de cocodrilo, esto con respecto al control del
antibiótico Trimetoprim/Sulfametoxazol (SXT). No paramétrico, Kruskal-
Wallis test. (P < 0.05)…………………………………………………………………………………
52
FIGURA 20. Porcentaje de viabilidad presente en el plasma sanguíneo de
C. moreletii incubado con células linfocíticas humanas tomadas de sangre
periférica…………………………………………………………………………………………………….
53

12
RESUMEN
Izaguirre Beltrán, Mariana. MCA. Facultad de Medicina Veterinaria y Zootecnia.
Universidad Veracruzana. Septiembre 2020. Evaluación de la actividad
antibacteriana del plasma sanguíneo del cocodrilo de pantano (Crocodylus
moreletii) sobre cepas de Staphylococcus aureus. Directora: Dra. Carolina
Barrientos Salcedo. Co-Director: Dr. Alfonso Alexander Aguilera. Asesora: Dra.
Concepción del Carmen Ahuja Aguirre.
El objetivo del estudio fue evaluar la actividad antibacteriana del plasma sanguíneo de
Crocodylus moreletii sobre aislados clínicos de Staphylococcus aureus, y determinar su
actividad citotóxica sobre cultivos de linfocitos humanos. Se obtuvieron muestras
sanguíneas del seno postoccipital de cocodrilos adultos, de las que se separó el plasma.
Se realizó un ensayo de reducción metabólica (MTT) usando diferentes concentraciones
de plasma crudo de cocodrilo (250-2000 mg/mL) para determinar su capacidad
citotóxica sobre cultivos de linfocitos humanos. A continuación, se realizó el ensayo de
microdilución en caldo en una placa de 96 pocillos para evaluar la actividad
antibacteriana de diversas concentraciones de plasma de cocodrilo (0.2-3.31 mg/mL)
sobre un aislado clínico humano de S. aureus y una cepa de referencia ATCC 1556,
ambos meticilino-resistentes; se dispuso de un pocillo para control de crecimiento con
medio de cultivo y bacterias y se utilizó otro pocillo como control del antibiótico
(trimetoprim con sulfametoxazol, 1.25 y 35.75 µg/mL) y bacterias. Los resultados se
analizaron mediante la prueba de Kruskall-Wallis. El ensayo de MTT mostró que el
plasma sanguíneo crudo de C. moreletii no tuvo efectos citotóxicos; por el contrario, se
observó proliferación celular de hasta 400%. El ensayo de microdilución en caldo
mostró que el plasma sanguíneo de cocodrilo tuvo actividad antibacteriana sobre las
cepas de S. aureus en la concentración más alta que fue 3.31 mg/mL de proteínas
totales respecto al control del antibiótico utilizado (P<0.05). En conclusión, el plasma
sanguíneo del cocodrilo de pantano tuvo actividad antibacteriana sobre S. aureus, y en
lugar de causar citotoxicidad hacia las células humana sanas estimuló su división.

13
ABSTRACT
Izaguirre Beltrán, Mariana. MCA. Facultad de Medicina Veterinaria y Zootecnia.
Universidad Veracruzana. September 2020. Evaluation of antibacterial activity
of swamp crocodile’s (Crocodylus moreletii) blood plasma on Staphylococcus
aureus strains. Director: Dr. Carolina Barrientos Salcedo. Co-Director: Dr.
Alfonso Alexander Aguilera. Advisor Dr. Concepción del Carmen Ahuja Aguirre.
The objective of this study was to evaluate the antibacterial activity of Crocodylus
moreletii blood plasma on clinical isolates of Staphylococcus aureus, and to determine
its cytotoxic activity on human lymphocyte cultures. Blood samples were taken from
the postoccipital sinus of adult crocodiles and the plasma was separated. A metabolic
reduction test (MTT) was carried out where different concentrations of crude crocodile
plasma (250-2000 mg/mL) were used to determine its cytotoxic capacity on human
lymphocyte cultures. Subsequently, the broth microdilution test was performed in a 96-
well plate to evaluate the antibacterial activity of different concentrations of crocodile
plasma (0.2-3.31 mg/mL) on a human clinical isolate of S. aureus and a reference
strain ATCC 1556, both methicillin resistant strains; one well was used for growth
control only with culture medium and bacteria, and another well was used as an
antibiotic control (trimethoprim with sulfamethoxazole, 1.25 and 35.75 µg/mL) and
bacteria. The MTT test showed that crude blood plasma of C. moreletii had no cytotoxic
effects; instead, it stimulated cell proliferation up to 400%. The broth microdilution test
showed antibacterial activity of the crocodile blood plasma on the S. aureus strains in
the highest concentration of plasma, which was 3.31 mg/mL of total proteins, compared
to the control of the antibiotic used (P <0.05). In conclusion, C. moreletii blood plasma
had antibacterial activity on S. aureus, and instead of causing cytotoxicity on healthy
human cells, it stimulated their division.

14
INTRODUCCIÓN
El poder que tienen los antibióticos para contribuir a la muerte bacteriana sin afectar al
huésped humano fue descubierto en 1940 (Waksman, 1947; Belloso, 2009). Sin
embargo, la resistencia a los antibióticos surgió una década después, cuando
Staphylococcus aureus dejó de ser sensible a la penicilina (Chambers y DeLeo, 2009),
dando como resultado un grave problema ocasionado principalmente por el uso
indebido de estos fármacos (OMS, 2001). Esto ha llevado a buscar agentes antibióticos
que resulten efectivos (Kommanee et al., 2012) y que no provoquen efectos
secundarios o toxicidad a largo plazo (Kollef, 2007; van Hal et al., 2012).
Staphylococcus aureus es un patógeno Gram positivo que en humanos es la principal
causa de infecciones septicémicas, osteoarticulares, pleuropulmonares, de piel y tejidos
blandos, endocarditis, e infecciones asociadas a dispositivos (Wertheim et al., 2005;
Tong et al., 2015). Las cepas de S. aureus resistentes a los antibióticos β-lactámicos
se conocen como S. aureus resistentes a la meticilina (MRSA) y se ha observado que
estas cepas son resistentes a múltiples fármacos como aminoglucósidos, macrólidos,
lincosamidas, estreptograminas, tetraciclinas, entre otros (Wang et al. 2008; Kumar et
al. 2010). Hasta hace unas décadas se consideraba que estas cepas se limitaban al
entorno hospitalario; sin embargo, desde principios de 1990 las cepas de MRSA se han
asociado a pacientes que no han tenido contacto previo con algún hospital, por lo que
se les ha denominado como cepas MRSA asociadas a la comunidad (David y Daum,
2010; DeLeo et al., 2010).
Por otro lado, el sistema inmune innato es la primera barrera de defensa del organismo
frente a agentes infecciosos (Hoffmann et al., 1999). Entre sus componentes se
encuentran péptidos antimicrobianos, células natural killer (NK), el sistema de
complemento, y leucocitos no específicos como macrófagos, neutrófilos, basófilos y
eosinófilos (Mondal y Rai, 2001; Merchant et al., 2003; Pata et al., 2011).

15
Los cocodrilianos son animales que habitan en ecosistemas con altas cargas de
patógenos potenciales (Johnston et al., 2010), pero no parecen ser susceptibles a ellos
(Merchant y Britton, 2006). Esto ha llevado a la idea de la existencia de compuestos
antimicrobianos altamente potentes en el sistema inmune de los cocodrilianos, por lo
que se han realizado estudios sobre las propiedades antibióticas de sus componentes
sanguíneos (Pata et al., 2011; Kommanee et al., 2012). Así, se ha demostrado actividad
antimicrobiana del plasma sanguíneo del cocodrilo siamés (Crocodylus siamensis) sobre
cepas de Staphylococcus aureus, Salmonella typhy, Escherichia coli, Pseudomonas
aeruginosa, Staphylococcus epidermidis y Vibrio cholerae (Kommanne et al., 2012).
La resistencia a antibióticos que presentan las cepas MRSA, tanto en humanos como
en animales, es una problemática a nivel mundial que hace necesario buscar
alternativas terapéuticas eficaces, como lo pueden ser ciertos componentes
encontrados en el plasma sanguíneo de los cocodrilianos.

16
1. ANTECEDENTES
1.1. Staphylococcus aureus: características y mecanismos de resistencia
Staphylococcus aureus es una bacteria Gram positiva, de forma esférica de
aproximadamente 1 µm de diámetro, dispuesta en racimos irregulares, no es móvil y
no forma esporas. Esta bacteria crece rápidamente en casi todos los medios
bacteriológicos bajo condiciones aerobias, desarrollándose con más rapidez a una
temperatura de 37 °C. Las colonias de S. aureus suelen ser de color gris a amarillo
dorado profundo (Jawetz et al., 1992).
Este patógeno es resistente a la desecación, aguantando temperaturas de 50 °C
durante 30 minutos y al cloruro de sodio al 9%. La resistencia de S. aureus se presenta
en varias clases: La primera es que en esta bacteria la producción de β lactamasa es
frecuente y está controlada por los plásmidos, lo que hace que este microorganismo
sea resistente a muchas penicilinas (penicilina G, ampicilina, ticarcilina, piperacilina)
(Novick et al., 2001). Los plásmidos son transmitidos mediante transducción de
bacteria a bacteria. La segunda forma se refiere a que la resistencia a la nafcilina,
meticilina y oxacilina es independiente de la producción de la β lactamasa; ésta es
regulada por una serie de genes que se encuentran en una región del cromosoma
denominada casete cromosómico estafilocócico mec (SCCmec por sus siglas en inglés)
(Chambers y DeLeo, 2009). Específicamente el gen mecA codifica una proteína fijadora
de penicilina (PBP2a) que es la que interviene en la resistencia (Archer y Climo, 1994).
Existen diferentes tipos de SCCmec; los tipos I, II y III se relacionan con infecciones
intrahospitalarias y pueden contener genes que codifican la resistencia a otros
antibióticos, mientras que el tipo IV se ha observado principalmente en cepas MRSA
extrahospitalarias (CA-MSRA) (Novick et al., 2001). Una tercera forma de resistencia
presentada por S. aureus se relaciona con un incremento de la síntesis de pared celular
y alteraciones de la misma; en este caso, en Estados Unidos de América (E.U.A.) S.

17
aureus se considera susceptible a la vancomicina si la concentración inhibitoria mínima
(MIC) es ≤2 µg/mL; de susceptibilidad intermedia si la MIC es de 4 a 8 µg/mL; y
resistente si la MIC es ≥16 µg/mL.(Wayne, 2011). En E.U.A. en 2002 se aislaron cepas
de S. aureus resistentes a vancomicina (VRSA), que contenían el gen de la resistencia
a la vancomicina VanA de los enterococos, junto con el gen mecA; este es un problema
importante en todo el mundo (van Hal et al., 2012). Finalmente, la resistencia mediada
por plásmidos a tetraciclinas, eritromicina, aminoglucósidos y otros fármacos es
frecuente en S. aureus (Read y Woods, 2014).
1.1.1. Estructura antigénica
Staphylococcus aureus contiene polisacáridos y proteínas antigénicos entre otras
sustancias importantes en la estructura de la pared celular. El peptidoglucano es un
polímero de polisacárido que contiene subunidades ligadas, lo que le proporciona rigidez
al exoesqueleto de la pared celular; éste es destruido por ácido potente o por la
exposición a lisozimas, desencadena la producción de interleucina-1 y anticuerpos
opsónicos por parte de los monocitos y puede ser quimioatrayente para los leucocitos
polimorfonucleares, y por su actividad endotoxínica activa el sistema del complemento
(Bronner et al., 2004).
Por su parte, la proteína A es un componente de la pared celular de las cepas de S.
aureus que Se encuentra situada en la superficie bacteriana; pertenece a un grupo de
adhesinas denominadas componentes de superficie microbianos que reconocen las
moléculas de matriz adhesiva (MSCRAMMS) (Winn et al., 2006). La adherencia
bacteriana a las células del hospedador es mediada por los componentes antes
mencionados y éstos son factores de virulencia importantes; entonces, la proteína A se
une a la porción Fc de las moléculas de IgG, en donde la porción Fab de la IgG unida a
la proteína A está libre para combinarse con un antígeno específico (Bronner et al.,
2004; Winn et al., 2006).
Algunas cepas de S. aureus tienen cápsulas que inhiben la fagocitosis por los leucocitos
polimorfonucleares a menos que haya anticuerpos específicos presentes; la mayor
parte de las cepas de S. aureus tienen coagulasa, que es un factor de aglutinación en
la superficie de la pared celular que permite que se una de manera no enzimática al
fibrinógeno, generando agregación de estas bacterias (Weems et al., 2006).

18
1.1.2. Enzimas y toxinas de Staphylococcus aureus
Staphylococcus aureus produce enfermedad a través de su capacidad para multiplicarse
y diseminarse rápidamente y por la producción de sustancias extracelulares, como
enzimas y toxinas que pueden funcionar como enzimas (Jonas et al., 2002).
1.1.2.1. Enzimas
Muchas de las enzimas producidas por S. aureus están sujetas al control de los
plásmidos y algunas otras están sujetas al control cromosómico y extracromosómico.
Al respecto, la catalasa es una enzima que convierte el peróxido de hidrógeno en agua
y oxígeno. La prueba de catalasa diferencia a los estafilococos de los estreptococos
(Weems et al., 2006). Por su parte, la coagulasa es una proteína semejante a una
enzima, que coagula el plasma oxalado o citratado uniéndose a la protrombina; en
conjunto inician la polimerización de fibrina. La coagulasa deposita esta fibrina en la
superficie de los estafilococos, alterando su ingestión por las células fagocíticas o su
destrucción dentro de estas células, por lo que la producción de coagulasa es sinónimo
del potencial invasor del patógeno (Chapman et al., 1934; Weems et al., 2006). Por
otro lado, el factor de aglutinación interviene en la adherencia de los microorganismos
al fibrinógeno y a la fibrina, y cuando se mezcla con el plasma el S. aureus forma
racimos que desencadenan una respuesta inmune potente en el hospedador (Weems
et al., 2006).
Existen otras enzimas producidas por S. aureus como la hialuronidasa o factor de
propagación; una estafilocinasa que produce fibrinólisis pero que tiene una acción
mucho más lenta que la estreptocinasa; proteinasas, lipasas y β lactamasa (Rivera et
al., 2007).
1.1.2.2. Toxinas
Dentro del grupo de las exotoxinas producidas por S. aureus están las toxinas α, β, δ
y γ. La toxina α es una proteína heterogénea que ejerce acción sobre una gran gama
de membranas celulares eucariotas, por lo que es considerada una hemolisina potente
(Bronner et al., 2004). La toxina β degrada esfingomielina, lo que la hace tóxica para
muchos tipos de células incluidos los eritrocitos humanos. La toxina δ destruye

19
membranas celulares y participa en las enfermedades diarreicas La hemolisina γ se
conforma por tres proteínas que interactúan con las dos proteínas que componen la
leucocidina de Panton-Valentine, formando seis potenciales toxinas de dos
componentes. La principal función de estas toxinas es producir lisis eficiente de los
leucocitos al causar la formación de poros en las membranas de las células,
incrementando la permeabilidad y desencadenando la liberación masiva de mediadores
inflamatorios, leucotrienos e histaminas (Bronner et al., 2004; Moreillon et al., 2005).
La toxina leucocidina de Panton-Valentine tiene dos componentes que pueden destruir
leucocitos humanos actuando sobre su membrana ; éste es un factor de virulencia
importante en el caso infecciones extrahospitalarias por MRSA (Moreillon et al., 2005).
Las toxinas exfoliativas incluyen a las toxinas epidermolíticas A y B, que son dos
proteínas. La primera es termoestable y es producto de un gen cromosómico, mientras
que la segunda es termolábil y es mediada por plásmidos; ambas producen la
descamación generalizada propia de la epidermólisis estafilocócica aguda, lo que ocurre
cuando disuelven la matriz de mucopolisacáridos de la epidermis, por lo que se
consideran superantígenos (Bronner et al., 2004; Francis et al., 2005).
La toxina-1 del síndrome de choque tóxico (TSST-1) se une a moléculas del complejo
mayor de histocompatibilidad (MHC) clase II estimulando a los linfocitos T y
favoreciendo las diversas manifestaciones del síndrome del choque tóxico. El gen de
TSST-1 se detecta en casi 20% de las cepas de S. aureus incluyendo MSRA (Francis et
al., 2005).
Las enterotoxinas A-E, G-J, K-R y U, V son producidas una o más de ellas, por
aproximadamente el 50% de las cepas de S. aureus producen una o más de ellas; estas
toxinas son termoestables y resistentes a la acción de las enzimas intestinales. Las
enterotoxinas, las toxinas exfoliativas y la TSST-1, se encuentran en un elemento
cromosómico denominado isla de la patogenicidad, que interacciona con elementos
genéticos como bacteriófagos para producir las toxinas (Francis et al., 2005).

20
1.1.3. Staphylococcus aureus en la salud humana
Staphylococcus aureus es una bacteria oportunista y patógena en el humano, de la que
aproximadamente el 30% de la población humana es portadora (Werthein et al., 2005).
Es la causa principal de septicemia, así como de infecciones osteoarticulares, de piel y
tejidos blandos, pleuropulmonares y relacionadas con dispositivos (Wertheim et al.,
2005; Tong et al., 2015). La resistencia a la penicilina surgió en este patógeno en 1950
debido a que sus plásmidos codificaron y adquirieron una betalactamasa,
diseminándose rápidamente; se dice que el 90 % de los aislamientos de S. aureus son
resistentes a este antibiótico (Chambers y DeLeo, 2009). Durante la década de 1950
se creía conocer los mecanismos bacterianos para generar resistencia a antibióticos, y
se pensaba que siempre existiría la posibilidad de sobrepasar este efecto mediante la
síntesis de nuevos antibióticos, lo que llevó en 1960 a la creación de la meticilina, un
antibiótico betalactámico del grupo de las penicilinas; sin embargo, para 1961 se
reportó la aparición de cepas de S. aureus resistentes a la meticilina (MRSA) (Jevons,
1961; Crisóstomo et al., 2001). La resistencia a la meticilina fue mediada por la
adquisición del gen mecA de origen cromosómico; esta resistencia se diseminó
rápidamente en los hospitales de todo el mundo y se considera una epidemia en algunos
países (Arias et al., 2008; Chambers y DeLeo, 2009).
Los hospitales de humanos fueron los primeros en informar la presencia de MRSA como
un agente patógeno nosocomial; estas cepas de S. aureus asociadas a hospitales se
han vuelto resistentes a la mayoría de los antibióticos comunes y su tratamiento
representa un desafío (Katayama et al., 2000; Crisóstomo et al., 2001; Napolitano,
2009). Existen cepas de S. aureus resistente a la meticilina asociado a la comunidad
(CA-MRSA) (Dickinson y Archer, 2000; Crisóstomo et al., 2001; Gradelski et al., 2001;
Ito et al., 2001), detectadas por primera vez en poblaciones de alto riesgo como
consumidores de drogas intravenosas, personas en sanatorios y personas con
enfermedades crónicas, aunque también se han informado casos en niños sanos
(Archer y Climo, 1994; Nishi et al., 1995). En México, en hospitales de tercer nivel la
frecuencia de cepas de SARM es elevada (50- 85 %) (Novales, 2011).
Hasta hace algunos años las cepas MRSA eran susceptibles a gran cantidad de
antibióticos diferentes de los betalactámicos, como la vancomicina, un glucopéptido
que inhibe la síntesis de la pared celular, que ha sido el antibiótico de primera línea en

21
el tratamiento de infecciones asociadas al MRSA en los últimos 40 años (van Hal y
Fowler, 2013). Sin embargo, existen diversas problemáticas asociadas al uso de este
antibiótico, tales como la aparición de cepas de S. aureus con sensibilidad disminuida
a la vancomicina y el desarrollo de insuficiencia renal aguda como efecto secundario
(Kollef, 2007; van Hal et al., 2012). Se ha reportado que la resistencia intermedia de
MRSA sobre la vancomicina sigue aumentando y han comenzado a aparecer cepas
resistentes a gran cantidad de antibióticos (Tenover y Goering, 2009; Boucher et al.,
2010; CDC, 2018).
1.2. Resistencia bacteriana
La resistencia que adquieren las bacterias a los antibióticos se define como el
mecanismo mediante el cual una bacteria disminuye la acción de los agentes
antibióticos; es decir, la bacteria tiene la capacidad de sobrevivir y crecer en presencia
de uno o más antibióticos, haciendo que dicha bacteria continúe causando la infección.
(Jackson et al., 1998). Esta capacidad puede ser intrínseca o adquirida; la primera es
una propiedad específica de las bacterias y su aparición es anterior al uso de
antibióticos. Por su parte la resistencia adquirida se produce cuando las bacterias son
sometidas al estrés que les produce un antibiótico y la adquieren a través de
mutaciones, o bien, por la transmisión de material genético producido por otras
bacterias (Couvalin, 1988).
La era moderna de los antibióticos comenzó con el descubrimiento de la penicilina por
Sir Alexander Fleming en 1928 (Piddock, 2012). Los antibióticos se recetaron por
primera vez para tratar infecciones graves en la década de 1940 (CDC, 2013). Sin
embargo, poco después, la resistencia a éstos se convirtió en un problema clínico
importante, por lo que se desarrollaron nuevos agentes terapéuticos (Sengupta et al.,
2013; Spellberg y Gilbert, 2014). Así, desde finales de la década de 1960 hasta
principios de la década de 1980 la industria farmacéutica introdujo antibióticos nuevos
para resolver el problema de la resistencia, pero al cabo del tiempo dejaron de ser
eficaces, y como resultado, las infecciones bacterianas se convirtieron nuevamente en
una amenaza (Spellberg y Gilbert, 2014). En septiembre de 2016 la Organización de
las Naciones Unidas (ONU) reconoció el aumento mundial de la resistencia bacteriana
a los antibióticos como una amenaza para la salud y el desarrollo humano (ONU, 2016).

22
La resistencia bacteriana a los antibióticos es un proceso biológico natural que puede
darse por transferencia genética o por mutación; así, los genes resistentes codifican
algunos mecanismos por los cuales los microorganismos pueden resistir los efectos
inhibitorios de antibióticos específicos (OMS, 2001). En las bacterias, la herencia de los
genes puede darse en la misma familia y entre diferentes familias, a través de la
transferencia de elementos genéticos móviles conocidos como plásmidos; esta
transferencia horizontal de genes (HGT por sus siglas en inglés: horizontal gene
transfer) permite transmitir resistencia a antibióticos, aún entre diferentes especies de
bacterias. Por su parte, la resistencia a través de mutaciones llega a ocurrir
espontáneamente (Gould y Bal, 2013).
El uso excesivo o inapropiado de antibióticos es la principal causa de resistencia
bacteriana (CDC, 2013; Read y Woods, 2014). Diversos estudios han demostrado una
relación directa entre el consumo de antibióticos y la aparición y diseminación de cepas
bacterianas resistentes; en muchos países, los antibióticos no están regulados y están
disponibles sin prescripción médica (Nature, 2013; Michael et al., 2014). Esta falta de
regulación da como resultado antibióticos de fácil acceso, abundantes y baratos, lo que
promueve su uso excesivo; la posibilidad de comprar dichos productos en línea también
los ha hecho accesibles en países donde están regulados (Michael et al., 2014). De
igual manera, los antibióticos prescritos incorrectamente también contribuyen a la
aparición de bacterias resistentes (CDC, 2013). Se ha demostrado que las indicaciones
de tratamientos, la elección de los antibióticos y la duración del tratamiento son
incorrectos en 30 a 50 % de los casos (CDC, 2013; Luyt et al., 2014). Las
concentraciones sub-inhibitorias y terapéuticas de antibióticos pueden promover el
desarrollo de resistencia a éstos al apoyar las alteraciones genéticas, como los cambios
en la expresión génica, la HGT y la mutagénesis. Los cambios en la expresión génica
inducida por antibióticos pueden aumentar la virulencia, mientras que el aumento de
la mutagénesis y la HGT promueven la resistencia a los antibióticos y su propagación
(Viswanathan, 2014).
Por otro lado, la gran cantidad de antibióticos utilizados en la producción animal es la
causa principal de la acumulación de genes de resistencia dañinos en el reservorio
animal (Aarestrup, 2015). El uso de antibióticos en el ganado, que en muchos países
supera el consumo humano (FDA, 2016), se asocia principalmente con su uso rutinario
como promotores del crecimiento o como sustitutos de bajo costo de medidas de

23
higiene para prevenir infecciones en los animales. La evidencia que vincula la
resistencia bacteriana a antibióticos entre animales y humanos, mediante el consumo
de productos cárnicos, es particularmente fuerte para los patógenos resistentes a las
quinolonas, como Campylobacter spp. y Salmonella spp. (Engberg et al., 2001). Se
estima que la resistencia de las bacterias a los antibióticos aumentará los brotes de
enfermedades infecciosas en el ganado, reduciendo su productividad y perturbando el
comercio internacional (Lowder et al., 2016).
1.3. Generalidades del sistema inmunológico
El sistema inmunológico es el conjunto de estructuras y procesos biológicos en el
interior de un organismo que le permite mantener la homeostasis o equilibrio interno
frente a agresiones externas, ya sean de naturaleza biológica (agentes patógenos) o
físico-químicas (contaminantes o radiaciones), e internas (p. ej., células cancerosas)
(Lozano-Soto et al., 2012).
El sistema inmunológico de los vertebrados tiene como objetivo combatir partículas
extrañas y mantener la integridad de los tejidos mediante sus cuatro funciones
centrales: 1) reconocimiento de células propias y extrañas; 2) discriminación entre
células propias y extrañas; 3) protección de las células propias; y 4) muerte o
eliminación de las partículas extrañas (Tizard, 2013).
El sistema inmunológico reconoce componentes de los microorganismos como
lipopolisacáridos, peptidoglicanos, lipoproteínas, proteínas flagelares y ciertas
secuencias de ADN no metiladas conocidos como patrones moleculares asociados a
patógenos o PAMP (por sus siglas en inglés: patogens associated to molecular
patterns); éstos pueden generar rápidamente una respuesta inmune no específica
denominada inmunidad innata y al mismo tiempo estimular a las células inmunes
específicas desarrollando una respuesta eficaz, duradera y protectora, lo que se conoce
como inmunidad adaptativa (Siroski, 2011).
Así, el sistema inmunológico de los vertebrados está compuesto por dos grandes
subsistemas que se relacionan entre sí: el sistema inmunológico innato y el sistema
inmunológico adaptativo (Medzhitov y Janeway, 2000; Siroski, 2011).

24
1.3.1. Sistema inmunológico innato (SII)
El SII se define como el conjunto de todas las defensas de un organismo que no tienen
memoria inmunológica y ha evolucionado a partir de las asociaciones entre el huésped
y el patógeno. La inmunidad innata está basada en mecanismos de defensa activos y
pasivos menos específicos y poco sofisticados de un hospedador; es considerada la
primera línea de defensa e interviene en la activación de la respuesta inmune
adaptativa, respondiendo rápidamente a la presencia de patógenos (minutos a horas)
(Medzhitov y Janeway, 2000; Siroski, 2011).
Los componentes del SII son las células fagocíticas, como los neutrófilos y macrófagos,
y las células natural killer. Los macrófagos pueden reconocer directamente a los
patógenos por medio de la identificación de los PAMP (Ulevitch y Tobias, 1999), que
son compartidos por un gran número de patógenos debido a que realizan funciones
esenciales en los microorganismos (Medzhitov y Janeway, 1997). El reconocimiento de
los PAMP por receptores específicos que se encuentran sobre los fagocitos lleva a la
absorción de los microorganismos así como a la activación de las células fagocíticas,
resultando en la liberación de proteínas bactericidas y enzimas que se encuentran pre-
almacenadas en gránulos dentro de los neutrófilos y también a la liberación de
quimoquinas y citoquinas a partir de los monocitos y macrófagos. Mientras que los
fagocitos actúan para combatir patógenos que existen libremente en el torrente
circulatorio o los tejidos, las células natural killer combaten a los virus que se alojan en
las células por medio de la destrucción de las células infectadas (Biron, 1997). Debido
a la falta de capacidad del SII para reconocer específicamente a los patógenos y proveer
una inmunidad específica, se desarrolla la respuesta inmune adaptativa (Siroski, 2011).
1.3.2. Sistema inmunológico adaptativo (SIA)
El SIA puede tardar varios días o semanas en activarse completamente y requiere de
exposición previa a un antígeno para generar una respuesta inmunológica completa,
utilizando respuestas tanto humorales como mediadas por células (Coico y Sunshine,
2015). El SIA tiene una gran variedad de receptores que reconocen específicamente a
los antígenos y posee una flexibilidad superior al SII en su adaptación a los cambios
moleculares que presentan los diferentes patógenos, lo que significa que la respuesta

25
se va adaptando para asegurar una respuesta inmune eficaz, generando una memoria
inmunológica de larga duración contra posibles reinfecciones (Andersson et al., 1981).
Esta respuesta inmune, mediada por linfocitos B y T, se desarrolla en alrededor de diez
días y responde a las características específicas expresadas por el patógeno invasor; a
estas células inmunitarias también se le conoce como inmunógenos y funcionan casi
exclusivamente contra el patógeno contra el cual se generaron dada la especificidad de
sus determinantes antigénicos (Robert y Ohta, 2009). Esto sucede cuando en el SII las
células dendríticas y los macrófagos ingieren a los microorganismos y presentan los
antígenos derivados de los mismos a los linfocitos T, lo que desencadena una expansión
clonal de linfocitos T específicos para cada antígeno (Ulevitch et al., 2004). A su vez,
las citoquinas y quimoquinas de las células T regulan la proliferación de células B y la
producción de anticuerpos, así como la actividad de las células del SII. Estas
intercomunicaciones entre el SIA y el SII facilitan una respuesta inmune amplia y
flexible (Siroski, 2011).
En las fases iniciales de la infección, las respuestas inflamatorias se derivan
exclusivamente del SII, mientras que conforme avanza el proceso infeccioso, las
respuestas inflamatorias surgen del efecto cooperativo entre ambos sistemas (Siroski,
2011). En el caso de producirse una reinfección, la respuesta inmune es rápida. Cuando
los anticuerpos disponibles son insuficientes para prevenir la reinfección se activa la
memoria inmunológica, en este caso, las células con memoria se activan y la respuesta
adaptativa se induce en un tiempo más corto y con mayor intensidad que en la infección
primaria, sin que aparezca algún signo o síntoma de infección (Ulevitch et al., 2004).
En general, la respuesta del SIA puede dividirse en dos tipos: 1) inmunidad humoral,
e 2) inmunidad mediada por células (IMC).
1.3.2.1. Inmunidad humoral
Este tipo de inmunidad se refiere a la producción de anticuerpos. La respuesta humoral
es más eficaz contra los patógenos extracelulares. Los anticuerpos específicos cumplen
tres funciones principales en la defensa del hospedador. La primera función es la de
neutralización, que implica la unión de los anticuerpos específicos a adhesinas
bacterianas para impedir la adhesión del patógeno a las células diana o tejidos
(Janeway y Medzhitov, 2002). La segunda función es la de opsonización, mediante la

26
cual los agentes patógenos ubicados en los espacios extracelulares son reconocidos por
los macrófagos y se produce la unión entre el receptor en la superficie del macrófago y
de la molécula del anticuerpo en la superficie del patógeno, que luego es ingerido y
degradado. La tercera función es la de activación del sistema del complemento, que se
refiere a que los componentes específicos del complemento que reconocen los
complejos antígeno-anticuerpo inducen la cascada del complemento que puede resultar
en la muerte directa del patógeno (Schmidt y Colten, 2000).
1.3.2.2. Inmunidad mediada por células (IMC)
Las respuestas de la IMC son más eficaces contra los patógenos intracelulares que
pueden encontrarse dentro de las vesículas fagocíticas o libres en el citoplasma de la
célula hospedera (Siroski, 2011). Los linfocitos T son las células encargadas de esta
respuesta; ésta sucede cuando las células presentadoras de antígenos procesan y
presentan dichos antígenos en su membrana mediante el Complejo Mayor de
Histocompatibilidad (CMH). Los linfocitos T citotóxicos (CD8+) reaccionan con el CMH
I y los linfocitos T colaboradores o helper (CD4+) con el CMH II que son reconocidos
por el receptor T que dichos linfocitos presentan en su membrana. Entonces, los
linfocitos T activan toda la cascada de señales y reacciones que hacen frente a la
infección (Abbas et al., 2002).
1.4. Sistema inmune de los reptiles
Los reptiles son los únicos amniotas ectotérmicos y, por lo tanto, se convierten en un
grupo fundamental para el estudio de la evolución del sistema inmune y del
funcionamiento del sistema inmune en un entorno ecológico (Zimmerman et al., 2010).
1.4.1. Sistema inmune innato (SII) en reptiles
El SII está compuesto por diversas moléculas y células que actúan como una primera
línea de protección no específica contra patógenos e incluye lisozimas, proteínas
antimicrobianas, el sistema del complemento y leucocitos no específicos (Siroski,
2011).

27
1.4.1.1. Lisozimas
Las lisozimas son enzimas que pueden causar la lisis de las bacterias por hidrólisis de
la pared celular y se han aislado de lagartos y varias especies de tortugas (Gayen et
al., 1977; Ingram y Molyneux, 1983; Araki et al., 1998; Thammasirirak et al., 2006).
Al comparar lisozimas de la tortuga china de caparazón blando (Pelodiscus sinensis), la
tortuga asiática de caparazón blando (Amyda cartilagenea) y la tortuga verde (Chelonia
mydas), se observó que, aunque cada una tenía diversos grados de capacidad lítica
frente a varias cepas de bacterias Gram-positivas y Gram-negativas, ninguna fue
efectiva contra las cepas de bacterias de las especies más patógenas para reptiles
(Thammasirirak et al., 2006).
1.4.1.2. Proteínas antimicrobianas
Los reptiles tienen proteínas antimicrobianas que son similares en estructura y función
a las defensinas; son proteínas que tienen un pliegue característico rico en lámina β,
así como seis cisteínas ligadas a disulfuro y se han encontrado en mamíferos y en aves
(Ganz, 2004). La clara de huevo de la tortuga boba (Caretta caretta) carece de lisozima
pero contiene una pequeña proteína catiónica estructural y funcionalmente similar a
una subfamilia de defensinas conocidas como β-defensinas. Estas proteínas muestran
una fuerte actividad antibacteriana contra Escherichia coli y Salmonella typhimurium,
así como actividad antiviral contra el virus Chandipura (Chattopadhyay et al., 2006).
También se encontró un péptido de tipo β-defensina, pelovaterina, en la cáscara de
huevo de la tortuga china de caparazón blando (Lakshminarayanan et al., 2008). Al
igual que en los mamíferos, las proteínas del complemento de cocodrilo pueden actuar
como opsoninas, aumentando la eficacia fagocítica. Aree et al. (2011) demostraron
estos efectos del suero del cocodrilo siamés (Crocodylus siamensis) sobre
Staphylococcus aureus y Escherichia coli. La incubación con suero indujo aumentos de
2 y 4 veces en la captación fagocítica de S. aureus y E. coli, respectivamente.
1.4.1.3. Sistema del complemento
El sistema del complemento consiste en una serie de proteínas que existen en el plasma
y matan a las bacterias invasoras, ya sea a través del proceso de opsonización o por
lisis. En la opsonización las proteínas cubren la membrana bacteriana permitiendo que

28
las bacterias sean reconocidas por los macrófagos, que luego envuelven a las bacterias
a través de la fagocitosis. En la lisis, las proteínas del complemento rompen la
membrana bacteriana y matan a las bacterias invasoras directamente (Seelen et al.,
2005). La cascada del complemento se puede lograr mediante tres vías diferentes: vía
clásica, alternativa y lectina. La vía clásica se activa con inmunoglobulinas IgG e IgM.
La ruta alternativa es activada por moléculas como un virus o un lipopolisacárido en la
superficie de las bacterias y no requiere anticuerpos; se denomina ruta alternativa sólo
porque la vía clásica se descubrió primero (Seelen et al., 2005). La ruta de la lectina
se activa por residuos de manosa de proteínas que se encuentran en la superficie de
las bacterias (Coico y Sunshine, 2015). Se ha confirmado la presencia de vías
alternativas y clásicas en reptiles y aunque no se ha encontrado evidencia directa para
la vía de lectina, se cree que los reptiles también la tienen debido a su presencia en
peces sin mandíbula, tiburones, teleósteos, anfibios, aves y mamíferos (Sunyer y
Lambris, 1998).
1.4.1.4. Leucocitos no específicos
Existe una serie de leucocitos no específicos presentes en los reptiles, incluidos
macrófagos, monocitos, heterófilos, basófilos y eosinófilos. Los monocitos y macrófagos
son células fagocíticas que procesan y presentan antígenos y también liberan citocinas
(Coico y Sunshine, 2015). La temperatura puede afectar la actividad fagocítica de los
macrófagos en los reptiles. Se ha reportado que los niveles más altos de fagocitosis y
citotoxicidad de los macrófagos esplénicos de los lagartos de pared (Hemidactylus
flavivis) ocurrieron a 25 °C, con función macrófaga alterada tanto a temperaturas más
altas como más bajas (Mondal y Rai, 2001). Los heterófilos son equivalentes a los
neutrófilos de mamíferos y pueden variar ampliamente entre especies; éstos ayudan a
suprimir la invasión microbiana y también participan en la respuesta inflamatoria en
reptiles (Montali, 1988). Los basófilos contienen inmunoglobulinas específicas de
antígeno en sus superficies y cuando se activan por un antígeno, se degranulan y
liberan histamina, pero esta liberación depende de la concentración y la temperatura
del antígeno. Por otro lado, los eosinófilos se han descrito en reptiles, pero se sabe
poco acerca de su función en este grupo. En los mamíferos, los eosinófilos juegan un
papel en la defensa contra las infecciones parasitarias produciendo radicales de
peróxido y superóxido, pero no se ha determinado si desempeñen o no un papel similar
en reptiles (Coico y Sunshine, 2015).

29
1.4.2. Respuesta inflamatoria
Una función importante del sistema inmune innato es responder a una lesión o infección
a través de la respuesta inflamatoria. La respuesta varía según el tipo de patógeno
invasor. En reptiles, los patógenos extracelulares inducen la formación de granulomas
heterofílicos, donde los heterófilos se acumulan, degranulan y sufren necrosis, lo que
estimula una fuerte respuesta de macrófagos (Montali, 1988). Por el contrario, en
respuesta a un patógeno intracelular, los reptiles forman un granuloma histiocítico,
formado por los macrófagos que se centralizan y experimentan necrosis. Los
granulomas crónicos pueden formarse a partir de granulomas heterófilos e histiocíticos
(Montali, 1988). A diferencia de los mamíferos, los reptiles no forman un exudado de
pus líquido como parte de la respuesta inflamatoria, sino que forman una masa caseosa
que consiste principalmente en heterófilos degranulados y degenerados (Montali,
1988). Las primeras etapas de la respuesta inflamatoria después de la lesión son
similares a las de los mamíferos, con gran migración de heterófilos a la zona. Sin
embargo, estos heterófilos pueden durar mucho más tiempo en el sitio de la lesión que
los neutrófilos de mamíferos (Tucunduva et al., 2001).
La respuesta inflamatoria está dirigida por una serie de citoquinas y quimiocinas que
incluyen el factor de necrosis tumoral alfa (TNF-alfa), la interleucina-6 (IL-6) y la
interleucina-1 (IL-1) en los mamíferos. Las citoquinas son sustancias de bajo peso
molecular producidas por diferentes tipos celulares que median las interacciones
celulares, mientras que las quimioquinas son un tipo de citocinas pequeñas que dirigen
la migración de células y linfocitos fagocíticos (Coico y Sunshine, 2015). Sin embargo,
la información sobre citoquinas y quimiocinas en reptiles es limitada (Zimmerman et
al., 2010).
1.5. Actividad de los componentes sanguíneos de los cocodrilianos sobre
células de mamíferos
Se han realizado estudios acerca de la citotoxicidad que pudieran tener los
componentes sanguíneos de los cocodrilianos sobre células humanas. Al respecto,
Jangpromma et al. (2016) estudiaron la viabilidad y proliferación celular del plasma y
del suero sanguíneo del C. siamensis sobre una línea celular de keratinocitos humanos
(HaCaT) y reportaron que a una concentración de 3.9-1000 µg/mL no se observaron

30
efectos citotóxicos en un ensayo MTT; además, se descubrió que el plasma y el suero
promueven la proliferación celular en un 114-155% y 119-146% respectivamente. Es
importante señalar que, según la temporada, los cocodrilos con frecuencia pelean entre
sí, lo que a menudo provoca heridas graves en la piel; sin embargo, estas lesiones
parecen sanar rápidamente y están casi libres de infección, aunque estos animales
pueden transportar una gran carga de coliformes fecales de su entorno acuático
(Buthelezi et al., 2012; Van-Hoek, 2014) Considerando estos hechos, parece factible
que el plasma y el suero de C. siamensis contengan una variedad de sustancias con
propiedades antimicrobianas o cicatrizantes (Jangpromma et al. 2016).
Otro estudio realizado por Kommanee et al. (2012), en donde a pesar de que el objetivo
fue determinar la actividad antibacteriana del plasma sanguíneo crudo igualmente del
C. siamensis, se realizó también un ensayo de MTT, donde se probó la citotoxicidad del
plasma sobre células de macrófagos de ratones (RAW264.47) y reportaron que no era
tóxico ya que las células tuvieron una viabilidad del 99%.
1.6. Actividad antimicrobiana de los componentes sanguíneos de cocodrilianos
Los cocodrilos viven con infecciones causadas por bacterias oportunistas, y aunque no
son totalmente inmunes a éstas, normalmente no sufren efectos adversos, lo que
sugiere que la resistencia a estos microorganismos es efectiva (Merchant et al., 2003,
2004). Al ser animales temperamentales, los cocodrilos frecuentemente presentan
heridas graves como consecuencia de disputas sociales (Webb et al., 1987; Piña et al.,
2003), incluyendo la pérdida de extremidades enteras. A pesar de la gravedad de
muchas heridas, generalmente hay pocos signos de infección, lo que ha llevado a
preguntarse cómo es que estos animales a veces sobreviven a heridas tan graves sin
mostrar signos evidentes de enfermedad, particularmente cuando viven en ambientes
que contienen microorganismos que potencialmente pueden causar infección (Merchant
et al., 2003; 2005; Siuntawineti et al., 2004; Merchant y Britton, 2006).
Existen reportes acerca de la eficacia antimicrobiana de algunos componentes
sanguíneos de los cocodrilianos. Siroski et al. (2010) realizaron un estudio en donde se
demostró que el plasma sanguíneo del caimán de hocico ancho (Caiman latirostris)
presentó actividad antibacteriana sobre la cepa de Escherichia coli ATCC 11105,
comparándose con la actividad antibacteriana del plasma de gallo bankiva (Gallus

31
gallus) y de humano, demostrándose que la actividad del plasma del caimán fue
superior al inhibir completamente la proliferación de E. coli. En un estudio similar,
Leelawongtawon et al. (2010) demostraron que el suero fresco y el suero liofilizado del
cocodrilo siamés (Crocodylus siamensis) tiene actividad antimicrobiana sobre cepas de
Klebsiella aerogenes, Escherichia coli, Klebsiella pneumoniae, Salmonella typhimurium,
Pseudomonas aeruginosa, y sobre cepas fúngicas de Cryptococcus neoformans y
Aspergillus niger. Kommanee et al. (2012) demostraron actividad antimicrobiana
significativa del plasma sanguíneo semipurificado del cocodrilo americano sobre cepas
bacterianas humanas como Staphylococcus aureus ATCC 25923, Salmonella typhy
ATCC 11778, Escherichia coli O157:H7, Vibrio cholerae non01, Pseudomonas
aeruginosa, Salmonella typhy y Vibrio cholerae.
Pata et al. (2011) purificaron cuatro péptidos antibacterianos, denominados Leucrocin
I a IV, a partir de extractos de glóbulos blancos de cocodrilo siamés y observaron que
estas leucrocinas exhibieron fuerte actividad antibacteriana sobre Staphylococcus
epidermidis, Salmonella typhy y Vibrio cholerae. Por su parte, Machha et al. (2011)
demostraron la actividad inhibitoria del extracto de leucocitos refinado del cocodrilo
americano (Alligator mississippiensis) sobre aislamientos de patógenos resistentes a
antibóticos como Pseudomonas aeruginosa, Enterococcus faecium y Klebsiella
pneumoniae. Barksdale et al. (2016) observaron fuerte actividad in vitro de péptidos
antimicrobianos catiónicos del plasma de cocodrilo americano contra cepas
de Staphylococcus aureus, Escherichia coli, Pseudomonas aeruginosa y Acinetobacter
baumannii resistentes a múltiples fármacos.
Por otro lado, Prajanban et al. (2017) estudiaron los efectos antimicrobianos de los
nuevos péptidos cOT2 y sOT2 derivados de las ovotranferrinas de Crocodylus siamensis
y demostraron actividad antibacteriana potente sobre Bacillus megaterium, B. pumilus,
E. coli ATCC25922, E. coli O157: H7 y Vibrio cholerae (aislado clínico de Tailandia, con
un rango de CIM 7-32.2 µM). Por otro lado, recientemente Tankrathoj et al. (2018)
identificaron el primer gen de catelicidina de Crocodylus siamensis y el péptido RN15
derivado del dominio de catelina exhibiendo actividad antibacteriana contra bacterias
Gram positivas como Bacillus megaterium y Bacillus subtilis y Gram negativas como E.
coli exhibiendo una concentración inhibitoria mínima CIM baja 12.5 y 25 µg/mL para
las Gram positivas respectivamente; también se observaron cambios en las morfologías
celulares de las bacterias después de estar expuestas al péptido 15 minutos.

32
HIPÓTESIS
El plasma sanguíneo del cocodrilo de pantano (Crocodylus moreletii) presenta actividad
antibacteriana sobre cepas de Staphylococcus aureus.
OBJETIVO GENERAL
Establecer la actividad antibacteriana del plasma sanguíneo del cocodrilo de pantano
(Crocodylus moreletii) sobre cepas de Staphylococcus aureus.
OBJETIVOS ESPECÍFICOS
1 Conocer la actividad antibacteriana del plasma sanguíneo del cocodrilo de
pantano sobre cepas de Staphylococcus aureus con resistencia a la meticilina
de importancia clínica.
2 Determinar la citotoxicidad del plasma sanguíneo del cocodrilo de pantano sobre
cultivos de linfocitos humanos de sangre periférica.

33
2. MATERIALES Y MÉTODOS
2.1. Características del estudio
Se realizó un estudio experimental que se llevó a cabo de febrero de 2019 a marzo de
2020, de manera conjunta en las Facultades de Medicina Veterinaria y Zootecnia y de
Bioanálisis de la región Veracruz en la Universidad Veracruzana. El estudio comprendió
de cuatro fases: 1) Obtención del plasma sanguíneo de C. moreletii; 2) Evaluación de
la reactividad de linfocitos humanos al plasma de cocodrilo; 3) Aislamiento y
confirmación de las cepas de S. aureus y evaluación de susceptibilidad y resistencia
antimicrobiana; y 4) Evaluación de la actividad antibacteriana del plasma sanguíneo de
cocodrilo sobre S. aureus.
2.2. Obtención del plasma sanguíneo de Crocodylus moreletii
Se obtuvieron muestras sanguíneas de 10 ejemplares adultos de cocodrilo de pantano
(Crocodylus moreletii) aparentemente sanos, mantenidos en cautiverio en la Unidad de
Manejo Ambiental (UMA) Cacahuatal (INE/CITES7DGVS-CR-IN-0434-VER/99),
localizada en el municipio de La Antigua, Veracruz.
Los animales fueron sacados de los estanques y se colocaron en jaulas de contención
(Fig. 1). Las muestras se obtuvieron mediante punción del seno venoso postoccipital
utilizando aguja calibre 22 y jeringa desechable de 10 mL, desinfectando previamente
el área con iodopovidona (Fig. 2). La sangre se colocó en tubos de plástico de 10 mL
heparinizados. Las muestras se invirtieron delicadamente de 8 a 10 veces para
homogeneizar el anticoagulante con la sangre; posteriormente fueron trasladadas en
red fría al laboratorio para su procesamiento dentro de las 2-4 h después de recolectada
la muestra.

34
FIGURA 1. Ejemplares de C. moreletii sacados de los estanques para la toma de
muestra sanguínea.
FIGURA 2. Toma de muestra sanguínea de C. moreleti.
2.2.1. Procesamiento de las muestras sanguíneas
Las muestras se mantuvieron durante todo el procesamiento a 4 °C y se centrifugaron
a 1100 g x 10 min para obtener el plasma. Se evitó mezclar o agitar la muestra después

35
de centrifugarlas para evitar la re-suspensión del pellet celular. Se tomó el plasma que
corresponde a la parte superior del tubo centrifugado, evitando acercarse a la capa de
la interfase. El plasma obtenido se alicuotó y se almacenó a 4 °C para su posterior uso
(Fig. 3) (ISBER, 2008)
FIGURA 3. Plasma sanguíneo de C. moreletii alicuotado.
2.2.2. Cuantificación de proteínas del plasma sanguíneo de cocodrilo y
humano
Para determinar la cantidad de proteína en el plasma sanguíneo se utilizó el protocolo
del kit comercial PierceTM BCA Protein Assay Kit de Thermo ScientificTM y las muestras
fueron analizadas por espectrofotometría. Tomando como referencia la curva de
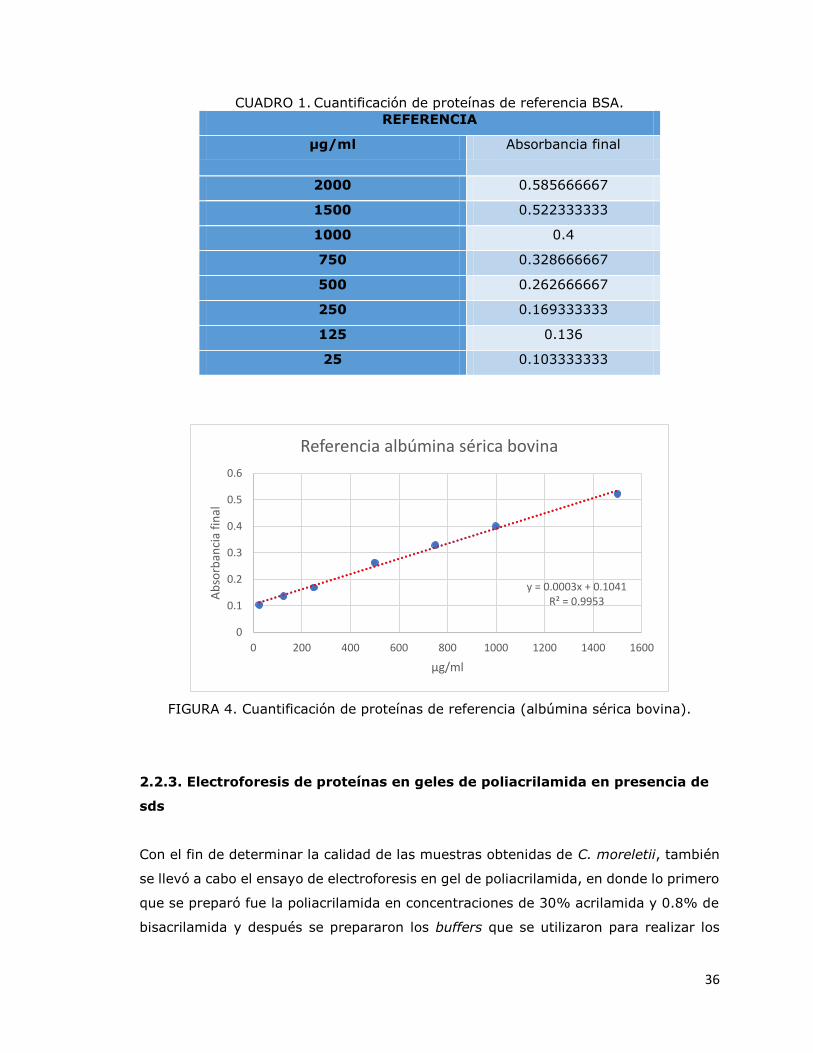
proteínas totales de la albúmina (Cuadro 1 y Fig. 4), se pudo determinar la
concentración de proteínas totales de los plasmas de humano y cocodrilo (Cuadro 2).
Los valores puntuales de proteínas totales de C. moreletii y humano fueron expresados
en miligramos por mililitro, donde para C. moreletii tuvo un valor de 66.3 mg/mL,
mientras que para humano fue 56.1 mg/mL de proteínas totales.
36
CUADRO 1. Cuantificación de proteínas de referencia BSA.
REFERENCIA
µg/ml Absorbancia final
2000 0.585666667
1500 0.522333333
1000 0.4
750 0.328666667
500 0.262666667
250 0.169333333
125 0.136
25 0.103333333
FIGURA 4. Cuantificación de proteínas de referencia (albúmina sérica bovina).
2.2.3. Electroforesis de proteínas en geles de poliacrilamida en presencia de
sds
Con el fin de determinar la calidad de las muestras obtenidas de C. moreletii, también
se llevó a cabo el ensayo de electroforesis en gel de poliacrilamida, en donde lo primero
que se preparó fue la poliacrilamida en concentraciones de 30% acrilamida y 0.8% de
bisacrilamida y después se prepararon los buffers que se utilizaron para realizar los
y = 0.0003x + 0.1041R² = 0.9953
0
0.1
0.2
0.3
0.4
0.5
0.6
0 200 400 600 800 1000 1200 1400 1600
Ab
sorb
anci
a fi
nal
µg/ml
Referencia albúmina sérica bovina

37
geles, uno de 1.5 M tris-HCl pH 8.8 y otro de 0.5 M tris HCl pH 6.8, así como el buffer
amortiguador de la cámara y el buffer de muestra reductor; todos estos reactivo se
hicieron de acuerdo a los protocolos descritos por Coligan et al. (1992).
Con los reactivos listos, se procedió a realizar los geles. El primero fue el gel separador
al 15%. Para éste, se colocaron en un vaso de precipitado de 10 mL 3.75 mL de
poliacrilamida, 1.875 mL de buffer 4X Tris-HCl/SDS pH 8.8, 1.875 mL de H2Od, 25 µl
de persulfato de amonio al 10% y finalmente 10 µl de TEMED; todos los reactivos se
homogeneizaron y con ayuda de una pipeta de transferencia se vertió la preparación
entre los cristales del molde y de inmediato se agregó una delgada capa de 1-2 mm de
agua destilada dejando polimerizar durante 15-20 min. Una vez que la interfase del
polímero y el agua se tornó visible, se decantó el exceso y se enjuagó con agua
destilada. Posteriormente, en ese mismo molde se realizó el gel de apilamiento al 5%,
para el que en un vaso de precipitado de 10 mL se agregaron 700 µl de poliacrilamida,
1.5 mL de H2Od, 250 µl de buffer 4X Tris HCl/SDS pH 6.8, 2.5 µl d TEMED y finalmente
20 µl de persulfato de amonio al 10%; los reactivos se homogeneizaron y la mezcla se
vertió con una pipeta de transferencia al molde de cristal rápidamente, acomodando el
peine de manera que no quedaran burbujas y se dejó polimerizar de 15-20 min.
Después, se retiró el peine y se lavaron los pozos del gel con agua destilada. El molde
con el gel se colocó en la cámara de electroforesis y ésta se llenó con el buffer
amortiguador de cámara 1X, sumergiendo totalmente el tanque interior entre los
cristales y de esta manera el gel quedó listo para ser cargado con las muestras.
La preparación de la muestra se realizó de manera independiente y el primer paso fue
la extracción de proteínas por medio del reactivo ProteoJet Mammalian Cell Lysis
(Thermo Fisher). Para esto, el plasma liofilizado de cocodrilo se diluyó en agua destilada
por medio de un sonicador BransonTM; el plasma diluido se colocó en viales en diferentes
concentraciones para un volumen final de 1 mL por vial. Las concentraciones de plasma
utilizadas fueron: 250, 128, 64, 32, 16, 8, 4 y 2 mg por vial. Los viales se centrifugaron
a 2500 g por 10 min y se les retiró el sobrenadante; se les agregó el reactivo ProteoJet
Mammalian Cell Lysis (Thermo Fisher) a razón de 1 mL por cada 100 µl de pellet
húmedo, se homogeneizó de manera suave durante 1 min y se centrifugó a 14000 g
por 15 minutos. Finalmente, se tomó el sobrenadante de cada muestra para analizarlo.
Una vez teniendo las muestras listas, se mezclaron con el buffer reductor tomando 15
µl de muestra y 15 µl de buffer reductor para reducir o romper los enlaces de disulfuro

38
de las proteínas, se incubó cada vial a 95 °C por 5 minutos y una vez pasado este
tiempo las muestras estaban listas para ser montadas en los pozos de gel.
Finalmente se colocaron las muestras en los pozos del ge. En el primer pozo se
colocaron 3 µl del marcador de peso molecular de proteínas de la marca Fermentas
PageRulerTM Unstained Protein Ladder, mientras que en los siguientes pozos se
colocaron las muestras de manera descendente comenzando con la de mayor
concentración de proteína, mencionadas anteriormente. Con el gel listo se cerró con la
tapa y se conectaron los electrodos a la fuente de poder, dejando correr el gel 90
minutos a 200 V, se sacó el molde de cristal y con mucho cuidado se separaron los
cristales y el gel se colocó en un recipiente con azul de Coomassie R-250 para teñirse,
en donde se dejó 24 horas El gel teñido se colocó en una solución fijadora con ácido
acético glacial durante 2 horas y finalmente se observó en un pequeño negatoscopio
(Fig. 5).
FIGURA 5. Electroforesis en gel de poliacrilamida para lectura de proteínas en plasma
de C. moreletii con marcador de peso molecular.

39
2.3. Conteo y evaluación de reactividad de linfocitos por ensayo MTT
Primero se evaluó la viabilidad y reactividad de linfocitos humanos tomados de sangre
periférica después de ser mezclados con el plasma sanguíneo de cocodrilo. Para esto,
se hizo un cultivo de linfocitos humanos separados con Ficoll® Paque Plus GE, EE.UU
por gradiente de centrifugación a partir de 10 mL de sangre total, utilizando medio de
cultivo RPMI 1640 suplementado con suero fetal bovino al 10%, antibiótico-antimicótico
1% y glutamina 1%. El cultivo se incubó en atmósfera de CO2 al 5% y humedad
constante.
Para el cultivo de linfocitos de sangre periférica humana, se obtuvieron 10 mL de sangre
periférica humana y se almacenaron en tubos con heparina sódica (5 mL por tubo);
posteriormente se realizó la separación de los mononucleares por gradiente de Ficoll
1:1. Para esto, en la campana de flujo laminar se colocaron 5 mL de ficoll en dos tubos
Falcon de 15 mL y a continuación se depositaron 5 mL de sangre periférica por tubo,
esto se hizo de manera muy suave a través de las paredes del tubo con una pipeta de
transferencia estéril de 3 mL (Fig.6). Los tubos con sangre y Ficoll se centrifugaron a
3500 rpm por 60 min. Después, con una pipeta de transferencia estéril se tomó de la
parte superior del tubo el plasma sanguíneo y éste se depositó en un tubo Falcon para
ser almacenado a -4 °C. El resto de cada muestra que quedó en cada tubo Falcon se
transfirió a otro tubo Falcon de 15 mL, al que se le adicionaron 2 mL de solución PBS
1X y se centrifugó a 1100 rpm por 10 min. Después se retiró el sobrenadante con una
pipeta de transferencia y el pellet resultante se depositó en una botella de cultivo de
25 cm3 con 10 mL de medio de cultivo RPMI, previamente atemperado a 37 °C y
suplementado. Se homogeneizó suavemente y se dejó incubar en una estufa de cultivo
celular a 37 °C con 5% de CO2 por 24 h.

40
FIGURA 6. Adición del Ficoll® Paque Plus GE, EE.UU a los tubos con sangre
heparinizada.
Después del cultivo por 24 h se realizó el conteo celular con azul de Trypan al 0.4%.
Esta tinción permitió además distinguir los linfocitos vivos de los muertos. En un tubo
Eppendorf de 1.5 mL se agregaron 10 μl de azul de Trypan al 0.4% y 10 μl del cultivo
homogeneizado, dejándolo incubar durante 1 min; a continuación, se colocó sobre la
cámara de Neubauer un cubreobjetos y se depositaron 10 μl de la suspensión celular
premezclada con azul de Trypan en la entrada superior y los otros 10 μl en la entrada
inferior, hasta que se alcanzó la confluencia para examinarse al microscopio óptico
(Olympus®, E.U.), y se contaron las células presentes (Fig. 7). Posteriormente, los
linfocitos teñidos de azul en su totalidad o su membrana se consideraron muertos (azul
de Trypan positivos, AT+), mientras que los que no teñidos se consideraron vivos (azul
de Trypan negativos, AT-). Este conteo se realizó cada 24 h para obtener una curva de
crecimiento y establecer las mejores condiciones de estabilidad del cultivo, que fueron
las que se emplearon para todos los conteos de linfocitos posteriores.

41
FIGURA 7. Linfocitos humanos observados por medio de microscopio óptico y cámara
de Neubauer.
Para determinar el efecto citotóxico del plasma de cocodrilo sobre los linfocitos
humanos de sangre periférica se realizó un ensayo de la reducción metabólica del
Bromuro de 3-(4,5- dimetiltazol-2-ilo) -2,5- difeniltetrazol (MTT) (Mosmann, 1983).
Para esto, se emplearon cultivos óptimos de 160,000 células por pocillo, en placas de
cultivo de 96 pozos; se colocaron 100 µl de células y se adicionaron 100 µl de plasma
de cocodrilo con seis diferentes concentraciones, se dejó incubar por 1 h en estufa de
cultivo a 37 °C, y una vez pasado el tiempo, se le añadió a cada pozo 10 µl de reactivo
MTT dejándose incubar 4 h a 37 °C. Posteriormente, se le añadió a cada pozo 100 µl
de detergente y se volvió a incubar 4 h a 37 °C. Finalmente, se midió la citotoxicidad
por medio de espectrofotometría a 570 nm; se realizaron dos repeticiones
independientes por cada muestra.
2.4. Aislamiento y confirmación de las cepas de Staphylococcus aureus
En esta sección es importante señalar que se obtuvieron seis aislados clínicos de S.
aureus de importancia clínica que fueron proporcionados por el Cepario Microbiano de
la Facultad de Bioanálisis de la UV región Veracruz; sin embargo, para los ensayos de
microdilución en caldo descritos más adelante, solamente se utilizó el aislado MRSA
20090127 y una cepa de referencia ATCC 1556.

42
Dado que estos aislados no estaban almacenados correctamente, se prepararon los
agares sólidos para su debido almacenamiento. Para esto, se realizó el pesaje de los
agares sal y manitol y BHI, y se preparó un litro de cada uno. El agar sal y manitol se
vertió en cajas de cultivo individuales, mientras que el agar BHI se vertió en tubos de
16 x 100 a razón de 5 mL por tubo los cuales una vez esterilizados y solidificados se
almacenaron en refrigeración a -4 °C para su posterior uso (Fig. 8).
Por otro lado, en las placas que tenían agar sal y manitol se hizo una resiembra de las
cepas de S. aureus que estaban en refrigeración y se incubaron a 37 °C por 24 h, para
observar el crecimiento y asegurar que no hubiera contaminación (Fig. 9).
Posteriormente se hizo una siembra de esas cepas en los tubos de cultivo con agar BHI,
se dejaron incubar a 37 °C por 24 h (Fig. 10) y finalmente se hizo el almacenamiento,
cubriendo el crecimiento bacteriano de los tubos con aceite mineral estéril como se
puede observar en la Figura 10.
Ahí mismo fueron identificados los aislados bacterianos por los métodos tradicionales
de siembra en gelosa. Para su reactivación, los cultivos fueron inoculados en caldo de
infusión de cerebro-corazón (BHI), el cual fue incubado en estufa de cultivo a 37 °C por
4 h hasta observar turbidez en el medio, esto se realizó por duplicado. Posteriormente,
éstos aislados fueron inoculados por medio de un hisopo que se humedeció con el
medio líquido y se sembró en, en agar sangre y se incubaron a 37 °C por 24 h en estufa
de cultivo para reactivar su metabolismo. Al terminar el período de incubación se
tomaron las colonias aisladas para re-suspenderlas y fomentar su crecimiento en agar
sal y manitol por 24 h a 37 °C, donde se observó la morfología colonial, la fermentación
del manitol y en agar sangre se pudo observar la presencia de hemólisis (Fig. 11 y 12).
Para poder confirmar que los aislados clínicos fueran de S. aureus se llevaron a cabo
diferentes pruebas como tinción de Gram, para observar la caracterización morfológica,
las pruebas de catalasa, coagulasa, y finalmente la prueba de la susceptibilidad y
resistencia antimicrobiana.

43
FIGURA 8. A) Preparando cajas de Petri de agar sal y manitol; B) Tubos de cultivo con
agar BHI; C) Cajas de cultivo con agar sal y manitol y agar BHI.
FIGURA 9. Observación de cepas de Staphylococcus aureus después de 24 h en estufa
de cultivo.
A B C

44
FIGURA 10. A) Siembra de S. aureus en tubos de cultivo para su almacenamiento; B)
Cepa de S. aureus en tubo de cultivo con aceite mineral esterilizado.
FIGURA 11. A) Staphylococcus aureus en agar sal y manitol, donde las colonias doradas
(flecha roja) representan la fermentación del manitol; B) S. aureus en agar sangre
donde se observa la hemólisis beta. En forma de halos claros alrededor de las colonias.
A B

45
2.4.1. Caracterización morfológica
Se realizó una tinción de Gram (Fig. 12), a cada uno de los aislados clínicos para
confirmar que las bacterias tuvieran las características morfológicas de Staphylococcus
aureus, para esto se tomaron con ayuda de un aplicador de madera 3 a 4 colonias de
la placa con agar sal y manitol y se colocaron sobre un portaobjetos y se fijaron a este
pasando el portaobjetos por la flama del mechero rápidamente, posterior a eso se
empleó cristal violeta como primer colorante durante 1 min y se enjuagó con agua
corriente, después se utilizó yodo-Lugol como fijador y dejó 1 min y se enjuagó con
agua corriente, después se decoloró con alcohol-cetona por 15 seg, se enjuagó con
agua corriente y finalmente como colorante de contraste se utilizó safranina por 1 min
y se enjuagó con agua corriente, la morfología colonial se observó bajo el microscopio
óptico.
FIGURA 12. Morfología colonial de Staphylococcus aureus.
2.4.2 Prueba de catalasa en lámina
Por otro lado, se llevó a cabo la prueba de catalasa (Fig. 13), para lo que se colocó una
gota de peróxido de hidrógeno (H2O2) sobre un portaobjetos y se tomó con un aplicador
de madera de 3 a 4 colonias de la placa de cultivo de sal y manitol y se homogeneizó
hasta observar la descomposición del peróxido convirtiéndolo en agua, más oxígeno el
cual se evidencia en forma de burbujas.

46
FIGURA 13. Realizando prueba de catalasa.
2.4.3. Prueba de coagulasa
Se realizó la prueba de coagulasa. En un tubo de ensaye de 13 x 100 se colocaron 100
µl de plasma humano citrado y se le agregó 100 µl de caldo BHI inoculado con S.
aureus; el tubo se tapó con una torunda de algodón y se dejó incubar a 37 °C por 24
h, finalmente se observó el coágulo que forma la enzima coagulasa característico de S.
aureus (Fig. 14).
FIGURA 14. Formación de la fibrina (flecha roja) en fibrinógeno en plasma humano.

47
2.4.4. Prueba de susceptibilidad y resistencia antimicrobiana
La identificación de la susceptibilidad y resistencia antimicrobiana se realizó a los
aislados clínicos de S. aureus, y se midió mediante el método de difusión en agar
de Kirby-Bauer (Bauer et al., 1966). Primero se prepararon los inóculos para la escala
McFarland en medio líquido, se propagaron de 3 a 5 colonias de cada cepa en caldo de
infusión cerebro-corazón y se incubaron a 35 °C de 2 a 4 h, hasta observar turbidez,
misma que se midió a través de un espectrofotómetro de luz visible Genesys 20 a una
longitud de onda de (λ) de 625 nm, utilizando como blanco el caldo inocuo, con un
marco de absorbancia de 0.08 a 0.13 nm, el equivalente a 0.5 en la escala de
McFarland, con una replicación de precisión ± 0.10, ya a que a dicha lectura presenta
una óptima distribución de la cepa en el medio sólido.
Posteriormente, ya con el inóculo listo para cada cepa, se prepararon dos cajas de Petri
para cada aislado de S. aureus, con medio agar Mueller-Hinton (HM) pues este medio
permite un crecimiento óptimo de S. aureus así como una correcta expresión del
fenotipo de resistencia bacteriana. Entonces, un hisopo humedecido del patrón de
McFarland se deslizó por todo el agar tres veces, rotando la caja de Petri en un ángulo
de 90°, formando un césped uniforme, dejando cada caja de Petri de 3 a 5 minutos.
Posteriormente se colocaron los discos con los antibióticos (ATB) con pinzas estériles,
se presionaron ligeramente sobre el agar y se dejaron incubar a 35 °C por 24 h; en
una caja de Petri se aplicaron los antibióticos de sulfametoxazol/trimetoprim (SXT) en
concentración de 1.25 y 35.75 µg/mL respectivamente (Fig. 15); y en otra se aplicó
cefoxitín (FOX) en concentración de 30 µg/mL (Fig. 16). La lectura de los halos de
inhibición se midió en mm con ayuda de una regla y las medidas se compararon con
los puntos de corte descritos en el manual CLSI-2018-M100-S28ed Performance
Standards for Antimicrobial Susceptibility Testing, establecidos para la especie de
Staphylococcus aureus que sean resistentes a la meticilina o su equivalente actual a
oxacilina.
Los puntos de corte que considera el CLSI en cultivos de S. aureus son: para
sulfametoxazol/trimetoprim <10 mm resistente, >16 mm sensible; y para cefoxitín
<21 mm resistente, >22 mm sensible.

48
FIGURA 15. Halos de inhibición (mm) en cultivo de S. aureus con discos de
sulfonamidas/trimetoprim (17 mm), donde el halo gris que se observa indica la
inhibición bacteriana que provoca cada antibiótico.
FIGURA 16. No se observaron halos de inhibición (mm) en cultivo de S. aureus con
discos de cefoxitín.

49
2.5. Evaluación de la actividad antibacteriana por el método de microdilución
en caldo
Se realizó el ensayo de microdilución en caldo con dos cultivos MRSA, la cepa de
referencia ATCC 1556 y el aislado clínico MRSA 20090127, este último procedente del
Hospital Regional de la Ciudad de Veracruz. Se tomaron con el asa bacteriológica de 3
a 4 colonias de cada cultivo y se propagaron en caldo BHI, dejándolas en la estufa de
cultivo 3 horas a 37 °C hasta observar turbidez; esta turbidez se midió a través de un
espectrofotómetro de luz visible Genesys 20 a una longitud de onda de (λ) de 625 nm,
utilizando como blanco el caldo inocuo, con un marco de absorbancia de 0.08 a 0.13
nm, el equivalente a 0.5 en la escala de McFarland. Ya con el inóculo listo se hizo una
dilución 1/100, donde se colocaron 100 µl del caldo con bacterias en 9900 µl de caldo
nutritivo inocuo.
Una vez con el inóculo listo se procedió a cargar la placa de 96 pozos con fondo plano,
agregándole 100 µl de caldo nutritivo a cada pocillo, y posteriormente a la primera
columna se le agregó 100 µl de plasma de cocodrilo, se homogenizó con ayuda de la
micropipeta y se hicieron 5 diluciones seriadas; esto se hizo tomando 100 µl del primer
pocillo y agregándolo al siguiente, y de ese segundo pocillo se tomaron 100 µl y se
agregaron al pocillo siguiente y así consecutivamente hasta terminar las diluciones;
desechando los últimos 100 µl restantes. El siguiente paso consistió en agregar 100 µl
de caldo nutritivo con 1.5x106 UFC/mL de bacterias. Las concentraciones finales de
plasma de cocodrilo quedaron de la siguiente manera, 3315 µg/mL, 1657 µg/mL, 828
µg/mL, 414 µg/mL y 207 µg/mL. Los controles positivos o control del antibiótico se
colocaron en la columna siguiente con 100 µl de caldo nutritivo inoculado con bacterias
más el antibiótico trimetoprim con sulfametoxazol en concentración de 1.25 y 35.75
µg/mL respectivamente; y como control negativo o control de crecimiento en la
siguiente columna se utilizaron 100 µl de caldo nutritivo inoculado con bacterias más
100 µl de caldo nutritivo inocuo. La placa se dejó incubar 20 horas a 37 °C y se leyó
por espectrofotometría a 600 mn (Fig. 17).

50
FIGURA 177. Placa con las diferentes diluciones de plasma de cocodrilo.
2.6. Análisis estadístico
Las variables a analizar fueron las diferentes concentraciones de plasma de cocodrilo y
la presencia o ausencia de actividad antimicrobiana. El análisis estadístico de los datos
se realizó mediante la prueba no paramétrica de Kruskal-Wallis.
Donde:
ni= Tamaño de la muestra para la población i
Ti= Suma de rangos para la población i
n= Número total de observaciones
𝐻 =12
𝑛(𝑛 + 1)∑
𝑇𝑖2
𝑛𝑖
𝑘
𝑖=1
− 3(𝑛 + 1)
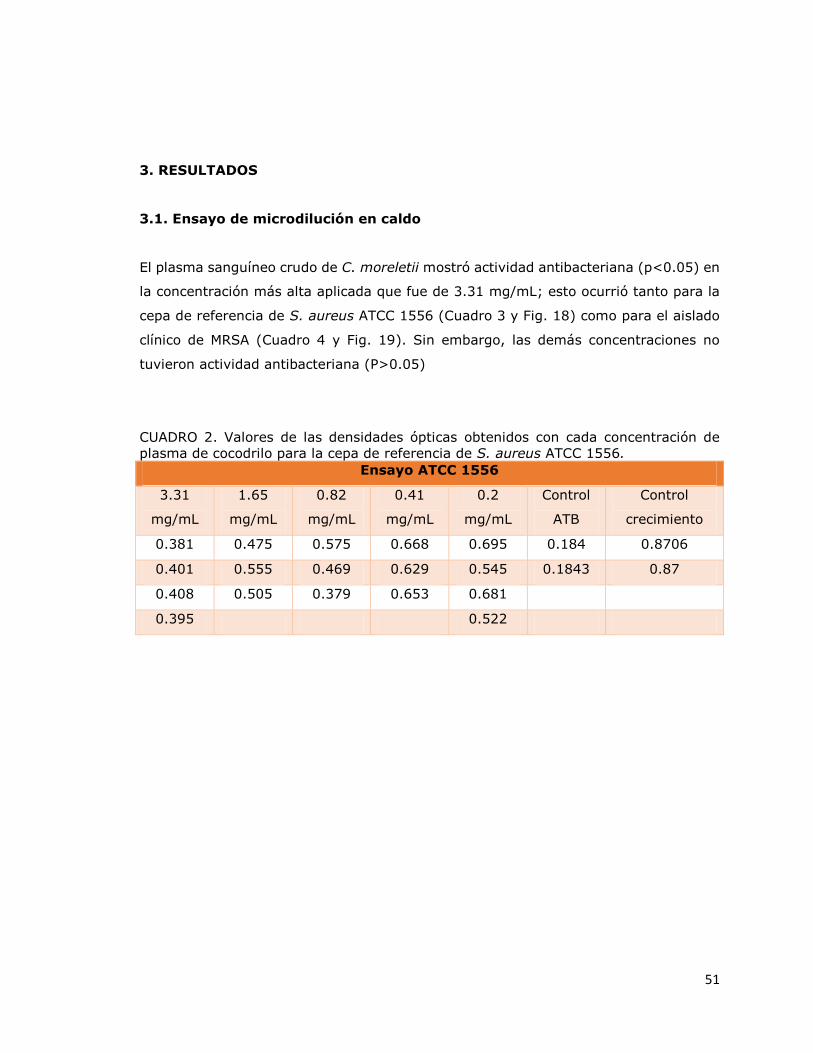
51
3. RESULTADOS
3.1. Ensayo de microdilución en caldo
El plasma sanguíneo crudo de C. moreletii mostró actividad antibacteriana (p<0.05) en
la concentración más alta aplicada que fue de 3.31 mg/mL; esto ocurrió tanto para la
cepa de referencia de S. aureus ATCC 1556 (Cuadro 3 y Fig. 18) como para el aislado
clínico de MRSA (Cuadro 4 y Fig. 19). Sin embargo, las demás concentraciones no
tuvieron actividad antibacteriana (P>0.05)
CUADRO 2. Valores de las densidades ópticas obtenidos con cada concentración de
plasma de cocodrilo para la cepa de referencia de S. aureus ATCC 1556.
Ensayo ATCC 1556
3.31
mg/mL
1.65
mg/mL
0.82
mg/mL
0.41
mg/mL
0.2
mg/mL
Control
ATB
Control
crecimiento
0.381 0.475 0.575 0.668 0.695 0.184 0.8706
0.401 0.555 0.469 0.629 0.545 0.1843 0.87
0.408 0.505 0.379 0.653 0.681
0.395 0.522
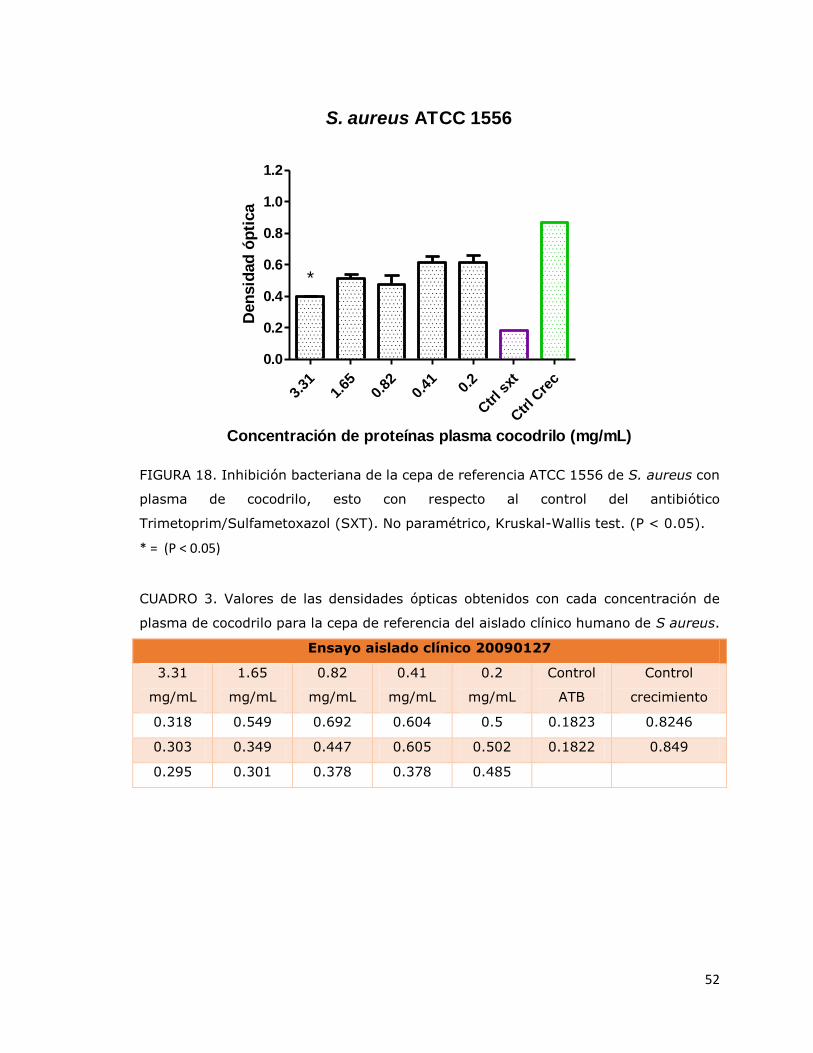
52
S. aureus ATCC 1556
3.31
1.65
0.82
0.41 0.
2
Ctrl s
xt
Ctrl C
rec
0.0
0.2
0.4
0.6
0.8
1.0
1.2
*
Concentración de proteínas plasma cocodrilo (mg/mL)
Den
sid
ad
óp
tica
FIGURA 18. Inhibición bacteriana de la cepa de referencia ATCC 1556 de S. aureus con
plasma de cocodrilo, esto con respecto al control del antibiótico
Trimetoprim/Sulfametoxazol (SXT). No paramétrico, Kruskal-Wallis test. (P < 0.05).
* = (P < 0.05)
CUADRO 3. Valores de las densidades ópticas obtenidos con cada concentración de
plasma de cocodrilo para la cepa de referencia del aislado clínico humano de S aureus.
Ensayo aislado clínico 20090127
3.31
mg/mL
1.65
mg/mL
0.82
mg/mL
0.41
mg/mL
0.2
mg/mL
Control
ATB
Control
crecimiento
0.318 0.549 0.692 0.604 0.5 0.1823 0.8246
0.303 0.349 0.447 0.605 0.502 0.1822 0.849
0.295 0.301 0.378 0.378 0.485

53
S. aureus aislado clínico humano20090127
3.31
1.65
0.82
0.41 0.
2
Ctrl s
tx
Ctrl C
rec
0.0
0.2
0.4
0.6
0.8
1.0
1.2
*
Concentración de proteínas plasma cocodrilo (mg/mL)
Den
sid
ad
óp
tica
FIGURA 19. Inhibición bacteriana del aislado clínico humano de S. aureus 20090127
con plasma de cocodrilo, esto con respecto al control del antibiótico
Trimetoprim/Sulfametoxazol (SXT). No paramétrico, Kruskal-Wallis test. (P < 0.05).
* = (P < 0.05)
3.2. Ensayo MTT
La citotoxicidad del plasma crudo de C. moreletii se determinó por el ensayo MTT,
después de que las células se cultivaran con 0-2000 µg/mL de proteínas totales de
plasma. No se observaron efectos citotóxicos de los componentes sanguíneos de C.
moreletii. Además, se observó que el plasma promovió la proliferación celular hasta un
400% (Fig. 20).
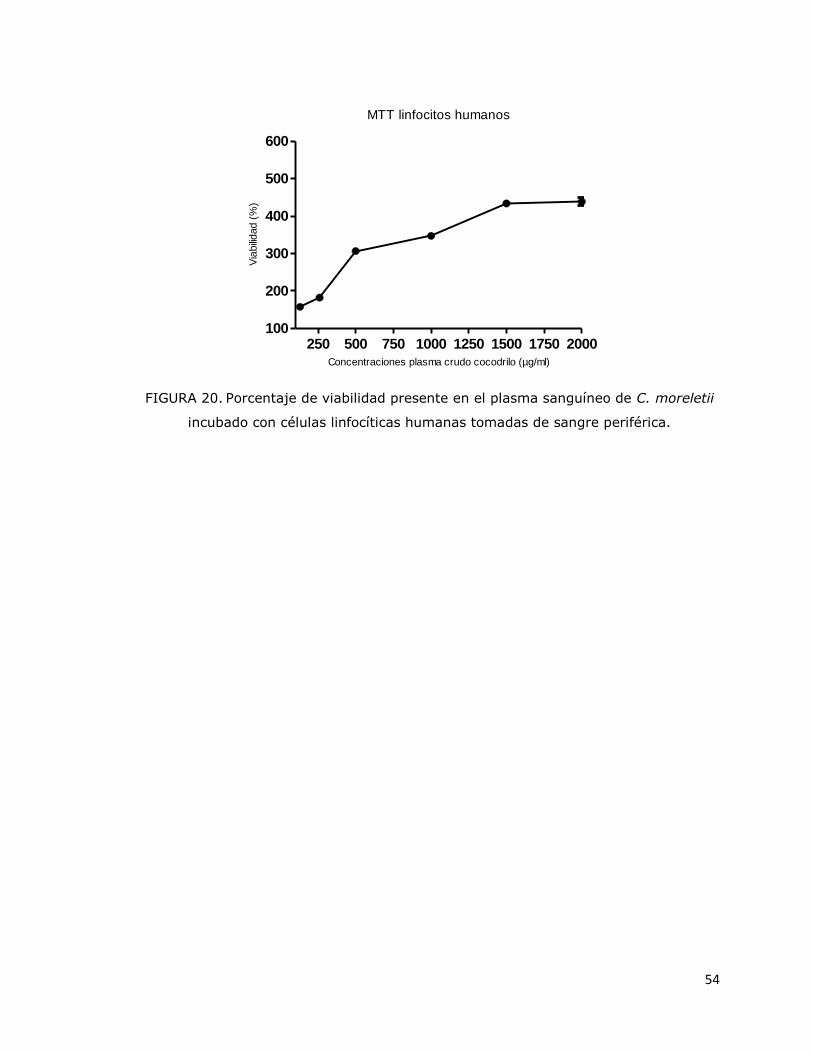
54
MTT linfocitos humanos
250 500 750 1000 1250 1500 1750 2000100
200
300
400
500
600
Concentraciones plasma crudo cocodrilo (µg/ml)
Via
bilid
ad
(%
)
FIGURA 20. Porcentaje de viabilidad presente en el plasma sanguíneo de C. moreletii
incubado con células linfocíticas humanas tomadas de sangre periférica.

55
4. DISCUSIÓN
La actividad antibacteriana del plasma sanguíneo de cocodrilianos ha sido reportada en
algunas especies como el C. siamensis, en el que Kommanee et al. (2012) observaron
que a concentraciones de 50 µg/mL de plasma semipurificado hubo inhibición de más
del 80% del crecimiento de algunas cepas bacterianas patógenas como Salmonella
typhi ATCC 11778, Vibrio Cholerae non01, Staphylococcus epidermidis ATCC 12228, un
aislado clínico de Pseudomonas aeruginosa y un aislado clínico de Salmonella typhi. En
el presente estudio se observó que el plasma sanguíneo crudo de C. moreletii resultó
eficaz como antimicrobiano sobre cepas MRSA en la concentración de 3.31 mg/mL de
proteínas totales del plasma. No existen datos previos sobre la actividad antibacteriana
del plasma sanguíneo de C. moreletii con los que comparar los resultados de este
estudio. Algunos autores sugieren que el compuesto antibacteriano del plasma podría
entrar progresivamente en el espacio citoplasmático, perturbando y dañando la
superficie bacteriana (Kommanee et al., 2012; Barksdale et al., 2016).
En este estudio se utilizaron concentraciones de proteínas totales del plasma sanguíneo
de C. moreletii, lo que podría explicar por qué la concentración que tuvo una actividad
antibacteriana significativa (3.31 mg/mL) fue mucho más alta que las reportadas por
Kommanee et al. (2012), quienes sólo utilizaron 50 µg/mL de plasma que había sido
semipurificado. Se infiere que al ser plasma completo existen una gran variedad de
componentes presentes y que sólo un porcentaje de éstos serían los responsables de
la actividad antibacteriana.
Los péptidos antimicrobianos se han aislado de una amplia variedad de organismos, así
que se podría suponer que ciertos péptidos presentes en el plasma de C. moreletii sean
responsables de la actividad antibacteriana mostrada en este estudio. El contacto inicial
entre el péptido y el organismo objetivo sería electrostático, ya que la mayoría de las
superficies bacterianas son aniónicas, por lo que la membrana bacteriana sería
propensa a ser atacada por dichos péptidos cargados positivamente (Leelawongtawon
et al., 2010).

56
Existen datos de péptidos catiónicos aislados en C. siamensis como el Apo 1, Apo2 y
A1P, que mostraron actividad antibacteriana de amplio espectro sobre aislados clínicos
de E. coli, P. aeruginosa, e inclusive sobre S. aureus resistente a antibióticos (Barksdale
et al. 2016).
La resistencia a los antibióticos ha aumentado constantemente durante las últimas
décadas, y los péptidos antimicrobianos se consideran una terapia alternativa ya que
poseen una alta actividad bactericida y disminución de la capacidad que tienen las
bacterias para volverse resistentes. En general, este hallazgo sugiere una nueva
perspectiva sobre la viabilidad de utilizar los componentes que ofrece el plasma
sanguíneo de C. moreletii como una opción alternativa para combatir infecciones
causadas por bacterias. En ese sentido, se considera que el siguiente paso en la
investigación de las propiedades de los componentes del plasma sanguíneo de C.
moreletii puede ser aislar péptidos con posible actividad antimicrobiana.
Los datos del ensayo de citotoxicidad demostraron que las diferentes concentraciones
(250-2000 µg/mL) de proteínas totales del plasma de C. moreletii no tuvieron efecto
citotóxico sobre los cultivos de linfocitos humanos. Este resultado no se había reportado
anteriormente para esta especie de cocodriliano. Sin embargo, coincide con lo que
observaron Kommanee et al. (2012) al evaluar el efecto de diferentes concentraciones
de plasma de C. siamensis (6.2-1000 µg/mL) sobre cultivos de células fagocíticas de
ratones RAW264.7 en un ensayo MTT, dado que no tuvieron efecto sobre las mismas.
En el presente estudio, además de no causar citotoxicidad, los componentes del plasma
sanguíneo de C. moreletii promovieron la proliferación celular de los linfocitos humanos
hasta un 400%. Al respecto, en un estudio realizado por Jangpromma et al. (2016),
donde se probó la capacidad citotóxica del plasma sanguíneo de C. siamensis sobre
queratinocitos humanos, se demostró que aparte de no causar citotoxicidad, hubo
proliferación celular de hasta 155%. Esto podría deberse a que el plasma de los
cocodrilianos tendría propiedades pronunciadas de cicatrización relacionadas con una
mayor proliferación y migración celular y podría estar relacionado con las plaquetas
suspendidas en el plasma.
Hasta donde se sabe, todavía no se han divulgado informes que cubran la actividad
proliferativa de componentes sanguíneos de esta especie, este es el primer informe;

57
sin embargo, la reparación de heridas ocurre en todo tipo de tejido, por lo que el plasma
sanguíneo de C. moreletii mostró una capacidad de proliferación celular que podría
apoyar a la regeneración celular de tejidos dañados.

58
5. CONCLUSIONES
El plasma sanguíneo crudo de C. moreletii mostró actividad antibacteriana contra cepas
de Staphylococcus aureus. Cuando fue puesto en contacto con linfocitos humanos sanos
no sólo no causó citotoxicidad, sino que estimuló la multiplicación de estas células.

59
REFERENCIAS
Aarestrup, F. M. 2015. The livestock reservoir for antimicrobial resistance: a personal
view on changing patterns of risks, effects of interventions and the way forward. Philos.
Trans. R. Soc. B, 370(1670): 20140085.
Andersson, U., Bird, A. G., Britton, S., y Palacios, R. 1981. Humoral and cellular
immunity in humans studied at the cell level from birth to two years of age. Immuno.l
Rev., 57 (1): 5-38.
Araki, T., Yamamoto, T., y Torikata, T. 1998. Reptile lysozyme: the complete amino
acid sequence of soft-shelled turtle lysozyme and its activity. Biosci. Biotechnol.
Biochem., 62(2): 316-324.
Archer, G. L., y Climo, M. W.1994. Antimicrobial susceptibility of coagulase-negative
staphylococci. Antimicrob. Agents Chemother., 38(10): 2231.
Aree, K., Siruntawineti, J., y Chaeychomsri, W. 2011. Crocodylus siamensis serum and
macrophage phagocytic activity. J. Med. Assoc. Thai.= Chotmaihet thangphaet, 94:
S131-8.
Arias, C. A., Rincon, S., Chowdhury, S., Martínez, E., Coronell, W., Reyes, J., y Murray,
B. E. 2008. MRSA USA300 clone and VREF--a US-Colombian connection?. N. Engl. J.
Med., 359(20): 2177-2179.
Barksdale, S. M., Hrifko, E. J., Chung, E. M. C., y Van-Hoek, M. L. 2016. Peptides from
American alligator plasma are antimicrobial against multi-drug resistant bacterial
pathogens including Acinetobacter baumannii. BMC. Microbiol., 16(1): 189.
Bauer, A. W., Kirby, W. M. M., Sherris, J. C., y Turck, M. 1966. Antibiotic susceptibility
testing by a standardized single disk method. Am. J. Clin. Pathol., 45: 493-496.
Belloso, W. H. 2009. Historia de los antibióticos. Rev. Hosp. Ital. B. Aires, 29: 102-11.
Biron, C. A. 1997. Natural killer cell regulation during viral infection. Bichem. Soc.
Trans., 25: 687-690.
Boucher, H., Miller, L. G., y Razonable, R. R. 2010. Serious infections caused by
methicillin-resistant Staphylococcus aureus. Clin. Infect. Dis., 51: S183-S197.

60
Bronner, S., Monteil, H., y Prévost, G. 2004. Regulation of virulence determinants in
Staphylococcus aureus: complexity and applications. FEMS. Microbiol. Rev., 28(2):
183-200.
Buthelezi, S., Southway, C., Govinden, U., Bodenstein, J., & du Toit, K. 2012. An
investigation of the antimicrobial and anti-inflammatory activities of crocodile
oil. Journal. Ethnopharmacol., 143(1), 325-330.
CDC. Centers for Disease Control and Prevention. 2013.
https://www.cdc.gov/mrsa/healthcare/index.html. Recuperado 3 de octubre de 2018.
CDC. Centers for Disease Control and Prevention. 2018.
https://www.cdc.gov/drugresistance/biggest_threats.html. Recuperado 3 de octubre
de 2018.
Chambers, H. F., y DeLeo, F. R. 2009. Waves of resistance: Staphylococcus aureus in
the antibiotic era. Nat. Rev. Microbiol., 7(9): 629.
Chapman, G. H., Berens, C., Peters, A., y Curcio, L. 1934. Coagulase and hemolysin
tests as measures of the pathogenicity of staphylococci. J. Bacteriol., 28(4): 343.
Chattopadhyay, S., Sinha, N. K., Banerjee, S., Roy, D., Chattopadhyay, D., y Roy, S.
2006. Small cationic protein from a marine turtle has β‐defensin‐like fold and
antibacterial and antiviral activity. Proteins, 64(2): 524-531.
Coico, R., y Sunshine, G. 2015. Immunology: a short course. USA: John Wiley & Sons.
Coligan, J.E.; Kruisbeek, A.M.; Margulies, D.H.; Shevach, E.M., and Strober, W. 1992.
Antibody detection and preparation. En: Current protocols in immunology. John Wiley
and sons. New York, unit 2,4.
Couvalin, A. J. (1988). El final de la edad de oro de los antibióticos. Ther Nat.
314(3):50-2.
Crisóstomo, M. I., Westh, H., Tomasz, A., Chung, M., Oliveira, D. C., y De-Lencastre,
H. 2001. The evolution of methicillin resistance in Staphylococcus aureus: similarity of
genetic backgrounds in historically early methicillin-susceptible and-resistant isolates
and contemporary epidemic clones. PNAS, 98(17), 9865-9870.
David, M. Z., y Daum, R. S. 2010. Community-associated methicillin-resistant
Staphylococcus aureus: epidemiology and clinical consequences of an emerging
epidemic. Clin. Microbiol. Rev., 23(3), 616-687.
DeLeo, F. R., Otto, M., Kreiswirth, B. N. y Chambers, H. F. 2010. Community-associated
methicillin-resistant Staphylococcus aureus. Lancet, 375: 1557-68.

61
Dickinson, T. M., y Archer, G. L. 2000. Phenotypic Expression of Oxacillin Resistance in
Staphylococcus epidermidis: Roles of mecATranscriptional Regulation and Resistant-
Subpopulation Selection. Antimicrob. Agents. Chemother., 44(6): 1616-1623.
Engberg, J., Aarestrup, F. M., Taylor, D. E., Gerner-Smidt, P., y Nachamkin, I. 2001.
Quinolone and macrolide resistance in Campylobacter jejuni and C. coli: resistance
mechanisms and trends in human isolates. Emer. Infect. Dis., 7(1): 24.
Francis, J. S., Doherty, M. C., Lopatin, U., Johnston, C. P., Sinha, G., Ross, T., Cai, M.,
Hansel, N., Perl, T., Ticehurst, J. R., Thomas, D., Bartlett, J. G., Nuermberger, E., y
Carroll, K. 2005. Severe community-onset pneumonia in healthy adults caused by
methicillin-resistant Staphylococcus aureus carrying the Panton-Valentine leukocidin
genes. Clin. Infect. Dis., 40(1): 100-107.
FDA. Food and Drug Administration. 2016.
https://www.fda.gov/downloads/forindustry/userfees/animaldruguserfeeactadufa/uc
m588085.pdf. Recuperado 28 de octubre de 2018.
Ganz, T. 2004. Defensins: antimicrobial peptides of vertebrates. Comptes rendus
biologies, 327(6): 539-549.
Gayen, S. K., Som, S., Sinha, N. K., y Sen, A. 1977. Lysozyme in egg whites of tortoises
and turtle: Purification and properties of egg white lysozyme of Trionyx gangeticus
cuvier. Arch. Biochem. Biophys., 183(2): 432-442.
Gould, I. M., y Bal, A. M. 2013. New antibiotic agents in the pipeline and how they can
help overcome microbial resistance. Virulence, 4(2), 185-191.
Gradelski, E., Valera, L., Aleksunes, L., Bonner, D., y Fung-Tomc, J. 2001. Correlation
between genotype and phenotypic categorization of staphylococci based on methicillin
susceptibility and resistance. J. Clin. Microbiol., 39(8), 2961-2963.
Hoffmann, J. A., Kafatos, F. C., Janeway, C. A., y Ezekowitz, R. A. B. 1999. Phylogenetic
perspectives in innate immunity. Science, 284(5418): 1313-1318.
Ingram, G. A., y Molyneux, D. H. 1983. The humoral immune response of the spiny-
tailed agamid lizard (Agama caudospinosum) to injection with Leishmania agamae
promastigotes. Vet. Immunol. Immunopathol., 4(4): 479-491.
ISBER. International Society for Biological and Environmental Repositories. 2008. Best
Practices for Repositories. Collection, Storage, Retrieval and Distribution of Biological
Materials for Research. Cell Preserv. Technol. 6: 1.
Ito, T., Katayama, Y., Asada, K., Mori, N., Tsutsumimoto, K., Tiensasitorn, C., y
Hiramatsu, K. 2001. Structural comparison of three types of staphylococcal cassette

62
chromosome mec integrated in the chromosome in methicillin-resistant Staphylococcus
aureus. Antimicrob. Agents. Chemother., 45(5): 1323-1336.
Jackson, L. C., Machado, L., y Hamilton, M. L. 1998. Principios generales de la
terapéutica antimicrobiana. Acta. Med., 8(1): 13-27.
Janeway, C. A., y Medzhitov, R. 2002. Innate immune recognition. Annu. Rev.
Immunol., 20(1): 197-216.
Jangpromma, N., Preecharram, S., Srilert, T., Maijaroen, S., Mahakunakorn, P.,
Nualkaew, N., y Klaynongsruang, S. (2016). In vitro and in vivo wound healing
properties of plasma and serum from Crocodylus siamensis blood. J. Microbiol.
Biotechnol, 26(6), 1140-1147.
Jawetz, E., Adelberg, E. A., Brooks, G. F., Butel, J. S., Melnick, J. L., y Ornston, L. N.
1992. Microbiología médica de Jawetz, Melnick y Adelberg. Publisher: McGrawHill, USA.
Pp. 185-193.
Jevons, M. P. 1961. Celbenin-resistant staphylococci. Br. Med. J., 124: 124-125.
Johnston, M. A., Porter, D. E., Scott, G. I., Rhodes, W. E., y Webster, L. F. 2010.
Isolation of faecal coliform bacteria from the American alligator (Alligator
mississippiensis). J. Appl. Microbiol., 108(3): 965-973.
Jonas, D., Speck, M., Daschner, F. D., y Grundmann, H. 2002. Rapid PCR-based
identification of methicillin-resistant Staphylococcus aureus from screening swabs. J.
Clin. Microbiol., 40(5): 1821-1823.
Katayama, Y., Ito, T., y Hiramatsu, K. 2000. A new class of genetic element,
staphylococcus cassette chromosome mec, encodes methicillin resistance in
Staphylococcus aureus. Antimicrob. Agents. Chemother., 44(6): 1549-1555.
Kollef, M. H. 2007. Limitations of vancomycin in the management of resistant
staphylococcal infections. Clin. Infect. Dis., 45: 191-195.
Kommanee, J., Preecharram, S., Daduang, S., Temsiripong, Y., Dhiravisit, A., Yamada,
Y., y Thammasirirak, S. 2012. Antibacterial activity of plasma from crocodile
(Crocodylus siamensis) against pathogenic bacteria. Ann. Clin. Microbiol.
Antimicrob., 11(1): 22.
Kumar, R., Yadav, B. R., y Singh, R. S. 2010. Genetic determinants of antibiotic
resistance in Staphylococcus aureus isolates from milk of mastitic crossbred
cattle. Curr. Microbiol., 60(5): 379-386.
Lakshminarayanan, R., Vivekanandan, S., Samy, R. P., Banerjee, Y., Chi-Jin, E. O.,
Teo, K. W., Jois, S., Kini, R. M., y Valiyaveettil, S. 2008. Structure, self-assembly, and

63
dual role of a β-defensin-like peptide from the Chinese soft-shelled turtle eggshell
matrix. J. Am. Chem. Soc., 130(14): 4660-4668.
Leelawongtawon, R., Siruntawineti, J., Chaeychomsri, W., y Sattaponpan, C. 2010.
Antibacterial and antifungal activities from Siamese crocodile blood. J. Med. Assoc.
Thai= Chotmaihet thangphaet, 93: 58-64.
Lowder, S. K., Skoet, J., y Raney, T. 2016. The number, size, and distribution of farms,
smallholder farms, and family farms worldwide. World Dev., 87: 16-29.
Lozano-Soto, F., Engel, P., Rodríguez, S., Jaraquemada, D., y Gallart-Gallart, M. T.
2012. Introducción al sistema inmunológico, sus principales elementos y la respuesta
inmunitaria. Publisher: Elsevier, España.
Luyt, C. E., Bréchot, N., Trouillet, J. L., y Chastre, J. 2014. Antibiotic stewardship in the
intensive care unit. Crit. Care., 18(5): 480.
Machha, V., Spencer, P., y Merchant, M. 2011. Effects of Leukocyte Extract from the
American Alligator (Alligator mississippiensis) on Antibiotic-Resistant Bacteria. Open
Zoology Journal, 4: 9-13.
Medzhitov, R., y Janeway, C. A. 1997. Innate immunity: the virtues of a nonclonal
system of recognition. Cell, 91(3): 295-298.
Medzhitov, R., y Janeway, C. A. 2000. Inmunidad innata. New England Journal of
Medicine, 343(5): 338-344.
Merchant, M., y Britton, A. 2006. Characterization of serum complement activity of
saltwater (Crocodylus porosus) and freshwater (Crocodylus johnstoni)
crocodiles. Comp. Biochem. Physiol. Part A: Molecular & Integrative
Physiology, 143(4): 488-493.
Merchant, M. E., Roche, C., Elsey, R. M., y Prudhomme, J. 2003. Antibacterial properties
of serum from the American alligator (Alligator mississippiensis). Comp. Biochem.
Physiol. Part B: Biochemistry and Molecular Biology, 136(3): 505-513.
Merchant, M., Thibodeaux, D., Loubser, K., y Elsey, R. M. 2004. Amoebacidal effects of
serum from the American alligator (Alligator mississippiensis). J. Parasitol., 90(6):
1480-1483.
Merchant, M. E., Verret, B., y Elsey, R. M. 2005. Role of divalent metal ions in serum
complement activity of the American alligator (Alligator mississippiensis). Comp.
Biochem. Physiol. Part B: Biochemistry and Molecular Biology, 141(3): 289-293.
Michael, C. A., Dominey-Howes, D., y Labbate, M. 2014. The antimicrobial resistance
crisis: causes, consequences, and management. Front. Public Health, 2: 145.

64
Mondal, S., y Rai, U. 2001. In vitro effect of temperature on phagocytic and cytotoxic
activities of splenic phagocytes of the wall lizard, Hemidactylus flaviviridis. Comp.
Biochem. Physiol. Part A: Molecular & Integrative Physiology, 129(2-3): 391-398.
Montali, R. J. 1988. Comparative pathology of inflammation in the higher vertebrates
(reptiles, birds and mammals). J. Comp. Pathol., 99: 1-27.
Moreillon, P., Que, Y., y Glauser, M. P. 2005. Staphylococcus aureus. Principles and
Practice of Infectious Diseases, 6a ed. Publisher: Churchill-Livingston, USA. Pp. 2321-
2351.
Mosmann, T. 1983. Rapid colorimetric assay for celular growth and survival: Application
to proliferation and cytotoxicity assay. J. Immunol. Methods, 65: 55-63.
Napolitano, L. M. 2009. Severe soft tissue infections. Infect. Dis. Clin. N. Am., 23(3):
571-591.
Nature, E. 2013. The antibiotic alarm. Nature, 495(7440): 141.
Nishi, J., Miyanohara, H., Nakajima, T., Kitajima, I., Yoshinaga, M., Maruyama, I., &
Miyata, K. 1995. Molecular typing of the methicillin resistance determinant (mec) of
clinical strains of Staphylococcus based on mec hypervariable region length
polymorphisms. J. Lab. Clin. Med., 126(1): 29-35.
Novales, M. G. M. 2011. Resistencia antimicrobiana del Staphylococcus aureus en
México. Bol. Méd. Hosp. Infan. Méx., 68(4): 262-270.
Novick, R. P., Schlievert, P., y Ruzin, A. 2001. Pathogenicity and resistance islands of
staphylococci. Microb. Infect., 3(7): 585-594.
Pata, S., Yaraksa, N., Daduang, S., Temsiripong, Y., Svasti, J., Araki, T., y
Thammasirirak, S. (2011). Characterization of the novel antibacterial peptide Leucrocin
from crocodile (Crocodylus siamensis) white blood cell extracts. Dev. Comp.
Immunol., 35(5): 545-553.
Piddock, L. J. 2012. The crisis of no new antibiotics—what is the way forward?. Lancet
Infect. Dis., 12(3): 249-253.
Piña, C. I., Larriera, A., y Cabrera, M. R. 2003. Effect of incubation temperature on
incubation period, sex ratio, hatching success, and survivorship in Caiman latirostris
(Crocodylia, Alligatoridae). J. Herpetol., 37(1): 199-203.
Prajanban, B. O., Jangpromma, N., Araki, T., y Klaynongsruang, S. (2017).
Antimicrobial effects of novel peptides cOT2 and sOT2 derived from Crocodylus
siamensis and Pelodiscus sinensis ovotransferrins. Biochim. Biophys. Acta (BBA)-
Biomembranes, 1859(5), 860-869.

65
Read, A. F., & Woods, R. J. 2014. Antibiotic resistance management. Evol. Med. Public
Health., 2014(1): 147.
Rivera, J., Vannakambadi, G., Höök, M., y Speziale, P. 2007. Fibrinogen-binding
proteins of Gram-positive bacteria. Thromb. Haemost., 98(3): 503-511.
Robert, J., y Ohta, Y. 2009. Comparative and developmental study of the immune
system in Xenopus. Dev. Dyn., 238(6): 1249-1270.
Schmidt, B. Z., y Colten, H. R. 2000. Complement: a critical test of its biological
importance. Immunol. Rev., 178: 166-176.
Seelen, M. A., Roos, A., y Daha, M. R. 2005. Role of complement in innate and
autoimmunity. J. Nephrol., 18(6): 642-653.
Sengupta, S., Chattopadhyay, M. K., y Grossart, H. P. 2013. The multifaceted roles of
antibiotics and antibiotic resistance in nature. Front. Microbiol., 4: 47.
Siroski, P. 2011. “Caracterización del Sistema del complemento e identificación de
componentes del sistema inmune innato del yacaré overo Caiman latirostris”. Tesis
Doctoral. Universidad Nacional del Litoral. Mayo, 2011.
Siroski, P., Merchant, M., Parachu Marco, V., Piña, C., y Ortega, H. 2010.
Characterization of serum complement activity of the broad snouted caiman (Caiman
latirostris, Crocodilia: Alligatoridae). Zool. Stud., 49(1): 64-70.
Siuntawineti, J., Chaeychomsri, W., Vajarasathira, B., Ong-aat, T., y Temsiripong, Y.
(2004). Efficacy of Siamese crocodile (Crocodylus siamensis) serum on bacterial growth
inhibition. In Proceedings of the 17th Working Meeting of the Crocodile Specialist
Group (pp. 465-468).
Spellberg, B., y Gilbert, D. N. 2014. The future of antibiotics and resistance: a tribute
to a career of leadership by John Bartlett. Clin. Infect. Dis., 59: 71-75.
Sunyer, J. O., y Lambris, J. D. 1998. Evolution and diversity of the complement system
of poikilothermic vertebrates. Immunol. Rev. 166(1); 39-57.
Tenover, F. C., y Goering, R. V. 2009. Methicillin-resistant Staphylococcus aureus strain
USA300: origin and epidemiology. J. Antimicrob. Chemother., 64(3): 441-446.
Thammasirirak, S., Ponkham, P., Preecharram, S., Khanchanuan, R., Phonyothee, P.,
Daduang, S., Araki, T., y Svasti, J. 2006. Purification, characterization and comparison
of reptile lysozymes. Comp. Biochem. Physiol. Part C: Toxicology &
Pharmacology, 143(2): 209-217.

66
ONU. The United Nations. 2016. https://news.un.org/en/story/2016/09/539912-un-
global-leaders-commit-act-antimicrobial-resistance. Recuperado 3 de octubre de 2018.
Tizard, I.R. 2013. Inmunología veterinaria-E-Libro. Publicher: Elsevier, USA.
Tong, S. Y., Davis, J. S., Eichenberger, E., Holland, T. L., y Fowler, V. G. 2015.
Staphylococcus aureus infections: epidemiology, pathophysiology, clinical
manifestations, and management. Clin. Microbial. Rev., 28(3): 603-661.
Tucunduva, M., Borelli, P., y Silva, J. R. M. C. 2001. Experimental study of induced
inflammation in the Brazilian boa (Boa constrictor constrictor). J. Comp.
Pathol., 125(2-3): 174-181.
Ulevitch, R. J., y Tobias, P. S. 1999. Recognition of gram-negative bacteria and
endotoxin by the innate immune system. Curr. Opin. Immunol., 11(1): 19-22.
Ulevitch, R. J., Mathison, J. C., y DaSilva-Correia, J. 2004. Respuestas inmunes innatas
durante la infección. Vacuna, 22 : 25-30.
van Hal, S. J., y Fowler Jr, V. G. 2013. Is it time to replace vancomycin in the treatment
of methicillin-resistant Staphylococcus aureus infections?. Clin. Infect. Dis., 56(12):
1779-1788.
van Hal, S. J., Lodise, T. P., y Paterson, D. L. 2012. The clinical significance of
vancomycin minimum inhibitory concentration in Staphylococcus aureus infections: a
systematic review and meta-analysis. Clin. Infect. Dis., 54(6): 755-771.
Van Hoek, M. L. 2014. Antimicrobial peptides in reptiles. Pharmaceuticals, 7(6): 723-
753.
Viswanathan, V. K. 2014. Off-label abuse of antibiotics by bacteria. Journal Gut
Microbes, 5: 3-4.
Waksman, S. A. 1947. What is an antibiotic or an antibiotic
substance?. Mycologia, 39(5): 565-569.
Wang, Y., Wu, C. M., Lu, L. M., Ren, G. W. N., Cao, X. Y., y Shen, J. Z. 2008. Macrolide–
lincosamide-resistant phenotypes and genotypes of Staphylococcus aureus isolated
from bovine clinical mastitis. Vet. Microbial., 130(1-2): 118-125.
Wayne, P. A. 2011. Clinical and laboratory standards institute. Performance standards
for antimicrobial susceptibility testing.
Webb, G. J. W., Manolis, S. C., y Whitehead, P. J. 1987. Wildlife management:
crocodiles and alligators. Publisher: Surrey Beatty and Sons, Australia. , Australia.

67
Weems, J. J., Steinberg, J. P., Filler, S., Baddley, J. W., Corey, G. R., Sampathkumar,
P., Winston, L., John, J. F., Kubin, C. J., Talwani, R., y Moore, T. 2006. Phase II,
randomized, double-blind, multicenter study comparing the safety and
pharmacokinetics of tefibazumab to placebo for treatment of Staphylococcus aureus
bacteremia. Antimicrob. Agents Chemother., 50(8): 2751-2755.
Wertheim, H. F., Melles, D. C., Vos, M. C., van Leeuwen, W., van Belkum, A., Verbrugh,
H. A., y Nouwen, J. L. 2005. The role of nasal carriage in Staphylococcus aureus
infections. Lancet Infect. Dis., 5(12): 751-762.
Winn, W., Allen, S., Janda, W. M., Koneman, E. W., Procop, C. W., Schreckenberger,
P. C., y Woods, G. 2006. Gram-positive cocci Part 1: Staphylococci and related Gram-
positive cocci. Koneman's Color Atlas and Textbook of Diagnostic Microbiology.
Publisher: Lippincott Williams & Wilkins, USA. Pp. 623-671.
Organización Mundial de la Salud (World Health Organization). 2001. Estrategia
mundial de la OMS para contener la resistencia a los antimicrobianos (No.
WHO/CDS/CSR/DRS/2001.2). Ginebra: Organización Mundial de la Salud.
Zimmerman, L. M., Vogel, L. A., y Bowden, R. M. 2010. Understanding the vertebrate
immune system: insights from the reptilian perspective. J. Exp. Biol., 213(5): 661-
671.


















