
Folding and self-assembly of the domains of βB2-crystallin from rat eye lens
6
COMMUNICATION Folding and Self-assembly of the Domains of bB2-Crystallin from Rat Eye Lens Karin Wieligmann, Eva-Maria Mayr and Rainer Jaenicke* Institut fu ¨ r Biophysik und Physikalische Biochemie Universita ¨t Regensburg D-93040 Regensburg, Germany bB2-Crystallin from vertebrate eye lens forms domain-swapped dimers, with subunits consisting of two all-b domains connected by an eight-resi- due extended linker peptide. Topologically, the two domains show great similarity; however, they differ widely in their stability. As shown by urea-induced equilibrium unfolding experiments, the isolated monomeric C-terminal domain is more stable than complete bB2. In contrast, the N- terminal domain exhibits marginal stability only in its dimeric state; upon subunit dissociation, at low protein concentration, unfolding takes place. The folding and association of intact bB2 follows a sequential uni- bimolecular mechanism according to N 2 2I 2U, whereas the isolated domains may be quantitatively described by the two-state model (N . U). # 1999 Academic Press Keywords: association; crystallins; domains; folding; stability *Corresponding author a-Crystallin and the superfamily of the bg-crys- tallins are major structural proteins in the ver- tebrate eye lens. They are responsible for both lens transparency and the high refractive index required for image formation on the retina. The b and g-crystallins are all-b two-domain proteins with a high level of symmetry due to the identical overall topology of their N and C-terminal domains. Each of them consists of two Greek-key motifs that intercalate two four-stranded anti-par- allel b-sheets (Blundell et al., 1981; Wistow et al., 1983; White et al., 1989; Bax et al., 1990; Lapatto et al., 1991). In spite of their autonomous folding behaviour and high intrinsic stability, so far iso- lated domains of crystallins or their homologues have not been discovered either in vertebrate eye lens or at lower levels of evolution (Wistow & Piatigorsky, 1988; Jaenicke, 1994, 1999). On the other hand, higher states of association have been found as a consequence of molecular domain swapping (Schlunegger et al., 1997; Jaenicke, 1999). In the case of bB2-crystallin, mutual interactions between the N and C-terminal domains of two polypeptide chains cause dimer formation (Bax et al., 1990). Given the close similarity of the domains and their tendency to form pairs, one may ask what are the contributions of the separate domains and their mutual interactions to the over- all intrinsic stability of bB2-crystallin. Both problems refer to the equilibrium unfolding properties (e.g. in the presence of urea) and to the folding pathway, which may be determined either by rate-limiting isomerization reactions at the domain or subunit level or by subunit association. In order to answer these questions, recombinant bB2-crystallin from rat eye lens and its isolated N and C-terminal domains were studied. Intact bB2-crystallin (bB2) and its isolated N and C-terminal domains (bB2N and bB2C) were recom- binantly expressed in Escherichia coli and purified to homogeneity (see the legend to Figure 1). In the case of bB2N, the fragment with and without the C-terminal ‘‘connecting peptide’’ was prepared. The proteins do not differ in their physicochemical properties (data not shown); in the following, we restrict ourselves to bB2N, with His88 as the C-terminal end of the connecting peptide (Table 1; numbering of the residues is according to Bax et al. (1990) and Trinkl et al. (1994)). In order to characterize the native and denatured states, absorption, fluorescence emission and circu- lar dichroism (CD) spectroscopy were applied. The UV-absorption spectra of the three proteins reflect the differences in the content of aromatic residues (Figure 1(a)): The maximum absorbance at 280 nm and the shoulder at 290 nm in the case of bB2 and E-mail address of the corresponding author: [email protected] Abbreviations used: bB2, bB2N and bB2C: intact bB2, recombinant N-terminal and C-terminal domains of bB2; CD, circular dichroism. Article No. jmbi.1999.2554 available online at http://www.idealibrary.com on J. Mol. Biol. (1999) 286, 989–994 0022-2836/99/090989–06 $30.00/0 # 1999 Academic Press
-
Upload
karin-wieligmann -
Category
Documents
-
view
216 -
download
0
Transcript of Folding and self-assembly of the domains of βB2-crystallin from rat eye lens

Article No. jmbi.1999.2554 available online at http://www.idealibrary.com on J. Mol. Biol. (1999) 286, 989±994
COMMUNICATION
Folding and Self-assembly of the Domains of
bbbB2-Crystallin from Rat Eye LensKarin Wieligmann, Eva-Maria Mayr and Rainer Jaenicke*
Institut fuÈ r Biophysik undPhysikalische BiochemieUniversitaÈt RegensburgD-93040 Regensburg, Germany
*Corresponding author
E-mail address of the [email protected]
Abbreviations used: bB2, bB2Nrecombinant N-terminal and C-terCD, circular dichroism.
0022-2836/99/090989±06 $30.00/0
bB2-Crystallin from vertebrate eye lens forms domain-swapped dimers,with subunits consisting of two all-b domains connected by an eight-resi-due extended linker peptide. Topologically, the two domains show greatsimilarity; however, they differ widely in their stability. As shown byurea-induced equilibrium unfolding experiments, the isolated monomericC-terminal domain is more stable than complete bB2. In contrast, the N-terminal domain exhibits marginal stability only in its dimeric state;upon subunit dissociation, at low protein concentration, unfolding takesplace. The folding and association of intact bB2 follows a sequential uni-bimolecular mechanism according to N2� 2 I� 2U, whereas the isolateddomains may be quantitatively described by the two-state model(N . U).
# 1999 Academic Press
Keywords: association; crystallins; domains; folding; stability
a-Crystallin and the superfamily of the bg-crys-tallins are major structural proteins in the ver-tebrate eye lens. They are responsible for both lenstransparency and the high refractive indexrequired for image formation on the retina. Theb and g-crystallins are all-b two-domain proteinswith a high level of symmetry due to the identicaloverall topology of their N and C-terminaldomains. Each of them consists of two Greek-keymotifs that intercalate two four-stranded anti-par-allel b-sheets (Blundell et al., 1981; Wistow et al.,1983; White et al., 1989; Bax et al., 1990; Lapattoet al., 1991). In spite of their autonomous foldingbehaviour and high intrinsic stability, so far iso-lated domains of crystallins or their homologueshave not been discovered either in vertebrate eyelens or at lower levels of evolution (Wistow &Piatigorsky, 1988; Jaenicke, 1994, 1999). On theother hand, higher states of association have beenfound as a consequence of molecular domainswapping (Schlunegger et al., 1997; Jaenicke, 1999).In the case of bB2-crystallin, mutual interactionsbetween the N and C-terminal domains of twopolypeptide chains cause dimer formation (Baxet al., 1990). Given the close similarity of the
ding author:burg.de
and bB2C: intact bB2,minal domains of bB2;
domains and their tendency to form pairs, onemay ask what are the contributions of the separatedomains and their mutual interactions to the over-all intrinsic stability of bB2-crystallin.
Both problems refer to the equilibrium unfoldingproperties (e.g. in the presence of urea) and to thefolding pathway, which may be determined eitherby rate-limiting isomerization reactions at thedomain or subunit level or by subunit association.In order to answer these questions, recombinantbB2-crystallin from rat eye lens and its isolated Nand C-terminal domains were studied.
Intact bB2-crystallin (bB2) and its isolated N andC-terminal domains (bB2N and bB2C) were recom-binantly expressed in Escherichia coli and puri®edto homogeneity (see the legend to Figure 1). In thecase of bB2N, the fragment with and without theC-terminal ``connecting peptide'' was prepared.The proteins do not differ in their physicochemicalproperties (data not shown); in the following, werestrict ourselves to bB2N, with His88 as theC-terminal end of the connecting peptide (Table 1;numbering of the residues is according to Bax et al.(1990) and Trinkl et al. (1994)).
In order to characterize the native and denaturedstates, absorption, ¯uorescence emission and circu-lar dichroism (CD) spectroscopy were applied. TheUV-absorption spectra of the three proteins re¯ectthe differences in the content of aromatic residues(Figure 1(a)): The maximum absorbance at 280 nm
and the shoulder at 290 nm in the case of bB2 and# 1999 Academic Press
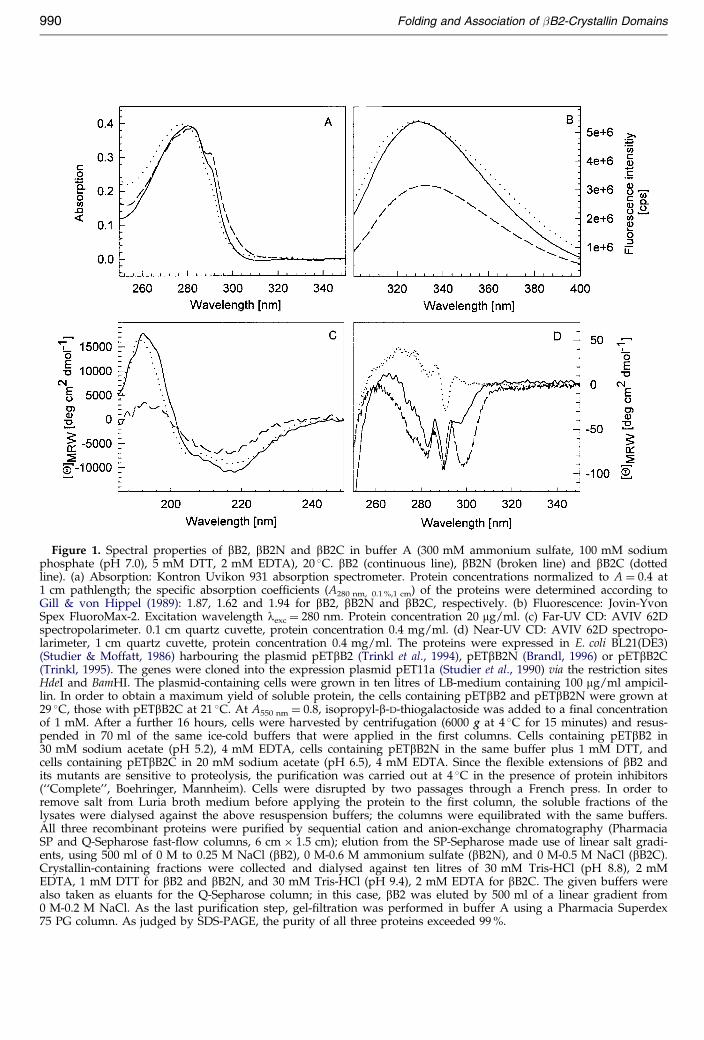
Figure 1. Spectral properties of bB2, bB2N and bB2C in buffer A (300 mM ammonium sulfate, 100 mM sodiumphosphate (pH 7.0), 5 mM DTT, 2 mM EDTA), 20 �C. bB2 (continuous line), bB2N (broken line) and bB2C (dottedline). (a) Absorption: Kontron Uvikon 931 absorption spectrometer. Protein concentrations normalized to A � 0.4 at1 cm pathlength; the speci®c absorption coef®cients (A280 nm, 0.1 %,1 cm) of the proteins were determined according toGill & von Hippel (1989): 1.87, 1.62 and 1.94 for bB2, bB2N and bB2C, respectively. (b) Fluorescence: Jovin-YvonSpex FluoroMax-2. Excitation wavelength lexc � 280 nm. Protein concentration 20 mg/ml. (c) Far-UV CD: AVIV 62Dspectropolarimeter. 0.1 cm quartz cuvette, protein concentration 0.4 mg/ml. (d) Near-UV CD: AVIV 62D spectropo-larimeter, 1 cm quartz cuvette, protein concentration 0.4 mg/ml. The proteins were expressed in E. coli BL21(DE3)(Studier & Moffatt, 1986) harbouring the plasmid pETbB2 (Trinkl et al., 1994), pETbB2N (Brandl, 1996) or pETbB2C(Trinkl, 1995). The genes were cloned into the expression plasmid pET11a (Studier et al., 1990) via the restriction sitesHdeI and BamHI. The plasmid-containing cells were grown in ten litres of LB-medium containing 100 mg/ml ampicil-lin. In order to obtain a maximum yield of soluble protein, the cells containing pETbB2 and pETbB2N were grown at29 �C, those with pETbB2C at 21 �C. At A550 nm � 0.8, isopropyl-b-D-thiogalactoside was added to a ®nal concentrationof 1 mM. After a further 16 hours, cells were harvested by centrifugation (6000 g at 4 �C for 15 minutes) and resus-pended in 70 ml of the same ice-cold buffers that were applied in the ®rst columns. Cells containing pETbB2 in30 mM sodium acetate (pH 5.2), 4 mM EDTA, cells containing pETbB2N in the same buffer plus 1 mM DTT, andcells containing pETbB2C in 20 mM sodium acetate (pH 6.5), 4 mM EDTA. Since the ¯exible extensions of bB2 andits mutants are sensitive to proteolysis, the puri®cation was carried out at 4 �C in the presence of protein inhibitors(``Complete'', Boehringer, Mannheim). Cells were disrupted by two passages through a French press. In order toremove salt from Luria broth medium before applying the protein to the ®rst column, the soluble fractions of thelysates were dialysed against the above resuspension buffers; the columns were equilibrated with the same buffers.All three recombinant proteins were puri®ed by sequential cation and anion-exchange chromatography (PharmaciaSP and Q-Sepharose fast-¯ow columns, 6 cm � 1.5 cm); elution from the SP-Sepharose made use of linear salt gradi-ents, using 500 ml of 0 M to 0.25 M NaCl (bB2), 0 M-0.6 M ammonium sulfate (bB2N), and 0 M-0.5 M NaCl (bB2C).Crystallin-containing fractions were collected and dialysed against ten litres of 30 mM Tris-HCl (pH 8.8), 2 mMEDTA, 1 mM DTT for bB2 and bB2N, and 30 mM Tris-HCl (pH 9.4), 2 mM EDTA for bB2C. The given buffers werealso taken as eluants for the Q-Sepharose column; in this case, bB2 was eluted by 500 ml of a linear gradient from0 M-0.2 M NaCl. As the last puri®cation step, gel-®ltration was performed in buffer A using a Pharmacia Superdex75 PG column. As judged by SDS-PAGE, the purity of all three proteins exceeded 99 %.
990 Folding and Association of �B2-Crystallin Domains

Table 1. Molecular masses of bB2-crystallin and its isolated domains bB2N and bB2C in buffer A
Sedimentation analysis
Protein Amino acid residues Molecular massa Mcalc (kDa) s20,w (S)b Mw (kDa)c State of association
bB2 205 (ÿ16-185) 23.38 3.04 � 0.05 42.4 � 2.2 DimerbB2N 107 (ÿ16-88) 12.03 1.85 � 0.08 19.8 � 3.1 Dimerd
bB2C 99 (88-185) 11.64 1.21 � 0.05 10.6 � 1.0 Monomer
Determination of the molecular mass of bB2, bB2N and bB2C. Sedimentation velocity and sedimentation equilibrium measure-ments were performed in a Beckman model E analytical ultracentrifuge, making use of 12 mm double-sector cells with sapphire win-dows in an AN-G rotor. Initial protein concentrations, 300 mg/ml. In order to control homogeneity, the meniscus depletiontechnique was applied (Yphantis, 1964). Experiments were performed at room temperature using a scanning wavelength of 280 nm.s-values were determined at 15,000 and 44,000 rev/min, plotting ln r versus time and correcting for water viscosity and 20 �C. Sedi-mentation equilibria at 24,000 and 30,000 rev/min were calculated from ln c versus r2 plots. The partial speci®c volumes were calcu-lated from the amino acid composition: 0.719 , 0.717 and 0.720 cm3 gÿ1 for bB2, bB2N and bB2C, respectively.
a Molecular masses calculated from the amino acid sequences using the UWGCG program.b s20,w, sedimentation coef®cient at 300 mg/ml from sedimentation velocity experiments.c Mw, molecular mass from high-speed sedimentation equilibria; correlation coef®cient for ln r versus r2 linearization >0.999.d At protein concentrations <50 mg/ml, dissociation to the monomer becomes detectable.
Folding and Association of �B2-Crystallin Domains 991
bB2N are attributable to the relatively high contentof tryptophan residues (®ve in bB2 and three inbB2N). In contrast, the spectrum of bB2C (withseven tyrosine and two tryptophan residues) isdetermined mainly by the absorption of tyrosine;accordingly, its maximum absorbance is shifted to277 nm, and no shoulder at 290 nm is detectable.The ¯uorescence emission spectra of all three pro-teins (Figure 1(b)) are dominated by the trypto-phan content, which provides a means to monitorconformational alterations upon unfolding/foldingof the domains on the one hand, and domain andsubunit interactions upon dissociation/association,on the other. At low protein concentration(�20 mg/ml) and low ionic strength (100 mMsodium phosphate (pH 7.0), 5 mM DTT, 2 mMEDTA) bB2N undergoes partial denaturation, indi-cated by a ¯uorescence maximum at 346 nm com-pared to the maximum for the unfolded protein at355(�1) nm. In the presence of high salt (buffer A;300 mM ammonium sulfate plus 100 mM sodiumphosphate (pH 7.0), 5 mM DTT, 2 mM EDTA), sig-ni®cant stabilization is accomplished. Therefore,this solvent was used as standard solvent in thepresent study, also for intact bB2 and bB2C.
As illustrated by the dichroic absorption in thefar-UV, all three proteins show all-b characteristics(Figure 1(c)). The near-UV CD spectra are indica-tive of well-de®ned tertiary structures for all threeproteins (Figure 1(d)). Considering the differences,it is tempting to attribute the minimum at 330 nmobserved for the N-terminal domain to speci®cstructural characteristics of this protein; it may beconnected with the anomalous association beha-viour that is observed exclusively for bB2N.
The state of association of recombinant bB2 andits isolated domains was determined by analyticalultracentrifugation (Table 1). The results show thatin buffer A, and at an initial concentration of300 mg/ml, bB2N forms a stable dimer, whereasbB2C is monomeric; no tendency to form higheroligomers was detected. Below a critical concen-tration of �50 mg/ml and at low salt, bB2N dis-sociates to the monomer. Figure 2(a) illustrates the
concentration-dependence of the maximum of ¯u-orescence emission, clearly demonstrating that atlow concentration the protein loses its native struc-ture.
The urea-induced equilibrium denaturation con-®rms this observation. Shifting the protein concen-tration from 200 to 5 mg/ml, the midpoint ofdenaturation is decreased from curea � 1.2 M to0.7 M (Figure 2(b)). A comparison of the stabilitiesof intact bB2 and its isolated domains is given inFigure 3; the equilibrium transitions of all threeproteins are fully reversible. bB2C is found to exhi-bit enhanced stability compared to its intrinsicstability within the complete molecule. Obviously,the stabilization of bB2N is accompanied by a sig-ni®cant strain of the polypeptide chain within thecomplete dimeric bB2 molecule. Apart from themutual destabilizing and stabilizing effects, thetransitions of the single domains show a lowdegree of cooperativity, in agreement with unfold-ing transitions reported for other small single-chainproteins (Myers et al., 1995).
In order to gain insight into the involvement ofquaternary interactions in the unfolding and fold-ing of bB2, equilibrium unfolding transitions weremeasured at varying concentrations of protein. Asshown by ¯uorescence emission at 316 nm, thecooperativity increases with increasing protein con-centration (Figure 4(a)), in contrast to the two-statemodel for the folding reaction of a dimeric protein,which would predict identical cooperativities andshifts of the midpoints of transition at varying pro-tein concentrations. Monitoring the urea-induceddenaturation at high protein concentration also byfar-UV CD, the perturbations of the secondary andtertiary structure are found to parallel each other(data not shown). The comparison of the ¯uor-escence spectra of intact bB2 at 5 and 200 mg/ml inFigure 4(b) shows that at low protein concentrationa monomeric intermediate becomes signi®cantlypopulated. At high concentration, all spectracoincide in a single point of identical ¯uorescenceemission, in agreement with the two-state model:the equilibrium favours the dimer, so that the
monomer escapes detection.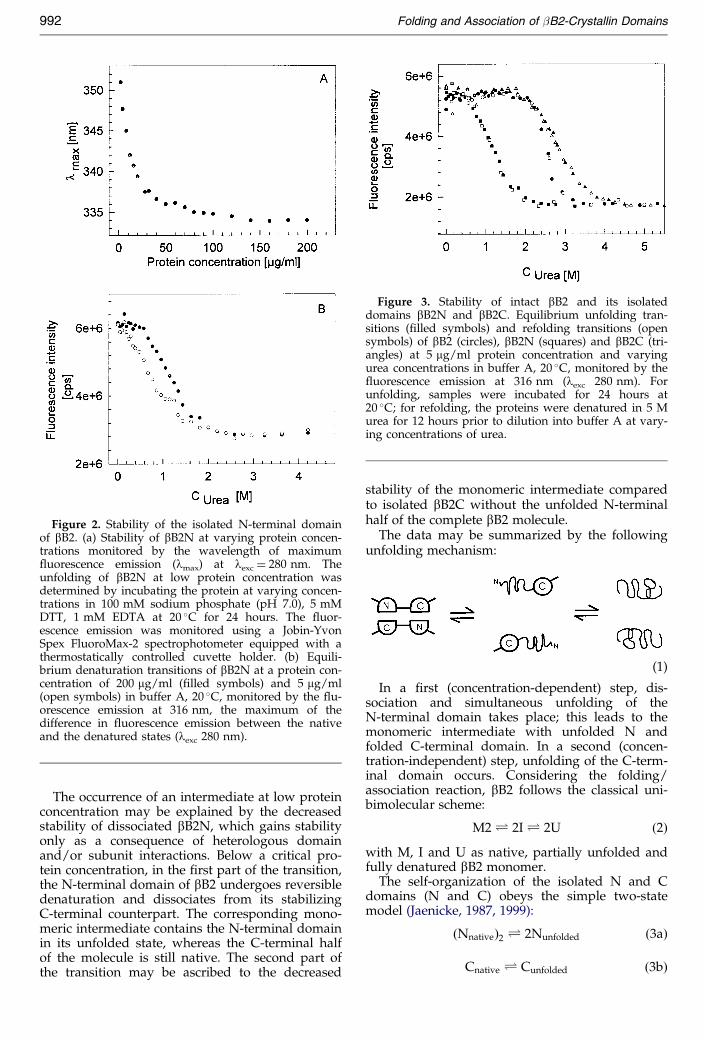
C � C �3b�
Figure 2. Stability of the isolated N-terminal domainof bB2. (a) Stability of bB2N at varying protein concen-trations monitored by the wavelength of maximum¯uorescence emission (lmax) at lexc � 280 nm. Theunfolding of bB2N at low protein concentration wasdetermined by incubating the protein at varying concen-trations in 100 mM sodium phosphate (pH 7.0), 5 mMDTT, 1 mM EDTA at 20 �C for 24 hours. The ¯uor-escence emission was monitored using a Jobin-YvonSpex FluoroMax-2 spectrophotometer equipped with athermostatically controlled cuvette holder. (b) Equili-brium denaturation transitions of bB2N at a protein con-centration of 200 mg/ml (®lled symbols) and 5 mg/ml(open symbols) in buffer A, 20 �C, monitored by the ¯u-orescence emission at 316 nm, the maximum of thedifference in ¯uorescence emission between the nativeand the denatured states (lexc 280 nm).
Figure 3. Stability of intact bB2 and its isolateddomains bB2N and bB2C. Equilibrium unfolding tran-sitions (®lled symbols) and refolding transitions (opensymbols) of bB2 (circles), bB2N (squares) and bB2C (tri-angles) at 5 mg/ml protein concentration and varyingurea concentrations in buffer A, 20 �C, monitored by the¯uorescence emission at 316 nm (lexc 280 nm). Forunfolding, samples were incubated for 24 hours at20 �C; for refolding, the proteins were denatured in 5 Murea for 12 hours prior to dilution into buffer A at vary-ing concentrations of urea.
992 Folding and Association of �B2-Crystallin Domains
The occurrence of an intermediate at low proteinconcentration may be explained by the decreasedstability of dissociated bB2N, which gains stabilityonly as a consequence of heterologous domainand/or subunit interactions. Below a critical pro-tein concentration, in the ®rst part of the transition,the N-terminal domain of bB2 undergoes reversibledenaturation and dissociates from its stabilizingC-terminal counterpart. The corresponding mono-meric intermediate contains the N-terminal domainin its unfolded state, whereas the C-terminal halfof the molecule is still native. The second part of
the transition may be ascribed to the decreasedstability of the monomeric intermediate comparedto isolated bB2C without the unfolded N-terminalhalf of the complete bB2 molecule.
The data may be summarized by the followingunfolding mechanism:
(1)
In a ®rst (concentration-dependent) step, dis-sociation and simultaneous unfolding of theN-terminal domain takes place; this leads to themonomeric intermediate with unfolded N andfolded C-terminal domain. In a second (concen-tration-independent) step, unfolding of the C-term-inal domain occurs. Considering the folding/association reaction, bB2 follows the classical uni-bimolecular scheme:
M2� 2I� 2U �2�with M, I and U as native, partially unfolded andfully denatured bB2 monomer.
The self-organization of the isolated N and Cdomains (N and C) obeys the simple two-statemodel (Jaenicke, 1987, 1999):
�Nnative�2 � 2Nunfolded �3a�
native unfolded

Figure 4. Stability of intact bB2-crystallin at varyingprotein concentrations in buffer A, 20 �C. (a) Equilibriumdenaturation transitions of bB2 at 200 mg/ml (®lled sym-bols) and 5 mg/ml (open symbols), monitored by ¯uor-escence emission at 316 nm (lexc 280 nm). Continuouslines are meant only to guide the eye of the reader.Inset: Wavelengths of maximum ¯uorescence emission(lmax) of bB2 at varying protein concentrations in bufferA containing 2.15 M urea. (b) Fluorescence spectra ofbB2 after 24 hours incubation at 0 M (Ð), 0.9 M(Ð Ð), 2.2 M (± ± ±), 2.6 M ( � �Ð � �Ð) and 5 M( � � � � � ) urea, 20 �C. Upper frame. 200 mg/ml; lowerframe, 5 mg/ml.
Folding and Association of �B2-Crystallin Domains 993
For bB2N at low concentration, the equilibrium isshifted to the dissociated and unfolded monomer.
Acknowledgements
This work was supported by the Deutsche For-schungsgemeinschaft, the Fonds der Chemischen Indu-strie and the German-Israel Foundation. We thank MariaBrandl and Sabine Trinkl for providing plasmids.
References
Bax, B., Lapatto, R., Nalini, V., Driessen, H., Lindley,P. F., Makadevan, D., Blundell, T. L. & Slingsby, C.(1990). X-ray analysis of bB2-crystallin and evolu-tion of oligomeric lens proteins. Nature, 347, 776-780.
Blundell, T. L., Lindley, P., Miller, L., Moss, D.,Slingsby, C., Tickley, I., Turnell, B. & Wistow, G.(1981). The molecular structure and stability of theeye lens: X-ray analysis of c-crystallin II. Nature,289, 771-777.
Brandl, M. (1996). Charakterisierung der N-terminalenDomaÈne von bB2-Kristallin aus der Augenlinse,Diploma thesis, University of Regensburg.
Gill, S. C. & von Hippel, P. (1989). Calculation of pro-tein extinction coef®cients from amino acidsequence data. Anal. Biochem. 182, 319-326.
Jaenicke, R. (1987). Folding and association of proteins.Prog. Biophys. Mol. Biol. 49, 127-237.
Jaenicke, R. (1994). Eye-lens proteins: structure, super-structure, stability and genetics. Naturwissenschaften,81, 423-429.
Jaenicke, R. (1999). Folding and stability of domainproteins. Prog. Biophys. Mol. Biol. 71, 155-241.
Lapatto, R., Nalini, V., Bax, B., Driessen, H., Lindley,P. F., Blundell, T. & Slingsby, C. (1991). High-resol-ution structure of an oligomeric eye lens b-crystal-lin. Loops, arches, linkers and interfaces in bB2dimer compared to a monomeric g-crystallin. J. Mol.Biol. 222, 1067-1083.
Myers, J. K., Pace, C. N. & Scholtz, J. M. (1995). Dena-turant m values and heat capacity changes: relationto changes in accessible surface areas of proteinunfolding. Protein Sci. 4, 2138-2148.
Schlunegger, M. P., Bennett, M. J. & Eisenberg, D.(1997). Oligomer formation by 3D domain swap-ping: a model for protein assembly and misassem-bly. Advan. Protein Chem. 50, 61-122.
Studier, F. W. & Moffatt, B. A. (1986). Use of the bac-teriophage T7 RNA polymerase to direct selectivehigh-level expression of cloned genes. J. Mol. Biol.189, 113-130.
Studier, F. W., Rosenberg, A. H., Dunn, J. J. &Dubendorff, J. W. (1990). Use of T7 RNA polymer-ase to direct expression of cloned genes. MethodsEnzymol. 185, 60-89.
Trinkl, S. (1995). Ein¯uû von Strukturelementen aufAssoziation und StabilitaÈ t von bB2-Kristallin. Dis-sertation, University of Regensburg.
Trinkl, S., Glockshuber, R. & Jaenicke, R. (1994). Dimeri-zation of bB2-crystallin: the role of the linker pep-tide and the N- and C-terminal extensions. ProteinSci. 3, 1392-1400.
White, H. E., Driessen, H. P. C., Slingsby, C., Moss, D. S.
& Lindley, P. F. (1989). Packing interactions in the
994 Folding and Association of �B2-Crystallin Domains
eye lens. Structural analysis, internal symmetry andlattice interactions of bovine gIVa-crystallin. J. Mol.Biol. 201, 217-235.
Wistow, G. J. & Piatigorsky, J. (1988). Lens crystallins:the evolution and expression of proteins from ahighly specialized tissue. Annu. Rev. Biochem. 57,
479-504.Wistow, G. J., Turnell, B., Summers, L., Slingsby, C.,Moss, D., Miller, L., Lindley, P. & Blundell, T.(1983). X-ray analysis of the eye lens protein gII-crystallin at 1.9 AÊ resolution. J. Mol. Biol. 170, 175-202.
Yphantis, D. Y. (1964). Equilibrium ultracentrifugation
of dilute solutions. Biochemistry, 3, 297-317.Edited by R. Huber
(Received 23 October 1998; received in revised form 8 January 1999; accepted 15 January 1999)


![Characterization of an antibody that recognizes peptides ... · in αA-crystallin (Asp 58 and Asp 151) [3], αB-crystallin (Asp 36 and Asp 62) [4], and βB2-crsytallin (Asp 4) [5]](https://static.fdocument.pub/doc/165x107/5ff1e68e89243b57b64135f8/characterization-of-an-antibody-that-recognizes-peptides-in-a-crystallin-asp.jpg)















