
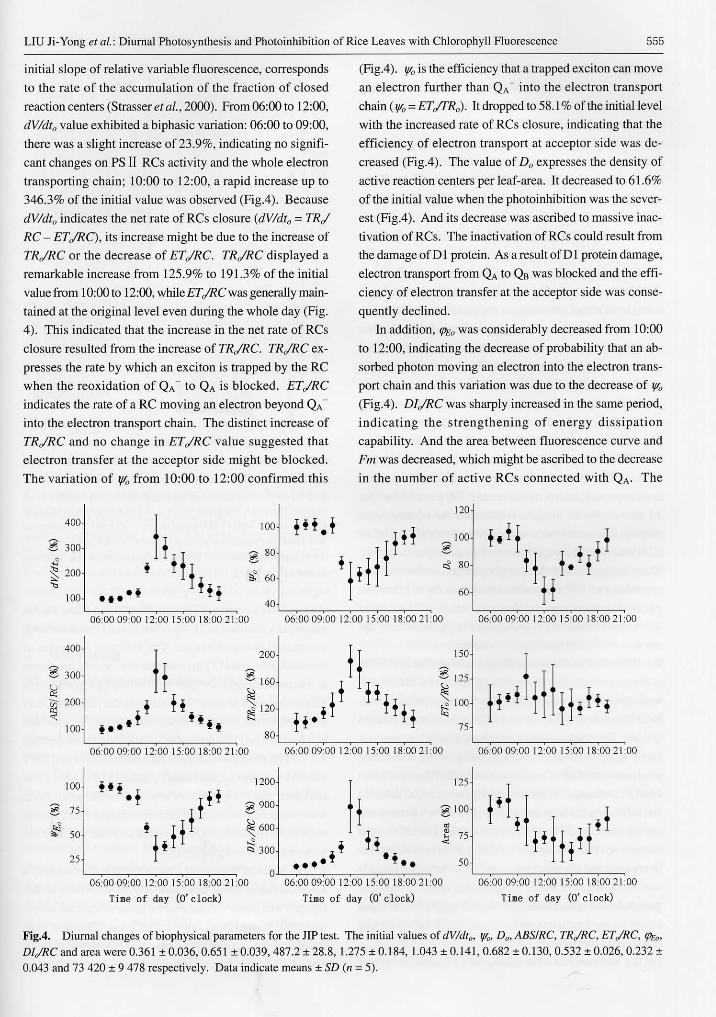
Diurnal Photosynthesis and Photoinhibition of Rice … is uncertain whether the speedup of ATP con-...
8
Received 8 Sept. 2003 Accepted 7 Jan. 2004 Supported by the National Natural Science Foundation of China (30200021). * Author for correspondence. E-mail: <[email protected]>. http://www.chineseplantscience.com Acta Botanica Sinica 植 物 学 报 2004, 46 (5): 552- 559 Diurnal Photosynthesis and Photoinhibition of Rice Leaves with Chlorophyll Fluorescence LIU Ji-Yong 1 , QIU Bao-Sheng 1, 2* , LIU Zhi-Li 2 , YANG Wan-Nian 1 (1. College of Life Sciences, Central China Normal University, Wuhan 430079, China; 2. College of Life Sciences, Nanjing University, Nanjing 210093, China) Abstract: The diurnal photosynthesis and photoinhibition in the photoperiod-sensitive genic male-sterile rice (Oryza sativa L.), Nongken 58S (NK58S), were investigated. From 06:00 to 09:00, no remarkable photoinhibition occurred, and the down-regulation of photosynthesis might be due to the running of xanthophylls cycle. From 10:00 to 12:00, the specific energy flux for dissipation (DI o /RC) and the net rate of reaction centers (RCs) closure (dV/dt o ) were increased, while the probability of electron transport at the acceptor side (y o ) and the density of active RCs (D o ) were decreased. These indicated that the photoinhibition of NK58S was exacerbated with the inactivation of PSⅡ RCs. Fluorescence dark relaxation analysis and inhibitor treatment suggested that all of state transition, xanthophyll cycle and inactivation of PSⅡ RCs could contribute to protect NK58S against photodamage. Compared with the inactivation of PSⅡRCs, xanthophyll cycle had an immediate response to high light stress, which functioned mainly in the period of relatively low light intensity. However, the inactivation of PSⅡ RCs played an important role in protecting the remaining active RCs when xanthophyll cycle was saturated. Key words: photoperiod-sensitive genic male-sterile rice; Nongken 58S; JIP test; photoinhibition; reaction center inactivation; D1 protein turnover; xanthophylls cycle Higher plants are usually encountered with light stress in their habitats. Strong irradiance will inevitably induce photoinhibition with the decrease of photochemical effi- ciency when the absorbed energy is much higher than that utilized by the photosynthetic organs (Long et al., 1994). The photoinhibition will occur much frequently when strong irradiance is combined with high temperature, drought or other stresses in summer midday. Furthermore, the photoinhibition could result in photooxidative damage, pig- ment bleaching and even irreversible damage to the photo- synthetic apparatus. Numerous protective mechanisms have been developed to minimize the sunlight absorption in long-term evolutionary history, such as alteration in leaves angle, chloroplast movement and reflective cuticle covering. Alternatively, state transition, xanthophylls cycle and PSⅡ repair cycle contribute internally to avoiding the damage to the photosynthetic apparatus (Long et al., 1994; Horton et al., 1996). However, various species might differ in their protective mechanisms for photoinhibition. Photoinhibition is characterized by the decline of pho- tosynthetic quantum efficiency and photochemical efficiency, and Fv/Fm value is widely used as an index to evaluate the extent of photoinhibition (Krause and Weis, 1991; Govindjee, 1995; Maxwell and Johnson, 2000). Nevertheless, many processes including PSⅡ reaction center repair cycle and thermal dissipation associated with xanthophylls cycle could lead to the decrease of Fv/Fm (Long et al. , 1994). In addition, the location of photoinhibition varies with the specific inhibitory mechanisms, donor side or acceptor side of PSⅡ reaction center (Long et al., 1994; Andersson and Aro, 2001). These could not be distinguished just by evaluating the changes of Fv/Fm. Then, a thorough investigation on the primary photochemical reactions as well as the whole electron trans- port chain by the fluorescence techniques is required. This is the story of “JIP test” . Fast fluorescence rise of dark-adapted leaves recorded with high time-resolution fluorimeters such as PEA shows a polyphasic O-J-I-P transient. The polyphasic O-J-I-P rise reflects the kinetic process of PSⅡ closure and has also been found to be very sensitive to stresses caused by changes in different environmentalconditions, such as light intensity, temperature, drought, atmospheric CO 2 or ozone elevation and chemical influences (Strasser and Tsimilli- Michael, 2001). But it is hard to evaluate the PSⅡ photo- chemical behaviors just from its shape change. The JIP- test has been introduced and further developed for the quantitative analysis of O-J-I-P transient (Strasser and Strasser, 1995). It translates the shape changes of O-J-I-P transient to quantitative changes of various parameters and
-
Upload
duongthuan -
Category
Documents
-
view
213 -
download
0
Transcript of Diurnal Photosynthesis and Photoinhibition of Rice … is uncertain whether the speedup of ATP con-...

Received 8 Sept. 2003 Accepted 7 Jan. 2004Supported by the National Natural Science Foundation of China (30200021).
* Author for correspondence. E-mail: <[email protected]>.
http://www.chineseplantscience.com
Acta Botanica Sinica植 物 学 报 2004, 46 (5): 552-559
Diurnal Photosynthesis and Photoinhibition of Rice Leaves with Chlorophyll Fluorescence
LIU Ji-Yong1, QIU Bao-Sheng1, 2* , LIU Zhi-Li2, YANG Wan-Nian1
(1. College of Life Sciences, Central China Normal University, Wuhan 430079, China;2. College of Life Sciences, Nanjing University, Nanjing 210093, China)
Abstract : The diurnal photosynthesis and photoinhibition in the photoper iod-sensit ive genicmale-sterile rice (Oryza sativa L.), Nongken 58S (NK58S), were investigated. From 06:00 to 09:00, noremarkable photoinhibition occurred, and the down-regulation of photosynthesis might be due to therunning of xanthophylls cycle. From 10:00 to 12:00, the specific energy flux for dissipation (DIo/RC) andthe net rate of reaction centers (RCs) closure (dV/dto) were increased, while the probability of electrontransport at the acceptor side (ψo) and the density of active RCs (Do) were decreased. These indicatedthat the photoinhibition of NK58S was exacerbated with the inactivation of PSⅡ RCs. Fluorescence darkrelaxation analysis and inhibitor treatment suggested that all of state transition, xanthophyll cycle andinactivation of PSⅡ RCs could contribute to protect NK58S against photodamage. Compared with theinactivation of PSⅡRCs, xanthophyll cycle had an immediate response to high light stress, wh ichfunctioned mainly in the period of relatively low light intensity. However, the inactivation of PSⅡ RCsplayed an important role in protecting the remaining active RCs when xanthophyll cycle was saturated.Key words: photoperiod-sensitive genic male-sterile rice; Nongken 58S; JIP test; photoinhibition;
reaction center inactivation; D1 protein turnover; xanthophylls cycle
Higher plants are usually encountered with light stressin their habitats. Strong irradiance will inevitably inducephotoinhibit ion with the decrease of photochemical effi-ciency when the absorbed energy is much higher than thatutilized by the photosynthetic organs (Long et al., 1994).The photoinhibition will occur much frequently when strongirradiance is combined with high temperature, drought oro ther s t ress es in s ummer midday . Furthermore, thephotoinhibition could result in photooxidative damage, pig-ment bleaching and even irreversible damage to the photo-syn thetic apparatus. Numerous p rotective mechanismshave been developed to minimize the sunlight absorptionin long-term evolu tionary history, such as alterat ion inleaves ang le, chloroplast movement and reflective cuticlecovering. Alternatively, state transition, xanthophylls cycleand PSⅡ repair cycle contribute internally to avoiding thedamage to the photosynthetic apparatus (Long et al., 1994;Horton et al., 1996). However, various species might differin their protective mechanisms for photoinhibition.
Photoinhibition is characterized by the decline of pho-tos ynthet ic quan tum efficiency and pho tochemicalefficiency, and Fv/Fm value is widely used as an index toevaluate the extent o f photoinhibit ion (Krause and Weis,1991; Govindjee, 1995; Maxwell and Johnson, 2000).Nevertheles s, many processes includ ing PSⅡ reaction
center repair cycle and thermal dissipation associated withxan thophylls cycle could lead to the decrease o f Fv/Fm(Long et a l ., 1994). In add it ion , the locat ion o fpho to inh ib it ion varies with the s pecific inh ib itorymechanisms, donor side or acceptor side of PSⅡ reactioncenter (Long et al., 1994; Andersson and Aro, 2001). Thesecould not be distinguished just by evaluating the changesof Fv/Fm. Then, a thorough investigation on the primaryphotochemical reactions as well as the whole electron trans-port chain by the fluorescence techniques is required. Thisis the story of “JIP test”.
Fast fluorescence rise of dark-adapted leaves recordedwith high time-resolution fluorimeters such as PEA showsa polyphasic O-J-I-P transient. The polyphasic O-J-I-P risereflects the kinetic process of PSⅡ closure and has alsobeen found to be very sensit ive to s tres ses caused bychanges in different environmental conditions, such as lightintensity, temperature, drought, atmospheric CO2 or ozoneelevation and chemical in fluences (Strasser and Tsimilli-Michael, 2001). But it is hard to evaluate the PSⅡ pho to-chemical behav iors just from its shape change. The JIP-tes t has been in troduced and further developed for thequantitative analysis of O-J-I-P transient (St rass er andStrasser, 1995). It translates the shape changes of O-J-I-Ptransient to quantitative changes of various parameters and





Acta Botanica Sinica 植物学报 Vol.46 No.5 2004558
down-regulation o f photosynthetic activity in NK58S.Nevertheless, each mechanism plays different roles to dis-sipate excess excitation energy and protects NK58S againstphotodamage. It has been suggested that state transitionis much important only in low light (Krause and Weis, 1991).Thus, we have not paid much attention to it in this paper.The xanthophyll cycle was initiated immediately in the firsthour when NK58S was exposed to strong irradiance in-doors (Fig.6). Thereafter, certain amounts of PSⅡ RC wereinactivated. The inactivation of PSⅡ RC is inevitable whenthe fully operation of xanthophyll cycle could not dissipateexcess excitation energy effectively. This is consistent withthe suggestion that xanthophyll cycle is the firs t line ofdefense against PSⅡ over-excitat ion (Long et a l., 1994).The JIP test has also shown that the photoinhibition ofNK58S is not marked from 06:00 to 09:00 and the down-regulation of photosynthetic activity is mainly related withthe xanthophyll cycle (Fig .4). The inactivation of PS Ⅱ
RC occurs when ligh t intensity is increased to 1 200µmol·m-2·s-1 at 10:00, and both Do and ψo values started todecrease (Fig.4). This is also consistent with Quick andStitt (1989). They had suggested that the fast componentof fluorescence quenching was not well suited to dissipatelight energy at super-saturating intensities.
Our resu lts sugges t that the damage-repair cycle ofPSⅡ RCs could contribute to protecting NK58S againstphotodamage. The inactivation of PSⅡ RC has ever beenregarded simply as damage to plants. Its protective effectsagainst photodamage have been known gradually. Theaccumulated inactivated RCs could act as potent quenchers,convert absorbed excitation energy to heat, and protectthe remaining centers against further damage (Long et al.,1994). However, the exact mechanism for inactivated RCsto dissipate excess excitation energy is still uncertain. Therepair of photodamaged PSⅡ requires a number of p ro-cesses including the dephosphorylation of D1 protein, itsdegradation, de novo synthesis, and reassembly into func-tional PSⅡ complexes (Zhang et al., 2000; Andersson andAro, 2001). The damaged D1 protein is t riggered to bedegraded via a multistep proteolytic reaction requiring GTPand ATP and catalyzed by proteases Deg P2 and FtsH(Goldberg, 1992; Spetea et a l., 1999; Andersson and Aro,2001). It is uncertain whether the s peedup of ATP con-sumption in the damage-repair cycle of PSⅡ RC could con-tribute NK58S to dissipating excess excitation energy.
It has been s uggested that the net degradation of D1protein may res ult from the strong irradiance used indoorwhich the field grown p lants seldom encounter in theirhabitats. The indoor treatment of strong irradiance is 1 200
µmol·m-2·s -1 in this study and is far below the maximallight intensity in the field (2 000 µmol·m-2·s-1). This sug-gests that the inactivation of PSⅡ RCs in NK58S is not dueto the difference of light intensities indoor and outdoor.
In this study, we have investigated the photoinhibitionof field grown NK58S with JIP test, from which the conclu-sion is well in accordance with that of laboratory experi-ment with the utilization of inhibitor. The JIP-test has beenproved to be a very useful tool for the in vivo investigationof the adaptive behavior of the photosynthetic apparatusand, especially, of PSⅡ to a wide variety and combinationof stressors owing to its convenient, rapid and non-inva-sive features (Lu and Vonshak, 1999; Strasser and Tsimilli-Michael, 2001). However, the recorded fluorescence tran-sients carry much more information than those used by theJIP test. The more powerful methods of numerical simula-tions utilizing the rich in formation are expected. This isdependent on our profound understanding to the fluores-cence signal and the appearance of more powerful fluorim-eters in the future.
References:
Andersson B, Aro E M. 2001. Photodamage and D1 prot ein
turnover in photosystem Ⅱ. Aro E M, Andersson B. Regu-
lation of Photosynthesis. The Netherlands: Kluwer AcademicPublishers. 377–393.
Aro E M, McCaffery S, Anderson J M. 1993. Photoinhibition
and D1 protein degradat ion in peas acclimated to differentgrowth irradiances. Plant Physiol, 103: 835–843.
Goldberg A L. 1992. T he mechanism and functions of ATP-
dependent proteases in bact erial and animal cells. Eur JBiochem, 203: 9–23.
Govindjee. 1995. Sixty-three years since Kautsky: chlorophyll
a fluorescence. Aust J Plant Physiol, 22: 131–160.Hong S S, Xu D Q. 1997. Difference in response of chlorophyll
fluorescence parameters to st rong light between wheat and
soybean leaves. Chin Sci Bull, 42: 684–688.Hort on P, Ruban A V, Walters R G. 1996. Regulat ion of light
harvesting in green plants. Annu Rev Plant Physiol Plant Mol
Biol, 47: 655–684.Keren N, Berg A, van Kan P J M, Levanon H, Ohad I. 1997.
Mechanism of photosystem Ⅱ inactivation and D1 protein
degradation at low light: the role of back electron flow. ProcNatl Acad Sci USA, 94: 1579–1584.
Krause G H, Weis E. 1991. Chlorop hyll fluorescence and
photosynt hesis: the basics. Annu Rev Plant Phys iol PlantMol Biol, 42: 313–349.
Long S P, Humphries S, Falkowski P G. 1994. Photoinhibition
of photosynthesis in Nature. Annu Rev Plant Physiol Plant

LIU Ji-Yong et al.: Diurnal Photosynthesis and Photoinhibition of Rice Leaves with Chlorophyll Fluorescence 559
Mol Biol, 45: 633–662.Lu C M, Qiu N W, Lu Q T, Wang B S, Kuang T Y. 2003. PSⅡ
photochemistry, thermal energy dissipation, and the xantho-
phyll cycle in Kalanchoë daigremontiana exposed to a com-bination of water stress and high light . Phys iol Plant, 113:
173–182.
Lu C M, Vonshak A. 1999. Characterization of PSⅡ photo-chemistry in salt-adapted cyanobacterium Spirulina platensis
cells. New Phytol, 141/2: 231–239.
Mattoo A K, Giardi M T, Raskind A, Edelman M. 1999. Dy-namic metabolism of photosyst em Ⅱ reaction center p ro-
teins and pigments. Physiol Plant, 107: 454–461.
Maxwell K, Johnson G N. 2000. Chlorophyll fluorescence - apractical guide. J Exp Bot, 51: 659–668.
Meng Q-W, Weis E, Zou Q, Zhao S-J. 1999. Photoinhibition and
phot op rotect ion in Ginkgo biloba leaves : influence oftemperature, CO2 and O2. Acta Bot Sin, 41: 398–404.
Quick W P, Stitt M. 1989. An examination of factors contribut-
ing to non-photochemical quenching of chlorophyll fluores-cence in barley leaves. Biochim Biophys Acta, 977: 287–296.
Spetea C, Hundal T, Lohmann F, Andersson B. 1999. GTP
bound to chloroplast thylakoid membranes is required forlight-induced multi-enzyme degradation of the photosystem
Ⅱ D1 protein. Proc Natl Acad Sci USA, 96: 6547–6552.
Strasser B J , Strasser R J. 1995. Measuring fast fluorescencetransients to address environmental questions: the JIP test.
Mathis P. Photosynthesis: From Light to Biosphere, Vol. 5.
The Netherlands: Kluwer Academic Publishers. 977–980.Strasser R J, Srivastava A, Govindjee. 1995. Polyphasic chloro-
phyll a fluorescence transient in plants and cyanobacteria.
Photochem Photobiol, 61: 32–42.Strasser R J, Tsimilli-Michael M . 2001. Stress in p lants, from
daily rhythm to global changes, detected and quantified by the
JIP-test. Chimie Nouvelle (SRC), 75: 3321–3326.Strasser R J, Tsimilli-Michael M, Srivastava A. 2000. The fluo-
rescence transient as a tool to characterise and screen photo-
synthetic samples. Yunus M, Pathre U, Mohanty P. ProbingPhotosynt hesis: Mechanisms, Regulation and Adaptation.
London: Taylor & Francis. 445–483.
Tong Z. 1998. Studies on photoperiod-sensitive genic malesterilerice in terms of developmental biology. Acta Bot Sin, 40:189–
199. (in Chinese with English abstract)
Zhang L X, Paakkarinen V, van Wijk K J, Aro E M . 2000.Biogenesis of the chloroplast-encoded D1 protein: regulation
of translation elongation, insertion, and assembly into photo-
system Ⅱ. Plant Cell, 12: 1769–1781.
(Managing editor: HE Ping)


















