
Distinct roles of neuropilin 1 signaling for radial and tangential extension of callosal axons
11
Distinct Roles of Neuropilin 1 Signaling for Radial and Tangential Extension of Callosal Axons YUMIKO HATANAKA, 1,2 * TOMOKO MATSUMOTO, 2 YUCHIO YANAGAWA, 3 HAJIME FUJISAWA, 4 FUJIO MURAKAMI, 2 AND MASAYUKI MASU 1 * 1 Department of Molecular Neurobiology, Graduate School of Comprehensive Human Sciences, University of Tsukuba, Tsukuba, Ibaraki 305-8577, Japan 2 Laboratory of Neuroscience, Graduate School of Frontier Biosciences, Osaka University, Suita, Osaka 565-0178, Japan 3 Department of Genetic and Behavioral Neuroscience, Graduate School of Medicine, Gunma University, Maebashi, Gunma 371- 8511, Japan 4 Division of Biological Science, Graduate School of Science, Nagoya University, Chikusa-ku, Nagoya, Aichi 464-8602, Japan ABSTRACT Cortical excitatory neurons migrate from their origin in the ventricular zone (VZ) toward the pial surface. During migra- tion, these neurons exhibit a stellate shape in the interme- diate zone (IZ), transform into bipolar cells, and then initiate radial migration, extending a trailing process, which may lead to an axon. Here we examined the role of neuropilin 1 (NRP1) in these developmental events. Both NRP1 mRNA and protein were highly expressed in the IZ, where stellate- shaped cells were located. DiI labeling experiments showed that neuronal migration occurred normally in Nrp1 mutant mice up to embryonic day (E) 14.5, the latest day to which the mutant survives, with only subtle axonal defasciculation. However, interference with Nrp1 signaling at a later stage caused pathfinding errors: when a dominant negative form of Nrp1 was electroporated into the cortical VZ cells at E12.5 or E15.5 and examined perinatally, guidance errors were found in tangential axonal extension toward the midline. In contrast, no significant effect was noted on the migration of cortical excitatory neurons. These findings indicate that NRP1 plays an important role in the guidance of callosal axons originating from cortical excitatory neurons but does not support a role in their migration. Moreover, insofar as radial axonal extension within the cortical plate was unaf- fected, the present findings imply that molecular mecha- nisms for the axonal extension of excitatory neurons within the cortical plate are distinct from those in the white matter. J. Comp. Neurol. 514:215–225, 2009. © 2009 Wiley-Liss, Inc. Indexing terms: cerebral cortex; electroporation; intermediate zone; migration; mouse During development, many neurons are generated in dis- crete regions lining the ventricular space and migrate long distances to reach their final destinations. They also extend axons and dendrites to make synaptic connections that lead to functional neural circuits. These events essentially occur sequentially, but in certain cases the neuronal migration and the axonal extension occur simultaneously. For example, cer- ebellar granule cells extend their “T-shaped” parallel fibers during their migration from the external to internal granular layer (for review see Altman and Bayer, 1997; Hatten and Heintz, 1995). Facial branchiomotor neurons, which are born in rhombomere (r) 4, migrate caudally to travel through r5 and r6, concomitant with the extension of axons into the second branchial arch (see Chandrasekhar, 2004). These findings suggest that there may be a link between migration of neurons and their axonal extensions. The cerebral cortex is one of the structures in which both neuronal migration and axonal projection are well docu- mented. The cerebral cortex consists of a six-layered struc- ture, exhibiting a characteristic arrangement of excitatory neurons, which are a principal neuronal type in the cerebral cortex. They extend axons toward the white matter, where these axons turn perpendicularly to make ascending or de- scending projections. These neurons originate from the cor- Additional Supporting Information may be found in the online version of this article. Grant sponsor: Ministry of Education, Culture, Sports, Science and Tech- nology, Grant number: 17500200 (to Y.H.); Grant number: 17024007 (to Y.H.); Grant number: 18300102 (to Y.Y.); Grant number: 18019007 (to Y.Y.); Grant sponsor: University of Tsukuba Research Projects (to Y.H.). Yumiko Hatanaka’s current address is Graduate School of Biological Sciences, Nara Institute of Science and Technology, Takayama-cho, Ikoma, Nara 630-0192, Japan. *Correspondence to: Yumiko Hatanaka, Graduate School of Biological Sciences, Nara Institute of Science and Technology, Takayama-cho, Ikoma 630-0192, Japan. E-mail: [email protected] or to Masayuki Masu, Department of Molecular Neurobiology, Graduate School of Comprehen- sive Human Sciences, University of Tsukuba, Tsukuba, Ibaraki 305-8577, Japan. E-mail: [email protected] Received 11 December 2007; Revised 2 May 2008; Accepted 27 January 2009 DOI 10.1002/cne.22021 Published online February 23, 2009 in Wiley InterScience (www.interscience. wiley.com). The Journal of Comparative Neurology 514:215–225 (2009) Research in Systems Neuroscience © 2009 Wiley-Liss, Inc.
-
Upload
yumiko-hatanaka -
Category
Documents
-
view
212 -
download
0
Transcript of Distinct roles of neuropilin 1 signaling for radial and tangential extension of callosal axons

Distinct Roles of Neuropilin 1 Signaling for Radial andTangential Extension of Callosal Axons
YUMIKO HATANAKA,1,2* TOMOKO MATSUMOTO,2 YUCHIO YANAGAWA,3 HAJIME FUJISAWA,4
FUJIO MURAKAMI,2 AND MASAYUKI MASU1*1Department of Molecular Neurobiology, Graduate School of Comprehensive Human Sciences, University of Tsukuba, Tsukuba,Ibaraki 305-8577, Japan2Laboratory of Neuroscience, Graduate School of Frontier Biosciences, Osaka University, Suita, Osaka 565-0178, Japan3Department of Genetic and Behavioral Neuroscience, Graduate School of Medicine, Gunma University, Maebashi, Gunma 371-8511, Japan4Division of Biological Science, Graduate School of Science, Nagoya University, Chikusa-ku, Nagoya, Aichi 464-8602, Japan
ABSTRACTCortical excitatory neurons migrate from their origin in theventricular zone (VZ) toward the pial surface. During migra-tion, these neurons exhibit a stellate shape in the interme-diate zone (IZ), transform into bipolar cells, and then initiateradial migration, extending a trailing process, which maylead to an axon. Here we examined the role of neuropilin 1(NRP1) in these developmental events. Both NRP1 mRNAand protein were highly expressed in the IZ, where stellate-shaped cells were located. DiI labeling experiments showedthat neuronal migration occurred normally in Nrp1 mutantmice up to embryonic day (E) 14.5, the latest day to whichthe mutant survives, with only subtle axonal defasciculation.However, interference with Nrp1 signaling at a later stagecaused pathfinding errors: when a dominant negative form
of Nrp1 was electroporated into the cortical VZ cells at E12.5or E15.5 and examined perinatally, guidance errors werefound in tangential axonal extension toward the midline. Incontrast, no significant effect was noted on the migration ofcortical excitatory neurons. These findings indicate thatNRP1 plays an important role in the guidance of callosalaxons originating from cortical excitatory neurons but doesnot support a role in their migration. Moreover, insofar asradial axonal extension within the cortical plate was unaf-fected, the present findings imply that molecular mecha-nisms for the axonal extension of excitatory neurons withinthe cortical plate are distinct from those in the white matter.J. Comp. Neurol. 514:215–225, 2009.© 2009 Wiley-Liss, Inc.
Indexing terms: cerebral cortex; electroporation; intermediate zone; migration; mouse
During development, many neurons are generated in dis-crete regions lining the ventricular space and migrate longdistances to reach their final destinations. They also extendaxons and dendrites to make synaptic connections that leadto functional neural circuits. These events essentially occursequentially, but in certain cases the neuronal migration andthe axonal extension occur simultaneously. For example, cer-ebellar granule cells extend their “T-shaped” parallel fibersduring their migration from the external to internal granularlayer (for review see Altman and Bayer, 1997; Hatten andHeintz, 1995). Facial branchiomotor neurons, which are bornin rhombomere (r) 4, migrate caudally to travel through r5 andr6, concomitant with the extension of axons into the secondbranchial arch (see Chandrasekhar, 2004). These findingssuggest that there may be a link between migration of neuronsand their axonal extensions.
The cerebral cortex is one of the structures in which bothneuronal migration and axonal projection are well docu-mented. The cerebral cortex consists of a six-layered struc-ture, exhibiting a characteristic arrangement of excitatoryneurons, which are a principal neuronal type in the cerebral
cortex. They extend axons toward the white matter, wherethese axons turn perpendicularly to make ascending or de-scending projections. These neurons originate from the cor-
Additional Supporting Information may be found in the online version ofthis article.
Grant sponsor: Ministry of Education, Culture, Sports, Science and Tech-nology, Grant number: 17500200 (to Y.H.); Grant number: 17024007 (toY.H.); Grant number: 18300102 (to Y.Y.); Grant number: 18019007 (to Y.Y.);Grant sponsor: University of Tsukuba Research Projects (to Y.H.).
Yumiko Hatanaka’s current address is Graduate School of BiologicalSciences, Nara Institute of Science and Technology, Takayama-cho,Ikoma, Nara 630-0192, Japan.
*Correspondence to: Yumiko Hatanaka, Graduate School of BiologicalSciences, Nara Institute of Science and Technology, Takayama-cho, Ikoma630-0192, Japan. E-mail: [email protected] or to Masayuki Masu,Department of Molecular Neurobiology, Graduate School of Comprehen-sive Human Sciences, University of Tsukuba, Tsukuba, Ibaraki 305-8577,Japan. E-mail: [email protected]
Received 11 December 2007; Revised 2 May 2008; Accepted 27 January2009
DOI 10.1002/cne.22021Published online February 23, 2009 in Wiley InterScience (www.interscience.
wiley.com).
The Journal of Comparative Neurology 514:215–225 (2009)
Research in Systems Neuroscience
© 2009 Wiley-Liss, Inc.

tical ventricular zone (VZ) and migrate radially toward the pialsurface (see Kriegstein and Noctor, 2004; Nadarajah and Par-navelas, 2002). During their migration, they show morpholog-ical changes in accordance with their location; a radial glia-like shape in the VZ, stellate (or multipolar; Tabata andNakajima, 2003) shape in the intermediate zone (IZ)/subventricular zone (SVZ), and bipolar shape in the upper partof the cortical wall (Hatanaka et al., 2004; Hatanaka and Mu-rakami, 2002; Noctor et al., 2004; Shoukimas and Hinds, 1978;Tabata and Nakajima, 2003). Stellate-shaped cells in the IZ/SVZ are characterized by many thin, short processes withrandomly oriented cell bodies. When they migrate across theIZ/SVZ, a thin axon-like tangential process seems to extendthrough the IZ (Hatanaka et al., 2004). Thus, cellular migrationof cortical excitatory neurons appears to take place concur-rently with axonal extension in the IZ/SVZ.
These findings tempted us to explore the molecular mecha-nisms underlying the concurrent occurrence of cellular migrationand axonal extension in the IZ/SVZ. There are several moleculesknown to be expressed in the IZ/SVZ (Englund et al., 2005; Hiraiet al., 2002; Kudo et al., 2005; Nieto et al., 2004; Tarabykin et al.,2001), including neuropilin 1 (NRP1; Takagi et al., 1995). NRP1 isa component of a receptor complex that mediates the signalingof chemorepulsive activities from class 3 semaphorins and con-tributes to directional axonal guidance (He and Tessier-Lavigne,1997; Kolodkin et al., 1997). In addition to axonal guidance,several studies indicate its role in neuronal migration. For exam-ple, prospective cortical interneurons originating from the basalforebrain express NRP1 and -2 during migration, and those lack-ing either neuropilin accumulated in the striatum, failing to enterthe cerebral cortex (Marı́n et al., 2001). These neurons also useneuropilin signaling to find proper paths in the cerebral cortex(Tamamaki et al., 2003a). The appropriate positioning of sympa-thetic neurons is also controlled by NRP1 signaling, which sup-presses migration and thus promotes the condensation of theseneurons at defined target sites (Kawasaki et al., 2002). NRP1 isexpressed in the IZ during the embryonic development of themouse cerebral cortex at the stage when cortical neurons mi-grate radially and extend axons (Bagnard et al., 1998; Takagi etal., 1995). This raises the idea that NRP1 may be involved in boththe axonal guidance and the cellular migration of cortical exci-tatory neurons when they traverse the IZ.
In this study, we examined NRP1 expression pattern and itspotential role in axonal pathfinding and cellular migration forcortical excitatory neurons. We found that both NRP1 mRNAand NRP1 protein were highly expressed in stellate-shapedcells in the IZ. Analyses of Nrp1 mutant mice and overexpres-sion of a dominant negative form (DN) of NRP1 in migratingcells indicated an essential role for NRP1 during the guidanceof callosal axons extending from excitatory neurons.
MATERIALS AND METHODSExperimental animals
Timed-pregnant ICR mice were purchased from Nihon-SLC.The day on which the vaginal plug was observed in the morn-ing was designated E0. Nrp1 mutants produced in the ICRbackground (Kitsukawa et al., 1997) were provided by RIKENBRC (generation >N15). Their genotype was determined byPCR as described previously (Kitsukawa et al., 1997). NRP1immunoreactivity is absent in these mutants (Kitsukawa et al.,
1997; present study). Moreover, the lack of NRP1 signalingwas confirmed by the lack of growth cone collapse in dorsalroot ganglion neurons when elicited by Sema3A (Kitsukawa etal., 1997). Heterozygous glutamate decarboxylase 67(GAD67)-green fluorescent protein (GFP) knock-in mouse em-bryos with ICR background were used (Tamamaki et al.,2003b; Tanaka et al., 2006). More than 96% of �-aminobutyricacid (GABA)-immunoreactive neurons at embryonic stages(E13.5 and E15.5) express GFP (Tanaka et al., 2006), whereasimmuonoreactivities for GABA have been detected in 95%and 75% of GFP-expressing cells at E13.5 and E15.5, respec-tively (Tanaka, Maekawa, Yanagawa, Obata, Murakami, un-published observations). All experiments were conducted incompliance with the guidelines for use of laboratory animalsof the University of Tsukuba and Osaka University.
In situ hybridizationE12.5 embryos or E15.5 brains were fixed in 4% parafor-
maldehyde (PFA)/0.1 M phosphate buffer, pH 7.4 (PB), over-night, immersed in 30% sucrose in PBS, embedded in OCTcompound (Sakura Finetechnical, Co. Ltd., Tokyo, Japan),and cut coronally at a thickness of 16 �m. Sections werecollected on silane-coated glass slides and subjected to insitu hybridization. Hybridization was performed as describedpreviously (Hatanaka and Jones, 1998), with some modifica-tions. In brief, a digoxigenin-labeled probe for Nrp1(Kawakami et al., 1996) was used without alkaline hydrolysisand hybridized overnight at 60°C.
PlasmidseGFP (Clontech, Logan, UT), Venus (Nagai et al., 2002), and
a dominant negative form of Nrp1 (Nrp1DN) were cloned intopCAGGS vector (Niwa et al., 1991). Nrp1DN was prepared bydeleting extracellular regions containing the a1, a2, b1, and b2domains (Serini et al., 2003), thus lacking both a ligand-binding domain (a1 and a2 or b1 and b2, Giger et al., 1998; b1and b2, Renzi et al., 1999; a1 and a2, Shimizu et al., 2000) anda cell adhesion domain (b1 and b2, Shimizu et al., 2000). Theresulting construct contained a signal sequence (fromSema3A) and myc-tag epitope, followed by amino acids 590–923 (NCBI; BC060129), and interfered with the biological re-sponse to Sema3 proteins, presumably by interrupting theoligomerization of functional receptors (Serini et al., 2003).
Antibody characterizationAnti-NRP1 was a rabbit polyclonal antibody raised against
the latter half of the b2 domain and c/MAM domain of recom-binant mouse NRP1. It was affinity purified by using the re-combinant as a ligand (Kawakami et al., 1996). No stainingwas seen on sections of Nrp1 mutant mice (Kitsukawa et al.,1997; present study). Anti TAG-1 (4D7/TAG1 from Develop-mental Studies Hybridoma Bank) is a mouse monoclonal IgMantibody obtained with rat fetal brain as an immunogen(Yamamoto et al., 1986). It binds to a 135-kDa glycoprotein(Dodd et al., 1988) and recognizes specific neural cell types inthe fetal brain (Yamamoto et al., 1986). Anti-L1 is a rat mono-clonal IgG antibody obtained by immunizing a glycoproteinfraction from the cerebellum of 8–10-day-old C57BL/6J mice(Boehringer Mannheim, Indianapolis, IN: 1450999; or Chemi-con, Temecula, CA: MAB5272). It binds to a 140-kDa and a200-kDa protein (Rathjen and Schachner, 1984) and recog-nizes a subset of axonal tracts in the cerebral cortex (Jones et
Research in Systems Neuroscience The Journal of Comparative Neurology
216 Y. HATANAKA ET AL.

al., 2002). No binding has been seen on sections of L1 knock-out mice (Fransen et al., 1998). Anti-myc (Myc-Tag; Cell Sig-naling, Beverly, MA: 2272) is a rabbit polyclonal antibodyraised against a synthetic peptide corresponding to aminoacids 410–419 from human c-Myc. It was purified by proteinA and peptide affinity chromatography. The antibody stainedonly ectopically expressed myc-tagged proteins in COS7 cellsand neural cells.
ImmunohistochemistryCryostat sections were made from E12.5 and 13.5 em-
bryos or E15.5 and 18.5 brains fixed in 4% PFA/PB for 1hours, as described above. Anti-NRP1 (1 �g/ml) and Cy3-conjugated anti-rabbit IgG (1/300; Jackson Immunore-search, West Grove, PA) were used. For double immuno-staining, anti-TAG-1 (supernatant) and biotinylated anti-mouse IgM (1/500; Vector, Burlingame, CA), or anti-L1 (5–10�g/ml) and biotinylated anti-rat IgG (1/500; Jackson Immu-noresearch) were used. They were subsequently treatedwith Cy2-conjugated streptavidin (1/300; Jackson Immu-noresearch). To examine Nrp1DN expression, anti-NRP1 oranti-myc antibody (1/1,000) was used.
Slice cultureE12.5 embryos either heterozygous or homozygous for
Nrp1 mutants were removed from dams. About 0.5 �l ofpCAGGS:Venus solution (5 �g/�l in PBS) was injected into thelateral ventricles. After electric pulses (20 V, pulse duration 50msec, interval 75 msec, twice) were delivered with forceps-shaped electrodes (diameter 5 mm; CUY650P5; Unique Me-dial Imada, Miyagi, Japan), the forebrain was dissected out,embedded in 4% low-melting-temperature agarose (AgaroseLM; Nacalai, Kyoto, Japan) in PBS, and cut into 300-�m-thickcoronal slices with a vibrating blade microtome (VT1000S,Leica Microsystems, Tokyo, Japan). Slices were placed on acollagen-coated membrane (Transwell-COL; No. 3492; Corn-ing Coster, Cambridge, MA) and cultured in Dulbecco’s mod-ified Eagle’s medium and F12 medium containing 10% fetalbovine serum, N-2 supplement (Gibco, Grand Island, NY),penicillin (100 IU/ml), and streptomycin (100 �g/ml) under95% O2 and 5% CO2 at 37°C (Jossin et al., 2003; Miyata et al.,2002). After 48 hours in culture, slices were fixed in 4%PFA/PB for 2 hours and cut into 20-�m sections as describedabove.
DiI labelingNrp1 mutants at E13.5 and E14.5 were collected and fixed in
4% PFA/PB for several days. Brains were dissected out, embed-ded in 4% low-melting-temperature agarose, and cut coronallyat a thickness of 200 �m. Slices from all rostrocaudal levels ofthe cerebral cortex were collected. A small crystal of 1,1�-dioctadecyl-3,3,3�,3�-tetramethylindocarbocyanine perchlorate(DiI; Molecular Probes, Eugene, OR) was placed into the dorsaland lateral part of the IZ of these slices. They were stored in thedark for a few weeks in the same fixative at 4°C and examinedunder a fluorescence microscope or a confocal microscope(LSM 510; Zeiss, Jena, Germany).
In utero electroporationPlasmids (eGFP or a mixture of eGFP and Nrp1DN) were
dissolved in PBS at a concentration of 5 �g/�l in each. In uteroelectroporation was performed as described previously (Ha-
tanaka et al., 2004). Successful electroporation was judged byexamining the embryos or pups under a fluorescence dissectionmicroscope (Leica MZ FL III). Those showing adequate fluores-cent signal were collected: heads of E14.5 or brains of E18.5mice were immersion fixed in 4% PFA/PB, and postnatal brainswere fixed by intracardial perfusion with the fixative and re-moved. After being treated overnight in the same fixative, theywere processed for cryosections as described above. Sections(50 �m) were collected in PBS and mounted onto MAS-coatedglass slides (Matsunami, Osaka, Japan) or further subjected toimmunohistochemistry. Some sections were treated with pro-pidium iodide (1 �g/ml) or 4�,6-diamidino-2-phenylindole dihy-drochloride (1 �g/ml). For single-cell tracing, fixed brains werewashed with PBS, then cut coronally into 200-�m-thick sliceswith a vibrating blade microtome.
Dissociation cultureTwenty-four hours after in utero electroporation with eGFP
at E12.5, labeled cortices were cut out, dissociated, plated at8 � 104 cells per well on a poly-L-lysine- and laminin-coatedLab-tek Permanox slide (eight wells; Nunc, Naperville, IL), andcultured as previously described (Hatanaka and Murakami,2002). After 24 hours in culture, cells were fixed with 4%PFA/PB (pH 7.4) for 10 minutes, then processed for immuno-cytochemistry.
Image processingImages were acquired with a CCD camera (AxioCam HRc,
Zeiss; or CoolSnap HQ, Roper) on an Olympus BX-60 mi-croscope or a Keyence VB-7000 camera on a Zeiss Axio-plan2 microscope. Z-series confocal images of sectionswere captured and assembled into a single image in ZeissLSM510 software (version 3.2). All digital files were im-ported into Adobe Photoshop CS2. After minor contrast andbrightness adjustment, montages of images were con-structed. For quantification of either the GFP� or theNRP1� area in the IZ of GAD67-GFP mouse, the IZ region inan image was first cut out in Photoshop CS2. GFP� area ingreen and NRP1� area in red channels were split off, then mea-sured in ImageJ (1.36b, http://rsb.info.nih.gov/ij/Java 1.4.2_12).For quantification of the effect of NRP1DN on axonal guidance,images were captured from a coronal section taken from theintermediate level along the anteroposterior axis from eachbrain. GFP� axons were viewed through a �63 oil immersionobjective, and a stack of images was created from a series of40–50 images along the z-axis at 0.5-�m intervals. The numberof individually recognizable GFP� axons projecting medially wascounted from each section, and the percentage of GFP� axonsentering the ipisilateral CP was calculated. Data are presentedas mean � SEM. Statistical analysis was via Student’s t-test.
RESULTSExcitatory neurons expressed NRP1 in the IZNRP1 is expressed in the IZ of the mouse cerebral cortex
(Bagnard et al., 1998; Kawakami et al., 1996), which raisesthe possibility that excitatory neurons express NRP1 duringthe course of their migration. To test this, we first examinedNrp1 mRNA expression in these neurons. In situ hybridiza-tion indicated that Nrp1 was expressed in the preplate (PP)at E12.5 and the IZ at E15.5 (Fig. 1A,B), consistently withprevious studies (Bagnard et al., 1998; Kawakami et al.,
Research in Systems NeuroscienceThe Journal of Comparative Neurology
217ROLE OF NEUROPILIN 1 IN CALLOSAL AXON GUIDANCE
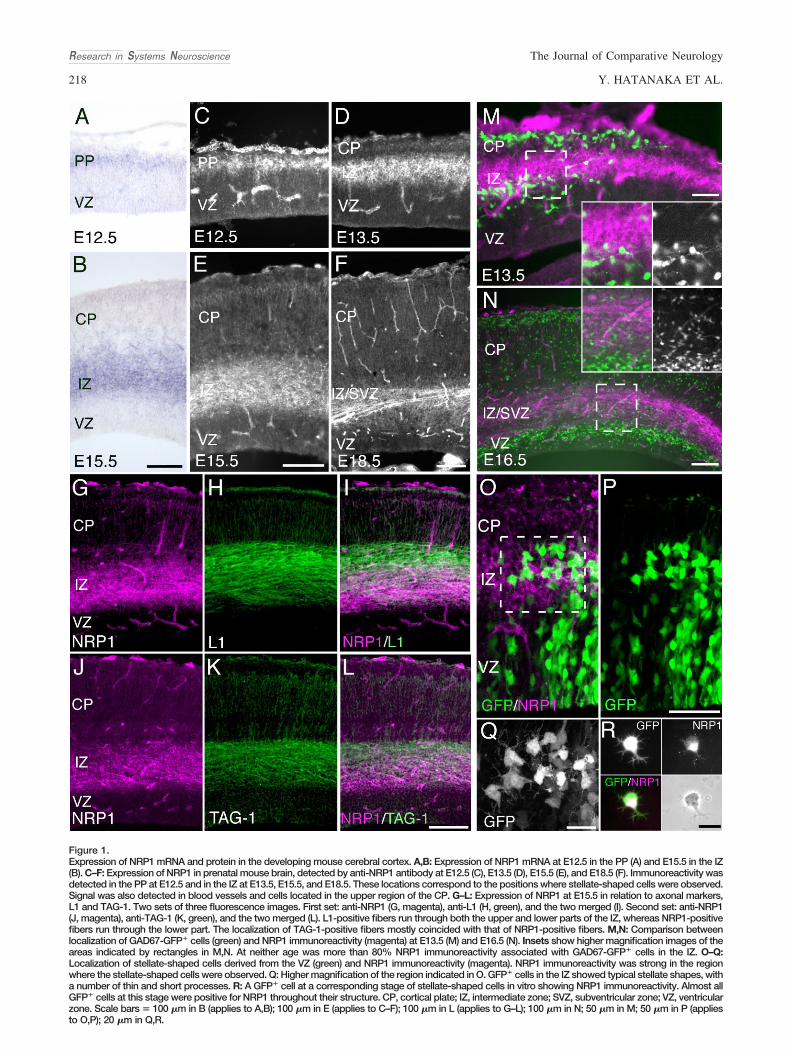
Figure 1.Expression of NRP1 mRNA and protein in the developing mouse cerebral cortex. A,B: Expression of NRP1 mRNA at E12.5 in the PP (A) and E15.5 in the IZ(B). C–F: Expression of NRP1 in prenatal mouse brain, detected by anti-NRP1 antibody at E12.5 (C), E13.5 (D), E15.5 (E), and E18.5 (F). Immunoreactivity wasdetected in the PP at E12.5 and in the IZ at E13.5, E15.5, and E18.5. These locations correspond to the positions where stellate-shaped cells were observed.Signal was also detected in blood vessels and cells located in the upper region of the CP. G–L: Expression of NRP1 at E15.5 in relation to axonal markers,L1 and TAG-1. Two sets of three fluorescence images. First set: anti-NRP1 (G, magenta), anti-L1 (H, green), and the two merged (I). Second set: anti-NRP1(J, magenta), anti-TAG-1 (K, green), and the two merged (L). L1-positive fibers run through both the upper and lower parts of the IZ, whereas NRP1-positivefibers run through the lower part. The localization of TAG-1-positive fibers mostly coincided with that of NRP1-positive fibers. M,N: Comparison betweenlocalization of GAD67-GFP� cells (green) and NRP1 immunoreactivity (magenta) at E13.5 (M) and E16.5 (N). Insets show higher magnification images of theareas indicated by rectangles in M,N. At neither age was more than 80% NRP1 immunoreactivity associated with GAD67-GFP� cells in the IZ. O–Q:Localization of stellate-shaped cells derived from the VZ (green) and NRP1 immunoreactivity (magenta). NRP1 immunoreactivity was strong in the regionwhere the stellate-shaped cells were observed. Q: Higher magnification of the region indicated in O. GFP� cells in the IZ showed typical stellate shapes, witha number of thin and short processes. R: A GFP� cell at a corresponding stage of stellate-shaped cells in vitro showing NRP1 immunoreactivity. Almost allGFP� cells at this stage were positive for NRP1 throughout their structure. CP, cortical plate; IZ, intermediate zone; SVZ, subventricular zone; VZ, ventricularzone. Scale bars � 100 �m in B (applies to A,B); 100 �m in E (applies to C–F); 100 �m in L (applies to G–L); 100 �m in N; 50 �m in M; 50 �m in P (appliesto O,P); 20 �m in Q,R.
Research in Systems Neuroscience The Journal of Comparative Neurology
218 Y. HATANAKA ET AL.

1996). Because cortical interneurons, mostly GABA-ergiccells, originating from the basal forebrain have not arrivedat the medial part of the cortex at E12.5 (Tanaka et al.,2003), this finding suggests that excitatory neurons expressNrp1.
Then, we examined the distribution of NRP1 protein by immu-nohistochemistry (Fig. 1C–F). High-level NRP1 expression wasfound in the PP at E12.5 and in the IZ from E13.5 through E15.5.Immunoreactivity in the IZ, however, has been seen to decreaseat E18.5 (Kawakami et al., 1996). Although the immunoreactivitywas punctate at earlier stages, it showed a fasciculated axon-like pattern at this stage (Fig. 1F).
To determine the precise localization of NRP1 within the IZ,we compared the NRP1 expression with either L1 or TAG-1expression. L1 is expressed both in corticofugal and inthalamocortical axons, whereas TAG-1 is expressed in corti-cofugal axons (Jones et al., 2002). As shown in Figure 1G–L,most NRP1 expression occurred where TAG-1 was ex-pressed, supporting the notion that cortical efferents expressNRP1.
Cortical GABAergic interneurons migrate tangentially intothe cortex through the IZ/SVZ and/or MZ (for reviews seeMarı́n and Rubenstein, 2001; Nadarajah and Parnavelas,2002; Parnavelas, 2000). Because some GABAergic inter-neurons also express NRP1 (Marı́n et al., 2001; Tamamaki etal., 2003a), the immunoreactivity observed from E13 on-ward might be attributable to GABAergic interneurons. Toexamine further the expression of NRP1 by cortical excita-tory neurons, we performed immunostaining for NRP1 insections for GAD67-GFP knock-in mice, in which GABAer-gic cells express GFP (Tamamaki et al., 2003b; Tanaka etal., 2006). We found that the somata and processes ofGABAergic cells accounted for only 18% and 14.5% ofNRP1-immunoreactive area in the IZ at E13.5 and at E16.5,respectively (Fig. 1M,N). These findings, together with theresults of TAG-1 staining, support the idea that the excita-tory neurons express NRP1 protein.
Excitatory neurons change their shape from a radial glia-likeshape to stellate-shape upon leaving the VZ (Hatanaka et al.,2004; Noctor et al., 2004; Tabata and Nakajima, 2003). Thepattern of NRP1 expression is consistent with the idea thatexcitatory neurons, specifically stellate-shaped cells in the IZ,express high levels of NRP1. To test this, we electroporatedeGFP into E12.5 VZ cells and examined NRP1 immunoreac-tivity in GFP� cells at E13.5. As shown in Figure 1O–Q, thedistribution of GFP� stellate-shaped cells exhibiting a numberof thin and short processes coincided with that of NRP1expression. However, the presence of numerous stellate-shaped cells and punctate immunoreactivity for NRP1 in theIZ prevented us from examining the precise distribution ofNRP1 at the single-cell level. To address this point, we applieda dissociation culture of GFP� cells. One day after electropo-ration at E12.5, GFP� cells were dissociated and cultured for24 hours in vitro. During culture, many GFP� cells extendedthin and short processes, which probably corresponded to thestage of stellate-shaped cells in vivo. Staining with anti-NRP1antibody showed that almost all GFP� cells exhibited immu-noreactivity for NRP1 in the entire cell body, including neurites(Fig. 1R). Taken together, these findings indicate that excita-tory neurons in the IZ express NRP1 at a high level duringcortical development.
Both axonal extension and cellular migration ofearly-born neurons were largely normal in Nrp1
mutant miceStellate-shaped cells in the IZ have many short processes,
one of which sometimes extends tangentially through the IZ.This process will eventually become an axon (Hatanaka et al.,2004; Shoukimas and Hinds, 1978). The stellate-shaped cellsalso extend leading processes while moving toward the pialsurface (Hatanaka et al., 2004; Noctor et al., 2004; Shoukimasand Hinds, 1978; Tabata and Nakajima, 2003). The expressionof NRP1 in stellate-shaped cells in the IZ prompted us toinvestigate the role of NRP1 in the axonal extension andmigration of cortical excitatory neurons.
To examine the role of NRP1, we first used Nrp1 mutantmice (Kitsukawa et al., 1997). We labeled VZ cells of Nrp1mutant mice at E12.5 with Venus by electroporation and cul-tured organotypically. After 2 days in culture, the CP devel-oped well, and many Venus� cells were found in the CP (Supp.Info. Fig. 1; n � 2). This result raises the possibility that cellsoriginating in the VZ at E12 or later reach the CP in theabsence of NRP1.
We noticed that Nrp1 homozygous mutant mice obtained fromheterozygotes that had been back-crossed with wild-type ICRmice over 15 generations survived up to E14.5, although someabnormalities, including impaired capillary invasion in the centralnervous system (Kawasaki et al., 1999), a thinner cortical wall,and the accumulation of edema fluid in the back, were apparent.This allowed us to examine the phenotype of cortical neurons inE13.5 and E14.5 Nrp1 mutant mice.
The CP appears in the lateral portion of the cerebral cortexby E13.5 in wild-type mice. A CP-like structure also appearedin Nrp1 mutant mice at E13.5 in the lateral portion (Fig. 2A),suggesting proper migration of VZ-derived cells into the CP.Because the extension of corticofugal axons has already oc-curred by E13.5 in wild-type mice, the morphology of excita-tory projection neurons was examined by retrograde labelingwith a small DiI crystal placed into the IZ (Fig. 2B). DiI fluores-cence indicated that cells extending their axons toward theinternal capsule showed a shape of projection neurons ex-tending thick processes toward the pial surface (Fig.2C,D,G,H). There were no apparent differences in orientationand shape of soma of the labeled cells among heterozygous(n � 5; Fig. 2G), homozygous (n � 4; Fig. 2H), and wild-type(n � 3; data not shown) mice. Anterograde labeling of axonalextensions from neurons in the dorsal cortex were also indis-tinguishable; they ran through the IZ dorsal to the lateralcortex. Similar results were obtained for E14.5 embryos. Inboth heterozygous (n � 3) and homozygous mice (n � 3),trailing processes extended from the bottom of the cell bodystraight toward the VZ (Fig. 2E,F,I–L). Taken together with ourprevious observation that the trailing process represents themigratory path of excitatory neurons in the CP (Hatanaka andMurakami, 2002), this observation supports the view that ra-dial migration occurred normally in homozygous mice. Al-though subtle axonal defasciculation in Nrp1 mutant mice wasobserved at E14.5 (Fig. 2I,J), trajectories of axons extendingfrom the dorsal cortex appeared unaffected. These resultsindicated that both the corticofugal axonal extension and thecellular migration of early-born neurons were largely normal inthe absence of NRP1, at least up to E14.5.
Research in Systems NeuroscienceThe Journal of Comparative Neurology
219ROLE OF NEUROPILIN 1 IN CALLOSAL AXON GUIDANCE

Cellular migration at later developmental stagesappeared normal, but their axonal pathfinding
was disrupted by overexpression of Nrp1DN
Nrp1 mutant mice die by E15.5 in the ICR background. Toexamine the role of NRP1 in axonal extension and cellular
migration at later developmental stages, we utilized a domi-nant negative form of Nrp1 (Nrp1DN; Serini et al., 2003), whichlacks extracellular domains, two CUB domains (a1 and a2)and two FN domains (b1 and b2; Fig. 3A). This constructinterferes with either NRP1 or NRP2 binding to Sema3 pro-teins, probably by impairing the homo- and heterooligomer-
Figure 2.Axonal extension and migration of excitatory neurons in Nrp1 mutant mice. A: A 200-�m coronal slice of a Nrp1 mutant mouse at E13.5.A CP-like structure (arrows) was observed in the lateral portion of the cerebral wall under reflected light. B–L: Retrograde labeling of thecortical projection neurons by inserting DiI into the IZ. B: Illustration indicating the position of DiI in the dorsal and lateral cortex. C,D,G,H:DiI labeling in heterozygous (C,G) and homozygous (D,H) mice at E13.5. Labeled cells in the lateral portion (indicated as rectangles) weremagnified (G,H). E,F,I–L: DiI labeling in heterozygous (E,I,K) and homozygous (F,J,L) mice at E14.5. Labeled axons in the IZ (I,J) and cellsin the lateral portion (K,L) were magnified. Neuronal migration seemed to occur normally in either genotype at both ages. Extension of thetrailing process straight toward the ventricle (arrows) suggests the proper occurrence of radial migration in homozygous mice. Only axonsin the IZ showed loose bundles at E14.5 in homozygous mice. LV, lateral ventricle. Scale bars � 500 �m in A; 300 �m in F (applies to C–F);50 �m in L (applies to G–L).
Research in Systems Neuroscience The Journal of Comparative Neurology
220 Y. HATANAKA ET AL.
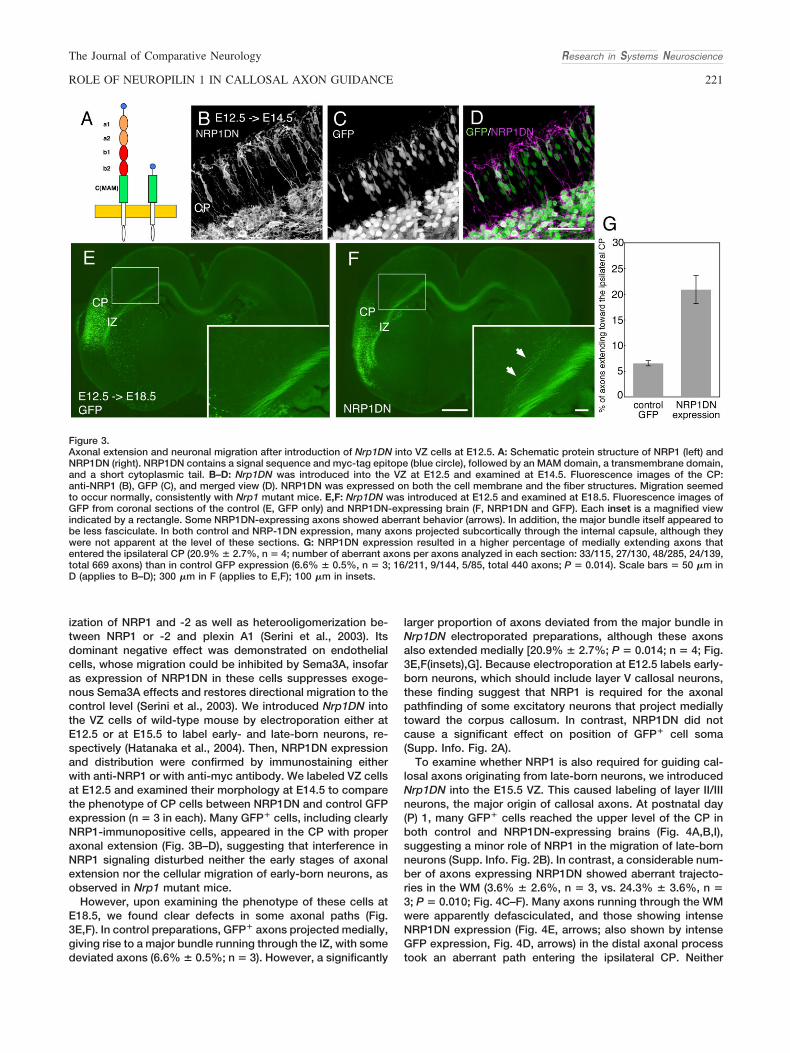
ization of NRP1 and -2 as well as heterooligomerization be-tween NRP1 or -2 and plexin A1 (Serini et al., 2003). Itsdominant negative effect was demonstrated on endothelialcells, whose migration could be inhibited by Sema3A, insofaras expression of NRP1DN in these cells suppresses exoge-nous Sema3A effects and restores directional migration to thecontrol level (Serini et al., 2003). We introduced Nrp1DN intothe VZ cells of wild-type mouse by electroporation either atE12.5 or at E15.5 to label early- and late-born neurons, re-spectively (Hatanaka et al., 2004). Then, NRP1DN expressionand distribution were confirmed by immunostaining eitherwith anti-NRP1 or with anti-myc antibody. We labeled VZ cellsat E12.5 and examined their morphology at E14.5 to comparethe phenotype of CP cells between NRP1DN and control GFPexpression (n � 3 in each). Many GFP� cells, including clearlyNRP1-immunopositive cells, appeared in the CP with properaxonal extension (Fig. 3B–D), suggesting that interference inNRP1 signaling disturbed neither the early stages of axonalextension nor the cellular migration of early-born neurons, asobserved in Nrp1 mutant mice.
However, upon examining the phenotype of these cells atE18.5, we found clear defects in some axonal paths (Fig.3E,F). In control preparations, GFP� axons projected medially,giving rise to a major bundle running through the IZ, with somedeviated axons (6.6% � 0.5%; n � 3). However, a significantly
larger proportion of axons deviated from the major bundle inNrp1DN electroporated preparations, although these axonsalso extended medially [20.9% � 2.7%; P � 0.014; n � 4; Fig.3E,F(insets),G]. Because electroporation at E12.5 labels early-born neurons, which should include layer V callosal neurons,these finding suggest that NRP1 is required for the axonalpathfinding of some excitatory neurons that project mediallytoward the corpus callosum. In contrast, NRP1DN did notcause a significant effect on position of GFP� cell soma(Supp. Info. Fig. 2A).
To examine whether NRP1 is also required for guiding cal-losal axons originating from late-born neurons, we introducedNrp1DN into the E15.5 VZ. This caused labeling of layer II/IIIneurons, the major origin of callosal axons. At postnatal day(P) 1, many GFP� cells reached the upper level of the CP inboth control and NRP1DN-expressing brains (Fig. 4A,B,I),suggesting a minor role of NRP1 in the migration of late-bornneurons (Supp. Info. Fig. 2B). In contrast, a considerable num-ber of axons expressing NRP1DN showed aberrant trajecto-ries in the WM (3.6% � 2.6%, n � 3, vs. 24.3% � 3.6%, n �3; P � 0.010; Fig. 4C–F). Many axons running through the WMwere apparently defasciculated, and those showing intenseNRP1DN expression (Fig. 4E, arrows; also shown by intenseGFP expression, Fig. 4D, arrows) in the distal axonal processtook an aberrant path entering the ipsilateral CP. Neither
Figure 3.Axonal extension and neuronal migration after introduction of Nrp1DN into VZ cells at E12.5. A: Schematic protein structure of NRP1 (left) andNRP1DN (right). NRP1DN contains a signal sequence and myc-tag epitope (blue circle), followed by an MAM domain, a transmembrane domain,and a short cytoplasmic tail. B–D: Nrp1DN was introduced into the VZ at E12.5 and examined at E14.5. Fluorescence images of the CP:anti-NRP1 (B), GFP (C), and merged view (D). NRP1DN was expressed on both the cell membrane and the fiber structures. Migration seemedto occur normally, consistently with Nrp1 mutant mice. E,F: Nrp1DN was introduced at E12.5 and examined at E18.5. Fluorescence images ofGFP from coronal sections of the control (E, GFP only) and NRP1DN-expressing brain (F, NRP1DN and GFP). Each inset is a magnified viewindicated by a rectangle. Some NRP1DN-expressing axons showed aberrant behavior (arrows). In addition, the major bundle itself appeared tobe less fasciculate. In both control and NRP-1DN expression, many axons projected subcortically through the internal capsule, although theywere not apparent at the level of these sections. G: NRP1DN expression resulted in a higher percentage of medially extending axons thatentered the ipsilateral CP (20.9% � 2.7%, n � 4; number of aberrant axons per axons analyzed in each section: 33/115, 27/130, 48/285, 24/139,total 669 axons) than in control GFP expression (6.6% � 0.5%, n � 3; 16/211, 9/144, 5/85, total 440 axons; P � 0.014). Scale bars � 50 �m inD (applies to B–D); 300 �m in F (applies to E,F); 100 �m in insets.
Research in Systems NeuroscienceThe Journal of Comparative Neurology
221ROLE OF NEUROPILIN 1 IN CALLOSAL AXON GUIDANCE
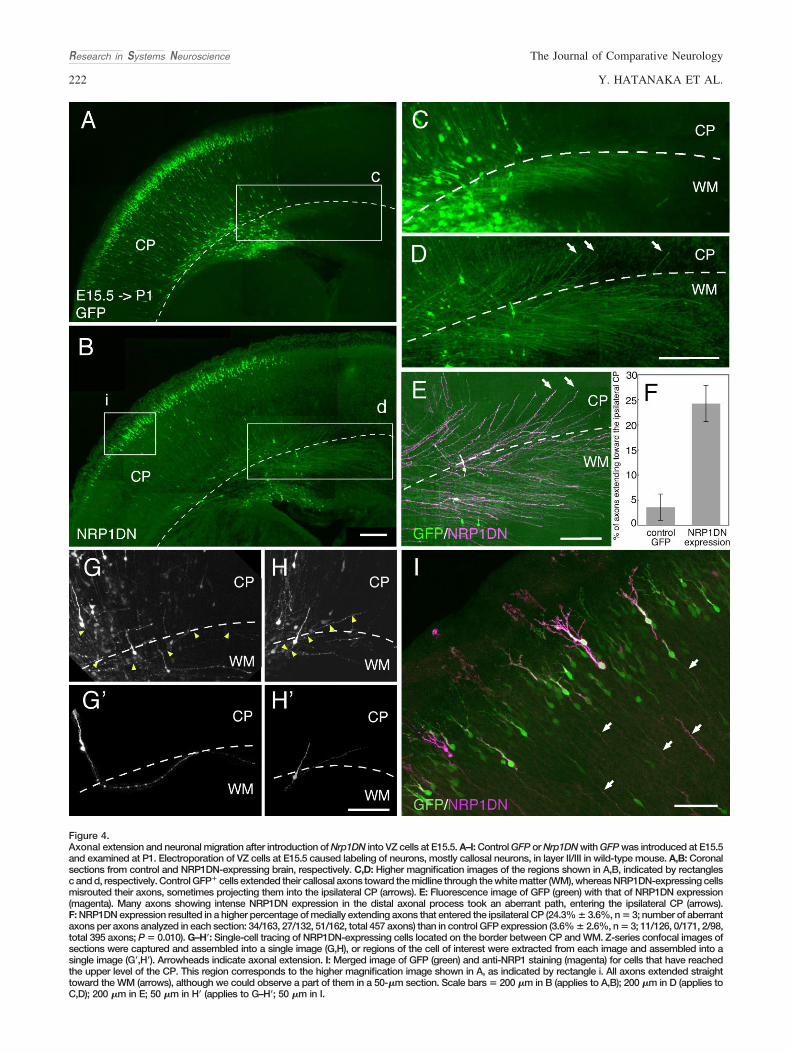
Figure 4.Axonal extension and neuronal migration after introduction of Nrp1DN into VZ cells at E15.5. A–I: Control GFP or Nrp1DN with GFP was introduced at E15.5and examined at P1. Electroporation of VZ cells at E15.5 caused labeling of neurons, mostly callosal neurons, in layer II/III in wild-type mouse. A,B: Coronalsections from control and NRP1DN-expressing brain, respectively. C,D: Higher magnification images of the regions shown in A,B, indicated by rectanglesc and d, respectively. Control GFP� cells extended their callosal axons toward the midline through the white matter (WM), whereas NRP1DN-expressing cellsmisrouted their axons, sometimes projecting them into the ipsilateral CP (arrows). E: Fluorescence image of GFP (green) with that of NRP1DN expression(magenta). Many axons showing intense NRP1DN expression in the distal axonal process took an aberrant path, entering the ipsilateral CP (arrows).F: NRP1DN expression resulted in a higher percentage of medially extending axons that entered the ipsilateral CP (24.3% � 3.6%, n � 3; number of aberrantaxons per axons analyzed in each section: 34/163, 27/132, 51/162, total 457 axons) than in control GFP expression (3.6% � 2.6%, n � 3; 11/126, 0/171, 2/98,total 395 axons; P � 0.010). G–H�: Single-cell tracing of NRP1DN-expressing cells located on the border between CP and WM. Z-series confocal images ofsections were captured and assembled into a single image (G,H), or regions of the cell of interest were extracted from each image and assembled into asingle image (G�,H�). Arrowheads indicate axonal extension. I: Merged image of GFP (green) and anti-NRP1 staining (magenta) for cells that have reachedthe upper level of the CP. This region corresponds to the higher magnification image shown in A, as indicated by rectangle i. All axons extended straighttoward the WM (arrows), although we could observe a part of them in a 50-�m section. Scale bars � 200 �m in B (applies to A,B); 200 �m in D (applies toC,D); 200 �m in E; 50 �m in H� (applies to G–H�; 50 �m in I.
Research in Systems Neuroscience The Journal of Comparative Neurology
222 Y. HATANAKA ET AL.

control nor NRP1DN-expressing axons seemed to project lat-erally toward the internal capsule. The axonal pathfindingerror was also confirmed at the single-cell level (Fig. 4G–H�).NRP1DN-expressing cells, located at the border between theCP and the WM, turned their axonal extension from the WMtoward the CP (arrowheads); however, their radial axonal ex-tension observed within the CP seemed to be unaffected (Fig.4G–H�; see also Fig. 4B,I, arrowheads).
DISCUSSIONNRP1 plays important roles in axonal extension and cellular
migration (Huber et al., 2005; Kawasaki et al., 2002; Kitsukawaet al., 1997; Marı́n et al., 2001; Schwarz et al., 2004; Tamamakiet al., 2003a). The potential involvement of NRP1 in the axonalguidance and migratory process of excitatory neurons in thecerebral cortex has been inferred from its abundant expres-sion in the IZ. In the present study, we clarified that theNRP1-immunoreactive region coincided with the localizationof migrating excitatory neurons during the period when theyexhibited a stellate shape. Analysis of Nrp1 mutant mice andNrp1DN expression experiments indicated that NRP1 is in-volved in the guidance of callosal axons but failed to supportits role in the migration of cortical excitatory neurons. More-over, the present finding that pathfinding errors were ob-served only in the WM suggests differential roles for NRP1 inaxonal extension within the CP and the WM.
Involvement of NRP1 in callosal axon guidanceTo examine the role of NRP1 in excitatory neurons, we used
Nrp1 mutant mice and forced Nrp1DN expression in thesecells by electroporation. By introducing the Nrp1DN plasmidinto the cortical VZ at two different developmental stages,E12.5 or E15.5, we could dissect the role of NRP1 in twopopulations: early (E12.5)-born neurons, which are mainlylayer V and VI neurons that project subcortically and cal-losally, and layer II/III late (E15.5)-born neurons, which projectaxons mainly callosally in rodents (Hatanaka et al., 2004;Jones, 1984; Koester and O’Leary, 1993; Molnar and Cheung,2006). Analyses of the cortical axonal tracts of Nrp1 mutantsand growth pattern of axons from NRP1DN-expresing cellsshowed a difference in axonal guidance errors between early-and late-born neurons. The direction and fasciculation of ax-onal growth from early-born neurons at early stages werelargely normal: corticofugal axons extended from the dorsal tolateral direction through the IZ and then extended toward theinternal capsule. However, at a later stage, a notable guidanceerror occurred for axons extending medially, suggesting aspecific effect of NRP1 signaling on callosal projection. Con-sistently with this result, introducing Nrp1DN into late-bornneurons caused a significant axonal guidance error towardthe corpus callosum. The fact that introducing a dominantnegative form of plexinA1 (Takahashi et al., 1999), whichshould attenuate the NRP1 signaling, in late-born neuronscaused a similar guidance error (Hatanaka et al., unpublishedobservation) also suggests involvement of NRP1 signaling incallosal axonal guidance. These results suggest that NRP1plays an important role in the fasciculation and guidance ofcallosal axons, irrespective of their origin.
Although a considerable number of NRP1DN-expressingcallosal axons showed aberrant behavior, many GFP-coexpressing callosal axons still projected normally. This may
be because only high-level expression NRP1DN affects ax-onal extension.
Medially projecting neurons also include corticocorticalneurons that project their axons to ipsilateral fields situatednearby. However, it seems unlikely that the affected axonsobserved here are ipsilaterally projecting axons, because 1)the corticocortical axons from layer VI neurons extend laterthan that of callosal axons, and they course tangentiallythrough layers V and VI away from the white matter (Arimatsuand Ishida, 2002), and 2) the corticocortical neurons in layerII/III tend to be situated superficially compared with callosalneurons (Jones, 1984). Indeed, we observed that labeling ofVZ cells at a later stage (E16.5) caused mostly superficiallylocated neurons and an increase in the proportion of labeledaxons projecting to the ipsilateral fields (Hatanaka et al., un-published observation), supporting the view that the majorityof layer II/III neurons labeled at E15.5 are callosal neurons.
One might argue that the guidance errors of callosal axonswere caused by suppression of NRP2 rather than NRP1, be-cause NRP1DN can also bind to NRP2, which is expressed inthe developing cerebral cortex (Bagnard et al., 1998; Chen etal., 1997). However, this is unlikely, because callosal axons inNrp2 knockout mice did not show aberrant behavior as ob-served here (Hatanaka et al., unpublished observation).
Dissection of the pathfinding errors in the callosalaxon
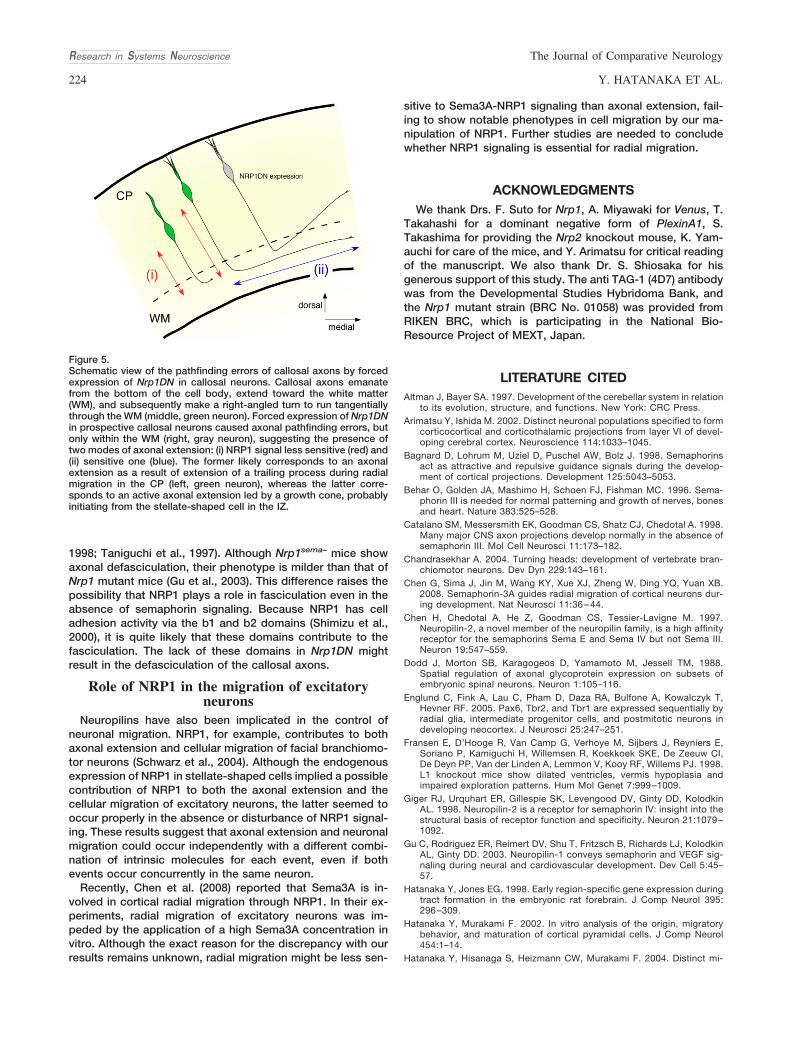
Axons of excitatory projection neurons in the cerebral cor-tex emanate from the bottom of the cell body, extend towardthe white matter, and subsequently make a right-angled turnto run tangentially through the WM. In the present study, wefound that expression of NRP1DN in callosal neurons causedaxonal pathfinding errors, but only within the WM (Fig. 5). Alikely explanation for this finding is that radial axonal exten-sion within the CP occurs as a result of radial migration ofexcitatory neurons rather than extension led by an axonalgrowth cone (Hatanaka and Murakami, 2002). We have previ-ously obtained evidence that the trailing processes of radiallymigrating excitatory neurons turn into axons (Hatanaka andMurakami, 2002). On the other hand, tangential axonal exten-sion led by an axonal growth cone may be initiated fromstellate-shaped cells in the IZ. Thus, the present results canbe interpreted to indicate that the tangential extension ofcallosal axons in the IZ is more sensitive to NRP1 signalingthan the radial extension of axons as trailing processes in theCP.
Role of NRP1 in the axonal fasciculation ofcortical efferents
NRP1DN-expressing axons from late-born neurons showeddefasciculation and misrouting. Our findings that weak-to-moderate Nrp1DN expression caused defasciculation andthat strong expression resulted in complete misrouting sug-gest that all these phenotypes are due primarily to defascicu-lation.
Although NRP1 exerts a repellent activity via interactionswith semaphorins, a recent study indicated that it is involvedin the fasciculation of neurites from sympathetic ganglioncells in a Sema3A-dependent manner (Kawasaki et al., 2002).Consistently, defasciculation of axonal tracts was observed inNrp1 mutant mice (Huber et al., 2005; Kitsukawa et al., 1997)and Sema3A mutant mice (Behar et al., 1996; Catalano et al.,
Research in Systems NeuroscienceThe Journal of Comparative Neurology
223ROLE OF NEUROPILIN 1 IN CALLOSAL AXON GUIDANCE
1998; Taniguchi et al., 1997). Although Nrp1sema– mice showaxonal defasciculation, their phenotype is milder than that ofNrp1 mutant mice (Gu et al., 2003). This difference raises thepossibility that NRP1 plays a role in fasciculation even in theabsence of semaphorin signaling. Because NRP1 has celladhesion activity via the b1 and b2 domains (Shimizu et al.,2000), it is quite likely that these domains contribute to thefasciculation. The lack of these domains in Nrp1DN mightresult in the defasciculation of the callosal axons.
Role of NRP1 in the migration of excitatoryneurons
Neuropilins have also been implicated in the control ofneuronal migration. NRP1, for example, contributes to bothaxonal extension and cellular migration of facial branchiomo-tor neurons (Schwarz et al., 2004). Although the endogenousexpression of NRP1 in stellate-shaped cells implied a possiblecontribution of NRP1 to both the axonal extension and thecellular migration of excitatory neurons, the latter seemed tooccur properly in the absence or disturbance of NRP1 signal-ing. These results suggest that axonal extension and neuronalmigration could occur independently with a different combi-nation of intrinsic molecules for each event, even if bothevents occur concurrently in the same neuron.
Recently, Chen et al. (2008) reported that Sema3A is in-volved in cortical radial migration through NRP1. In their ex-periments, radial migration of excitatory neurons was im-peded by the application of a high Sema3A concentration invitro. Although the exact reason for the discrepancy with ourresults remains unknown, radial migration might be less sen-
sitive to Sema3A-NRP1 signaling than axonal extension, fail-ing to show notable phenotypes in cell migration by our ma-nipulation of NRP1. Further studies are needed to concludewhether NRP1 signaling is essential for radial migration.
ACKNOWLEDGMENTSWe thank Drs. F. Suto for Nrp1, A. Miyawaki for Venus, T.
Takahashi for a dominant negative form of PlexinA1, S.Takashima for providing the Nrp2 knockout mouse, K. Yam-auchi for care of the mice, and Y. Arimatsu for critical readingof the manuscript. We also thank Dr. S. Shiosaka for hisgenerous support of this study. The anti TAG-1 (4D7) antibodywas from the Developmental Studies Hybridoma Bank, andthe Nrp1 mutant strain (BRC No. 01058) was provided fromRIKEN BRC, which is participating in the National Bio-Resource Project of MEXT, Japan.
LITERATURE CITEDAltman J, Bayer SA. 1997. Development of the cerebellar system in relation
to its evolution, structure, and functions. New York: CRC Press.Arimatsu Y, Ishida M. 2002. Distinct neuronal populations specified to form
corticocortical and corticothalamic projections from layer VI of devel-oping cerebral cortex. Neuroscience 114:1033–1045.
Bagnard D, Lohrum M, Uziel D, Puschel AW, Bolz J. 1998. Semaphorinsact as attractive and repulsive guidance signals during the develop-ment of cortical projections. Development 125:5043–5053.
Behar O, Golden JA, Mashimo H, Schoen FJ, Fishman MC. 1996. Sema-phorin III is needed for normal patterning and growth of nerves, bonesand heart. Nature 383:525–528.
Catalano SM, Messersmith EK, Goodman CS, Shatz CJ, Chedotal A. 1998.Many major CNS axon projections develop normally in the absence ofsemaphorin III. Mol Cell Neurosci 11:173–182.
Chandrasekhar A. 2004. Turning heads: development of vertebrate bran-chiomotor neurons. Dev Dyn 229:143–161.
Chen G, Sima J, Jin M, Wang KY, Xue XJ, Zheng W, Ding YQ, Yuan XB.2008. Semaphorin-3A guides radial migration of cortical neurons dur-ing development. Nat Neurosci 11:36 – 44.
Chen H, Chedotal A, He Z, Goodman CS, Tessier-Lavigne M. 1997.Neuropilin-2, a novel member of the neuropilin family, is a high affinityreceptor for the semaphorins Sema E and Sema IV but not Sema III.Neuron 19:547–559.
Dodd J, Morton SB, Karagogeos D, Yamamoto M, Jessell TM. 1988.Spatial regulation of axonal glycoprotein expression on subsets ofembryonic spinal neurons. Neuron 1:105–116.
Englund C, Fink A, Lau C, Pham D, Daza RA, Bulfone A, Kowalczyk T,Hevner RF. 2005. Pax6, Tbr2, and Tbr1 are expressed sequentially byradial glia, intermediate progenitor cells, and postmitotic neurons indeveloping neocortex. J Neurosci 25:247–251.
Fransen E, D’Hooge R, Van Camp G, Verhoye M, Sijbers J, Reyniers E,Soriano P, Kamiguchi H, Willemsen R, Koekkoek SKE, De Zeeuw CI,De Deyn PP, Van der Linden A, Lemmon V, Kooy RF, Willems PJ. 1998.L1 knockout mice show dilated ventricles, vermis hypoplasia andimpaired exploration patterns. Hum Mol Genet 7:999 –1009.
Giger RJ, Urquhart ER, Gillespie SK, Levengood DV, Ginty DD, KolodkinAL. 1998. Neuropilin-2 is a receptor for semaphorin IV: insight into thestructural basis of receptor function and specificity. Neuron 21:1079 –1092.
Gu C, Rodriguez ER, Reimert DV, Shu T, Fritzsch B, Richards LJ, KolodkinAL, Ginty DD. 2003. Neuropilin-1 conveys semaphorin and VEGF sig-naling during neural and cardiovascular development. Dev Cell 5:45–57.
Hatanaka Y, Jones EG. 1998. Early region-specific gene expression duringtract formation in the embryonic rat forebrain. J Comp Neurol 395:296 –309.
Hatanaka Y, Murakami F. 2002. In vitro analysis of the origin, migratorybehavior, and maturation of cortical pyramidal cells. J Comp Neurol454:1–14.
Hatanaka Y, Hisanaga S, Heizmann CW, Murakami F. 2004. Distinct mi-
Figure 5.Schematic view of the pathfinding errors of callosal axons by forcedexpression of Nrp1DN in callosal neurons. Callosal axons emanatefrom the bottom of the cell body, extend toward the white matter(WM), and subsequently make a right-angled turn to run tangentiallythrough the WM (middle, green neuron). Forced expression of Nrp1DNin prospective callosal neurons caused axonal pathfinding errors, butonly within the WM (right, gray neuron), suggesting the presence oftwo modes of axonal extension: (i) NRP1 signal less sensitive (red) and(ii) sensitive one (blue). The former likely corresponds to an axonalextension as a result of extension of a trailing process during radialmigration in the CP (left, green neuron), whereas the latter corre-sponds to an active axonal extension led by a growth cone, probablyinitiating from the stellate-shaped cell in the IZ.
Research in Systems Neuroscience The Journal of Comparative Neurology
224 Y. HATANAKA ET AL.

gratory behavior of early- and late-born neurons derived from thecortical ventricular zone. J Comp Neurol 479:1–14.
Hatten ME, Heintz N. 1995. Mechanisms of neural patterning and speci-fication in the developing cerebellum. Annu Rev Neurosci 18:385– 408.
He Z, Tessier-Lavigne M. 1997. Neuropilin is a receptor for the axonalchemorepellent semaphorin III. Cell 90:739 –751.
Hirai S, Kawaguchi A, Hirasawa R, Baba M, Ohnishi T, Ohno S. 2002.MAPK-upstream protein kinase (MUK) regulates the radial migration ofimmature neurons in telencephalon of mouse embryo. Development129:4483– 4495.
Huber AB, Kania A, Tran TS, Gu C, De Marco Garcia N, Lieberam I,Johnson D, Jessell TM, Ginty DD, Kolodkin AL. 2005. Distinct roles forsecreted semaphorin signaling in spinal motor axon guidance. Neuron48:949 –964.
Jones EG. 1984. Laminar distribution of cortical efferent cells. In: Peters A,Jones EG. editor. Cerebral cortex: New York: Plenum. p 521–553.
Jones L, Lopez-Bendito G, Gruss P, Stoykova A, Molnar Z. 2002. Pax6 isrequired for the normal development of the forebrain axonal connec-tions. Development 129:5041–5052.
Jossin Y, Ogawa M, Metin C, Tissir F, Goffinet AM. 2003. Inhibition of Srcfamily kinases and non-classical protein kinases C induce a reeler-likemalformation of coritical plate development. J Neurosci 23:9953–9959.
Kawakami A, Kitsukawa T, Takagi S, Fujisawa H. 1996. Developmentallyregulated expression of a cell surface protein, neuropilin, in the mousenervous system. J Neurobiol 29:1–17.
Kawasaki T, Kitsukawa T, Bekku Y, Matsuda Y, Sanbo M, Yagi T, FujisawaH. 1999. A requirement for neuropilin-1 in embryonic vessel formation.Development 126:4895– 4902.
Kawasaki T, Bekku Y, Suto F, Kitsukawa T, Taniguchi M, Nagatsu I,Nagatsu T, Itoh K, Yagi T, Fujisawa H. 2002. Requirement of neuropilin1-mediated Sema3A signals in patterning of the sympathetic nervoussystem. Development 129:671– 680.
Kitsukawa T, Shimizu M, Sanbo M, Hirata T, Taniguchi M, Bekku Y, Yagi T,Fujisawa H. 1997. Neuropilin-semaphorin III/D-mediated chemorepul-sive signals play a crucial role in peripheral nerve projection in mice.Neuron 19:995–1005.
Koester SE, O’Leary DD. 1993. Connectional distinction between callosaland subcortically projecting cortical neurons is determined prior toaxon extension. Dev Biol 160:1–14.
Kolodkin AL, Levengood DV, Rowe EG, Tai YT, Giger RJ, Ginty DD. 1997.Neuropilin is a semaphorin III receptor. Cell 90:753–762.
Kriegstein AR, Noctor SC. 2004. Patterns of neuronal migration in theembryonic cortex. Trends Neurosci 27:392–399.
Kudo C, Ajioka I, Hirata Y, Nakajima K. 2005. Expression profiles of EphA3at both the RNA and protein level in the developing mammalian fore-brain. J Comp Neurol 487:255–269.
Marı́n O, Rubenstein JL. 2001. A long, remarkable journey: tangentialmigration in the telencephalon. Nat Rev Neurosci 2:780 –790.
Marı́n O, Yaron A, Bagri A, Tessier-Lavigne M, Rubenstein JL. 2001.Sorting of striatal and cortical interneurons regulated by semaphorin-neuropilin interactions. Science 293:872– 875.
Miyata T, Kawaguchi A, Saito K, Kuramochi H, Ogawa M. 2002. Visualiza-tion of cell cycling by an improvement in slice culture methods. J Neu-rosci Res 69:861– 868.
Molnar Z, Cheung AF. 2006. Towards the classification of subpopulationsof layer V pyramidal projection neurons. Neurosci Res 55:105–115.
Nadarajah B, Parnavelas JG. 2002. Modes of neuronal migration in thedeveloping cerebral cortex. Nat Rev Neurosci 3:423– 432.
Nagai T, Ibata K, Park ES, Kubota M, Mikoshiba K, Miyawaki A. 2002. Avariant of yellow fluorescent protein with fast and efficient maturationfor cell-biological applications. Nat Biotechnol 20:87–90.
Nieto M, Monuki ES, Tang H, Imitola J, Haubst N, Khoury SJ, CunninghamJ, Gotz M, Walsh CA. 2004. Expression of Cux-1 and Cux-2 in thesubventricular zone and upper layers II–IV of the cerebral cortex.J Comp Neurol 479:168 –180.
Niwa H, Yamamura K, Miyazaki J. 1991. Efficient selection for high-
expression transfectants with a novel eukaryotic vector. Gene 108:193–199.
Noctor SC, Martinez-Cerdeno V, Ivic L, Kriegstein AR. 2004. Corticalneurons arise in symmetric and asymmetric division zones and migratethrough specific phases. Nat Neurosci 7:136 –144.
Parnavelas JG. 2000. The origin and migration of cortical neurones: newvistas. Trends Neurosci 23:126 –131.
Rathjen FG, Schachner M. 1984. Immunocytological and biochemicalcharacterization of a new neuronal cell surface component (L1 antigen)which is involved in cell adhesion. EMBO J 3:1–10.
Renzi MJ, Feiner L, Koppel AM, Raper JA. 1999. A dominant negativereceptor for specific secreted semaphorins is generated by deleting anextracellular domain from neuropilin-1. J Neurosci 19:7870 –7880.
Schwarz Q, Gu C, Fujisawa H, Sabelko K, Gertsenstein M, Nagy A,Taniguchi M, Kolodkin AL, Ginty DD, Shima DT, Ruhrberg C. 2004.Vascular endothelial growth factor controls neuronal migration andcooperates with Sema3A to pattern distinct compartments of the facialnerve. Genes Dev 18:2822–2834.
Serini G, Valdembri D, Zanivan S, Morterra G, Burkhardt C, Caccavari F,Zammataro L, Primo L, Tamagnone L, Logan M, Tessier-Lavigne M,Taniguchi M, Puschel AW, Bussolino F. 2003. Class 3 semaphorinscontrol vascular morphogenesis by inhibiting integrin function. Nature424:391–397.
Shimizu M, Murakami Y, Suto F, Fujisawa H. 2000. Determination of celladhesion sites of neuropilin-1. J Cell Biol 148:1283–1293.
Shoukimas GM, Hinds JW. 1978. The development of the cerebral cortexin the embryonic mouse: an electron microscopic serial section anal-ysis. J Comp Neurol 179:795– 830.
Tabata H, Nakajima K. 2003. Multipolar migration: the third mode of radialneuronal migration in the developing cerebral cortex. J Neurosci 23:9996 –10001.
Takagi S, Kasuya Y, Shimizu M, Matsuura T, Tsuboi M, Kawakami A,Fujisawa H. 1995. Expression of a cell adhesion molecule, neuropilin,in the developing chick nervous system. Dev Biol 170:207–222.
Takahashi T, Fournier A, Nakamura F, Wang LH, Murakami Y, Kalb RG,Fujisawa H, Strittmatter SM. 1999. Plexin-neuropilin-1 complexes formfunctional semaphorin-3A receptors. Cell 99:59 – 69.
Tamagnone L, Artigiani S, Chen H, He Z, Ming GI, Song H, Chedotal A,Winberg ML, Goodman CS, Poo M, Tessier-Lavigne M, Comoglio PM.1999. Plexins are a large family of receptors for transmembrane,secreted, and GPI-anchored semaphorins in vertebrates. Cell 99:71–80.
Tamamaki N, Fujimori K, Nojyo Y, Kaneko T, Takauji R. 2003a. Evidencethat Sema3A and Sema3F regulate the migration of GABAergic neu-rons in the developing neocortex. J Comp Neurol 455:238 –248.
Tamamaki N, Yanagawa Y, Tomioka R, Miyazaki J, Obata K, Kaneko T.2003b. Green fluorescent protein expression and colocalization withcalretinin, parvalbumin, and somatostatin in the GAD67-GFP knock-inmouse. J Comp Neurol 467:60 –79.
Tanaka D, Nakaya Y, Yanagawa Y, Obata K, Murakami F. 2003. Multimodaltangential migration of neocortical GABAergic neurons independent ofGPI-anchored proteins. Development 130:5803–5813.
Tanaka DH, Maekawa K, Yanagawa Y, Obata K, Murakami F. 2006. Mul-tidirectional and multizonal tangential migration of GABAergic inter-neurons in the developing cerebral cortex. Deveolopment 133:2167–2176.
Taniguchi M, Yuasa S, Fujisawa H, Naruse I, Saga S, Mishina M, Yagi T.1997. Disruption of semaphorin III/D gene causes severe abnormalityin peripheral nerve projection. Neuron 19:519 –530.
Tarabykin V, Stoykova A, Usman N, Gruss P. 2001. Cortical upper layerneurons derive from the subventricular zone as indicated by Svet1gene expression. Development 128:1983–1993.
Yamamoto M, Boyer AM, Crandall JE, Edwards M, Tanaka H. 1986.Distribution of stage-specific neurite-associated proteins in the devel-oping murine nervous system recognized by a monoclonal antibody.J Neurosci 6:3576 –3594.
Research in Systems NeuroscienceThe Journal of Comparative Neurology
225ROLE OF NEUROPILIN 1 IN CALLOSAL AXON GUIDANCE












![Neuronal circuits and physiological roles of the basal ... · addictive drugs [4]. Dopaminergic neurons in the sub-stantia nigra pars compacta project their axons towards medium spiny](https://static.fdocument.pub/doc/165x107/5f09eefe7e708231d4293437/neuronal-circuits-and-physiological-roles-of-the-basal-addictive-drugs-4.jpg)




